
Expression of a novel non-coding mitochondrial RNA in human proliferating cells
12
7336–7347 Nucleic Acids Research, 2007, Vol. 35, No. 21 Published online 25 October 2007 doi:10.1093/nar/gkm863 Expression of a novel non-coding mitochondrial RNA in human proliferating cells Jaime Villegas 1,2,3 , Veronica Burzio 1,2,3 , Claudio Villota 1,2 , Eduardo Landerer 1,3 , Ronny Martinez 1 , Marcela Santander 1 , Rodrigo Martinez 1 , Rodrigo Pinto 3 , Marı´a I. Vera 3 , Enrique Boccardo 4 , Luisa L. Villa 4 and Luis O. Burzio 1,2,3, * 1 Bios Chile Ingenierı´a Gene ´ tica S.A., 2 Institute for Fundamental and Applied Biology, Fundacio ´ n Ciencia para la Vida, Avenida Zan ˜ artu 1482, 3 Laboratorio de Biologı´a Celular y Molecular and Department of Urology, Facultad de Ciencias de la Salud, Universidad Andre ´ s Bello, Repu ´ blica 252, Santiago, Chile and 4 Ludwig Institute for Cancer Research, Sao Paulo, Brazil Received August 20, 2007; Revised September 21, 2007; Accepted September 26, 2007 ABSTRACT Previously, we reported the presence in mouse cells of a mitochondrial RNA which contains an inverted repeat (IR) of 121 nucleotides (nt) covalently linked to the 5’ end of the mitochondrial 16S RNA (16S mtrRNA). Here, we report the structure of an equivalent transcript of 2374 nt which is over- expressed in human proliferating cells but not in resting cells. The transcript contains a hairpin structure comprising an IR of 815 nt linked to the 5’ end of the 16S mtrRNA and forming a long double- stranded structure or stem and a loop of 40 nt. The stem is resistant to RNase A and can be detected and isolated after digestion with the enzyme. This novel transcript is a non-coding RNA (ncRNA) and several evidences suggest that the transcript is synthesized in mitochondria. The expression of this transcript can be induced in resting lymphocytes stimulated with phytohaemag- glutinin (PHA). Moreover, aphidicolin treatment of DU145 cells reversibly blocks proliferation and expression of the transcript. If the drug is removed, the cells re-assume proliferation and over-express the ncmtRNA. These results suggest that the expression of the ncmtRNA correlates with the replicative state of the cell and it may play a role in cell proliferation. INTRODUCTION The mitochondrial DNA (mtDNA) is a closed-circular, double-stranded molecule that displays an exceptional economy of organization (1–3). In humans, the 16 500 bp of the genome encode the 12S and 16S ribosomal RNAs, 22 transfer RNAs (tRNAs) and 13 polypeptides (2,4–7). The H-strand encodes the 12S and 16S ribosomal RNAs, 14 tRNAs and 12 polypeptides, while the L-strand codes for 8 tRNAs and the ND6 subunit of NAD dehydrogenase (2,4–8). Between the tRNA Phe and tRNA Pro genes is the D-loop that has evolved as the major control region. Besides the H-strand origin of replication, the D-loop contains the major promoters that regulate transcription of the H- (HSP) and the L-strand (LSP) (9–11). Both strands are transcribed as polycistronic RNAs, which are then processed to release the individual mRNAs, tRNAs and rRNAs (3). The human mitochondrial RNA polymerase as well as transcription factors have been extensively described (6–8). Previously, we described the presence in mouse cells of a novel mitochondrial RNA containing an IR of 121 nt linked to the 5 0 end of the 16S mtrRNA (12,13). The IR generates a perfect double-stranded structure of 121 bp and a loop of 120 nt. In situ hybridization (ISH) revealed that this ncRNA is over-expressed in spermatogenic cells, especially in mouse proliferating spermatogonia (14). Similar results were obtained with human spermatogonia using a probe complementary to the 16S mtrRNA (14). These results suggest that human cells might contain a transcript with similar structural features to the mouse RNA, and that its expression correlates with cell proliferation. In this work, we report that the human RNA is over-expressed in several human proliferating cells but not in resting cells. The structure of this transcript of 2374 nt reveals the presence of an IR of 815 nt linked to the 5 0 end of the 16S mtrRNA. Together with the 16S mtrRNA, the IR forms a long double-stranded structure The authors wish it to be known that, in their opinion, the first three authors should be regarded as joint First Authors *To whom correspondence should be addressed. Tel: +56 2 473 6133; Fax: +56 2 2394250; Email: [email protected] ß 2007 The Author(s) This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/ by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited. by guest on March 7, 2014 http://nar.oxfordjournals.org/ Downloaded from
Transcript of Expression of a novel non-coding mitochondrial RNA in human proliferating cells

7336ndash7347 Nucleic Acids Research 2007 Vol 35 No 21 Published online 25 October 2007doi101093nargkm863
Expression of a novel non-coding mitochondrialRNA in human proliferating cellsJaime Villegas123 Veronica Burzio123 Claudio Villota12 Eduardo Landerer13
Ronny Martinez1 Marcela Santander1 Rodrigo Martinez1 Rodrigo Pinto3
Marıa I Vera3 Enrique Boccardo4 Luisa L Villa4 and Luis O Burzio123
1Bios Chile Ingenierıa Genetica SA 2Institute for Fundamental and Applied Biology Fundacion Ciencia para laVida Avenida Zanartu 1482 3Laboratorio de Biologıa Celular y Molecular and Department of Urology Facultad deCiencias de la Salud Universidad Andres Bello Republica 252 Santiago Chile and 4Ludwig Institute for CancerResearch Sao Paulo Brazil
Received August 20 2007 Revised September 21 2007 Accepted September 26 2007
ABSTRACT
Previously we reported the presence in mouse cellsof a mitochondrial RNA which contains an invertedrepeat (IR) of 121 nucleotides (nt) covalently linkedto the 5rsquo end of the mitochondrial 16S RNA (16SmtrRNA) Here we report the structure of anequivalent transcript of 2374nt which is over-expressed in human proliferating cells but not inresting cells The transcript contains a hairpinstructure comprising an IR of 815 nt linked to the5rsquo end of the 16S mtrRNA and forming a long double-stranded structure or stem and a loop of 40 ntThe stem is resistant to RNase A and can bedetected and isolated after digestion with theenzyme This novel transcript is a non-coding RNA(ncRNA) and several evidences suggest that thetranscript is synthesized in mitochondria Theexpression of this transcript can be induced inresting lymphocytes stimulated with phytohaemag-glutinin (PHA) Moreover aphidicolin treatment ofDU145 cells reversibly blocks proliferation andexpression of the transcript If the drug is removedthe cells re-assume proliferation and over-expressthe ncmtRNA These results suggest that theexpression of the ncmtRNA correlates with thereplicative state of the cell and it may play a role incell proliferation
INTRODUCTION
The mitochondrial DNA (mtDNA) is a closed-circulardouble-stranded molecule that displays an exceptional
economy of organization (1ndash3) In humans the16 500 bp of the genome encode the 12S and 16Sribosomal RNAs 22 transfer RNAs (tRNAs) and 13polypeptides (24ndash7) The H-strand encodes the 12S and16S ribosomal RNAs 14 tRNAs and 12 polypeptideswhile the L-strand codes for 8 tRNAs and the ND6subunit of NAD dehydrogenase (24ndash8) Between thetRNAPhe and tRNAPro genes is the D-loop that hasevolved as the major control region Besides the H-strandorigin of replication the D-loop contains the majorpromoters that regulate transcription of the H- (HSP)and the L-strand (LSP) (9ndash11) Both strands aretranscribed as polycistronic RNAs which are thenprocessed to release the individual mRNAs tRNAs andrRNAs (3) The human mitochondrial RNA polymeraseas well as transcription factors have been extensivelydescribed (6ndash8)
Previously we described the presence in mouse cells of anovel mitochondrial RNA containing an IR of 121 ntlinked to the 50 end of the 16S mtrRNA (1213) The IRgenerates a perfect double-stranded structure of 121 bpand a loop of 120 nt In situ hybridization (ISH) revealedthat this ncRNA is over-expressed in spermatogenic cellsespecially in mouse proliferating spermatogonia (14)Similar results were obtained with human spermatogoniausing a probe complementary to the 16S mtrRNA (14)
These results suggest that human cells might contain atranscript with similar structural features to the mouseRNA and that its expression correlates with cellproliferation In this work we report that the humanRNA is over-expressed in several human proliferating cellsbut not in resting cells The structure of this transcript of2374 nt reveals the presence of an IR of 815 nt linked tothe 50 end of the 16S mtrRNA Together with the 16SmtrRNA the IR forms a long double-stranded structure
The authors wish it to be known that in their opinion the first three authors should be regarded as joint First Authors
To whom correspondence should be addressed Tel +56 2 473 6133 Fax +56 2 2394250 Email lburziobioschilecl
2007 The Author(s)
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (httpcreativecommonsorglicenses
by-nc20uk) which permits unrestricted non-commercial use distribution and reproduction in any medium provided the original work is properly cited
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
or stem which is resistant to RNase A digestion On theother hand in silico analysis revealed that this transcript isa new member of the expanding family of non-codingRNAs (ncRNA) (15ndash17) and therefore we named thismolecule non-coding mitochondrial RNA or ncmtRNAMoreover several experimental evidences stronglysuggest the mitochondrial origin of this transcriptISH using a probe specific for the ncmtRNA confirmedover-expression of this transcript in proliferating cellsThe expression of this transcript can be induced in restinglymphocytes stimulated with phytohaemagglutinin (PHA)(18) together with DNA synthesis and the expression ofthe proliferation markers proliferating cell nuclear antigen(PCNA) Ki-67 and phosphohistone H3 (19ndash21) On theother hand treatment of DU145 cells with aphidicolin(2223) reversibly blocks cell proliferation as well as theexpression of the ncmtRNA These results suggest that thencmtRNA is a new marker of cell proliferation
MATERIALS AND METHODS
Cell culture
HeLa SiHa DU145 MCF7 H441 Caco-2 and 4295(melanoma) cells were cultured in Dulbeccorsquos ModifiedEaglersquos Medium (DMEM Gibco) containing 10 fetalcalf serum (FCS) 2mM glutamine 50 mgml of penicillin50 mgml of streptomycin and 01mM non-essential aminoacids The human promyelocitic leukemia cell line HL-60was cultured in Iscoversquos modified Dulbeccorsquos medium(IMDM Gibco) supplemented with 20 fetal bovineserum 2mM glutamine 50 mgml of penicillin 50 mgml ofstreptomycin and 01mM non-essential amino acids Allhuman cell lines were maintained in a humidified cellculture chamber at 378C and 5 CO2 To inhibitmitochondrial transcription HeLa cells were cultured asdescribed with the addition of 50 ngml of ethidiumbromide plus 50 mgml of uridine and 1mM sodiumpyruvate (24) For aphidicolin treatment (2223) DU145cells were cultured for 16 h with 5 mgml of aphidicolinfollowed by 10 h without the drug Then the cells wereagain treated with 5 mgml of aphidicolin for another 16 hTo release the cells from the aphidicolin block they werecultured for 48 h in regular medium without the drug
Human peripheral blood lymphocytes obtained fromhealthy donors were isolated by Ficoll gradient centrifu-gation as described before (18) The cells were suspendedin RPMI 1640 medium supplemented with 10 FCS (18)and the number of cells per ml was determined in aNeubauer chamber The cells were cultured for 24 48 and72 h at 378C with 5 CO2 with or without 10 mgml ofPHA (18) About 100 000 cells in 200 ml of medium werecultured in 96-wells plates and pulsed for 16 h with BrdUstarting at 0 24 and 48 h Cells were collected at 24 48 and72 h of culture and BrdU incorporation was measuredcolorimetrically by ELISA according to manufacturerrsquosinstructions (Proliferation ELISA Roche)
Human tissue samples
Formalin-fixed paraffin-embedded human tissue sampleswere obtained from diagnostic biopsies or resection
specimens from patients at the Hospital Barros LucoTrudeau (Santiago Chile) The tissues were used inagreement with the ethical guidelines approved by theethical committee of the hospital and our institutions
RNA and DNA isolation
Total RNA from cells was extracted with TRIzol(Invitrogen) as described before (121325) The polyA+
fraction was obtained with the Oligotex mRNA Midi kit(Qiagen) according to the manufacturerrsquos instructionsTo eliminate mtDNA contamination RNA preparationswere treated with TURBO DNA-free (Ambion) accordingto the manufacturerrsquos instructions Human mtDNA wasprepared from human lymphocytes HeLa SiHa Hep G2and HL-60 cells (26)
RT-PCR
Reverse transcription was carried out with 50ndash100 ng offreshly prepared RNA 50 ng of random hexamers orsequence-specific primers and 200 U of reverse transcrip-tase (M-MLV or SuperScript II Invitrogen) (1213) ThecDNA was amplified by PCR and analyzed by electro-phoresis as described before (121327) The primers usedto amplify the sense 16S mtrRNA were P1 (r) 50 AAGGTGGAGTGGGTTTGGGGC (position 11ndash31) P9 (f)50 TAGGCCTAAAAGCAGCCACCAA (position501ndash522) P10 (f) 50 ACCGTGCAAAGGTAGCATAATCAC (position 912ndash935) P11 (r) 50 AATAGGATTGCGCTGTTATCCCTA (position 1260ndash1283) P12 (r) 50 CTGTTCTTGGGTGGGTGTG (position 1536ndash1554) For theantisense 16S mtrRNA P2 (f) 50 GGGGTCTTAGCTTTGGCTCTCC (position 1326ndash1347) P3 (f) 50 TTGGTGGCTGCTTTTAGGCCTA (position 1207ndash1227) P4 (f)50 GGTTGATTGTAGATATTGGGCT (position 833ndash854)P5 (f) 50 GGTAAGATTTGCCGAGTTC (position741ndash759) P6 (f) 50 GTGATTATGCTACCTTTGCACGGT (position 626ndash649) P7 (r) 50 ACCATTTACCCAAATAAAGTATAG (position 1483ndash1506) P8 (r) 50 GGACCAATCTATCACCCTATA (position 942ndash962) To iden-tify the position of each primer the sequence from nt 1 to1559 of the sense or antisense 16S mtRNA was used asreference For the region between the IR and the 16SmtrRNA P13 (r) 50 AGGTTTAGCCAAACCATT(807ndash824) For the 12S mtrRNA P14 (f) 50 AGCCTATATACCGCCATCTTC (position 604ndash624) P15 (r) 50 AAGTATACTTGAGGAGGGTGA (position 843ndash863)P16 (f) 50 GTGTACTGGAAAGTGCACTTG (position927ndash947) For COX I P17 (f) 50 GAACAGGTTGAACAGTCTACCCT (position 371ndash393) and P18 (r) 50 TTCCGAAGCCTGGTAGGATAAGA (position 738ndash760)For the 18S rRNA P19 (f) 50 GATGCGTGCATTTATCAGATC (position 309ndash329) and P20 (r) 50 AGTGGACTCATTCCAATTACA (position 652ndash672) For GAPDHP21 (f) 50 ACTCTGGTAAAGTGGATATTGT (position131ndash152) and P22 (r) 50 ATGATGTTCTGGAGAGCCC(position 662ndash680) For b-actin mRNA P23 (f)50 AAGAGAGGCATCCTCACCCTG (position 181ndash202)P24 (r) 50 GGCGACGTAGCACAGCTTCTCC (position639ndash660) (f) and (r) represent forward and reverseprimers respectively The sequence of the primers for
