
Predicting folding pathways between RNA conformational structures guided by RNA stacks

1
2
3
456
7
8910111213141516171819202122
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
Biochimica et Biophysica Acta xxx (2009) xxx–xxx
BBAGRM-00153; No. of pages: 11; 4C: 8
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbagrm
ARTICLE IN PRESS
F
Identification and characterization of barley RNA-directed RNA polymerases
Christian Toft Madsen a,1, Jennifer Stephens b, Csaba Hornyik b, Jane Shaw b, David B. Collinge c,Christophe Lacomme b,2, Merete Albrechtsen a,⁎a Department of Genetics and Biotechnology, Faculty of Agricultural Sciences, University of Aarhus, Thorvaldsensvej 40, 1871-Frederiksberg C, Denmarkb Plant Pathology, Scottish Crop Research Institute, Invergowrie, Dundee DD2 5DA, UKc Department of Plant Biology and Biotechnology, Faculty of Life Sciences, University of Copenhagen, Thorvaldsensvej 40, 1871-Frederiksberg C, Denmark
U
⁎ Corresponding author. Tel.: +45 3533 2582; fax: +E-mail address: [email protected] (M. Albre
1 Current address: Department of Biology, University o5, 2200-Copenhagen N, Denmark.
2 Current address: Institute of Molecular Plant ScienceUniversity of Edinburgh, King's Buildings, Mayfield Road
1874-9399/$ – see front matter © 2009 Elsevier B.V. Aldoi:10.1016/j.bbagrm.2009.03.003
Please cite this article as: C.T. Madsen et alActa (2009), doi:10.1016/j.bbagrm.2009.03
Oa b s t r a c t
a r t i c l e i n f oArticle history:Received 9 October 2008Received in revised form 21 March 2009Accepted 23 March 2009Available online xxxx
Keywords:RNA-directed RNA polymeraseRNA interferenceDefence gene inductionSalicylic acidBarley
DPRORNA-directed RNA polymerases (RDRs) play crucial roles in the RNA silencing response of plants by
enhancing and maintaining silencing signals. At least two members of the RDR group, namely RDR1 andRDR6, are implicated in defence against plant viruses. RDRs have so far only been characterized in dicotspecies. In this report, we identified and characterized HvRDR1, HvRDR2 and HvRDR6 genes in the monocotplant barley (Hordeum vulgare). We analysed their expression under various biotic and abiotic stressesincluding fungal and viral infections, salicylic acid treatment as well as during plant development. Thedifferent classes and subclasses of barley RDRs displayed contrasting expression patterns during pathogenchallenge and development suggesting their involvement in specific regulatory pathways. Their response toheat and salicylic acid treatment suggests a conserved pattern of expression of these genes between monocotand dicot plant species. The existence of two HvRDR1 and two HvRDR6 genes suggests an evolutionaryselection for specialization in response to biotic and abiotic stresses after gene duplication.
© 2009 Elsevier B.V. All rights reserved.
E58
1. Introduction59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
NCOR
RECRNA silencing is an endogenous physiological mechanism which
fulfils the role of monitoring the genome for integrity and inappropri-ate expression of endogenous and exogenous RNAs, transposableelements and viruses [1]. It provides a sequence-specific inhibition ofgene expression, either at the transcriptional, mRNA stability ortranslational levels (for a review see [2]) and eliminates RNAs that aresensed as “aberrant” through direct recognition of, or formation of,double stranded RNA (dsRNA). dsRNA molecules are further pro-cessed into short interfering (si)RNAs 21 to 25 nucleotides (nt) inlength by a group of RNaseIII-like enzymes called Dicer-like (DCL)enzymes [3,4]. Dicer-processed siRNAs are incorporated into aneffector complex called the RNA-induced silencing complex (RISC),containing the Argonaute (AGO) class of proteins, in order to mediatehomology-dependant RNA degradation by base pairing between thesiRNA and the target RNA [5,6]. Arabidopsis thaliana encodes 4 DCLand 10 AGO proteins involved in the recognition and processing ofdsRNA [7–9]. In addition, the RNA-directed RNA polymerase (RDR)class of proteins also has important roles in RNA silencing pathways
78
79
80
81
82
83
84
45 3533 2589.chtsen).f Copenhagen, Ole Maaløes vej
s, School of Biological Sciences,, Edinburgh EH9 3JR, UK.
l rights reserved.
., Identification and character.003
T[10–12]. RDRs produce a secondary pool of dsRNA which serves assubstrate for DCL enzymes to amplify the initial siRNA signal [13].
The A. thaliana genome contains 6 RDR genes, some of which areknown to have important roles in post-transcriptional gene silencing,virus and viroid infections, RNA-directed DNA methylation andheterochromatin formation in plants [14]. However, a role has yetto be assigned to AtRDR3a, ATRDR3b and AtRDR3c. AtRDR1, theexpression of which can be induced by virus infection or salicylic acid(SA) in A. thaliana and Nicotiana tabacum, has been shown to play arole in antiviral defence [15,16]. AtRDR2 produces dsRNA fromtandem repeat units and retroelements for subsequent DCL3processing and transcriptional silencing in heterochromatic regions[12]. AtRDR6 (formerly annotated SGS2/SDE1) produces the initiatingprimary dsRNA in single-copy transgene silencing, in addition tobeing involved in direct-repeat transgene RNA silencing pathways[10,11], in the long-term maintenance of systemic silencing [10], aswell as in the spreading of the silencing potential along the targetRNA sequence known as transitivity [17]. Furthermore, AtRDR6 isimplicated in transacting siRNA (tasiRNA) biogenesis [18,19] and thequality control of mRNA maturation by converting improperlymatured RNA lacking either a 5′ cap-structure or a 3′ polyadenylatedtail into dsRNA [20–22].
Plants deficient in RDR activity frequently exhibit increasedsusceptibility to some viral species. Furthermore, the RDRs appear tohave overlapping functions in antiviral defence. For example, the Ar-abidopsis rdr6 mutant exhibits increased susceptibility to Cucumbermosaic virus (CMV) but not to Turnip vein-clearing virus (TVCV),
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.
