
The Powdery Mildew Effector CSEP0027 Interacts With Barley ...
Upload
independentCategory
view
0download
0
Sex Plant Reprod (1995) 8:49-59 �9 Springer-Verlag 1995
P.B. Holm �9 S. Knudsen �9 P. Mouritzen �9 D. Negri F.L. Olsen. C. Rou6
Regeneration of the barley zygote in ovule culture
Received: 2 May 1994 / Revision accepted: 3 May 1994
Abstract An ovule culture technique has been estab- lished for barley that allows the regeneration of plants from zygotes. An average of 1.3 plantlets per ovule could be regenerated from more than 60% of the cul- tured ovules and about 75 % of the regenerated plantlets developed into normal, fertile plants. The same regener- ation frequencies were obtained in intact ovules and in ovules where the two integuments had been removed from the micropylar region. Unfertilized ovules and ovules where the fertilized eggs had been destroyed by a microinjection needle did not give rise to embryo-like structures. Plants could be regenerated from the zygote at the same frequency at developmental stages from im- mediately after fertilization until the formation of bicel- lular embryos. This tissue culture system appeared to be largely independent of genotype since similar regenera- tion frequencies were obtained in two different barley cultivars, Igri and Alexis, that in anther and microspore culture behave differently. The same technique has also been applied successfully in the wheat cultivar Walter.
Key words Hordeum vulgare �9 Egg cell In vitro culture �9 Embryogenesis �9 Plant regeneration
introduction
Regeneration of fertilized egg cells and young zygotic embryos by means of in situ culture of intact ovules is
P. B. Holm ( 9 ) - S. Knudsen - P. Mouritzen �9 F. L. Olsen Carlsberg Research Laboratory, 10, Gamle Carlsberg Vej, DK-2500 Copenhagen, Denmark, FAX no: +45-33274741
D. Negri Secobra Recherches S.A., Centre de Bois Henry, F-78580 Maule, France
C. Rou~ Tepral, Brasseries Kronenbourg, 68, route d'Oberhausbergen, F-67200 Strasbourg, France
well established for many plant species (for review, see Rangan 1982). The initial interests in this tissue culture system were primarily to rescue hybrid embryos and to identify media and conditions supporting embryo mat- uration. In a few cases, this technique has also been used to generate haploids by cultivation of unfertilized ovules (for review, see Yang and Zhou 1982).
In the last few years there has been an increasing interest in developing techniques allowing the mani- pulation of egg cells and for the regeneration of in vivo or in vitro fertilized egg cells into plants. These tech- niques are required for the establishment of a more controlled experimental approach to essential proces- ses such as fertilization, zygote development, and early embryogenesis. The egg cell is also considered a pro- mising target for many approaches of relevance for bio- technology and plant breeding such as in vitro fertil- ization and transformation. Substantial efforts have thus been devoted to the isolation of viable embryo sacs and protoplasts of the egg by combined enzymatic di- gestion and microdissection approaches (for review, see Huang and Russell 1992). In general, however, the vi- ability of isolated cells is reported to be poor and only in two cases have regenerable protoplasts of egg cells been isolated. In maize, Kranz et al. (1991) isolated pro- toplasts of the unfertilized egg by dissection of enzymat- ically digested ovules. The protoplasts were in vitro fer- tilized by electrofusion with sperm cells, isolated from mature pollen by osmotic shock. The fusion products were regenerated into plants by co-cultivation with a nonmorphogenic maize suspension (Kranz and L6rz 1993). In barley, protoplasts of the fertilized egg could be isolated by a simple dissection of the ovule and re- generated into fully fertile plants by a co-cultivation with microspores undergoing embryogenesis. The same technique could also be applied to wheat (Holm et al. 1994).
Cultivation and regeneration of the zygote by means of ovule culture represent an alternative to the more complex and tedious isolation and cultivation of isolat- ed egg protoplasts. Many studies have been devoted to

50
developing ovule culture systems that also allow the eggs or small embryos to be accessible to manipulation. Keijzer et al. (1988) outlined a strategy for in vitro fertil- ization of Torenia, where sperm cells were microinjected into the embryo sac. T6pfer and Steinbiss (1985) showed that barley zygotes cultured in ovules could be regener- ated into plants and Steinbiss et al. (1985) suggested that a transformation system for barley might be established by microinjection into the fertilized egg. A similar strat- egy was formulated for wheat (Mathias 1987), involving injection of DNA into the embryo sac on ovaries still attached to the spike. In maize, recent studies have been directed towards mapping the three-dimensional posi- tion of the egg cell in ovules as a prerequisite for mi- croinjection of DNA into the egg (Wagner et al. 1990). Culture techniques have also been established for mi- crodissected ovules consisting primarily of the embryo sac (Campenot et al. 1992) or embryo sacs surrounded by a few layers of nucellus cells (M61 et al. 1993). In wheat, Comeau et al. (1992) have described ovule cul- ture techniques allowing the rescue of proembryos 1-6 days after pollination.
Manipulation of the fertilized egg in situ in isolated ovules by microinjection requires that the egg is accessi- ble and can be viewed with high resolution. For labor- intensive manipulations such as microinjection, it is also essential that plants can be regenerated from the egg cell with a high frequency. The present study has been ad- dressed to developing an ovule culture technique for barley that can meet these two requirements. It has been an additional objective of this study to investigate if the regeneration capacity of the egg cell in situ relates to its developmental stage from immediately after fertilization until the first division of the zygote. The analyses were performed in two barley cultivars, the winter barley Igri and the spring malting barley Alexis that show very different performances in anther and microspore cul- ture.
Materials and methods
Plant material
Barley plants of the winter barley cultivar Igri and the spring malting barley cultivar Alexis were grown in growth cabinets at 15~176 day/night temperatures using a 16-h light period at a light intensity of ca 350 laE x m -2 x s 1. The florets were emasculat- ed 3-4 days before anthesis and pollinated by hand using a fine- tipped brush.
mixture (no. K3129, Sigma Chemical Company, St. Louis, Mo., USA). Vitamin-free casamino acids (no. 2238, Merck, Darmstadt, Germany) were added at a concentration of 250 mg/l together with 20 ml/1 coconut water (no. 570-5180AJ, Gibco Laboratories, Life Technologies, Grand Island, N.Y., USA). The medium was made up 2 x concentrated, the pH adjusted to pH 5.8 with 1 N NaOH and the solution sterilized by filtration (0.22 i, tm pore size). Solid Kao 90 medium was made by mixing the double concentrat- ed Kao 90 medium with 0.8% autoclaved SeaPlaque agarose (FMC BioProducts, Rockland, Me., USA) in a 1 : i ratio. For re- generation, the MS medium described above was used but con- taining only 0.4 mg/1 BAP and 30 g/1 maltose. The medium was also solidified with SeaPlaque agarose as described above. Unless otherwise stated, all chemicals were from Merck or Sigma.
