
Melatonin regulates glucocorticoid receptor: an answer to its antiapoptotic action in thymus
Upload
independentCategory
view
0download
0
BRAIN RESEARCH
E L S E V I E R Brain Research 679 (1995) 72-81
Research report
Glucocorticoid receptor mRNA in Alzheimer's diseased hippocampus
Danie l M. Wetze l a,* M. Churchil l B o h n b A n n Marie Kazee a,c Robert W. Hamil l a,b
a Departments of Neurology, University of Rochester Medical Center and Monroe Community Hospital, 435 East Henrietta Road, Rochester NY 14620 USA
b Neurobiology and Anatomy, University of Rochester Medical Center, Rochester NY 14620 USA c Pathology and Laboratory Medicine, University of Rochester Medical Center, Rochester NY 14620 USA
Accepted 31 January 1995
Abs~ad
Hypothalamic-pituitary-adrenal (HPA) axis dysfunction is a common finding in Alzheimer's dementia. Since there is loss of hippocampal corticosteroid receptors in animal models of aging, and since hippocampal cell loss occurs in Alzheimer's disease (AD), it has been suggested that a loss of hippocampal glucocorticoid receptors (GR) may underlie some aspects of HPA axis dysfunction in patients with AD. Levels of corticosteroid receptor protein are not reliably determined in postmortem human brain due to rapid lability. In contrast, levels of mRNA coding for GR are stable in postmortem tissue. We report here initial observations from in situ hybridization experiments which indicate that regional levels of glucocorticoid receptor mRNA in hippocampus, as determined by film autoradiography, are significantly higher in AD hippocampus than in controls. While neuronal levels of GR mRNA in AD, revealed by emulsion autoradiography, were equal in control and AD tissue. Taken together these results suggest that adrenal dysfunction in AD may relate to defects in receptor function rather than corticosteroid receptor loss in the hippocampus.
Keywords: Hypothalamic-pituitary-adrenal axis; In situ hybridization; Dementia; Corticosteroid receptor
1. Introduction
Human, primate and rodent studies suggest that the hippocampus is involved in feedback regulation of the hypothalamic-pituitary-adrenal (HPA) axis [17,34,52]. In patients with Cushings syndrome there is a negative correlation between hippocampal formation volume and plasma cortisol levels [41]. Recent evidence demonstrates that impaired performance on memory tasks, with presumed hippocampal involvement, is as- sociated with elevated basal cortisol levels in healthy aged humans [21]. Studies in Alzheimer's disease (AD) reveal an altered HPA axis in a subset of patients. Increased plasma and urinary cortisol levels and posi- tive dexamethasone suppression tests (i.e. failure of
* Corresponding author. University of Vermont College of Medicine, Department of Neurology, Given Building, Burlington, VT 05405-0068, USA. Fax: (1) (802) 656 5844. E-mail: [email protected]
0006-8993/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)00230-8
plasma cortisol levels to fall to below 50 nM) following dexamethasone administration occur in 35-65% of pa- tients with this neurodegenerative dementia [22,28,40]. Hippocampal atrophy as determined by computerized tomographic scans in living AD patients has been linked to an abnormal cortisol response [8]. It has been hy- pothesized that glucocorticoid receptor loss in AD hippocampus may be one mechanism through which HPA dysfunction occurs [31]. However, due to the lability of the receptor binding site, fundamental knowledge of G R protein distribution and levels in human brain are unavailable [12,13,33,35]. Recent ex- periments suggest that mRNA in postmortem human brain is stable [11], and studies in rodents indicate that GR mRNA is only minimally influenced by post- mortem delay [49]. The distribution of G R mRNA has been examined in postmortem human brain and is similar to that of GR protein in the primate brain [37,50]. Since it has not yet been possible to quantita- tively measure GR protein levels in postmortem hu- man brain, measurement of mRNA levels for this

D.M. Wetzel et al. ~Brain Research 679 (1995) 72-81 73
receptor is the only means presently available to exam- ine cellular events related to glucocorticoid receptors or even estimate receptor' protein.
Collectively, current evidence suggests that GR mRNA might be decreased in AD. The present studies utilizing in situ hybridization with human specific ribo- probes sought to confirm the hypothesis that GR mRNA levels are decreased in AD hippocampus. We found, however, that mRNA levels for GR were in- creased or equal in AD relative to controls.
2. Materials and methods
2.1. Patients
Postmortem human brains were obtained from 6 individuals with a diagnosis of Alzheimer's disease and 4 control subjects enrolled in the Rochester Alzheimer's Disease Protect (RADP), a longitudinal clinical and neuropathological study of aging and AD. AD patients met NINCDS-ADRDA criteria for proba- ble AD and DSM III criteria for primary degenerative dementia of the Alzheimer type. Control subjects were selected from cognitively intact elderly subjects without neurologic or psychiatric disorders who came to au- topsy and for whom neuropathological examination revealed no Alzheimer's changes or other confounding neuropathological processes. Acceptable cases were classified as normal or AD, with the severity of AD being further clinically categorized by the clinical de- mentia rating (CDR), min:i-mental state exam (MMSE) and functional performance scales. All retrospective classifications were on the basis of a review of clinical records, and interviews with patient's physician and family, and a neuropathological survey of the brain which adhered to the NIA age-adjusted consensus cri- teria for definition of AD [19].
