
Ras regulates neuronal polarity via the PI3-kinase/Akt/GSK-3β/CRMP2 pathway

Taurocholate feeding prevents CCl4-induced damage oflarge cholangiocytes through PI3-kinase-dependent mechanism
LUCA MARUCCI,6* GIANFRANCO ALPINI,1,2,4* SHANNON S. GLASER,3
DOMENICO ALVARO,5 ANTONIO BENEDETTI,6 HEATHER FRANCIS,1
JO LYNNE PHINIZY,1 MARCO MARZIONI,2 JEREMY MAULDIN,2
JULIE VENTER,2 BRANDY BAUMANN,2 LAURA UGILI,6 AND GENE LESAGE1
1Department of Internal Medicine and 2Medical Physiology, 3Division of Research and Education,Scott & White Hospital and The Texas A&M University System Health Science Center,College of Medicine and 4Central Texas Veterans Health Care System, Temple, Texas 76504;5Division of Gastroenterology, University of Rome, La Sapienza, Rome, Italy 00100; and6Department of Gastroenterology, University of Ancona, Ancona, Italy 60131Submitted 24 June 2002; accepted in final form 23 September 2002
Marucci, Luca, Gianfranco Alpini, Shannon S. Gla-ser, Domenico Alvaro, Antonio Benedetti, HeatherFrancis, Jo Lynne Phinizy, Marco Marzioni, JeremyMauldin, Julie Venter, Brandy Baumann, Laura Ugili,and Gene LeSage. Taurocholate feeding prevents CCl4-induced damage of large cholangiocytes through PI3-kinase-dependent mechanism. Am J Physiol Gastrointest LiverPhysiol 284: G290–G301, 2003. First published October 2,2002; 10.1152/ajpgi.00245.2002.—Bile acids are cytoprotec-tive in hepatocytes by activating phosphatidylinositol-3-ki-nase (PI3-K) and its downstream signal AKT. Our aim was todetermine whether feeding taurocholate to CCl4-treated ratsreduces cholangiocyte apoptosis and whether this cytoprotec-tive effect is dependent on PI3-K. Cholangiocyte prolifera-tion, secretion, and apoptosis were determined in cholangio-cytes from bile duct ligation (BDL), CCl4-treated BDL rats,and CCl4-treated taurocholate-fed rats. In vitro, we testedwhether CCl4 induces apoptosis and whether loss of cholan-giocyte proliferation and secretion is dependent on PI3-K.The CCl4-induced cholangiocyte apoptosis and loss of cholan-giocyte proliferation and secretion were reduced in CCl4-treated rats fed taurocholate. CCl4-induced cholangiocyteapoptosis, loss of cholangiocytes secretion, and proliferationwere prevented by preincubation with taurocholate. Tauro-cholate cytoprotective effects were ablated by wortmannin.Taurocholate prevented, in vitro, CCl4-induced decrease ofphosphorylated AKT protein expression in cholangiocytes.The cytoprotective effects of taurocholate on CCl4 effects oncholangiocyte proliferation and secretion were abolished bywortmannin. Taurocholate protects cholangiocytes fromCCl4-induced apoptosis by a PI3-K-dependent mechanism.Bile acids are important in the prevention of drug-inducedductopenia in cholangiopathies.
cytoprotection; biliary epithelium; bile acids
CHOLANGIOCYTES LINE THE INTRAHEPATIC biliary tree, aninterconnecting network of actively secreting conduits
extending from the ducts of Hering to the septal bileducts (9, 26). Cholangiocytes participate in hormone-regulated ductal secretion (8, 33, 35). Secretin stimu-lates ductal secretion (8, 33, 35) by interaction withbasolateral secretin receptors [expressed only bycholangiocytes in rat liver (13)] through an increase incAMP levels (6, 10, 12, 27, 33, 35). In the normal state,cholangiocytes are mitotically dormant (6, 26, 33) butproliferate in response to liver injury/toxins includingbile duct ligation (BDL) (8, 22) or acute carbon tetra-chloride (CCl4) administration (35). In these models ofductal hyperplasia, cholangiocyte proliferation is asso-ciated with changes in basal and secretin-stimulatedductal secretion (8, 22, 26, 35). In BDL rats, cholangio-cyte proliferation is associated with an increase inbasal and secretin-stimulated ductal secretion, whereasin rats with ductopenia (after acute CCl4 administra-tion), reduced cholangiocyte proliferation is coupledwith inhibition of secretin-stimulated ductal secretion(8, 22, 26, 35). Thus the choleretic or lack of cholereticresponse to secretin may be an index to functionallyevaluate cholangiocyte hyperplasia (6–9, 22, 33) orductopenia (9, 35), respectively.
We have developed techniques for isolating subpopu-lations of small and large cholangiocytes (6, 10, 12, 35)and intrahepatic bile duct units (4) from different por-tions of the rat intrahepatic biliary epithelium andhave shown that cholangiocytes are morphologicallyand functionally heterogeneous with secretin-stimu-lated ductal secretion restricted to large cholangiocytes(lining large ducts) but not small cholangiocytes (liningsmall bile ducts) (4, 6, 10, 12, 35). In BDL rats, cholan-giocyte proliferation and increased ductal secretionoccur in large but not small cholangiocytes (6, 26). Asingle dose of CCl4 to BDL rats induces a transientcholangiocyte loss (by apoptosis) only in large ducts, an
*L. Marucci and G. Alpini share the first authorship.Address for reprint requests and other correspondence: G.
LeSage, Professor of Medicine, Univ. of Texas Houston MedicalSchool, 6431 Fannin St., MSB 4.234, Houston TX 77030 (E-mail:[email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be herebymarked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734solely to indicate this fact.
Am J Physiol Gastrointest Liver Physiol 284: G290–G301, 2003.First published October 2, 2002; 10.1152/ajpgi.00245.2002.
0193-1857/03 $5.00 Copyright © 2003 the American Physiological Society http://www.ajpgi.orgG290

event associated with a reduction of proliferative andsecretory activity in large cholangiocytes (35). In con-trast, small cholangiocytes are resistant to apoptosisand, de novo, proliferate and secrete in response toCCl4 compensating for loss of large cholangiocyte func-tion (35). The different sensitivity of small and largecholangiocytes to CCl4 toxicity may be due to the ex-pression of the cytochrome P-4502E1 (the enzyme thatinitiates CCl4 hepatotoxicity) (15) in large but notsmall cholangiocytes (35). This unique animal modelpresents several features common to human pathology(9). In fact, loss and/or damage of large interlobularducts by degenerative or inflammatory processes is amajor factor in the pathogenesis of vanishing bile ductsyndromes [e.g., primary biliary cirrhosis (PBC) andprimary sclerosing cholangitis (PSC)] (9). Further-more, several studies suggest that apoptosis is an im-portant mechanism of cholangiocyte death resulting inductopenia (35, 39).
Bile acids are internalized into cholangiocytes by theapically located Na�-dependent bile acid transporter(ABAT) (5, 28) thus modulating cholangiocyte secretionand growth (3, 7). In vitro, taurocholate increased DNAsynthesis, and secretin-stimulated cAMP levels incholangiocytes (3). Taurocholate uptake into cholangio-cytes was required for taurocholate stimulation of growthand secretion (3). Chronic in vivo feeding of taurocholateto normal rats increased secretin-stimulated ductal se-cretion and cholangiocyte proliferation (7).
The regulatory protein phosphatidylinositol-3-ki-nase (PI3-K) is involved in different signaling path-ways and controls key functions of a number of cells(17, 36). PI3-K is considered one of the main intracel-lularfactorsresponsibleforthetransmissionofantiapop-totic signals and controlling the survival of cells (19,24). For instance, overexpression of PI3-K in cells isaccompanied by a marked antiapoptotic effect and asignificant increase in cell survival following radiation(19, 24). PI3-K have also been implicated in controllingDNA synthesis and cell cycle (1). A number of studieshas shown that bile acids may affect PI3-K activity (36,42, 46). The hydrophobic bile acid taurochenodeoxy-cholate activates PI3-K-dependent survival pathwaysin hepatocytes, which prevent its inherent toxicity (42).In addition, cAMP-mediated cytoprotection againstbile acid-induced apoptosis in cultured rat hepatocytesappears to involve PI3-K (46). In view of these findings,we tested the hypothesis that in BDL rats, tauro-cholate, both in vivo (by chronic feeding) and in vitro,prevents the CCl4-induced damage of large cholangio-cyte functions through a PI3-K-dependent pathway.