CTED
PROO
F
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
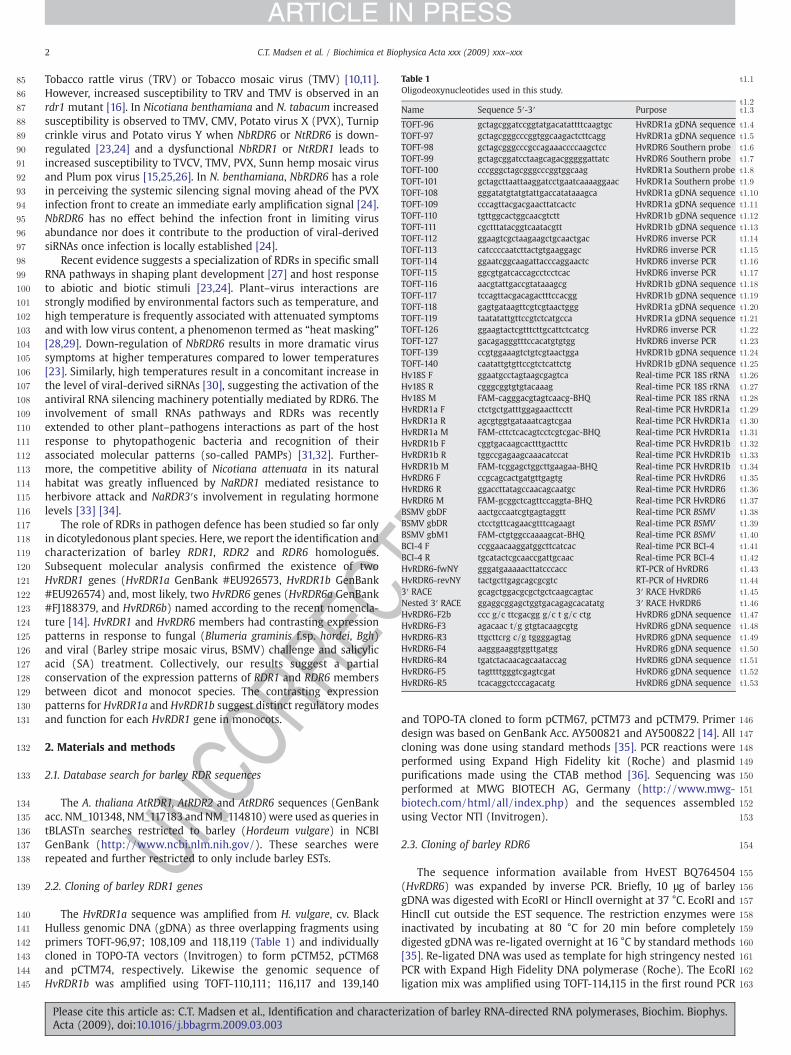
Table 1 t1:1
Oligodeoxynucleotides used in this study.t1:2t1:3Name Sequence 5′-3′ Purpose
t1:4TOFT-96 gctagcggatccggtatgacatattttcaagtgc HvRDR1a gDNA sequencet1:5TOFT-97 gctagcgggcccggtggcaagactcttcagg HvRDR1a gDNA sequencet1:6TOFT-98 gctagcgggcccgccagaaaccccaagctcc HvRDR6 Southern probet1:7TOFT-99 gctagcggatcctaagcagacgggggattatc HvRDR6 Southern probet1:8TOFT-100 cccgggctagcgggcccggtggcaag HvRDR1a Southern probet1:9TOFT-101 gctagcttaattaaggatcctgaatcaaaaggaac HvRDR1a Southern probet1:10TOFT-108 gggatatgtatgtattgaccatataaagca HvRDR1a gDNA sequencet1:11TOFT-109 cccagttacgacgaacttatcactc HvRDR1a gDNA sequencet1:12TOFT-110 tgttggcactggcaacgtctt HvRDR1b gDNA sequencet1:13TOFT-111 cgctttatacggtcaatacgtt HvRDR1b gDNA sequencet1:14TOFT-112 ggaagtcgctaagaagctgcaactgac HvRDR6 inverse PCRt1:15TOFT-113 catccccaatcttactgtgaaggagc HvRDR6 inverse PCRt1:16TOFT-114 ggaatcggcaagattacccaggaactc HvRDR6 inverse PCRt1:17TOFT-115 ggcgtgatcaccagcctcctcac HvRDR6 inverse PCRt1:18TOFT-116 aacgtattgaccgtataaagcg HvRDR1b gDNA sequencet1:19TOFT-117 tccagttacgacagactttccacgg HvRDR1b gDNA sequencet1:20TOFT-118 gagtgataagttcgtcgtaactggg HvRDR1a gDNA sequencet1:21TOFT-119 taatatattgttccgtctcatgcca HvRDR1a gDNA sequencet1:22TOFT-126 ggaagtactcgtttcttgcattctcatcg HvRDR6 inverse PCRt1:23TOFT-127 gacagagggtttccacatgtgtgg HvRDR6 inverse PCRt1:24TOFT-139 ccgtggaaagtctgtcgtaactgga HvRDR1b gDNA sequencet1:25TOFT-140 caatattgtgttccgtctcattctg HvRDR1b gDNA sequencet1:26Hv18S F ggaatgcctagtaagcgagtca Real-time PCR 18S rRNAt1:27Hv18S R cgggcggtgtgtacaaag Real-time PCR 18S rRNAt1:28Hv18S M FAM-cagggacgtagtcaacg-BHQ Real-time PCR 18S rRNAt1:29HvRDR1a F ctctgctgatttggagaacttcctt Real-time PCR HvRDR1at1:30HvRDR1a R agcgtggtgataaatcagtcgaa Real-time PCR HvRDR1at1:31HvRDR1a M FAM-cttctcacagtcctcgtcgac-BHQ Real-time PCR HvRDR1at1:32HvRDR1b F cggtgacaagcactttgactttc Real-time PCR HvRDR1bt1:33HvRDR1b R tggccgagaagcaaacatccat Real-time PCR HvRDR1bt1:34HvRDR1b M FAM-tcggagctggcttgaagaa-BHQ Real-time PCR HvRDR1bt1:35HvRDR6 F ccgcagcactgatgttgagtg Real-time PCR HvRDR6t1:36HvRDR6 R ggaccttatagccaacagcaatgc Real-time PCR HvRDR6t1:37HvRDR6 M FAM-gcggctcagttccaggta-BHQ Real-time PCR HvRDR6t1:38BSMV gbDF aactgccaatcgtgagtaggtt Real-time PCR BSMVt1:39BSMV gbDR ctcctgttcagaacgtttcagaagt Real-time PCR BSMVt1:40BSMV gbM1 FAM-ctgtggccaaaagcat-BHQ Real-time PCR BSMVt1:41BCI-4 F ccggaacaaggatggcttcatcac Real-time PCR BCI-4t1:42BCI-4 R tgcatactcgcaaccgattgcaac Real-time PCR BCI-4t1:43HvRDR6-fwNY gggatgaaaaacttatcccacc RT-PCR of HvRDR6t1:44HvRDR6-revNY tactgcttgagcagcgcgtc RT-PCR of HvRDR6t1:453′ RACE gcagctggacgcgctgctcaagcagtac 3′ RACE HvRDR6t1:46Nested 3′ RACE ggaggcggagctggtgacagagcacatatg 3′ RACE HvRDR6t1:47HvRDR6-F2b ccc g/c ttcgacgg g/c t g/c ctg HvRDR6 gDNA sequencet1:48HvRDR6-F3 agacaac t/g gtgtacaagcgtg HvRDR6 gDNA sequencet1:49HvRDR6-R3 ttgcttcrg c/g tggggagtag HvRDR6 gDNA sequencet1:50HvRDR6-F4 aagggaaggtggttgatgg HvRDR6 gDNA sequencet1:51HvRDR6-R4 tgatctacaacagcaataccag HvRDR6 gDNA sequencet1:52HvRDR6-F5 tagttttgggtcgagtcgat HvRDR6 gDNA sequencet1:53HvRDR6-R5 tcacaggctcccagacatg HvRDR6 gDNA sequence
2 C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
UNCO
RRE
Tobacco rattle virus (TRV) or Tobacco mosaic virus (TMV) [10,11].However, increased susceptibility to TRV and TMV is observed in anrdr1 mutant [16]. In Nicotiana benthamiana and N. tabacum increasedsusceptibility is observed to TMV, CMV, Potato virus X (PVX), Turnipcrinkle virus and Potato virus Y when NbRDR6 or NtRDR6 is down-regulated [23,24] and a dysfunctional NbRDR1 or NtRDR1 leads toincreased susceptibility to TVCV, TMV, PVX, Sunn hemp mosaic virusand Plum pox virus [15,25,26]. In N. benthamiana, NbRDR6 has a rolein perceiving the systemic silencing signal moving ahead of the PVXinfection front to create an immediate early amplification signal [24].NbRDR6 has no effect behind the infection front in limiting virusabundance nor does it contribute to the production of viral-derivedsiRNAs once infection is locally established [24].
Recent evidence suggests a specialization of RDRs in specific smallRNA pathways in shaping plant development [27] and host responseto abiotic and biotic stimuli [23,24]. Plant–virus interactions arestrongly modified by environmental factors such as temperature, andhigh temperature is frequently associated with attenuated symptomsand with low virus content, a phenomenon termed as “heat masking”[28,29]. Down-regulation of NbRDR6 results in more dramatic virussymptoms at higher temperatures compared to lower temperatures[23]. Similarly, high temperatures result in a concomitant increase inthe level of viral-derived siRNAs [30], suggesting the activation of theantiviral RNA silencing machinery potentially mediated by RDR6. Theinvolvement of small RNAs pathways and RDRs was recentlyextended to other plant–pathogens interactions as part of the hostresponse to phytopathogenic bacteria and recognition of theirassociated molecular patterns (so-called PAMPs) [31,32]. Further-more, the competitive ability of Nicotiana attenuata in its naturalhabitat was greatly influenced by NaRDR1 mediated resistance toherbivore attack and NaRDR3′s involvement in regulating hormonelevels [33] [34].
The role of RDRs in pathogen defence has been studied so far onlyin dicotyledonous plant species. Here, we report the identification andcharacterization of barley RDR1, RDR2 and RDR6 homologues.Subsequent molecular analysis confirmed the existence of twoHvRDR1 genes (HvRDR1a GenBank #EU926573, HvRDR1b GenBank#EU926574) and, most likely, two HvRDR6 genes (HvRDR6a GenBank#FJ188379, and HvRDR6b) named according to the recent nomencla-ture [14]. HvRDR1 and HvRDR6 members had contrasting expressionpatterns in response to fungal (Blumeria graminis f.sp. hordei, Bgh)and viral (Barley stripe mosaic virus, BSMV) challenge and salicylicacid (SA) treatment. Collectively, our results suggest a partialconservation of the expression patterns of RDR1 and RDR6 membersbetween dicot and monocot species. The contrasting expressionpatterns for HvRDR1a and HvRDR1b suggest distinct regulatory modesand function for each HvRDR1 gene in monocots.