2.2. Neuroanatomical nomenclature and tissue process- ing
In this study the hippocampal nomenclature of Lorento de Nb [20] as adopted by Amaral and Insausti [1] was used. At autopsy, the right hippocampus was dissected, sliced into ~ 1 cm thick slabs and subse- quently frozen on dry ice. Coronal cryostat sections were taken approximately at the level of the lateral geniculate nucleus.
spans nucleotides 2,374 to 2,846 and corresponds to 3' translated and non-translated regions of human GR cDNA. For RNA probe transcription, linearized plas- mid templates were incubated with the appropriate polymerase (i.e. SP6, T7) for production of single- stranded anti-sense or sense strand RNA probes fol- lowing the manufacturer's procedures (Promega, Madi- son, WI). Depending on the specific activity of the probe required, up to 250/~Ci of [35S]UTP (1,200-1,400 Ci/mmol, New England Nuclear) was evaporated to dryness in an eppendorf tube using a speedvac before additions of buffer, unlabeled nucleotides, plasmid DNA and polymerase. Incorporation was measured using DE81 filters. Specific activity was estimated from these values. To verify full length of transcripts, a sample was checked using denaturing polyacrylamide gel electrophoresis (4% poly~tcrylamide, 8 M Urea).
Cryostat sections (20 /z) were picked up onto 3- aminopropyltriethoxysilane (Sigma) treated slides [29]. Sections were stored at -80°C until use. For hy- bridization, slides were warmed to room temperature (~ 1 h) and sections were permanently affixed to the slides by brief (5 min) fixation with a phosphate buffered 4% formaldehyde. The in situ hybridization (ISH) procedure utilized was adapted from Van Eeke- len et al. [44] for GR mRNA. Hybridization buffer consisted of 50% deionized formamide, 0.3 M NaCl, 10 mM Tris-HCl, pH 8.0, 1 mM EDTA, 10% dextran sulfate, 1 x Denhardts solution, 100 mM Dq~r, 500 ~g /ml denatured yeast tRNA, 500/zg/ml poly A and
S-labeled sense or antisense RNA probe added to a final concentration of 0.2 x 106 cpm//~l. Hybridization was overnight at 55 ° C. Coverslips were removed in 4 x SSC with 20 mM DTI". Following four 5 min rinses in 4 x SSC (with 10 mM DTr in all except the last), slides were incubated in RNase (20/.~g/ml) in 500 mM NaCI and 1 x TE for 30 min at 37°C. Rinses of increasing stringency were as follows, (i) two 2 x SSC rinses for 5 min at RT, (ii) 1 X SSC for 10 min at RT, (iii) 0.5x SSC at RT for 10 min, 3) 0.1× SSC at hybridization temperature for 30 min, (iv) 0.1 x SSC at RT for 3 min followed by dehydration in graded alco- hols. Air dried slides were placed in a metal X-ray cassette and exposed to /3-max film (Amersham). Ini- tial exposures were for 48-200 h. Following any reex- posures, slides were subsequently processed for emul- sion autoradiography.
2.4. Image analysis of film autoradiograms
2.3. In situ hybridization
For studies of human GR mRNA, an EcoRI-XbaI fragment of human GR cDNA (pOB7) provided by R. Evans [15] was subcloned into the polycloning site of pGEM3Z (Promega Biotcch). This 472 bp fragment
Video images of autoradiograhic films were digi- tized and quantitatively analyzed with an Imaging Re- search Inc. system (St. Catharines, Ontario, Canada) using M1 software. Calibrated 14C plastic autoradio- graphic standards (American Radiolabeled Chemicals, St. Louis, MO) were incorporated in all experiments to

74 D.M. Wetzel et aL / Brain Research 679 (1995) 72-81
verify film response (linearity) and permit comparison between experiments. The use of plastic 14C standards for calibrating and quantifying 35S-probes has previ- ously been decribed [24].
attribute most grains to a given neuron. Since this was not possible in dentate gyrus, due to the high packing density of the neurons, this region was not analyzed with grain counting.
2.5. Emulsion autoradiography 2. 7. Estimates of neuron density
For an accurate determination of the cellular local- ization of mRNA, emulsion based autoradiography was used, combined with grain counting using light mi- croscopy. Emulsion dipped slides (NTB-2 diluted 1:1 with distilled H 2 0 ) were initially allowed to gel in a humidified atmosphere, dried in a desiccated atmo- sphere 2 h at room temperature, and stored desiccated at 4 ° C, in light tight boxes. Slides were all processed (emulsion dipped) in the same batch. Slides used for grain counting were exposed for 7-10 days, an expo- sure time below emulsion saturation, as determined by pilot experiments. Another set of slides were exposed for up to 21 days for photographic purposes. Emulsion coated slides were developed with Kodak D19 for 2 min at 65 ° C, rinsed 30 s in distilled H 2 0 , fixed twice for 6 min each in Kodak fixer (not rapid fix) and rinsed in distilled water for 30 min. Sections were counter- stained with Cresyl violet.
