
Proliferating Cholangiocytes: A Neuroendocrine Compartment in the Diseased Liver
17
SPECIAL REPORTS AND REVIEWS Proliferating Cholangiocytes: A Neuroendocrine Compartment in the Diseased Liver DOMENICO ALVARO,* ,‡ MARIA GRAZIA MANCINO,* SHANNON GLASER, §,# EUGENIO GAUDIO,** MARCO MARZIONI, HEATHER FRANCIS, § and GIANFRANCO ALPINI ¶,#,‡‡ *Division of Gastroenterology, Department of Clinical Medicine, Rome, Italy; ‡ Polo Pontino, Latina, Italy; ¶ Central Texas Veterans Health Care System, # Department of Medicine and ‡‡ Systems Biology and Translational Medicine, § Division of Research and Education, Scott & White Hospital and The Texas A&M University System Health Science Center, College of Medicine, Temple, Texas; **Anatomy, University “La Sapienza,” Rome, Italy; Department Gastroenterology, Università Politecnica delle Marche, Ancona, Italy See Charatcharoenwitthaya P et al on page 32 for companion article in Clin Gastroenterol Hepatol. In the last 15 years, the intrahepatic biliary tree has become the object of extensive studies, which high- lighted the extraordinary biologic properties of cholangiocytes involved in bile formation, prolifer- ation, injury repair, fibrosis, angiogenesis, and reg- ulation of blood flow. Proliferation is a “typical” property of cholangiocytes and is key as a mecha- nism of repair responsible for maintaining the in- tegrity of the biliary tree. Cholangiocyte prolifera- tion occurs virtually in all pathologic conditions of liver injury where it is associated with inflamma- tion, regeneration, and repair, thus conditioning the evolution of liver damage. Interestingly, prolif- erating cholangiocytes acquire the phenotype of neuroendocrine cells, and secrete different cyto- kines, growth factors, neuropeptides, and hor- mones, which represent potential mechanisms for cross talk with other liver cells. Many studies sug- gest the generation of a neuroendocrine compart- ment in the injured liver, mostly constituted by cells with cholangiocyte features, which function- ally conditions the progression of liver disease. These insights on cholangiocyte pathophysiology will provide new potential strategies for the man- agement of chronic liver diseases. The purpose of this review is to summarize the recent findings on the mechanisms regulating cholangiocyte prolifer- ation and the significance of the neuroendocrine regulation of cholangiocyte biology. T he intrahepatic biliary tree is a complex 3-dimen- sional network of interconnected ducts lined by cholangiocytes (Figure 1). 1,2 The nomenclature of the finer branches of the biliary tree has been recently re- vised. 3 The finest proximal branch of the intrahepatic biliary tree is the canal of Hering, which represents the link between the bile canaliculus and the biliary tree. 4,5 The canal of Hering is localized intralobularly extending into the parenchyma for 1/3 the distance to the terminal hepatic vein, is lined by hepatocytes and cholangiocytes, contains the bipotential hepatic progenitor cells (HPC), and serves as the site for influx and localization of intra- and extrahepatic adult stem cells. 5–7 The canal of Hering continues into bile ductules, entirely lined by cholangio- cytes, which may traverse the limiting plate including both intralobular and intraportal localization. 6,7 Bile ductules converge into interlobular bile ducts, which are located in the portal space, and then continue into ducts of major size. 6,7 Sympathetic and parasympathetic nerves are located around the intrahepatic and extrahepatic bile ducts. 8 Sympathetic (originating from the celiac ganglion) and parasympathetic (from the vagus nerve) nerves are orga- nized in the anterior plexus around the hepatic artery and a posterior plexus surrounding portal vein and bile duct. 8 Other than the classical neurotransmitters, many different neuropeptides including neuropeptide tyrosine, C-terminal flanking peptide of neuropeptide tyrosine, -calcitonin gene-related peptide, somatostatin, vasoac- Abbreviations used in this paper: 6-OHDA, 6-hydroxydopamine; AC, adenylate cyclase; ADPKD, abnormal dominant polycystic kidney dis- ease; BDL, bile duct ligation; cAMP, adenosine 3’,5’-cyclic monophos- phate; [Ca 2 ] i , intracellular Ca 2 ; ER, endoplasmic reticulum; ERK, extracellular regulated kinases; HPC, hepatic progenitor cells; IGF-1, insulin-like growth factor-1; IGFBP, IGF binding protein; IP 3 , inositol 1,4,5-trisphosphate; IRS, insulin receptor substrates; NGF, nerve growth factor; PI3K, phosphatidylinositol 3-kinase; PBC, primary biliary cirrhosis; PBP, peribiliary vascular plexus; PKA, protein kinase A; PKC, protein kinase C; PSC, primary sclerosing cholangitis; VEGF, vascular endothelial growth factor. © 2007 by the AGA Institute 0016-5085/07/$32.00 doi:10.1053/j.gastro.2006.07.023 GASTROENTEROLOGY 2007;132:415– 431
Transcript of Proliferating Cholangiocytes: A Neuroendocrine Compartment in the Diseased Liver

S
PD
DH
*MHd
IblcaupnttlttenkmcgmcaTwattar
Tc
GASTROENTEROLOGY 2007;132:415– 431
PECIAL REPORTS AND REVIEWS
roliferating Cholangiocytes: A Neuroendocrine Compartment in theiseased Liver
OMENICO ALVARO,*,‡ MARIA GRAZIA MANCINO,* SHANNON GLASER,§,# EUGENIO GAUDIO,** MARCO MARZIONI,�
EATHER FRANCIS,§ and GIANFRANCO ALPINI¶,#,‡‡
Division of Gastroenterology, Department of Clinical Medicine, Rome, Italy; ‡Polo Pontino, Latina, Italy; ¶Central Texas Veterans Health Care System, #Department ofedicine and ‡‡Systems Biology and Translational Medicine, §Division of Research and Education, Scott & White Hospital and The Texas A&M University System
�
ealth Science Center, College of Medicine, Temple, Texas; **Anatomy, University “La Sapienza,” Rome, Italy; Department Gastroenterology, Università Politecnicaelle Marche, Ancona, ItalyfivblTihcaaccbdlo
aSpnaddC�
aepei1gcpe
See Charatcharoenwitthaya P et al on page 32for companion article in Clin GastroenterolHepatol.
n the last 15 years, the intrahepatic biliary tree hasecome the object of extensive studies, which high-
ighted the extraordinary biologic properties ofholangiocytes involved in bile formation, prolifer-tion, injury repair, fibrosis, angiogenesis, and reg-lation of blood flow. Proliferation is a “typical”roperty of cholangiocytes and is key as a mecha-ism of repair responsible for maintaining the in-
egrity of the biliary tree. Cholangiocyte prolifera-ion occurs virtually in all pathologic conditions ofiver injury where it is associated with inflamma-ion, regeneration, and repair, thus conditioninghe evolution of liver damage. Interestingly, prolif-rating cholangiocytes acquire the phenotype ofeuroendocrine cells, and secrete different cyto-ines, growth factors, neuropeptides, and hor-ones, which represent potential mechanisms for
ross talk with other liver cells. Many studies sug-est the generation of a neuroendocrine compart-ent in the injured liver, mostly constituted by
ells with cholangiocyte features, which function-lly conditions the progression of liver disease.hese insights on cholangiocyte pathophysiologyill provide new potential strategies for the man-gement of chronic liver diseases. The purpose ofhis review is to summarize the recent findings onhe mechanisms regulating cholangiocyte prolifer-tion and the significance of the neuroendocrineegulation of cholangiocyte biology.
he intrahepatic biliary tree is a complex 3-dimen-sional network of interconnected ducts lined by
holangiocytes (Figure 1).1,2 The nomenclature of the
ner branches of the biliary tree has been recently re-ised.3 The finest proximal branch of the intrahepaticiliary tree is the canal of Hering, which represents the
ink between the bile canaliculus and the biliary tree.4,5
he canal of Hering is localized intralobularly extendingnto the parenchyma for 1/3 the distance to the terminalepatic vein, is lined by hepatocytes and cholangiocytes,ontains the bipotential hepatic progenitor cells (HPC),nd serves as the site for influx and localization of intra-nd extrahepatic adult stem cells.5–7 The canal of Heringontinues into bile ductules, entirely lined by cholangio-ytes, which may traverse the limiting plate includingoth intralobular and intraportal localization.6,7 Bileuctules converge into interlobular bile ducts, which are
ocated in the portal space, and then continue into ductsf major size.6,7
Sympathetic and parasympathetic nerves are locatedround the intrahepatic and extrahepatic bile ducts.8
ympathetic (originating from the celiac ganglion) andarasympathetic (from the vagus nerve) nerves are orga-ized in the anterior plexus around the hepatic arterynd a posterior plexus surrounding portal vein and bileuct.8 Other than the classical neurotransmitters, manyifferent neuropeptides including neuropeptide tyrosine,-terminal flanking peptide of neuropeptide tyrosine,-calcitonin gene-related peptide, somatostatin, vasoac-
Abbreviations used in this paper: 6-OHDA, 6-hydroxydopamine; AC,denylate cyclase; ADPKD, abnormal dominant polycystic kidney dis-ase; BDL, bile duct ligation; cAMP, adenosine 3’,5’-cyclic monophos-hate; [Ca2�]i, intracellular Ca2�; ER, endoplasmic reticulum; ERK,xtracellular regulated kinases; HPC, hepatic progenitor cells; IGF-1,
nsulin-like growth factor-1; IGFBP, IGF binding protein; IP3, inositol,4,5-trisphosphate; IRS, insulin receptor substrates; NGF, nerverowth factor; PI3K, phosphatidylinositol 3-kinase; PBC, primary biliaryirrhosis; PBP, peribiliary vascular plexus; PKA, protein kinase A; PKC,rotein kinase C; PSC, primary sclerosing cholangitis; VEGF, vascularndothelial growth factor.
© 2007 by the AGA Institute0016-5085/07/$32.00
doi:10.1053/j.gastro.2006.07.023

tbdnp
phbbatcsbPbHltpn
dmmtxt
dcqboactccipcletptRstaeocdancItdc
Fbdoaomc
Fba
416 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
ive intestinal peptide, enkephalin, and bom-esin have been detected in the liver including bileucts.9 –16 Most of the afferent nerves run via the splanch-ic branches of the vagus nerve and also to the righthrenic nerve.8
The intrahepatic biliary tree is nourished by theeribiliary vascular plexus (PBP), which originates fromepatic artery branches and flows into hepatic sinusoidsy means of connecting small venules with portal veinranches.17,18 The PBP is represented by a rich and bil-minar vascular network near the hilus of the liver andhe larger portal tracts, whereas it is formed by feweronnected vessels in the smaller portal tracts.17,18 Anyubstance reabsorbed from bile by cholangiocytes flowsack toward hepatocytes through the PBP. Therefore, theBP represents the anatomical basis for a cross talketween cholangiocytes and hepatocytes.17,18 Canals ofering and part of bile ductules lack of a direct vascu-
arization,17,18 and this could be one of the mechanismshat maintain the HPC compartment, located in thisortion of the biliary tree, in a quiescent state underormal conditions.
Cholangiocyte Proliferation inExperimental Models and Human LiverDiseasesCholangiocytes in the adult liver are mitotically
ormant,1,19 supported on a conventional basementembrane20 and surrounded by the connective tissueatrix21 and the PBP.18 The constitutive expression of
he cyclin dependent kinase inhibitors p27,22 bcl2,23 Bcl-L,23 and Mcl-124 are some of the factors responsible for
igure 1. As shown in this rat liver cholangiogram, the intrahepaticiliary epithelium is a complex 3-dimensional network of interconnecteducts lined by cholangiocytes. The rat cholangiogram shows a portionf the intrahepatic biliary tree at the periphery of the liver. The contrastgent, renographin, was injected in a retrograde fashion into bile ductsf a normal rat liver. The image of bile ducts was obtained employing aicrofocus magnification mammography instrument. Original magnifi-
ation, 320�. Reproduced with permission from ref. 2.
he maintenance of this resting state. Cholangiocytes o
isplay primary cilia in their apical pole (Figure 2), whichould be of relevance for maintaining cholangiocytes in auiescent state because, as recently proposed,25 ciliaending maintains intracellular Ca2� [Ca2�]i at a thresh-ld where cAMP, an intracellular modulator of prolifer-tion,26 –31 is inhibited. Many different insults determiningell damage, disruption of the normal cell/matrix interac-ion, or release of cytokines or soluble mediators triggerholangiocyte proliferation.21,32–34 The peculiar capacity ofholangiocytes to proliferate is evidenced in specific exper-mental conditions as well as in many different humanathological conditions.1,32–36 In most of these conditions,holangiocyte proliferation is a part of the so-called “ductu-ar reaction,” a term coined by Popper36 to identify thexpanded population of epithelial cells at the interface ofhe biliary tree and the hepatocytes, and which refers toroliferation of pre-existing ductules, progenitor cell ac-ivation, and appearance of intermediate hepatocytes.ecently, the term “ductular proliferation”3 has been
uggested instead of “ductular reaction;” however, thiserm is not completely satisfactory because the prolifer-ting reactive ductules may not simply arise from prolif-ration of pre-existing bile ductular cells, as they may alsoriginate from activated and differentiated progenitorells, from cells that entered from the circulation andifferentiate toward liver cells or, more rarely, from bili-ry metaplasia of hepatocytes.32–36 Besides the differentomenclatures, cholangiocyte proliferation, as the mainomponent of “ductular reaction,” has been classified inV type.21,32 Type I or “typical” cholangiocyte prolifera-ion results in an increased number of intrahepatic bileucts confined to portal spaces.1,37,38 The proliferatingholangiocytes form a well-differentiated 3-dimensional
igure 2. Liver from rats with BDL for 3 weeks. The luminal surface ofile ducts is shown, with lining cholangiocytes that are rich of microvillind sparse, long cilia (enlarged box). (Scanning electron microscopy,
riginal magnification, 3,500�).