2. Materials and methods
2.1. Database search for barley RDR sequences
The A. thaliana AtRDR1, AtRDR2 and AtRDR6 sequences (GenBankacc. NM_101348, NM_117183 andNM_114810)were used as queries intBLASTn searches restricted to barley (Hordeum vulgare) in NCBIGenBank (http://www.ncbi.nlm.nih.gov/). These searches wererepeated and further restricted to only include barley ESTs.
2.2. Cloning of barley RDR1 genes
The HvRDR1a sequence was amplified from H. vulgare, cv. BlackHulless genomic DNA (gDNA) as three overlapping fragments usingprimers TOFT-96,97; 108,109 and 118,119 (Table 1) and individuallycloned in TOPO-TA vectors (Invitrogen) to form pCTM52, pCTM68and pCTM74, respectively. Likewise the genomic sequence ofHvRDR1b was amplified using TOFT-110,111; 116,117 and 139,140
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
1and TOPO-TA cloned to form pCTM67, pCTM73 and pCTM79. Primer1design was based on GenBank Acc. AY500821 and AY500822 [14]. All1cloning was done using standard methods [35]. PCR reactions were1performed using Expand High Fidelity kit (Roche) and plasmid1purifications made using the CTAB method [36]. Sequencing was1performed at MWG BIOTECH AG, Germany (http://www.mwg-1biotech.com/html/all/index.php) and the sequences assembled1using Vector NTI (Invitrogen).
12.3. Cloning of barley RDR6
1The sequence information available from HvEST BQ7645041(HvRDR6) was expanded by inverse PCR. Briefly, 10 μg of barley1gDNA was digested with EcoRI or HincII overnight at 37 °C. EcoRI and1HincII cut outside the EST sequence. The restriction enzymes were1inactivated by incubating at 80 °C for 20 min before completely1digested gDNAwas re-ligated overnight at 16 °C by standard methods1[35]. Re-ligated DNA was used as template for high stringency nested1PCR with Expand High Fidelity DNA polymerase (Roche). The EcoRI1ligation mix was amplified using TOFT-114,115 in the first round PCR
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
235
236
237
238
239
240
241
242
243
244
245
246
247
248
249
250
251
252
253
254
255
256
257
258
259
260
261
262
263
264
265
266
267
268
269
270
271
272
273
274
275
276
277
278
279
280
281
3C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
UNCO
RREC
(30 cycles of 94 °C 20 s, 68 °C 30 s, and 72 °C 5 min) and 1 μl from thefirst round PCR was then used as a template in the second round PCRusing nested primers TOFT-112,113 (as above but with 65 °C asannealing temp). In a similar approach, the HincII religation mix wasamplified using TOFT-115,127 in the first round PCR (64 °C asannealing temp). TOFT-113,126 was used in the second round PCR(64 °C as annealing temp). PCR products were TOPO-TA cloned(Invitrogen) and sequenced; two clones originating from the EcoRIand HincII digests revealed a high degree of similarity to AtRDR6 andOsRDR6. A further 780 bp fragment of genomic sequence wasidentified by genome walking as described by [37] with somemodifications (M. Schiller et al, unpublished) including the use ofTaKaRa LA Taq as polymerase (TAKARA BIO Inc.). Intron–exonboundaries were confirmed by RT-PCR on barley RNA using primersHvRDR6-fwNY and HvRDR6-revNY followed by sequencing. Toamplify the 3′ end of the RDR6 transcript, RNA was extracted usingAmbion RiboPure kit and used to make RACE-ready cDNA withInvitrogen's GeneRacer Kit. PCR primers used were 3′ RACE withGeneRacer 3′ primer, followed by Nested 3′ RACE with GeneRacer 3′nested primer. Finally, the assembled sequence (GenBank FJ188379)was confirmed by sequencing of four overlapping fragments amplifiedfrom gDNA using primers HvRDR6-F2b and TOFT-99, HvRDR6-F3 andHvRDR6-R3, HvRDR6-F4 and HvRDR6-R4, and HvRDR6-F5 andHvRDR6-R5 (Table 1).
2.4. In silico analysis of RDR sequences
The putative barley RDR sequences obtained were compared toRDR1, RDR2 and RDR6 sequences from A. thaliana, N. benthamiana,Triticumaestivum, Zeamays andOryza sativa using TreePuzzle softwarewith default support values. Only coding sequences were included inthe analysis. Protein sequences were aligned using ClustalW onlinesoftware. The output file was run in TreePuzzle using quartet puzzlingmaximum likelihoodmethod, JTT amino acid substitutionmatrix. Rateheterogeneity among sites was modelled using a gamma distributionwith four categories. The boxshade software 3.21 from the EMBnet(http://www.ch.embnet.org/software/BOX_form.html) was used forvisualising alignments.
2.5. Nucleic acid isolation
Barley gDNA (cv. Black Hulless) was isolated by grinding 0.25 gof leaf material in liquid N2 and further incubated with 500 μl ofextraction buffer (100 mM Tris–HCl pH 8.5, 100 mM NaCl, 50 mMEDTA 2% (w/v) SDS). Nucleic acids were extracted with 500 μlphenol/chloroform and 500 μl chloroform. DNA was precipitatedwith 40 μl 3 M Na Acetate pH 5.6 and 1 ml 96% Ethanol for 30 minat −20 °C, pelleted at 13.000 rpm for 15 min at 4 °C andredissolved in 600 μl MilliQ H2O. RNA was removed by precipitationwith 300 μl 8 M LiCl at 0 °C for 1 h. gDNA was precipitated from thesupernatant with 0.6 volume of isopropanol. gDNA pellets werewashed twice with 80% Ethanol and redissolved in 20 μl MilliQ H2O.RNA used for quantitative real-time PCR was extracted using theRNeasy mini kit (Qiagen) using the on-column DNase treatment assuggested by the manufacturer.
2.6. Southern blotting
Southern blotting was performed by digesting 10 μg of gDNA (cv.Black Hulless). The digest was transferred after agarose gel electro-phoresis to a Hybond-XL nylon membrane (Amersham Biosciences)overnight in 20×SSC buffer (3 M NaCl, 0.3 M Na Citrate. pH 7.0) usingstandard techniques [35]. The gDNA was UV-crosslinked (0.6 J/cm2)before overnight hybridization at 65 °C in 1% (w/v) BSA, fraction V(Sigma), 7% SDS, 1 mM EDTA, 0.5 M sodium-phosphate pH 7.0. Probeswere 100 ng Gfx purified (Amersham Biosciences) PCR fragment
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
TEDPR
OOF
formed by amplification from pCTM52 (HvRDR1a) using TOFT-100,101 or from pCTM53 (HvRDR6a) using TOFT-98, 99. Probes werelabelled withα-32P dCTP (370MBq/ml, Amersham Biosciences) usingthe Amersham Megaprime kit as recommended by the manufacturer.The membrane was washed twice in 2×SSC, 0.5% SDS, twice in 2×SSC,0.1% SDS and 4 times in 0.1×SSC, 0.5% SDS before autoradiography at−80 °C using Kodak Biomax MS films.
2.7. Barley inoculation
We obtained Barley stripe mosaic virus (BSMV) cDNA clones fromDr. Andy Jackson, University of California, Berkeley, USA. BMSVinoculation was achieved as described previously [38]. The BSMVinoculation experiments shown in Fig. 3B were conducted understandard greenhouse conditions with light intensities not exceeding1200 μmol m−2 s−1 and the temperature close to 20 °C. Climatechamber experiments shown in Fig. 3C were conducted at 20 °C and28 °C respectively running a 16 h light/8 h dark cycle with lightintensity between 240 and 380 μmol m−2 s−1. Powdery mildewisolates 206 and 213 exhibiting respectively an incompatible and acompatible interaction on barley cv. Clansman were obtained fromNIAB. Powdery mildew inoculation was effectuated as previouslydescribed [39].
2.8. Salicylic acid (SA) treatment
Primary leaves of 7-day old barley cv. Black Hulless seedlingsgrown in a climate chamber at 20 °C, light regime as above, wereused. Leaf segments of 5 cm length were floated on water, 50 μM SA,200 μM SA, or 500 μM SA and incubated in a growth chamber for24 h before being harvested into liquid nitrogen. RNA was extractedand DNase treated as described above and analysed by quantitativereal-time RT-PCR.