Isolation of ovules
Spikes were harvested 1-24 h after pollination and the ovaries removed from the florets and placed on Kao 90 medium. Ovaries from one spike were sterilized for 10 rain in 5 ml 10% sodium hypochlorite (no. 23039, BDH, Poole, UK) with 50 gl 10% Tween 20 (no. 82218, Merck, Schuchardt, Germany). After two washes in sterile water, the ovaries were transferred to Kao 90 and dissected under a dissection microscope (40x magnification, transmitted light). Ovaries were placed with the placental side facing down- wards and the pedicel and the two lodicules were removed by using a pair of fine-tipped forceps. A cross section though the ovarian tissue was made with the tip of the forceps at a median position and along one side down to the micropylar end of the ovary. The tip of the ovary was removed, exposing the micropylar end of the ovule. In some preparations, the inner and outer integu- ments were dissected away from the micropylar part of the ovule. A few experiments were performed where the tip of the ovule containing the egg was cut off and cultured on liquid Kao 90 with 20% Ficoll 400 (no. 17-0400-02, Pharmacia LKB Biotechnology AB, Uppsala, Sweden). The Ficoll was used as a buoyancy in- creasing agent, which allowed the ovules to float on top of the liquid medium. In the majority of cases, the dissected ovaries were transferred to 2.5-cm Whatman filter paper discs (no. 1004025, Whatman, Maidstone, UK) on agarose-solidified medium. The ovaries were opened by cleaving them slightly to the one side of their longitudinal axis and the ovules lifted out and placed on filter paper with their placental side facing downwards. Steritin culture dishes (5 cm, Bibby Sterilin, Stone, Staffs, UK) were sealed with Nesco film (Nippon Shoji Kaisha, Ltd., Osaka, Japan), wrapped in aluminum foil and incubated at 23~ After 3 weeks, the dishes were opened and the developing embryo-like structures were dissected out of the ovule. After 3-4 weeks, when the em- bryos started to germinate, they were transferred to regeneration medium in Sterilin dishes and cultured under low light conditions at 23~ When additional growth had occurred, the plantlets were moved to fresh regeneration medium in 10-cm-high cylindrical containers (Greiner, Bibby Sterilin). For the winter barley Igri, the plantlets were transferred to 4~ for 8 weeks for vernalization when large plantlets had formed. Thereafter, the plantlets were transferred to soil and cultured in a greenhouse equipped with artificial light and heating/cooling facilities (15/10~ 16 h light period).
Media
In all ovule cultures, a medium referred to as Kao 90 was used. This contains the macro- and micro-elements of the modified Mu- rashige and Skoog (MS) medium (Murashige and Skoog 1962) described by Olsen (1987). In this medium, the NH4NO 3 concen- tration is reduced to 165 mg/l while 750 mg/1 of glutamine is added. The medium also contained 1 mg/1 6-benzylaminopurine (BAP), maltose (90 g/l) as the only carbohydrate source, and the vitamins and organic acids from the Kao medium (Kao and Michayluk 1975). The vitamins were purchased as a ready-to-use
Optical analysis of the egg in vivo
Ovaries where one or both integuments had been removed from the micropylar part of the ovule were mounted for optical analysis in the following way: the outermost 2-3 mm of a 24 x 40 mm cover slip was covered with a rim of silicone glue (Fugendichtung Silicon, Practicus Chemie,Grevenbroich, Germany) and the cover slips sterilized by placing them under UV illumination for a min- imum of 30 min. The ovaries were mounted in the glue with the exposed ovule tip towards the center of the cover slip and oriented to ensure that the longitudinal axis of the egg cell was parallel to

the surface of the coverslip. Kao 90 medium was added in suffi- cient amounts to cover the ovule tip. The dissected ovules were analyzed under differential interference contrast on a Zeiss Ax- iovert microscope using a 40X objective with a variable focal distance. The microscope was equipped with a VE-1000 video enhancement system (DAGE-MTI, Inc., Ind., USA). Needles to be used for disrupting the egg were pulled from borosilicate micro- capillaries (microelectrode capillaries no. 1403512, 1.5 mm in out- er diameter, Hilgenberg, Malsfeld, Germany) using a vertical puller (Bachofer, Reutlingen, Germany) and the tips bevelled with a Narishige EG-4 microgrinder (Narishige, Setagaya-Ku, Tokyo, Japan).
Regeneration in relation to the stage of the zygote
Ovules were sampled at 1 h intervals from 2 h after pollination until 28 h after pollination and the integuments removed from the micropylar part of half the ovules of each spike. The analysis was performed for both Igri and Alexis. To ensure optimal synchrony between the individual zygotes, florets from the tip and the base of the spike were not used, since they are delayed in development compared to the florets of the median part of the spike. Fifteen to 20 ovules were cultured for each interval of each of the four com- binations, i.e., Igri and Alexis with and without integuments in the micropylar part. The embryo-like structures were dissected out after 21-28 days of culture and the number of ovules with embryo- like structures recorded as responding ovules. The number of em- bryo-like structures in each ovule was counted and if loose aggre- gates of embryo-like structures were present, the individual struc- tures were separated. Individual embryo-like structures from each ovule were cultured separately and the number of plants regener- ated from each ovule was counted.
Light microscopy
Freshly isolated as well as cultured ovules were fixed in 4% glu- taraldehyde in 50 mM phosphate buffer, pH 7.0 for 90 min. After three washes in the same buffer, the material was stored in buffer at 4~ overnight and the next day postfixed for 2 h in 1% OsO4 in phosphate buffer. After a wash for 15 min in buffer, the cells were dehydrated in a graded ethanol series (30 min in each concentra- tion), washed twice in absolute ethanol, once in propytenoxide, and thereafter infiltrated with Spurr's low viscosity epoxy resin (Spurr 1969). The resin was polymerized at 70~ overnight in flat embedding molds. The material was sectioned at 2-5 txm thick- ness with a glass knife, using a Reichert OmU3 ultramicrotome. The sections were placed on water droplets on slides which were then baked on a hot plate. Permanent slides were made by mount- ing in Eukitt (acrylic resin in xylene, O. Kindler, Freiburg, Ger- many). The sections were analyzed under phase contrast optics in a Zeiss Axioplan microscope.