nmiactcddiuctsdocctrtmsesmapp“cta
acasrlbtaaTtorspb(dprn
mpd
plhtaodahttshcimaec
tmoic“gdchit“ns
csdhtptscmst
January 2007 PROLIFERATING CHOLANGIOCYTES 417
etwork of tubular structures with a well-defined lu-en.1,37,38 In the rat, “typical” cholangiocyte proliferation
s observed after bile duct ligation (BDL),38 partial hep-tectomy,19 chronic �-naphthylisothiocyanate feeding,39
hronic L-proline treatment,40 prolonged oral adminis-ration of lithocholate, chenodeoxycholate, or tauro-holate.41,42 According to different studies, the “typical”uctal proliferation originates from pre-existing bileucts located within portal areas because: (1) proliferat-
ng ducts in BDL rats retain immunohistochemical,ltrastructural, and functional characteristics of normalholangiocytes and appear, morphologically, as elonga-ion of pre-existing ducts37,38,43,44; (2) thymidine labelingtudies were consistent with proliferation of pre-existingucts45,46; and (3) in humans with extrahepatic biliarybstruction of recent onset, proliferating parenchymalells and cholangiocytes express their respective normalytokine profile.47 Recently, a 3-dimensional reconstruc-ion of the biliary tree by microscopic computed tomog-aphy demonstrated that: (1) the volume of the biliaryree increases 20-fold after �-naphthylisothiocyanate ad-
inistration; and (2) cholangiocyte hyperplasia involvesprouting of new side branches rather than elongation ornlargement of pre-existing ducts.43,44 However, sometudies show that proliferating ductular cells express
arkers of progenitor cells (the full-length �-fetoproteinnd stem cell factor, c-kit) indicating that, in “typical”roliferation as well, a participation of a stem cell com-artment cannot be completely ruled out.48 In humans, atypical” proliferation is observed in acute obstructiveholestasis,32,35,37,47,49 extrahepatic biliary atresia,21 andhe early phases of chronic cholestatic liver diseases inssociation with “atypical proliferation.”1,3,32,35,37,50
Type II or “atypical” proliferation is characterized byn irregular proliferation of intrahepatic bile ducts notonfined to portal areas but sprouting into periportalnd parenchymal regions (“biliary piecemeal necro-is”).1,32–35,37 The proliferating ductular structures are ar-anged in a 3-dimensional network of tortuous and irregu-ar conduits without a well-defined lumen and surroundedy edema and inflammatory infiltrate.1,3,33,37 This implieshat the new-formed bile ducts are functionally immaturend the lack of anatomic contact with the canalicular spacesnd PBP around them51–53 further supports this concept.ype II ductular proliferation occurs in rodents in associa-
ion with oval cell proliferation after chronic administrationf chemicals such as D-galactosamine54 or carbon tetrachlo-ide55,56 and in humans, in alcoholic liver diseases, long-tanding extrahepatic biliary obstruction, focal nodular hy-erplasia, and chronic cholestatic liver diseases [eg, primaryiliary cirrhosis (PBC), primary sclerosing cholangitisPSC)].1,21,33–37,57–59 Although evidence exists that type IIuctular proliferation may originate from ductular meta-lasia or transformation of hepatic liver cell cords (ie, ret-odifferentiation of hepatocytes into cholangiocytes), and
ot from replication of pre-existing cholangiocytes,3,59–62 fost studies support the concept that the “atypical”roliferation arise from both proliferation of pre-existinguctules and expansion of the HPC compartment.63– 65
Type III ductular proliferation consists in the massiveroliferation of ductular hepatocytes, or progenitor cells, in
iver with submassive hepatic necrosis.21,32 Type IV ductularyperplasia (also called “oval cell” proliferation) occurs inhe early stages of carcinogenesis in rat liver and is caused by
number of chemicals including ethionine, 2-acetylamin-fluorene, and furan.32,66–70 This type of proliferation in-uces the formation of disorganized tubular structures withpoorly defined duct lumen, which randomly sprout intoepatic lobules creating a distorted hepatic architec-ure.32,66–70 Oval cells, so called for their shape, are related toerminal biliary ductules and canals of Hering, and repre-ent a subpopulation of nonparenchymal cells that areeterogeneous with regard to the expression of proteins andell surface markers.3,66–70 Oval cells express phenotypes ofmmature or neoplastic hepatocytes (ie, �-fetoprotein, albu-
in, and G-6-PO4) and of cholangiocytes (specific cytoker-tins),66–71 and may differentiate into hepatocytes, bile ductpithelial cells, enterocytes, and exocrine pancreaticells.66,70
From the findings described previously, it is evidenthat the classification of cholangiocyte proliferation is
ostly based on experimental studies. In human pathol-gy, the distinction between “typical” proliferation (orig-
nating from the sprouting of pre-existing ducts andlassically associated with acute bile obstruction) andatypical” ductular proliferation (closely related to pro-enitor cell activation and associated with vanishing syn-romes) is problematic, and the use of these terms isurrently discouraged.3,34,72,73 In fact, in both conditions,epatocellular and cholangiocyte damage occurs, induc-
ng regeneration of progenitor cells, which differentiateoward the most damaged cell type. In addition, the termatypical” includes a concept of premalignancy in diag-ostic histopathology, and this is not the case in mostituations.3,34,72,73
The Significance of Ductal Reaction inLiver DiseasesIn the course of chronic cholestatic liver diseases,
holangiocyte proliferation aims to repair and compen-ate for the anatomic and functional loss of injureducts.1,37 In fact, proliferating cholangiocytes display en-anced secretory activities by which they compensate forhe impaired function of injured cells.1,19,27,38,39 In theseathologies, HPC activation contributes to ductal reac-ion, as described by immunohistochemical and ultra-tructural studies.63,74,75 In these studies, intermediateells between progenitor cells and cholangiocytes areuch more numerous than intermediate hepatocytes,
uggesting that in biliary diseases, HPC mainly differen-iate towards the biliary lineage.63,76,77 Independently
rom ductular reaction, during immuno-mediated
csedctcrdtPdastttrtteaptto
“dt
crevacpvrtdawsdome
otrrtt
hi(caFHlmto
lepigiacrtvlrtacaoiftio
mpPeNtt
418 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
holangiopathies such as PBC and PSC, cholangiocyteshow enhanced proliferative activities together with annhanced rate of apoptosis.50,78 – 81 The course of theseiseases is characterized by a balance between cholangio-yte death and compensatory cholangiocyte prolifera-ion.1,37 An unbalanced equilibrium between cholangio-yte death (apoptosis or necrosis) and the proliferation ofesidual cholangiocytes determines the evolution towarductopenia, which characterizes the final evolution ofhese pathologies.32,37,50 As specifically evaluated inBC,78 the early compensated (nonductopenic) stages ofisease are characterized by a similar rate of apoptosisnd proliferation, whereas in the terminal ductopenictages apoptosis predominates favoring the evolutionoward ductopenia. Therefore, in these pathologic condi-ions, where the mechanisms of injury cannot be removed,he mechanism of repair mainly driven by proliferation ofesidual cholangiocytes is not sufficient to block the evolu-ion toward ductopenia. Therefore, conceptually, stimula-ion of cholangiocyte proliferation could represent a strat-gy for the management of human ductopenic pathologiesffecting the intrahepatic biliary tree. On the contrary, sup-ression of proliferation should be the potential strategy forhe management of pathologies characterized by uncon-rolled proliferative conditions such as cholangiocarcinomar polycystic liver diseases.23,32,50
Other than in chronic cholestatic liver diseases,ductular reaction” is seen in many different chronic liveriseases, which are very often associated with inflamma-ion, fibrosis, and HPC proliferation.3,32,37,71
From animal models, we have learned that progenitorells are activated when injury is associated with impairedegeneration of the mature cell types involved.69,72,82 Ev-ry acute and chronic human liver disease caused byiruses, alcohol, toxic substances, and metabolic errors, isssociated with damage and loss of hepatocytes and/orholangiocytes.1,37 Hence, it is conceivable that HPC ex-ansion and ductular reaction have been described in aariety of human liver diseases,3,21,33,36,37,71–77,82 whereeactive ductules in continuity with intermediate hepa-ocytes are seen at the periphery of the necrotic areas. Theegree of progenitor cell activation, of ductular reaction,nd the number of intermediate hepatocytes correlatesith the degree of inflammatory activity81,83 and with the
tage of the disease,60,72,77 indicating that progenitor cellsifferentiate towards hepatocytes when a certain thresh-ld of damage is reached,81 and that the HPC compart-ent activation is linked with the evolution of the dis-
ase.The emerging hypothesis is that the replicative arrest
f hepatocytes, induced by viral infection or inflamma-ion or exacerbated by insulin resistance and steatosis,esults in HPC expansion, the latter resulting not only inegeneration of hepatocytes, but also in a ductular reac-ion that contributes to periportal fibrogenesis. Consis-
ently, in livers with cirrhosis, in nonalcoholic and alco- rolic fatty liver disease, and in other chronic liver diseasesncluding chronic HCV, the number of hepatitis C virusHCV) and the extent of the ductular reaction significantlyorrelated with the severity of fibrosis and steatosis as wells with the degree of replicative arrest in hepatocytes.64,65,75
rom these findings, it was suggested that the extent ofPC expansion and of ductal reaction conditions the evo-
ution of most chronic liver diseases.63–65,84 For this reason,odulation of ductal reaction and cholangiocyte prolifera-
ion could represent a strategy for controlling the evolutionf chronic liver diseases.
Intracellular Signaling RegulatingCholangiocyte ProliferationcAMP-Related PathwayscAMP levels are increased in cholangiocytes pro-
iferating after BDL,26 a hyperplastic model where alsoxtracellular regulated kinases (ERK) and p70S6K phos-horylation is enhanced.85 A wide set of subsequent stud-
es showed that most of the agents stimulating cholan-iocyte proliferation increase intracellular cAMP levelsncluding acetylcholine,30 forskolin,29,86 dobutamine,86
nd clenbuterol.86 On the other hand, inhibition ofholangiocyte proliferation induced, in normal or BDLats, by carbon tetrachloride,55,56 gastrin,27,28 somatosta-in,26 serotonin,87 6-hydroxydopamine (6-OHDA),86 andagotomy30 is associated with decreased cAMP intracel-ular levels. The mechanistic relevance of cAMP in theegulation of cholangiocyte growth is demonstrated byhe fact that the in vivo treatment of normal rats with thedenylate cyclase (AC) stimulator forskolin88 inducesholangiocyte proliferation in association with increasedctivity of protein kinase A (PKA) and phosphorylationf Src-Tyrosine 139 and ERK1/2.29 This signaling cascade
s also the pathway that allows estrogens,89 nerve growthactor (NGF),90 and insulin-like growth factor-1 (IGF-1)91
o trigger cholangiocyte proliferation, whereas serotonin-nduced inhibition of cell growth is due to the inhibitionf cAMP/PKA/Src/ERK1/2 cascade.87
Phosphatidylinositol 3-Kinase (PI3K)PathwayThe PI3K signaling plays a role not only in pro-
oting growth but also survival in all cell types.92 Thehosphorylation of AKT (an immediate downstream ofI3K) is increased when cholangiocyte proliferation islicited by biliary obstruction93 or upon treatment withGF,90 IGF-I,91 and bile salts.94 In keeping, inhibition of
he PI3K/AKT pathway by wortmannin results in inhibi-ion of cholangiocyte proliferation.91,95
Intracellular Ca2� and RelatedSignalingThe intracellular Ca2� signaling plays a pivotal
ole in mediating the modulatory effects of different

agtdos
coseawhmbpttPSsottcp�dl
vliAttist
bltaHtspc4c
cplccuatmacaadacHIdtcaiguottdscHmamaopchftcascntisuc
January 2007 PROLIFERATING CHOLANGIOCYTES 419
gents on cholangiocyte proliferation. First of all, cholan-iocytes express inositol 1,4,5-trisphosphate (IP3)-recep-ors but not ryanodine receptors.96,97 Very importantly,ysregulation of IP3-mediated Ca2� signaling with a lossf IP3-receptors in cholangiocytes has been recently de-cribed as a common final pathway for cholestasis.96
A number of different agents activate Ca2� signaling inholangiocytes, but the stimulatory or inhibitory effectsn proliferation and secretion depend on the down-tream intracellular pathways elicited. Acetylcholine, forxample, increases cholangiocyte proliferation via Ca2�
nd calcineurin-mediated positive modulation of ACith enhanced cAMP intracellular levels.30 Acetylcholine,owever, acts by protein kinase C (PKC)-independentechanisms. On the other hand, when PKC is recruited