MATERIALS AND METHODS
Materials
Reagents were purchased from Sigma (St Louis, MO) un-less otherwise indicated. Control chow (i.e., AIN 76) andchow containing 1% taurocholate with AIN 76 were preparedfrom Dyets (Bethlehem, PA). Control chow (AIN 76) has thesame composition of rat chow containing 1% taurocholate butit does not contain taurocholate. Porcine secretin was pur-
chased from Peninsula Laboratories (Belmont, CA). RIA kitsfor the determination of intracellular cAMP levels were pur-chased from Amersham (Arlington Heights, IL). The mouseanti-cytokeratin 19 (CK-19) antibody was purchased fromAmersham. The monoclonal mouse antibody against PCNAwas purchased from DAKO (Kyoto, Japan). The monoclonalantibody (C-20) against total Akt was purchased from SantaCruz Biotechnology (Santa Cruz, CA). The affinity-purifiedgoat polyclonal antibody mapping at the COOH terminus ofH3 histone of human origin was purchased from Santa CruzBiotechnology. The monoclonal antibody against phosphory-lated Akt (phospho-Akt, Ser473, 4E2) was purchased fromCell Signaling Technology (Beverly, MA). The nuclear dye4,6-diamidino-2-phenylindole (DAPI) was purchased fromMolecular Probes (Eugene, OR). A biotinylated antibody re-acting with annexin-V was purchased from Boehringer-Mannheim (Mannheim, Germany).
Animal Model
Male Fisher 344 rats (150–175 g) were purchased fromCharles River (Wilmington, MA), maintained in a tempera-ture-controlled environment (22°C) with a 12:12-h light-darkcycle, and fed ad libitum standard rat chow. Studies wereperformed in 1) rats with BDL or bile duct incannulation[BDI; for bile collection (8)] for 9 days (controls); and 2) 1 wkBDL or BDI rats, which after the intragastric administrationof a single dose of CCl4/mineral oil 1:1 (0.5 ml/100 g body wt),were fed control diet (AIN 76) or 1% taurocholate for 2 daysfor a total of 9 days of BDL or BDI. We omitted the group ofBDL or BDI rats treated with mineral oil (0.5 ml/100 g bodywt), because in previous studies, we have shown (35) thatadministration of a single dose of mineral oil (0.5 ml/100 gbody wt) does not alter cholangiocyte proliferation and secre-tion. The animals were killed on day 2 after CCl4 adminis-tration plus taurocholate or control diet. BDL or BDI wasperformed as described (8). Before each experimental proce-dure, the animals were anesthetized with pentobarbital so-dium (50 mg/kg weight ip).
Cholangiocyte Purification
Pure subpopulations of small and large cholangiocyteswere purified from the selected group of animals as described(4, 6, 10) using a mouse monoclonal antibody against anunidentified membrane antigen expressed by all rat intrahe-patic cholangiocytes (25). Cell number and viability wereassessed by trypan blue exclusion. Cholangiocyte purity wasassessed by histochemistry for �-GT (43).
Studies in BDL Rats Treated in Vivo With CCl4 PlusTaurocholate or Control Feeding
Measurement of transaminase serum levels. We evaluatedthe effect of taurocholate or control feeding on CCl4-inducedliver damage assessed by transaminase serum level determi-nation. In BDL rats and 1-wk-old BDL rats treated with CCl4and subsequently fed 1% taurocholate or control diet, theserum levels of aspartate aminotransferase (AST) and ala-nine aminotransferase (ALT) were measured by commer-cially available kits (Sigma).
In Situ Morphological Studies and TUNEL Analysis
After anesthesia, small pieces of liver tissue were takenfrom each animal and fixed in 10% formalin (pH 7.4). Sec-tions (5-�m thick) were cut from two liver blocks for eachanimal, stained for hematoxylin and eosin, and examinedunder a light microscope equipped with a reticular frame
G291BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

inserted in the ocular lens (35). An in situ cell death detectionkit (Boehringer-Mannheim, Monza, Italy) was also used. Thekit (previously used by us for determining cholangiocyteapoptosis) (35) was used according to the instructions of thevendor. In each liver section, the number of apoptotic bodiespresent in cholangiocytes and the diameter (distance be-tween basement membranes) of the bile duct (in which apop-tosis was observed) were evaluated. In each liver section, atleast 50 nonoverlapping fields were analyzed. The incidenceof apoptosis was expressed as number of apoptotic bodies per100 cholangiocytes.
Measurement of Caspase 8, 9, and 3 Activities
Intracellular activity of caspase 8, 9, and 3 of small andlarge cholangiocytes from the selected groups of animals wasmeasured by enzymatic kits according to the instructionssupplied by the vendor (Medical & Biological Laboratories,Nagoya, Japan). After isolation, cholangiocytes were centri-fuged at 1,500 rpm for 10 min, incubated in lysis buffer on icefor 10 min, and centrifuged at 10,000 g for 10 min. After thesupernatant was centrifuged, that containing the cytosolicfraction, was transferred to a clean tube. For each sample,100 �g of proteins or BSA (negative control) were added to 50�l of 2� reaction buffer. The activity of caspase 8, 9, and 3was measured by proteolytic cleavage of substrates such asDEVD-p-nitoranilide (pNA; caspase 3 substrate), IETD-pNA(caspase 8 substrate), and LEHD-pNA (caspase 9 substrate),respectively, added to each sample. The assay is based on thephotometric detection of the chromophore pNA after cleavagefrom the substrates. The pNA light emission was quantifiedusing a microtiter plate reader at 406 nm. Values are ex-pressed as fold increase compared with negative control.
Assessment of Cholangiocyte Proliferation
In situ morphometric analysis of bile duct mass. In codedfrozen liver sections (n � 6 for each group) from the selectedgroup of animals, intrahepatic mass of large ducts was eval-uated by point count analysis (33, 48) by determining thenumber of ducts stained for CK-19, a specific marker for ratcholangiocytes (33). Immunohistochemistry for CK-19 wasperformed as described (33). After sections were stained, theywere counterstained with hematoxylin and examined in acoded fashion with a microscope (Olympus BX 40, OlympusOptical, Tokyo, Japan). The duct diameter was estimatedfrom the distance between basement membranes. The datawere expressed for each duct as the mean of two diameterstaken perpendicular to each other.
Assessment of Ductal Secretion
Bile flow. After anesthesia, BDI rats were surgically pre-pared for bile collection (8). When steady-state bile flow wasachieved, secretin (100 nM) was infused for 30 min followedby a final infusion of Krebs-Ringer Henseleit for 60 min. Bilewas collected at 10-min intervals, placed in preweighed tubes
and immediately stored at �70°C before determining bicarbon-ate concentration. Biliary bicarbonate concentration (measuredas total CO2) was determined by a Natelson microgasometerapparatus (Scientific Industries, Bohemia, NY).
Intracellular cAMP levels. Small and large cholangiocyteswere incubated for 1 h at 37°C to restore surface proteinsdamaged by treatment with proteolytic enzymes (27) andsubsequently stimulated with 0.2% BSA (basal) or secretin(100 nM) in the presence of 0.2% BSA for 5 min at 22°C (6, 10,22, 27, 33, 35, 45). Following ethanol extraction, cAMP levelswere measured by RIA using a commercially available kit(Amersham).
In Vitro Studies in Large Cholangiocytes from BDL Rats
PI3-K regulation of apoptosis, proliferation and secretion.To evaluate the intracellular mechanisms by which tauro-cholate protects against CCl4-induced damage of largecholangiocytes, we evaluated in vitro the effect of tauro-cholate on CCl4-induced apoptosis [assessed by both an-nexin-V (a specific marker of early stage apoptosis) (23) orDAPI staining (29, 32, 35)], proliferation (measured by bothH3 histone and PCNA protein expression) (32), and secretorycapacity (by measurement of cAMP levels) (4, 6, 10–12, 22,29, 32, 35, 45) of large cholangiocytes in the presence orabsence of wortmannin, a specific PI3-K inhibitor (36). Be-cause downstream effectors of PI3-K-dependent cell survivalinclude Akt [which may inactivate Bad phosphorylation andupregulate Bcl-2 by transcriptional activation (38)], we eval-uated whether taurocholate protection of CCl4-induced dam-age of large ducts (35) was associated with increased proteinexpression of phosphorylated AKT. The comparability of theprotein used was assessed by immunoblots for �-actin (11).The intensity of the bands was determined by scanning videodensitometry using the ChemiImager 4000 low-light imagingsystem (Alpha Innotech, San Leandro, CA).
