
C-isotope composition of CO2 respired by shoots and roots: fractionation during dark respiration?

b i om a s s a n d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 9
Avai lab le a t www.sc iencedi rec t .com
ht tp : / /www.e lsev ier . com/ loca te /b iombioe
Effect of antioxidants and associate changes in antioxidantenzymes in controlling browning and necrosis ofproliferating shoots of elite Jatropha curcas L
Pratibha Misra*, Dibya D. Toppo, Neha Gupta, Debasis Chakrabarty, Rakesh Tuli**
National Botanical Research Institute, Council of Scientific and Industrial Research (CSIR), Rana Pratap Marg, Lucknow 226001, India
a r t i c l e i n f o
Article history:
Received 30 November 2009
Received in revised form
15 July 2010
Accepted 22 July 2010
Available online 21 August 2010
Keywords:
Antioxidant enzyme
Isoenzymes
Jatropha curcas
Long-term culture
Shoot necrosis
Abbreviations: AdS, adenine sulphate; APXCys, cysteine; EDTA, ethylene diaminetetraaform; GSSG, glutathione, oxidized form; H2ONADPH, b-nicotinamide adenine dinucleotidvinylpyrrolidone; PMSF, phenazine methosumutase; Toco, a-tocopherol; TCA, trichloroa* Corresponding author. Tel.: þ91 522 229792** Corresponding author. Present address:Punjab, India.
E-mail addresses: [email protected]/$ e see front matter ª 2010 Elsevdoi:10.1016/j.biombioe.2010.07.027
a b s t r a c t
A high yielding elite plant of Jatropha curcas was established under aseptic condition from
field-grown nodal explants. Shoots were proliferated in MS medium supplemented with
0.5 mg dm�3 benzyladenine and 0.1 mg dm�3 indolebutyric acid along with 10 mg dm�3
adenine sulphate and a combination of 15 mg dm�3 each of L-glutamine and L-arginine.
However, within 15e20 d of culture incubation, tissue browning/necrosis leading to poor
plant regeneration in vitro was observed. A set of different antioxidants, namely, reduced
glutathione, ascorbic acid, tocopherol and cysteine were used in the medium individually
and in combination to solve the problem of tissue browning and necrosis. The addition of
antioxidants proved beneficial for the growth of the shoots. The optimum medium
comprised of 25 mg dm�3 reduced glutathione and 10 mg dm�3 ascorbic acid, where
proliferating shoots having highest leaf canopy area, remained fresh, green and regener-
ative up to 40 d of culture incubation without any subculture. The activities of antioxidant
enzymes, such as superoxide dismutase was higher in control shoots, indicating that
tissue browning/necrosis was associated with oxidative stress which was further sup-
ported by higher contents of H2O2 and phenolics in control shoots compared to the other
treatments. Similarly glutathione reductase, ascorbate peroxidase and guiacol peroxidase
was higher in treated shoots than control indicating that these shoots have developed
antioxidant enzymatic protective system which determine the ability to survive in oxida-
tive stress and up regulation of these enzymes would help to reduce the built up of reactive
oxygen species.
ª 2010 Elsevier Ltd. All rights reserved.
, ascorbate peroxidase; AA, ascorbic acid; BA, 6-benzyladenine; BSA, bovine serum albumin;cetic acid; GPX, guiacol peroxidase; GR, glutathione reductase; GSH, glutathione, reduced
2, hydrogen peroxide; IBA, indole-3-butyric acid; MS, Murashige and Skoog’s (1962) medium;e phosphate, reduced form; NBT, nitroblue tetrazolium; OH, hydroxyl radicals; PVP, poly-lfate; ROS, reactive oxygen species; SDS, sodium dodecyl sulphate; SOD, superoxide dis-cetic acid.3; fax: þ91 522 2205836/2205839.National Agrifood Biotechnology Institute (Department of Biotechnology), Mohali-160071,
s.in, [email protected] (P. Misra).ier Ltd. All rights reserved.
b i om a s s an d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 91862
1. Introduction
Biofuel is a locally-available source of energy that not only can
provide energy to meet the increased energy demand derived
from the economic development of developing countries, but
also contributes to climate change mitigation and rural deve-
lopment. Jatropha curcas (Fam. Euphorbiaceae) plant has been
acknowledged as the preferred crop for the purpose which is
drought resistant, perennial and fast growing on poor soil [1]. It
produces oil from the seeds, which can be combusted as fuel
without being refined [2]. It burns with clear smoke-free flame,
tested successfully as fuel for simple diesel engine [3]. Although
there are other sources available as biofuel crops, but being
a non food crop Jatropha cultivation as biodiesel crop ranked
first among all, considering social, economic and humanitarian
reasons [4]. Jatrophahas a lot of interspecific genetic diversity [5]
as well as diversity among different accessions of Indian vari-
eties [6]. The inter-variety spread of oil yield between Jatropha
varieties ranges between 30 and 37%. Therefore, use of elite
variety for planting material can assure a 20% more biodiesel
yield. The present study overcomes hitherto problem of
browning of in vitro cultures of Jatropha [7] and achieves a robust
protocol for large scale propagation of tissue raised plants of its
elite variety. There are few reports on in vitro shoot regenera-
tion from different seedling explants [8,9], nodal explants [10]
and from leaf explants [11]. In most of the reports in vitro
clonal propagation was achieved through organogenesis
[1,10,12], but few are through somatic embryogenesis [13,14].