y Ca2� signaling, the effects on cAMP and cholangiocyteroliferation depend on the PKC isoforms involved. Sero-onin,87 ursodeoxycholate, and tauroursodeoxycholate98 ac-ivate PKC�, whereas gastrin activates PKC�, PKC�I, andKC�II,27,28 and this results in inhibition of cAMP/PKA/rc/ERK1/2 signaling, cholangiocyte proliferation,27,28 andecretin-stimulates ductal secretion, a functional markerf cholangiocyte proliferation.1,19,26–28,30,38,55,56,99 Therefore,he activation of specific PKC isoforms by [Ca2�]i is impor-ant in determining the differential increase or decrease ofAMP levels and the proliferative response of the intrahe-atic biliary epithelium. In support of this concept, the1-adrenergic receptor agonist, phenylephrine, stimulateductal secretion by PKC�II-dependent activation of cAMP
evels.100
The differential effect of agonist-induced [Ca2�]i acti-ation on cAMP levels and, therefore cholangiocyte pro-iferation, may be also due to the activation of specific ACsoforms. It is established that the AC isoforms, AC1 andC8, are induced by Ca2�, whereas AC5 and AC6 comprise
he calcium-inhibited family of AC isoforms.101 Consis-ently, in cholangiocytes stimulation of AC1 and AC8 andnhibition of AC5 and AC6 resulted in enhanced secretin-timulated ductal secretion and cholangiocyte prolifera-ion.102
Heterogeneity of Proliferative Capacityof the Intrahepatic Biliary TreeCanals of HeringStudies on the canals of Hering, which are lined
y hepatocytes, cholangiocytes, and HPC,3,6 have beenimited by the technical difficulties in studying this por-ion of the biliary tree. Distinction between the prolifer-tion of HPC and cholangiocytes is often problematic.owever, studies have shown that the functional regula-
ion of the biliary tree by neural/neuroendocrine factorstarts at the levels of the canals of Hering. First of all,ositivity for the parathyroid hormone-related peptide,hromogranin A, neural adhesion molecule, neurothrophin/5, and the neurothrophin receptor B in cells lining the
anals of Hering has been demonstrated.34,103–107 As for hholangiocytes lining the interlobular bile ducts, the ex-ression of neural and neuroendocrine markers in cells
ining the canals of Hering is marked during pathologiconditions of activation.34,103–107 Recently, the M3 acetyl-holine receptor has been detected with a positive mod-latory effect exerted by acetylcholine on proliferation,nd this has been postulated to play a role in the activa-ion of HPC compartment during injury.108 In the nor-
al liver, hepatocytes that lack M3 receptors producecetyl cholinesterase, which destroys acetylcholine in theanal of Hering, thus blocking the proliferation of HPCnd cholangiocytes.108 This also occurs after partial hep-tectomy.108 In contrast, during conditions of hepatocyteamage, acetylcholine is free to induce ductular reactionnd HPC expansion.108 The modulatory effect by acetyl-holine on the proliferation of cells lining the canals ofering is absent in the denervated transplanted liver.108
n keeping, the number of HPC and “atypical” reactiveuctular cells in the transplanted liver injured by hepa-itis was significantly lower than in innervated matchedontrol livers.108 This clinical observation was confirmedt the experimental level where vagotomy induces anmpaired regeneration of HPC and ductal reaction inalactosamine damaged rat liver.108 It is obvious to spec-late that the rapid progression of viral, toxic, ischemic,r immuno-mediated injury of the transplanted liveroward terminal and/or ductopenic conditions is facili-ated by impaired regenerative and repair capacity of theenervated transplanted liver. In contrast with the para-ympathetic system, the adrenergic system seems toounterregulate the activation of cells lining the canal ofering. First of all, adrenergic inhibition mobilizes he-atopoietic precursors into the circulation, and it has
lso been shown to promote liver regeneration.109,110 Inice fed antioxidant-depleted diets to induce liver injury,
dministration of prazosin (an �-1 adrenoceptor antag-nist) or 6-OHDA (an agent that induces chemical sym-athectomy) increases the numbers of oval cells.111 In-reased oval cell accumulation was accompanied by lessepatic necrosis and steatosis, lower serum aminotrans-
erases, and greater liver and whole body weights.111 Nei-her prazosin nor 6-OHDA affected the expression ofytokines, growth factors, or growth factor receptors thatre known to regulate progenitor cells.111 Therefore,tress-related sympathetic activity modulates progenitorell accumulation in damaged livers and sympatheticervous system (SNS) blockade with �-adrenoceptor an-agonists enhances HPC accumulation. These findingsndicate that modulation of the parasympathetic andympathetic system could represent a strategy for mod-lation of liver damage through the regulation of cellompartments residing in the canals of Hering.112
Bile DuctulesMost of the studies related to biliary proliferation
ave been allowed by the recent technological advances

t(�naomtaatAalnnmdwssllscetatpFppccoch
ta�pculBlbomtta
lmtddmpcdcdelgPsdpompmfa
po“nrSaewrdsprnsm
wmvtaoaii
420 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
hat made it possible to isolate small cholangiocytesapproximately 8 �m in diameter and lining small ducts
15–20 �m)2,113,114 and intrahepatic bile duct units fromormal and proliferating rat livers.26,39,99,113,114 In humannd rodent liver, small cholangiocytes express high levelsf the antiapoptotic proteins, annexin V and bcl2, whichay explain why small cholangiocytes are more resistant
han large cholangiocytes to injury.115–117 Bile ductulesre frequent targets of damage during drug-induced hep-totoxicity, as they are exposed to the largest concentra-ion of drugs/metabolites excreted by hepatocytes.118,119
t the experimental level, bile ductules do not proliferatefter BDL,2 which is the classical model of massive pro-iferation of intrahepatic bile ducts, and, in addition, doot respond to secretin or somatostatin because they doot express these receptors.26,113,114 Only during experi-ental models leading to damage of larger (�15 �m
iameter)114 bile ducts (eg, by carbon tetrachloride)55,56
ith increased apoptosis and reduced proliferative andecretory activities of large ducts,55,56 bile ductules tran-iently compensate for the anatomical and functionaloss of large ducts, with the de novo expression of pro-iferative and secretory activities including response toecretin.2,26,55,56,120,121 Recent studies indicate that smallholangiocytes lining bile ductules express receptors forndothelin (ETA and ETB),122 endogenous opioid pep-ides,123 insulin, acetylcholine (M3), and �1-adrenergicgonists (Alpini, unpublished observations, 2005), buthe role of these agents in modulating cholangiocyteroliferation in bile ductules has not yet been defined.inally, small cholangiocytes exhibited the de novo ex-ression of the apical sodium-dependent bile acid trans-orter during chronic feeding with the bile salts, tauro-holate, and taurolithocholate, suggesting a role in theholehepatic recirculation of bile salts in conditions ofverload.120,121,124 Whether bile salts modulate smallholangiocyte proliferation as they do in large bile ductsas yet to be elucidated.
Bile DuctsThe bulk of information currently available on
he neurohormonal regulation of cholangiocyte prolifer-tion relates mainly to large interlobular ducts (20 –100m), which are located in the portal spaces and are therimary target of damage in the course of most humanholangiopathies.1,32,37 In the BDL rat, a model widelysed for evaluating cholangiocyte proliferation,1,26 –28,30,38
arge ducts undergo massive “typical” proliferation afterDL with increased secretory activities.26 The increased
uminal pressure has been considered for many years toe the primary trigger for cholangiocyte proliferationccurring after BDL or obstructive cholestasis in hu-ans.32,45,125 However, selective ligation of one lobe of
he liver induces also proliferation of cholangiocytes inhe nonligated lobe, suggesting that humoral factors play
key role in modulating cholangiocyte proliferation fol- aowing obstructive cholestasis.126 A new lesson on theechanisms regulating cholangiocyte proliferation in
he course of cholestasis originates from polycystic liverisease.127 Cholangiocytes lining interlobular bile ductsisplay primary cilia in their apical site and express aechanoreceptor, polycystin-1 (PC-1), a Ca2� channel,
olycystin-2 (PC-2) and the Ca2�-inhibitable adenylyl cy-lase isoform 6 (AC6).127 Perfusion of intrahepatic bileuct units at flow rates of sufficient magnitude to bendilia results in a 40% increase in [Ca2�]i and a 4-foldecrease of forskolin-stimulated intracellular cAMP lev-ls.127 The flow-induced changes in [Ca2�]i and cAMPevels are significantly reduced or abolished when cholan-iocyte cilia are removed by chloral hydrate or whenC-1, PC-2, and AC6 are individually down-regulated byiRNAs.127 A possible suggestion is that cilia movementetermines a threshold of [Ca2�]i where cholangiocyteroliferation is inhibited. When bile flow is stopped asccurs in the BDL or human cholestasis or when cilia areorphologically abnormal, such as occurs in dominant
olycystic kidney disease (ADPKD),128 the lack of ciliaovement induces a decrease of [Ca2�]i, which could
avor the activation of AC, the increase of cAMP levels,nd therefore, cholangiocyte proliferation.
Proliferating Cholangiocytes AcquirePhenotype of Neuroendocrine CellsIn Figure 3, we depict the peptides secreted by
roliferating cholangiocytes and their interactions withther cells. Proliferating cholangiocytes, especially in theatypical” proliferation, acquire phenotypical features ofeuroendocrine cells including (1) the expression of neu-oendocrine markers (chromogranin A, glycolipid A2-B4,-100 protein, and neural cell adhesion molecule) andcquisition of neuroendocrine granules;34 –36,71,105 (2) thexpression of parathyroid hormone-related peptide,hich is encoded by a growth factor regulated “early
esponse” gene and which is involved in the growth andifferentiation of cholangiocytes105; (3) increased expres-ion and secretion of serotonin,87 endogenous opioideptides,123 and neurotrophins and their correspondingeceptors87,90; and (4) enhanced response to hormones/europeptides such as secretin,19,26,38,99 gastrin,27,28 somato-tatin,26,129 acetylcholine,30,130 and adrenergic100 and dopa-
inergic131 agonists.Proliferating cholangiocytes communicate and interact
ith other cells in the liver due to the secretion ofediators and immune factors that stimulate and acti-
ate several cell subtypes.132,133 This represents the basiso explain why cholangiocyte proliferation is frequentlyssociated with portal inflammation and fibrosis.133 Firstf all, other than the target of damage, cholangiocytes arective players in immune processes by interacting withnflammatory cells in the liver.132,134 –140 Cholangiocytes,n fact, express adhesion molecules by which they localize
nd intensify the immune response.132,136 –138 Further-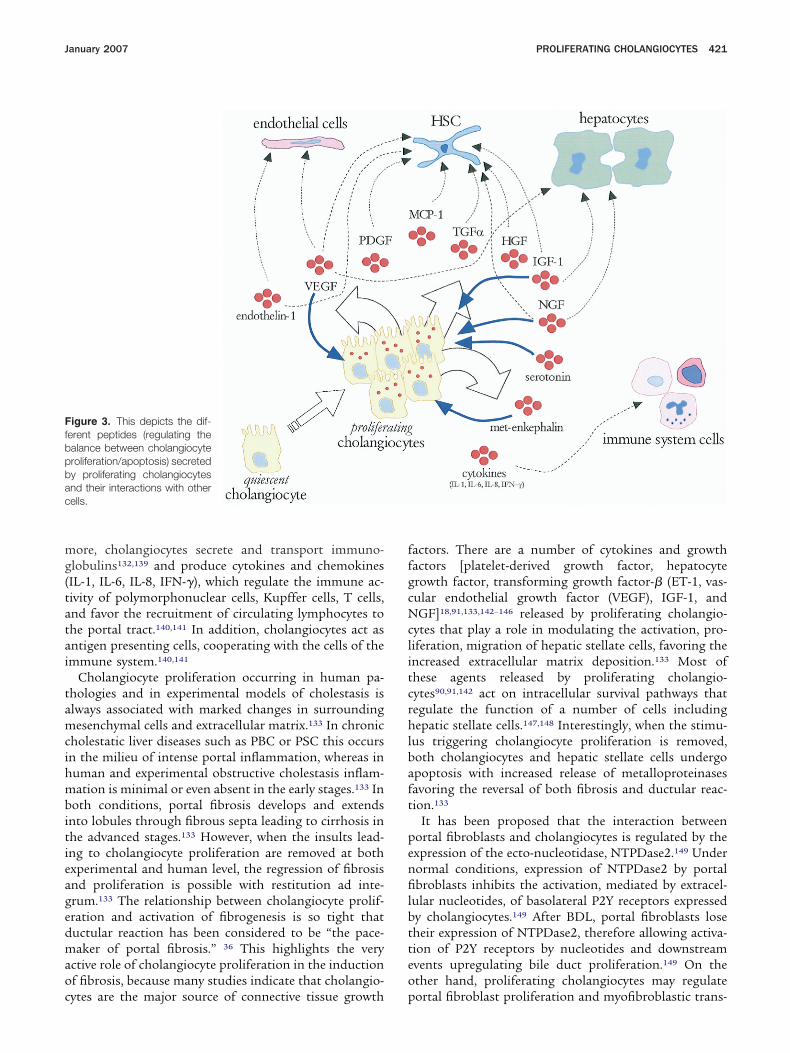
mg(tatai
tamcihmbitieagedmaoc
ffgcNclitcrhlbaft
penfilbtteo
Ffbpbac
January 2007 PROLIFERATING CHOLANGIOCYTES 421
ore, cholangiocytes secrete and transport immuno-lobulins132,139 and produce cytokines and chemokinesIL-1, IL-6, IL-8, IFN-�), which regulate the immune ac-ivity of polymorphonuclear cells, Kupffer cells, T cells,nd favor the recruitment of circulating lymphocytes tohe portal tract.140,141 In addition, cholangiocytes act asntigen presenting cells, cooperating with the cells of themmune system.140,141
Cholangiocyte proliferation occurring in human pa-hologies and in experimental models of cholestasis islways associated with marked changes in surroundingesenchymal cells and extracellular matrix.133 In chronic
holestatic liver diseases such as PBC or PSC this occursn the milieu of intense portal inflammation, whereas inuman and experimental obstructive cholestasis inflam-ation is minimal or even absent in the early stages.133 In
oth conditions, portal fibrosis develops and extendsnto lobules through fibrous septa leading to cirrhosis inhe advanced stages.133 However, when the insults lead-ng to cholangiocyte proliferation are removed at bothxperimental and human level, the regression of fibrosisnd proliferation is possible with restitution ad inte-rum.133 The relationship between cholangiocyte prolif-ration and activation of fibrogenesis is so tight thatuctular reaction has been considered to be “the pace-aker of portal fibrosis.” 36 This highlights the very
ctive role of cholangiocyte proliferation in the inductionf fibrosis, because many studies indicate that cholangio-
igure 3. This depicts the dif-erent peptides (regulating thealance between cholangiocyteroliferation/apoptosis) secretedy proliferating cholangiocytesnd their interactions with otherells.