Nucleic Acids Research 2007 Vol 35 No 21 7337
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
the 16S 12S and COX 1 transcripts were deduced from thehuman mtDNA (GenBank Accession No V00662)The sequence of the primers for the 18S rRNA GAPDHmRNA and b-actin mRNA were deduced from theGenBank Accession Nos M10098 M33197 andNM_001101 respectively Amplified DNA fragmentswere purified (Wizard SU Gel and PCR Clean-up systemPromega) cloned in pGEM-T Easy (Promega) or pTOPO(Invitrogen) and the purified recombinant plasmids weresequenced as described before (1213)
RNase digestion
About 1 mg of total RNA from human cells treated withTURBO DNA-free in 50 ml of 2 SSC (28) was incubatedwith RNase A at a final concentration of 50 mgml for15min at 258C The digestion products were extractedwith phenolndashchloroform and after adding 10 mg ofglycogen to the aqueous phase the RNA was precipitatedwith ethanol and recovered by centrifugation (1213)
S1 protection assay
S1 protection assay was carried out using a single-stranded digoxigenin-labeled DNA probe complementaryto the region between the IR and the 16S mtrRNA of thencmtRNA First an amplicon was obtained by RT-PCRusing primers 1 and 2 and this fragment was used as atemplate in an asymmetric PCR reaction that containedonly primer 1 and digoxigenin-11-dUTP (Roche Non-Radioactive ISH) S1 protection was carried out asdescribed (27) using 50 mg RNA and 50 ng digoxigenin-labeled probe After S1 digestion reactions were ethanol-precipitated and analyzed on native 25 agarose gelswhich were transferred onto HybondTM-XL membranes(Amersham) After blocking with 002 Tween 20 plus5 skim milk in PBS the position of the digoxigenin-labeled probe was revealed with anti-digoxigenin antibodyconjugated with alkaline phosphatase (Roche) and devel-oping in BCIPNBT alkaline phosphatase substratemixture (DAKO) for 10min
5rsquoRACE
HeLa cells RNA dissolved in 2 SSC was digested withRNase A and the double-stranded structure was recoveredafter phenol extraction as described before The cDNAwas synthesized with primer P8 (Figure 2a) and tailed atthe 30 end with dCTP using terminal deoxynucleotidyl-transferase (TdT Promega) (29) Amplification by PCRwas carried out with the specific primer 8 (Figure 2a) andthe anchor primer provided by the manufacturer Theamplicon obtained of 250 bp was purified and bothstrands were sequenced directly
Northern blot
Total RNA of HeLa and MCF7 cells (3 mg each) wereelectrophoresed at 75V for 90min in a native 10agarose gel prepared in TAE buffer or in an agarose gelsunder denaturing conditions containing 22M formalde-hyde (27) The RNAs were transferred in 20 SSC to anylon membrane (HybondTM-XL Amersham) for 18 h
and exposed to UV light for 5min The probe used wasprimer 13 (Figure 2a) labeled with 32P Briefly in a finalvolume of 20 ml 100 ng of the oligonucleotide were mixedwith 4 ml of enzyme buffer (100mM cacodylate buffer pH68 1mM CoCl2 and 01mM DTT) 2 ml of [a-32P]-dCTP(3000Cimmol) and 10U of TdT and incubated at 378Cfor 30min The probe was purified using a Sephadex G-50spin column (Amersham) Hybridization was carried outfor 18 h at 378C with 5ml of 05M sodium phosphate pH71 2mM EDTA 7 SDS and 01 sodium pyropho-sphate (30) containing 32P-labeled primer 13 at a ratio of105 cpmcm2 of membrane surface area The blots werewashed twice for 5min at room temperature with 2 SSCplus 01 SDS and once with 05 SSC plus 01 SDSRadioactivity on the membranes was visualized with aphosphor imager (MolecularTM Imager FX PhosphorImaging System BioRad)
About 1 mg of RNA digested with RNAse A asdescribed before was subjected to electrophoresis on a15 agarose gel containing 22M formaldehyde andtransferred to a nylon membrane (27) The membrane wasprobed with an amplicon of 250 bp corresponding to thedouble-stranded region of the ncmtRNA and obtained byRT-PCR using primers 8 and 5 (Figure 2a) and[a-32P]-dCTP The probe was extracted with phenol andprecipitated with isopropanol Hybridization was carriedout with a solution containing 8 million cpmml in4 SSC 10 dextran sulfate 150 mgml yeast tRNA150 mgml herring sperm DNA 50 formamide and1Denhardtrsquos solution (27) After hybridization at 658Covernight the membrane was washed for 10min at roomtemperature with 2 SSC and 1SSC 20min at 558Cwith 05 SSC and 20min at room temperature with02 SSC Then the membrane was exposed to X-ray filmsat 708C (1227)
Mitochondria isolation
HeLa cells were grown as indicated and about 5 108 cellswere harvested and recovered by centrifugation at 300gfor 5min at 48C The cells were resuspended in about 10volumes of a hypotonic solution containing 10mM KCl015mM MgCl2 and 10mM TrisndashHCl pH 68 incubatedfor 10min on ice and dounce homogenized with the tightpestle (1331) The homogenization was monitored byphase microscopy until 70 of the cells were broken andthe mitochondrial fraction was obtained as describedbefore (31) The final mitochondria fraction was resus-pended in 2ndash3ml of 025M sucrose 2mM MgCl2 and04mM sodium phosphate buffer at pH 68 and treatedwith RNAse A at a final concentration of 50 mgml for15min at room temperature (32) The mitochondriafraction was recovered by centrifugation at 10 000g for15min suspended in 100 ml of PBS containing 100 U ofRNaseOut (Invitrogen) and mitochondrial RNA wasextracted with TRIzol as described before
ISH
Cells cultured in 8-well chamber slides (Lab-TekNUNC) for 24ndash48 h were washed three times with PBSand fixed in 4 paraformaldehyde in PBS for 10min at
7338 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
room temperature The slides were then washed threetimes with PBS for 5min and incubated with 02N HClfor 10min at room temperature Hybridization wascarried out essentially as described before (1214) Thecells were hybridized for 18 h at 378C with 200 ml of thehybridization solution containing 35 pmol of the anti-sense probe (primer 6 or 13) or the corresponding senseprobes (Figure 2a) previously labeled at the 30 end withdigoxigenin-11-dUTP (Roche Applied Science) (1214)The slides were washed with 2SSC and 1SSC for10min at room temperature 02 SSC for 30min at 378Cand finally with 02 SSC for 10min at room temperature(14) Then the cells were incubated for 2 h at roomtemperature with anti-digoxigenin antibody conjugated toalkaline phosphatase (Roche Applied Science) diluted1500 in the blocking buffer (1 BSA 03 Triton X-100in PBS) The color reaction was carried out with a BCIPNBT substrate mixture (DAKO) plus levamisol for 30min(14) Resting or PHA-stimulated human lymphocyteswere applied on silanized slides air dried fixed with 4paraformaldehyde and subjected to the same hybridiza-tion protocol For FISH after hybridization the cells wereincubated with anti-digoxigenin antibody conjugated toFITC (Roche Applied Science) For tissue samplesparaffin sections of 5 mm thick were collected in silanizedslides and deparaffinized by immersion in three consecu-tive xylene baths 10min each The sections wererehydrated with three washes in 100 90 70 and50 ethanol once in PBS and once in distilled water for10min Afterwards the sections were fixed with 4paraformaldehyde in PBS and incubated with 02N HClplus 4mgml of pepsin for 10min at room temperatureand subjected to ISH as described before
Immunocytochemistry and FISH
After ISH melanoma cells (4295) were incubated for30min in blocking buffer (1 BSA 03 Triton X-100 inPBS) and then incubated for 2 h at room temperaturewith anti-digoxigenin antibody conjugated with fluores-cein (Roche) and with anti-PCNA monoclonal antibody(DAKO) The cells were washed three times with 005Tween20 in PBS incubated for 2 h at room temperaturewith anti-mouse IgG conjugated with rhodamin washedagain and mounted with DABCO After hybridizationHeLa cells were incubated with anti-digoxigenin antibodyconjugated with rhodamin (Roche) and with anti-cyto-chrome c monoclonal antibody (Promega) or anti-endonuclease G polyclonal antibody (ChemiconInternational) After washing with 005 Tween20 inPBS the cells were incubated with either anti-mouse IgGconjugated with fluorescein or with anti-rabbit IgG con-jugated with fluorescein Fluorescence microscopy wasanalyzed with a Olympus BX51 microscope Confocalmicroscopy was performed with a LSM 5 Zeiss micro-scope equipped with a 63 objective The analysis wascarried out with the Zeiss LSM 5 Image Browser software
Fixed resting or PHA-stimulated lymphocytes wereincubated with the blocking solution and then withanti-PCNA (DAKO) or anti-Ki67 antigen (DAKO) oranti-phosphohistone H3 (Upstate) for 60min at
room temperature After washing in PBS the sectionswere incubated for 30min with anti-mouse IgG con-jugated to alkaline phosphatase (KPL) diluted 1250 in2 BSA in PBS The color reaction was developed for20min with BCIPNBT substrate mixture (DAKO)containing levamisol Previous to immunocytochemistrydeparaffinized tissue sections were incubated in coplin jarsfor 15min at 958C in Target Retrieval Solution (DAKO)diluted 110 with distilled water The incubation with anti-PCNA was as described before
RESULTS
The human ncmt RNA
As shown in Figure 1a (panel 1) ISH with probe 6targeted to the 16S mtrRNA (see Materials and Methodssection) revealed strong hybridization signals in HeLaHL-60 MCF7 DU145 H441 and Caco-3 human cellsAs expected hybridization with the control sense probe 10(see Materials and Methods section) was negative(Figure 1a panel 2) Moreover FISH with probe 6 incombination with immunocytochemistry of PCNA of themelanoma cell line 4295 revealed that each cell expressingthe transcript was also expressing PCNA (Figure 1b) Thehybridization with probe 10 was again negative(Figure 1b) As a positive control FISH was carriedwith probe 20 (see Materials and Methods section) todetect the 18S rRNA (Figure 1b)The target of probe 6 might be a transcript homologous
to the mouse ncRNA (12) Therefore to characterize thistranscript a hypothetical human mitochondrial RNA wasdeduced (Figure 2a) The transcript contains the completesequence of the 16S mtrRNA (Figure 2a red line) linked atits 50 end to a fragment of the RNA transcribed from the16S gene corresponding to the L-strand of the mtDNA orantisense 16S mtrRNA (Figure 2a blue line) Based on thisstructure primers were designed to amplify the putativetranscript by RT-PCR (Figure 2a) Primer 1 was posi-tioned at the theoretical loop between positions 11 and 31of the human sense 16S mtrRNA while the forwardprimer 2 was at the putative IR corresponding to position1326ndash1347 of the antisense 16S mtrRNA (Figure 2a)RT-PCR carried out with RNA from HeLa HL-60 andMCF7 cells yielded a single amplicon of 210 bp(Figure 2c odd lanes) No amplification was obtainedwhen reverse transcriptase was omitted from the reaction(Figure 2c even lanes) The sequence of the amplicon ofeach cell line revealed that an IR of 184 nt was linked to thefirst 31 nt at the 50 end of the 16S mtrRNA (SupplementaryFigure S1a and GenBank Accession No DQ386868)To determine whether the IR was longer than 184 nt a
PCR-walking strategy was utilized The cDNA fromHeLa cells was amplified between primer 1 and primers3 4 5 and 6 targeted to a putative longer IR (Figure 2a)Amplification fragments of 500 700 and 800 bp wereobtained when primer 1 was used in combination withprimers 3 4 and 5 respectively (Figure 2d lanes 1 3 and 5)No amplification product was obtained with primers 1 and6 (Figure 2d lanes 7 and 8) suggesting that the 50 of theIR is positioned between primers 5 and 6 (see below)
Nucleic Acids Research 2007 Vol 35 No 21 7339
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
The sequence of the amplicon of 800 bp revealed an IR of769 nt linked to the first 31 nt of the 16S mtrRNA(Supplementary Figure S1b and GenBank Access NoDQ386868) The sequence of the region between the 16SmtrRNA and the IR is identical to that found in the sameregion of the amplicon of 215 bp (compare SupplementaryFigures S1a and b) confirming that in both cases we wereamplifying the same transcript The sequences of the 500and 700 bp fragments indicated that they were part of the800 bp amplicon The same results and sequences wereobtained with RNA from HL-60 cells PHA-stimulatedlymphocytes and MCF7 cells (data not shown)
The stem of the ncmtRNA is resistant to RNase digestion
The 769 nt of the IR are fully complementary to the16S mtrRNA Therefore the transcript should contain
HeLa HL-60 MCF7C1
DU145 H441 Caco-2
(a)
1
1 1
2
2
2
2
1
1
2
Probe 6(b)
Probe 10
PCNA
Merge
18S r RNA
1
2
B1A1
A2
A3
Figure 1 Expression of the hypothetical human ncmtRNA (a) ISH ofthe indicated cells with probe 6 (panels 1) or the control sense probe 10(panels 2) Note cytoplasmic and perinuclear hybridization signals(100) (b) Melanoma cell line expressing the transcript are alsoexpressing PCNA The cells were subjected to ISH with probe 6 orprobe 10 together with immunocytochemistry of PCNA Themerge figure confirmed the co-expression of both molecules As apositive control the same cells were subjected to ISH for the 18SrRNA and immunocytochemistry of PCNA (63)
(d)
(c)
RT + minus + minus + minus
MM 1 2 3 4 5 6HeLa HL-60 MCF7
MM
RT + minus + minus + minus + minus
1 2 3 4 5 6
210 bp
700 bp 800 bp
2 3 4
1
65
7
11
89
63prime
2 3 4
1
65
7
11
8 10
63prime
RNas e A
(a)
12
13
13
500 bp
7 8
600 bp
10
Primers 1-3 1-4 1-5 1-6
9(b)
2 3 4 5 61 MM
800 bp 750 bp
370 bpRNase A minus + minus + minus +
(e)
500 bp
(f) GAPDH12S 18S
M1 2 3 4 5 6
RNase A minus + minus + minus +
500 bp
HL-
60M
CF
7
HeL
a
872 bp
603 bp
1078 bpg)
Figure 2 The human ncmtRNA (a) Theoretical structure of thehuman ncmtRNA The 50 end of the sense 16S mtrRNA (red line) islinked to a fragment of the antisense 16S mtrRNA or IR (blue line)forming a double-stranded structure and a loop of unknown lengthThe position of the reverse primers (under the lines) and the forwardprimers (over the lines) are indicated (see Materials and Methodssection) (b) Digestion of the loop and the single-stranded region of thencmtRNA by RNase A is also illustrated (c) Amplification of thecDNA obtained from three tumor cell lines using primers 1 and 2 Anamplicon of 210 bp was obtained only when the reaction was carriedout with reverse transcriptase (c odd lanes) (d) Amplification of thecDNA of HeLa cells by PCR using primers 1 in combination withprimers 3 (lanes 1 and 2) 4 (lanes 3 and 4) 5 (lanes 5 and 6) and 6(lanes 7 and 8) respectively Amplicons of 500 700 and 800 bp wereobtained No amplification products were generated with primer 1 and6 (lanes 7 and 8) or without reverse transcriptase (d even lanes) M100 bp ladder (e) RNA from HeLa cells in 2 SSC was incubatedwithout (odd lanes) or with 50 mgml of RNase A for 15min at 258C(even lanes) The RNA was recovered and amplified by RT-PCR usingprimers 1 and 5 (lanes 1 and 2) primers 10 and 11 (lanes 3 and 4) orwith primers 7 and 5 (lanes 5 and 6) Amplicons of 800 and 350 bp wereobtained with primer 1 and 5 and 10 and 11 respectively only withuntreated RNA (lanes 1 and 3 respectively) Amplification of thefragment of 750 bp obtained with primers 7 and 5 was not affected bythe nuclease treatment (lanes 5 and 6) (f) Amplification of the 12SmtrRNA (lanes 1 and 2) 18S rRNA (lanes 3 and 4) and GAPDHmRNA (lanes 5 and 6) after digestion with RNase A (mockexperiment odd lanes) (g) About 1 mg of RNA from the indicatedcell lines was digested with RNase A and the digestion products wereresolved by electrophoresis on a 15 agarose gel After blotting themembrane was probed with a 32P-labeled PCR fragment targeted to thedouble-stranded region of the ncmtRNA (see Materials Methodssection) A single hybridization band corresponding to a transcript of800 nt was detected