2.9. Quantitative real-time RT-PCR and semi-quantitativeRT-PCR analysis
RNAwas extracted from individual barley plants and DNase treatedas described above. Quantification of HvRDR1a, HvRDR1b, HvRDR6and BSMV RNA was performed using Taqman real-time RT-PCR asdescribed previously [38]. BCI-4 mRNA was quantified using SYBRGreen real-time RT-PCR as described [40]. Primer and probesequences are shown in Table 1. Primer/probe sets were validated asdescribed [38]. All samples were measured in duplicate and theresulting data were normalised to 18S rRNA using the ddCT method[41]. Statistical analysis consisted in determining if equal variance wasupheld between two sample groups (F-test) and a student t-test at95% confidence level to test whether a statistically significantdifference existed between two mean values. The specificity of theHvRDR1a and HvRDR1b primer/probe sets was evaluated usingdifferent concentrations of plasmids containing either the HvRDR1aor the HvRDR1b sequence. The HvRDR1a primer/probe set was notcompletely specific but produced an amplicon from the HvRDR1atarget 10 PCR cycles before amplification from the HvRDR1b target.The HvRDR1b primer/probe set was completely specific and did notamplify the HvRDR1a target at any concentration tested. Semi-quantitative RT-PCR analysis of HvRDR1a, HvRDR1b, HvRDR6, PR10mRNA and 26S rRNA analysis was performed as described previously[42]. A total of six leaves from three independent mock or Blumeriachallenged plants—barley cv Clansman—were sampled and pooledprior to RNA extraction.
2.10. Meta-analysis of RDR expression in Arabidopsis and barley
Probe set IDs for Arabidopsis RDR genes were obtained from theArabidopsis Co-expression Tools website (http://www.arabidopsis.
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

C
282
283
284
285
286
287
288
289
290
291
292
293
294
295
296
297
298
299
300
301
302
303
304
305
306
307
308
309
310
311
312
313
314
315
316
317
318
319
320
321
322
323
324
325
326
327
328
329
330
331
332
333
334
335
336
337
338
339
340
341
342
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
00
01
02
03
04
05
06
4 C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
UNCO
RRE
leeds.ac.uk/act/idexchanger.php). Probe sets for the barley RDRswereobtained from the Plant Expression Database (http://www.plexdb.org/). Expression profiles of RDRs on the Affymetrix Arabidopsis ATH122K array and the Barley1 22K genome arrays were investigated usingthe Genevestigator website (http://www.genevestigator.ethz.ch/).The expression profile of HvRDR1a (contig 44342) and putativeHvRDR2 (contig 14349) in response to Blumeria challenge wereinvestigated using the affymetrix data at PLEXdb/BarleyBase (http://www.plexdb.org) [43,44].
3. Results
3.1. Identification of barley RNA-directed RNA polymerases
The A. thaliana AtRDR1 and AtRDR6 sequences were used asqueries in a barley restricted BLAST search in the NCBI GenBankdatabase. This revealed two cDNA sequences with significantsimilarity to the Arabidopsis AtRDR1 gene. Wassenegger and co-workers recognized the same two barley cDNA sequences as RNA-directed RNA polymerases (RDRs) and suggested the nomenclatureHvRDR1a and HvRDR1b [14]. We use this nomenclature henceforth.We also identified a number of ESTs from different barley genotypeswith considerable similarity to RDR-like sequences from related plantspecies. Sequence information from the barley EST BQ764504 showedsignificant similarity to AtRDR6 and was considered a likely HvRDR6candidate. We expanded our search to the available barley microarraydataset and identified two contigs; 44342 and 14349, which exhibitedcomplete identity to HvRDR1a and resemblance to RDR2-likesequences, respectively. No HvRDR6-like contig was identified on thebarley 22K Affymetrix chip.
To confirm our classification of barley RDR members, we clonedand sequenced HvRDR1a, HvRDR1b and HvRDR6 from genomic (g)DNA. The sequence information available from EST BQ764504(HvRDR6) was expanded from 671 nt to 4822 nt by a combinationof high stringency inverse PCR, genome walking, and 3′ RACE. Theassembled genomic gene sequence was confirmed by amplificationand sequencing of four overlapping fragments from gDNA and hasbeen deposited at GenBank as acc. FJ188379. The HvRDR6 sequencecorresponds to most of the OsRDR6 exon 1 followed by an intronsequence and the complete OsRDR6 exon 2, and 206 bp of the 3′ UTR.Attempts to determine the sequence of the 5′ UTR and the extreme 5′end of exon 1 were unsuccessful. The expanded barley HvRDR6sequence extends over the conserved DxDGD motif in the catalyticdomain, strongly supporting that FJ188379 is correctly identified as abarley RDR6 orthologue (Fig. 1B and 1C) [14]. We also sequenced thegenomic clones of HvRDR1a and HvRDR1b and compared theassembled sequences to the cDNA sequences available from GenBankacc. AY500821 and AY50822 [14]. We note that HvRDR1a andHvRDR1b possess comparable gene architectures, i.e. 4 exons and 3introns each (Fig. 2A) and overall 89% identity at the nucleotide level(excluding intron sequences). The genomic sequences of HvRDR1aand HvRDR1b have been deposited at GenBank as Acc. EU926573and EU926574.
A phylogenetic analysis was performed on the putative HvRDR1a,HvRDR1b, HvRDR2 and HvRDR6 sequences together with establishedmonocot and dicot RDR sequences (Fig. 1A). This analysis supports thecorrect assignment of barley HvRDR-like sequences into groups 1, 2and 6.
3.2. Southern blotting of barley RNA-directed RNA polymerases
To corroborate the number of RDR1 genes in barley, we performedSouthern blotting using a probe originating from the first exon ofHvRDR1a (Fig. 2A). Based on the close similarity of HvRDR1a andHvRDR1b we expect our HvRDR1a derived probe to hybridize to bothHvRDR1 genes. We used two restriction enzymes cutting outside the
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
TEDPR
OOF
3probe hybridization region of both HvRDR1a and HvRDR1b (EcoRI and3HindIII) and a single restriction enzyme (XhoI) cutting exclusively3within the probe hybridization region of HvRDR1a. EcoRI is expected3to produce two bands of N1.7 kb and N0.6 kb, HindIII is expected to3produce two bands of N0.9 kb and N0.7 kb band, and XhoI is expected3to produce three bands of N4.8 kb, N4.4 kb and N0.2 kb, respectively.3The EcoRI and HindIII band patterns confirmed the presence of two3RDR1 like genes in barley, whereas the XhoI digest surprisingly also3produced only two bands (Fig. 2B). Presumably, the 4.9 kb XhoI band3is a double band containing the HvRDR1a fragment 3′ of the XhoI site3as well as the entire HvRDR1b gene (Fig. 2A). The 2.7 kb band can only3derive from probe hybridization 5′ to the XhoI site in HvRDR1a. In any3case, the Southern data do not indicate the presence of more than two3HvRDR1 genes. A similar Southern blot was used with an HvRDR63derived probe (Fig. 2C) to investigate the number of HvRDR6-like3genes in barley. Again we used two restriction enzymes cutting3outside the probe area (EcoRI and HindIII) and one enzyme cutting in3the middle of the probe hybridization region (BglII). We would expect3one band in the EcoRI and HindIII lanes of 2.8 kb and N3.4 kb3respectively, and two bands in the BglII lane of 2.0 kb and N1.1 kb if3barley harbours one RDR6 like gene. Surprisingly, we obtained an3additional band from both EcoRI and HindIII digests and four bands3from the BglII digest (Fig. 2C), suggesting the presence of two3paralogous RDR6 genes in barley. Accordingly, we rename the3FJ188379 sequence as HvRDR6a.