Although shoot multiplication and their growth is not
a problem in J. curcas, but the regenerating shoots turned
brown/necrosed within 15e20 d of culture incubation. Due to
browning of tissue in Jatropha, it becomes difficult to continue
long-term cultures. Tissue browning generally inhibits the
growth and adventitious shoot formation [15]. The problem of
browning in callus cultures of Jatropha has recently been pub-
lished [7]. Reactive oxygen species such as superoxide radical,
H2O2 and OH radicals have a role in lipid peroxidation,
membrane damage and consequently in leaf senescence. It has
been shown that during leaf senescence, proteins, phospho-
lipids and pigments may be degraded by free radicals as free
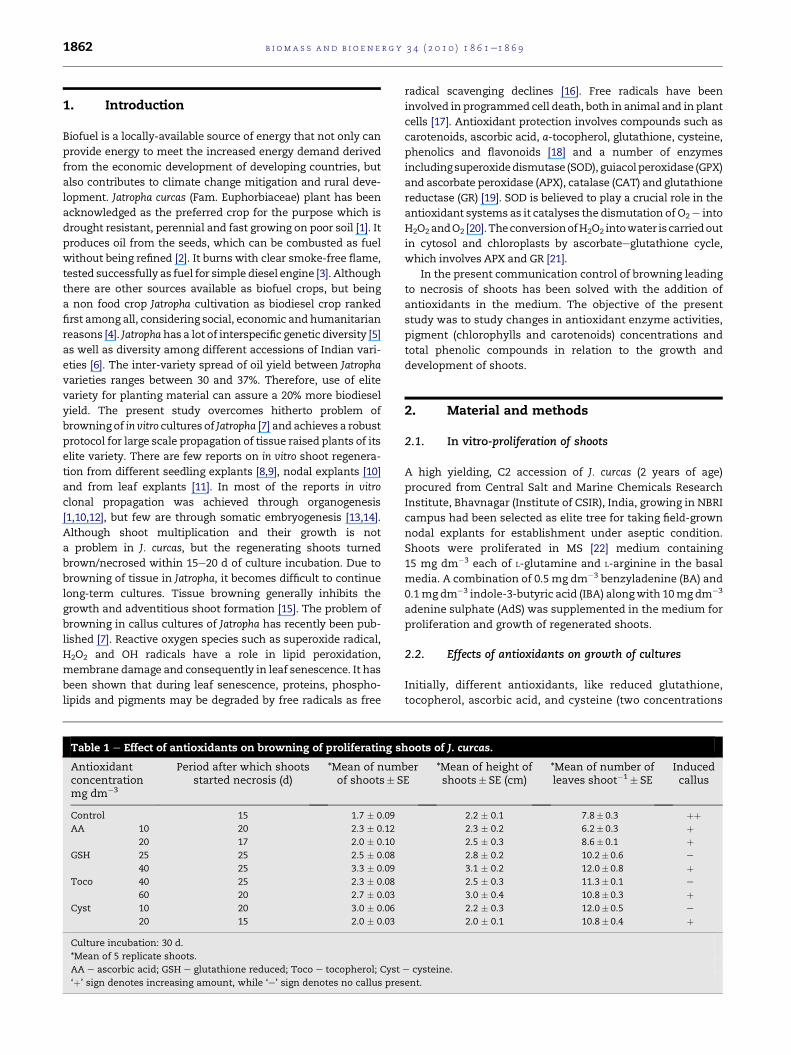
Table 1 e Effect of antioxidants on browning of proliferating s
Antioxidantconcentrationmg dm�3
Period after which shootsstarted necrosis (d)
*Mean of numof shoots� S
Control 15 1.7 � 0.09
AA 10 20 2.3 � 0.12
20 17 2.0 � 0.10
GSH 25 25 2.5 � 0.08
40 25 3.3 � 0.09
Toco 40 25 2.3 � 0.08
60 20 2.7 � 0.03
Cyst 10 20 3.0 � 0.06
20 15 2.0 � 0.03
Culture incubation: 30 d.
*Mean of 5 replicate shoots.
AA e ascorbic acid; GSH e glutathione reduced; Toco e tocopherol; Cyst
‘þ’ sign denotes increasing amount, while ‘e’ sign denotes no callus pres
radical scavenging declines [16]. Free radicals have been
involved in programmed cell death, both in animal and in plant
cells [17]. Antioxidant protection involves compounds such as
carotenoids, ascorbic acid, a-tocopherol, glutathione, cysteine,
phenolics and flavonoids [18] and a number of enzymes
includingsuperoxidedismutase (SOD), guiacolperoxidase (GPX)
and ascorbate peroxidase (APX), catalase (CAT) and glutathione
reductase (GR) [19]. SOD is believed to play a crucial role in the
antioxidant systems as it catalyses the dismutation of O2e into
H2O2andO2 [20]. TheconversionofH2O2 intowater iscarriedout
in cytosol and chloroplasts by ascorbateeglutathione cycle,
which involves APX and GR [21].
In the present communication control of browning leading
to necrosis of shoots has been solved with the addition of
antioxidants in the medium. The objective of the present
study was to study changes in antioxidant enzyme activities,
pigment (chlorophylls and carotenoids) concentrations and
total phenolic compounds in relation to the growth and
development of shoots.
2. Material and methods
2.1. In vitro-proliferation of shoots
A high yielding, C2 accession of J. curcas (2 years of age)
procured from Central Salt and Marine Chemicals Research
Institute, Bhavnagar (Institute of CSIR), India, growing in NBRI
campus had been selected as elite tree for taking field-grown
nodal explants for establishment under aseptic condition.
Shoots were proliferated in MS [22] medium containing
15 mg dm�3 each of L-glutamine and L-arginine in the basal
media. A combination of 0.5 mg dm�3 benzyladenine (BA) and
0.1mg dm�3 indole-3-butyric acid (IBA) alongwith 10mg dm�3
adenine sulphate (AdS) was supplemented in the medium for
proliferation and growth of regenerated shoots.
2.2. Effects of antioxidants on growth of cultures
Initially, different antioxidants, like reduced glutathione,
tocopherol, ascorbic acid, and cysteine (two concentrations
hoots of J. curcas.
berE
*Mean of height ofshoots� SE (cm)
*Mean of number ofleaves shoot�1� SE
Inducedcallus
2.2 � 0.1 7.8� 0.3 þþ2.3 � 0.2 6.2� 0.3 þ2.5 � 0.3 8.6� 0.1 þ2.8 � 0.2 10.2� 0.6 e
3.1 � 0.2 12.0� 0.8 þ2.5 � 0.3 11.3� 0.1 e
3.0 � 0.4 10.8� 0.3 þ2.2 � 0.3 12.0� 0.5 e
2.0 � 0.1 10.8� 0.4 þ
e cysteine.
ent.