2.6. Grain counting of emulsion dipped slides
Grain counting was performed by an experimenter, blind to case type, using phase contrast optics at a magnification of 400 X. The phase contrast optics combined with a light green filter facilitated discrimi- nation of cell type and the boundaries of neurons. From each region (i.e. subiculum,CA1, CA2 hippocam- pus, etc.) grains were manually counted over 100 neu- rons. Pilot experiments had determined autoradiogram exposure times of 7-10 days resulted in labelling below emulsion saturation with background counts (i.e. grain counts over adjacent white matter) below 0.03 grains//x 2. Grains, which were over the cell body, were counted up to a distance of ~ 5 microns outside of the cell membrane. No labeling was apparent over glia, or other non-neuronal structures. Since the density of neurons in the regions in which grain counting was done was low ( < 250 neurons /mm2, see Fig. 6) and the average neuron diameter was relatively small ( < 16 /.~, calculated from data in Fig. 6), it was possible to
Although a complete analysis of neuron density was beyond the scope of this project, we obtained a rough estimate of neuron density from Cresyl violet stained cryostat sections adjacent to those used for in situ hybridization. Sections were briefly fixed (30 min) in phosphate buffered 4% formaldehyde (pH 7.4) before staining. Microscopic images from an Olympus BH-2 microscope were digitized with a high resolution Ko- dak Megaplus camera. The M2 software from Imaging Research Inc was used for interactive morphometric analysis and microscope stage control, the latter as- sured that microscopic fields did not overlap. An ex- perimenter, blind as to case type, selected all neurons within a field which were than automatically counted and morphometrically evaluated. The criteria for selec- tion as a neuron was neuronal morphology without a darkly stained nucleus. Glia and microglia were easily recognized cells with dark densely staining nuclei and of much smaller size. Within a given hippocampal region (e.g. CA1) 10 fields (340.3 microns X 273.8 mi- crons) were used to determine neuron number / f ie ld and neuron size (crossectional area of cell body as stained by Cresyl violet).
2.8. Statistical analysis
Regression analysis and analysis of variance with repeated measures (ANOVA) were as provided for in General Linear Models module of the statistical soft- ware package SYSTAT (ver. 5.2) (SYSTAT.InteUigent software, Evanston, I11.).
3. Results
3.1. Postmortem interval
The mean postmortem interval (time of death until tissue was frozen) for both AD and control are shown in Table 1. No significant difference between the two
Table 1 Subject profile a
Group Age (yr) Brain weight (g) Postmortem delay (h) Illness duration (yrs)
Alzheimer (n = 6) 80.1 5:4.3 949.2 + 72.8 10.5 5:1.5 12.0 5:1.1 Control (n = 4) 82.5 5:5.2 1345 +_ 89.2 b 13.8 _+ 1.8 -
a Values are mean + standard error of the mean. b P < 0.01.

D.M. Wetzel et al. ~Brain Research 679 (1995) 72-81 7 5
[.
i
",2, "d
<
~.~ ~
~J
e'~ N 0
0 ~
N ~. ~
t ~ t g ~ t t % ~ ' t t ~ t t %
t - . ,
. o ' o "6
• = .~
.., r - :
z ~

76 D.M. Wetzel et al. / Brain Research 679 (1995) 72-81
Fig. I. Levels of glucocorticoid receptor mRNA in human control (C) and Alzheimer's diseased (D) hippocampus, subiculum and parahippocam- pal regions as determined by in situ hybridization with human specific antisense 35S-riboprobes. Human specific probes were subcloned and labeled to high specific activity using SP6/T7 initiated transcription. After hybridization sections were stringently rinsed, dried and apposed to /3-max film (Amersham) for a 168 h exposure. Sense controls had negligible signal. Panels A and B are sections stained with cresyl violet (CV). Note the higher levels of GR mRNA in the Alzhemier's brain CA1, subiculum (S) and parasubiculum regions (pS). DG = dentate gyrus.
groups ( F = 2.03; df = 8; P > 0.1) was found. The range for AD and control subjects was 6 to 17.2 h.
3.2. Clinical profiles
The mean age at time of death for AD and control groups are shown in Table 1. There was no significant difference between the two groups (F = 0.07; df = 8; P > 0.1). The age range for the AD group was 62 to 90 years and for controls 76 to 87 years. All AD patients had severe, end-stage disease (CDR 4-5). A brief clini- cal profile of the patients studied is presented in Table 2. All control subjects had heart disease. Some early investigations suggest elevated plasma cortisol may be associated with early stages of atherosclerosis [42]. Cor- tisol levels at the time of death were not available for control or AD patients.
3.3. Glucocorticoid receptor mRNA
3.3.1. Film autoradiography Hybridization of human hippocampal sections with
35S-antisense riboprobes to GR resulted in specific labeling of neuronal areas within the hippocampus, dentate gyrus, and subiculum (Fig. 1). In both control and AD tissue, high levels of specific signal for G R
mRNA were apparent over regions of dentate gyrus; less label was detectable in CA2 and CA3. In the control and AD tissue, GR mRNA was detectable in CA1 (although at a lower level). Results from quantita- tive image analysis of films from in situ hybridization are presented in Fig. 2. ANOVA revealed that GR mRNA levels were significantly higher in all subregions of Ammon's horn and subiculum in AD tissue, but not
40
35
• 30
w a 26
E 2o E o~ I s
0 e- lO
o CA3 CA2 CA1 SUB
Bra in R e g i o n
Fig. 2. Quantitative image analysis of fl-max films of in situ hy- bridizations for GR mRNA with ass-human specific riboprobes. * * P < 0.001, * P < 0.05.