ytes are the major source of connective tissue growth p
actors. There are a number of cytokines and growthactors [platelet-derived growth factor, hepatocyterowth factor, transforming growth factor-� (ET-1, vas-ular endothelial growth factor (VEGF), IGF-1, andGF]18,91,133,142–146 released by proliferating cholangio-
ytes that play a role in modulating the activation, pro-iferation, migration of hepatic stellate cells, favoring thencreased extracellular matrix deposition.133 Most ofhese agents released by proliferating cholangio-ytes90,91,142 act on intracellular survival pathways thategulate the function of a number of cells includingepatic stellate cells.147,148 Interestingly, when the stimu-
us triggering cholangiocyte proliferation is removed,oth cholangiocytes and hepatic stellate cells undergopoptosis with increased release of metalloproteinasesavoring the reversal of both fibrosis and ductular reac-ion.133
It has been proposed that the interaction betweenortal fibroblasts and cholangiocytes is regulated by thexpression of the ecto-nucleotidase, NTPDase2.149 Underormal conditions, expression of NTPDase2 by portalbroblasts inhibits the activation, mediated by extracel-
ular nucleotides, of basolateral P2Y receptors expressedy cholangiocytes.149 After BDL, portal fibroblasts loseheir expression of NTPDase2, therefore allowing activa-ion of P2Y receptors by nucleotides and downstreamvents upregulating bile duct proliferation.149 On thether hand, proliferating cholangiocytes may regulate
ortal fibroblast proliferation and myofibroblastic trans-
dc
gmwfdopfsmwbcfipcVatspmgEndgtchcfihvcosesciu
caaiTpgPfn
bPCcdpcIchpstsbctacc
thatpecopptdhwcttritdcscp3iwti
422 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
ifferentiation in a paracrine fashion via release of theytokine MCP-1.150
A recent emerging concept is that proliferating cholan-iocytes secrete vascular active substances,142 capable ofodulating the remodelling of bile duct vascular supplyith the aim to sustain the enhanced nutritional and
unctional demands of proliferating cells.17 This has beenemonstrated in the BDL model, where the proliferationf intrahepatic bile ducts is associated with an adaptiveroliferation of PBP.17 The adaptive proliferation of PBPollows, in order of time, the proliferation of bile ductsince after 1-week BDL, despite the enhanced bile duct
ass, there is no demonstrable proliferation of PBP,hich starts to be evident only after 2 weeks.10,144 Theasis for this cross talk between proliferating cholangio-ytes and endothelial cells has only recently been clari-ed.142 Proliferating cholangiocytes from BDL rats ex-ress and secrete VEGF-A and VEGF-C at greater levelsompared to normal quiescent cholangiocytes.142 TheEGF secreted by proliferating cholangiocytes enters, asn autocrine factor, in the complex loop of growth fac-ors, which sustain proliferation.142 Specifically, as de-cribed in many different cell types,151–153 VEGF couldlay a synergistic role with estrogens and IGF-1 on theodulation of cholangiocyte proliferation with conver-
ence on common signaling pathways, including theRK system. However, when VEGF is blocked by specificeutralizing antibodies, no proliferation of PBP occursespite BDL, suggesting that VEGF secreted by cholan-iocytes drives the proliferative adaptive response of PBPo cholestasis.142 Finally, VEGF secreted by proliferatingholangiocytes into the PBP and subsequently into theepatic sinusoids could play a role in protecting hepato-ytes from cholestatic injury, as supported by recentndings showing that VEGF plays a role in protectingepatocytes against different type of injuries.154 Anotherasoactive substance secreted by proliferating cholangio-ytes is endothelin, which is involved in the modulationf vascular bed functions.155 In support of this concept,tudies have shown that cholangiocytes secrete hepaticndothelin during the development of hepatopulmonaryyndrome after BDL, but not in thioacetamide-inducedirrhosis.145 Cholangiocyte-derived endothelin may be anmportant endocrine mediator of experimental hepatop-lmonary syndrome.145
The cross talk between hepatocytes and cholangiocytesould occur by substances released in bile by hepatocytesnd targeting the apical pole of cholangiocytes or bygents released in the PBP by cholangiocytes and target-ng hepatocytes after reaching the hepatic sinusoids.here is evidence that purines and insulin could be thelayers in this cross talk between hepatocytes and cholan-iocytes.156 Cholangiocytes express basolateral and apical2Y receptors, which could be activated in a paracrineashion by purines released in bile by hepatocytes or
earby located cholangiocytes or in an autocrine fashion fy purines released by the same cell.156 The activation of2Y receptors results in Ca2� signaling and activation ofa2�-dependent anion secretion, but whether cholangio-
yte proliferation is also influenced has not been eluci-ated yet. The cross talk between cholangiocytes andortal fibroblasts, mediated by purines and the P2Y re-eptors has been previously discussed.156 Insulin andGF-1, secreted in bile by hepatocytes,157,158 triggerholangiocytes because receptors for insulin and IGF-1ave been documented in the cholangiocyte apicalole.91,159 On the other hand, IGF-1, NGF, and VEGFecreted by proliferating cholangiocytes90,91,142 may re-urn back to hepatocytes via the PBP, favoring hepatocyteurvival during cholestasis.1,37 To this regard, VEGF haseen recently considered an important agent for hepato-yte protection during experimental injury.154 In addi-ion, the survival of hepatocyte preparations for the biolrtificial liver was markedly improved by coculture withholangiocytes, and thus further supports the role ofholangiocyte function for liver survival.160
Regulation of CholangiocyteProliferation by Hormones andNeuropeptidesFigure 4 illustrates the various agents involved in
he modulation of cholangiocyte proliferation. Differentormones modulate cholangiocyte proliferation: first ofll, estrogens, which have been considered for many yearso play a role in the development and progression ofathologies involving the biliary tree.161,162 This hypoth-sis was based on a number of clinical observations in-luding: (1) PBC, the most prevalent acquired cholangi-pathy,1,37 specifically affect females with a clinicalresentation typically occurring during the peri- andostmenopausal period163–167; (2) endocrine dysfunc-ions are frequent in PBC, including an increased inci-ence of menstrual disturbances and hysterectomy andigh incidence of osteoporosis in postmenopausalomen, which is a sign of estrogenic function deficien-
y168 and which is corrected by estrogen replacementherapy169; (3) chronic hepatic rejection leading to duc-openia more frequently interests male donor into femaleecipient170; (4) in Turner syndrome, bile duct pathologys frequent, and is improved by estrogen replacementreatment171; and (5) the progression of polycystic liverisease is significantly influenced by female sex, pregnan-ies, and estrogen replacement treatment.172 We havehown that rat cholangiocytes express both estrogen re-eptors (ER)-� and -� subtypes, and that cholangiocyteroliferation, after 3 weeks of BDL, is associated with a-fold increase in the expression of ER-� and a 30-fold
ncrease of ER-�.89,173–175 When BDL rats were treatedith tamoxifen or the pure ER antagonist, ICI 182,780,
he intrahepatic bile duct mass was markedly decreasedn comparison with control BDL rats.173 These data were
urther supported by the effects of ovariectomy that
nbctE
gdcpafsPpiintwlpspc(dtc
IcmipesisfiIpcPplcpcalcctsrta
Ftnttw
January 2007 PROLIFERATING CHOLANGIOCYTES 423
eutralized the induction of cholangiocyte proliferationy BDL.174 In vitro experiments in isolated cholangio-ytes demonstrated that 17�-estradiol-induced prolifera-ion was linked with activation of the ER/Src/Shc/RK1/2 pathway.89
In human studies, ER were not expressed by cholan-iocytes in the normal liver, but only in pathologic con-itions characterized by reactive or neoplastic cholangio-yte proliferation. Studies have been performed inatients with PBC and PSC,78 polycystic liver disease,128
nd cholangiocarcinoma.176 In PBC patients,78 positivityor ER-� was stably high (50 – 65%) in all histologictages. In contrast, positivity for ER-� increases fromBC stage I (1% positive cholangiocytes) to stage III (12%ositive cholangiocytes), correlates with and colocalizes
n the same cell with proliferating cell nuclear antigen,ndicating that those cells that undergo proliferation, butot quiescent cholangiocytes, express this receptor sub-ype.78 Consistently, in stage IV of PBC, cholangiocytesere negative for ER-� in association with impaired pro-
iferation and with maximal degree of ductopenia.78 Therogression of PBC toward the terminal ductopenictages is characterized by a different balance betweenroliferation and apoptosis.78 Although in the firstompensated stages I and II, cholangiocyte proliferationproliferating cell nuclear antigen) and death (terminaleoxynucleotidyl transferase–mediated deoxyuridineriphosphate nick-ending labeling) involve a similar per-
igure 4. Schematic drawing ofhe growth factors, gastrointesti-al hormones, and neuropeptideshat affect cholangiocyte prolifera-ion with decreases or increasesith biliary ductal mass.
entage of cholangiocytes, in the ductopenic PBC stages c
II and IV, proliferating cell nuclear antigen was signifi-antly lower than terminal deoxynucleotidyl transferase–ediated deoxyuridine triphosphate nick-ending label-
ng.78 This suggests that a relative deficiency ofroliferation with respect to apoptosis determines thevolution of the diseases toward the terminal ductopenictages. The association of this condition of relativelympaired proliferation with the disappearance of ER-�upports the hypothesis that a functional estrogenic de-ciency plays a role in the evolution toward ductopenia.
n addition, the ER-� positivity in cholangiocytes of PBCatients was markedly lower than PSC and alcoholicirrhosis, suggesting that the female predominance ofBC and its “typical” presentation during the postmeno-ausal period could be related with a defect in the pro-
iferative response of cholangiocytes to estrogens.78 Inells expressing ER, several studies showed that estrogenslay a major role, in concert with growth factors andytokines, in promoting the resistance to apoptotic dam-ge, in modulating reparative processes and in control-ing inflammation.176,177 The modulation of cholangio-yte proliferation exerted by estrogens and their receptorsould open new therapeutic perspectives stimulatingherapeutic trials aimed to evaluate the effects of ERelective modulators on PBC clinical expression. To thisegard, preliminary clinical observations indicate thatamoxifen (mixed agonist/antagonist for ER-� and onlyntagonist for ER-�) improves biochemical parameters of
holestasis in PBC patients.178
cthlcptmp1Aeswatbopt
atpucftltuOmscgpatIcts
ancpwirl
rn
eilhectttbpcmrc
ilbCtabociWslportsesmccEcpontft
tttgs
424 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
Also, for polycystic liver diseases there are a number oflinical observations suggesting that estrogens promotehe disease development and progression.128,179 –181 Weave recently shown that the biliary epithelium lining
arge hepatic cysts of patients with ADPKD lack primaryilia,128 suggesting that this is a primary event leading tohenotype of hyperproliferating and hypersecretory epi-helium. The ADPKD hepatic cyst epithelium displays
arked and diffuse immunohistochemical expressionattern for endoplasmic reticulum (ER), IGF-1, and IGF--R.128 IGF-1 was also found in the hepatic cyst fluid.128
lso, an immortalized cell line (LCDE) derived from thepithelium of hepatic cysts of patients with ADPKD,howed expression of both ER-� and ER-� subtypes andhen exposed to estrogens underwent proliferation withdditive effects exerted by estradiol and IGF-1.128 Consis-ently, specific antagonists of ER (ICI 182,780) or IGF1-Rlocking antibody (�IR3), inhibit the proliferative effectf 17�-estradiol and IGF-1, respectively,128 suggesting theotential benefit of a therapeutic strategy based on es-rogen and/or IGF-1 antagonism.
In comparison with benign cholangiocyte proliferationssociated with PBC, PSC, and alcoholic cirrhosis, neoplas-ic cholangiocyte proliferation showed a 4-fold higher ex-ression of ER-� and a similar expression of ER-�, as eval-ated in biopsies of patients with human intrahepaticholangiocarcinoma.78,182 This is consistent with many dif-erent studies showing an increased ER-�/ER-� ratio inumor versus normal tissues including ovary, prostate, co-on, and breast cancer.183–187 In these tissues, neoplasticransformation and progression have been associated withpregulation of ER-� and downregulation of ER-�.183–187
n the other hand, ER-� has been associated with a positiveodulation of cell proliferation whereas the opposite was
uggested for ER-�.183–187 Human intrahepatic cholangio-arcinoma also overexpress IGF-1 and its receptor. Estro-ens, IGF1 and their receptors, altogether, generate a com-lex loop favoring cell proliferation and resistance againstpoptosis. In this loop, the IGF1-R plays a central role sinceransfection of human cholangiocarcinoma cell lines withGF1-R antisense oligonucleotides induces impairment ofell proliferation.182 These findings suggest that modula-ion of ER and IGF1-R could represent a future therapeutictrategy for the management of cholangiocarcinoma.