Table 2 e Responses of in vitro-growing shoots of J. curcas after 40 d of incubation.
Treatments FW� SE(g shoot�1)
DW � SE(mg shoot�1)
*Mean ofnumber ofshoots � SE
*Mean of heightof shoots(cm) � SE
*Mean of numberof leaves
shoot�1 � SE
Canopy leaf area(cm2) � SE
Concentrationmg dm�3
Incubationperiod (d)
Control 20 0.52 � 0. 35.47 � 7.0 1.67 � 0.06 2.24 � 0.1 7.3 � 0.28 6.78 � 0.5
40 1.04 � 0. 51.33 � 8. 1.27 � 0.08 2.54 � 0. 6.2 � 0.34 4.13 � 0.4
40 GSHþ 10 AA 20 1.28 � 0.6 87.2 � 12.5 2.68 � 0.33 2.83 � 1.6 8.2 � 0.54 11.05 � 6.4
40 2.55 � 0.5 153.1 � 25.6 2.63 � 0.06 3.36 � 0.1 10.6 � 0.12 15.21 � 5.3
25 GSHþ 10 AA 20 0.97 � 0.6 69.6 � 3.8 3.52 � 0.63 3.33 � 1.9 8.4 � 0.48 9.11 � 5.3
40 1.74 � 0.6 132.3 � 42.5 3.67 � 0.23 3.44 � 0.1 11.6 � 0.78 21.05 � 6.4
25 GSHþ 40 Toco 20 0.99 � 0.6 72.4 � 3.8 2.82 � 0.33 3.50 � 2.0 9.2 � 0.56 12.35 � 7.1
40 2.27 � 0.2 153.0 � 10.0 3.00 � 0.58 3.20 � 0.1 12.6 � 0.12 15.26 � 7.1
25 GSHþ 10 Cyst 20 0.60 � 0.1 50.4 � 6.2 2.73 � 0.34 2.5 � 1.4 11.0 � 0.49 9.84 � 5.1
40 1.46 � 0.1 99.6 � 15.3 2.33 � 0.23 2.50 � 0.1 7.0 � 0.26 8.75 � 5.1
*Mean of 5 replicate shoots.
AA e ascorbic acid; GSH e glutathione reduced; Toco e tocopherol; Cyst e cysteine.
b i om a s s a n d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 9 1863
of each) were used individually in the medium to remove
browning of in vitro-proliferating shoots and for their further
growth and development (Table 1). Later, on the basis of their
responses in culture these antioxidants were used in
combination. Four different combinations used were;
50 mg dm�3 glutathione red and 10 mg dm�3 ascorbic acid
(St1); 25 mg dm�3 glutathione red and 10 mg dm�3 ascorbic
acid (St2); 25 mg dm�3 glutathione red and 20 mg dm�3
tocopherol (St3); 25 mg dm�3 glutathione red and 10 mg dm�3
cysteine (St4). Different parameters were recorded after 40 d
of culture incubation and presented in Table 2 and Table 3.
Tocopherol was used from Evion� capsules (400 mg, Merck
Co. Ltd.) with a sterilized syringe and dissolved in 70%
alcohol. It was used directly in sterilized medium.
Half strength basal medium having the optimized
concentration and combination of antioxidants was used for
rooting of isolated shoots. Different concentrations of IBA
were used for root induction. During initial 10e15 d of hard-
ening, high humidity was maintained by covering the plants
with acclimatization hoods and irrigated with tap water.
For next 15 d, acclimatization hoods were raised slightly
decreasing the humidity slowly. Finally, the plants were then
grown under open field conditions.
Table 3 e Chlorophyll estimation in different treatments of an
Cultureincubation (d)
Parameters studies(mg g�1 FW)
C � SE
0 Total chlorophyll 0.66 � 0.12
Chlorophyll a 0.47 � 0.08
Chlorophyll b 0.20 � 0.02
Carotenoids 0.23 � 0.03
15 Total chlorophyll 0.48 � 0.06
Chlorophyll a 0.40 � 0.02
Chlorophyll b 0.08 � 0.01
Carotenoids 0.19 � 0.01
30 Total chlorophyll 0.40 � 0.02
Chlorophyll a 0.29 � 0.02
Chlorophyll b 0.11 � 0.01
Carotenoids 0.14 � 0.01
The pH of all the media was adjusted to 5.8 prior to the
addition of agar and autoclaved at 121 �C for 20 min. All the
media were solidified with 7 g dm�3 agar. Cultures were
incubated at 25� 2 �C under 16 h light and 8 h dark period.
Light at 60 mmol photon m�2 s�1 was supplied by fluorescent
tube lights fitted on culture racks.
2.3. Total phenolics
The content of the total phenolic in leaf extract was deter-
mined (according to Folin-Ciocalteu method) [23] following
the preparation of the caliberation curve where 1 cm3 aliquot
0.024, 0.075, 0.105 and 0.3 mg cm�3 ethanolic gallic acid solu-
tion was mixed with 5 cm3 Folin-Ciocalteu reagent (diluted 10
fold) and 4 cm3 sodium carbonate (75 g dm�3). The absorbance
at 765 nmwasmeasured after 1 h at 20 �C and the caliberation
curve was drawn. To the same reagent (Folin-Ciocalteu
reagent & sodium carbonate) 1 cm3 methanolic plant extract
(10 g dm�3) was mixed as described above and after 1 h the
absorbance was measured. All determinations were per-
formed in triplicates. Total phenolic content in plant meth-
anolic extracts in Gallic Acid Equivalents (GAE) was measured
by the formula:
tioxidants in J. curcas at different time intervals.