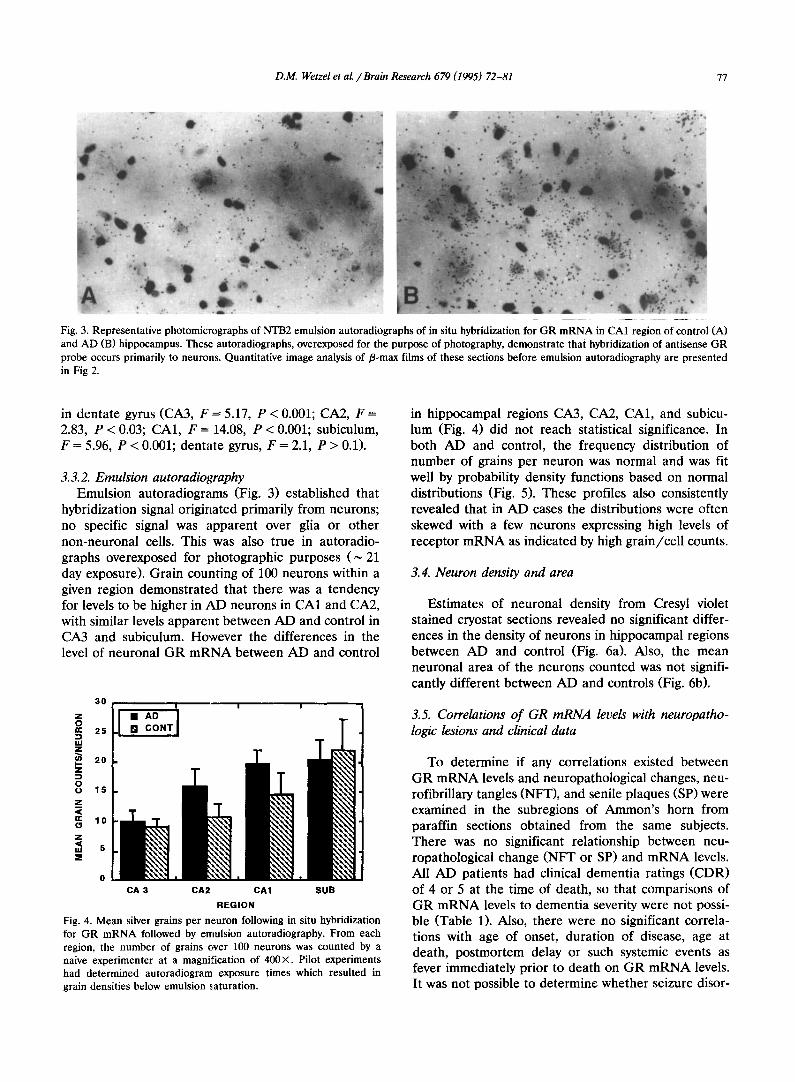
D.M. Wetzel et al. ~Brain Research 679 (1995) 72-81 77
Fig. 3. Representative photomicrographs of NTB2 emulsion autoradiographs of in situ hybridization for GR mRNA in CA1 region of control (A) and AD (B) hippocampus. These autoradiographs, overexposed for the purpose of photography, demonstrate that hybridization of antisense GR probe occurs primarily to neurons. Quantitative image analysis of/3-max films of these sections before emulsion autoradiography are presented in Fig 2.
in dentate gyrus (CA3, F = 5.17, P < 0.001; CA2, F = 2.83, P < 0.03; CA1, F = 14.08, P < 0.001; subiculum, F = 5.96, P < 0.001; dentate gyrus, F = 2.1, P > 0.1).
3.3.2. Emulsion autoradiography Emulsion autoradiograms (Fig. 3) established that
hybridization signal originated primarily from neurons; no specific signal was apparent over glia or other non-neuronal cells. This was also true in autoradio- graphs overexposed for photographic purposes ( ~ 21 day exposure). Grain counting of 100 neurons within a given region demonstrated that there was a tendency for levels to be higher in AD neurons in CA1 and CA2, with similar levels apparent between AD and control in CA3 and subiculum. However the differences in the level of neuronal GR mRNA between AD and control
30
Z O " 25 UJ z
20 I - z
O 1 5
z u < " 1 0
z < uJ 5 :E
0
CA 3 CA2 CA1 S U B
R E G I O N
Fig. 4. Mean silver grains per neuron following in situ hybridization for GR mRNA followed by emulsion autoradiography. From each region, the number of grains over 100 neurons was counted by a naive experimenter at a magnification of 400 ×. Pilot experiments had determined autoradiogram exposure times which resulted in grain densities below emulsion saturation.
in hippocampal regions CA3, CA2, CA1, and subicu- lum (Fig. 4) did not reach statistical significance. In both AD and control, the frequency distribution of number of grains per neuron was normal and was fit well by probability density functions based on normal distributions (Fig. 5). These profiles also consistently revealed that in AD cases the distributions were often skewed with a few neurons expressing high levels of receptor mRNA as indicated by high grain/cell counts.
3.4. Neuron density and area
Estimates of neuronal density from Cresyl violet stained cryostat sections revealed no significant differ- ences in the density of neurons in hippocampal regions between AD and control (Fig. 6a). Also, the mean neuronal area of the neurons counted was not signifi- cantly different between AD and controls (Fig. 6b).
3.5. Correlations of GR mRNA levels with neuropatho- logic lesions and clinical data
To determine if any correlations existed between GR mRNA levels and neuropathological changes, neu- rofibrillary tangles (NFT), and senile plaques (SP) were examined in the subregions of Ammon's horn from paraffin sections obtained from the same subjects. There was no significant relationship between neu- ropathological change (NFT or SP) and mRNA levels. All AD patients had clinical dementia ratings (CDR) of 4 or 5 at the time of death, so that comparisons of GR mRNA levels to dementia severity were not possi- ble (Table 1). Also, there were no significant correla- tions with age of onset, duration of disease, age at death, postmortem delay or such systemic events as fever immediately prior to death on GR mRNA levels. It was not possible to determine whether seizure disor-

78 D.M. Wetzel et al. / Brain Research 679 (1995) 72-81
03
n- < m 02 ¢ I,U o. Z O 0.1
¢ 0 n 0 ¢ o.