33.3. Expression patterns of HvRDR members during fungal and3viral infections
3RNA-directed RNA polymerases as well as other defence related3genes are often induced as a result of pathogen infection or salicylic3acid (SA) treatment in A. thaliana and N. tabacum [15,16,26]. To3investigate the expression patterns of HvRDR members during3pathogen infection, we infected barley cv. Clansman with a virulent3and an avirulent strain of the biotrophic fungus powdery mildew, B.3graminis f.sp. hordei (Bgh) resulting respectively in a compatible or an3incompatible interaction. In this cultivar, race-specific resistance is3mediated by the resistance gene Mla13 leading to a hypersensitive3response—incompatible interaction—upon challenge by the Bgh strain3206 harbouring the corresponding avirulence gene AvrMla13 [39,45].3Alternatively, a compatible interaction leading to fungal colonisation3is observed upon challenge by the Bgh strain 213 that does not carry3AvrMla13. Irrespective of the outcome of the interaction, Bgh induces3a number of defence related genes in barley including pathogenesis3related (PR) proteins in what appears to be an SA-independent3manner [46,47]. We followed the expression of each barley RDR3member HvRDR1a, HvRDR1b and HvRDR6 for 24 h after Bgh infection3by semi-quantitative RT-PCR (Fig. 3A). For comparison, accumulation3of transcripts of PR protein 10 (PR10) was also determined. To3differentiate between the two HvRDR1a and HvRDR1b members,3oligonucleotides were selected to amplify specifically each individual3gene. The absence of cross PCR amplification was validated as3described in Materials and methods. No obvious changes in3HvRDR1a transcript levels were observed except for a moderate3increase within the first hour of the interaction with the avirulent Bgh3isolate. HvRDR1b expression on the other hand showed a peak in3transcript accumulation by 18 h post inoculation (hpi) during both3incompatible and compatible interactions, preceding the strong3increase of PR10 transcript accumulation seen by 24 hpi (Fig. 3A).4In contrast, a decrease in HvRDR6 transcript accumulation by 24 h4after challenge was observed during both incompatible and compa-4tible interactions. No such changes in the expression of HvRDR1a,4HvRDR2 or HvRDR6a were observed after mock inoculation (Suppl.4Fig. 1). Furthermore, we analysed HvRDR1a and HvRDR2 expression4patterns from the publicly available Affymetrix/PLEXdb database,4comparing compatible and incompatible interactions of different
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

UNCO
RREC
TEDPR
OOF
407
408
409
410
411
412
413
414
415
416
Fig. 1. Characterization of barley RNA-directed RNA polymerase genes. (A) Phylogram of RDR sequences from selected monocot and dicot species constructed using TreePuzzlesoftware with support values. The following abbreviations are used. At: Arabidopsis thaliana, Nb: Nicotiana benthamiana, Hv: Hordeum vulgare, Ta: Triticum aestivum, Os: Oryza sativa,and Zm: Zea mays. Accession numbers as follow: AtRDR1 (GenBank NM_101348), ZmRDR1 (GenBank AY103827), HvRDR1a (GenBank EU926573), HvRDR1b (GenBank EU926574),TaRDR1 (GenBank BJ268864), OsRDR1 (GenBank NM_001054586), AtRDR6 (GenBank NM_114810), NbRDR6 (GenBank AY722008), HvRDR6a (GenBank FJ188379), OsRDR6(GenBank AP004357), TaRDR6 (GenBank CF134248), AtRDR2 (GenBank NM_117183), NbRDR2 (GenBank AY722009), HvRDR2 (Barleybase contig 14349), and OsRDR2 (GenBankAP008210). Only coding sequences were used for the analysis. Calculated distances are shown. (B) Partial sequence alignment of HvRDR1a, HvRDR1b, HvRDR6a, and OsRDR6extending over the putative catalytic motif markedwith asterisks. Identical nucleotides are in black and similar amino acids in grey. (C) The same region as in B translated into aminoacids showing the putative catalytic DLDGD motif marked with asterisks. Proposed intron–exon junctions are marked with black arrows in the case of HvRDR1a and HvRDR1b andwith grey arrows for HvRDR6a. Identical amino acids are marked in black and similar amino acids in grey.
5C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
barley near-isogenic lines—Pallas NILs-carrying different Mla resis-tance genes challenged with Bgh isolates K1 and 5874 [45] (Fig. 4A).A higher expression level of HvRDR1awas observed in comparison toHvRDR2. No specific trend of HvRDR1a expression patterns duringincompatible or compatible interactions was observed, in agreement
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
with our own data on HvRDR1a expression in a barley Mla13background (Fig. 3A). Similarly, no obvious trend of HvRDR2expression patterns was observed.
Next, we investigated the expression level of barley RDR membersduring virus infection. RDR expression patterns were in this case
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

CPR
OOF
417
418
419
420
421
422
423
424
425
426
427
428
429
430
431
432
433
434
435
436
437
438
439
440
441
442
443
444
445
446
447
448
449
450
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
Fig. 2. Genomic maps and Southern blots of barley RNA-directed RNA polymerases. (A) Genomic region of HvRDR1a, HvRDR1b, HvRDR6a and OsRDR6 drawn to scale. The size bar is500 bp. Exons are represented as open boxes and introns as thin black lines. The grey areas show the probe regions used for Southern blots. Thick black lines represent the regionsamplified by the qRT-PCR primers. Restriction sites important for the Southern blot and the inverse PCR amplification are marked as XI (XhoI), HIII (HindIII), EI (EcoRI), HII (HincII)and BII (BglII). (B) Southern blot of barley gDNA developed with HvRDR1a probe. Band sizes in kb as estimated by comparison to a lambda HindIII DNA marker are shown. (C)Southern blot of barley gDNA developed with HvRDR6a probe.
6 C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
UNCO
RRE
measured by quantitative real-time PCR (qRT-PCR) using 18S rRNA fornormalisation. We infected the first leaf of 6 day old barley seedlingscv. Black Hulless with Barley stripemosaic virus (BSMV). The third leafwas harvested for RNA extraction in three separate experiments ateither 10, 14 or 18 days post inoculation (dpi). The relative expressionof HvRDR1a, HvRDR1b and HvRDR6 from 5 independent virus-infectedplants was compared to the expression from 5 independentuninfected control plants (Fig. 3B). In the case of HvRDR1b andHvRDR6, there was a statistically significant increase in expressionlevels in virus-infected leaves compared to the control group(pb0.05). The approximately two-fold increase in expression wasobserved in three independent experiments. We did observe astatistically significant increase in HvRDR1a expression after virusinfection in one experiment (data not shown), but HvRDR1bexpression was more consistently induced by virus treatment. Thisprompted us to compare the relative expression levels of HvRDR1aand HvRDR1b. HvRDR1b transcript was on average 4 times moreabundant than the HvRDR1a transcript during virus infection in the14 dpi experiment (data not shown). Collectively, we conclude thatbarley RDRmembers appear to be differentially induced in a pathogenspecific manner.
3.4. Induction of barley RDR members during abiotic stress conditions
The effect of temperature on barley RDR expressionwas examined.Six day old barley plants kept in growth chambers at 20 °C or 28 °Cwere inoculated with BSMV. At 10 dpi, the third leaf was harvested forRNA extraction and analysis by qRT-PCR. As observed previously,HvRDR1a was not significantly induced by virus infection at 20 °C,however at 28 °C, a significant increase in response to virus infectionwas observed (pb0.05) (Fig. 3C). The HvRDR1b transcript was notsignificantly induced by virus infection in these experiments, althoughthe virus-induced increase at 20 °C was close to being significant. ForHvRDR6, a significant increase in expression upon virus infection wasobserved at 20 °C and a high basal HvRDR6 expression level at 28 °Cwas not significantly increased following virus infection (Fig. 3C). The
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
TED 4amount of virus was also determined by qRT-PCR and found to be
4significantly lower at 28 °C than at 20 °C (Fig. 3C). We did not observe4any obvious differences in virus symptoms at the two temperatures4(data not shown).4The effect of salicylic acid (SA) on RDR expression in barley was4examined in a detached leaf assay. Primary leaves from 7 day old4barley plants were floated for 24 h on water containing SA at4concentrations of 50 μM, 200 μM and 500 μM. The HvRDR1a4expression did not increase in response to SA treatment compared4to thewater control, on the contrary, a statistically significant decrease4was observed at the highest SA concentrations of 200 μM and 500 μM4(Fig. 3D). In contrast, HvRDR1b expression was significantly increased4in response to increasing SA concentrations (pb0.05). HvRDR64expression was unchanged at any SA concentration (Fig. 3D). As a4control for SA treatment, we also measured the expression of a Ca2+-4binding EF-hand protein; BCI-4 (GenBank AJ250283) which is4chemically induced by SA or structural mimic compounds [48]. We4observed an approximately 500-fold induction of BCI-4 at the highest4SA concentration compared to the water control (Fig. 3D), in4agreement with previous observations [48].