Treatments
St1 � SE St2 � SE St3 � SE St4 � SE
e e e e
e e e e
e e e e
e e e e
0.73 � 0.15 0.97 � 0.20 0.87 � 0.18 0.71 � 0.08
0.54 � 0.02 0.74 � 0.17 0.64 � 0.17 0.52 � 0.02
0.19 � 0.09 0.22 � 0.06 0.23 � 0.09 0.19 � 0.01
0.21 � 0.01 0.30 � 0.02 0.30 � 0.10 0.25 � 0.02
0.64 � 0.18 0.84 � 0.23 0.81 � 0.21 0.57 � 0.02
0.51 � 0.18 0.60 � 0.16 0.65 � 0.19 0.47 � 0.02
0.13 � 0.07 0.20 � 0.02 0.16 � 0.03 0.10 � 0.01
0.14 � 0.02 0.30 � 0.02 0.28 � 0.04 0.23 � 0.01

b i om a s s an d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 91864
C ¼ c,V=m
Where, C-total content of phenolic compounds, mg g�1 of
plant extract, in GAE; c-the concentration of gallic acid
deduced from the calibration curve, mg cm�3; V-volume of
extract, cm3; m-the DW of the plant material.
2.4. Estimation of chlorophyll and carotenoids
Fresh leaves (100mg) each from control, St1, St2, St3 and St4
treatments were crushed in 5 cm3 (80%v v�1) chilled acetone
preparedwith TDW. Following the standardmethod (according
to Arnon) [24], extract was centrifuged at 166.67 Hz for 10min.
Absorbanceofsupernatantwasreadat663,645,510and480 nm.
Total chlorophyll content along with its component i.e. chloro-
phyll a and chlorophyll b was calculated in mg g�1 FW using
following formulas (according to Machlachalan and Zalic) [25].
Total Chlorophyll ¼ ½20:2ðA645Þ þ 8:02ðA663Þ� � V1000� V
Chlorophyll a ¼ ½12:7ðA663Þ þ 2:63ðA645Þ� � V1000� V
Chlorophyll b ¼ ½22:9ðA645Þ � 4:68ðA663Þ� � V
1000� V
Carotenoid content was calculated in mg g�1 FW using
following formula (according to Duxbury and Yentsch) [26].
Carotenoid ¼ ½7:6ðA480Þ � 2:63ðA510Þ� � V1000� V
2.5. Measurement of H2O2
Fresh leaves (500 mg) from in vitro regenerated J. curcas shoots
were taken each from control (without antioxidant), and from
different combinations of antioxidants (St1eSt4), at 15 and
30 d culture incubation, for studying H2O2 content. The H2O2
content of the leaves was measured spectrophotometrically
after reaction with potassium iodide (KI) [27]. The reaction
mixture consisted of 0.5 cm3 of 0.1% TCA (v v�1), leaf extract
supernatant, 0.5 cm3 of 10% potassium phosphate buffer
(w v�1, pH 7.8) and 2 cm3 reagent of 100% KI (w v�1), in fresh
double distilled water. The blank probe consisted of 0.1% TCA
(v v�1) in the absence of leaf extract. After 1 h of reaction in
darkness, the absorbance was measured at 390 nm. The
amount of hydrogen peroxidewas calculated using a standard
curve prepared with known concentrations of H2O2.
2.6. Enzyme extraction
Samples of leaves from in vitro cultures of different treatments
of antioxidants were collected at 15 and 30 d of culture incu-
bation and extracted in 10% potassium phosphate buffer
(w v�1, pH 7.8), containing 1% PVP (w v�1) and 0.5% Triton X
100 (v v�1) and 0.01% EDTA (w v�1) for enzymatic study. The
homogenisation was done using liquid nitrogen in the ratio of
1:2 (w v�1) of tissue to buffer for enzymatic activity. The
homogenate was centrifuged in a refrigerated centrifuge at
250 Hz for 30 min at 4 �C (Eppendorf). The supernatant was
collected, aliquoted and stored at �80 �C for subsequent
analysis. All operations were conducted at 0e4 �C.
2.7. Protein determination
Total protein of all the samples was determined (according to
Bradford) [28] using bovine serum albumin (BSA) as a stan-
dard. Enzyme activities and isoenzyme patterns were deter-
mined by themethods specific to each enzyme and isoenzyme
as described below. All enzyme assays were performed at
25 �C and activity was measured spectrophotometrically on
PerkinElmer, Lambda 35, UV/VIS Spectrometer.
2.8. Antioxidant enzyme activity
For measuring superoxide dismutase (SOD, EC 1.15.1.1)
activity (according to Nishikimi et al.) [29], the reaction
mixture contained 52% sodium pyrophosphate buffer (w v�1,
pH 8.3), 0.1% EDTA (w v�1), 0.018% PMSF (w v�1), 0.03% NBT
(w v�1) and 0.078% NADH (w v�1). Reaction was started by
addingNADH. Reactionmixturewas incubated at 30 �C for 90 s
and stopped by adding 1 cm3 glacial acetic acid and stirred
vigorously. Colour intensity of the chromogen in the reaction
mixture was measured at 560 nm against sodium pyrophos-
phate buffer. One unit of enzyme activity is defined as the
amount of enzyme required to inhibit the NBT reduction by
50%, under the assay conditions.
Total GPX (EC 1.11.1.7) activity was determined (according
to Hemeda and Klim) [30] monitoring the formation of tetra-
guaiacol from guaiacol. The reaction mixture contained 10%
sodiumphosphate buffer (w v�1, pH 5.8), 1% guiacol (v v�1) and
0.3% H2O2 (v v�1). Increase of absorbance due to oxidation of
guiacol was recorded at 470 nm every 15 s for 3 min.
For measuring APX (EC 1.11.1.11) activity (according to
Nakano and Asada) [31], the reaction mixture contained 5%
potassium phosphate buffer (w v�1, pH 7.0), 1.0% sodium
ascorbic acid (w v�1), 0.1% H2O2 (v v�1) and 0.1% EDTA (w v�1).