A
t '
I
0
I I
30 60
Grains per cell
. . . . . . . . . . . . . . . 25
- 20
15
I0
- 5
I
9O
03
rr < 0.2 co n- uJ o. z
h- 0.1 0 0 a .
B
- / / -
.4
I I I
30 60 90
Grains per cell
25
20 0
10
Fig. 5. Representative frequency histograms from grain counts of in situ hybridization for GR mRNA in control (A) and AD (B) CA1 hippocampus. Insets are probability density functions, the straight line reflects a normal distribution. Probability functions were well fit by normal distribution (R 2 > 97% for all AD and control cases) suggesting uniform expression of GR mRNA in neurons within a given region.
3 0 0
25O
E 200
e,, .O 150
:3 4)
Z lOO
50
250
Ca 2 0 0
<
O 150
Q)
Z 100 e-. (Q 4)
~ ; S0
C a 3 C a 2 C a 1 Sublculum C a 3 C a 2 C a 1 Subiculum
Brain Region Brain Region
Fig. 6. Estimates of neuronal density (A) and neuronal area (B) in control and AD hippocampus. Cresyl violet stained sections adjacent to those used for ISH were used for these analyses.
ders were correlated with GR levels due to the small sample size. It was striking, however, that one subject who was receiving the anticonvulsant Dilantin (phenytoin) for seizure disorders, had the lowest level of GR in all regions of any of the cases. Published reports suggest that the teratogenic effects of pheny- toin may be mediated through GR related mechanisms [18]. Since anticonvulsants are commonly perscribed for seizure disorders associated with severe dementia, it will be important to consider the possible effects of these drugs in the interpretation of future results.
4. Discussion
The current studies extend observations regarding alterations within the hippocampal-HPA axis in Alzheimer's disease by demonstrating the unexpected finding of higher (film autoradiography) or equal (em- ulsion autoradiography) regional levels of GR mRNA in AD hippocampus compared to normal controls.
While this study was being completed we became aware of another group [38,39], using similar, although not indentical cRNA probes and procedures, and subse- quent emulsion autoradiography. This group did not use film autoradiography to quantify changes in GR mRNA. Their grain counting was based on a smaller number of neurons per region (8-16 neurons) than the present study (100 neurons) and although in general they found no significant diffrence between AD and control cases, a carefully examination of their Fig. 1 shows the neuronal level of GR mRNA is significantly higher in the CA1 hippocampal region AD. However the point to be made here is that their observations, taken together with the present study, indicate that in contrast to what has been hypothesized regarding the level of mRNA for GR in AD hippocampus (i.e. that GR mRNA levels should be decreased), message level is at least preserved and indeed may be upregulated.
One methodological explanation for the present data might be that with the hippocampal atrophy associated with the neurodegenerative processes in AD increased

D.M. Wetzel et al. /Brain Research 679 (1995) 72-81 79
neuron cell density might occur, and result in a flawed interpretation of the regional levels on the autoradio- graphs; i.e., increased cell packing might falsely elevate regional levels. Accordingly we examined cell density, but found no significant differences between the den- sity of neurons in AD versus control hippocampus. This finding is different from previous studies which report decreased neuronal density within the hip- pocampus of AD brain: estimates of neuronal density suggest decreases ranging from 28 to 47% in the CA1 region [5,7]. However, a recent report using modern stereological techniques has found that differences in neuronal density in various hippocampal regions are small and highly variable between AD and control (West et al., 1993). Also, ,decreases in neuronal density with normal aging in these hippocampal regions are small, although this matter remains controversial [48].
Regional film analysis may also reflect hybridization signal arising not only from neurons, but from other GR mRNA containing c, ells such as glia, microglia, macrophages, and vascular endothelial ceils, however we think this possibility :is unlikely since the level of expression of GR mRNA in such cells is an order of magnitude less [3,46]. Additionally, the present emul- sion autoradiography demonstrated that hybridization signal originated from neurons; even with long expo- sure times (21 days) no signal was derived from non- neuronal cells.
Prevailing hyotheses suggest that GR mRNA levels might be markedly less in AD hippocampus [31]. An age related decrease of hippocampal corticosteroid re- ceptors, independent of neuron loss, has been observed repeatedly in rodent aging models, as well as a de- crease in glucocorticoid receptor mRNA [25,30,32,54]. However the observation,,; of Eldridge et al. [10] who found that aged rats did not down regulate GR as well as younger rats in response to repeated stress may also be relevant in the present context. Alzheimer's disease may vary from normal aging in that it might represent a prolonged systemic stress which may alter normal GR regulation. The high circulating levels of cortisol found in AD patients would be expected to down regulate GR levels in hippocampus [17]. Thus, even in the absence of neuron loss, we still would have predicted that levels of mRNA for GR should have been slightly less in AD than in control. However, in the present study in which there were no significant differences between AD and control in the density of neurons in the hippocampal regions examined, GR mRNA levels were equal or higher in AD hippocampus.