43.5. HvRDR expression analysis during development and in4different organs
4According to the sequence-homology-based annotations, there are4two probe sets on the Barley 1 GeneChip corresponding to HvRDR1a4and HvRDR2. We used the publicly available datasets to analyse4HvRDR1a and HvRDR2 expression patterns during barley development4and across tissues [44] (Fig. 4B and D). HvRDR1a displayed a marked4increase in transcript accumulation in floral tissues including spike,4spikelet, floret and most strongly in the pistil (respectively N10-fold,4N15-fold, N20-fold, and N30-fold higher than in coleoptile and radicle4tissues from seedlings) (Fig. 4B). In contrast, a lower HvRDR1a4expression level was found in stamen and anthers (Fig. 4B) suggesting4a highly tissue-regulated mode of expression associated with flower-4ing (Fig. 4D) and between male and female organs (Fig. 4B). Despite a
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

NCOR
RECT
EDPR
OOF
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
500
Fig. 3. Expression analysis of barley RNA-directed RNA polymerases. (A) Semi-quantitative RT-PCR analysis of HvRDR1a, HvRDR1b, HvRDR6, PR10 and 26S rRNA expression at fivedifferent time points (0, 1, 5, 18 and 24 h post inoculation—hpi) during B. graminis f.sp. hordei (Bgh) infection. 0 h constitutes the control before fungal inoculation. Bgh strains 206and 213 are avirulent and virulent on barley cv. Clansman, respectively. The number of PCR amplification cycles was 25 for 26S, 30 for HvRDR6 and HvRDR1b and 35 for HvRDR1a,corresponding to the log-linear phase for all genes. (B) Relative expression of HvRDR1a, HvRDR1b and HvRDR6 in barley cv. Black Hulless at three different time points after Barleystripe mosaic virus (BSMV) infection. These experiments were conducted under greenhouse conditions with temperatures close to 20 °C. The expression was measured by qRT-PCRusing primer/probe sets positioned as shown in Fig. 2A. The relative expression of each RDR transcript was normalised to 18S rRNA. Mean values and standard deviations arecalculated from 5 individual plants. An asterisk (⁎) indicates that the sample is significantly different (pb0.05) from healthy untreated control plants. (C) Relative expression ofHvRDR1a, HvRDR1b, HvRDR6 and BSMV at 20 °C and 28 °C measured by qRT-PCR. These experiments were conducted under controlled climate chamber conditions. The controlconstitutes uninoculated barley cv. Black Hulless. Mean values and standard deviations are calculated from 5 individual plants. Columns marked with the same letters are notsignificantly different (pN0.05). (D) Relative expression of HvRDR1a, HvRDR1b, HvRDR6 and BCI-4 in response to salicylic acid treatment measured by qRT-PCR. Mean values andstandard deviations are calculated from 5 individual leaves. An asterisk (⁎) indicates sample is significantly different (pb0.05) from water control.
7C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
Ulower expression level than for HvRDR1a, HvRDR2 displayed acomparable pattern of transcript accumulation in the same tissueswith the highest expression levels observed (N10-fold higher than inradicle tissues from seedlings) in the pistil (Fig. 4B). In Arabidopsisindividual AtRDR1, AtRDR2, AtRDR3c and AtRDR6 display contrastingexpression levels in different tissues, with the highest expressionfound in cork, callus, seed and root tip respectively (Fig. 4C). Duringdevelopment, AtRDR1, AtRDR2 and AtRDR6 display a similar expression
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
pattern while AtRDR3c is expressed more strongly in germinated seed(N7-fold higher than in young flower) and mature siliques (N20-foldhigher than in young flower) (Fig. 4E). Expression increase for bothHvRDR1a and HvRDR2 was observed during stem elongation(Fig. 4D), and as AtRDR1 displays a strong increase in the hypocotylsand cork, this could suggest a conserved role for RDR1 in vascular-associated functions (Fig. 4B–E). Globally, an increase in expressionwas associated with reproductive tissues and/or flowering
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

UNCO
RREC
TEDPR
OOF
8 C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
Please cite this article as: C.T. Madsen et al., Identification and characterization of barley RNA-directed RNA polymerases, Biochim. Biophys.Acta (2009), doi:10.1016/j.bbagrm.2009.03.003

OOF
501
502
503
504
505
506
507
508
509
510
511
512
513
514
515
516
517
518
519
520
521
522
523
524
525
526
527
528
529
530
531
532
533
534
535
536
537
538
539
540
541
542
543
544
545
546
547
548
549
550
551
552
553
554
555
556
557
558
559
560
Table 2t2:1
Arabidopsis and barley RNA-directed RNA polymerase expression analysis.t2:2t2:3 Developmental
stage/TreatmentA. thaliana H. vulgare Referencesa
AtRDR1 AtRDR6 HvRDR1a HvRDR1b HvRDR6
t2:5 Reproductive phase ↑ ↑ ↑ n.a. n.d. Genevestigator and [43]t2:6 Vegetative phase – – – n.a. n.d.t2:7 Biotic stress Genevestigator and [43]t2:8 P. syringae ↓ ↓ n.a. n.a. n.a.t2:9 B. graminisb – ↑ ↓t2:10 TMVc ↑ – n.a. n.a. n.a.t2:11 BSMVd n.a. n.a. – ↑ ↑t2:12 Chemical stress Genevestigator and [43]t2:13 Syringolin – ↓ n.a. n.a. n.a.t2:14 Salicylic acid c/d ↑ – – ↑ –
t2:15 Methyl jasmonate ↓ – n.d. n.d. n.d.t2:16 Temperature stress Genevestigator and [43]t2:17 Heat/d ↑ ↑ ↑ – ↑t2:18 Cold ↓ ↓ n.d. n.d. n.d.t2:19 Nutritional stress Genevestigator and [43]t2:20 Low glucose ↑ – n.d. n.d. n.d.t2:21 Low nitrate ↑ ↑ n.d. n.d. n.d.
↑:Up-regulation; ↓:downregulation; –:no differential expression; n.a.:no available data; n.d.:not determined.t2:22 a All expression changes were determined from Genevestigator (http://www.genevestigator.ethz.ch/) and affymetrix data from [43] except those marked with superscripts as
described below.t2:23b Determined by RT-PCR (this study).t2:24c Determined by Northern blotting [16].t2:25d Determined by qRT-PCR (this study).t2:26
9C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
NCOR
REC
suggesting a high level of coordination of RDR expression in differentplant species.
4. Discussion
We report the further identification and characterization of RDRsin barley. Sequence analysis of HvRDR1a, HvRDR1b and HvRDR6 genesconfirmed their phylogenetic relationship with other RDR membersfrom closely related monocots and more distantly related dicots (Fig.1A). Southern blotting and sequence data confirmed that barleypossesses two copies of both RDR1 and RDR6 (Fig. 2). Our PCR-basedcloning approaches produced only one HvRDR6 sequence (GenBank#FJ188379), even though some of the primers used for amplifying theoverlapping HvRDR6 fragments from gDNA were designed torecognize sequences conserved between the rice and maize RDR6genes and the HvRDR6a sequence. This could mean that the twoHvRDR6 genes are identical throughout the sequenced region, or thattheir sequences differ at numerous nt positions. Since the sequencedHvRDR6 region includes an intron we favour the latter interpretation.We propose that the HvRDR6 sequence (FJ188379), be renamedHvRDR6a in accordance with the recent nomenclature [14]. Althoughwe consider it unlikely we cannot completely rule out that HvRDR6bcontributes to the expression patterns determined here for HvRDR6a;consequently we refer to the expression data as HvRDR6 throughoutthis report.
The expression of HvRDR1a, HvRDR1b and HvRDR6 was monitoredunder various biotic and abiotic stresses. Quantitative RT-PCRanalysis using sequence-specific oligonucleotides was undertakento discriminate the expression patterns between HvRDR1a andHvRDR1b. The two genes exhibited different expression patterns inresponse to Bgh challenge. No obvious change in expression was
UFig. 4. Microarray analysis of barley and Arabidopsis RDR gene expression in response toexpression profiles (Plexdb website) for HvRDR1a (red) and HvRDR2 (blue) in incompatibleMla-specified reciprocal interactions with Bgh isolates (5874 and K1) on Pallas NILs [44,45].linear scale and calculated based on three independent replications (+/− SE) as previouslyspecific gene expression for HvRDR1a (red) and HvRDR2 (blue) genes in barley and (C) in AThe left axis represents the change of gene expression on a linear scale (+/− SE). The numheat maps of gene expression during development for HvRDR1a (red) and HvRDR2 (blue) gand AtRDR6 (green) genes. The left axis represents the change of gene expression on a lineadevelopmental stage.
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
TEDPRobserved for HvRDR1a, as opposed to the HvRDR1b expression which
increased transiently by 18 hpi (Fig. 3A). In A. thaliana and Nicotianaspecies, RDR1 is induced by SA [15,16,25,26]. However, SA appears tohave a minor role in defence gene induction in cereals [49] as SAlevels remain largely unaffected in response to Bgh infection [46]. Inspite of the fact that HvRDR1b expression increased upon SAtreatment and precede PR-10 transcripts, HvRDR1b expressionfollowing Bgh challenge may not be associated to an SA-dependentresponse, as expression analysis of the SA-responsive gene BCI-4(PLEXdb contig AJ250283_at) was not significantly increased uponBgh challenge ([45], PLEXdb website and data not shown). Moreover,PR-proteins are selectively activated by the SA and jasmonic acidpathways in cereals as opposed to dicots. Indeed, PR-3 and PR-5 areinduced following treatment with either JA or SA, and PR-1 and PR-9are only activated by the JA pathway [50,51], suggesting that thecontribution of the SA-pathway in PR induction in cereals issignificantly different from dicots and is likely to vary betweendifferent cereal–pathogen interactions.