The reaction mixture started by addition of H2O2. The
decrease in absorbance was recorded at 290 nm at an interval
of 15 s up to 3 min per cycle period. The blank was determined
by the addition of phosphate buffer instead of extracts in the
reaction mixture.
For measuring the activity of GR (EC 1.6.4.2) (according to
Smith et al.) [32], the reaction mixture contained 20% potas-
sium phosphate buffer (w v�1, pH 7.5), containing 0.1% EDTA
(w v�1), 0.3% 5-50dithiobis (2-nitrobenzoic acid, w v�1) in 1.0%
potassium phosphate buffer (w v�1, pH 7.5), 0.2% NADPH
(w v�1) and 2.0% GSSG (w v�1). All the reagents were added
directly to cuvette and reaction was started by addition of
GSSG andwasmonitored for 5 min at 412 nm. Enzyme activity
was expressed as picomoles of GSSG reduced min�1 g�1 FW.
GSSG was replaced by water in the reaction mixture as blank.
2.9. Statistical analysis of data
Morphological changes were recorded by visual observation.
The values of data in case of in vitro shoots are presented as
mean� SE of five replicate cultures. All other analyses were
carried out in triplicate. Duncan’s multiple-range test was
used as a post hoc comparison of statistical significance
( p values<0.05). All statistical analyses were performed using
Statistical for Windows (version 5.1, StatSoft, Inc., Tulsa, OK).

b i om a s s a n d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 9 1865
3. Results and discussion
3.1. Effects of antioxidants on growth of cultures
Shoot proliferation started in MSmedium having 15mg dm�3
L-glutamine and L-arginine supplemented with 0.5 mg dm�3
BA and 0.1 mg dm�3 IBA along with 10 mg dm�3 AdS, but leaf
margins started turning yellowish brown within 15 d of
culture incubation (Fig. 1a, Table 1). Later, whole leaves
necrosed within 30 d (Fig. 1b). Therefore, initially, two
concentrations of each of four antioxidants were incorpo-
rated in the basal media individually. With the addition of
antioxidants, overall growth of shoots was observed better
and the shoots remained fresh, green and healthy for
a longer period as against the shoots without antioxidants.
Regeneration of shoots was not affected with the addition or
lack of antioxidants in the medium. Lower concentrations of
all the antioxidants used were more effective to keep the
cultures fresh and green, as compared to higher concentra-
tions except GSH where shoots grew more vigorous than the
control but those were associated with callus formation
(Fig. 1c). Both the concentrations of GSH were almost equally
effective to keep the shoots fresh and green up to 25 d beyond
which shoots started browning. The shoots growing in lower
concentrations of antioxidants did not show associated
callus except AA. The AA (10 or 20 mg dm�3) was least
effective to remove browning as well as regeneration of
shoots in in vitro-growing cultures (Fig. 1d, Table 1). Similar
results had been observed by Zheng et al. [33] in peanut
regeneration, where they found the highest regeneration
with the addition of GSH, then tocopherol and least in the
Fig. 1 e Effect of antioxidants on shoot growth of Jatropha curcas;
browning and necrosis of leaves in control after 30 d of incubat
glutathione; (d) 20 mg dmL3 ascorbic acid; (e) 40 mg dmL3 toco
glutathione D 10 mg dmL3 ascorbic acid; (h) 25 mg dmL3 gluta
tocopherol D 10 mg dmL3 ascorbic acid; (j) 10 mg dmL3 cystein
presence of ascorbic acid. They explained that AA is an
unsuitable antioxidant in MS medium due to the stimulation
of oxidation in the presence of iron in MS media. In the
present communication, addition of tocopherol resulted in
sturdier shoots with more number of leaves (Fig. 1e),
whereas addition of cysteine did not prove beneficial to
protect the browning of cultures, although an average of 3.2
shoots regenerated in its lower concentration (Fig. 1f).
Similar to our results Grewal et al. [34] also found increased
regeneration with the addition of cysteine in Indica rice.
From this experiment, GSH was selected as the most effec-
tive antioxidant, as in its presence, the shoots remained
fresh, green and healthy for about 25 d. The role of gluta-
thione is already known in promoting callus and shoot
growth in Malus pumila [35], as stimulant of shoot organo-
genesis in Gladiolus hybridus [36], and also in controlling
hyperhydricity, tissue necrosis and increased plant regen-
eration in Ramonda myconi [37].
In the next experiment GSH was used with addition of
lower concentrations of AA, tocopherol and cysteine in
separate four treatments (St1eSt4). Data has been recorded
at 20 and 40 d and presented in Table 2. In the presence of
higher concentration of glutathione (40 mg dm�3) along
with AA (10 mg dm�3) in St1, shoots grew up to 3.36 cm of
height but necrosis of shoots started after 25 d of culture
incubation (Fig. 1g). However, lowering the concentration
of glutathione to 25 mg dm�3 in St2 treatment resulted in
growth of proliferating shoots with highest leaf canopy
area. Shoots remained fresh, green and regenerative up to
40 d of culture incubation without any subculture (Fig. 1h).
The combination of 25 mg dm�3 GSHþ 40 mg dm�3
tocopherol (St3) also proved beneficial as a maximum
(a) yellowing of leaves in control after 15 d of incubation; (b)
ion; (cej) cultures after 30 d of incubation, (c) 50 mg dmL3
pherol; (f) 10 mg dmL3 cysteine; (g) 50 mg dmL3
thione D 10 mg dmL3 ascorbic acid; (i) 40 mg dmL3
e D 10 mg dmL3 ascorbic acid.

Fig. 2 e Propagation of J. curcas; (a) in vitro-proliferation of shoots; (b) in vitro rooting; (c) 4-wk-old in vitro-raised plants in pots.