7340 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

or stem which is resistant to RNase A digestion On theother hand in silico analysis revealed that this transcript isa new member of the expanding family of non-codingRNAs (ncRNA) (15ndash17) and therefore we named thismolecule non-coding mitochondrial RNA or ncmtRNAMoreover several experimental evidences stronglysuggest the mitochondrial origin of this transcriptISH using a probe specific for the ncmtRNA confirmedover-expression of this transcript in proliferating cellsThe expression of this transcript can be induced in restinglymphocytes stimulated with phytohaemagglutinin (PHA)(18) together with DNA synthesis and the expression ofthe proliferation markers proliferating cell nuclear antigen(PCNA) Ki-67 and phosphohistone H3 (19ndash21) On theother hand treatment of DU145 cells with aphidicolin(2223) reversibly blocks cell proliferation as well as theexpression of the ncmtRNA These results suggest that thencmtRNA is a new marker of cell proliferation
MATERIALS AND METHODS
Cell culture
HeLa SiHa DU145 MCF7 H441 Caco-2 and 4295(melanoma) cells were cultured in Dulbeccorsquos ModifiedEaglersquos Medium (DMEM Gibco) containing 10 fetalcalf serum (FCS) 2mM glutamine 50 mgml of penicillin50 mgml of streptomycin and 01mM non-essential aminoacids The human promyelocitic leukemia cell line HL-60was cultured in Iscoversquos modified Dulbeccorsquos medium(IMDM Gibco) supplemented with 20 fetal bovineserum 2mM glutamine 50 mgml of penicillin 50 mgml ofstreptomycin and 01mM non-essential amino acids Allhuman cell lines were maintained in a humidified cellculture chamber at 378C and 5 CO2 To inhibitmitochondrial transcription HeLa cells were cultured asdescribed with the addition of 50 ngml of ethidiumbromide plus 50 mgml of uridine and 1mM sodiumpyruvate (24) For aphidicolin treatment (2223) DU145cells were cultured for 16 h with 5 mgml of aphidicolinfollowed by 10 h without the drug Then the cells wereagain treated with 5 mgml of aphidicolin for another 16 hTo release the cells from the aphidicolin block they werecultured for 48 h in regular medium without the drug
Human peripheral blood lymphocytes obtained fromhealthy donors were isolated by Ficoll gradient centrifu-gation as described before (18) The cells were suspendedin RPMI 1640 medium supplemented with 10 FCS (18)and the number of cells per ml was determined in aNeubauer chamber The cells were cultured for 24 48 and72 h at 378C with 5 CO2 with or without 10 mgml ofPHA (18) About 100 000 cells in 200 ml of medium werecultured in 96-wells plates and pulsed for 16 h with BrdUstarting at 0 24 and 48 h Cells were collected at 24 48 and72 h of culture and BrdU incorporation was measuredcolorimetrically by ELISA according to manufacturerrsquosinstructions (Proliferation ELISA Roche)
Human tissue samples
Formalin-fixed paraffin-embedded human tissue sampleswere obtained from diagnostic biopsies or resection
specimens from patients at the Hospital Barros LucoTrudeau (Santiago Chile) The tissues were used inagreement with the ethical guidelines approved by theethical committee of the hospital and our institutions
RNA and DNA isolation
Total RNA from cells was extracted with TRIzol(Invitrogen) as described before (121325) The polyA+
fraction was obtained with the Oligotex mRNA Midi kit(Qiagen) according to the manufacturerrsquos instructionsTo eliminate mtDNA contamination RNA preparationswere treated with TURBO DNA-free (Ambion) accordingto the manufacturerrsquos instructions Human mtDNA wasprepared from human lymphocytes HeLa SiHa Hep G2and HL-60 cells (26)
RT-PCR
Reverse transcription was carried out with 50ndash100 ng offreshly prepared RNA 50 ng of random hexamers orsequence-specific primers and 200 U of reverse transcrip-tase (M-MLV or SuperScript II Invitrogen) (1213) ThecDNA was amplified by PCR and analyzed by electro-phoresis as described before (121327) The primers usedto amplify the sense 16S mtrRNA were P1 (r) 50 AAGGTGGAGTGGGTTTGGGGC (position 11ndash31) P9 (f)50 TAGGCCTAAAAGCAGCCACCAA (position501ndash522) P10 (f) 50 ACCGTGCAAAGGTAGCATAATCAC (position 912ndash935) P11 (r) 50 AATAGGATTGCGCTGTTATCCCTA (position 1260ndash1283) P12 (r) 50 CTGTTCTTGGGTGGGTGTG (position 1536ndash1554) For theantisense 16S mtrRNA P2 (f) 50 GGGGTCTTAGCTTTGGCTCTCC (position 1326ndash1347) P3 (f) 50 TTGGTGGCTGCTTTTAGGCCTA (position 1207ndash1227) P4 (f)50 GGTTGATTGTAGATATTGGGCT (position 833ndash854)P5 (f) 50 GGTAAGATTTGCCGAGTTC (position741ndash759) P6 (f) 50 GTGATTATGCTACCTTTGCACGGT (position 626ndash649) P7 (r) 50 ACCATTTACCCAAATAAAGTATAG (position 1483ndash1506) P8 (r) 50 GGACCAATCTATCACCCTATA (position 942ndash962) To iden-tify the position of each primer the sequence from nt 1 to1559 of the sense or antisense 16S mtRNA was used asreference For the region between the IR and the 16SmtrRNA P13 (r) 50 AGGTTTAGCCAAACCATT(807ndash824) For the 12S mtrRNA P14 (f) 50 AGCCTATATACCGCCATCTTC (position 604ndash624) P15 (r) 50 AAGTATACTTGAGGAGGGTGA (position 843ndash863)P16 (f) 50 GTGTACTGGAAAGTGCACTTG (position927ndash947) For COX I P17 (f) 50 GAACAGGTTGAACAGTCTACCCT (position 371ndash393) and P18 (r) 50 TTCCGAAGCCTGGTAGGATAAGA (position 738ndash760)For the 18S rRNA P19 (f) 50 GATGCGTGCATTTATCAGATC (position 309ndash329) and P20 (r) 50 AGTGGACTCATTCCAATTACA (position 652ndash672) For GAPDHP21 (f) 50 ACTCTGGTAAAGTGGATATTGT (position131ndash152) and P22 (r) 50 ATGATGTTCTGGAGAGCCC(position 662ndash680) For b-actin mRNA P23 (f)50 AAGAGAGGCATCCTCACCCTG (position 181ndash202)P24 (r) 50 GGCGACGTAGCACAGCTTCTCC (position639ndash660) (f) and (r) represent forward and reverseprimers respectively The sequence of the primers for
Nucleic Acids Research 2007 Vol 35 No 21 7337
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
the 16S 12S and COX 1 transcripts were deduced from thehuman mtDNA (GenBank Accession No V00662)The sequence of the primers for the 18S rRNA GAPDHmRNA and b-actin mRNA were deduced from theGenBank Accession Nos M10098 M33197 andNM_001101 respectively Amplified DNA fragmentswere purified (Wizard SU Gel and PCR Clean-up systemPromega) cloned in pGEM-T Easy (Promega) or pTOPO(Invitrogen) and the purified recombinant plasmids weresequenced as described before (1213)
RNase digestion
About 1 mg of total RNA from human cells treated withTURBO DNA-free in 50 ml of 2 SSC (28) was incubatedwith RNase A at a final concentration of 50 mgml for15min at 258C The digestion products were extractedwith phenolndashchloroform and after adding 10 mg ofglycogen to the aqueous phase the RNA was precipitatedwith ethanol and recovered by centrifugation (1213)
S1 protection assay
S1 protection assay was carried out using a single-stranded digoxigenin-labeled DNA probe complementaryto the region between the IR and the 16S mtrRNA of thencmtRNA First an amplicon was obtained by RT-PCRusing primers 1 and 2 and this fragment was used as atemplate in an asymmetric PCR reaction that containedonly primer 1 and digoxigenin-11-dUTP (Roche Non-Radioactive ISH) S1 protection was carried out asdescribed (27) using 50 mg RNA and 50 ng digoxigenin-labeled probe After S1 digestion reactions were ethanol-precipitated and analyzed on native 25 agarose gelswhich were transferred onto HybondTM-XL membranes(Amersham) After blocking with 002 Tween 20 plus5 skim milk in PBS the position of the digoxigenin-labeled probe was revealed with anti-digoxigenin antibodyconjugated with alkaline phosphatase (Roche) and devel-oping in BCIPNBT alkaline phosphatase substratemixture (DAKO) for 10min
5rsquoRACE
HeLa cells RNA dissolved in 2 SSC was digested withRNase A and the double-stranded structure was recoveredafter phenol extraction as described before The cDNAwas synthesized with primer P8 (Figure 2a) and tailed atthe 30 end with dCTP using terminal deoxynucleotidyl-transferase (TdT Promega) (29) Amplification by PCRwas carried out with the specific primer 8 (Figure 2a) andthe anchor primer provided by the manufacturer Theamplicon obtained of 250 bp was purified and bothstrands were sequenced directly
Northern blot
Total RNA of HeLa and MCF7 cells (3 mg each) wereelectrophoresed at 75V for 90min in a native 10agarose gel prepared in TAE buffer or in an agarose gelsunder denaturing conditions containing 22M formalde-hyde (27) The RNAs were transferred in 20 SSC to anylon membrane (HybondTM-XL Amersham) for 18 h
and exposed to UV light for 5min The probe used wasprimer 13 (Figure 2a) labeled with 32P Briefly in a finalvolume of 20 ml 100 ng of the oligonucleotide were mixedwith 4 ml of enzyme buffer (100mM cacodylate buffer pH68 1mM CoCl2 and 01mM DTT) 2 ml of [a-32P]-dCTP(3000Cimmol) and 10U of TdT and incubated at 378Cfor 30min The probe was purified using a Sephadex G-50spin column (Amersham) Hybridization was carried outfor 18 h at 378C with 5ml of 05M sodium phosphate pH71 2mM EDTA 7 SDS and 01 sodium pyropho-sphate (30) containing 32P-labeled primer 13 at a ratio of105 cpmcm2 of membrane surface area The blots werewashed twice for 5min at room temperature with 2 SSCplus 01 SDS and once with 05 SSC plus 01 SDSRadioactivity on the membranes was visualized with aphosphor imager (MolecularTM Imager FX PhosphorImaging System BioRad)
About 1 mg of RNA digested with RNAse A asdescribed before was subjected to electrophoresis on a15 agarose gel containing 22M formaldehyde andtransferred to a nylon membrane (27) The membrane wasprobed with an amplicon of 250 bp corresponding to thedouble-stranded region of the ncmtRNA and obtained byRT-PCR using primers 8 and 5 (Figure 2a) and[a-32P]-dCTP The probe was extracted with phenol andprecipitated with isopropanol Hybridization was carriedout with a solution containing 8 million cpmml in4 SSC 10 dextran sulfate 150 mgml yeast tRNA150 mgml herring sperm DNA 50 formamide and1Denhardtrsquos solution (27) After hybridization at 658Covernight the membrane was washed for 10min at roomtemperature with 2 SSC and 1SSC 20min at 558Cwith 05 SSC and 20min at room temperature with02 SSC Then the membrane was exposed to X-ray filmsat 708C (1227)
Mitochondria isolation
HeLa cells were grown as indicated and about 5 108 cellswere harvested and recovered by centrifugation at 300gfor 5min at 48C The cells were resuspended in about 10volumes of a hypotonic solution containing 10mM KCl015mM MgCl2 and 10mM TrisndashHCl pH 68 incubatedfor 10min on ice and dounce homogenized with the tightpestle (1331) The homogenization was monitored byphase microscopy until 70 of the cells were broken andthe mitochondrial fraction was obtained as describedbefore (31) The final mitochondria fraction was resus-pended in 2ndash3ml of 025M sucrose 2mM MgCl2 and04mM sodium phosphate buffer at pH 68 and treatedwith RNAse A at a final concentration of 50 mgml for15min at room temperature (32) The mitochondriafraction was recovered by centrifugation at 10 000g for15min suspended in 100 ml of PBS containing 100 U ofRNaseOut (Invitrogen) and mitochondrial RNA wasextracted with TRIzol as described before
ISH
Cells cultured in 8-well chamber slides (Lab-TekNUNC) for 24ndash48 h were washed three times with PBSand fixed in 4 paraformaldehyde in PBS for 10min at
7338 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
room temperature The slides were then washed threetimes with PBS for 5min and incubated with 02N HClfor 10min at room temperature Hybridization wascarried out essentially as described before (1214) Thecells were hybridized for 18 h at 378C with 200 ml of thehybridization solution containing 35 pmol of the anti-sense probe (primer 6 or 13) or the corresponding senseprobes (Figure 2a) previously labeled at the 30 end withdigoxigenin-11-dUTP (Roche Applied Science) (1214)The slides were washed with 2SSC and 1SSC for10min at room temperature 02 SSC for 30min at 378Cand finally with 02 SSC for 10min at room temperature(14) Then the cells were incubated for 2 h at roomtemperature with anti-digoxigenin antibody conjugated toalkaline phosphatase (Roche Applied Science) diluted1500 in the blocking buffer (1 BSA 03 Triton X-100in PBS) The color reaction was carried out with a BCIPNBT substrate mixture (DAKO) plus levamisol for 30min(14) Resting or PHA-stimulated human lymphocyteswere applied on silanized slides air dried fixed with 4paraformaldehyde and subjected to the same hybridiza-tion protocol For FISH after hybridization the cells wereincubated with anti-digoxigenin antibody conjugated toFITC (Roche Applied Science) For tissue samplesparaffin sections of 5 mm thick were collected in silanizedslides and deparaffinized by immersion in three consecu-tive xylene baths 10min each The sections wererehydrated with three washes in 100 90 70 and50 ethanol once in PBS and once in distilled water for10min Afterwards the sections were fixed with 4paraformaldehyde in PBS and incubated with 02N HClplus 4mgml of pepsin for 10min at room temperatureand subjected to ISH as described before
Immunocytochemistry and FISH
After ISH melanoma cells (4295) were incubated for30min in blocking buffer (1 BSA 03 Triton X-100 inPBS) and then incubated for 2 h at room temperaturewith anti-digoxigenin antibody conjugated with fluores-cein (Roche) and with anti-PCNA monoclonal antibody(DAKO) The cells were washed three times with 005Tween20 in PBS incubated for 2 h at room temperaturewith anti-mouse IgG conjugated with rhodamin washedagain and mounted with DABCO After hybridizationHeLa cells were incubated with anti-digoxigenin antibodyconjugated with rhodamin (Roche) and with anti-cyto-chrome c monoclonal antibody (Promega) or anti-endonuclease G polyclonal antibody (ChemiconInternational) After washing with 005 Tween20 inPBS the cells were incubated with either anti-mouse IgGconjugated with fluorescein or with anti-rabbit IgG con-jugated with fluorescein Fluorescence microscopy wasanalyzed with a Olympus BX51 microscope Confocalmicroscopy was performed with a LSM 5 Zeiss micro-scope equipped with a 63 objective The analysis wascarried out with the Zeiss LSM 5 Image Browser software