Large cholangiocytes from BDL rats were treated at 37°Cwith 1) 0.2% BSA (basal) for 2 h; 2) 5 �M CCl4 for 2 h with0.2% BSA; 3) taurocholate (20 �M for 2 h) plus CCl4 (5 �M for2 h) with 0.2% BSA; 4) wortmannin (100 nM for 1 h) plustaurocholate (20 �M for 2 h) plus CCl4 (5 �M for 2 h) with0.2% BSA; or 5) wortmannin (100 nM for 1 h) with 0.2% BSA.Cholangiocyte apoptosis was determined by observation ofnuclear fragmentation by staining with both annexin-V andDAPI as described by LeSage et al. (35) and and Que et al.(39). In this set of cells, we also evaluated cholangiocyteproliferation by measurement of both H3 histone and PCNAprotein expression (32). In separate sets of experiments,large cholangiocytes were incubated for 1 h at 37°C to restoresurface proteins damaged by treatment with proteolytic en-zymes (27) and subsequently were treated at room tempera-ture with 1) 0.2% BSA (basal) for 5 min; 2) taurocholate (20�M for 10 min, positive control) with 0.2% BSA; 3) 5 �M CCl4for 10 min in the absence or presence of secretin (100 nM for5 min) with 0.2% BSA; 4) taurocholate (20 �M for 10 min)
Table 1. Serum levels of AST and ALT in 1-wk BDL rats, 1-wk BDL rats fed bile acid control diet after asingle dose of CCl4, and 1-wk BDL rats fed with TC before a single administration of CCl4
Treatment AST Levels, Sigma Frankel Units/ml ALT Levels, Sigma Frankel Units/ml
BDL 1 wk (control) 421.76�66.32 1232.00�108.20BDL 1 wk � bile acid control diet � CCl4 2 days 1514.37�270.31* 2369.56�403.25*BDL 1 wk � TC � CCl4 2 days 625.66�133.35,ns 1689.35�505.45,ns
Data are means � SE of �21 values. TC, taurocholate; AST, aspartate aminotransferase; ALT, alanine aminotransferase; ns, notsignificant. *P � 0.05 vs. bile duct ligation (BDL) control rats.
G292 BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

plus CCl4 (5 �M for 10 min) in the absence or presence ofsecretin (100 nM for 5 min) with 0.2% BSA; 5) wortmannin(100 nM for 10 min) plus taurocholate (20 �M for 10 min)plus CCl4 (5 �M for 10 min) in the absence or presence ofsecretin (100 nM for 5 min) with 0.2% BSA; or 6) wortmannin(100 nM for 10 min) in the absence or presence of CCl4 (5 �Mfor 10 min) or taurocholate (20 �M for 10 min) with 0.2%BSA. Intracellular cAMP levels were measured by RIA (6, 10,22, 27, 33, 35, 45).
Total and phosphorylated AKT protein expression wasevaluated by immunoblots in large cholangiocytes from BDLrats treated at 37°C with 1) 0.2% BSA (basal) for 2 h; 2) 5 �MCCl4 for 2 h with 0.2% BSA; 3) taurocholate (20 �M for 2 h)plus CCl4 (5 �M for 2 h) with 0.2% BSA; 4) wortmannin (100nM for 1 h) plus taurocholate (20 �M for 2 h) plus CCl4 (5 �Mfor 2 h) with 0.2% BSA; or 5) wortmannin (100 nM for 1 h)with 0.2% BSA. The comparability of the protein used wasassessed by immunoblots for �-actin (11). The intensity of thebands was determined by scanning video densitometry usingthe ChemiImager 4000 low-light imaging system (Alpha In-notech).
Statistical Analysis
All data are expressed as means � SE. The differencesbetween groups were analyzed by Student’s t-test when twogroups were analyzed or by ANOVA if more than two groupswere analyzed.
RESULTS
Studies in BDL Rats Treated in Vivo with CCl4 PlusTaurocholate or Control Feeding
Measurement of transaminase serum levels. Transam-inase serum levels of BDL rats and BDL rats treatedwith CCl4 and subsequently fed taurocholate or controldiet are shown in Table 1. Consistent with the conceptthat acute CCl4 administration induces hepatic dam-age, CCl4 administration induced a marked increase inthe serum levels of AST and ALT compared with 1 wkBDL rats (Table 1). Taurocholate feeding preventedCCl4-induced increases in AST and ALT serum levels,which were similar to those of BDL rats (Table 1).
In Situ Morphological Studies and TUNEL Analysis
Low-power view of a rat liver section from a rat withBDL for 9 days shows normal parenchyma surround-ing the terminal hepatic vein (Fig. 1A). Two days afterCCl4 treatment plus control diet feeding, we foundnecrosis restricted to zone 3 (Fig. 1B). Taurocholatefeeding protected the liver from CCl4 toxicity, whereasthe hepatocytes surrounding the central vein did notappear damaged (Fig. 1C).
Morphological analysis of liver sections from BDLrats showed a few apoptotic bodies (0.18 � 0.06%),which appeared as small acidophilic globules oftencontaining nuclear material localized between two ad-jacent cholangiocytes, engulfed within their cytoplasm,or released into the biliary lumen without an associ-ated inflammatory response (not shown). The percent-age of apoptotic bodies per 100 cholangiocytes observedin large (diameter of 15 �m) bile ducts appearshigher in liver sections from BDL, CCl4-treated rats
(0.50 � 0.06%; Fig. 2A) compared with BDL rats(0.18 � 0.06%) and BDL, taurocholate-fed, CCl4-treated rats (0.23 � 0.05%; Fig. 2B). Similarly, termi-nal deoxyneucleotidyl transferase-mediated dUTP
Fig. 1. Hematoxylin and eosin staining of a rat liver section (5-�mthick) from a rat with bile duct ligation (BDL) for 9 days (A), a rat 2days after the administration of a single dose of CCl4 � controlfeeding (B), and a BDL rat 2 days after the administration of a singledose of CCl4 � taurocholate feeding (C). Two days after acute CCl4administration (B), a large area of necrosis (*) in proximity to aterminal hepatic vein (THV) is shown. Note that hepatocytes showevidence of micro- and macrovacuolar steatosis. C: taurocholatefeeding protects liver parenchyma from CCl4 toxicity. Original mag-nification, �25.
G293BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

nick-end labeling (TUNEL) analysis showed that CCl4increased the number of cholangiocytes undergoingapoptosis compared with BDL rats (Fig. 3). Tauro-cholate feeding prevented CCl4-induced increase in
cholangiocyte apoptosis, which remained at levels sim-ilar to that of BDL rats (Fig. 3). Taurocholate feedingdid not alter apoptosis in cholangiocytes from BDL rats(not shown). In agreement with previous studies (35),
Fig. 2. Light microscopic analysis of a rat liver section(5 �m thick) obtained from BDL rat 2 days after theadministration of a single dose of CCl4 � control feeding(A) and BDL rat 2 days after the administration of asingle dose of CCl4 � taurocholate feeding (B). A: 3apoptotic bodies (arrows) appear in the epithelium of abile duct (arrowhead). B: rat liver section from BDL rat2 days after the administration of a single dose ofCCl4 � taurocholate feeding showing proliferating bileducts without evidence of apoptosis. Original magnifi-cation, �250.
Fig. 3. Evaluation of apoptosis (by terminal deoxyneucleotidyl transferase-mediated dUTP nick-end labelinganalysis) in liver sections from BDL rats and BDL rats treated with CCl4 and subsequently fed 1% taurocholate(TC) or control diet. A: several apoptotic bodies (arrows) in the epithelium lining bile ducts. After TC administra-tion (B) or in 9-day BDL rats (C), a few apoptotic bodies (arrows) were observed. Original magnification, �1,000.
G294 BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

apoptosis was never observed in small (diameter of15 �m) bile ducts. In agreement with our previousstudies (34, 35), acute CCl4 treatment did not result inmorphological evidence of necrosis in cholangiocytes.