Fig. 3 e Estimation of total phenolic compounds in in vitro
shoots of J. curcas at 0, 15 and 30th d of culture incubation.
b i om a s s an d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 91866
number of 12.6 leaves could be developed in this treatment,
but the shoots formed started browning after 35 d of
culture incubation (Fig. 1i). In the presence of cysteine
(10 mg dm�3, St4) best regeneration of shoots was observed
but browning started after 23 d of culture incubation
(Fig. 1j). The importance of AA has been shown in
controlling lethal browning and disease incidence of tissue
culture shoots in Cavendish banana [38]. Tocopherol was
reported beneficial for stimulated shoot organogenesis in G.
hybridus [36], while cysteine in Dactylis glomerata [39]. On
the basis of our results obtained, St2 (25 mg dm�3
GSHþ 10 mg dm�3 AA) was selected as the best treatment
for biomass accumulation, where shoots remained fresh,
green, healthy and regenerative up to 40 d. After excision of
isolated grown-up shoots, the remaining regenerative
small shoots had been subcultured into fresh medium and
like this it became a repository of shoots for a long-term
period.
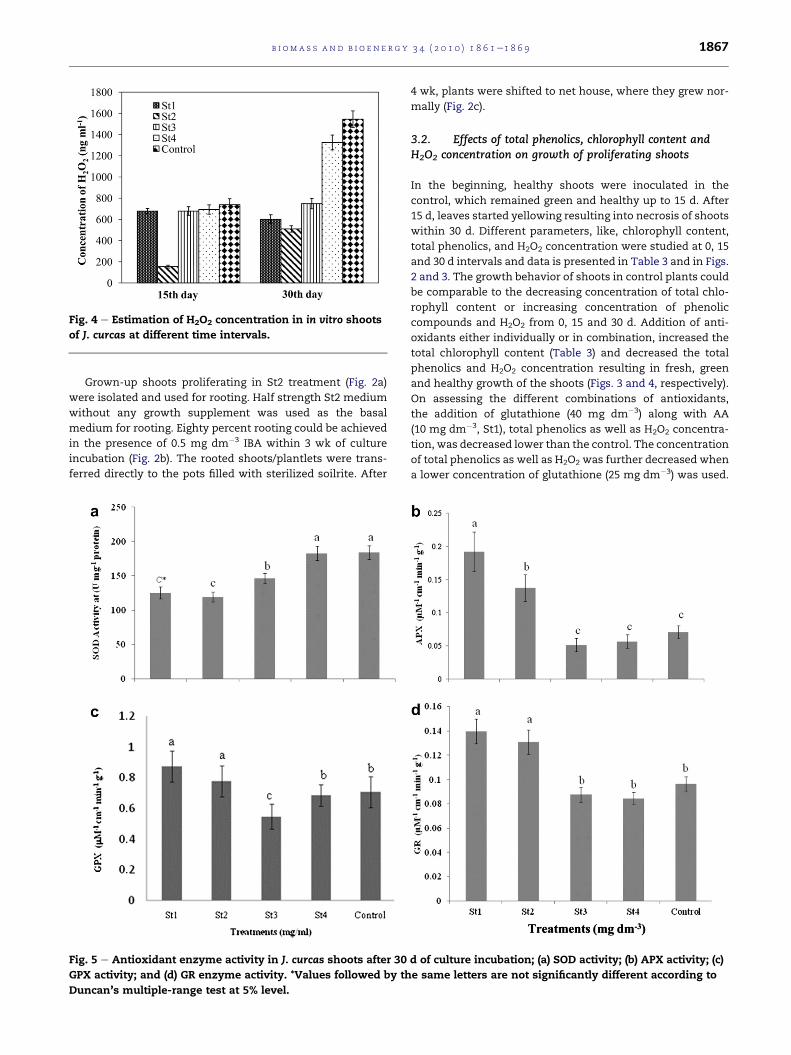
Fig. 4 e Estimation of H2O2 concentration in in vitro shoots
of J. curcas at different time intervals.
b i om a s s a n d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 9 1867
Grown-up shoots proliferating in St2 treatment (Fig. 2a)
were isolated and used for rooting. Half strength St2 medium
without any growth supplement was used as the basal
medium for rooting. Eighty percent rooting could be achieved
in the presence of 0.5 mg dm�3 IBA within 3 wk of culture
incubation (Fig. 2b). The rooted shoots/plantlets were trans-
ferred directly to the pots filled with sterilized soilrite. After
Fig. 5 e Antioxidant enzyme activity in J. curcas shoots after 30
GPX activity; and (d) GR enzyme activity. *Values followed by th
Duncan’s multiple-range test at 5% level.
4 wk, plants were shifted to net house, where they grew nor-
mally (Fig. 2c).
3.2. Effects of total phenolics, chlorophyll content andH2O2 concentration on growth of proliferating shoots
In the beginning, healthy shoots were inoculated in the
control, which remained green and healthy up to 15 d. After
15 d, leaves started yellowing resulting into necrosis of shoots
within 30 d. Different parameters, like, chlorophyll content,
total phenolics, and H2O2 concentration were studied at 0, 15
and 30 d intervals and data is presented in Table 3 and in Figs.
2 and 3. The growth behavior of shoots in control plants could
be comparable to the decreasing concentration of total chlo-
rophyll content or increasing concentration of phenolic
compounds and H2O2 from 0, 15 and 30 d. Addition of anti-
oxidants either individually or in combination, increased the
total chlorophyll content (Table 3) and decreased the total
phenolics and H2O2 concentration resulting in fresh, green
and healthy growth of the shoots (Figs. 3 and 4, respectively).