HvRDR1a expression appeared to be only slightly affected duringvirus infection at the lower temperature (20 °C), while HvRDR6expression and in most experiments also HvRDR1b expression wassignificantly increased. Importantly, HvRDR1a showed a significantincrease in expression at 28 °C in response to virus infection. In thatrespect, HvRDR1a and HvRDR6 expression strongly correlates with arole in antiviral defence. Strikingly, their viral-induced expressionwasenhanced at the higher temperature and was associated with asignificant decrease in BSMV accumulation, reminiscent of thepreviously mentioned “heat masking” phenomenon [28–30]. Thissuggests that HvRDR1a and HvRDR6 could fulfil a similar function inlimiting virus propagation at higher temperatures, as observed inplant–virus interactions in dicots [15,16,24,26].
pathogen challenge in specific tissues and during development. (A) Histograms of(Mla1/5874, Mla1/K1, Mla6/5874, Mla13/K1) and compatible (Mla6/K1, Mla13/5874)The signal intensities (A.U. x-axis) over time (0, 8, 16, 20, 24 and 32 hpi y-axis) are ondescribed [44,45]. (B) Scatterplots and heat maps (Genevestigator website) of tissue-
rabidopsis for AtRDR1 (red), AtRDR2 (blue), AtRDR3c (black) and AtRDR6 (green) genes.bers on the bottom axis represent the number of arrays per tissue. (D) Scatterplots andenes in barley and (E) in Arabidopsis for AtRDR1 (red), AtRDR2 (blue), AtRDR3c (black)r scale (+/− SE). The numbers on the bottom axis represent the number of arrays per
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

C
561
562
563
564
565
566
567
568
569
570
571
572
573
574
575
576
577
578
579
580
581
582
583
584
585
586
587
588
589
590
591
592
593
594
595
596
597
598
599
600
601
602
603
604
605
606
607
608
609
610
611
612
613
614
615
616
617
618
619
620
621
622
623
624
625
626
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62 Q1
63646566676869707172737475767778798081828384858687888990919293
10 C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
UNCO
RRE
We analysed publicly available Arabidopsis gene profiling data forAtRDR1 and AtRDR6 using the Arabidopsis Genevestigator resource(http://www.genevestigator.ethz.ch). AtRDR1 expression increased inresponse to SA treatment, low levels of nutrients (glucose and nitrate)as well as heat-treatment, thereby confirming that AtRDR1 displays acomparable expression pattern to both HvRDR1a and HvRDR1b.AtRDR6 is similarly upregulated in response to low nitrate level andheat. These data are summarized inTable 2which also includes expres-sion changes from biotic stress conditions determined elsewhere.
Exogenous application of SA resulted in distinct expressionpattern for HvRDR1a, HvRDR1b and HvRDR6. HvRDR1b expressionappears to be induced upon SA treatment similar to its A. thaliana andN. tabacum RDR1 homologues supporting an evolutionarily conservedfunction between monocots and dicots for HvRDR1b and HvRDR6based on gene expression. The apparent difference in HvRDR1a andHvRDR1b expressions suggests a specialization of their respectivefunction. Duplication of RDR1 and RDR6 gene members in barley mayhave been a recent event. This possibly reflects the divergence in theregulation of innate and induced disease resistance response topathogens between monocots and dicots [49], possibly requiring thecoordinated regulation of multiple RDR1 and RDR6 members togenerate an efficient RNA silencing response. Taken together, thesedata suggest that the selective induction of barley RDRs is dependenton specific pathogen-host combinations, triggering different defencepathways some of which are SA-dependent [46,48].
Analysis of the publicly available barley Affymetrix dataset(PLEXdb website) revealed that HvRDR1a and HvRDR2 exhibitcomparable tissue-specific expression patterns with a markedincrease in reproductive organs (spike, spikelet, floret and pistil). Incontrast, the lowest expression was observed in vegetative tissuessuch as seedlings and shoot (Fig. 4B). In Arabidopsis, AtRDR1, AtRDR2and AtRDR6 display a comparable expression pattern associated withthe early stage of flower development with a moderate increaseduring boulting in comparison to other developmental stages (Fig.4D, E). However, AtRDR3c expression contrasted significantly fromothers RDRs, displaying high expression levels in germinated seedand mature siliques. We observed a generally lower RDR2 expressionlevel in comparison to RDR1 and RDR6 during development andpathogen challenge (Fig. 4A–E). Another conserved trend is therelatively low level of induction observed for all HvRDRs (usually inthe range of a two-fold increase) in response to pathogen challengein comparison to defence genes such as PR-10 or the SA-responsivegene BCI-4 (500-fold for the latter). An exceptionally high increase inRDR expression (ranging from 3-fold to 8-fold) was observed forHvRDR1a and HvRDR2 during development in particular in thefemale (8-fold and 10-fold respectively for HvRDR1a and HvRDR2 inthe pistil) as opposed to the male (basal expression level in stamen,anther) reproductive organs. The functional significance of increasedRDR expression in reproductive tissues is supported by the observa-tion that small RNAs are generally more abundant in these tissues[27]. Recently, a significant enrichment of FLOWERING LOCUS C (FLC)3′ small RNAs was found in siliques compared to seedlings in Arabi-dopsis [27]. Interestingly, the biogenesis of FLC small RNAs requiredAtRDR2; the resulting RNAi-mediated chromatin modification isinvolved in suppressing FLC, a repressor of the transition to floweringin Arabidopsis [27].
The existence of multiple RDRs in plants reflects the operation ofseveral small RNA-directed silencing pathways and subsequentfunctional specialization of DCLs, AGOs and RDRs. Recently, smallRNAs have been identified as potentially key regulators of bacterialpathogenicity [31,52]. A pathogen-inducible endogenous 22-nt siRNAderiving from natural antisense transcripts nat-siRNAATGB2 wasshown to accumulate specifically in Arabidopsis in response to Pseu-domonas syringae AvrRpt2 and to contribute to RPS2-mediated race-specific resistance. This process is RDR6-dependant [52]. The biogen-esis of nat-siRNAATGB2 is believed to occur upon expression of two
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
TEDPR
OOF
6overlapping and antiparallel genes ATGB2 and PPRL, resulting in the6creation of dsRNA and siRNA targeting PPRL transcripts for RISC-6mediated degradation [52]. Since then, other classes of 30–40 nt long6siRNAs (lsiRNAs) have been identified that accumulate during6pathogen infection or specific growth conditions [32]. The biogenesis6of these lsiRNAs also requires RDR6 [32]. Strikingly, we observed a6decrease in HvRDR6 expression by 24 hpi after Bgh challenge. It is6tempting to speculate that a possible pathogen-induced downregula-6tion of HvRDR6 may be part of the Bgh invasion strategy by lowering6the endogenous siRNA and lsiRNA levels and affecting in turn the6defence response. Infection with the hemibiotrophic fungus Rhynch-6osporium secalis did not result in a noticeable change of HvRDR66expression levels on resistant and susceptible barley cultivars6(Supplementary data Fig. 2). Different fungi may be differently6affected by plant RDR genes and the suggested pathogen-induced6HvRDR6 downregulation may be highly pathogen specific. Further6analysis of RDR expression in response to challenge by a broader range6of fungi and viruses should help to validate the possible functional6specialization of barley RDRs.6These data collectively suggest the existence of conserved6intricate regulatory pathways of the silencing machinery involving6RDRs in very distinct modes of regulation of gene expression6associated with the innate defence response and plant development6in dicots and monocots.
6Acknowledgements
6We gratefully acknowledge A. O. Jackson (University of California,6Berkeley, USA) for providing the BSMV clones, B. S. Olsen (University6of Aarhus), Brian Harrower and Ingo Hein (SCRI) for technical6assistance and P. Hedley (SCRI) for useful comments. This work was6supported by the Danish National Research Foundation; CH was6supported by a Marie Curie IEF fellowship (contract number MEIF-CT-62005-025076).
6Appendix A. Supplementary data
6Supplementary data associated with this article can be found, in6the online version, at doi:10.1016/j.bbagrm.2009.03.003.