Results
The in vitro development of the barley zygote
The barley embryo sac is of the Polygonum type (Ma- heshwari 1950). The central cell constitutes mainly the embryo sac and contains a giant vacuole surrounded by a thin band of cytoplasm. The two central or polar nu- clei are located next to the synergids and the egg cell. In the barley cultivar Bomi grown at 18/13~ day/night temperature with a 16 h light period, the egg is fertilized 45 rain after pollination. At 55-60 rain after pollination, the two polar nuclei and the associated sperm nucleus
51
move away from the egg cell and into the central cell (Engell 1989). The absence of polar nuclei associated with the egg apparatus thus provides an excellent con- trol indicating that fertilization has taken place. This can easily be seen in dissected ovaries using a dissection microscope equipped with transmitted light (Holm et al. 1994).
Initial experiments were directed towards excising and culturing the micropylar tip of the ovule. By this approach, the egg would also be accessible for manipu- lation. The ovule tips were cultured floating on liquid Kao 90 medium with 20% Ficoll. The addition of Ficoll proved to be important , since the increased density of the medium prevented the ovule tip from sinking into the medium, in which case the development of embryo- like structures was generally arrested. Several series of experiments revealed that about 20% of the egg cells of Igri as well as Alexis developed into embryo-like struc- tures which produced plants. Ovule tips with embryos ranging in stage from bicellular to 0.2-0.3 mm in length were also cultured, and there was an almost linear, pos- itive correlation between the time after pollination and the frequency of embryo formation.
Cultivation of ovule tips on solid agarose-Kao 90 medium resulted in a slight increase in the frequency of embryo-like structures, but the structures which devel- oped appeared more embryo-like than when grown on liquid medium. It was also apparent that the egg was often damaged when cutting off the micropylar tip of the ovule. To improve embryo quality and to minimize dissection-induced damage, all subsequent experiments were performed on whole ovules. This meant that the egg cell would only be accessible by penetrating the in- teguments and the nucellus layers.
Removal of one integument drastically improved the optical resolution of the egg apparatus, in particular if interference contrast microscopy was performed. When both integuments were dissected away from the mi- cropylar tip, it was possible to see details such as the nucleus of the egg at a high resolution (Fig. 1A) in most of the mounted ovaries. After a few hours of culture, the nucellus tissue became more opaque and the further de- velopment of the egg and the embryo-like structures could only be moni tored in dissected ovules or in fixed and sectioned material. The series of photographs in Fig. 2 illustrates the development of the egg into em- bryogenic cell masses. Polyembryony was often encoun- tered and separation of cells or cell clusters giving rise to two or more embryo-like structures may occur early in development (Fig. 2B). Endosperm formation was not observed but the antipodal cells appeared to stay alive for a few days. At later stages of culture, the developing structures often consisted of aggregates of cells that were either completely or partly separated from each other (Fig. 2C,D). The smaller aggregates and the outer layers of the larger aggregates consisted of cells with dense cytoplasm, large nuclei, and few vacuoles, while the central part of the large aggregates was parenchy- ma-like with large vacuoles.