Fixed resting or PHA-stimulated lymphocytes wereincubated with the blocking solution and then withanti-PCNA (DAKO) or anti-Ki67 antigen (DAKO) oranti-phosphohistone H3 (Upstate) for 60min at
room temperature After washing in PBS the sectionswere incubated for 30min with anti-mouse IgG con-jugated to alkaline phosphatase (KPL) diluted 1250 in2 BSA in PBS The color reaction was developed for20min with BCIPNBT substrate mixture (DAKO)containing levamisol Previous to immunocytochemistrydeparaffinized tissue sections were incubated in coplin jarsfor 15min at 958C in Target Retrieval Solution (DAKO)diluted 110 with distilled water The incubation with anti-PCNA was as described before
RESULTS
The human ncmt RNA
As shown in Figure 1a (panel 1) ISH with probe 6targeted to the 16S mtrRNA (see Materials and Methodssection) revealed strong hybridization signals in HeLaHL-60 MCF7 DU145 H441 and Caco-3 human cellsAs expected hybridization with the control sense probe 10(see Materials and Methods section) was negative(Figure 1a panel 2) Moreover FISH with probe 6 incombination with immunocytochemistry of PCNA of themelanoma cell line 4295 revealed that each cell expressingthe transcript was also expressing PCNA (Figure 1b) Thehybridization with probe 10 was again negative(Figure 1b) As a positive control FISH was carriedwith probe 20 (see Materials and Methods section) todetect the 18S rRNA (Figure 1b)The target of probe 6 might be a transcript homologous
to the mouse ncRNA (12) Therefore to characterize thistranscript a hypothetical human mitochondrial RNA wasdeduced (Figure 2a) The transcript contains the completesequence of the 16S mtrRNA (Figure 2a red line) linked atits 50 end to a fragment of the RNA transcribed from the16S gene corresponding to the L-strand of the mtDNA orantisense 16S mtrRNA (Figure 2a blue line) Based on thisstructure primers were designed to amplify the putativetranscript by RT-PCR (Figure 2a) Primer 1 was posi-tioned at the theoretical loop between positions 11 and 31of the human sense 16S mtrRNA while the forwardprimer 2 was at the putative IR corresponding to position1326ndash1347 of the antisense 16S mtrRNA (Figure 2a)RT-PCR carried out with RNA from HeLa HL-60 andMCF7 cells yielded a single amplicon of 210 bp(Figure 2c odd lanes) No amplification was obtainedwhen reverse transcriptase was omitted from the reaction(Figure 2c even lanes) The sequence of the amplicon ofeach cell line revealed that an IR of 184 nt was linked to thefirst 31 nt at the 50 end of the 16S mtrRNA (SupplementaryFigure S1a and GenBank Accession No DQ386868)To determine whether the IR was longer than 184 nt a
PCR-walking strategy was utilized The cDNA fromHeLa cells was amplified between primer 1 and primers3 4 5 and 6 targeted to a putative longer IR (Figure 2a)Amplification fragments of 500 700 and 800 bp wereobtained when primer 1 was used in combination withprimers 3 4 and 5 respectively (Figure 2d lanes 1 3 and 5)No amplification product was obtained with primers 1 and6 (Figure 2d lanes 7 and 8) suggesting that the 50 of theIR is positioned between primers 5 and 6 (see below)
Nucleic Acids Research 2007 Vol 35 No 21 7339
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
The sequence of the amplicon of 800 bp revealed an IR of769 nt linked to the first 31 nt of the 16S mtrRNA(Supplementary Figure S1b and GenBank Access NoDQ386868) The sequence of the region between the 16SmtrRNA and the IR is identical to that found in the sameregion of the amplicon of 215 bp (compare SupplementaryFigures S1a and b) confirming that in both cases we wereamplifying the same transcript The sequences of the 500and 700 bp fragments indicated that they were part of the800 bp amplicon The same results and sequences wereobtained with RNA from HL-60 cells PHA-stimulatedlymphocytes and MCF7 cells (data not shown)
The stem of the ncmtRNA is resistant to RNase digestion
The 769 nt of the IR are fully complementary to the16S mtrRNA Therefore the transcript should contain
HeLa HL-60 MCF7C1
DU145 H441 Caco-2
(a)
1
1 1
2
2
2
2
1
1
2
Probe 6(b)
Probe 10
PCNA
Merge
18S r RNA
1
2
B1A1
A2
A3
Figure 1 Expression of the hypothetical human ncmtRNA (a) ISH ofthe indicated cells with probe 6 (panels 1) or the control sense probe 10(panels 2) Note cytoplasmic and perinuclear hybridization signals(100) (b) Melanoma cell line expressing the transcript are alsoexpressing PCNA The cells were subjected to ISH with probe 6 orprobe 10 together with immunocytochemistry of PCNA Themerge figure confirmed the co-expression of both molecules As apositive control the same cells were subjected to ISH for the 18SrRNA and immunocytochemistry of PCNA (63)
(d)
(c)
RT + minus + minus + minus
MM 1 2 3 4 5 6HeLa HL-60 MCF7
MM
RT + minus + minus + minus + minus
1 2 3 4 5 6
210 bp
700 bp 800 bp
2 3 4
1
65
7
11
89
63prime
2 3 4
1
65
7
11
8 10
63prime
RNas e A
(a)
12
13
13
500 bp
7 8
600 bp
10
Primers 1-3 1-4 1-5 1-6
9(b)
2 3 4 5 61 MM
800 bp 750 bp
370 bpRNase A minus + minus + minus +
(e)
500 bp
(f) GAPDH12S 18S
M1 2 3 4 5 6
RNase A minus + minus + minus +
500 bp
HL-
60M
CF
7
HeL
a
872 bp
603 bp
1078 bpg)
Figure 2 The human ncmtRNA (a) Theoretical structure of thehuman ncmtRNA The 50 end of the sense 16S mtrRNA (red line) islinked to a fragment of the antisense 16S mtrRNA or IR (blue line)forming a double-stranded structure and a loop of unknown lengthThe position of the reverse primers (under the lines) and the forwardprimers (over the lines) are indicated (see Materials and Methodssection) (b) Digestion of the loop and the single-stranded region of thencmtRNA by RNase A is also illustrated (c) Amplification of thecDNA obtained from three tumor cell lines using primers 1 and 2 Anamplicon of 210 bp was obtained only when the reaction was carriedout with reverse transcriptase (c odd lanes) (d) Amplification of thecDNA of HeLa cells by PCR using primers 1 in combination withprimers 3 (lanes 1 and 2) 4 (lanes 3 and 4) 5 (lanes 5 and 6) and 6(lanes 7 and 8) respectively Amplicons of 500 700 and 800 bp wereobtained No amplification products were generated with primer 1 and6 (lanes 7 and 8) or without reverse transcriptase (d even lanes) M100 bp ladder (e) RNA from HeLa cells in 2 SSC was incubatedwithout (odd lanes) or with 50 mgml of RNase A for 15min at 258C(even lanes) The RNA was recovered and amplified by RT-PCR usingprimers 1 and 5 (lanes 1 and 2) primers 10 and 11 (lanes 3 and 4) orwith primers 7 and 5 (lanes 5 and 6) Amplicons of 800 and 350 bp wereobtained with primer 1 and 5 and 10 and 11 respectively only withuntreated RNA (lanes 1 and 3 respectively) Amplification of thefragment of 750 bp obtained with primers 7 and 5 was not affected bythe nuclease treatment (lanes 5 and 6) (f) Amplification of the 12SmtrRNA (lanes 1 and 2) 18S rRNA (lanes 3 and 4) and GAPDHmRNA (lanes 5 and 6) after digestion with RNase A (mockexperiment odd lanes) (g) About 1 mg of RNA from the indicatedcell lines was digested with RNase A and the digestion products wereresolved by electrophoresis on a 15 agarose gel After blotting themembrane was probed with a 32P-labeled PCR fragment targeted to thedouble-stranded region of the ncmtRNA (see Materials Methodssection) A single hybridization band corresponding to a transcript of800 nt was detected
7340 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

the 16S 12S and COX 1 transcripts were deduced from thehuman mtDNA (GenBank Accession No V00662)The sequence of the primers for the 18S rRNA GAPDHmRNA and b-actin mRNA were deduced from theGenBank Accession Nos M10098 M33197 andNM_001101 respectively Amplified DNA fragmentswere purified (Wizard SU Gel and PCR Clean-up systemPromega) cloned in pGEM-T Easy (Promega) or pTOPO(Invitrogen) and the purified recombinant plasmids weresequenced as described before (1213)
RNase digestion
About 1 mg of total RNA from human cells treated withTURBO DNA-free in 50 ml of 2 SSC (28) was incubatedwith RNase A at a final concentration of 50 mgml for15min at 258C The digestion products were extractedwith phenolndashchloroform and after adding 10 mg ofglycogen to the aqueous phase the RNA was precipitatedwith ethanol and recovered by centrifugation (1213)
S1 protection assay
S1 protection assay was carried out using a single-stranded digoxigenin-labeled DNA probe complementaryto the region between the IR and the 16S mtrRNA of thencmtRNA First an amplicon was obtained by RT-PCRusing primers 1 and 2 and this fragment was used as atemplate in an asymmetric PCR reaction that containedonly primer 1 and digoxigenin-11-dUTP (Roche Non-Radioactive ISH) S1 protection was carried out asdescribed (27) using 50 mg RNA and 50 ng digoxigenin-labeled probe After S1 digestion reactions were ethanol-precipitated and analyzed on native 25 agarose gelswhich were transferred onto HybondTM-XL membranes(Amersham) After blocking with 002 Tween 20 plus5 skim milk in PBS the position of the digoxigenin-labeled probe was revealed with anti-digoxigenin antibodyconjugated with alkaline phosphatase (Roche) and devel-oping in BCIPNBT alkaline phosphatase substratemixture (DAKO) for 10min
5rsquoRACE
HeLa cells RNA dissolved in 2 SSC was digested withRNase A and the double-stranded structure was recoveredafter phenol extraction as described before The cDNAwas synthesized with primer P8 (Figure 2a) and tailed atthe 30 end with dCTP using terminal deoxynucleotidyl-transferase (TdT Promega) (29) Amplification by PCRwas carried out with the specific primer 8 (Figure 2a) andthe anchor primer provided by the manufacturer Theamplicon obtained of 250 bp was purified and bothstrands were sequenced directly
Northern blot
Total RNA of HeLa and MCF7 cells (3 mg each) wereelectrophoresed at 75V for 90min in a native 10agarose gel prepared in TAE buffer or in an agarose gelsunder denaturing conditions containing 22M formalde-hyde (27) The RNAs were transferred in 20 SSC to anylon membrane (HybondTM-XL Amersham) for 18 h
and exposed to UV light for 5min The probe used wasprimer 13 (Figure 2a) labeled with 32P Briefly in a finalvolume of 20 ml 100 ng of the oligonucleotide were mixedwith 4 ml of enzyme buffer (100mM cacodylate buffer pH68 1mM CoCl2 and 01mM DTT) 2 ml of [a-32P]-dCTP(3000Cimmol) and 10U of TdT and incubated at 378Cfor 30min The probe was purified using a Sephadex G-50spin column (Amersham) Hybridization was carried outfor 18 h at 378C with 5ml of 05M sodium phosphate pH71 2mM EDTA 7 SDS and 01 sodium pyropho-sphate (30) containing 32P-labeled primer 13 at a ratio of105 cpmcm2 of membrane surface area The blots werewashed twice for 5min at room temperature with 2 SSCplus 01 SDS and once with 05 SSC plus 01 SDSRadioactivity on the membranes was visualized with aphosphor imager (MolecularTM Imager FX PhosphorImaging System BioRad)
About 1 mg of RNA digested with RNAse A asdescribed before was subjected to electrophoresis on a15 agarose gel containing 22M formaldehyde andtransferred to a nylon membrane (27) The membrane wasprobed with an amplicon of 250 bp corresponding to thedouble-stranded region of the ncmtRNA and obtained byRT-PCR using primers 8 and 5 (Figure 2a) and[a-32P]-dCTP The probe was extracted with phenol andprecipitated with isopropanol Hybridization was carriedout with a solution containing 8 million cpmml in4 SSC 10 dextran sulfate 150 mgml yeast tRNA150 mgml herring sperm DNA 50 formamide and1Denhardtrsquos solution (27) After hybridization at 658Covernight the membrane was washed for 10min at roomtemperature with 2 SSC and 1SSC 20min at 558Cwith 05 SSC and 20min at room temperature with02 SSC Then the membrane was exposed to X-ray filmsat 708C (1227)
Mitochondria isolation
HeLa cells were grown as indicated and about 5 108 cellswere harvested and recovered by centrifugation at 300gfor 5min at 48C The cells were resuspended in about 10volumes of a hypotonic solution containing 10mM KCl015mM MgCl2 and 10mM TrisndashHCl pH 68 incubatedfor 10min on ice and dounce homogenized with the tightpestle (1331) The homogenization was monitored byphase microscopy until 70 of the cells were broken andthe mitochondrial fraction was obtained as describedbefore (31) The final mitochondria fraction was resus-pended in 2ndash3ml of 025M sucrose 2mM MgCl2 and04mM sodium phosphate buffer at pH 68 and treatedwith RNAse A at a final concentration of 50 mgml for15min at room temperature (32) The mitochondriafraction was recovered by centrifugation at 10 000g for15min suspended in 100 ml of PBS containing 100 U ofRNaseOut (Invitrogen) and mitochondrial RNA wasextracted with TRIzol as described before
ISH
Cells cultured in 8-well chamber slides (Lab-TekNUNC) for 24ndash48 h were washed three times with PBSand fixed in 4 paraformaldehyde in PBS for 10min at
7338 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
room temperature The slides were then washed threetimes with PBS for 5min and incubated with 02N HClfor 10min at room temperature Hybridization wascarried out essentially as described before (1214) Thecells were hybridized for 18 h at 378C with 200 ml of thehybridization solution containing 35 pmol of the anti-sense probe (primer 6 or 13) or the corresponding senseprobes (Figure 2a) previously labeled at the 30 end withdigoxigenin-11-dUTP (Roche Applied Science) (1214)The slides were washed with 2SSC and 1SSC for10min at room temperature 02 SSC for 30min at 378Cand finally with 02 SSC for 10min at room temperature(14) Then the cells were incubated for 2 h at roomtemperature with anti-digoxigenin antibody conjugated toalkaline phosphatase (Roche Applied Science) diluted1500 in the blocking buffer (1 BSA 03 Triton X-100in PBS) The color reaction was carried out with a BCIPNBT substrate mixture (DAKO) plus levamisol for 30min(14) Resting or PHA-stimulated human lymphocyteswere applied on silanized slides air dried fixed with 4paraformaldehyde and subjected to the same hybridiza-tion protocol For FISH after hybridization the cells wereincubated with anti-digoxigenin antibody conjugated toFITC (Roche Applied Science) For tissue samplesparaffin sections of 5 mm thick were collected in silanizedslides and deparaffinized by immersion in three consecu-tive xylene baths 10min each The sections wererehydrated with three washes in 100 90 70 and50 ethanol once in PBS and once in distilled water for10min Afterwards the sections were fixed with 4paraformaldehyde in PBS and incubated with 02N HClplus 4mgml of pepsin for 10min at room temperatureand subjected to ISH as described before