Measurement of Caspase 8, 9, and 3 Activities
The activities of caspase 8, 9, and 3 in small andlarge cholangiocytes from BDL rats was low (Fig. 4).Consistent with the concept that CCl4 damages largebut not small ducts (34, 35), acute CCl4 administrationinduces an increase in the activities of caspase 8, 9, and3 in large but not small cholangiocytes (Fig. 4). Tauro-cholate feeding prevented CCl4-induced increases incaspase 8, 9, and 3 activities in large cholangiocytes,which remained at values similar to that of BDL rats(Fig. 4).
Measurement of Cholangiocyte Proliferation
In situ morphometry. In agreement with previousstudies (8), in BDL rats, there was a marked increasein the number of large intrahepatic bile ducts re-stricted to portal areas (volume of liver occupied by bileducts, normal control 0.53 � 0.1% vs. BDL 6.9 � 0.2%;P 0.001). Parallel with other studies (8), in rats fedcontrol diet after CCl4 treatment, we found a reductionin the liver volume occupied by large bile ducts com-pared with BDL rats (3.2 � 0.3% vs. 6.9 � 0.2%,respectively; P 0.05). Taurocholate feeding pre-vented the CCl4-induced loss of large bile ducts, be-cause liver volume occupied by large bile ducts wassimilar in taurocholate-fed, CCl4-treated rats and BDLrats (5.9 � 0.2% vs. 6.9 � 0.2%, respectively).
Measurement of Ductal Secretion
We evaluated the in vitro (i.e., cAMP) and in vivo(i.e., bile flow and bicarbonate secretion) responses tosecretin as functional assays of cholangiocyte damage/integrity of bile ducts following CCl4 administrationwith and without taurocholate feeding.
Similar to other studies (35), secretin increased intra-cellular cAMP levels of large (but not small) cholangio-cytes from BDL rats (Fig. 5). CCl4 treatment inhibitedsecretin-induced cAMP levels of large cholangiocytesfrom BDL rats fed control diet, whereas small cholan-giocytes (normally unresponsive to secretin) (6, 35) denovo responded to secretin with increases in cAMPlevels (Fig. 5). Taurocholate but not control feedingprevented CCl4-induced decreases of large cholangio-cyte secretin-stimulated cAMP levels and preventedthe de novo cAMP response of small cholangiocytes toCCl4 (Fig. 5).
The effect of control or taurocholate feeding on CCl4-induced effects on basal and secretin-stimulated bileflow and bicarbonate concentration and secretion isshown in Table 2. Basal bile flow and bicarbonateconcentration and secretion of BDL rats were similarto those of previous studies (Table 2) (8, 35, 45). Secre-tin increased bile flow and bicarbonate concentrationand secretion in BDL rats (Table 2). In rats fed controldiet and treated with CCl4, secretin did not increasebile flow, bicarbonate concentration, or secretion (Ta-ble 2). Secretin-induced bile flow and bicarbonate con-centration and secretion were similar to control BDLvalues after CCl4 administration and taurocholatefeeding (Table 2), indicating restoration of ductal se-cretion by taurocholate. The data show that after CCl4administration, there is a lack of secretin secretoryresponses, whereas taurocholate prevention of CCl4-induced duct damage is associated with intact ductfunctional integrity and secretin response.
Studies in Large Cholangiocytes Treated in Vitrowith CCl4 and Taurocholate
Quantitative evaluation of apoptosis by annexin-Vand DAPI staining. When large cholangiocytes weretreated in vitro with 5 �M CCl4, we found an increasein apoptosis (evaluated by both annexin-V and DAPIstaining) compared with large cholangiocytes treatedwith 0.2% BSA (Fig. 6, A and B). In vitro taurocholatepretreatment (20�M) reduced large cholangiocyte apop-tosis induced by CCl4 treatment (Fig. 6, A and B). Theprotective effect of taurocholate on CCl4-inducedapoptosis of large cholangiocytes was abolished by wort-mannin (100 nM), a PI3-K inhibitor (Fig. 6, A and B).Taurocholate treatment did not alter cholangiocyteapoptosis of BDL rats (not shown). Wortmannin alonedid not alter apoptosis of large cholangiocytes (Fig. 6, Aand B). Normal cholangiocytes treated with 10 �Mbeauvericin (the positive control, not shown) (39) dis-played the same morphological features of apoptosis aswe observed in cholangiocytes from CCl4-treated rats.
Proliferative capacity of large cholangiocytes. Invitro, taurocholate increased both H3 histone and
Fig. 4. Measurement of caspase 8, 9, and 3 activities in small andlarge cholangiocytes from BDL rats and BDL rats that, after CCl4administration, were fed 1% TC or control diet. Acute CCl4 admin-istration induces an increase in the activities of caspase 8, 9, and 3 inlarge but not small cholangiocytes. TC feeding prevented CCl4-induced increases in caspase 8, 9, and 3 activities in large cholan-giocytes, which remained at values similar to those of BDL rats. *P 0.05 vs. caspase activities of large cholangiocytes from BDL rats andBDL rats treated with CCl4 and subsequently fed 1% TC. Values areexpressed as fold increases compared with negative control. Data aremeans � SE of 3 experiments.
G295BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

PCNA protein expression of large cholangiocytes com-pared with large cholangiocytes stimulated with BSA[93.60 � 8.65 (taurocholate-stimulated value forPCNA) vs. 54.31 � 5.16 (basal value for PCNA) arbi-trary units and 198.88 � 28.84 (taurocholate-stimu-lated value for H3 histone) vs. 68.01 � 6.92 (basalvalue for H3 histone) arbitrary units]. Taurocholate-induced increases in H3 histone and PCNA proteinexpression were inhibited by wortmannin [50.24 �5.73 vs. 54.31 � 5.16 (basal value) arbitrary units forPCNA and 90.41 � 19.80 vs. 68.01 � 6.92 (basal value)arbitrary units for H3 histone]. In vitro, 5 �M CCl4decreased both H3 histone and PCNA protein expres-sion of large cholangiocytes from BDL rats comparedwith large cholangiocytes treated with BSA (Fig. 7).The CCl4-induced loss of H3 histone and PCNA proteinexpression of large cholangiocytes was prevented by invitro 20 �M taurocholate treatment (Fig. 7). The pro-
tective effect of taurocholate on CCl4-induced inhibi-tion of H3 histone and PCNA protein expression oflarge cholangiocytes was prevented by pretreatment oflarge cholangiocytes with wortmannin (Fig. 7), a spe-cific PI3-K inhibitor (36). Wortmannin did not alterbasal PCNA protein expression of large cholangiocytes(Fig. 7).
Intracellular cAMP levels. Secretin increased cAMPlevels of large cholangiocytes from BDL rats (Fig. 8). Inagreement with previous studies (3), taurocholate in-creased both basal (69.8 � 5.44 vs. 41.27 � 4.64 fmol/100,000 cells; P 0.05) and secretin-stimulated(141.65 � 40.54 vs. 72.53 � 6.01 fmol/100,000 cells;P 0.05) cAMP levels of large cholangiocytes fromBDL rats. CCl4 decreased both basal (19.9 � 2.39 vs.41.27 � 4.64 fmol/100,000 cells) and secretin-stimu-lated cAMP levels of large cholangiocytes (Fig. 8).When large purified cholangiocytes were pretreated in
Fig. 5. Measurement of cAMP levels in small and largecholangiocytes from BDL rats and BDL rats fed TC orcontrol diet before a single administration of CCl4.After purification, small and large cholangiocytes werestimulated with 0.2% BSA (basal) or secretin (100 nM)in the presence of 0.2% BSA. Secretin increased intra-cellular cAMP levels of large (but not small) cholangio-cytes from BDL rats. CCl4 treatment decreased secre-tin-induced cAMP levels of large cholangiocytes of BDLrats fed control diet, whereas small cholangiocytes denovo responded to secretin with increases in cAMPlevels. TC but not control feeding prevented CCl4-in-duced decreases of large cholangiocyte secretin-stimu-lated cAMP levels and blocked the de novo cAMP re-sponse of small cholangiocytes to CCl4. Data aremeans � SE of 11–15 experiments. *P 0.05 vs.corresponding basal values.