6References
6[1] O. Voinnet, Nat. Rev., Genet. 6 (2005) 206–220.6[2] P. Brodersen, O. Voinnet, Trends Genet. 22 (2006) 268–280.6[3] P.D. Zamore, T. Tuschl, P.A. Sharp, D.P. Bartel, Cell 101 (2000) 25–33.6[4] E. Bernstein, A.A. Caudy, S.M. Hammond, G.J. Hannon, Nature 409 (2001)6363–366.6[5] J.B. Morel, C. Godon, P. Mourrain, C. Beclin, S. Boutet, F. Feuerbach, F. Proux, H.6Vaucheret, Plant Cell 14 (2002) 629–639.6[6] H. Vaucheret, F. Vazquez, P. Crete, D.P. Bartel, Genes Dev. 18 (2004) 1187–1197.6[7] S.E. Schauer, S.E. Jacobsen, D.W. Meinke, A. Ray, Trends Plant. Sci. 7 (2002)6487–491.6[8] C. Hunter, H. Sun, R.S. Poethig, Curr. Biol. 13 (2003) 1734–1739.6[9] R. Margis, A.F. Fusaro, N.A. Smith, S.J. Curtin, J.M. Watson, E.J. Finnegan, P.M.6Waterhouse, FEBS Lett. 580 (2006) 2442–2450.6[10] T. Dalmay, A. Hamilton, S. Rudd, S. Angell, D.C. Baulcombe, Cell 101 (2000)6543–553.6[11] P. Mourrain, C. Beclin, T. Elmayan, F. Feuerbach, C. Godon, J.B. Morel, D. Jouette,6A.M. Lacombe, S. Nikic, N. Picault, K. Remoue, M. Sanial, T.A. Vo, H. Vaucheret,6Cell 101 (2000) 533–542.6[12] Z. Xie, L.K. Johansen, A.M. Gustafson, K.D. Kasschau, A.D. Lellis, D. Zilberman, S.E.6Jacobsen, J.C. Carrington, PLoS Biol. 2 (2004) E104.6[13] K. Nishikura, Cell 107 (2001) 415–418.6[14] M. Wassenegger, G. Krczal, Trends Plant Sci. 11 (2006) 142–151.6[15] Z. Xie, B. Fan, C. Chen, Z. Chen, Proc. Natl. Acad. Sci. U. S. A. 98 (2001) 6516–6521.6[16] D. Yu, B. Fan, S.A. MacFarlane, Z. Chen, Mol Plant Microbe Interact 16 (2003)6206–216.6[17] F.E. Vaistij, L. Jones, D.C. Baulcombe, Plant Cell 14 (2002) 857–867.6[18] F. Vazquez, H. Vaucheret, R. Rajagopalan, C. Lepers, V. Gasciolli, A.C. Mallory, J.L.6Hilbert, D.P. Bartel, P. Crete, Mol. Cell 16 (2004) 69–79.6[19] E. Allen, Z. Xie, A.M. Gustafson, J.C. Carrington, Cell 121 (2005) 207–221.6[20] S. Gazzani, T. Lawrenson, C. Woodward, D. Headon, R. Sablowski, Science 3066(2004) 1046–1048.
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.

694695696697698699700701702703704705706707708709710711712713714715716717718719
720721722723724725726727728729730731732733734735736737738739740741742743744745
747
11C.T. Madsen et al. / Biochimica et Biophysica Acta xxx (2009) xxx–xxx
ARTICLE IN PRESS
[21] M.J. Axtell, C. Jan, R. Rajagopalan, D.P. Bartel, Cell 127 (2006) 565–577.[22] A.J. Herr, A. Molnar, A. Jones, D.C. Baulcombe, Proc. Natl. Acad. Sci. U. S. A. 103
(2006) 14994–15001.[23] F. Qu, X. Ye, G. Hou, S. Sato, T.E. Clemente, T.J. Morris, J. Virol. 79 (2005)
15209–15217.[24] F. Schwach, F.E. Vaistij, L. Jones, D.C. Baulcombe, Plant Physiol. 138 (2005)
1842–1852.[25] S.J. Yang, S.A. Carter, A.B. Cole, N.H. Cheng, R.S. Nelson, Proc. Natl. Acad. Sci. U. S. A.
101 (2004) 6297–6302.[26] J.M. Alamillo, P. Saenz, J.A. Garcia, Plant J 48 (2006) 217–227.[27] S. Swiezewski, P. Crevillen, F. Liu, J.R. Ecker, A. Jerzmanowski, C. Dean, Proc. Natl.
Acad. Sci. U. S. A. 104 (2007) 3633–3638.[28] J. Johnson, Phytopathology 12 (1922) 438–440.[29] R. Hull, Academic Press (2002).[30] G. Szittya, D. Silhavy, A. Molnar, Z. Havelda, A. Lovas, L. Lakatos, Z. Banfalvi, J.
Burgyan, EMBO J. 22 (2003) 633–640.[31] L. Navarro, P. Dunoyer, F. Jay, B. Arnold, N. Dharmasiri, M. Estelle, O. Voinnet, J.D.
Jones, Science 312 (2006) 436–439.[32] S. Katiyar-Agarwal, S. Gao, A. Vivian-Smith, H. Jin, Genes Dev. 21 (2007)
3123–3134.[33] S.P. Pandey, T. Baldwin, Plant J. 50 (2007) 40–53.[34] S.P. Pandey, E. Gaquerel, K. Gase, T. Baldwin, Plant Physiol. 147 (2008) 1212–1224.[35] J. Sambrook, E.F. Fritsch and T. Maniatis, Cold Spring Harbor Laboratory Press. Cold
Spring Harbor, New York. (1989).[36] G. Del Sal, G. Manfioletti, C. Schneider, Biotechniques 7 (1989) 514–520.[37] H. Guo, J. Xiong, Gene 381 (2006) 18–23.
UNCO
RREC
746
Please cite this article as: C.T. Madsen et al., Identification and characterActa (2009), doi:10.1016/j.bbagrm.2009.03.003
F
[38] M. Bruun-Rasmussen, C.T. Madsen, S. Jessing, M. Albrechtsen, Mol. Plant MicrobeInteract. 20 (2007) 1323–1331.
[39] I. Hein, M. Barciszewska-Pacak, K. Hrubikova, S. Williamson, M. Dinesen, .E.Soenderby, S. Sundar, A. Jarmolowski, K. Shirasu, C. Lacomme, Plant Physiol. 138(2005) 2155–2164.
[40] G.D. Constantin, B.N. Krath, S.A. MacFarlane, M. Nicolaisen, E. Johansen, O.S. Lund,Plant J. 40 (2004) 622–631.
[41] K.J. Livak, T.D. Schmittgen, Methods 25 (2001) 402–408.[42] C. Lacomme, K. Hrubikova, I. Hein, Plant J. 34 (2003) 543–553.[43] L. Shen, J. Gong, R.A. Caldo, D. Nettleton, D. Cook, R.P. Wise, J.A. Dickerson, Nucleic
Acids Res. 33 (2005) D614–D618.[44] A. Druka, G. Muehlbauer, I. Druka, R. Caldo, U. Baumann, N. Rostoks, A. Schreiber,
R. Wise, T. Close, A. Kleinhofs, A. Graner, A. Schulman, P. Langridge, K. Sato, P.Hayes, J. McNicol, D. Marshall, R. Waugh, Funct. Integr. Genomics 6 (2006)202–211.
[45] R.A. Caldo, D. Nettleton, R.P. Wise, Plant Cell 16 (2004) 2514–2528.[46] L. Vallélian-Bindschedler, J.-P. Métraux, P. Schweizer, Mol. Plant Microbe Interact.
11 (1998) 702–705.[47] R. Huckelhoven, J. Fodor, C. Preis, K.H. Kogel, Plant Physiol. 119 (1999) 1251–1260.[48] K. Besser, B. Jarosch, G. Langen, K.H. Kogel, Mol. Plant Pathol 1 (2000) 277–286.[49] K.H. Kogel, G. Langen, Cell. Microbiol. 7 (2005) 1555–1564.[50] P. Schweizer, A. Buchala, P. Silverman, M. Seskar, I. Raskin, J.P. Metraux, Plant
Physiol. 114 (1997) 79–88.[51] J. Geddes, F. Eudes, A. Laroche, L.B. Selinger, Proteomics 8 (2008) 545–554.[52] S. Katiyar-Agarwal, R. Morgan, D. Dahlbeck, O. Borsani, A. Villegas Jr., J.K. Zhu, B.J.
Staskawicz, H. Jin, Proc. Natl. Acad. Sci. U. S. A. 103 (2006) 18002–18007.
OTEDPR
O
ization of barley RNA-directed RNA polymerases, Biochim. Biophys.
Copyright © 2022 FDOKUMEN




