Immunocytochemistry and FISH
After ISH melanoma cells (4295) were incubated for30min in blocking buffer (1 BSA 03 Triton X-100 inPBS) and then incubated for 2 h at room temperaturewith anti-digoxigenin antibody conjugated with fluores-cein (Roche) and with anti-PCNA monoclonal antibody(DAKO) The cells were washed three times with 005Tween20 in PBS incubated for 2 h at room temperaturewith anti-mouse IgG conjugated with rhodamin washedagain and mounted with DABCO After hybridizationHeLa cells were incubated with anti-digoxigenin antibodyconjugated with rhodamin (Roche) and with anti-cyto-chrome c monoclonal antibody (Promega) or anti-endonuclease G polyclonal antibody (ChemiconInternational) After washing with 005 Tween20 inPBS the cells were incubated with either anti-mouse IgGconjugated with fluorescein or with anti-rabbit IgG con-jugated with fluorescein Fluorescence microscopy wasanalyzed with a Olympus BX51 microscope Confocalmicroscopy was performed with a LSM 5 Zeiss micro-scope equipped with a 63 objective The analysis wascarried out with the Zeiss LSM 5 Image Browser software
Fixed resting or PHA-stimulated lymphocytes wereincubated with the blocking solution and then withanti-PCNA (DAKO) or anti-Ki67 antigen (DAKO) oranti-phosphohistone H3 (Upstate) for 60min at
room temperature After washing in PBS the sectionswere incubated for 30min with anti-mouse IgG con-jugated to alkaline phosphatase (KPL) diluted 1250 in2 BSA in PBS The color reaction was developed for20min with BCIPNBT substrate mixture (DAKO)containing levamisol Previous to immunocytochemistrydeparaffinized tissue sections were incubated in coplin jarsfor 15min at 958C in Target Retrieval Solution (DAKO)diluted 110 with distilled water The incubation with anti-PCNA was as described before
RESULTS
The human ncmt RNA
As shown in Figure 1a (panel 1) ISH with probe 6targeted to the 16S mtrRNA (see Materials and Methodssection) revealed strong hybridization signals in HeLaHL-60 MCF7 DU145 H441 and Caco-3 human cellsAs expected hybridization with the control sense probe 10(see Materials and Methods section) was negative(Figure 1a panel 2) Moreover FISH with probe 6 incombination with immunocytochemistry of PCNA of themelanoma cell line 4295 revealed that each cell expressingthe transcript was also expressing PCNA (Figure 1b) Thehybridization with probe 10 was again negative(Figure 1b) As a positive control FISH was carriedwith probe 20 (see Materials and Methods section) todetect the 18S rRNA (Figure 1b)The target of probe 6 might be a transcript homologous
to the mouse ncRNA (12) Therefore to characterize thistranscript a hypothetical human mitochondrial RNA wasdeduced (Figure 2a) The transcript contains the completesequence of the 16S mtrRNA (Figure 2a red line) linked atits 50 end to a fragment of the RNA transcribed from the16S gene corresponding to the L-strand of the mtDNA orantisense 16S mtrRNA (Figure 2a blue line) Based on thisstructure primers were designed to amplify the putativetranscript by RT-PCR (Figure 2a) Primer 1 was posi-tioned at the theoretical loop between positions 11 and 31of the human sense 16S mtrRNA while the forwardprimer 2 was at the putative IR corresponding to position1326ndash1347 of the antisense 16S mtrRNA (Figure 2a)RT-PCR carried out with RNA from HeLa HL-60 andMCF7 cells yielded a single amplicon of 210 bp(Figure 2c odd lanes) No amplification was obtainedwhen reverse transcriptase was omitted from the reaction(Figure 2c even lanes) The sequence of the amplicon ofeach cell line revealed that an IR of 184 nt was linked to thefirst 31 nt at the 50 end of the 16S mtrRNA (SupplementaryFigure S1a and GenBank Accession No DQ386868)To determine whether the IR was longer than 184 nt a
PCR-walking strategy was utilized The cDNA fromHeLa cells was amplified between primer 1 and primers3 4 5 and 6 targeted to a putative longer IR (Figure 2a)Amplification fragments of 500 700 and 800 bp wereobtained when primer 1 was used in combination withprimers 3 4 and 5 respectively (Figure 2d lanes 1 3 and 5)No amplification product was obtained with primers 1 and6 (Figure 2d lanes 7 and 8) suggesting that the 50 of theIR is positioned between primers 5 and 6 (see below)
Nucleic Acids Research 2007 Vol 35 No 21 7339
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
The sequence of the amplicon of 800 bp revealed an IR of769 nt linked to the first 31 nt of the 16S mtrRNA(Supplementary Figure S1b and GenBank Access NoDQ386868) The sequence of the region between the 16SmtrRNA and the IR is identical to that found in the sameregion of the amplicon of 215 bp (compare SupplementaryFigures S1a and b) confirming that in both cases we wereamplifying the same transcript The sequences of the 500and 700 bp fragments indicated that they were part of the800 bp amplicon The same results and sequences wereobtained with RNA from HL-60 cells PHA-stimulatedlymphocytes and MCF7 cells (data not shown)
The stem of the ncmtRNA is resistant to RNase digestion
The 769 nt of the IR are fully complementary to the16S mtrRNA Therefore the transcript should contain
HeLa HL-60 MCF7C1
DU145 H441 Caco-2
(a)
1
1 1
2
2
2
2
1
1
2
Probe 6(b)
Probe 10
PCNA
Merge
18S r RNA
1
2
B1A1
A2
A3
Figure 1 Expression of the hypothetical human ncmtRNA (a) ISH ofthe indicated cells with probe 6 (panels 1) or the control sense probe 10(panels 2) Note cytoplasmic and perinuclear hybridization signals(100) (b) Melanoma cell line expressing the transcript are alsoexpressing PCNA The cells were subjected to ISH with probe 6 orprobe 10 together with immunocytochemistry of PCNA Themerge figure confirmed the co-expression of both molecules As apositive control the same cells were subjected to ISH for the 18SrRNA and immunocytochemistry of PCNA (63)
(d)
(c)
RT + minus + minus + minus
MM 1 2 3 4 5 6HeLa HL-60 MCF7
MM
RT + minus + minus + minus + minus
1 2 3 4 5 6
210 bp
700 bp 800 bp
2 3 4
1
65
7
11
89
63prime
2 3 4
1
65
7
11
8 10
63prime
RNas e A
(a)
12
13
13
500 bp
7 8
600 bp
10
Primers 1-3 1-4 1-5 1-6
9(b)
2 3 4 5 61 MM
800 bp 750 bp
370 bpRNase A minus + minus + minus +
(e)
500 bp
(f) GAPDH12S 18S
M1 2 3 4 5 6
RNase A minus + minus + minus +
500 bp
HL-
60M
CF
7
HeL
a
872 bp
603 bp
1078 bpg)
Figure 2 The human ncmtRNA (a) Theoretical structure of thehuman ncmtRNA The 50 end of the sense 16S mtrRNA (red line) islinked to a fragment of the antisense 16S mtrRNA or IR (blue line)forming a double-stranded structure and a loop of unknown lengthThe position of the reverse primers (under the lines) and the forwardprimers (over the lines) are indicated (see Materials and Methodssection) (b) Digestion of the loop and the single-stranded region of thencmtRNA by RNase A is also illustrated (c) Amplification of thecDNA obtained from three tumor cell lines using primers 1 and 2 Anamplicon of 210 bp was obtained only when the reaction was carriedout with reverse transcriptase (c odd lanes) (d) Amplification of thecDNA of HeLa cells by PCR using primers 1 in combination withprimers 3 (lanes 1 and 2) 4 (lanes 3 and 4) 5 (lanes 5 and 6) and 6(lanes 7 and 8) respectively Amplicons of 500 700 and 800 bp wereobtained No amplification products were generated with primer 1 and6 (lanes 7 and 8) or without reverse transcriptase (d even lanes) M100 bp ladder (e) RNA from HeLa cells in 2 SSC was incubatedwithout (odd lanes) or with 50 mgml of RNase A for 15min at 258C(even lanes) The RNA was recovered and amplified by RT-PCR usingprimers 1 and 5 (lanes 1 and 2) primers 10 and 11 (lanes 3 and 4) orwith primers 7 and 5 (lanes 5 and 6) Amplicons of 800 and 350 bp wereobtained with primer 1 and 5 and 10 and 11 respectively only withuntreated RNA (lanes 1 and 3 respectively) Amplification of thefragment of 750 bp obtained with primers 7 and 5 was not affected bythe nuclease treatment (lanes 5 and 6) (f) Amplification of the 12SmtrRNA (lanes 1 and 2) 18S rRNA (lanes 3 and 4) and GAPDHmRNA (lanes 5 and 6) after digestion with RNase A (mockexperiment odd lanes) (g) About 1 mg of RNA from the indicatedcell lines was digested with RNase A and the digestion products wereresolved by electrophoresis on a 15 agarose gel After blotting themembrane was probed with a 32P-labeled PCR fragment targeted to thedouble-stranded region of the ncmtRNA (see Materials Methodssection) A single hybridization band corresponding to a transcript of800 nt was detected
7340 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

room temperature The slides were then washed threetimes with PBS for 5min and incubated with 02N HClfor 10min at room temperature Hybridization wascarried out essentially as described before (1214) Thecells were hybridized for 18 h at 378C with 200 ml of thehybridization solution containing 35 pmol of the anti-sense probe (primer 6 or 13) or the corresponding senseprobes (Figure 2a) previously labeled at the 30 end withdigoxigenin-11-dUTP (Roche Applied Science) (1214)The slides were washed with 2SSC and 1SSC for10min at room temperature 02 SSC for 30min at 378Cand finally with 02 SSC for 10min at room temperature(14) Then the cells were incubated for 2 h at roomtemperature with anti-digoxigenin antibody conjugated toalkaline phosphatase (Roche Applied Science) diluted1500 in the blocking buffer (1 BSA 03 Triton X-100in PBS) The color reaction was carried out with a BCIPNBT substrate mixture (DAKO) plus levamisol for 30min(14) Resting or PHA-stimulated human lymphocyteswere applied on silanized slides air dried fixed with 4paraformaldehyde and subjected to the same hybridiza-tion protocol For FISH after hybridization the cells wereincubated with anti-digoxigenin antibody conjugated toFITC (Roche Applied Science) For tissue samplesparaffin sections of 5 mm thick were collected in silanizedslides and deparaffinized by immersion in three consecu-tive xylene baths 10min each The sections wererehydrated with three washes in 100 90 70 and50 ethanol once in PBS and once in distilled water for10min Afterwards the sections were fixed with 4paraformaldehyde in PBS and incubated with 02N HClplus 4mgml of pepsin for 10min at room temperatureand subjected to ISH as described before
Immunocytochemistry and FISH
After ISH melanoma cells (4295) were incubated for30min in blocking buffer (1 BSA 03 Triton X-100 inPBS) and then incubated for 2 h at room temperaturewith anti-digoxigenin antibody conjugated with fluores-cein (Roche) and with anti-PCNA monoclonal antibody(DAKO) The cells were washed three times with 005Tween20 in PBS incubated for 2 h at room temperaturewith anti-mouse IgG conjugated with rhodamin washedagain and mounted with DABCO After hybridizationHeLa cells were incubated with anti-digoxigenin antibodyconjugated with rhodamin (Roche) and with anti-cyto-chrome c monoclonal antibody (Promega) or anti-endonuclease G polyclonal antibody (ChemiconInternational) After washing with 005 Tween20 inPBS the cells were incubated with either anti-mouse IgGconjugated with fluorescein or with anti-rabbit IgG con-jugated with fluorescein Fluorescence microscopy wasanalyzed with a Olympus BX51 microscope Confocalmicroscopy was performed with a LSM 5 Zeiss micro-scope equipped with a 63 objective The analysis wascarried out with the Zeiss LSM 5 Image Browser software
Fixed resting or PHA-stimulated lymphocytes wereincubated with the blocking solution and then withanti-PCNA (DAKO) or anti-Ki67 antigen (DAKO) oranti-phosphohistone H3 (Upstate) for 60min at
room temperature After washing in PBS the sectionswere incubated for 30min with anti-mouse IgG con-jugated to alkaline phosphatase (KPL) diluted 1250 in2 BSA in PBS The color reaction was developed for20min with BCIPNBT substrate mixture (DAKO)containing levamisol Previous to immunocytochemistrydeparaffinized tissue sections were incubated in coplin jarsfor 15min at 958C in Target Retrieval Solution (DAKO)diluted 110 with distilled water The incubation with anti-PCNA was as described before
RESULTS
The human ncmt RNA
As shown in Figure 1a (panel 1) ISH with probe 6targeted to the 16S mtrRNA (see Materials and Methodssection) revealed strong hybridization signals in HeLaHL-60 MCF7 DU145 H441 and Caco-3 human cellsAs expected hybridization with the control sense probe 10(see Materials and Methods section) was negative(Figure 1a panel 2) Moreover FISH with probe 6 incombination with immunocytochemistry of PCNA of themelanoma cell line 4295 revealed that each cell expressingthe transcript was also expressing PCNA (Figure 1b) Thehybridization with probe 10 was again negative(Figure 1b) As a positive control FISH was carriedwith probe 20 (see Materials and Methods section) todetect the 18S rRNA (Figure 1b)The target of probe 6 might be a transcript homologous
to the mouse ncRNA (12) Therefore to characterize thistranscript a hypothetical human mitochondrial RNA wasdeduced (Figure 2a) The transcript contains the completesequence of the 16S mtrRNA (Figure 2a red line) linked atits 50 end to a fragment of the RNA transcribed from the16S gene corresponding to the L-strand of the mtDNA orantisense 16S mtrRNA (Figure 2a blue line) Based on thisstructure primers were designed to amplify the putativetranscript by RT-PCR (Figure 2a) Primer 1 was posi-tioned at the theoretical loop between positions 11 and 31of the human sense 16S mtrRNA while the forwardprimer 2 was at the putative IR corresponding to position1326ndash1347 of the antisense 16S mtrRNA (Figure 2a)RT-PCR carried out with RNA from HeLa HL-60 andMCF7 cells yielded a single amplicon of 210 bp(Figure 2c odd lanes) No amplification was obtainedwhen reverse transcriptase was omitted from the reaction(Figure 2c even lanes) The sequence of the amplicon ofeach cell line revealed that an IR of 184 nt was linked to thefirst 31 nt at the 50 end of the 16S mtrRNA (SupplementaryFigure S1a and GenBank Accession No DQ386868)To determine whether the IR was longer than 184 nt a
PCR-walking strategy was utilized The cDNA fromHeLa cells was amplified between primer 1 and primers3 4 5 and 6 targeted to a putative longer IR (Figure 2a)Amplification fragments of 500 700 and 800 bp wereobtained when primer 1 was used in combination withprimers 3 4 and 5 respectively (Figure 2d lanes 1 3 and 5)No amplification product was obtained with primers 1 and6 (Figure 2d lanes 7 and 8) suggesting that the 50 of theIR is positioned between primers 5 and 6 (see below)
Nucleic Acids Research 2007 Vol 35 No 21 7339
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