Gastrin exerts trophic effects on different epitheliand induces the growth of different normal188,189 andeoplastic tissues190 expressing gastrin receptors. Ratholangiocytes and cholangiocarcinoma cell lines ex-ress CCK-B/gastrin receptors,27,28,191 and similar tohat was observed in pancreatic cell lines,192 gastrin
nhibits hyperplastic growth of cholangiocytes in BDLats and cholangiocarcinoma growth in 3 different cellines.27,28,191
SSTR2 somatostatin receptors have been identified inat and human cholangiocytes as well as cholangiocarci-
oma cell lines.26,129,193 Somatostatin inhibits the prolif- cration of large cholangiocytes in BDL rats by selectivelynteracting with SSTR2 receptors and decreasing intracel-ular cAMP levels.26 Somatostatin and its analogues in-ibit the growth of human cholangiocarcinoma cell linesxpressing SSTR2 receptor subtypes,193 thus encouraginglinical studies to assess their impact in the diagnosis andreatment of this malignancy. Recently, we have shownhat cholangiocytes express prolactin receptors,194 andhat prolactin increased rat cholangiocyte proliferationy activation of the IP3/Ca2�/PKC pathway.194 Finally,reliminary data from our group indicate that ratholangiocytes from BDL rats express the thyroid hor-one receptors, TR-�1 and TR-�1,195 and that the thy-
oid hormone inhibits cholangiocyte proliferation by in-reased IP3 levels and PKC� phosphorylation.195
NeuropeptidesThe general concept emerging from recent studies
s that cholangiocyte proliferation is positively modu-ated by parasympathetic and sympathetic innervationut negatively modulated by the serotoninergic system.holangiocytes express the M3 subtype receptor for ace-
ylcholine, which positively modulates the proliferativend secretory activities of cholangiocytes.30,130 In BDL,ut not normal rats, vagotomy induces the disappearancef M3 acetylcholine receptor, impairment of cholangio-yte proliferation, and activation of apoptotic cell deathn association with decreased cAMP intracellular levels.30
hen BDL � vagotomy rats were treated with for-kolin, an activator of adenylyl cyclase,88 cAMP intracel-ular levels were maintained and vagotomy failed to im-air cholangiocyte proliferation and did not induce ap-ptosis.30 Also, the sympathetic innervation is ofelevance for hepatic proliferation as first investigated inhe partial hepatectomy model.196 Recently, we demon-trated that proliferating cholangiocytes from BDL ratsxpress the �1 and �2 adrenergic receptors.86 Chemicalympathetic denervation of the liver by 6-OHDA treat-
ent increased cholangiocyte apoptosis and decreasedholangiocyte proliferation in association with decreasedAMP levels, and reduced phosphorylation of PKA,RK1/2, and AKT.86 Very importantly, 6-OHDA-inducedhanges in cholangiocyte proliferative and secretory ca-acity were prevented by the �1 adrenergic receptor ag-nist, dobutamine, and the �2 adrenergic receptor ago-ist clenbuterol (adrenergic agonists which both coupleo Gs-proteins to activate adenylyl cyclase)86,197 and byorskolin (an adenylyl cyclase activator)88 administra-ion.86
Cholangiocytes express the serotonin 1A and 1B recep-ors.87 Their activation markedly inhibits the growth andhe choleretic activity of the intrahepatic biliary tree inhe BDL rat.87 Serotonin inhibition of cholangiocyterowth is mediated by activation of the IP3/Ca2�/PKCignaling pathway and the consequent inhibition of
AMP/PKA/Src/ERK1/2.87 Cholangiocytes secrete seroto-
nldsc
tipettnrtmdfptfioh“l(hahicdirotfs
hccigdsptmotdtt
scaacddlcd
January 2007 PROLIFERATING CHOLANGIOCYTES 425
in, the blockage of which enhances cholangiocyte pro-iferation in the course of cholestasis.87 Therefore, theseata indicate the existence of an autocrine loop based onerotonin that limits the growth of the biliary tree in theourse of chronic cholestasis.87
All these studies highlight the role of neurotransmit-ers and neuropeptides in modulating the functions ofntrahepatic biliary tree. However, it appears that onlyroliferating but not quiescent cholangiocytes are influ-nced in their functions by neuropeptides and neuro-ransmitters. This is in keeping with what happens in theransplanted liver where the absolute denervation doesot compromise liver functions.198 In addition, althougheinnervation starts very soon after orthotropic liverransplantation in rodents, it is complete (after 3– 4
onths), only around the hilus,199,200 and therefore, aeficient innervation characterizes the transplanted liveror all its life. Although the normal function of trans-lanted liver is not significantly affected by denerva-ion,198 the repair capacity against injury could be inef-cient, and this should explain the very rapid progressionf recurrent or ex novo diseases. In keeping with this, itas been recently shown that the number of HPC andatypical” reactive ductular cells in human transplantivers (denervated) is lower than in matched controlsinnervated).71,82,108 Specifically, transplanted livers withepatitis show significantly less HPCs and “atypical” re-ctive ductular cells than innervated control livers withepatitis.71,108 In addition, a replicative senescence and
nefficient proliferative response to injury account for theharacteristic cytological alterations of cholangiocytesuring early chronic rejection.71,108 Therefore, an emerg-
ng hypothesis is that the proliferative and secretoryesponse of the biliary epithelium to viral, ischemic, toxic,r immunologic injury should be markedly impaired inhe transplanted liver by the lack of innervation, thusavoring the evolution to chronic damage and decompen-ation rather than to restitution at integrum.
Conclusion and Future PerspectivesIn conclusion, accumulating evidence indicates
ow, in the course of chronic liver diseases, HPC andholangiocytes are activated constituting a neuroendo-rine compartment regulated by many different agentsncluding hormones, neuropeptides, cytokines, androwth factors. This compartment produces and secretesifferent mediators that regulate not only its own expan-ion but also the survival, apoptosis, and function ofarenchymal and mesenchymal hepatic cells, as well ashe components of the immune systems. The develop-
ent and progression of liver damage and mechanismsf repair are all conditioned by this compartment, andhis occurs in the course of cholangiopathies, as well asuring viral and nonviral chronic liver diseases. In addi-ion, recent studies also suggest that clinical manifesta-
ions of chronic liver diseases including portal hyperten-ion and hepatopulmonary syndrome could beonditioned by soluble mediators generated from thectivated neuroendocrine compartments. The functionnd pathology of transplanted liver are also stronglyonditioned by the inefficient activation of this neuroen-ocrine compartment as a consequence of the completeenervation. Understanding how we can manage the pro-
iferation of HPC and cholangiocytes could represent ahallenge for the treatment of acute and chronic liveriseases.
References
1. Alpini G, Prall RT, LaRusso NF. The pathobiology of biliary epi-thelia. In: Arias IM, Boyer JL, Chisari FV, Fausto N, Jakoby W,Schachter D, Shafritz DA, eds. The liver: biology and pathobiol-ogy. 4th ed. Philadelphia, PA: Lippincott Williams & Wilkins;2001:421–435.
2. Kanno N, LeSage G, Glaser S, Alvaro D, Alpini G. Functionalheterogeneity of the intrahepatic biliary epithelium. Hepatology2000;31:555–561.
3. Roskams TA, Theise ND, Balabaud C, Bhagat G, Bhathal PS,Bioulac-Sage P, Brunt EM, Crawford JM, Crosby HA, Desmet V,Finegold MJ, Geller SA, Gouw AS, Hytiroglou P, Knisely AS, KojiroM, Lefkowitch JH, Nakanuma Y, Olynyk JK, Park YN, Portmann B,Saxena R, Scheuer PJ, Strain AJ, Thung SN, Wanless IR, WestAB. Nomenclature of the finer branches of the biliary tree: ca-nals, ductules, and ductular reactions in human livers. Hepatol-ogy 2004;39:1739–1745.
4. Hering E. Ueber den Bau der Wirbelthierleber. Arch Mikrosk Anat1867;3:88–114.
5. Theise ND, Saxena R, Portmann BC, Thung SN, Yee H, ChiribogaL, Kumar A, Crawford JM. The canals of Hering and hepatic stemcells in humans. Hepatology 1999;30:1425–1433.
6. Crawford JM. Normal and abnormal development of the biliarytree. In: Alpini G, Alvaro D, LeSage G, Marzioni M, LaRusso NF,eds. The pathophysiology of biliary epithelia. Georgetown, TX:Landes Biosciences; 2004:1–13.
7. Saxena R, Theise ND, Crawford JM. Microanatomy of the humanliver: exploring the hidden interfaces. Hepatology 1999;30:1339–1346.
8. Reilly FD, McCuskey PA, McCuskey RS. Intrahepatic distributionof nerves in the rat. Anat Rec 1978;191:55–68.
9. McCuskey RS. Anatomy of efferent hepatic nerves. Anat Rec ADiscov Mol Cell Evol Biol 2004;280:821–826.
10. Berthoud HR. Anatomy and function of sensory hepatic nerves.Anat Rec A Discov Mol Cell Evol Biol 2004;280:827–835.
11. el-Salhy M, Stenling R, Grimelius L. Peptidergic innervation andendocrine cells in the human liver. Scand J Gastroenterol 1993;28:809–815.
12. Goehler LE, Sternini C, Brecha NC. Calcitonin gene-related pep-tide immunoreactivity in the biliary pathway and liver of theguinea-pig: distribution and colocalization with substance P. CellTissue Res 1988;253:145–150.
13. Inoue N, Hirata K, Yamada M, Hamamori Y, Matsuda Y, Akita H,Yokoyama M. Lysophosphatidylcholine inhibits bradykinin-inducedphosphoinositide hydrolysis and calcium transients in culturedbovine aortic endothelial cells. Circ Res 1992;71:1410–1421.
14. Burt AD, Tiniakos D, MacSween RN, Griffiths MR, Wisse E, PolakJM. Localization of adrenergic and neuropeptide tyrosine-contain-ing nerves in the mammalian liver. Hepatology 1989;9:839–845.
15. Schultzberg M, Dalsgaard CJ. Enteric origin of bombesin immu-noreactive fibres in the rat coeliac-superior mesenteric ganglion.Brain Res 1983;269:190–195.
16. Barbaro B, Glaser S, Francis H, Taffetani S, Marzioni M, LeSage
G, Alpini G. Nerve regulation of cholangiocyte functions. In:
426 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
Alpini G, Alvaro D, LeSage G, Marzioni M, LaRusso NF, eds. Thepathophysiology of biliary epithelia. Georgetown, TX: LandesBiosciences; 2004:199–209.
17. Gaudio E, Onori P, Pannarale L, Alvaro D. Hepatic microcircula-tion and peribiliary plexus in experimental biliary cirrhosis: amorphological study. Gastroenterology 1996;111:1118–1824.
18. Gaudio E, Onori P, Franchitto A, Sferra R, Vetuschi A, Morini S,Alpini G, Alvaro D. Vascularization of the intrahepatic biliary treeand its role in the regulation of cholangiocyte growth. In: AlpiniG, Alvaro D, LeSage G, Marzioni M, LaRusso NF, eds. Thepathophysiology of biliary epithelia. Georgetown, TX: LandesBiosciences; 2004:41–45.
19. LeSage G, Glaser S, Gubba S, Robertson WE, Phinizy JL, LasaterJ, Rodgers RE, Alpini G. Regrowth of the rat biliary tree after 70%partial hepatectomy is coupled to increased secretin-inducedductal secretion. Gastroenterology 1996;111:1633–1644.
20. Schaffner F, Popper H. Electron microscopic studies of normaland proliferated bile ductules. Am J Pathol 1961;38:393–410.
21. Demetris AJ. Participation of cytokines and growth factors inbiliary cell proliferation and mito-inhibition during ductular reac-tion. In: Alpini G, Alvaro D, LeSage G, Marzioni M, LaRusso NF,eds. The pathophysiology of biliary epithelia. Georgetown, TX:Landes Biosciences; 2004:167–182.
22. Tadlock L, Yamagiwa Y, Hawker J, Marienfeld C, Patel T. Trans-forming growth factor-beta inhibition of proteasomal activity: apotential mechanism of growth arrest. Am J Physiol Cell Physiol2003;285:C277–C285.
23. Harnois DM, Que FG, Celli A, LaRusso NF, Gores GJ. Bcl-2 isoverexpressed and alters the threshold for apoptosis in a cho-langiocarcinoma cell line. Hepatology 1997;26:884–890.
24. Okaro AC, Deery AR, Hutchins RR, Davidson BR. The expressionof antiapoptotic proteins Bcl-2, Bcl-X(L), and Mcl-1 in benign,dysplastic, and malignant biliary epithelium. J Clin Pathol 2001;54:927–932.
25. Masyuk AI, Splinter PL, Masyuk TV, Stroope AJ, LaRusso NF.Cholangiocyte cilia act as sensory organelles affecting the in-tracellular signaling pathways. FASEB J 2005;19:A148.
26. Alpini G, Glaser S, Ueno Y, Pham L, Podila PV, Caligiuri A,LeSage G, LaRusso NF. Heterogeneity of the proliferative capac-ity of rat cholangiocytes after bile duct ligation. Am J PhysiolGastrointest Liver Physiol 1998;274:G767–G775.
27. Glaser S, Benedetti A, Marucci L, Alvaro D, Baiocchi L, Kanno N,Caligiuri A, Phinizy JL, Chowdhury U, Papa E, LeSage G, Alpini G.Gastrin inhibits cholangiocyte growth in bile duct-ligated rats byinteraction with cholecystokinin-B/Gastrin receptors via D-myo-inositol 1,4,5-triphosphate-, Ca(2�)-, and protein kinase C alpha-dependent mechanisms. Hepatology 2000;32:17–25.
28. Glaser S, Alvaro D, Ueno Y, Francis H, Marzioni M, Phinizy JL,Baumann B, Mancino MG, Venter J, LeSage G, Alpini G. Gastrinreverses established cholangiocyte proliferation and enhancedsecretin-stimulated ductal secretion of BDL rats by activation ofapoptosis through increased expression of Ca2�-dependentPKC isoforms. Liver Int 2003;23:78–88.
29. Francis H, Glaser S, Ueno Y, LeSage G, Marucci L, Benedetti A,Taffetani S, Marzioni M, Alvaro D, Venter J, Reichenbach R, FavaG, Phinizy JL, Alpini G. cAMP stimulates the secretory andproliferative capacity of the rat intrahepatic biliary epitheliumthrough changes in the PKA/Src/MEK/ERK1/2 pathway.J Hepatol 2004;41:528–537.
30. LeSage G, Alvaro D, Benedetti A, Glaser S, Marucci L, BaiocchiL, Eisel W, Caligiuri A, Phinizy JL, Rodgers R, Francis H, Alpini G.Cholinergic system modulates growth, apoptosis, and secretionof cholangiocytes from bile duct-ligated rats. Gastroenterology1999;117:191–199.
31. Sand TE, Thoresen GH, Refsnes M, Christoffersen T. Growth-
regulatory effects of glucagon, insulin, and epidermal growthfactor in cultured hepatocytes. Temporal aspects and evidencefor bidirectional control by cAMP. Dig Dis Sci 1992;37:84–92.
32. Alvaro D, Gigliozzi A, Attili AF. Regulation and deregulation ofcholangiocyte proliferation. J Hepatol 2000;33:333–340.
33. Desmet V, Roskams T, Van Eyken P. Pathology of the biliary treein cholestasis: ductular reaction. In: Manns MP, Boyer JL,Jansen PLM, Reichen J, eds. Cholestatic liver diseases. NewYork: Kluwer Academic Publishers; 1998:143–154.
34. Roskams T, Van den Oord JJ, De Vos R, Desmet VJ. Neuro-endocrine features of reactive bile ductules in cholestatic liverdisease. Am J Pathol 1990;137:1019–1025.
35. Desmet V, Roskams T, Van Eyken P. Ductular reaction in theliver. Pathol Res Pract 1995;191:513–524.
36. Popper H, Kent G, Stein R. Ductular reaction in the liver inhepatic injury. J Mt Sinai Hosp 1957;24:551–556.
37. Lazaridis KN, Strazzabosco M, LaRusso NF. The cholangiopa-thies: disorders of biliary epithelia. Gastroenterology 2004;127:1565–1577.
38. Alpini G, Lenzi R, Sarkozi L, Tavoloni N. Biliary physiology in ratswith bile ductular cell hyperplasia. Evidence for a secretoryfunction of proliferated bile ductules. J Clin Invest 1988;81:569–578.
39. LeSage G, Glaser S, Ueno Y, Alvaro D, Baiocchi L, Kanno N,Phinizy JL, Francis H, Alpini G. Regression of cholangiocyteproliferation after cessation of ANIT feeding is coupled withincreased apoptosis. Am J Physiol Gastrointest Liver Physiol2001;281:G182–G190.