Table 2. Basal and secretin-stimulated bile flow, bicarbonate concentration, and secretion in 1-wk BDL rats,1-wk BDL rats fed bile acid control diet after a single dose of CCl4, and 1-wk BDL ratsfed with TC before a single administration of CCl4
Treatment
Bile Flow Bicarbonate Concentration Bicarbonate Secretion
Basal,�l �min�1 �kg�1
Secretin,�l �min�1 �kg�1
Basal,�Eq �min�1 �kg�1
Secretin,�Eq �min�1 �kg�1
Basal,�Eq �min�1 �kg�1
Secretin,�Eq �min�1 �kg�1
BDL 1 wk (control) 98.94�9.73 159.11�11.72* 26.64�2.34 59.72�4.65† 2.67�0.40 9.67�1.59‡BDL 1 wk � bile acid control diet �
CCl4 2 days 106.97�5.43 127.60�12.70,ns 35.21�1.10 36.53�1.51,ns 3.78�0.25 4.55�0.39,nsBDL 1 wk � TC � CCl4 2 days 117.77�10.01 159.11�11.72* 31.59�3.50 51.10�5.40† 3.84�0.84 9.19�1.45‡
Data are means � SE of 3–12 rats. *P � 0.05 vs. corresponding basal value of bile flow. †P � 0.05 vs. corresponding basal value ofbicarbonate secretion. ‡P � 0.05 vs. corresponding basal value of control rats.
G296 BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

vitro with 20 �M taurocholate, CCl4 did not abolishsecretin-stimulated cAMP levels of large cholangio-cytes (Fig. 8). The protective effect of taurocholate onCCl4-induced decrease in secretin-stimulated cAMP
levels was abolished by in vitro pretreatment of largecholangiocytes with 100 nM wortmannin (Fig. 8). Wort-mannin (100 nM) did not alter basal or secretin-stimu-lated cAMP levels of large cholangiocytes (not shown).
Fig. 6. Assessment of apoptosis by an-nexin-V (A) and DAPI staining (B) inlarge cholangiocytes from BDL ratstreated for 2 h at 37°C with 1) 0.2%BSA (basal);2) 5 �M CCl4 in the ab-sence or presence of 20 �M TC � 100nM wortmannin with 0.2% BSA; or 3)wortmannin (100 nM) with 0.2% BSA.There was a significant (P 0.05) in-crease in apoptosis in large cholangio-cytes treated with CCl4 compared withlarge cholangiocytes treated with 0.2%BSA. TC in vitro pretreatment reducedlarge cholangiocyte apoptosis inducedby CCl4. The protective effect of TC onCCl4-induced apoptosis of large cholan-giocytes was abolished by wortmannin(100 nM), a phosphatidylinositol-3-ki-nase inhibitor (PI3-K). Data are means �SE of at least 3 experiments. DAPI,4,6-diamidino-2-phenylindole.
Fig. 7. Measurement of PCNA and H3 histone proteinexpression in large cholangiocytes from BDL ratstreated for 37°C with 1) 0.2% BSA (basal) for 2 h; 2) 5�M CCl4 for 2 h with 0.2% BSA; 3) TC (20 �M for 1 h) �CCl4 (5 �M for 2 h) with 0.2% BSA; 4) wortmannin (100nM for 1 h) � TC (20 �M for 1 h) � CCl4 (5 �M for 2 h)with 0.2% BSA; or 5) wortmannin (100 nM for 1 h) with0.2% BSA. There was a significant (P 0.05) decreasein both PCNA and H3 histone protein expression oflarge cholangiocytes treated with CCl4 compared withlarge cholangiocytes treated with 0.2% BSA. TC treat-ment prevented the decrease in both PCNA and H3histone protein expression of large cholangiocyte in-duced by CCl4. The effect of TC on CCl4-induced de-crease in both PCNA and H3 histone protein expressionof large cholangiocytes was abolished by wortmannin(100 nM), a PI3-K inhibitor. Data are means � SE of atleast 3 experiments. *P 0.05 vs. H3 histone andPCNA protein expression of large cholangiocytestreated with 0.2% BSA.
G297BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org
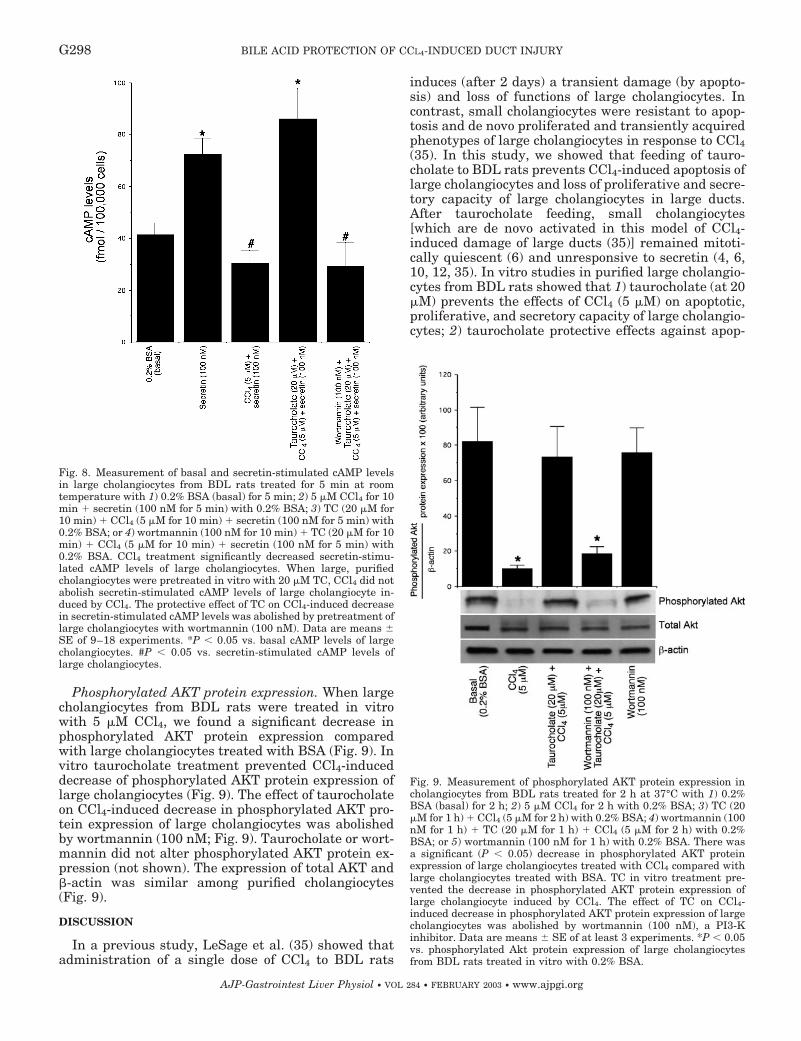
Phosphorylated AKT protein expression. When largecholangiocytes from BDL rats were treated in vitrowith 5 �M CCl4, we found a significant decrease inphosphorylated AKT protein expression comparedwith large cholangiocytes treated with BSA (Fig. 9). Invitro taurocholate treatment prevented CCl4-induceddecrease of phosphorylated AKT protein expression oflarge cholangiocytes (Fig. 9). The effect of taurocholateon CCl4-induced decrease in phosphorylated AKT pro-tein expression of large cholangiocytes was abolishedby wortmannin (100 nM; Fig. 9). Taurocholate or wort-mannin did not alter phosphorylated AKT protein ex-pression (not shown). The expression of total AKT and�-actin was similar among purified cholangiocytes(Fig. 9).