The sequence of the amplicon of 800 bp revealed an IR of769 nt linked to the first 31 nt of the 16S mtrRNA(Supplementary Figure S1b and GenBank Access NoDQ386868) The sequence of the region between the 16SmtrRNA and the IR is identical to that found in the sameregion of the amplicon of 215 bp (compare SupplementaryFigures S1a and b) confirming that in both cases we wereamplifying the same transcript The sequences of the 500and 700 bp fragments indicated that they were part of the800 bp amplicon The same results and sequences wereobtained with RNA from HL-60 cells PHA-stimulatedlymphocytes and MCF7 cells (data not shown)
The stem of the ncmtRNA is resistant to RNase digestion
The 769 nt of the IR are fully complementary to the16S mtrRNA Therefore the transcript should contain
HeLa HL-60 MCF7C1
DU145 H441 Caco-2
(a)
1
1 1
2
2
2
2
1
1
2
Probe 6(b)
Probe 10
PCNA
Merge
18S r RNA
1
2
B1A1
A2
A3
Figure 1 Expression of the hypothetical human ncmtRNA (a) ISH ofthe indicated cells with probe 6 (panels 1) or the control sense probe 10(panels 2) Note cytoplasmic and perinuclear hybridization signals(100) (b) Melanoma cell line expressing the transcript are alsoexpressing PCNA The cells were subjected to ISH with probe 6 orprobe 10 together with immunocytochemistry of PCNA Themerge figure confirmed the co-expression of both molecules As apositive control the same cells were subjected to ISH for the 18SrRNA and immunocytochemistry of PCNA (63)
(d)
(c)
RT + minus + minus + minus
MM 1 2 3 4 5 6HeLa HL-60 MCF7
MM
RT + minus + minus + minus + minus
1 2 3 4 5 6
210 bp
700 bp 800 bp
2 3 4
1
65
7
11
89
63prime
2 3 4
1
65
7
11
8 10
63prime
RNas e A
(a)
12
13
13
500 bp
7 8
600 bp
10
Primers 1-3 1-4 1-5 1-6
9(b)
2 3 4 5 61 MM
800 bp 750 bp
370 bpRNase A minus + minus + minus +
(e)
500 bp
(f) GAPDH12S 18S
M1 2 3 4 5 6
RNase A minus + minus + minus +
500 bp
HL-
60M
CF
7
HeL
a
872 bp
603 bp
1078 bpg)
Figure 2 The human ncmtRNA (a) Theoretical structure of thehuman ncmtRNA The 50 end of the sense 16S mtrRNA (red line) islinked to a fragment of the antisense 16S mtrRNA or IR (blue line)forming a double-stranded structure and a loop of unknown lengthThe position of the reverse primers (under the lines) and the forwardprimers (over the lines) are indicated (see Materials and Methodssection) (b) Digestion of the loop and the single-stranded region of thencmtRNA by RNase A is also illustrated (c) Amplification of thecDNA obtained from three tumor cell lines using primers 1 and 2 Anamplicon of 210 bp was obtained only when the reaction was carriedout with reverse transcriptase (c odd lanes) (d) Amplification of thecDNA of HeLa cells by PCR using primers 1 in combination withprimers 3 (lanes 1 and 2) 4 (lanes 3 and 4) 5 (lanes 5 and 6) and 6(lanes 7 and 8) respectively Amplicons of 500 700 and 800 bp wereobtained No amplification products were generated with primer 1 and6 (lanes 7 and 8) or without reverse transcriptase (d even lanes) M100 bp ladder (e) RNA from HeLa cells in 2 SSC was incubatedwithout (odd lanes) or with 50 mgml of RNase A for 15min at 258C(even lanes) The RNA was recovered and amplified by RT-PCR usingprimers 1 and 5 (lanes 1 and 2) primers 10 and 11 (lanes 3 and 4) orwith primers 7 and 5 (lanes 5 and 6) Amplicons of 800 and 350 bp wereobtained with primer 1 and 5 and 10 and 11 respectively only withuntreated RNA (lanes 1 and 3 respectively) Amplification of thefragment of 750 bp obtained with primers 7 and 5 was not affected bythe nuclease treatment (lanes 5 and 6) (f) Amplification of the 12SmtrRNA (lanes 1 and 2) 18S rRNA (lanes 3 and 4) and GAPDHmRNA (lanes 5 and 6) after digestion with RNase A (mockexperiment odd lanes) (g) About 1 mg of RNA from the indicatedcell lines was digested with RNase A and the digestion products wereresolved by electrophoresis on a 15 agarose gel After blotting themembrane was probed with a 32P-labeled PCR fragment targeted to thedouble-stranded region of the ncmtRNA (see Materials Methodssection) A single hybridization band corresponding to a transcript of800 nt was detected
7340 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
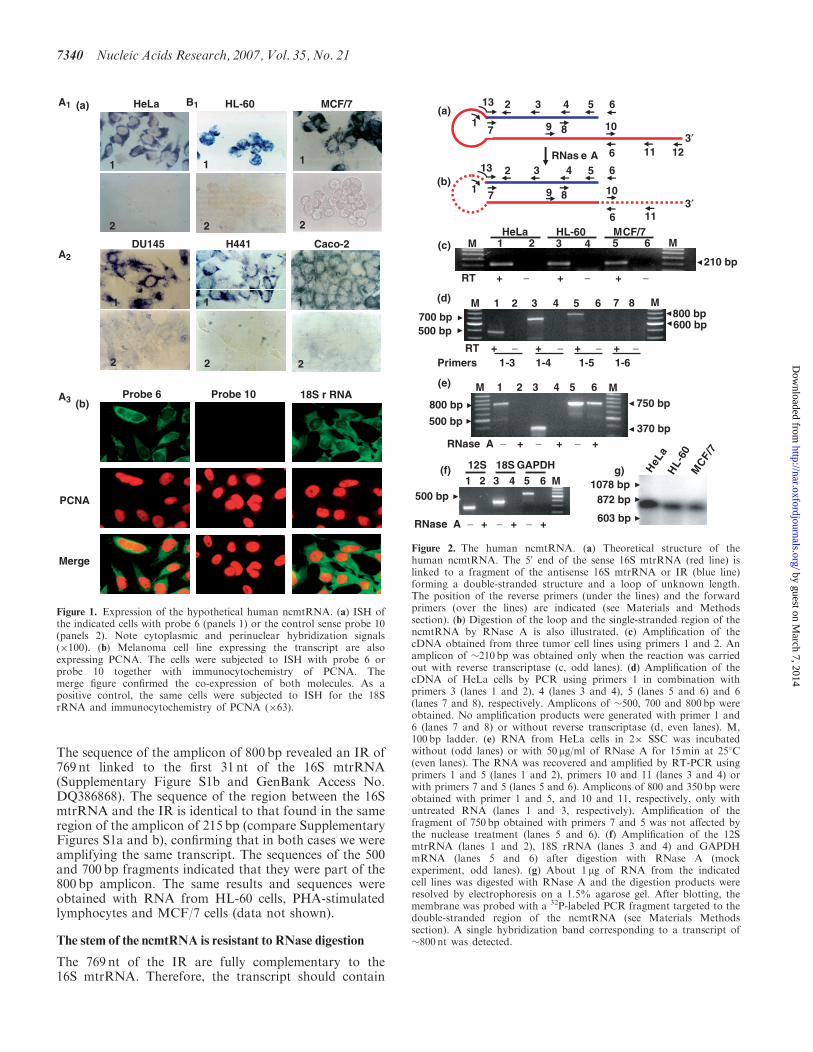
The sequence of the amplicon of 800 bp revealed an IR of769 nt linked to the first 31 nt of the 16S mtrRNA(Supplementary Figure S1b and GenBank Access NoDQ386868) The sequence of the region between the 16SmtrRNA and the IR is identical to that found in the sameregion of the amplicon of 215 bp (compare SupplementaryFigures S1a and b) confirming that in both cases we wereamplifying the same transcript The sequences of the 500and 700 bp fragments indicated that they were part of the800 bp amplicon The same results and sequences wereobtained with RNA from HL-60 cells PHA-stimulatedlymphocytes and MCF7 cells (data not shown)
The stem of the ncmtRNA is resistant to RNase digestion
The 769 nt of the IR are fully complementary to the16S mtrRNA Therefore the transcript should contain
HeLa HL-60 MCF7C1
DU145 H441 Caco-2
(a)
1
1 1
2
2
2
2
1
1
2
Probe 6(b)
Probe 10
PCNA
Merge
18S r RNA
1
2
B1A1
A2
A3
Figure 1 Expression of the hypothetical human ncmtRNA (a) ISH ofthe indicated cells with probe 6 (panels 1) or the control sense probe 10(panels 2) Note cytoplasmic and perinuclear hybridization signals(100) (b) Melanoma cell line expressing the transcript are alsoexpressing PCNA The cells were subjected to ISH with probe 6 orprobe 10 together with immunocytochemistry of PCNA Themerge figure confirmed the co-expression of both molecules As apositive control the same cells were subjected to ISH for the 18SrRNA and immunocytochemistry of PCNA (63)
(d)
(c)
RT + minus + minus + minus
MM 1 2 3 4 5 6HeLa HL-60 MCF7
MM
RT + minus + minus + minus + minus
1 2 3 4 5 6
210 bp
700 bp 800 bp
2 3 4
1
65
7
11
89
63prime
2 3 4
1
65
7
11
8 10
63prime
RNas e A
(a)
12
13
13
500 bp
7 8
600 bp
10
Primers 1-3 1-4 1-5 1-6
9(b)
2 3 4 5 61 MM
800 bp 750 bp
370 bpRNase A minus + minus + minus +
(e)
500 bp
(f) GAPDH12S 18S
M1 2 3 4 5 6
RNase A minus + minus + minus +
500 bp
HL-
60M
CF
7
HeL
a
872 bp
603 bp
1078 bpg)
Figure 2 The human ncmtRNA (a) Theoretical structure of thehuman ncmtRNA The 50 end of the sense 16S mtrRNA (red line) islinked to a fragment of the antisense 16S mtrRNA or IR (blue line)forming a double-stranded structure and a loop of unknown lengthThe position of the reverse primers (under the lines) and the forwardprimers (over the lines) are indicated (see Materials and Methodssection) (b) Digestion of the loop and the single-stranded region of thencmtRNA by RNase A is also illustrated (c) Amplification of thecDNA obtained from three tumor cell lines using primers 1 and 2 Anamplicon of 210 bp was obtained only when the reaction was carriedout with reverse transcriptase (c odd lanes) (d) Amplification of thecDNA of HeLa cells by PCR using primers 1 in combination withprimers 3 (lanes 1 and 2) 4 (lanes 3 and 4) 5 (lanes 5 and 6) and 6(lanes 7 and 8) respectively Amplicons of 500 700 and 800 bp wereobtained No amplification products were generated with primer 1 and6 (lanes 7 and 8) or without reverse transcriptase (d even lanes) M100 bp ladder (e) RNA from HeLa cells in 2 SSC was incubatedwithout (odd lanes) or with 50 mgml of RNase A for 15min at 258C(even lanes) The RNA was recovered and amplified by RT-PCR usingprimers 1 and 5 (lanes 1 and 2) primers 10 and 11 (lanes 3 and 4) orwith primers 7 and 5 (lanes 5 and 6) Amplicons of 800 and 350 bp wereobtained with primer 1 and 5 and 10 and 11 respectively only withuntreated RNA (lanes 1 and 3 respectively) Amplification of thefragment of 750 bp obtained with primers 7 and 5 was not affected bythe nuclease treatment (lanes 5 and 6) (f) Amplification of the 12SmtrRNA (lanes 1 and 2) 18S rRNA (lanes 3 and 4) and GAPDHmRNA (lanes 5 and 6) after digestion with RNase A (mockexperiment odd lanes) (g) About 1 mg of RNA from the indicatedcell lines was digested with RNase A and the digestion products wereresolved by electrophoresis on a 15 agarose gel After blotting themembrane was probed with a 32P-labeled PCR fragment targeted to thedouble-stranded region of the ncmtRNA (see Materials Methodssection) A single hybridization band corresponding to a transcript of800 nt was detected
7340 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

a double-stranded structure or stem of at least 769 bpresistant to RNase A digestion as predicted in Figure 2bOn the other hand the loop and the single-stranded regionthat extends beyond the hairpin structure should bedigested by the enzyme (Figure 2b dotted lines) To testthis possibility RNA from HeLa and HL-60 cells wasdigested with RNase A as described in Materials andMethods section The cDNA from the non-digested ordigested RNA was then amplified by PCR using theprimers described in Figure 2a The amplicon of 800 bpobtained with primers 1 and 5 was not amplified afterRNase digestion (Figure 2e lanes 1 and 2) and the samewas true for the fragment of 360 bp obtained with primers10 and 11 (Figure 2e lanes 3 and 4) indicating that thesingle-stranded regions of the transcript were digestedSimilarly the 12S mitochondrial rRNA 18S rRNA andGAPDH mRNA were not amplified after digestion(Figure 2f lanes 2 4 and 6) On the other handamplification of a fragment of 750 bp corresponding tothe double-stranded structure of the ncmtRNA andobtained using primers 7 and 5 (Figure 2a) was notaffected by RNase A digestion (Figure 2e lanes 5 and 6)The sequence of this fragment was identical to that of theamplicon of 800 bp obtained with primers 1 and 5 exceptfor the absence of the first 31 nt of the 16S mtrRNANorthern blot also confirmed the presence of the stemafter RNase A digestion A single hybridization band of800 nt was detected in digested RNA of HeLa HL-60and MCF7 cells (Figure 2g)
To determine the 50 end of the transcript using 50
RACE HeLa cell RNA was digested with RNase A andthe nuclease-resistant stem was isolated as describedbefore and used as template for 50 RACE The sequenceof the amplicon of 250 bp obtained by PCR with primer 8and the anchor primer (see Materials and Methodssection) revealed that the 769 nt of the IR were extendedby 46 additional nt (Supplementary Figure S1b under-lined sequence) indicating that the total length of the IRwas 815 nt
The 16S mtrRNA is contiguous with the IR
Similarly to the 12S and 16S mtrRNAs (33) thencmtRNA is also polyadenylated Total RNA fromHeLa HL-60 and MCF7 cells was separated intopolyA+ and polyA fractions (see Materials andMethods section) The cDNA obtained from bothfractions was then amplified with primers 1 and 2(Figure 2a) As shown in Figure 3a amplification of the215 bp fragment revealed that the transcript was enrichedin the polyA+ fraction (lanes 2 6 and 10) compared tothe polyA fraction (Figure 3a lanes 4 8 and 12) ThecDNA was synthesized from total RNA from HeLa andMCF7 cells using either oligo dT or primer 12 which iscomplementary to the 30 end of the 16S mtrRNA(Figure 2a) Then each cDNA was amplified by PCRbetween primer 1 positioned at the loop and primer 3positioned on the IR In both cases the expected ampliconof 500 bp was obtained (Figure 3b lanes 2 4 6 and 8)indicating that the cDNA primed with oligo dT or withprimer 12 comprised the complete 16S mtrRNA plus
the IR No amplification product was obtained withoutreverse transcriptase (Figure 3b odd lanes)If the IR of 815 nt is linked to the 1559 nt of the 16S
mtrRNA one would expect a transcript of 2374 nt TotalRNA from HeLa and MCF7 cells was subjected to
1 2 3 4 5 6 7 8HeLa MCF7
polyA + + minus minus + + minus minus + + minus minus
215 bp
M 1 2 3 4 5 6 11 1210987
HeLa HL-60 MCF7
M M
OligodT + + minus minus + + minus minusPrimer 1535 minus minus + + minus - + +
500 bp
RT minus + minus + minus + minus +
600 bp
HeLa
MC
18S
28S
(c)
(b)
(a)
1 2 3 4(e)
(d)
23 kbp
13 kbp10 kbp
06 kbp
MC
F7
F7
F7F7
HeLa
23 Kbp
M
16S
28S
18S16S
22 Kb
HeLa
MC
MC
HeLa
Figure 3 The IR is contiguous with the 16S mtrRNA (a) Total RNAfrom HeLa (lanes 1 to 4) HL-60 (lanes 5 to 8) and MCF7 (lanes 9 to12) cells were separated into polyA+ and polyA fractions A total of100 ng of each fraction was used to synthesize cDNA which was thenamplified by PCR using primers 1 and 2 to generate the 215 bpamplicon as indicated Odd lanes correspond to reactions carried out inthe absence of RT (b) cDNA was synthesized from the polyA+fraction of HeLa or MCF7 cells using oligo dT (lanes 1 2 5 and 6) orprimer 12 (lanes 3 4 7 and 8) A single amplicon of 500 bp wasobtained after PCR amplification of the cDNAs with primers 1 and 3only when reverse transcriptase was included in the reaction mixture(even lanes) (c) About 3 mg of HeLa and MCF7 cells RNA wasresolved by electrophoresis on a 1 native agarose gel and subjected tonorthern blot The membrane was probed with 32P-labeled primer 13(Figure 2a) The probe hybridized with a single transcript whichmigrated below the 1353 bp DNA marker (M= DNAHindIII andfDNAHaeIII) The size of this transcript deduced from the dsDNAladder corresponds to 2280 nt (d) Northern blot was carried out withRNA from the indicated cells under denaturing electrophoreticconditions In this case probe 13 hybridized with a single band thatmigrated on top of the 18S rRNA and corresponding to a transcript of2200 nt (e) For S1 protection assay an asymmetric PCR fragment 215nts containing the 31 nt of the sense 16S mtrRNA plus 184 nt of the IRwas synthesized and labeled with digoxigenin (see Materials andMethods section) After denaturation at 1008C for 5min the probe wasincubated overnight at 508C either alone (lane 2) with 20 mg of HeLaRNA (lane 3) or with 20 mg of yeast RNA (lane 4) After hybridizationthe products were digested with S1 nuclease and the products resolvedby 25 native agarose gel electrophoresis and blotted to a nylonmembrane The products of digestions were reveled with anti-digoxigenin antibody conjugated to alkaline phosphatase (seeMaterials and Methods section) Lane 1 represents the probe alonewithout treatment
Nucleic Acids Research 2007 Vol 35 No 21 7341
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
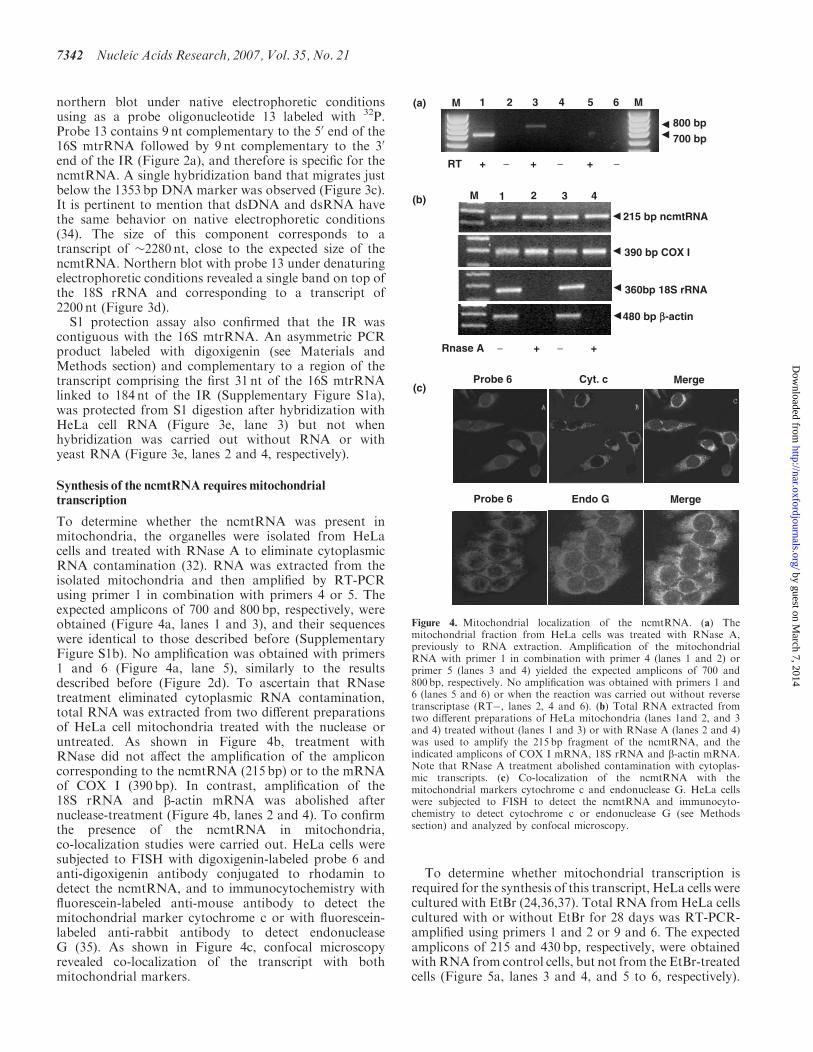
northern blot under native electrophoretic conditionsusing as a probe oligonucleotide 13 labeled with 32PProbe 13 contains 9 nt complementary to the 50 end of the16S mtrRNA followed by 9 nt complementary to the 30
end of the IR (Figure 2a) and therefore is specific for thencmtRNA A single hybridization band that migrates justbelow the 1353 bp DNA marker was observed (Figure 3c)It is pertinent to mention that dsDNA and dsRNA havethe same behavior on native electrophoretic conditions(34) The size of this component corresponds to atranscript of 2280 nt close to the expected size of thencmtRNA Northern blot with probe 13 under denaturingelectrophoretic conditions revealed a single band on top ofthe 18S rRNA and corresponding to a transcript of2200 nt (Figure 3d)S1 protection assay also confirmed that the IR was