The higher levels of GR mRNA in AD hippocam- pus may be the end result of many different processes. The hippocampal disconnection associated with AD [16] or the loss of cholinergic basal forebrain innerva- tion [6] may result in decreased neuronal activity which might alter regulation of GR. Recent animal experi-
ments suggest that basal forebrain lesions upregulate GR mRNA levels in some regions of hippocampus [51].
Neuronal pathology, in particular NFT's, may inter- fere with nuclear translocation of activated GR which normally acts to down regulate receptor levels. Hy- potheses suggest that the cytoskeleton may play an important role in translocation, perhaps in assocation with various molecular chaperons (i.e. heat shock pro- tein 90 and 70) known to interact with GR [26,27]. The inability of hippocampal GR to function normally sec- ondary to NFT's, resulting in a defective feedback mechanism, would also be parsimonious with the HPA dysfunction present in a subset of AD patients, and the normal and elevated mRNA levels observed in the present study. We examined this possibility by deter- mining whether any correlations existed between re- gional NFT numbers and regional GR mRNA levels, but we were unable to find an association. However since we did not examine this question at a cellular level, it is still possible that individual neurons with high NFT levels have disrupted GR regulation. Also, our patients had severe disease and their NFT burden was extremely high, possibly creating a ceiling effect. Additional studies in subjects in the early stages of the disease may be needed to clarify this issue.
Cytokines associated with cell degeneration and the mild inflammatory response observed in AD [45] may act directly or indirectly to increase GR levels. In peripheral tissues glucocortiocoid receptors are in- volved in inflammtory responses and glucocortioids can modulate the levels of some cytokines and cytokine receptors [14,43,47]. Increased levels of IL-lfl, IL-la, IL-6, and TNF have been reported in AD [23].
Finally, increased GR mRNA may reflect apoptotic cell death in a small population of neurons. A major role of glucocorticoids and glucocorticoid receptors in apoptosis in lymphocytes and other cell types has been established [4,36,53]. In a lymphoblast cell line, a brief increase in GR receptor and GR mRNA levels imme- diately precedes cell death [2,9]. Since AD is a chronic disorder progressing over decades the pathological pro- cess is indolent, and cell desfunction and death grad- ual. Thus, at any given time, only a small number of cells will exhibit an increased signal.
Our observations, taken together with those of Seckl et al. [39], indicate that levels of GR mRNA are normal or slightly increased in regions of the hip- pocampus of AD patients when compared to controls, and suggest that more complex processes than simply an increased vulnerability of GR containing neurons, are involved. Especially since granule cells of the den- tate gyrus, which are largely spared in AD, contain high levels of GR. Further studies of this phenomena, particularly within the context of glucocorticoid in- volvement in inflammatory reactions and modulatory roles of cytokines, and/or the role of atlered cyto-

80 D.M. Wetzel et al. / Brain Research 679 (1995) 72-81
skeletal proteins, may yield new insights into the pathogenesis of Alzheimer's disease and perhaps other neurodegenerative diseases as well.
Acknowledgements
We thank Drs. R. Evans for kindly supplying GR cDNA. We acknowledge the skillful technical assis- tance of Lori Shimoda and Syed Hussain and the secretarial assistance of Sharon Mills. The scientific advice of Drs. Kathy Maquire-Zeiss, James Powers, Lowell Lapham and the members of the Rochester Alzheimer's Disease Project is gratefully appreciated. This research was funded by grants from the National Institutes of Aging - AG03644 and AG 09016 (R.W. Hamill) and by the Adler Foundation (R.W. Hamill).
References
[1] Amaral, D.G. and Insausti, R., Hippocampal formation., In 0. Paxinos, (Eds.), The Human Nervous System, Academic Press, New York, 1990, pp. 711-755.
[2] Antakly, T., Thompson, E.B. and O'Donnell, D., Demonstration of the intracellular localization and up-regulation of glucocorti- coid receptor by in situ hybridization and immunocytochemistry, Cancer Res. Suppl., 49 (1989) 2230s-2234s.
[3] Bohn, M.C., Howard, E., Vielkind, U. and Krozowski, Z., Glial cells express both mineralocorticoid and glucocorticoid recep- tors, J. Steroid Biochem. Mol. Biol., 40 (1991) 105-11.
[4] Cohen, J.J. and Duke, R,, Glucocorticoid activation of a cal- cium-dependent endonuclease in thymocyte nuclei leads to cell death., J. Immunol., 132 (1984) 38-42.
[5] Coleman, P.D. and Flood, D.G., Neuron numbers and dendritic extent in normal aging and Alzheimer's disease, Neurobiol. Aging, 8 (1987) 521-545.
[6] Coyle, J.T., Price, D.L. and DeLong, M.R., Alzheimer's disease: A disorder of cortical cholinergic innervation, Science, 219 (1983) 1184-1190.
[7] Davies, D.C., Horwood, N., Issacs, S.L. and Mann, D.M.A., The effect of age and Alzheimer's disease on pyramidal neuron density in individual fields of the hippocampal formation, Acta Neuropathol., 83 (1992) 510-517.
[8] DeLeon, M.J., McRae, T., Tsai, J.R., George, A.E., Marcus, D.L., Freedman, M., Wolf, A.P. and McEwen, B., Abnormal cortisol response in Alzheimer's disease linked to hippocampal atrophy, Lancet, 13 (1988) 391.
[9] Eisen, L.P., Elsasser, M.S. and Harmon, J.M., Positive regula- tion of the glucocorticoid receptor in human T-cells sensitive to the cytolytic effects of glucocorticoids, J. Biol. Chem., 263 (1988) 12044-12048.