DISCUSSION
In a previous study, LeSage et al. (35) showed thatadministration of a single dose of CCl4 to BDL rats
induces (after 2 days) a transient damage (by apopto-sis) and loss of functions of large cholangiocytes. Incontrast, small cholangiocytes were resistant to apop-tosis and de novo proliferated and transiently acquiredphenotypes of large cholangiocytes in response to CCl4(35). In this study, we showed that feeding of tauro-cholate to BDL rats prevents CCl4-induced apoptosis oflarge cholangiocytes and loss of proliferative and secre-tory capacity of large cholangiocytes in large ducts.After taurocholate feeding, small cholangiocytes[which are de novo activated in this model of CCl4-induced damage of large ducts (35)] remained mitoti-cally quiescent (6) and unresponsive to secretin (4, 6,10, 12, 35). In vitro studies in purified large cholangio-cytes from BDL rats showed that 1) taurocholate (at 20�M) prevents the effects of CCl4 (5 �M) on apoptotic,proliferative, and secretory capacity of large cholangio-cytes; 2) taurocholate protective effects against apop-
Fig. 9. Measurement of phosphorylated AKT protein expression incholangiocytes from BDL rats treated for 2 h at 37°C with 1) 0.2%BSA (basal) for 2 h; 2) 5 �M CCl4 for 2 h with 0.2% BSA; 3) TC (20�M for 1 h) � CCl4 (5 �M for 2 h) with 0.2% BSA; 4) wortmannin (100nM for 1 h) � TC (20 �M for 1 h) � CCl4 (5 �M for 2 h) with 0.2%BSA; or 5) wortmannin (100 nM for 1 h) with 0.2% BSA. There wasa significant (P 0.05) decrease in phosphorylated AKT proteinexpression of large cholangiocytes treated with CCl4 compared withlarge cholangiocytes treated with BSA. TC in vitro treatment pre-vented the decrease in phosphorylated AKT protein expression oflarge cholangiocyte induced by CCl4. The effect of TC on CCl4-induced decrease in phosphorylated AKT protein expression of largecholangiocytes was abolished by wortmannin (100 nM), a PI3-Kinhibitor. Data are means � SE of at least 3 experiments. *P 0.05vs. phosphorylated Akt protein expression of large cholangiocytesfrom BDL rats treated in vitro with 0.2% BSA.
Fig. 8. Measurement of basal and secretin-stimulated cAMP levelsin large cholangiocytes from BDL rats treated for 5 min at roomtemperature with 1) 0.2% BSA (basal) for 5 min; 2) 5 �M CCl4 for 10min � secretin (100 nM for 5 min) with 0.2% BSA; 3) TC (20 �M for10 min) � CCl4 (5 �M for 10 min) � secretin (100 nM for 5 min) with0.2% BSA; or 4) wortmannin (100 nM for 10 min) � TC (20 �M for 10min) � CCl4 (5 �M for 10 min) � secretin (100 nM for 5 min) with0.2% BSA. CCl4 treatment significantly decreased secretin-stimu-lated cAMP levels of large cholangiocytes. When large, purifiedcholangiocytes were pretreated in vitro with 20 �M TC, CCl4 did notabolish secretin-stimulated cAMP levels of large cholangiocyte in-duced by CCl4. The protective effect of TC on CCl4-induced decreasein secretin-stimulated cAMP levels was abolished by pretreatment oflarge cholangiocytes with wortmannin (100 nM). Data are means �SE of 9–18 experiments. *P 0.05 vs. basal cAMP levels of largecholangiocytes. #P 0.05 vs. secretin-stimulated cAMP levels oflarge cholangiocytes.
G298 BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

tosis and loss of large cholangiocyte functions wereabolished by the PI3-K inhibitor wortmannin; and 3)taurocholate prevented CCl4-induced decrease incholangiocyte phosphorylated AKT protein expression.
Bile acids are internalized into cholangiocytes by theNa�-dependent ABAT (5, 28), thus modulating secre-tory and proliferative activities of cholangiocytes (3, 5,7). Alpini et al. (5) previously demonstrated that ABATand bile acid transport activity are expressed by largebut not small cholangiocytes of normal rats. Tauro-cholate, for example, increases proliferative and secre-tory activities of normal, large cholangiocytes (3). In-hibition of ABAT-mediated bile acid uptake ablates thetaurocholate effects on cholangiocyte proliferation andsecretion (3), so it is likely that bile acid internalizationby ABAT is required for bile acid signaling in cholan-giocytes. Thus the taurocholate cytoprotective effectsin CCl4-induced cholangiocyte injury may requireABAT-mediated bile acid uptake, and this issue will beaddressed in future studies by examining ABAT ex-pression and the effects of ABAT inhibitors.
Studies showed that bile acids affect PI3-K activityin hepatocytes (36, 42, 46); however, no informationexists regarding the role of PI3-K on the regulation ofcholangiocyte functions. Taurochenodeoxycholate, forexample, activates PI3-K-dependent survival path-ways in rat hepatocytes, which prevent its inherenttoxicity (42). In addition, cAMP-mediated cytoprotec-tion against bile acid-induced apoptosis in cultured rathepatocytes is mediated by PI3-K (46). PI3-K (which isactivated by taurocholate) (49) represents a potentactivator of survival signals, raising the possibilitythat nontoxic, hydrophobic bile acids do not triggerapoptosis in hepatocytes because they activate a PI3-K-dependent survival signaling pathway (42). Re-cently, Webster and Anwer (47) showed that hepato-cyte growth factor (HGF) inhibits bile acid-inducedhepatocyte apoptosis in primary cultures of rat hepa-tocytes. This antiapoptotic effect was completely re-versed by PI3-K inhibitors, indicating that HGF-in-duced survival from bile acid-induced apoptosis inhepatocytes is mediated via the PI3-K signaling path-way. Consistent with these previous studies, we foundthat taurocholate feeding may reduce CCl4-inducedhepatocyte injury, because markers of hepatocyte in-jury (serum ALT and AST levels), which were signifi-cantly elevated in CCl4-treated BDL rats, were similarin taurocholate-fed CCl4-treated and BDL control rats.The present data demonstrate that in cholangiocytes,taurocholates exert a protective effect against CCl4-induced apoptosis of large cholangiocytes through aPI3-K mechanism. Downstream effectors of PI3-K-de-pendent survival include PKC isoforms and Akt, whichseems to inactivate Bad phosphorylation and upregu-late Bcl-2 by transcriptional activation (38). In supportof this, we have shown that in vitro CCl4-inducedapoptosis of large cholangiocytes was associated withdecreased phosphorylated Akt protein expression andthat taurocholate protection against CCl4-induceddamage of large cholangiocytes is associated withmaintained phosphorylated Akt protein expression.
A number of studies has shown that the secretinreceptor and cAMP levels play an important role in theregulation of cholangiocyte apoptosis/proliferation (3,6, 7, 9, 22, 29, 30, 32–35). This concept is supported byseveral studies showing that there is upregulation ofsecretin receptor and basal and secretin-stimulatedcAMP levels in all of the hyperplastic models of cholan-giocyte proliferation so far studied (3, 6, 7, 9, 22, 29, 30,32–35), whereas reduced cholangiocyte replication[e.g., following total vagotomy (29), ursodeoxycholate,or tauroursodeoxycholate feeding (2) or gastrin treat-ment (22), or acute gavage CCl4 administration (34,35)] is associated with depressed secretin receptor andsecretin-stimulated cAMP levels. Furthermore, up-regulation of cAMP levels (by chronic forskolin treat-ment) increases cholangiocyte proliferation and thenumber of bile ducts of normal rats through a PKA-dependent mechanism (31). In BDL, vagotomized rats(29), chronic forskolin treatment prevented the de-crease in cAMP levels, maintained cholangiocyte pro-liferation and secretion, and decreased cholangiocyteapoptosis induced by vagotomy in BDL rats (29). Fi-nally, recent studies (40) have shown that changes inthe activity of Gs and Gi protein subunits are associ-ated with alteration of cholangiocytes of BDL rats. Inprimary hepatocyte cultures, elevated cAMP intracel-lular levels not only protected against Fas-inducedapoptosis, but also rescued the cells from functionaldeterioration (41). Furthermore, forskolin and cAMPanalogs have been shown to prevent apoptosis inducedby hydrophobic bile acids in cultured rat hepatocytes(46). In neuronal tissues, the intracellular cAMP sys-tem is fundamental for tissue differentiation and sur-vival, for promotion of regeneration of injured tissue,and for prevention of apoptosis (14, 16, 18). Severalgrowth factors (e.g., nerve growth factor and pituitaryadenylate cyclase-activating protein) that promoteneural tissue differentiation and prevent apoptosis actvia cAMP pathways through yet unidentified signalingfactors (e.g., mitogen-activated protein kinases andcAMP-dependent transcription factors) (44). In supportof the fact that cAMP system plays an important role inthe regulation of cell proliferation/apoptosis, our datashow that taurocholate, by maintaining basal and se-cretin-stimulated cAMP levels of cholangiocytes fromBDL rats, prevents CCl4-induced cholangiocyte apop-tosis (35) and the inhibition of cholangiocyte prolifera-tion and secretion (35).