contiguous with the 16S mtrRNA An asymmetric PCRproduct labeled with digoxigenin (see Materials andMethods section) and complementary to a region of thetranscript comprising the first 31 nt of the 16S mtrRNAlinked to 184 nt of the IR (Supplementary Figure S1a)was protected from S1 digestion after hybridization withHeLa cell RNA (Figure 3e lane 3) but not whenhybridization was carried out without RNA or withyeast RNA (Figure 3e lanes 2 and 4 respectively)
Synthesis of the ncmtRNA requires mitochondrialtranscription
To determine whether the ncmtRNA was present inmitochondria the organelles were isolated from HeLacells and treated with RNase A to eliminate cytoplasmicRNA contamination (32) RNA was extracted from theisolated mitochondria and then amplified by RT-PCRusing primer 1 in combination with primers 4 or 5 Theexpected amplicons of 700 and 800 bp respectively wereobtained (Figure 4a lanes 1 and 3) and their sequenceswere identical to those described before (SupplementaryFigure S1b) No amplification was obtained with primers1 and 6 (Figure 4a lane 5) similarly to the resultsdescribed before (Figure 2d) To ascertain that RNasetreatment eliminated cytoplasmic RNA contaminationtotal RNA was extracted from two different preparationsof HeLa cell mitochondria treated with the nuclease oruntreated As shown in Figure 4b treatment withRNase did not affect the amplification of the ampliconcorresponding to the ncmtRNA (215 bp) or to the mRNAof COX I (390 bp) In contrast amplification of the18S rRNA and b-actin mRNA was abolished afternuclease-treatment (Figure 4b lanes 2 and 4) To confirmthe presence of the ncmtRNA in mitochondriaco-localization studies were carried out HeLa cells weresubjected to FISH with digoxigenin-labeled probe 6 andanti-digoxigenin antibody conjugated to rhodamin todetect the ncmtRNA and to immunocytochemistry withfluorescein-labeled anti-mouse antibody to detect themitochondrial marker cytochrome c or with fluorescein-labeled anti-rabbit antibody to detect endonucleaseG (35) As shown in Figure 4c confocal microscopyrevealed co-localization of the transcript with bothmitochondrial markers
To determine whether mitochondrial transcription isrequired for the synthesis of this transcript HeLa cells werecultured with EtBr (243637) Total RNA from HeLa cellscultured with or without EtBr for 28 days was RT-PCR-amplified using primers 1 and 2 or 9 and 6 The expectedamplicons of 215 and 430 bp respectively were obtainedwith RNA from control cells but not from the EtBr-treatedcells (Figure 5a lanes 3 and 4 and 5 to 6 respectively)
(a)
RT + minus + minus + minus
MM 1 65432
700 bp800 bp
1 2
215 bp ncmtRNA
390 bp COX I
360bp 18S rRNA
480 bp b-actin
43M(b)
(c)Probe 6
Probe 6
Cyt c Merge
MergeEndo G
Rnase A minus + minus +
Figure 4 Mitochondrial localization of the ncmtRNA (a) Themitochondrial fraction from HeLa cells was treated with RNase Apreviously to RNA extraction Amplification of the mitochondrialRNA with primer 1 in combination with primer 4 (lanes 1 and 2) orprimer 5 (lanes 3 and 4) yielded the expected amplicons of 700 and800 bp respectively No amplification was obtained with primers 1 and6 (lanes 5 and 6) or when the reaction was carried out without reversetranscriptase (RT lanes 2 4 and 6) (b) Total RNA extracted fromtwo different preparations of HeLa mitochondria (lanes 1and 2 and 3and 4) treated without (lanes 1 and 3) or with RNase A (lanes 2 and 4)was used to amplify the 215 bp fragment of the ncmtRNA and theindicated amplicons of COX I mRNA 18S rRNA and b-actin mRNANote that RNase A treatment abolished contamination with cytoplas-mic transcripts (c) Co-localization of the ncmtRNA with themitochondrial markers cytochrome c and endonuclease G HeLa cellswere subjected to FISH to detect the ncmtRNA and immunocyto-chemistry to detect cytochrome c or endonuclease G (see Methodssection) and analyzed by confocal microscopy
7342 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
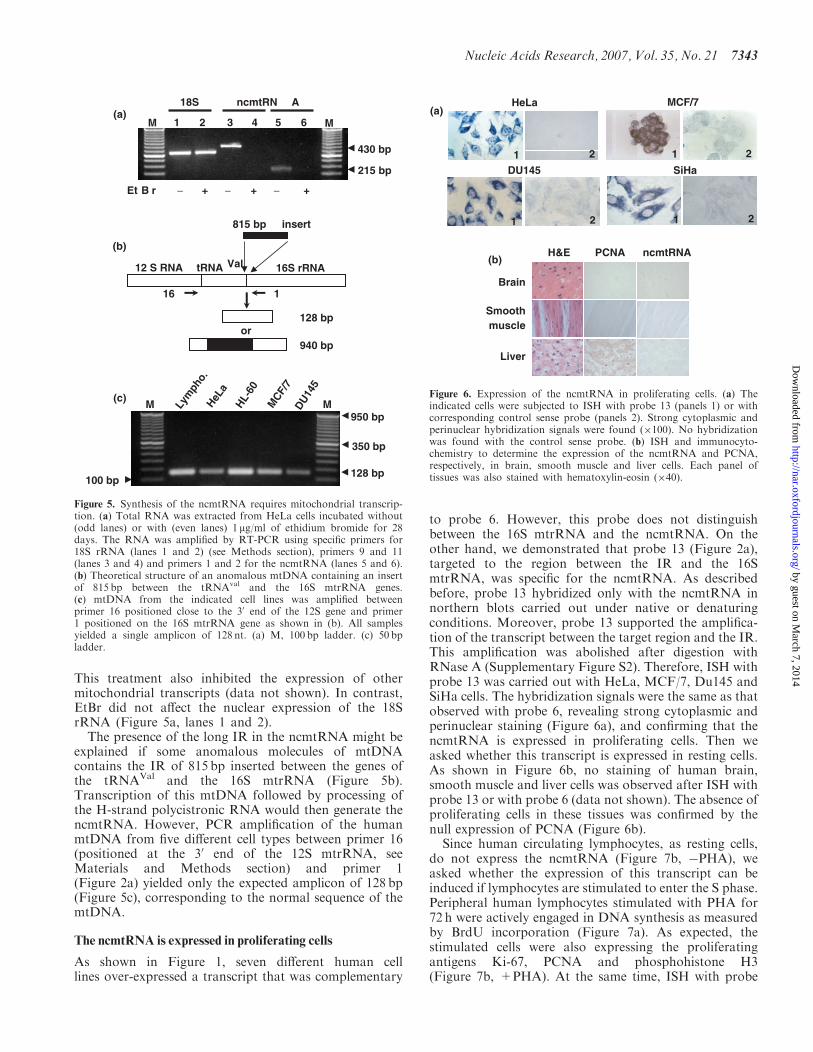
This treatment also inhibited the expression of othermitochondrial transcripts (data not shown) In contrastEtBr did not affect the nuclear expression of the 18SrRNA (Figure 5a lanes 1 and 2)
The presence of the long IR in the ncmtRNA might beexplained if some anomalous molecules of mtDNAcontains the IR of 815 bp inserted between the genes ofthe tRNAVal and the 16S mtrRNA (Figure 5b)Transcription of this mtDNA followed by processing ofthe H-strand polycistronic RNA would then generate thencmtRNA However PCR amplification of the humanmtDNA from five different cell types between primer 16(positioned at the 30 end of the 12S mtrRNA seeMaterials and Methods section) and primer 1(Figure 2a) yielded only the expected amplicon of 128 bp(Figure 5c) corresponding to the normal sequence of themtDNA
The ncmtRNA is expressed in proliferating cells
As shown in Figure 1 seven different human celllines over-expressed a transcript that was complementary
to probe 6 However this probe does not distinguishbetween the 16S mtrRNA and the ncmtRNA On theother hand we demonstrated that probe 13 (Figure 2a)targeted to the region between the IR and the 16SmtrRNA was specific for the ncmtRNA As describedbefore probe 13 hybridized only with the ncmtRNA innorthern blots carried out under native or denaturingconditions Moreover probe 13 supported the amplifica-tion of the transcript between the target region and the IRThis amplification was abolished after digestion withRNase A (Supplementary Figure S2) Therefore ISH withprobe 13 was carried out with HeLa MCF7 Du145 andSiHa cells The hybridization signals were the same as thatobserved with probe 6 revealing strong cytoplasmic andperinuclear staining (Figure 6a) and confirming that thencmtRNA is expressed in proliferating cells Then weasked whether this transcript is expressed in resting cellsAs shown in Figure 6b no staining of human brainsmooth muscle and liver cells was observed after ISH withprobe 13 or with probe 6 (data not shown) The absence ofproliferating cells in these tissues was confirmed by thenull expression of PCNA (Figure 6b)Since human circulating lymphocytes as resting cells
do not express the ncmtRNA (Figure 7b PHA) weasked whether the expression of this transcript can beinduced if lymphocytes are stimulated to enter the S phasePeripheral human lymphocytes stimulated with PHA for72 h were actively engaged in DNA synthesis as measuredby BrdU incorporation (Figure 7a) As expected thestimulated cells were also expressing the proliferatingantigens Ki-67 PCNA and phosphohistone H3(Figure 7b +PHA) At the same time ISH with probe
M M
Et B r minus + minus + minus +
18S ncmtRN A(a)
654321
430 bp
215 bp
12 S RNA tRNA Val
815 bp insert
(b)
or
16S rRNA
128 bp
940 bp
16 1
M DU14
5
Lym
pho
HeLa
HL-6
0
MCF
7
M(c)
128 bp
950 bp
350 bp
100 bp
Figure 5 Synthesis of the ncmtRNA requires mitochondrial transcrip-tion (a) Total RNA was extracted from HeLa cells incubated without(odd lanes) or with (even lanes) 1 mgml of ethidium bromide for 28days The RNA was amplified by RT-PCR using specific primers for18S rRNA (lanes 1 and 2) (see Methods section) primers 9 and 11(lanes 3 and 4) and primers 1 and 2 for the ncmtRNA (lanes 5 and 6)(b) Theoretical structure of an anomalous mtDNA containing an insertof 815 bp between the tRNAval and the 16S mtrRNA genes(c) mtDNA from the indicated cell lines was amplified betweenprimer 16 positioned close to the 30 end of the 12S gene and primer1 positioned on the 16S mtrRNA gene as shown in (b) All samplesyielded a single amplicon of 128 nt (a) M 100 bp ladder (c) 50 bpladder
Fig 6
(a)
HampE ncmtRNAPCNA
Smoothmuscle
Liver
Brain
(b)
HeLa
DU1451 2 21
21
MCF7
SiHa
1 2
Figure 6 Expression of the ncmtRNA in proliferating cells (a) Theindicated cells were subjected to ISH with probe 13 (panels 1) or withcorresponding control sense probe (panels 2) Strong cytoplasmic andperinuclear hybridization signals were found (100) No hybridizationwas found with the control sense probe (b) ISH and immunocyto-chemistry to determine the expression of the ncmtRNA and PCNArespectively in brain smooth muscle and liver cells Each panel oftissues was also stained with hematoxylin-eosin (40)
Nucleic Acids Research 2007 Vol 35 No 21 7343
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

13 revealed that the PHA-stimulated lymphocytes werealso over-expressing the ncmtRNA (Figure 7b +PHA)At low magnification hybridization of PHA-stimulatedlymphocytes with probe 13 show a strong signal in most ofthe cells (Figure 7c ncmtRNA) while no staining wasobserved with the corresponding control sense probe(Figure 7c Control sense) Similar results were obtainedwith lymphocytes obtained from 11 different healthydonors (data not shown)Next we asked whether the arrest of the cell cycle at G1
with aphidicolin would affect the expression of thencmtRNA As shown in Figure 7d treatment of DU145cells with the drug induces both a block on cellproliferation and a marked down regulation of thistranscript when compared with the untreated controlcells When the DU145 cells are removed from the drug-containing medium to normal culture medium for 2 dayscell proliferation was re-initiated and the level of expres-sion of the transcript is recovered and become similar tothe expression rate of the control cells (Figure 7d)
DISCUSSION
The ncmtRNA
This is the first report on a novel non-coding mitochon-drial RNA over-expressed in human proliferating cells butnot in resting cells Based on the hypothetical structureshown in Figure 2a we established that the humanncmtRNA contains an IR of 815 nt covalently linked tothe first 865 nt of the 16S mtrRNA To confirm theexistence of the ncmtRNA additional experiments werecarried out First northern blot with probe 13 revealeda transcript of 2280 nt (native conditions) or 2200 nt(denaturing conditions) which are close to the expectedsize of 2374 nt of the ncmtRNA Second amplification ofthe region comprising the loop and the IR of thencmtRNA was possible after using a cDNA primed witholigo dT or with a primer targeted to the 30 end of thetranscript This experiment was similar to the strategyused to establish that different regions of the long ncRNATsix were contiguous (38) Third the long double-stranded structure of 815 bp was isolated after RNase Adigestion and its presence in the digestion products wasdemonstrated by amplification between the 50 and 30 endsof the stem or by northern blot analysis Fourth S1protection assay demonstrated the presence in thencmtRNA of the sequence region between the IR andthe 16S mtrRNA Altogether these results support theconclusion that the ncmtRNA contains an IR of 815 ntcontiguous with the 16S mtrRNA
Translation in silico of the 2374 nt of the ncmtRNA(GenBank Accession No DQ386868) using either thestandard or the vertebrate mitochondrial codes yieldedseveral open reading frames corresponding to peptidesranging from 15 to 60 amino acids These results agreewith the definition of ncRNAs in which open readingframes greater than 100 amino acids are rarely present(15ndash17)
A search of the ncmtRNA was conducted in humanEST database (httpwwwncbinlmnihgovBLASTBlastcgi) Although several mitochondrial cDNA entrieswere present including the 16S mtrRNA the sequence ofthe ncmtRNA or at least part of the sequence containingthe region between the IR and the 16S mtrRNA was notfound We believe that the absence of this sequence is dueto both the double-stranded structure of the transcript andreplication slippage of the polymerases commonly used toconstruct EST libraries Escherichia coliDNA polymerasesI II and III as well as polymerases of phages T4 and T7have poor strand separation activity resulting in replicationslippage of DNA templates carrying hairpin structures(3940) As a consequence deletion mutations of hairpinsequences have been observed in vitro and in vivo (4142)Thermophilic DNA polymerases including Taq polymer-ase also exhibit replication slippage with templates contain-ing hairpin structures (43) with fateful consequences foramplification and DNA sequencing (44ndash46)
Synthesis of the ncmtRNA requires mtDNA transcription
Several results strongly suggest that the ncmtRNA issynthesized in mitochondria The sequence of the
(a)
02
06
1012
24 48 72 h
minus PHA+ PHA
Ab
sorb
ance
at45
0 n
m
+ PHA minus PHA
PCNA
Ki-67
ncmtRNA
(b)
(c) ncmtRNAControlsense
PO4-H3
04
08
Control + Aphidilcolin Recovery(d)
Figure 7 Reversibility of the ncmtRNA expression (a) Humanlymphocytes were incubated without (white bars) or with PHA(hatched bars) for 24 48 and 72 h At each time period the cellswere incubated with BrdU for 16 h and the incorporation of thenucleoside was determined by ELISA (see Materials and Methodssection) The incorporation of BrdU is expressed as the absorbanceat 450 nm (b) About 100 000 lymphocytes incubated without (PHA)or with PHA (+PHA) for 72 h were subjected to immunocytochemistryto determine the expression of Ki-67 PCNA and phospho-histoneH3 (PO4-H3) and to ISH with probe 13 to determine the expressionof the ncmtRNA (40) (c) PHA-stimulated lymphocytes for 72 h werehybridized with probe 13 (ncmtRNA) or with the correspondingsense control probe (Control sense) (20) (d) DU145 cells wereincubated twice with aphidicolin (see Methods section) andsubjected to ISH together with a control culture The blocked cellswere changed to normal medium for 48 h and then subjected to ISH(Recovery) (40)
7344 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

transcript exhibits a minimum of 999 identity withseveral haplotypes of the human mtDNA (see httpwwwncbinlmnihgovgquery) Moreover we showedthat the transcript was present in isolated mitochondriaand confocal microscopy demonstrated that thencmtRNA co-localized with cytochrome c and endonu-clease G two specific markers of the organelle (35) On theother hand treatment of HeLa cells with EtBr abolishedthe presence of the ncmtRNA as well as other mitochon-drial transcripts indicating that the synthesis of thencmtRNA requires mitochondrial transcription(243637) These results also indicate that the ncmtRNAis not a transcription product of a mitochondrialpseudogene (47) Searching for the sequence of thencmtRNA in the human genome (wwwncbinlmnihgovBLASTBlast) yielded only fragments of the16S mtrRNA (sense and antisense orientations) havingan identity of 82ndash94 with contigs corresponding toseveral human chromosomes However the sequence ofthe ncmtRNA was not found (data not shown) This resultis consistent with the results of amplification of thetranscript by RT-PCR As shown here we have neverobserved amplification between the loop and the IR whenthe reactions were carried out without reversetranscriptase