On assessing the different combinations of antioxidants,
the addition of glutathione (40 mg dm�3) along with AA
(10 mg dm�3, St1), total phenolics as well as H2O2 concentra-
tion, was decreased lower than the control. The concentration
of total phenolics as well as H2O2 was further decreased when
a lower concentration of glutathione (25 mg dm�3) was used.
d of culture incubation; (a) SOD activity; (b) APX activity; (c)
e same letters are not significantly different according to

Fig. 6 e APX isoenzymes in J. curcas shoots in different
treatments of antioxidants (from L to R-C, St1, St2, St3 and
St4) after 30 d of culture incubation.
b i om a s s an d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 91868
The results are compatible with the data presented in Table 3
showing higher chlorophyll content in St2 than St1. Total
chlorophyll content was more after 15 d than in 30 d, in all the
treatments including the control. Addition of tocopherol to
the medium again resulted in decrease of total phenolics as
well as H2O2 than the control but the decrease was less than
St1 and St2, while total chlorophyll content (higher but not
the highest) resulted in greening and better growth of leaves/
shoots. However, the addition of cysteine, could not sustain
good growth, freshness and green colour of the shoots due to
higher phenolic content and higher H2O2 concentration
which caused stress to the regenerating shoots. Total chlo-
rophyll content was also the least in St4 resulting in the
yellowing of shoots.
3.3. Antioxidant enzyme activity
High SOD activity increases concentration of H2O2 in the
tissue which triggers programmed cell death ultimately
causing browning and tissue necrosis. In J. curcas shoots, SOD
activity was found very high in the control which subse-
quently increases H2O2 content that could possibly lead to
oxidative damage, a ‘negative’ consequence of its presence
and a common response to many environmental stresses
leading to shoot browning/necrosis. Addition of different
antioxidants reduces SOD activity resulting in fresh, green
and healthy shoots. St2 treatment showing the lowest SOD
activity was found as the optimum treatment for the growth
of the cultures (Fig. 5a). Similar results of decreased SOD
activity have been reported inducing shoot organogenesis in
gladiolus [36]. It has been reported in chrysanthemum that
a decrease in the activity of a number of antioxidant enzymes
that normally prevent the buildup of free radicals can be
accounted for senescence in plants [19].
APX, GR and GPX decreased in treatments St1 and St2 as
compared to control (Fig. 5b and d). APX plays a key role in the
ascorbateeglutathione cycle by reducing H2O2 to water using
the reducing power of ascorbate and producing mono-
dehydroascorbate (MDHA). Since >90% of total leaf APX
activity is localized in the chloroplast [40], this activity can
represent the best estimation of chloroplast-based detoxifi-
cation of ROS via the Mehlerperoxidase pathway as sink for
reducing equivalents. Native PAGE is also supporting the
activity data, and a single band was clearly visible in all the
treatments (Figs. 5b and 6). Since it has already been observed
that chlorophyll content was highly reduced in control indi-
cating chloroplast degradation under necrotic condition
potentially increasing the probability of generating ROS
(Table 3). According to Marchner and Possingham oxidative
stress resulted in a reduction in the number of chloroplasts
[41]. GPX is involved in the removal of hydrogen peroxide and
organic peroxides [42] showing the increased activity in St1
and St2 as compared to control (Fig. 5c). Similarly, GR is
involved in the recycling of reduced glutathione to maintain
a constant intracellular concentration of GSH [43]. Elevated
concentrations of GSH are associatedwith increased oxidative
stress tolerance which is found true in our study (Fig. 5d).
The finding that control shoots had significantly lower
chlorophyll and higher phenolic contents than healthy shoots
which led us to conclude that control shoots after 30 d of
culture caused a substantial decline in photosynthesis
because the antioxidant enzyme system was swamped by
a massive accumulation of H2O2, the production of which is
favoured when metabolic pathways are down-regulated. The
resulting imbalance between H2O2 production and antioxi-
dants defences resulted in oxidative damage of the photo-
synthetic machinery in the chloroplasts resulting in tissue
browning/necrosis. However, supplementation of antioxi-
dants like GSH/AA resulting in increase antioxidant metabo-
lism in determining the ability of the plants to survive in
oxidative stress, an up regulation of these enzymes would
help to reduce the built up of H2O2.
The problem of necrosis is the main constraint in the large
scale propagation of J. curcas to obtain quality planting mate-
rial including higher biomass in long-term culture. In the
present study, we have manipulated the problem by using
different antioxidants and their combinations in the medium
so as to obtain long-term proliferating cultures of J. curcas
which remained fresh, green, healthy and regenerative up to
40 d of culture incubation.
Acknowledgements
The authors are thankful to the Council of Scientific and
Industrial Research, New Delhi, Govt. of India, for providing
the financial support to carry out this work.

b i om a s s a n d b i o e n e r g y 3 4 ( 2 0 1 0 ) 1 8 6 1e1 8 6 9 1869
r e f e r e n c e s
[1] Debnath M, Bisen PS. Jatropha curcas L., a multipurposestress resistant plant with a potential for ethnomedicine andrenewable energy. Curr Pharm Biotech 2008;9:288e306.
[2] Chhetri AB, Martin S, Tango MS, Budge SM, Watts KC,Islam MR. Non-edible plant oils as new sources for biodieselproduction. Int J Mol Sci 2008;9:169e80.
[3] Forson FK. Performance of Jatropha oil blends in a dieselengine. Renew Energy 2004;29:1135e45.
[4] Runge CF, Senauer B. Biofuel: corn isn’t the king of thisgrowing domain. Nature 2007;450:478.
[5] Pamidiamarri DVNS, Pandya N, Reddy MP, Radhakrishnan T.Comparative study of interspecific genetic divergence andphylogenic analysis of genus Jatropha by RAPD and AFLP. MolBiol Rep 2009;36:901e7.
[6] Basha SD, Sujatha M. Inter and intra-population variability ofJatropha curcas (L.) characterized by RAPD and ISSR markersand development of population-specific SCAR markers.Euphytica 2007;156:375e86.
[7] He Y, Guo X, Lu R, Niu B, Pasapula V, Hou P, et al. Changes inmorphology and biochemical indices in browning callusderived from Jatropha curcas hypocotyls. Plant Cell TissueOrgan Cult 2009;98:11e7.
[8] Sujatha M, Mukta N. Morphogenesis and plant regenerationfrom tissue cultures of Jatropha curcas. Plant Cell TissueOrgan Cult 1996;44:135e41.