40. Vacanti JP, Folkman J. Bile duct enlargement by infusion ofL-proline: potential significance in biliary atresia. J Pediatr Surg1979;14:814–848.
41. Alpini G, Glaser S, Ueno Y, Rodgers R, Phinizy JL, Francis H,Baiocchi L, Holcomb LA, Caligiuri A, LeSage G. Bile acid feedinginduces cholangiocyte proliferation and secretion: evidence forbile acid-regulated ductal secretion. Gastroenterology 1999;116:179–186.
42. Palmer RH, Ruban Z. Production of bile duct hyperplasia andgallstones by lithocholic acid. J Clin Invest 1966;45:1255–1267.
43. Masyuk TV, Masyuk AI, Ritman EL, LaRusso NF. Three-dimen-sional reconstruction of the rat intrahepatic biliary tree: physio-logical implications. In: Alpini G, Alvaro D, LeSage G, MarzioniM, LaRusso NF, eds. The pathophysiology of biliary epithelia.Georgetown, TX: Landes Biosciences; 2004:60–67.
44. Masyuk TV, Ritman EL, LaRusso NF. Quantitative assessmentof the rat intrahepatic biliary system by three-dimensional re-construction. Am J Pathol 2001;158:2079–2088.
45. Slott PA, Liu MH, Tavoloni N. Origin, pattern, and mechanism ofbile duct proliferation following biliary obstruction in the rat.Gastroenterology 1990;99:466–477.
46. Grisham JW. A morphologic study of deoxyribonucleic acid syn-thesis and cell proliferation in regenerating rat liver; autoradiog-raphy with thymidine-H3. Cancer Res 1962;22:842–849.
47. James J, Lygidakis NJ, van Eyken P, Tanka AK, Bosch KS,Ramaekers FC, Desmer V. Application of keratin immunocyto-chemistry and Sirius red staining in evaluating intrahepaticchanges with acute extrahepatic cholestasis due to hepaticduct carcinoma. Hepatogastroenterology 1989;36:151–155.
48. Omori M, Evarts RP, Omori N, Hu Z, Marsden ER, ThorgeirssonSS. Expression of alpha-fetoprotein and stem cell factor/c-kitsystem in bile duct ligated young rats. Hepatology 1997;25:1115–1122.
49. Miller DJ, Keeton DG, Webber BL, Pathol FF, Saunders SJ.Jaundice in severe bacterial infection. Gastroenterology 1976;71:94–97.
50. Celli A, Que FG. Dysregulation of apoptosis in the cholangiopa-thies and cholangiocarcinoma. Semin Liver Dis 1998;18:177–
185.
January 2007 PROLIFERATING CHOLANGIOCYTES 427
51. Desmet VJ. Current problems in diagnosis of biliary disease andcholestasis. Semin Liver Dis 1986;6:233–245.
52. Fausto N, Webber EM. Liver regeneration. In: Arias IM, Boyer JL,Fausto N, Jakoby WB, Schachter D, Shafritz DA, eds. The liver:biology and pathobiology. 3rd ed. New York: Raven, 1994:1059–1084.
53. Wegmann R, Corcos V, Caroli J. Histoenzymologie des ductulesbiliaires chez l’embryon humain normal et au tours des cirrho-ses humaines. Arch Ma1 Appar Dig 1967;54:215–228.
54. Kuhlmann WD, Wurster K. Correlation of histology and alpha1-fetoprotein resurgence in rat liver regeneration after experi-mental injury by galactosamine. Virchows Archiv A, Pathol AnatHistol 1980;387:47–57.
55. LeSage G, Benedetti A, Glaser S, Marucci L, Tretjak Z, CaligiuriA, Rodgers R, Phinizy JL, Baiocchi L, Francis H, Lasater J, UgiliL, Alpini G. Acute carbon tetrachloride feeding selectively dam-ages large, but not small, cholangiocytes from normal rat liver.Hepatology 1999;29:307–319.
56. LeSage G, Glaser S, Marucci L, Benedetti A, Phinizy JL, RodgersR, Caligiuri A, Papa E, Tretjak Z, Jezequel AM, Holcomb LA, AlpiniG. Acute carbon tetrachloride feeding induces damage of largebut not small cholangiocytes from BDL rat liver. Am J PhysiolGastrointest Liver Physiol 1999;276:G1289–G1301.
57. Nagore N, Howe S, Boxer L, Scheuer PJ. Liver cell rosettes: structuraldifferences in cholestasis and hepatitis. Liver 1989;9:43–51.
58. Knowles DM, Wolff M. Focal nodular hyperplasia of the liver: aclinicopathologic study and review of the literature. Hum Pathol1976;7:533–545.
59. Yamada S, Howe S, Scheuer PJ. Three-dimensional reconstruc-tion of biliary pathways in primary biliary cirrhosis: a computer-assisted study. J Pathol 1987;152:317–323.
60. Crosby HA, Hubscher S, Fabris L, Joplin R, Sell S, Kelly D, StrainAJ. Immunolocalization of putative human liver progenitor cellsin livers from patients with end-stage primary biliary cirrhosisand sclerosing cholangitis using the monoclonal antibody OV-6.Am J Pathol 1998;152:771–779.
61. De Vos R, Desmet V. Ultrastructural characteristics of novelepithelial cell types identified in human pathologic liver speci-mens with chronic ductular reaction. Am J Pathol 1992;140:1441–1450.
62. Tan J, Hytiroglou P, Wieczorek R, Park YN, Thung SN, Arias B,Theise ND. Immunohistochemical evidence for hepatic progen-itor cells in liver diseases. Liver 2002;22:365–373.
63. Roskams T. Progenitor cell involvement in cirrhotic human liver dis-eases: from controversy to consensus. J Hepatol 2003;39:431–434.
64. Lunz JG, 3rd, Tsuji H, Nozaki I, Murase N, Demetris AJ. Aninhibitor of cyclin-dependent kinase, stress-induced p21Waf-1/Cip-1, mediates hepatocyte mito-inhibition during the evolutionof cirrhosis. Hepatology 2005;41:1262–1271.
65. Clouston AD, Powell EE, Walsh MJ, Richardson MM, DemetrisAJ, Jonsson JR. Fibrosis correlates with a ductular reaction inhepatitis C: roles of impaired replication, progenitor cells andsteatosis. Hepatology 2005;41:809–818.
66. Ren P, Silberg DG, Sirica AE. Expression of an intestine-specifictranscription factor (CDX1) in intestinal metaplasia and in sub-sequently developed intestinal type of cholangiocarcinoma inrat liver. Am J Pathol 2000;156:621–627.
67. Lowes KN, Brennan BA, Yeoh GC, Olynyk JK. Oval cell numbersin human chronic liver diseases are directly related to diseaseseverity. Am J Pathol 1999;154:537–541.
68. Petropoulos CJ, Yaswen P, Panzica M, Fausto N. Cell lineages inliver carcinogenesis: possible clues from studies of the distri-bution of alpha-fetoprotein RNA sequences in cell populationsisolated from normal, regenerating, and preneoplastic rat livers.Cancer Res 1985;45:5762–5768.
69. Sirica AE, Cihla HP. Isolation and partial characterizations of
oval and hyperplastic bile ductular cell-enriched populationsfrom the livers of carcinogen and noncarcinogen-treated rats.Cancer Res 1984;44:3454–3466.
70. Farber E. Similarities in the sequence of early histological changesinduced in the liver of the rat by ethionine, 2-acetylamino-fluorene,and 3’-methyl-4-dimethylaminoazobenzene. Cancer Res 1956;16:142–148.
71. Roskams T, Cassiman D, De Vos R, Libbrecht L. Neuroregula-tion of the neuroendocrine compartment of the liver. Anat Rec ADiscov Mol Cell Evol Biol 2004;280:910–923.
72. Yang S, Koteish A, Lin H, Huang J, Roskams T, Dawson V, DiehlAM. Oval cells compensate for damage and replicative senes-cence of mature hepatocytes in mice with fatty liver disease.Hepatology 2004;39:403–411.
73. Roskams T, Desmet V. Ductular reaction and its diagnosticsignificance. Semin Diagn Pathol 1998;15:259–269.
74. Haque S, Haruna Y, Saito K, Nalesnik MA, Atillasoy E, Thung SN,Gerber MA. Identification of bipotential progenitor cells in hu-man liver regeneration. Lab Invest 1996;75:699–705.
75. Roskams TA, Libbrecht L, Desmet VJ. Progenitor cells in dis-eased human liver. Semin Liver Dis 2003;23:385–396.
76. Ros JE, Roskams TA, Geuken M, Havinga R, Splinter PL, Pe-tersen BE, LaRusso NF, van der Kolk DM, Kuipers F, Faber KN,Muller M, Jansen PL. ATP binding cassette transporter geneexpression in rat liver progenitor cells. Gut 2003;52:1060–1067.
77. Roskams T, De Vos R, Van Eyken P, Myazaki H, Van Damme B,Desmet V. Hepatic OV-6 expression in human liver disease andrat experiments: evidence for hepatic progenitor cells in man.J Hepatol 1998;29:455–463.
78. Alvaro D, Invernizzi P, Onori P, Franchitto A, De Santis A, Cro-signani A, Sferra R, Ginanni-Corradini S, Mancino MG, MaggioniM, Attili AF, Podda M, Gaudio E. Estrogen receptors in cholan-giocytes and the progression of primary biliary cirrhosis. J Hepa-tol 2004;41:905–912.
79. Harada K, Furubo S, Ozaki S, Hiramatsu K, Sudo Y, NakanumaY. Increased expression of WAF1 in intrahepatic bile ducts inprimary biliary cirrhosis relates to apoptosis. J Hepatol 2001;34:500–506.
80. Tinmouth J, Lee M, Wanless IR, Tsui FW, Inman R, HeathcoteEJ. Apoptosis of biliary epithelial cells in primary biliary cirrhosisand primary sclerosing cholangitis. Liver 2002;22:228–234.
81. Libbrecht L, Desmet V, Van Damme B, Roskams T. Deep in-tralobular extension of human hepatic “progenitor cells” corre-lates with parenchymal inflammation in chronic viral hepatitis:can “progenitor cells” migrate? J Pathol 2000;192:373–378.
82. Roskams T, Yang SQ, Koteish A, Durnez A, DeVos R, Huang X,Achten R, Verslype C, Diehl AM. Oxidative stress and oval cellaccumulation in mice and humans with alcoholic and nonalco-holic fatty liver disease. Am J Pathol 2003;163:1301–1311.
83. Xiao JC, Ruck P, Adam A, Wang TX, Kaiserling E. Small epithelialcells in human liver cirrhosis exhibit features of hepatic stem-like cells: immunohistochemical, electron microscopic and im-munoelectron microscopic findings. Histopathology 2003;42:141–149.
84. Demetris AJ, Seaberg EC, Wennerberg A, Ionellie J, Michalopou-los G. Ductular reaction after submassive necrosis in humans.Special emphasis on analysis of ductular hepatocytes. Am JPathol 1996;149:439–448.
85. Svegliati-Baroni G, Ridolfi F, Caradonna Z, Alvaro D, Marzioni M,Saccomanno S, Candelaresi C, Trozzi L, Macarri G, Benedetti A,Folli F. Regulation of ERK/JNK/p70S6K in two rat models ofliver injury and fibrosis. J Hepatol 2003;39:528–537.
86. Glaser S, Alvaro D, Francis H, Ueno Y, Marucci L, Benedetti A,De Morrow S, Marzioni M, Mancino MG, Phinizy JL, ReichenbachR, Fava G, Summers R, Venter J, Alpini G. Adrenergic receptor
agonists prevent bile duct injury induced by adrenergic denerva-
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
428 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
tion by increased cAMP levels and activation of Akt. Am J PhysiolGastrointest Liver Physiol 2006;290:G813–G826.
87. Marzioni M, Glaser S, Francis H, Marucci L, Benedetti A, AlvaroD, Taffetani S, Ueno Y, Roskams T, Phinizy JL, Venter J, Fava G,LeSage G, Alpini G. Autocrine/paracrine regulation of the growthof the biliary tree by the neuroendocrine hormone serotonin.Gastroenterology 2005;128:121–137.
88. Komalavilas P, Lincoln TM. Phosphorylation of the inositol1,4,5-trisphosphate receptor. Cyclic GMP-dependent protein ki-nase mediates cAMP and cGMP dependent phosphorylation inthe intact rat aorta. J Biol Chem 1996;271:21933–21938.
89. Alvaro D, Onori P, Metalli VD, Svegliati-Baroni G, Folli F, Fran-chitto A, Alpini G, Mancino MG, Attili AF, Gaudio E. Intracellularpathways mediating estrogen-induced cholangiocyte prolifera-tion in the rat. Hepatology 2002;36:297–304.
90. Gigliozzi A, Alpini G, Baroni GS, Marucci L, Metalli VD, Glaser S,Francis H, Mancino MG, Ueno Y, Barbaro B, Benedetti A, AttiliAF, Alvaro D. Nerve growth factor modulates the proliferativecapacity of the intrahepatic biliary epithelium in experimentalcholestasis. Gastroenterology 2004;127:1198–1209.
91. Alvaro D, Metalli VD, Alpini G, Onori P, Franchitto A, Barbaro B,Glaser S, Francis H, Cantafora A, Blotta I, Attili AF, Gaudio E. Theintrahepatic biliary epithelium is a target of the growth hor-mone/insulin-like growth factor 1 axis. J Hepatol 2005;43:875–883.
92. Cantley LC. The phosphoinositide 3-kinase pathway. Science2002;296:1655–1657.
93. Marzioni M, Francis H, Benedetti A, Ueno Y, Fava G, Venter J,Reichenbach R, Mancino MG, Summers R, Alpini G, Glaser S.Ca2�-dependent cytoprotective effects of ursodeoxycholic andtauroursodeoxycholic acid on the biliary epithelium in a ratmodel of cholestasis and loss of bile ducts. Am J Pathol 2006;168:398–409.
94. Alpini G, Glaser S, Francis H, Marzioni M, Venter J, Phinizy J,LeSage G. Bile acid interaction with cholangiocytes. In: Alpini G,Alvaro D, LeSage G, Marzioni M, LaRusso NF, eds. The patho-physiology of biliary epithelia. Georgetown, TX: Landes Bio-sciences; 2004:112–122.