Cytoprotection provided by activation of cAMP andAKT appear to occur through cooperative effects ofthese two pathways. cAMP does not lead to directactivation of PI3-K (46). The dual requirement forPI3-K and cAMP for cytoprotection against apoptosismay therefore reflect the need for cooperative action ona single downstream effector. Because PI3-K activatesprotein translocation, the movement of some effectormolecule to a subcellular location where cAMP exertsits effect, may be required. The lipid products of PI3-Khydrolysis are necessary to promote membrane inter-action of PKB, where it can subsequently activatecAMP’s cytoprotective actions (46). Studies (21) have
G299BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

also shown that PI3-K is involved in taurocholate reg-ulation of bile secretion. PI3-K is involved in the reg-ulation of vesicle trafficking and cytoskeleton organi-zation in the process of bile formation (20). Theincrease in canalicular bile acid secretion after tauro-cholate administration in vivo results from rapid trans-porter activation or recruitment to the canalicular do-main (21), and PI3-K is required for intracellulartrafficking of ATP-dependent transporters to the can-alicular membrane (36).
Defining the signaling pathways involved in cholan-giocyte apoptosis and cytoprotection in specificallysized ducts has important pathophysiological implica-tions, because several human cholestatic liver diseases(e.g., PBC) are characterized by spotty rather thandiffuse proliferation/loss of certain sized ducts (i.e.,small interlobular bile ducts) (9). The present studiesallow a better understanding of the mechanisms bywhich different-sized ducts are damaged and how cer-tain bile acids protect against toxin-induced ductope-nia in cholestatic liver diseases (e.g., PBC and PSC) (9).Failure of bile acids to activate cytoprotective mecha-nisms in cholangiocytes may promote apoptosis andloss of bile ducts in these diseases. Because ursodeoxy-cholic acid is a major therapeutically used bile acidwith antiapoptotic effects (37), a direct outgrowth ofthe studies is to evaluate the potential protective ef-fects of this bile in CCl4 (34, 35)- or vagotomy (29)-induced duct damage.
This work was supported by a grant from MURST (MM06215421/2)progetto nazionale 2000 to Dr. D. Alvaro, by a grant award, MURSTMM06215421, to the Dept. of Gastroenterology, Ancona, by a grantaward to Drs. G. LeSage and G. Alpini from Scott & White Hospital andThe Texas A&M Univ. System, by an National Institutes of Health(NIH) Grant DK-54208 to Dr. G. LeSage and by a veteran’s affairs meritaward, and NIH Grant DK-58411 to Dr. G. Alpini.
REFERENCES
1. Aagaard-Tillery KM and Jelinek DF. Phosphatidylinositol3-kinase activation in normal human B lymphocytes. J Immunol156: 4543–4554, 1996.
2. Alpini G, Baiocchi L, Glaser S, Ueno Y, Marzioni M, Fran-cis H, Phinizy JL, Angelico M, and LeSage G. Ursodeoxy-cholate and tauroursodeoxycholate inhibit cholangiocyte growthand secretion of BDL rats through activation of PKC alpha.Hepatology 35: 1041–1052, 2002.
3. Alpini G, Glaser S, Robertson W, Phinizy JL, Rodgers RE,Caligiuri A, and LeSage G. Bile acids stimulate proliferativeand secretory events in large but not small cholangiocytes. Am JPhysiol Gastrointest Liver Physiol 273: G518–G529, 1997.
4. Alpini G, Glaser S, Robertson W, Rodgers RE, Phinizy JL,Lasater J, and LeSage GD. Large but not small intrahepaticbile ducts are involved in secretin-regulated ductal bile secre-tion. Am J Physiol Gastrointest Liver Physiol 272: G1064–G1074, 1997.
5. Alpini G, Glaser SS, Rodgers R, Phinizy JL, RobertsonWE, Lasater J, Caligiuri A, Tretjak Z, and LeSage GD.Functional expression of the apical Na�-dependent bile acidtransporter in large but not small rat cholangiocytes. Gastroen-terology 113: 1734–1740, 1997.
6. Alpini G, Glaser SS, Ueno Y, Pham L, Podila PV, CaligiuriA, LeSage G, and LaRusso NF. Heterogeneity of the prolifer-ative capacity of rat cholangiocytes after bile duct ligation. Am JPhysiol Gastrointest Liver Physiol 274: G767–G775, 1998.
7. Alpini G, Glaser SS, Ueno Y, Rodgers R, Phinizy JL, Fran-cis H, Baiocchi L, Holcomb LA, Caligiuri A, and LeSage
GD. Bile acid feeding induces cholangiocyte proliferation andsecretion: evidence for bile acid-regulated ductal secretion. Gas-troenterology 116: 179–186, 1999.
8. Alpini G, Lenzi R, Sarkozi L, and Tavoloni N. Biliary phys-iology in rats with bile ductular cell hyperplasia. Evidence for asecretory function of proliferated bile ductules. J Clin Invest 81:569–578, 1988.
9. Alpini G, Prall RT, and LaRusso NF. The pathobiology ofbiliary epithelia. In: The Liver: Biology & Pathobiology (4th ed.),edited by Arias IM, Boyer JL, Chisari FV, Fausto N, Jakoby W,Schachter D, and Shafritz DA. Philadelphia, PA: Lippincott,Williams & Wilkins, 2001, p. 421–435.
10. Alpini G, Roberts S, Kuntz SM, Ueno Y, Gubba S, PodilaPV, LeSage G, and LaRusso NF. Morphological, molecular,and functional heterogeneity of cholangiocytes from normal ratliver. Gastroenterology 110: 1636–1643, 1996.
11. Alpini G, Ueno Y, Glaser S, Phinizy JL, Francis H, andLeSage G. Bile acid feeding stimulates proliferative activity ofboth small and large cholangiocytes through activation andmembrane translocation of protein kinase C alpha. Hepatology34: 868–876, 2001.
12. Alpini G, Ulrich C, Roberts S, Phillips JO, Ueno Y, PodilaPV, Colegio O, LeSage GD, Miller LJ, and LaRusso NF.Molecular and functional heterogeneity of cholangiocytes fromrat liver after bile duct ligation. Am J Physiol Gastrointest LiverPhysiol 272: G289–G297, 1997.
13. Alpini G, Ulrich C II, Phillips J, Pham L, Miller L, andLaRusso N. Upregulation of secretin receptor gene expressionin rat cholangiocytes after bile duct ligation. Am J PhysiolGastrointest Liver Physiol 266: G922–G928, 1994.
14. Ambron RT and Walters ET. Priming events and retrogradeinjury signals. A new perspective on the cellular and molecularbiology of nerve regeneration. Mol Neurobiol 13: 61–79, 1996.
15. Clawson GA. Mechanisms of carbon tetrachloride hepatotoxic-ity. Path Immunopathol Res 8: 104–112, 1989.
16. Crowder RJ and Freeman RS. Phosphatidylinositol 3-kinaseand Akt protein kinase are necessary and sufficient for thesurvival of nerve growth factor-dependent sympathetic neurons.J Neurosci: 2933–2943, 1998.
17. Davy DA, Campbell HD, Fountain S, de Jong D, andCrouch MF. The flightless I protein colocalizes with actin- andmicrotubule-based structures in motile Swiss 3T3 fibroblasts:evidence for the involvement of PI 3-kinase and Ras-relatedsmall GTPases. J Cell Sci 114: 549–562, 2001.
18. Eggenberger M, McKinney RA, Fischer JA, and Muff R.Differential expression of calcitonin and parathyroid hormone/parathyroid hormone-related protein receptors in P19 embryoniccarcinoma cells treated with retinoic acid. Endocrinology 139:1023–1030, 1998.
19. Fan S, Ma YX, Wang JA, Yuan RQ, Meng Q, Cao Y, LaterraJJ, Goldberg ID, and Rosen EM. The cytokine hepatocytegrowth factor/scatter factor inhibits apoptosis and enhancesDNA repair by a common mechanism involving signalingthrough phosphatidyl inositol 3� kinase. Oncogene 19: 2212–2223, 2000.
20. Folli F, Alvaro D, Gigliozzi A, Bassotti C, Kahn CR, Pon-tiroli AE, Capocaccia L, Jezequel AM, and Benedetti A.Regulation of endocytic-transcytotic pathways and bile secretionby phosphatidylinositol 3-kinase in rats. Gastroenterology 113:954–965, 1997.
21. Gatmaitan ZC, Nies AT, and Arias IM. Regulation and trans-location of ATP-dependent apical membrane proteins in ratliver. Am J Physiol Gastrointest Liver Physiol 272: G1041–G1049, 1997.