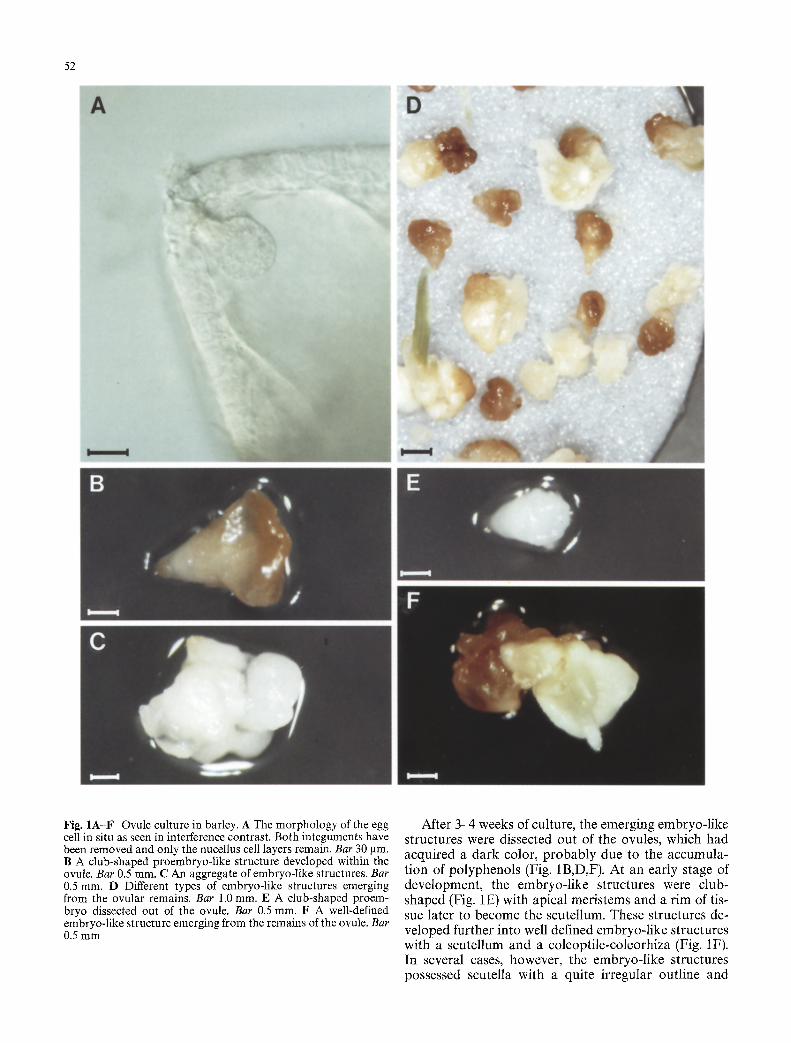
52
Fig. 1A-F Ovule culture in barley. A The morphology of the egg cell in situ as seen in interference contrast. Both integuments have been removed and only the nucellus cell layers remain. Bar 30 gin. B A club-shaped proembryo-like structure developed within the ovule. Bar 0.5 ram. C An aggregate of embryo-like structures. Bar 0.5 ram. D Different types of embryo-like structures emerging from the ovular remains. Bar 1.0 mm. E A club-shaped proem- bryo dissected out of the ovule. Bar 0.5 ram. F A well-defined embryo-like structure emerging from the remains of the ovule. Bar 0.5 mm
After 3 4 weeks of culture, the emerging embryo-l ike structures were dissected out of the ovules, which had acquired a dark color, p robab ly due to the accumula- tion of polyphenols (Fig. 1B,D,F). At an early stage of development , the embryo-l ike structures were club- shaped (Fig. 1E) with apical meris tems and a r im of tis- sue later to become the scutellum. These structures de- veloped further into well defined embryo-l ike structures with a scutellum and a coleopti le-coleorhiza (Fig. 1F). In several cases, however, the embryo-l ike structures possessed scutella with a quite irregular outline and
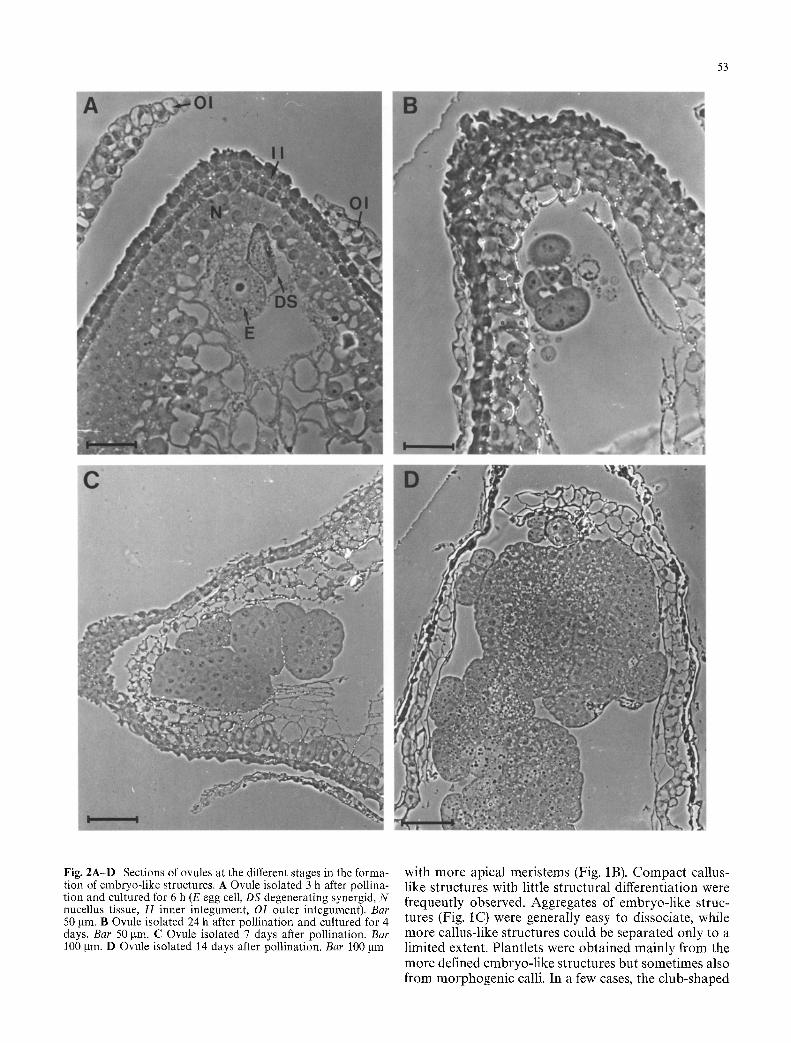
53
Fig. 2A-D Sections of ovules at the different stages in the forma- tion of embryo-like structures. A Ovule isolated 3 h after pollina- tion and cultured for 6 h (E egg cell, DS degenerating synergid, N nucellus tissue, I I inner integument, OI outer integument). Bar 50 gin. B Ovule isolated 24 h after pollination and cultured for 4 days. Bar 50 gin. C Ovule isolated 7 days after pollination. Bar 100 gin. D Ovule isolated 14 days after pollination. Bar 100 gm
with more apical meris tems (Fig. 1B). Compac t callus- like structures with little structural differentiation were frequently observed. Aggregates of embryo- l ike struc- tures (Fig. IC) were generally easy to dissociate, while more callus-like structures could be separated only to a limited extent. Plantlets were obta ined mainly f rom the more defined embryo- l ike structures but sometimes also f rom morphogen ic calli. In a few cases, the c lub-shaped
54
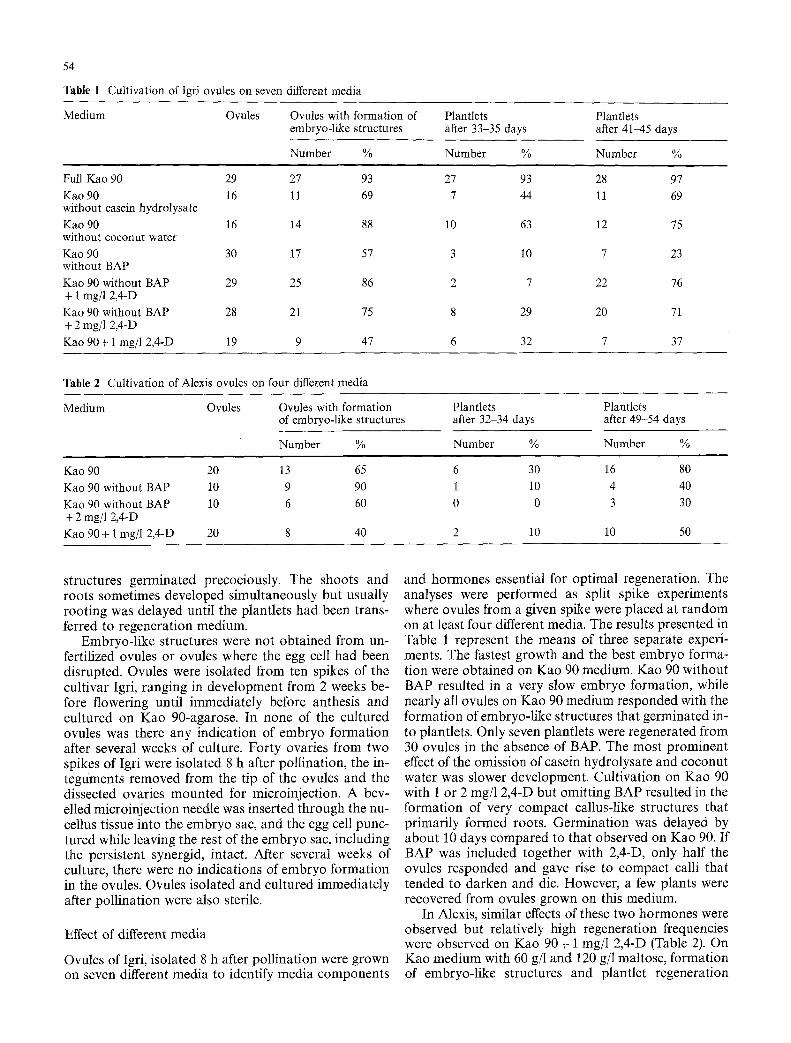
Table 1 Cultivation of Igri ovules on seven different media
Medium Ovules Ovules with formation of embryo-like structures
Number %
Plantlets Plantlets after 33-35 days after 41-45 days
Number % Number %
Full Kao 90 29 27 93 Kao 90 16 11 69 without casein hydrolysate Kao 90 16 14 88 without coconut water Kao 90 30 17 57 without BAP Kao 90 without BAP 29 25 86 + 1 rag/1 2,4-D Kao 90 without BAP 28 21 75 + 2 mg/1 2,4-D Kao 90+ 1 mg/l 2,4-D 19 9 47
27 93 28 97 7 44 l l 69
10 63 12 75
3 10 7 23
2 7 22 76
8 29 20 71
6 32 7 37
Table 2 Cultivation of Alexis ovules on four different media
Medium Ovules Ovules with formation of embryo-like structures
Number %
Plantlets Plantlets after 32-34 days after 49-54 days
Number % Number %
Kao 90 20 13 65 Kao 90 without BAP 10 9 90 Kao 90 without BAP 10 6 60 + 2 rag/1 2,4-D Kao 90 + 1 rag/1 2,4-D 20 8 40
6 30 16 80 1 10 4 40 0 0 3 30
2 10 10 50
structures germinated precociously. The shoots and roots sometimes developed simultaneously but usually rooting was delayed until the plantlets had been trans- ferred to regeneration medium.