95. Alpini G, Glaser S, Alvaro D, Ueno Y, Marzioni M, Francis H,Baiocchi L, Stati T, Barbaro B, Phinizy JL, Mauldin J, LeSage G.Bile acid depletion and repletion regulate cholangiocyte growthand secretion by a phosphatidylinositol 3-kinase-dependentpathway in rats. Gastroenterology 2002;123:1226–1237.
96. Shibao K, Hirata K, Robert ME, Nathanson MH. Loss of inositol1,4,5-trisphosphate receptors from bile duct epithelia is a com-mon event in cholestasis. Gastroenterology 2003;125:1175–1187.
97. Hernandez E, Nathanson MH. Calcium signaling in cholangio-cytes. In: Alpini G, Alvaro D, LeSage G, Marzioni M, LaRusso NF,eds. The pathophysiology of biliary epithelia. Georgetown, TX:Landes Biosciences, 2004:105–111.
98. Alpini G, Baiocchi L, Glaser S, Ueno Y, Marzioni M, Francis H,Phinizy JL, Angelico M, LeSage G. Ursodeoxycholate and taur-oursodeoxycholate inhibit cholangiocyte growth and secretion ofBDL rats through activation of PKC alpha. Hepatology 2002;35:1041–1052.
99. Alpini G, Ulrich C, Roberts S, Phillips JO, Ueno Y, Podila PV,Colegio O, LeSage G, Miller LJ, LaRusso NF. Molecular andfunctional heterogeneity of cholangiocytes from rat liver afterbile duct ligation. Am J Physiol Gastrointest Liver Physiol 1997;272:G289–G297.
00. LeSage G, Alvaro D, Glaser S, Francis H, Marucci L, Roskams T,Phinizy JL, Marzioni M, Benedetti A, Taffetani S, Barbaro B, FavaG, Ueno Y, Alpini G. Alpha-1 adrenergic receptor agonists mod-ulate ductal secretion of BDL rats via Ca(2�)- and PKC-depen-
dent stimulation of cAMP. Hepatology 2004;40:1116–1127.01. Beazely MA, Watts VJ. Regulatory properties of adenylate cy-clases type 5 and 6: a progress report. Eur J Pharmacol 2006;535:1–12.
02. Melero S, Strazzabosco M, Spirli C, Fiorotto R, Glaser S, FrancisH, Alpini G. Expression and regulation of adenylyl cyclase iso-forms (ACs) in rat cholangiocytes; effects of lypopolysaccharide(LPS) and �-naphthylisothiocyanate (ANIT) treatment. Hepatol-ogy;38:A1103.
03. Roskams T, De Vos R, van den Oord JJ, Desmet V. Cells withneuroendocrine features in regenerating human liver. APMISSuppl 1991;23:32–39.
04. Roskams T, Moshage H, Depla E, Willems M, Desmet V, Yap P.Parathyroid hormone-related peptide is expressed and rapidlyinducible in human liver cell cultures that have a bile ductphenotype. J Hepatol 1995;23:160–165.
05. Roskams T, Campos RV, Drucker DJ, Desmet VJ. Reactivehuman bile ductules express parathyroid hormone-related pep-tide. Histopathology 1993;23:11–19.
06. Cassiman D, Denef C, Desmet VJ, Roskams T. Human and rathepatic stellate cells express neurotrophins and neurotrophinreceptors. Hepatology 2001;33:148–158.
07. Libbrecht L, Cassiman D, Desmet V, Roskams T. Expression ofneural cell adhesion molecule in human liver development andin congenital and acquired liver diseases. Histochem Cell Biol2001;116:233–239.
08. Cassiman D, Libbrecht L, Sinelli N, Desmet V, Denef C,Roskams T. The vagal nerve stimulates activation of the hepaticprogenitor cell compartment via muscarinic acetylcholine recep-tor type 3. Am J Pathol 2002;161:521–530.
09. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Li Z, TorbensonM, Thomas SA, Diehl AM. Norepinephrine induces hepatic fibro-genesis in leptin deficient ob/ob mice. Biochem Biophys ResCommun 2003;308:284–292.
10. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Torbenson M,Smedh U, Moran TH, Li Z, Huang J, Thomas SA, Diehl AM.Hepatic fibrogenesis requires sympathetic neurotransmitters.Gut 2004;53:438–445.
11. Oben JA, Roskams T, Yang S, Lin H, Sinelli N, Li Z, TorbensonM, Huang J, Guarino P, Kafrouni M, Diehl AM. Sympatheticnervous system inhibition increases hepatic progenitors andreduces liver injury. Hepatology 2003;38:664–673.
12. Saxena R, Theise N. Canals of Hering: recent insights andcurrent knowledge. Semin Liver Dis 2004;24:43–48.
13. Alpini G, Roberts S, Kuntz SM, Ueno Y, Gubba S, Podila PV,LeSage G, LaRusso NF. Morphological, molecular, and func-tional heterogeneity of cholangiocytes from normal rat liver.Gastroenterology 1996;110:1636–1643.
14. Alpini G, Glaser S, Robertson W, Rodgers RE, Phinizy JL, LasaterJ, LeSage G. Large but not small intrahepatic bile ducts areinvolved in secretin-regulated ductal bile secretion. Am J PhysiolGastrointest Liver Physiol 1997;272:G1064–G1074.
15. Celli A, Que FG, Gores GJ, LaRusso NF. Glutathione depletion isassociated with decreased Bcl-2 expression and increased ap-optosis in cholangiocytes. Am J Physiol Gastrointest LiverPhysiol 1998;275:G749–G757.
16. Charlotte F, L’Hermine A, Martin N, Geleyn Y, Nollet M, GaulardP, Zafrani ES. Immunohistochemical detection of bcl-2 proteinin normal and pathological human liver. Am J Pathol 1994;144:460–465.
17. Katayanagi K, Van de Water J, Kenny T, Nakanuma Y, Ansari AA,Coppel R, Gershwin ME. Generation of monoclonal antibodiesto murine bile duct epithelial cells: identification of annexin V asa new marker of small intrahepatic bile ducts. Hepatology1999;29:1019–1025.
18. Desmet VJ. Vanishing bile duct syndrome in drug-induced liver
disease. J Hepatol 1997;26:31–35.
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
January 2007 PROLIFERATING CHOLANGIOCYTES 429
19. Desmet VJ. Drug-induced liver disease: pathogenetic mecha-nisms and histopathological lesions. Eur J Med 1993;2:36–47.
20. LeSage G, Glaser S, Francis H, Phinizy JL, Alpini G. Functionalheterogeneity of the intrahepatic cholangiocytes. In: Bittar E,ed. Liver biology in health and disease. Principles of medicalbiology. Oxford: Elsevier, 2005:21–48.
21. Glaser S, Francis H, Marzioni M, Taffetani S, Phinizy JL, LeSageG, Alpini G. Functional heterogeneity of the intrahepatic biliaryepithelium. In: Alpini G, Alvaro D, LeSage G, Marzioni M, La-Russo NF, eds. The pathophysiology of biliary epithelia. George-town, TX: Landes Biosciences, 2004:245–254.
22. Caligiuri A, Glaser S, Rodgers RE, Phinizy JL, Robertson W, PapaE, Pinzani M, Alpini G. Endothelin-1 inhibits secretin-stimulatedductal secretion by interacting with ETA receptors on largecholangiocytes. Am J Physiol Gastrointest Liver Physiol 1998;275:G835–G846.
23. Marzioni M, Alpini G, Saccomanno S, De Minicis S, Glaser S,Francis H, Trozzi L, Venter J, Orlando F, Fava G, Candelaresi C,Macarri G, Benedetti A. Endogenous opioids modulate thegrowth of the biliary tree in the course of cholestasis. Gastro-enterology 2006;130:1831–1847.
24. Alpini G, Ueno Y, Glaser S, Marzioni M, Phinizy JL, Francis H,LeSage G. Bile acid feeding increased proliferative activity andapical bile acid transporter expression in both small and largerat cholangiocytes. Hepatology 2001;34:868–876.
25. LeSage G, Glaser S, Alpini G. Regulation of cholangiocyte pro-liferation. Liver 2001;21:73–80.
26. Polimeno L, Azzarone A, Zeng QH, Panella C, Subbotin V, Carr B,Bouzahzah B, Francavilla A, Starzl TE. Cell proliferation andoncogene expression after bile duct ligation in the rat: evidenceof a specific growth effect on bile duct cells. Hepatology 1995;21:1070–1078.
27. Masyuk AI, Masyuk TV, Splinter PL, Huang BQ, Stroope AJ,LaRusso NF. Cholangiocyte cilia detect changes in luminal fluidflow and transmit them into intracellular Ca2� and cAMP sig-naling. Gastroenterology 2006;131:911–920.
28. Alvaro D, Onori P, Torrice A, Cardinale V, Barbaro B, FranchittoA, Alpini G, Battisti G, Jefferson DM, Strazzabosco M, AngelicoM, Attili A, Francis H, Glaser S, Gaudio E. Estrogens and IGF1promote the proliferation of hepatic cyst epithelium in autoso-mal dominant polycystic kidney disease (ADPK). Hepatology2005;42:A15.
29. Tietz PS, Alpini G, Pham LD, LaRusso NF. Somatostatin inhibitssecretin-induced ductal hypercholeresis and exocytosis bycholangiocytes. Am J Physiol Gastrointest Liver Physiol 1995;269:G110–G118.
30. Alvaro D, Alpini G, Jezequel AM, Bassotti C, Francia C, Fraioli F, RomeoR, Marucci L, Le Sage G, Glaser S, Benedetti A. Role and mecha-nisms of action of acetylcholine in the regulation of rat cholangiocytesecretory functions. J Clin Invest 1997;100:1349–1362.
31. Glaser S, Alvaro D, Roskams T, Phinizy JL, Stoica G, Francis H,Ueno Y, Barbaro B, Marzioni M, Mauldin J, Rashid S, MancinoMG, LeSage G, Alpini G. Dopaminergic inhibition of secretin-stimulated choleresis by increased PKC-gamma expression anddecrease of PKA activity. Am J Physiol Gastrointest Liver Physiol2003;284:G683–G694.
32. Fava G, Glaser S, Francis H, Alpini G. The immunophysiology ofbiliary epithelium. Semin Liver Dis 2005;25:251–264.
33. Alpini G, Alvaro D, Marzioni M, LeSage G, LaRusso N, eds.Georgetown, TX: Landes Bioscience, 2004:210–218.
34. Reynoso-Paz S, Coppel RL, Mackay IR, Bass NM, Ansari AA,Gershwin ME. The immunobiology of bile and biliary epithelium.Hepatology 1999;30:351–357.
35. Hreha G, Jefferson DM, Yu CH, Grubman SA, Alsabeh R, Geller
SA, Vierling JM. Immortalized intrahepatic mouse biliary epithe-lial cells: immunologic characterization and immunogenicity.Hepatology 1999;30:358–371.
36. Wu CT, Davis PA, Luketic VA, Gershwin ME. A review of thephysiological and immunological functions of biliary epithelialcells: targets for primary biliary cirrhosis, primary sclerosingcholangitis and drug-induced ductopenias. Clin Dev Immunol2004;11:205–213.
37. Leon MP, Kirby JA, Gibbs P, Burt AD, Bassendine MF. Immuno-genicity of biliary epithelial cells: study of the expression of B7molecules. J Hepatol 1995;22:591–595.
38. Morita M, Watanabe Y, Akaike T. Inflammatory cytokines up-regulate intercellular adhesion molecule-1 expression on pri-mary cultured mouse hepatocytes and T-lymphocyte adhesion.Hepatology 1994;19:426–431.
39. Sugiura H, Nakanuma Y. Secretory component and immuno-globulins in the intrahepatic biliary tree and peribiliary gland innormal livers and hepatolithiasis. Gastroenterol Jpn 1989;24:308–314.
40. Ayres RC, Neuberger JM, Shaw J, Joplin R, Adams DH. Intercel-lular adhesion molecule-1 and MHC antigens on human intra-hepatic bile duct cells: effect of pro-inflammatory cytokines. Gut1993;34:1245–1249.
41. Cruickshank SM, Southgate J, Selby PJ, Trejdosiewicz LK. Ex-pression and cytokine regulation of immune recognition ele-ments by normal human biliary epithelial and established livercell lines in vitro. J Hepatol 1998;29:550–558.
42. Gaudio E, Barbaro B, Alvaro D, Glaser S, Francis H, Ueno Y,Meininger CJ, Franchitto A, Onori P, Marzioni M, Taffetani S,Fava G, Stoica G, De Morrow S, Venter J, Reichenbach R, DeMorrow S, Summers R, Alpini G. Vascular endothelial growthfactor stimulates rat cholangiocyte proliferation via an autocrinemechanism. Gastroenterology 2006;130:1270–1282.
43. Grappone C, Pinzani M, Parola M, Pellegrini G, Caligiuri A,DeFranco R, Marra F, Herbst H, Alpini G, Milani S. Expression ofplatelet-derived growth factor in newly formed cholangiocytesduring experimental biliary fibrosis in rats. J Hepatol 1999;31:100–109.
44. Cramer T, Schuppan D, Bauer M, Pfander D, Neuhaus P, HerbstH. Hepatocyte growth factor and c-Met expression in rat andhuman liver fibrosis. Liver Int 2004;24:335–344.
45. Luo B, Tang L, Wang Z, Zhang J, Ling Y, Feng W, Sun JZ,Stockard CR, Frost AR, Chen YF, Grizzle WE, Fallon MB. Cholan-giocyte endothelin 1 and transforming growth factor beta1 pro-duction in rat experimental hepatopulmonary syndrome. Gastro-enterology 2005;129:682–695.
46. Matsumoto K, Nakamura T. Hepatocyte growth factor: molecu-lar structure and implications for a central role in liver regener-ation. J Gastroenterol Hepatol 1991;6:509–519.
47. Sato M, Suzuki S, Senoo H. Hepatic stellate cells: uniquecharacteristics in cell biology and phenotype. Cell Struct Funct2003;28:105–112.
48. Elsharkawy AM, Oakley F, Mann DA. The role and regulation ofhepatic stellate cell apoptosis in reversal of liver fibrosis. Apo-ptosis 2005;10:927–939.