22. Glaser S, Benedetti A, Marucci L, Alvaro D, Baiocchi L,Kanno N, Caligiuri A, Phinizy JL, Chowdhury U, Papa E,LeSage G, and Alpini G. Gastrin inhibits cholangiocyte growthin bile duct-ligated rats by interaction with cholecystokinin-B/gastrin receptors via D-myo-inositol 1,4,5-triphosphate-, Ca(2�)-,and protein kinase C alpha-dependent mechanisms. Hepatology32: 17–25, 2000.
23. Heidenreich S, Schmidt M, August C, Cullen P, Rade-maekers A, and Pauels HG. Regulation of human monocyte
G300 BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org

apoptosis by the CD14 molecule. J Immunol 159: 3178–3188,1997.
24. Ibuki Y and Goto R. Suppression of apoptosis by UVB irradi-ation: survival signaling via PI3-kinase/Akt pathway. BiochemBiophys Res Commun 279: 872–878, 2000.
25. Ishii M, Vroman B, and LaRusso NF. Isolation and morpho-logical characterization of bile duct epithelial cells from normalrat liver. Gastroenterology 97: 1236–1247, 1989.
26. Kanno N, LeSage G, Glaser S, Alvaro D, and Alpini G.Functional heterogeneity of the intrahepatic biliary epithelium.Hepatology 31: 555–561, 2000.
27. Kato A, Gores GJ, and LaRusso NF. Secretin stimulatesexocytosis in isolated bile duct epithelial cells by a cyclic AMP-mediated mechanism. J Biol Chem 267: 15523–15529, 1992.
28. Lazaridis KN, Pham L, Tietz P, Marinelli RA, deGroen PC,Levine S, Dawson PA, and LaRusso NF. Rat cholangiocytesabsorb bile acids at their apical domain via the ileal sodium-dependent bile acid transporter. J Clin Invest 100: 2714–2721,1997.
29. LeSage EG, Alvaro D, Benedetti A, Glaser S, Marucci L,Baiocchi L, Eisel W, Caligiuri A, Phinizy JL, Rodgers R,Francis H, and Alpini G. Cholinergic system modulatesgrowth, apoptosis, and secretion of cholangiocytes from bileduct-ligated rats. Gastroenterology 117: 191–199, 1999.
30. LeSage G, Glaser S, and Alpini G. Regulation of cholangiocyteproliferation. Liver 21: 73–80, 2001.
31. LeSage G, Glaser S, Alvaro D, Kanno N, Phinizy JL,Chowdhury U, and Alpini G. cAMP stimulates secretory andproliferative capacity of the rat intrahepatic biliary epitheliumthrough the PKA system (Abstract). Gastroenterology 118: A148,2000.
32. LeSage G, Glaser S, Ueno Y, Alvaro D, Baiocchi L, KannoN, Phinizy JL, Francis H, and Alpini G. Regression of cholan-giocyte proliferation after cessation of ANIT feeding is associatedwith increased apoptosis. Am J Physiol Gastrointest LiverPhysiol 281: G182–G190, 2001.
33. LeSage G, Glaser SS, Gubba S, Robertson WE, Phinizy JL,Lasater J, Rodgers RE, and Alpini G. Regrowth of the ratbiliary tree after 70% partial hepatectomy is coupled to in-creased secretin-induced ductal secretion. Gastroenterology 111:1633–1644, 1996.
34. LeSage GD, Benedetti A, Glaser S, Marucci L, Tretjak Z,Caligiuri A, Rodgers R, Phinizy JL, Baiocchi L, Francis H,Lasater J, Ugili L, and Alpini G. Acute carbon tetrachloridefeeding selectively damages large, but not small, cholangiocytesfrom normal rat liver. Hepatology 29: 307–319, 1999.
35. LeSage GD, Glaser SS, Marucci L, Benedetti A, PhinizyJL, Rodgers R, Caligiuri A, Papa E, Tretjak Z, JezequelAM, Holcomb LA, and Alpini G. Acute carbon tetrachloridefeeding induces damage of large but not small cholangiocytesfrom BDL rat liver. Am J Physiol Gastrointest Liver Physiol 276:G1289–G1301, 1999.
36. Misra S, Ujhazy P, Gatmaitan Z, Varticovski L, and AriasIM. The role of phosphoinositide 3-kinase in taurocholate-in-duced trafficking of ATP-dependent canalicular transporters inrat liver. J Biol Chem 273: 26638–26644, 1998.
37. Poupon RE, Poupon R, and Balkau B. Ursodiol for thelong-term treatment of primary biliary cirrhosis. The UDCA-PBC Study Group. N Engl J Med 330: 1342–1347, 1994.
38. Pugazhenthi S, Nesterova A, Sable C, Heidenreich KA,Boxer LM, Heasley LE, and Reusch JE. Akt/protein kinase Bup-regulates Bcl-2 expression through cAMP-response element-binding protein. J Biol Chem 275: 10761–10766, 2000.
39. Que FG, Gores GJ, and LaRusso NF. Development and initialapplication of an in vitro model of apoptosis in rodent cholangio-cytes. Am J Physiol Gastrointest Liver Physiol 272: G106–G115,1997.
40. Rodriguez-Henche N, Guijarro LG, Couvineau A, CarreroI, Arilla E, Laburthe M, and Prieto JC. G proteins in rat liverproliferation during cholestasis. Hepatology 20: 1041–1047,1994.
41. Rouquet N, Carlier K, Briand P, Wiels J, and Joulin V.Multiple pathways of Fas-induced apoptosis in primary cultureof hepatocytes. Biochem Biophys Res Commun 229: 27–35, 1996.
42. Rust C, Karnitz LM, Paya CV, Moscat J, Simari RD, andGores GJ. The bile acid taurochenodeoxycholate activates aphosphatidylinositol 3-kinase-dependent survival signaling cas-cade. J Biol Chem 275: 20210–20216, 2000.
43. Rutemburg AM, Kim H, Fishbein JW, Hanker JS, Was-serkrug HL, and Seligman AM. Histochemical and ultrastruc-tural demonstration of �-glutamyl transpeptidase activity.J Histochem Cytochem 17: 517–526, 1969.
44. Tanaka J, Koshimura K, Murakami Y, Sohmiya M,Yanaihara N, and Kato Y. Neuronal protection from apoptosisby pituitary adenylate cyclase-activating polypeptide. RegulPept 72: 1–8, 1997.
45. Tietz P, Alpini G, Pham LD, and LaRusso NF. Somatostatininhibits secretin-induced ductal choleresis in vivo and exocytosisby cholangiocytes. Am J Physiol Gastrointest Liver Physiol 269:G110–G118, 1995.
46. Webster CR and Anwer MS. Cyclic adenosine monophos-phate-mediated protection against bile acid-induced apoptosis incultured rat hepatocytes. Hepatology 27: 1324–1331, 1998.
47. Webster CR and Anwer MS. Phosphoinositide 3-kinase, butnot mitogen-activated protein kinase, pathway is involved inhepatocyte growth factor-mediated protection against bile acid-induced apoptosis in cultured rat hepatocytes. Hepatology 33:608–615, 2001.
48. Weibel ER and Gomez D. A principle for counting tissuestructure on random sections. Appl Physiol 17: 3443–3448, 1962.
49. Yao R and Cooper GM. Requirement for phosphatidylinosi-tol-3 kinase in the prevention of apoptosis by nerve growthfactor. Science 267: 2003–2006, 1995.
G301BILE ACID PROTECTION OF CCL4-INDUCED DUCT INJURY
AJP-Gastrointest Liver Physiol • VOL 284 • FEBRUARY 2003 • www.ajpgi.org
Copyright © 2022 FDOKUMEN



![Synthesis, structure and properties of heterotrinuclear carboxylate complexes [Fe 2M(Ca, Sr, Ba)O(CCl 3COO) 6(THF) n ]](https://static.fdokumen.com/doc/165x107/6333b73328cb31ef600d6251/synthesis-structure-and-properties-of-heterotrinuclear-carboxylate-complexes-fe.jpg)
















![Synthesis and biological evaluation of pyrido[3′,2′:4,5]furo[3,2-d]pyrimidine derivatives as novel PI3 kinase p110α inhibitors](https://static.fdokumen.com/doc/165x107/63259095584e51a9ab0ba457/synthesis-and-biological-evaluation-of-pyrido3245furo32-dpyrimidine.jpg)