Embryo-l ike structures were not obtained from un- fertilized ovules or ovules where the egg cell had been disrupted. Ovules were isolated from ten spikes of the cultivar Igri, ranging in development from 2 weeks be- fore flowering until immediately before anthesis and cultured on Kao 90-agarose. In none of the cultured ovules was there any indication of embryo formation after several weeks of culture. Forty ovaries from two spikes of Igri were isolated 8 h after pollination, the in- teguments removed from the tip of the ovules and the dissected ovaries mounted for microinjection. A bev- elled microinjection needle was inserted through the nu- cellus tissue into the embryo sac, and the egg cell punc- tured while leaving the rest of the embryo sac, including the persistent synergid, intact. After several weeks of culture, there were no indications of embryo formation in the ovules. Ovules isolated and cultured immediately after poll ination were also sterile.
Effect of different media
Ovules of Igri, isolated 8 h after pollination were grown on seven different media to identify media components
and hormones essential for optimal regeneration. The analyses were performed as split spike experiments where ovules from a given spike were placed at random on at least four different media. The results presented in Table 1 represent the means of three separate experi- ments. The fastest growth and the best embryo forma- tion were obtained on Kao 90 medium. Kao 90 without BAP resulted in a very slow embryo formation, while nearly all ovules on Kao 90 medium responded with the formation of embryo-like structures that germinated in- to plantlets. Only seven plantlets were regenerated from 30 ovules in the absence of BAP. The most prominent effect of the omission of casein hydrolysate and coconut water was slower development. Cultivation on Kao 90 with 1 or 2 rag/1 2,4-D but omitting BAP resulted in the formation of very compact callus-like structures that primarily formed roots. Germinat ion was delayed by about 10 days compared to that observed on Kao 90. If BAP was included together with 2,4-D, only half the ovules responded and gave rise to compact calli that tended to darken and die. However, a few plants were recovered from ovules grown on this medium.
In Alexis, similar effects of these two hormones were observed but relatively high regeneration frequencies were observed on Kao 90 + 1 mg/l 2,4-D (Table 2). On Kao medium with 60 g/l and 120 g/1 maltose, formation of embryo-like structures and plantlet regeneration
Alexis
100%'
o ``9` 9 0 % ' O ~'~ o9 80%'
O9 ,... ~ 70%
I-~ 600/0
~ 40% o9~ -5 6 3o0~
~= ~ 20~ 0 . o
10%
O% 2-6 6-10 10-14 14-18 18-24 24-30
1~176 ~ - " l
~ 4~ IMl IH_] LI~] lll~ II -~'i 3o%13 IH] I ~ Illl IHJ II
2~ IH] 1H] I1~1 IH] II o 1 ~ 1 7 6 1 7 6 IIEI III-
2-6 6-10 10-14 14-18 18-24 24-30
I~ 2.50" e-
=_= o= 2,00. O O9
.~ ,,- _~ 1.s0.
,= ~ o l O O w,-. O9
d ~ 0,50. z
0,00 2-6 6-10 10-14 14-18 18-24 24-30
"~ 0
z i i i
2-6 6-10 10-14 14-18 18-24 24-30
Hours after Pollination
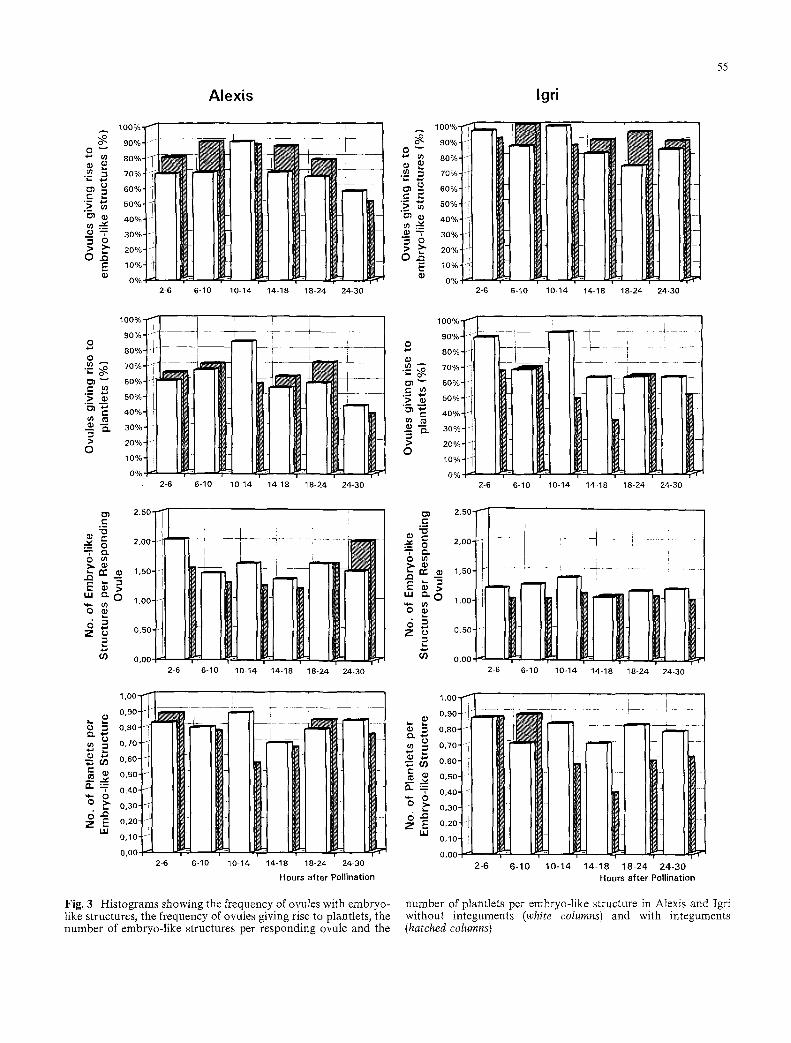
Fig. 3 Histograms showing the frequency of ovules with embryo- like structures, the frequency of ovules giving rise to plantlets, the number of embryo-like structures per responding ovule and the
55
Igri
A
o9~
o ~ E
100%"
90%'
80%'
70%
60%"
50%'
40%
30%-
20%-
10%-
OOA 2 - 6 6-10 10-14 14-18 18-24 24-30
9 O9 A
"5 0
100%-
90%-
80%.