[10] EIdridge, J.C,, Brodish, A., Kute, T.E. and Landfield, P.W., Apparent age-related resistance of type II hippocampal cortico- steroid receptors to down-regulation during chronic escape training, J. Neurosci., 9 (1989) 3237-3242.
[11] Finch, C.E. and Morgan, D.G., RNA and protein metabolism in the aging brain, Annu. Rev. Neurosci., 13 (1990) 75-87.
[12] Flugge, G., Schniewind, A. and Fuchs, E., The corticosterone receptive system in the brain of Tupaia belangeri visualized by in vivo autoradiography, Exp. Brain Res., 72 (1988) 417-424.
[13] Gerlach, J.L., McEwen, J.S., Pfaff, D.W., Moskovitz, S., Ferin, M., Carmel, P.W. and Zimmerman, E.A., Cells in regions of rhesus monkey brain and pituitary retain radioactive estradiol, corticosterone and cortisol differentially, Brain Res., 103 (1976) 603-612.
[14] Gottschatl, P.E., Koves, K., Mizuno, K., Tatsuno, I. and Arimura, A., Glucocorticoid upregulation of interleukin 1 receptor ex- pression in a glioblastoma cell line, Am. J. Physiol., 261 (1991) 362-368.
[15] Hollenberg, S.M., Weinberger, C., Ong, E.S., Cerelli, G., Oro, A., Lebo, R., Thompson, E.B., Rosenfeld, M.G. and Evans, R.M., Primary structure and expression of a functional human glucocorticoid receptor cDNA, Nature, 318 (1985) 635-641.
[16] Hyman, B.T., Van Hoesen, G.W., Damasio, A.R. and Barnes, C.L., Alzheimer's disease: cell-specific pathology isolates the hippocampal formation, Science, 225 (1984) 1168-1170.
[17] Jacobson, L. and Sapolsky, R., The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenocorti- cal axis, Endocrine Rev., 12 (1991) 118-134.
[18] Kay, E.D., Goldman, A.S. and Daniel, J.C., Common biochemi- cal pathway of dysmorphogenesis in murin ebryos: use of the glucocorticoid pathway by phenytoin, Teratogen. Carciongen. Mutagen., 10 (1990) 31-39.
[19] Khachaturian, Z.S., Diagnosis of Alzheimer's disease, Arch. Neurol., 42 (1985) 1097-1105.
[20] Lorento de No, R., Studies on the structure of the cerebral cortex. II. Continuation of the study of ammonic system, J. Psychol. Neurol., 46 (1934) 113-177.
[21] Lupien, S., Lecours, A.R., Lussier, I., Schwartz, G., Nair, N.P.V. and Meaney, M.J., Basal cortisol levels and cognitive deficits in human aging, J. Neurosci., 14 (1994) 2893-2903.
[22] Maeda, K., Tanimoto, K., Terada, T., Takeshi, S. and Tatsuya, T., Elevated urinary free cortisol in patients with dementia, Neurobiol. Aging, 12 (1991) 161-163.
[23] McGeer, P.L. and Rogers, J., Anti-inflammatory agents as a therapeutic approach to Alzheimer's disease, Neurology, 42 (1992) 447-9.
[24] Miller, J.A., The calibration of 35S or 32p with 14C-labeled brain paste or 14C-plastic standards for quantiative autoradiography using LKB Ultrofilm or Amersham Hyperfilm, Neurosci. Lett., 121 (1991) 211-214.
[25] Peiffer, A., Barden, N. and Meaney, M.J., Age-related changes in glucocorticoid receptor binding and mRNA levels in the rat brain and pituitary, Neurobiol. Aging, 12 (1991) 475-9.
[26] Picard, D. and Yamamoto, K.R., Two signals mediate hormone-dependent nuclear localization of the glucocorticoid receptor, EMBO J., 6 (1987) 3333-3340.
[27] Pratt, W., Glucocorticoid Receptor Structure and the Initial Events in Signal Transduction, Molecular Endocrinology and Steroid Hormone Action, Alan Liss, New York, 1990, 119-132.
[28] Raskind, M., Peskind, E., Rivard, M.-F., Veith, R. and Barnes, R., Dexamethasone suppression test and cortisol circadian rhythm in primary degenerative dementia, Am. J. Psychiatry, 139 (1982) 14681471.
[29] Rentrop, M., Knapp, B., Winter, H. and Schweizer, J., Aminoalkysilane-treated glass slides as support for in situ hy- bridization of keratin cDNAs to frozen tissue sections under varying fixation and pretreatment conditions, Histochem. J., 18 (1986) 271-276.
[30] Reul, M.M.H.M., Tonnaer, J. and De Kloet, E.R., Neurotrophic ACTH analogue promotes plasticity of type 1 corticosteroid receptor in brain of senescent male rats, Neurobiol. Aging, 9 (1988) 253-260.
[31] Sapolsky, R.M., Stress, the Aging Brain, and Mechanisms of Neuron Death, MIT Press, Cambrige, MA, 1992.
[32] Sapolsky, R.M., Krey, L. and McEwen, B., Corticosterone re-

D.M. Wetzel et al. /Brain Research 679 (1995) 72-81 81
ceptors decline in a site-specific manner in the aged rat brain, Brain Res., 289 (1983) 235-240.
[33] Sapolsky, R.M. and Meaney, M.J., Postmortem decay in gluco- corticoid binding in human and primate brain, Brain Res., 448 (1988) 182-185.