49. Jhandier MN, Kruglov EA, Lavoie EG, Sevigny J, Dranoff JA.Portal fibroblasts regulate the proliferation of bile duct epitheliavia expression of NTPDase2. J Biol Chem 2005;280:22986–22992.
50. Kruglov EA, Nathanson RA, Nguyen T, Dranoff JA. Secretion ofMCP-1/CCL2 by bile duct epithelia induces myofibroblastictransdifferentiation of portal fibroblasts. Am J Physiol Gastroin-test Liver Physiol 2006;290:G765–G771.
51. Hyder SM, Huang JC, Nawaz Z, Boettger-Tong H, Makela S,Chiappetta C, Stancel GM. Regulation of vascular endothelialgrowth factor expression by estrogens and progestins. Environ
Health Perspect 2000;108(suppl 5):785–790.
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
1
430 ALVARO ET AL GASTROENTEROLOGY Vol. 132, No. 1
52. Hyder SM, Stancel GM. Regulation of angiogenic growth factorsin the female reproductive tract by estrogens and progestins.Mol Endocrinol 1999;13:806–811.
53. Rosenthal R, Wohlleben H, Malek G, Schlichting L, Thieme H,Bowes Rickman C, Strauss O. Insulin-like growth factor-1 con-tributes to neovascularization in age-related macular degenera-tion. Biochem Biophys Res Commun 2004;323:1203–1208.
54. LeCouter J, Moritz DR, Li B, Phillips GL, Liang XH, Gerber HP,Hillan KJ, Ferrara N. Angiogenesis-independent endothelial pro-tection of liver: role of VEGFR-1. Science 2003;299:890–893.
55. Agapitov AV, Haynes WG. Role of endothelin in cardiovasculardisease. J Renin Angiotensin Aldosterone Syst 2002;3:1–15.
56. Schlenker T, Romac JM, Sharara AI, Roman RM, Kim SJ, La-Russo N, Liddle RA, Fitz JG. Regulation of biliary secretionthrough apical purinergic receptors in cultured rat cholangio-cytes. Am J Physiol Gastrointest Liver Physiol 1997;273:G1108–G1117.
57. Thissen JP, Pucilowska JB, Underwood LE. Differential regula-tion of insulin-like growth factor I (IGF-I) and IGF binding pro-tein-1 messenger ribonucleic acids by amino acid availabilityand growth hormone in rat hepatocyte primary culture. Endocri-nology 1994;134:1570–1576.
58. Thissen JP, Verniers J. Inhibition by interleukin-1 beta and tumornecrosis factor-alpha of the insulin-like growth factor I messen-ger ribonucleic acid response to growth hormone in rat hepato-cyte primary culture. Endocrinology 1997;138:1078–1084.
59. LeSage G, Marucci L, Alvaro D, Glaser S, Benedetti A, MarzioniM, Patel T, Francis H, Phinizy JL, Alpini G. Insulin inhibits secre-tin-induced ductal secretion by activation of PKC alpha andinhibition of PKA activity. Hepatology 2002;36:641–651.
60. Auth MK, Woitaschek D, Beste M, Schreiter T, Kim HS, Opper-man E, Joplin RE, Baumann U, Hilgard P, Nadalin S, Markus BH,Blaheta RA. Preservation of the synthetic and metabolic capac-ity of isolated human hepatocytes by co-culture with humanbiliary epithelial cells. Liver Transpl 2005;11:410–419.
61. Ahlqvist J. Can endocrine factors influence pathogenetic mech-anisms in chronic active hepatitis (CAH) and primary biliarycirrhosis (PBC)? A hypothesis. Hepatogastroenterology 1980;27:64–67.
62. Joplin R, Neuberger JM. Antigen expression in bile duct epitheliain cholestatic liver disease. In: Manns MO, Boyer JL, JansenPLM, Reichen J, eds. Cholestatic liver diseases. Falk sympo-sium 102. Dordrecht: Kluwer Academic, 1998:226–238.
63. Floreani A, Titta M, Plebani M, Faggian D, Chiaramonte M,Naccarato R. Sex hormone changes in post-menopausal womenwith primary biliary cirrhosis (PBC) and with cryptogenic chronicliver disease. Clin Exp Obstet Gynecol 1991;18:229–234.
64. Floreani A, Paternoster D, Mega A, Farinati F, Plebani M, BaldoV, Grella P. Sex hormone profile and endometrial cancer risk inprimary biliary cirrhosis: a case–control study. Eur J ObstetGynecol Reprod Biol 2002;103:154–157.
65. Stellon AJ, Williams R. Increased incidence of menstrual abnor-malities and hysterectomy preceding primary biliary cirrhosis. BrMed J (Clin Res Ed) 1986;293:297–298.
66. Wolke AM, Schaffner F, Kapelman B, Sacks HS. Malignancy inprimary biliary cirrhosis. High incidence of breast cancer inaffected women. Am J Med 1984;76:1075–1078.
67. Goudie BM, Burt AD, Boyle P, Macfarlane G, Birnie GG, Mills PR,Gillis CR, MacSween RN, Watkinson G. Breast cancer in womenwith primary biliary cirrhosis. Br Med J (Clin Res Ed) 1985;291:1597–1598.
68. Hodgson SF, Dickson ER, Wahner HW, Johnson KA, Mann KG,Riggs BL. Bone loss and reduced osteoblast function in primarybiliary cirrhosis. Ann Intern Med 1985;103:855–860.
69. Olsson R, Mattsson LA, Obrant K, Mellstrom D. Estrogen-pro-gestogen therapy for low bone mineral density in primary biliary
cirrhosis. Liver 1999;19:188–192.70. Wiesner RH, Batts KP, Krom RA. Evolving concepts in thediagnosis, pathogenesis, and treatment of chronic hepatic al-lograft rejection. Liver Transpl Surg 1999;5:388–400.
71. Elsheikh M, Hodgson HJ, Wass JA, Conway GS. Hormone re-placement therapy may improve hepatic function in women withTurner’s syndrome. Clin Endocrinol 2001;55:227–231.
72. Gabow PA, Johnson AM, Kaehny WD, Manco-Johnson ML, DuleyIT, Everson GT. Risk factors for the development of hepaticcysts in autosomal dominant polycystic kidney disease. Hepa-tology 1990;11:1033–1037.
73. Alvaro D, Alpini G, Onori P, Perego L, Svegliata Baroni G, Fran-chitto A, Baiocchi L, Glaser S, Le Sage G, Folli F, Gaudio E.Estrogens stimulate proliferation of intrahepatic biliary epithe-lium in rats. Gastroenterology 2000;119:1681–1691.
74. Alvaro D, Alpini G, Onori P, Franchitto A, Glaser S, Le Sage G,Gigliozzi A, Vetuschi A, Morini S, Attili AF, Gaudio E. Effect ofovariectomy on the proliferative capacity of intrahepatic ratcholangiocytes. Gastroenterology 2002;123:336–344.
75. Alvaro D, Alpini G, Onori P, Franchitto A, Glaser S, Le Sage G,Folli F, Attili AF, Gaudio E. Alfa and beta estrogen receptors andthe biliary tree. Mol Cell Endocrinol 2002;193:105–108.
76. Tiidus PM. Influence of estrogen on skeletal muscle damage,inflammation, and repair. Exerc Sport Sci Rev 2003;31:40–44.
77. Koh KK. Effects of estrogen on the vascular wall: vasomotorfunction and inflammation. Cardiovasc Res 2002;55:714–726.
78. Reddy A, Prince M, James OF, Jain S, Bassendine MF. Tamox-ifen: a novel treatment for primary biliary cirrhosis? Liver Int2004;24:194–197.
79. Pirenne J, Aerts R, Yoong K, Gunson B, Koshiba T, Fourneau I,Mayer D, Buckels J, Mirza D, Roskams T, Elias E, Nevens F,Fevery J, McMaster P. Liver transplantation for polycystic liverdisease. Liver Transpl 2001;7:238–245.
80. Ramos A, Torres VE, Holley KE, Offord KP, Rakela J, Ludwig J.The liver in autosomal dominant polycystic kidney disease.Implications for pathogenesis. Arch Pathol Lab Med 1990;114:180–184.
81. Sherstha R, McKinley C, Russ P, Scherzinger A, Bronner T,Showalter R, Everson GT. Postmenopausal estrogen therapyselectively stimulates hepatic enlargement in women with au-tosomal dominant polycystic kidney disease. Hepatology 1997;26:1282–1286.
82. Alvaro D, Barbaro B, Franchitto A, Onori P, Glaser S, Alpini G,Francis H, Marucci L, Sterpetti P, Ginanni-Corradini S, OnettiMuda, A, Dostal D, De Santis A, Attili AF, Benedetti A, Gaudio E.Estrogens and insulin-like growth factor 1 modulate neoplasticcell growth in human cholangiocarcinoma. Am J Pathol 2006;169:877–888.
83. Bardin A, Hoffmann P, Boulle N, Katsaros D, Vignon F, Pujol P,Lazennec G. Involvement of estrogen receptor beta in ovariancarcinogenesis. Cancer Res 2004;64:5861–5869.
84. Lau KM, LaSpina M, Long J, Ho SM. Expression of estrogenreceptor (ER)-alpha and ER-beta in normal and malignant pros-tatic epithelial cells: regulation by methylation and involvementin growth regulation. Cancer Res 2000;60:3175–3182.
85. Platet N, Cathiard AM, Gleizes M, Garcia M. Estrogens and theirreceptors in breast cancer progression: a dual role in cancerproliferation and invasion. Crit Rev Oncol Hematol 2004;51:55–67.
86. Bardin A, Boulle N, Lazennec G, Vignon F, Pujol P. Loss ofERbeta expression as a common step in estrogen-dependenttumor progression. Endocr Relat Cancer 2004;11:537–551.
87. Ikeda K, Inoue S. Estrogen receptors and their downstreamtargets in cancer. Arch Histol Cytol 2004;67:435–442.
88. Yassin RR, Clearfield HR, Little KM. Gastrin’s trophic effect inthe colon: identification of a signalling pathway mediated by
protein kinase C. Peptides 1993;14:1119–1124.
1
1
1
1
1
1
1
1
1
1
1
2
G5(
WRD
January 2007 PROLIFERATING CHOLANGIOCYTES 431
89. Quintero E, Ohning GV, Del Rivero M, Wong HC, Walsh JH, Guth PH.Gastrin mediates the increase in gastric cell growth in uremic rats.Am J Physiol Gastrointest Liver Physiol 1995;268:G586–G591.
90. Song DH, Rana B, Wolfe JR, Crimmins G, Choi C, Albanese C,Wang TC, Pestell RG, Wolfe MM. Gastrin-induced gastric ade-nocarcinoma growth is mediated through cyclin D1. Am J PhysiolGastrointest Liver Physiol 2003;285:G217–G222.
91. Kanno N, Glaser S, Chowdhury U, Phinizy JL, Baiocchi L,Francis H, LeSage G, Alpini G. Gastrin inhibits cholangiocar-cinoma growth through increased apoptosis by activation ofCa2�-dependent protein kinase C-alpha. J Hepatol 2001;34:284–291.
92. Detjen K, Fenrich MC, Logsdon CD. Transfected cholecystokininreceptors mediate growth inhibitory effects on human pancre-atic cancer cell lines. Gastroenterology 1997;112:952–959.
93. Tan CK, Podila PV, Taylor JE, Nagorney DM, Wiseman GA, GoresGJ, LaRusso NF. Human cholangiocarcinomas express soma-tostatin receptors and respond to somatostatin with growthinhibition. Gastroenterology 1995;108:1908–1916.
94. Taffetani S, Francis H, Glaser S, Phinizy JL, Marucci L, BenedettiA, Baumann B, Reichenbach R, Venter J, Alpini G. The pituitaryhormone, prolactin stimulates normal cholangiocyte prolifera-tion by a Ca2� and PKC mediated mechanism. Hepatology2003;38:A1096.
95. Fava G, Glaser S, Phinizy JL, Francis H, Marucci L, Benedetti A,Taffetani S, Venter J, Baumann B, Reichenbach R, Alpini G.Thyroid hormone inhibits cAMP dependent proliferation ofcholangiocytes from bile duct ligated rats by a IP /Ca2�/PKC-
3Dependent Mechanism. Hepatology 2003;38:A1097. 2
96. Kiba T, Tanaka K, Numata K, Hoshino M, Inoue S. Facilitation ofliver regeneration after partial hepatectomy by ventromedialhypothalamic lesions in rat. Pflugers Arch 1994;428:26–29.
97. Kilts JD, Gerhardt MA, Richardson MD, Sreeram G, MackensenGB, Grocott HP, White WD, Davis RD, Newman MF, Reves JG,Schwinn DA, Kwatra MM. Beta(2)-adrenergic and several otherG protein-coupled receptors in human atrial membranes acti-vate both G(s) and G(i). Circ Res 2000;87:705–709.
98. Colle I, Van Vlierberghe H, Troisi R, De Hemptinne B. Trans-planted liver: consequences of denervation for liver functions.Anat Rec A Discov Mol Cell Evol Biol 2004;280:924–931.
99. Boon AP, Hubscher SG, Lee JA, Hines JE, Burt AD. Hepaticreinnervation following orthotopic liver transplantation in man.J Pathol 1992;167:217–222.
00. Dhillon AP, Sankey EA, Wang JH, Wightman AK, Mathur S,Burroughs AK, Scheuer PJ. Immunohistochemical studies onthe innervation of human transplanted liver. J Pathol 1992;167:211–216.
Received May 25, 2006; Accepted July 12, 2006.Address requests for reprints to: Domenico Alvaro, MD, Division of
astorenterology, Department of Clinical Medicine, via R. Rossellini1, Rome 00137, Italy. e-mail: [email protected]; fax:39) 06 4453319.
Portion of this work was supported by a Grant Award from Scott &hite Hospital to Shannon Glaser and Heather Francis, by a VAesearch Scholar Award and a VA Merit Award, and the NIH GrantsK58411 and DK062975 (to G.A.), and by a Grant from MIUR (PRIN
005 #2005067975_002; to D.A.).



