70%.
60%"
50%"
40%.
30%
20%-
10%-
0% 2-6 6-10
l - i : : �84
10-14 14-18 18-24 24-30
.=_= J
i
0
ILl
e~ O9
t r m
2
2 , 5 0 -
2,00-
1,50-
1,00-
0,50-
0,00- 2-6 6-10 10-14 14-18 18-24 24-30
= . .
&
J 4-,
E "6
09
LU
2-6 6-10 10-14 14-18 18-24 24-30 Hours after Pollination
number of plantlets per embryo-like structure in Alexis and Igri without integuments (white columns) and with integuments (hatched columns)

56
Table 3 Summary table: frequency of formation of embryo-like structures and plantlet regeneration in Alexis and Igri with and without integuments in micropylar region
Cultivar Ovules Ovules giving rise to Embryo-like structures Plantlets
Embryo-like Plantlets Per Per responding Per embryo- Per responding structures % % ovule ovule like structure ovule
Igri 232 87 without integuments
Igri 205 92 with integuments
Alexis 246 72 without integuments
Alexis 261 77 with integuments
72 1.05 1.21 0.80 1.16
55 0.94 1.02 0.62 1.05
63 1.18 1.65 0.81 1.53
61 1.12 t.46 0.75 1.39
were also frequent. It remains to be determined whether regeneration frequencies are inferior or superior to those found on Kao 90.
Relationship between the age of the egg and regeneration frequency
The first division of the barley zygote takes place 24- 28 h after fertilization. A large-scale experiment was carried out to evaluate if the frequency of formation of embryo-like structures and plantlet regeneration de- pended on the developmental stage of the zygote.
The histograms in Fig. 3 illustrate the frequencies of ovules giving rise to embryo-like structures and plants, the number of embryo-like structures per responding ovule, and the number of plants per embryo-like struc- ture from 2 to 30 h after pollination in dissected and intact ovules of Igri and Alexis. The observations were grouped for 4 h intervals for the period from 2 to 18 h after pollination while the period from 18 h to 30 h was divided into 6 h intervals. This was performed to facili- tate the statistical analysis of the data and to minimize variation originating from small sample sizes. The means of the data for each of the four experimental con- ditions are given in Table 3.
Statistical analysis using a Tukey's Studentized Range (HSD) test for variable showed the following: in Igri the frequency of ovules with embryo-like structures was significantly higher than in Alexis (significant at the 1% level) but there was no effect of removing the integu- ments. In contrast, the number of embryo-like struc- tures per ovule was significantly higher in Alexis than in Igri. In Alexis 68, 19, 7, and 4% of the ovules contained 1, 2, 3, and 4 embryo-like structures respectively, while for Igri the corresponding frequencies were 87, 12, 1, and 0%. Individual embryo-like structures of the aggregates were as viable as those developed as single structures in an ovule. In a few cases, up to seven embryos and plantlets were obtained from single ovules and all ger- minated into plantlets. There was in both cultivars a significant increase in the number of embryo-like struc- tures per ovule when integuments were removed. Re-
moval of the integuments also resulted in an improved quality of the embryos as measured by their ability to form plants.
Minor differences among the four experimental con- ditions, (better response in Igri, a greater tendencey to polyembryony in Alexis, and the increase in polyembry- ony as well as embryo quality following removal of the integuments) tended to balance each other. There was thus no significant difference between the two varieties with respect to the frequency of ovules giving rise to plants and the effect of removing the integuments was only at the limit of the 5% significance level. Finally, statistical analysis did not reveal significant differences between the six intervals from 2 to 30 h after pollination for any of the measured parameters with the exception of the parameter "Ovules giving rise to plantlets" (signif- icant at the 5% level) where there was a reduction in frequency in the interval from 24 to 30 h after pollina- tion. It can be concluded from the combined data on the culture of 944 ovules of Igri and Alexis that 81% of the ovules gave rise to embryo-like structures and 63% to plants. On average, 1.3 embryo-like structure formed per responding ovule and 70% of these germinated into plantlets.
Fertility of the regenerated plants
Several hundred plantlets were regenerated from each of the two genotypes but albino plants were never ob- served among the primary regenerants. In a few cases only did an albino shoot develop from callus present at the base of the primary regenerant.
On average, 75% of the regenerated plantlets sur- vived and developed further. In nearly all cases these plantlets also survived the vernalization (Igri) and the transfer to soil. Most regenerated plantlets were dis- carded, but 43 Igri and 25 Alexis plants were grown to maturity in the greenhouse. The fertility of the individu- al spike was measured by counting the number of florets with and without grains. In Alexis, 19, 65, 15, 1, and 0% of the spikes showed a fertility of 100, 75-100, 50-75, 25 50, and 0-25% while in Igri the corresponding fre-

quencies were 8, 41, 21, 14, and 17%. The slightly lower fertility in Igri resulted because all spikes of each plant were evaluated for their fertility, while in Alexis only the first 15 spikes formed on each plant were included. Igri regenerants formed about three times as many spikes per plant as seed-grown control material and spikes ap- pearing late generally had low fertility. The high num- ber of spikes resulted in part from a cutting back of the Igri regenerants prior to vernalization which facilitates the production of more tillers. It may also be attributed to the presence of multiple apical meristems in the em- bryo-like structures. The fertility of the Alexis and Igri regenerants was similar to that observed in seed-grown control material and there was only a slight variation in fertility among the regenerants. No completely sterile plants were found.