[34] Sapolsky, R.M., Zola-Morgan, S. and Squire, L.R., Inhibition of glucocorticoid secretion by the hippocampal formation in the primate, J. Neurosci., 11 (1991)3695-3707.
[35] Sarrieau, A., Dussaillant, M., Agid, F., Philibert, D., Agid, Y. and Rostene, D., Radioaatographic localization of glucocorti- costeroid and progesterone binding sites in the human post- mortem brain, J. Steroid Biochem., 25 (1986) 717-721.
[36] Schwartzman, R.A. and Cidlowski, J.A., Internucleosomal de- oxyribonucleic acid cleavage avtivity in apoptotic thymocytes: detection and endocrine regulation, Endocrinology, 128 (1991) 1190-1197.
[37] Seckl, J.R., Dickson, K.L., Yates, C. and Fink, G., Distribution of glucocorticoid and mineralocorticoid receptor messenger RNA expression in human postmortem hippocampus, Brain Res., 56l (1991) 332-337.
[38] Seckl, J.R., French, K.L., O'Donnell, D., Meaney, M.J., Nair, N.P.V., Yates, C.M. and Fink, G., Glucocorticoid receptor gene expression is unaltered in hippocampal neurons in Alzheimer's disease, Mol. Brain Res., 18 (1993) 239-245.
[39] Seckl, J.R., O'Donnell, D., Meaney, M.J., Yates, C. and Fink, G., Hippocampal glucocor):icoid and mineralocorticoid receptor gene expression is unaltered in Alzheimer's disease, Soc. Neu- rosci. Abstr., 18 (1992) 479.
[40] Skare, S., Pew, B. and Dysken, M., The dexamethasone suppres- sion test in dementia: a review of the literature, J. Geriatric Psych. Neurol., 3 (1990) 12,l-138.
[41] Starkman, M.N., Gerbaski, S.S., Berent, S. and Schteingart, D.E., Hippocampal formation volume, memory dysfunction and cortisol levels in patients with Cushings syndrome, Biol. Psychia- try, 32 (1992) 756-765.
[42] Troxler, R.G., Sprague, E.A., Albanese, R.A., Fuchs, R. and Thompson, A.J., The assocriation of elevated plasma cortisol and early atherosclerosis as demonstrated by coronary angiography, Atherosclerosis, 26 (1977) 151-162.
[43] Vacca, A., Martinotti, S., Screpanti, I., Maroder, M., Felli, M.P., Farina, A.R. et al., Transcriptional regulation of the interleukin
2 gene by glucocorticoid hormones, J. Biol. Chem., 265 (1990) 8075-8080.
[44] van Eekelen, J.A.M., Jiang, W., De Kloet, E.R. and Bohn, M.C., Distribution of the mineralocorticoid and the glucocorticoid receptor mRNAs in the rat hippocampus, Neurosci. Res., 21 (1988) 88-94.
[45] Vandenabeele, P. and Fiers, W., Is amyloidogenesis during Alzheimer's disease due to an IL-1-/IL-6-mediated acute phase response' in the brain?, Immunol. Today, 12 (1991) 217-9.
[46] Vielkind, U., Walencewicz, A., Levine, J.M. and Bohn, M.C., Type II glucocorticoid receptors are expressed in oligodendro- cytes and astrocytes, J. Neurosci. Res., 27 (1990) 360-73.
[47] Waage, A., Slupphaug, G. and Shalaby, R., Glucocorticoids inhibit the production of IL6 from monocytes, endothelial cells and fibroblasts, Eur. J. Immunol., 20 (1990) 2439-2443.
[48] West, M.J. and Gunderson, H.J.G., Unbiased stereological esti- mation of the number of neurons in the human hippocampus, J. Comp. Neurol., 296 (1990) 1-22.
[49] Wetzel, D.M., Bohn, M.C. and Hamill, R.W., Postmortem sta- bility of mRNA for glucocortcoid and mineralocorticoid recep- tor in rodent brain, Brain Res., 649 (1994) 117-121.
[50] Wetzel, D.M., Hussain, S.S., Bohn, M.C. and Hamill, R.W., The distribution of glucocorticoid and mineralocorticoid receptor mRNA's in human hippocampus, Ann. Neurol., 30 (1991) 249.
[51] Yau, J.L.W., Dow, R.C., Fink, G. and Seckl, J.R., Medial septal cholinergic lesions increase hippocampal mineralocorticoid and glucocorticoid receptor messenger RNA expression, Brain Res., 577 (1992) 155-160.
[52] Young, E.A., Haskett, R.F., Murphy-Weinbert, V., Watson, S.J. and Akil, H., Loss of glucocorticoid fast feedback in depression, Arch. Gen. Psychol., 48 (1991) 693-699.
[53] Zacharchuk, C.M., Mercep, M., Chakraborti, P.K., Simons, S.S. and Ashwell, J.D., Programmed T lymphocyte death. Cell acti- vation- and steroid-induced pathways are mutually antagonistic, J. Immunol., 145 (1990) 4037-4045.
[54] Zoli, M., Ferraguti, F., Gustafsson, J.-A., Toffano, G., Fuxe, K. and Agnati, L.F., Selective reduction of glucocorticoid receptor immunoreactivity in the hippocampal formation and central amygdaloid nucleus of the aged rat, Brain Res, 545 (1991) 199-207.
Copyright © 2022 FDOKUMEN




