Since there is not a contiguous sequence in the mtDNAencoding the ncmtRNA it is reasonable to hypothesizethat the synthesis of this transcript involves post-transcriptional reactions Although other possibilitiescannot be discarded the synthesis of the ncmtRNAmight involve either a trans splicing reaction or a RNAligase reaction as reported in other organisms (48ndash50)Although the synthesis of the ncmtRNA warrants futurework it is interesting to mention the similarity betweenthe structures of this transcript with the presence ofdouble-stranded RNAs in HeLa cell mitochondria This isa heterogeneous fraction sedimenting from 4S to 17S andresistant to RNase A digestion (2851) Unfortunately nofurther sequence analysis of these transcripts wasreported and perhaps one of these double-strandedstructures corresponds to the ncmtRNA
Besides the ribosomal RNAs 16S and 12S and thetRNAs the ncmtRNA is not the only ncRNA present inmitochondria Recently a new family of small ncRNAswas reported to be present in isolated mouse mitochon-dria consisting of six transcripts ranging in size between23 and 68 nt (52) The authors also reported the presenceof similar molecules in isolated chloroplast of Nicotianatabacum Unfortunately no information on the biologicalfunction was provided (52)
Expression of the ncmtRNA and cell proliferation
The most striking results of the present work are the closecorrelations between expression of the ncmtRNA and cellproliferation Besides the cell lines reported here over-expression of the ncmtRNA was observed in 16 additionaltumor and normal cell lines (data not shown) In contrastthis transcript was not expressed in non-proliferating cellsincluding resting circulating lymphocytes Howeverthe expression of this transcript can be induced in
PHA-stimulated lymphocytes together with DNA synth-esis and the expression of the proliferation markers Ki-67PCNA and phosphohistone H3 (19ndash21) Moreovertreatment of DU145 cells with aphidicolin confirmed theprevious results After treatment with the drug the cellsstop proliferating as described before (2223) At thesame time the expression of the ncmtRNA was down-regulated However after removal of the drug theexpression of this transcript was re-established togetherwith cell proliferation Altogether these results stronglysuggest that the ncmtRNA is a new marker of cellproliferation However the precise function that thistranscript may play in the cell cycle remains unclear andwarrants future studiesSimilarly to the present work other ncRNAs have been
reported to be involved in cell proliferation ncRNAs areinvolved in different cellular and molecular eventsincluding differentiation and development imprintingregulation of X chromosome silencing and human diseases(17) Moreover microRNA an important group withinthe expanding family of ncRNAs (53) are also involved incell proliferation and the regulation of the cell cycleIn Drosophila melanogaster germline stem cell division isregulated by microRNAs This conclusion was reachedafter observing a reduction of the germline stem cellproliferation in the dicer-1 mutant the RNase III essentialfor microRNA production (54) Although themicroRNAs involved were not identified the authorsproposed that the function of these molecules is to inhibitDacapo which in turn negatively regulates the G1Stransition (54) In human lymphoma c-Myc up-regulatesthe expression of six microRNAs encoded by the c13orf24cluster of chromosome 13 (55) Two of these moleculesmicroR-17-5p and microR-20a negatively regulate thetranslation of the transcription factor E2F1 which in turnis also up-regulated by c-Myc The authors propose thatthe microRNAs play a fine-tuning role to regulateproliferative signals (55) MicroRNAs also regulate cellproliferation or differentiation Thus microRNA-133enhances proliferation of the mouse myoblast C2C12cells while microRNA-1 promotes differentiation andmyogenesis (56)
SUPPLEMENTARY DATA
Supplementary Data are available at NAR Online
ACKNOWLEDGEMENTS
We thank Dr Giuseppe Attardi California Institute ofTechnology Pasadena CA for suggesting the treatmentof HeLa cells with ethidium bromide We also thankDr Marc Shuman from UCSF for providing severalhuman cell lines and for his enthusiastic support Wethank Dr Bernardita Mendez Dr Pablo ValenzuelaDr Arturo Yudelevich and Dr Mario Rosemblatt fortheir continuous support and enriched discussions Thiswork was supported by Millennium Scientific Initiative N8P04-071-F Grant D04I1338 FONDEF CONICYTSantiago Chile and Grants DID-32-03 DID-26-04 and
Nucleic Acids Research 2007 Vol 35 No 21 7345
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

DID-57-04 Universidad Nacional Andres Bello Fundingto pay the Open Access publication charges for this articlewas provided by MIFAB P04-071-F
Conflict of interest statement None declared
REFERENCES
1 NassMM NassS and AfzeliusBA (1965) The generaloccurrence of mitochondrial DNA Exp Cell Res 37 516ndash539
2 AndersonS BankierAT BarrellBG de BrujinMHCoulsonAR DrouinJ EperonIC NierlichDP RoeBAet al (1981) Sequence and organization of the human mitochondrialgenome Nature 290 457ndash465
3 OjalaD MontoyaJ and AttardiG (1981) tRNA punctuationmodel of RNA processing in human mitochondria Nature 290470ndash474
4 ChomynA and AttardiG (1992) Recent advances on mitochon-drial biogenesis In ErnsterL (ed) Molecular Mechanisms inBiogenesis Elsevier Science Publishers B V Amsterdampp 483ndash509
5 ShadelGS and ClaytonDA (1997) Mitochondrial DNA main-tenance in vertebrates Ann Rev Biochem 66 409ndash435
6 TaanmanJ-W (1999) The mitochondrial genome structuretranscription translation and replication Biochim Biophys Acta1410 103ndash123
7 Fernandez-SilvaP EnriquezJA and MontoyaJ (2003)Replication and transcription of mammalian mitochondrial DNAExp Physiol 88 41ndash56
8 FalkenbergM LarssonN-G and GustafssonCM (2007) DNAreplication and transcription in mammalian mitochondria AnnRev Biochem 76 301ndash3021
9 ChangDD and ClaytonDA (1984) Precise identification ofindividual promoters for transcription of each strand of humanmitochondrial DNA Cell 36 635ndash643
10 BogenhagenDF ApplegateEF and YozaBK (1984)Identification of a promoter for transcription of the heavy strand ofhuman mtDNA in vitro transcription and deletion mutagenesisCell 36 1105ndash1113
11 ShueyDJ and AttardiG (1985) Characterization of an RNApolymerase activity from HeLa cell mitochondria which initiatestranscription at the heavy strand rRNA promoter and the lightstrand promoter in human mitochondrial DNA J Biol Chem 2601952ndash1958
12 VillegasJ ZarragaAM MullerI MontecinosL WernerEBritoM MenesesAM and BurzioLO (2000) A novel chimericmitochondrial RNA localized in the nucleus of mouse spermDNA Cell Biol 19 579ndash588
13 VillegasJ MullerI ArredondoJ PintoR and BurzioLO(2002a) A putative RNA editing from U to C in a mousemitochondrial transcript Nucleic Acids Res 30 1895ndash1901
14 VillegasJ ArayaP Bustos-ObregonE and BurzioLO (2002b)Localization of the 16S mitochondrial rRNA in the nucleus ofmammalian spermatogenic cells Mol Hum Reprod 8 977ndash983
15 EddySR (2001) Non-coding RNA genes and the modern RNAworld Nat Rev Genet 2 919ndash929
16 PangKC StephenS EngstromPG Tajul-ArifinK ChenWWahlestedtC LenhardB HayashizakiY and MattickJS (2005)RNAdb ndash a comprehensive mammalian noncoding RNA databaseNucleic Acids Res 33 D125ndashD130
17 CostaFF (2005) Non-coding RNAs new players in eukaryoticbiology Gene 357 83ndash94
18 DergunovaNN BulychevaTI ArtemenkoEG ShpakovaAPPegovaAN GemjianEG DudnikOA ZatsepinaOV andMalashenkoOS (2002) A major nucleolar protein B23 as a markerof proliferation activity of human peripheral lymphocytes ImmunolLett 83 67ndash72
19 FussJ and LinnS (2002) Human DNA polymerase epsiloncolocalizes with proliferating nuclear antigen and DNA replicationlate but not early in S phase J Biol Chem 277 8658ndash8666
20 SchmidtMH BrollR BruchHP and DuchrowM (2002)Proliferation marker pKi-67 affects the cell cycle in a self-regulatedmanner J Cell Biochem 87 334ndash341
21 ChadeeDN HendzelMJ TylipskiCP AllisCDBazett-JonesDP WrightJA and DavieJR (1999) IncreasedSer-10 phosphorylation of histone H3 in mitogen-stimulated andoncogene-transformed mouse fibroblast J Biol Chem 27424914ndash24920
22 OrsquoConnorPM FerrisDK PaganoM DraettaG PinesJHunterT LongoDL and KohnKW (1993) G2 delay inducedby nitrogen mustard in human cells affects cyclinAcdk2 and cyclinB1cdc2-kinase complex differently J Biol Chem 268 8298ndash8308
23 WetzlerM TalpazM YeeG StassSA Van EttenRAAndreeffM GoodacreAM KleinerHD MahadeviaRK et al(1995) Cell cycle-related shifts in subcellular localization of BCRassociation with mitotic chromosomes and with heterochromatinProc Natl Acad Sci USA 92 3488ndash3492
24 KingMP and AttardiG (1989) Human cells lacking mtDNArepopulation with exogenous mitochondria by complementationScience 246 500ndash503
25 ChomczynskiP and SacchiN (1987) Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloroformextraction Anal Biochem 162 156ndash159
26 WalshPS MetzgerDA and HiguchiR (1991) Chelex 199 asa medium for simple extraction of DNA for PCR-based typingfrom forensic material Biotechniques 10 506ndash513
27 SambrookJ FritschET and ManiatisT (1989) MolecularCloning A Laboratory Manual 2nd edn Cold Spring HarborLaboratory Press Cold Spring Harbor New York
28 YoungPG and AttardiG (1975) Characterization of double-stranded RNA from HeLa cell mitochondria Biochem BiophysRes Commun 65 1201ndash1207
29 FrohmanMA DushMK and MartinGR (1988) Rapidproduction of full-length cDNAs from rare transcriptsamplification using single gene-specific oligonucleotide primerProc Natl Acad Sci USA 85 8998ndash9002
30 ChurchGM and GilbertW (1984) Genomic sequencingProc Natl Acad Sci USA 81 1991ndash1995
31 KruseB MurdterNN and AttardiG (1995) Transcription systemusing a HeLa cell mitochondrial lysate Methods Mol Biol 37179ndash197
32 GainesG and AttardiG (1984) Highly efficient RNA-synthesizingsystem that uses isolated human mitochondria new initiation eventsand in vivo-like processing patterns Mol Cell Biol 4 1605ndash1617
33 Van EttenRA BirdJW and ClaytonDA (1983) Identificationof the 30-ends of the two mouse mitochondrial ribosomal RNAsThe 30-end of 16 S ribosomal RNA contains nucleotides encoded bythe gene for transfer RNALeuUUR J Biol Chem 25810104ndash10110
34 Rodriguez-CousinoN EstebanLM and EstebanR (1991)Molecular cloning and characterization of W double-strandedRNA a linear molecule present in Saccharomyces cerevisiae J BiolChem 266 12772ndash12778
35 CoteJ and Ruiz-CarrilloA (1993) Primers for mitochondrialDNA replication generated by endonuclease G Science 261765ndash769
36 HayashiJ TakemitsuM and NonakaI (1992) Recovery of themissing tumorigenicity in mitochondrial DNA-less HeLa cells byintroduction of mitochondrial DNA from normal cells Somat CellMol Genet 18 123ndash129
37 HayakawaT NodaM YasudaK YorifujiH TaniguchiSMiwaI SakuraH TerauchiY HayashiJ et al (1998) Ethidiumbromide-induced inhibition of mitochondrial gene transcriptionsuppresses glucose-stimulated insulin release in the mousepancreatic b-cell line bHC9 J Biol Chem 273 20300ndash20307
38 LeeJT DavidowLS and WarshawskyD (1999) Tsix a geneantisense to Xist at the X-inactivation centre Nat Genet 21400ndash404
39 CanceillD and EhrlichD (1996) Copy-choice recombinationmediated by DNA polymerase III haloenzyme from Escherichia coliProc Natl Acad Sci USA 93 6647ndash6652
40 CanceillD ViqueraE and EhrlichSD (1999) Replicationslippage of different DNA polymerases is inversely related to theirstrand displacement efficiency J Biol Chem 274 27481ndash27490
41 GlickmanBW and RipleyLS (1984) Structural intermediates ofdelation mutagenesis a role for palindromic DNA Proc Natl AcadSci USA 81 512ndash516
7346 Nucleic Acids Research 2007 Vol 35 No 21
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from
42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from

42 MadsenCS GhivizzaniSC and HauswirthWW (1993) In vivoand in vitro evidence for slipped mispairing in mammalianmitochondria Proc Natl Acad Sci USA 90 7671ndash7675
43 VigueraE CanceillD and EhrlichSD (2001) In vitro replicationslippage by DNA polymerases from thermophilic organismsJ Mol Biol 312 323ndash333
44 HewY GrzelczakZ LauC and KeeleyFW (1999)Identification of a large region of secondary structure in the30-unstranslated region of chicken elastin mTNA with implicationsfor the regulation of mRNA stability J Biol Chem 27414415ndash14421
45 ViswanathanVK KrcmarikK and CianciottoNP (1999)Template secondary structure promotes polymerase jumping duringPCR amplification Biotechniques 27 508ndash511
46 GeisztM LekstromK and LetoTL (2004) Analysis of mRNAtranscripts from the NAD(P)H oxidase 1 (Nox1) geneEvidence against production of the NADPH oxidase homolog-1short (NOH-1S) transcript variant J Biol Chem 27951661ndash51668
47 WoischnikM and MoraesCT (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome GenomeRes 12 885ndash893
48 BlancV AlfonzoJD AphasizhevR and SimpsonL (1999) Themitochondrial RNA ligase from Leishmania tarentolae can joinRNA molecules bridged by a complementary RNA J Biol Chem274 24289ndash24296
49 AgorioA ChalarC CardozoS and SalinasG (2003) AlternativemRNAs arising from trans-splicing code for mitochondrial andcytosolic variants of Echinococcus granulosus thioredoxin glu-tathione reductase J Biol Chem 278 12920ndash12928
50 SimpsonL SbisegoS and AphasizhevR (2003) Uridine inserta-tiondelation RNA editing in trypanosome mitochondria a complexbusiness RNA 9 265ndash276
51 AloniY and AttardiG (1971) Symmetrical in vivo transcription ofmitochondrial DNA in HeLa cells Proc Natl Acad Sci USA 681757ndash1761
52 LungB ZemannA MadejMJ SchuelkeM TechritzS RufSBockR and HuttenhoferA (2006) Identification of small non-coding RNAs from mitochondria and chloroplasts Nucleic AcidsRes 34 3842ndash3852
53 BartelDP (2004) Micro RNAs genomics biogenesis mechanismsand function Cell 116 281ndash297
54 HatfieldSD SccherbataHR FischerKA NakaharaKCarthewRW and Ruohola-BakerH (2005) Stem cell division isregulated by the microRNA pathway Nature 435 974ndash978
55 OrsquoDonellKA WentzelEA ZellerKI DangCV andMendellJT (2005) c-Myc-regulated microRNAs modulate E2F1expression Nature 435 839ndash843
56 ChenJ-F MandelEM ThomsonJM WuQ CallisTEHammondSM ConlonFL and WangD-Z (2006) The role ofmicroRNA-1 and microRNA-133 in skeletal muscle proliferationand differentiation Nat Genet 38 228ndash233
Nucleic Acids Research 2007 Vol 35 No 21 7347
by guest on March 7 2014
httpnaroxfordjournalsorgD
ownloaded from




