Regeneration of plants from ovule cultures of the wheat zygote
A few experiments were carried out to ascertain if the same ovule culture technique could be applied to wheat. The wheat cultivar Walter was grown and the ovules processed and cultured as described for barley. In three experiments spikes were hand-pollinated and the ovaries removed (26, 8, and 8 h after pollination). Thir- teen, 18, and 14 ovules were cultured and in each series an embryo-like structure formed. The structures germi- nated readily and the three plants were grown to matu- rity. All were morphologically normal and fertile.
Discussion
The present study has devised an ovule culture tech- nique that allowed an efficient regeneration of the bar- ley zygote into fertile plants. Similar regeneration fre- quencies were obtained from ovules cultured immedi- ately after fertilization until the formation of bicellular embryos, while unpollinated ovules did not give rise to embryos and plants. Removal of the integuments around the micropylar region of the ovule induced a slightly higher frequency of polyembryony, and the em- bryo-like structures from the dissected ovules tended to give rise to more plants than embryo-like structures from undissected ovules. The culture technique may al- so be largely genotype independent, since similar regen- eration frequencies were observed in the two cultivars Igri and Alexis. These two cultivars show in contrast very difference performances in microspore culture where a mean of 11.7 green plants and 0.1 albino plants could be regenerated per anther of Igri while 0.06 green plants and 0.61 albino plants were regenerated per an- ther of Alexis (P. Mouritzen and P.B. Holm, manuscript submitted). A similar difference was observed when whole anthers were cultured (P. Mouritzen and P.B. Holm, unpublished observation). While albinism in the regenerants was a major problem in the microspore-
57
derived plants of Alexis, albino plants were never ob- served among the primary regenerants from the egg cell.
These data confirm and extend the results and con- clusions presented by T6pfer and Steinbiss (1985). In their study, small samples of ovules were cultured from 11 different spring and winter cultivars using liquid Bar- ley Medium II (Norstog 1973). Poorly differentiated em- bryos were found in 5-28% of the ovules and about 20% of these developed into plants. Embryos could be obtained from ovules isolated 90 rain after pollination while unpollinated ovules were sterile. Cultivation of ovules of cross-pollinated material and genetic analysis of the regenerants proved that the regenerants arose from the zygote and not from maternal tissue.
The significant increase in the formation of embryo- like structures and plantlets obtained in this study com- pared to that obtained by T6pfer and Steinbiss (1985) may to a large extent can be attributed to cultivation on solid medium. In our experience, submerged ovule cul- tures seldom produced embryogenic structures of a good quality, while acceptable results were obtained if the ovules were floating on the surface of Kao 90 + 20% Ficoll. Cultivation on Whatman filter paper discs was also believed to have an effect since it reduced the accu- mulation of liquid around the ovules. The different me- dia used in the two studies may also be important. T6pfer and Steinbiss (1985) found no effect from adding casamino acids, coconut water, or BAP, while these proved important in the present study. Casamino acids, coconut water and cytokinins are often used for ovule culture (for review, see Rangan 1982) and particular concentration ranges of coconut water, casamino acids, glutamine, ammonium ions, and abscisic acid have also been found important for sustaining the growth and development of isolated barley embryos (Ziebur et al. 1950; Norstog 1961, 1970, 1973; Norstog and Klein 1972; Cameron-Mills and Duffus 1977; Umbeck and Norstog 1979).
Unfertilized barley eggs in ovule culture were not re- generable. The same conclusion was reached by T6pfer and Steinbiss (1985). Isolated, unfertilized protoplasts of maize (Kranz et al. 1991) and barley (Holm et al. 1994) were never observed to divide. In contrast, in vitro fertil- ized egg protoplasts of maize (Kranz et al. 1991) and in vivo fertilized egg protoplasts of barley (Holm et al. 1994), grown in medium only, divided a few times and formed microcalli that thereafter degenerated. This indi- cates that division of the egg cell requires the stimulus of fertilization. However, haploids can in some species be regenerated from ovules and ovaries, isolated and cul- tured at different developmental stages (see Yang and Zhou 1982 for review). In barley, haploid plants have been regenerated from ovaries of florets where the pol- len was either binucleate or young trinucleate (San Noeum 1979; Castillo and Cistu6 1993). tn the former study the best response was obtained from a doubled haploid, where 0.11% of the ovaries produced one or more plants. The embryogenic structures appeared to originate primarily from the egg but regeneration of

58
structures from antipodals and synergids was also re- ported. Castillo and Cistu6 (1993) obtained callus for- mation from about 8% of cultured barley ovaries when using N6 medium supplemented with 4-chloro-2- methylphenoxyacetic acid, indole-3-acetic acid, 6-ben- zyladenine, 90 g/1 sucrose, and 100 g/1 coconut water. Plants could be regenerated from half the calli and were confirmed to be haploid by chromosome counts. The best response was obtained when using ovaries from flowers at the trinucleate stage of pollen.
Protoplasts of in vitro and in vivo fertilized egg of maize and barley respectively could only undergo an embryogenic development if they were co-cultured with a maize suspension or barley microspores undergoing embryogenesis (Kranz and L6rz 1993; Holm et al. 1994). In barley, co-cultivation was required for up to 30 days, at which stage 1-2 mm large embryo-like structures had developed. The development of the egg in ovule culture, which is very similar to in vivo development, must thus depend upon a similar support from the ovule tissues. The fact that an embryo-like development of the fertil- ized egg also can be achieved by culturing tips of ovules further suggests than only a minimum of integument and nucellus tissue is required. In addition, the embryo- genic development of the egg does not appear to depend on the egg cell being left undisturbed in its original loca- tion. The mechanical isolation of the egg protoplasts involved a puncturing of the giant vacuole of the central cell followed by a gentle tapping on the micropyle with a pair of forceps. This resulted in the liberation of the egg as a spherical protoplast into the central cell and the egg protoplast could be squeezed out of the ovule fol- lowing removal of the nucellus tissues in the micropylar region (Holm et al. 1994). Ovules treated in a similar way and thereafter cultured gave rise to embryo-like structures at the same frequency as ovules where the egg had been left in situ (unpublished observation).
The present results show that it is possible to affect the embryogenic development of the egg by changing the media and adding hormones such as BAP and 2,4-D. This suggests that ovule cultures may be used as sensitive bioassays for analyzing the effect of particular compounds expected to affect embryogenesis. The fact that the cells of the egg apparatus and the polar nuclei can be seen with high optical resolution in the dissected ovules may also allow a manipulation of these cells by, e.g., microinjection where DNA, sperm cells or other materials are introduced into the egg, the synergids, and the central cell. Microinjection into the egg is further facilitated by the fact that the barley egg in vivo is large- ly a protoplast that is covered only by wall material in the micropylar region (Holm et al. 1994). Since the re- generation from the zygote appeared to be independent of its developmental stage from fertilization to the first division of the zygote, this technique also allows the manipulation of the egg at particular stages of its cell cycle.
Acknowledgements We are indebted to Bente Jensen and Judita Gartner for expert technical assistance and to Marie Dam for taking excellent care of the plants. Ann Soft Steinholz is thanked for producing the color prints and Jan Ruud Hansen for perform- ing the statistical analysis. Seeds of the wheat variety Waiter were generously provided by Dr. Sven Bode Andersen, Department of Crop Science, The Royal Veterinary and Agriculture University, Denmark. The work was supported by Eureka grant EU 270 and is ABIN (Adaptation of Barley for Industrial Needs) publication no. 139.
References
Cameron-Mills V, Duffus CM (1977) The in vitro culture of imma- ture barley embryos on different culture media. Ann Bot 41:1117-1127
Campenot MK, Zhang G, Cutler AJ, Cass DD (1992) Zea mays embryo sacs in culture. I. Plant regeneration from 1 day after pollination embryos. Am J Bot 79:1369-1373
Castillo AM, Cistu~ L (1993) Production of gynogenic haploids of Hordeum vulgare L. Plant Cell Rep 12:139-143
Comeau A, Nadeau P, Plourde A, Simard R, Maes O, Kelly S, Harper L, Lettre J, Landry B, St-Pierre CA (1992) Media for the in ovulo culture of proembryos of wheat and wheat-deri- ved interspecific hybrids or haploids. Plant Scie 81:117-125
Engell K (1989) Embryology of barley: time course and analysis of controlled fertilization and early embryo formation based on serial sections. Nord J Bot 9:265580
Holm PB, Knudsen S, Mouritzen P, Negri D, Olsen FL, Rou6 C (1994) Regeneration of fertile barley plants from mechanically isolated protoplasts of the fertilized egg cell. Plant Cell 6: 531-543
Huang B-Q, Russell SD (1992) Female germ unit: organization, isolation and function. Int Rev Cytol 140:233-293
Kao KN, Michayluk MR (1975) Nutritional requirements for growth of Vicia hajastana cells and protoplasts at a very low population density in liquid media. Planta 126:105-110
Keijzer CJ, Reinders MC, Leferik-ten Kloster HB (1988) A micro- manipulation method for artificial fertilization in Torenia. In: Cresti M, Gori P, Pacini E (eds) Sexual reproduction in higher plants. Springer, Berlin Heidelberg New York, pp 119-124
Kranz E, L6rz H (1993) In vitro fertilization with isolated single gametes results in zygotic embryogenesis and fertile maize plants. Plant Cell 5:739-746
Kranz E, Bautor J, L6rz H (1991) In vitro fertilization of single isolated gametes of maize mediated by electrofusion. Sex Plant Reprod 4:12-16
Maheshwari P (1950) An introduction to the embryology of an- giosperms. McGraw-Hill, New York
Mathias RJ (1987) Plant microinjection techniques. In: Setlow JK (ed) Genetic engineering principles and methods. Vol 9, Ple- num Press, New York, pp 199-277
M61 R, Matthys-Rochon E, Dumas C (1993) In vitro culture of fertilized embryo sacs of maize: zygotes and two-celled em- bryos can develop into plants. Planta 189:213-217
Murashige T, Skoog F (1962. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:472497
Norstog K (1961) The growth and differentiation of cultured bar- ley embryos. Am J Bot 48:876-884
Norstog K (1970) Induction of embryolike structures by kinetin in cultured barley embryos. Dev Biol 23:665-670
Norstog K (1973) New synthetic medium for the culture of prema- ture barley embryos. In Vitro 8:307-308
Norstog K, Klein RM (1972) Development of cultured barley embryos. II. Precocious germination and dormancy. Can J Bot 50:1887-1894

59
Olsen FL (1987) Induction of microspore embryogenesis in cultur- ed anthers of Hordeum vulgate. The effects of ammonium nitra- te, glutamine and asparagine as nitrogen sources. Carlsberg Res Commun 52:393-404
Rangan TS (1982) Ovary ovule and nucellus culture In: Johri BM (ed) Experimental embryology of vascular plants. Springer, New York, pp 105-129
San Noeum L-H (1979) In vitro induction of gynogenesis in higher plants. Proceedings Conference on Broadening of the Genetic Base of Crops, Wageningen, 1978. Pudoc, Wageningen, pp 327-329
Spurr AR (1969) A low-viscosity epoxy resin embedding medium for electron microscopy. J Ultrastruct Res 26:31-43
Steinbiss H-H, Stabel P, T6pfer R, Hirtz RD, Schell J (1985) Transformation of plant cells by microinjection of DNA. In: Chapmann GP, Mantell SH, Daniels RW (eds) The experi-
mental manipulation of ovule tissue. Longman, New York, pp 64-75
T6pfer R, Steinbiss H-H (1985) Plant regeneration from cultured fertilized barley ovules. Plant Sci 41:49-54
Umbeck PF, Norstog K (1979) Effects of abscisic acid and ammo- nium ion on morphogenesis of cultured barley embryos. Bull Torrey Bot Club 106:110-116
Wagner VT, Dumas C, Mogensen HL (1990) Quantitative three- dimensional study on the position of the female gametophyte and its constitutive cells as a prerequisite for corn (Zea mays) transformation. Theor Appl Genet 79:72~76
Yang HY, Zhou C (1982) In vitro induction of haploid plants from unpollinated ovaries and ovules. Theor Appl Genet 63:97-104
Ziebur NC, Brink RA, Graft LH, Stahmann MA (1950) The effect of casein hydrolysate on the growth in vitro of immature Hor- deum embryos. Am J Bot 37:144-148
Copyright © 2022 FDOKUMEN




