[9] Wei Q, Lu W-D, Liao Y, Pan S-L, Xu Y, Tang L, et al. Plantregeneration from epicotyl explant of Jatropha curcas. J PlantPhysiol Mol Biol 2004;30:475e8.
[10] Datta MM, Mukherjee P, Ghosh B, Jha TB. In vitro clonalpropagation of biodiesel plant (Jatropha curcas L.). Curr Sci2007;93:1438e42.
[11] Deore AJ, Johnson TS. High frequency plant regenerationfrom leaf-disc cultures of Jatropha curcas L.: an importantbiodiesel crop. Plant Biotech Rep 2008;2:7e11.
[12] Sujatha M, Makkar HPS, Becker K. Shoot bud proliferationfrom axillary nodes and leaf sections of non-toxic Jatrophacurcas L. Plant Growth Regul 2005;47:83e90.
[13] Sardana J, Batra A, Ali DJ. An expeditious method forregeneration of somatic embryos in Jatropha curcas L.Phytomorphology 2000;50:239e42.
[14] Jha TB, Mukherjee P, Datta MM. Somatic embryogenesis inJatropha curcas Linn., an important biofuel plant. PlantBiotech Rep 2007;1:135e40.
[15] Wei T, Newton RJ. Increase of polyphenol oxidase anddecrease of polyamines correlate with tissue browning inVirginia pine (Pinus virginiana Mill). Plant Sci 2004;167:621e8.
[16] Prochazkova D, Sairam RK, Srivastava GC, Singh DV.Oxidative stress and antioxidant activity as the basis ofsenescence in maize leaves. Plant Sci 2001;161:765e71.
[17] Dhindsa RA, Plumb-Dhindsa P, Thorpe PA. Leaf senescence:correlated with increased permeability and lipidperoxidation, and decreases levels of superoxide dismutaseand catalase. J Expl Bot 1981;126:93e101.
[18] Munne-Bosch S, Penuelas J. Photo- and antioxidativeprotection during summer leaf senescence in Pistacialentiscus L. grown under mediterranean field conditions. AnnBot 2003;92:385e91.
[19] Chakrabarty D, Chatterjee J, Datta SK. Oxidative stress andantioxidant activity as the basis of senescence inchrysanthemum florets. Plant Growth Regul 2007;53:107e15.
[20] Bowler C, van Camp W, van Montagu M, Inze D. Superoxidedismutase in plants. CRC Crit Rev Plant Sci 1994;13:199e208.
[21] Noctor G, Foyer CH. Ascorbate and glutathione: keepingactive oxygen under control. Annu Rev Plant Physiol PlantMol Biol 1998;49:249e79.
[22] Murashige T, Skoog F. A revised medium for rapid growthand bioassays with tobacco tissue cultures. Physiol Plant1962;15:473e97.
[23] Folin O, Ciocalteu V. On tyrosine and tryptophandetermination in proteins. J Biol Chem 1927;27:627e50.
[24] Arnon D. Copper enzymes in isolated chloroplasts e
polyphenoloxidase in Beta vulgaris. Plant Physiol 1949;24:1e15.
[25] Machlalan S, Zalic S. Plastid structure, chlorophyllconcentration and Tee amino acid composition ofa chlorophyll mutant barley. Can J Bot 1963;41:1053e62.
[26] Duxbury AC, Yentsch CS. Plankton pigment monograph. JMarine Res 1956;15:92.
[27] Alexieva V, Sergiev I, Mapelli S, Karanov E. The effect ofdrought and UV on growth and stress markers in pea andwheat. Plant Cell Environ 2001;24:1337e44.
[28] Bradford MM. A rapid and sensitive method for thequantization of microgram quantities of protein utilizing theprinciple of protein-dye binding. AnnBiochem1976;72:248e54.
[29] Nishikimi M, Appaji N, Yagi K. The occurrence of superoxideanion in the reaction of reduced phenazine methosulfateand molecular oxygen. Biochem Biophy Res Commun 1972;46:849e54.
[30] Hemeda HM, Klein BP. Effects of naturally occurringantioxidants on peroxidase activity of vegetable extracts. JFood Sci 1990;55:184e6.
[31] Nakano Y, Asada K. Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts. PlantCell Physiol 1981;22:867e80.
[32] Smith IK, Vierheller TL, Thorne CA. Assay of glutathionereductase in crude tissue homogenates using 5,50-dithiobis(2-nitrobenzoic acid). Ann Biochem 1988;175:408e13.
[33] Zheng Q, Ju B, Liang L, Xiao X. Effects of antioxidants on theplant regeneration and GUS expressive frequency of peanut(Arachis hypogaea) explants by Agrobacterium tumefaciens.Plant Cell Tissue Organ Cult 2005;81:83e90.
[34] Grewal D, Gill R, Gosal S. Role of cysteine in enhancingandrogenesis and regeneration of Indica rice (Oryza sativa L.).Plant Growth Regul 2006;49:43e7.
[35] Nomura K, Matsumoto S, Masuda K, Inoue M. Reducedglutathione promotes callus growth and shoot developmentin a shoot tip culture of apple root stock M.26. Plant Cell Rep1998;17:597e600.
[36] Dutta Gupta S, Datta S. Antioxidant enzyme activities duringin vitro morphogenesis of gladiolus and the effect ofapplication of antioxidants on plant regeneration. Biol Plant2003;47:179e83.
[37] Toth S, Scott P, Sorvari S, Toldi O. Effective and reproducibleprotocols for in vitro culturing and plant regeneration of thephysiological model plant Ramonda myconi (L.) Rchb. Plant Sci2004;166:1027e34.
[38] Ko WH, Su CC, Chen CL, Chao CP. Control of lethal browningof tissue culture plantlets of Cavendish banana cv.Formosana with ascorbic acid. Plant Cell Tissue Organ Cult2009;96:137e41.
[39] Lee KW, Lee SH, Lee DG, Woo HS, Kim DH, Choi MS, et al.Effect of plant growth regulators and antioxidants on callusinduction and plant regeneration from seed culture oforchardgrass. J Korean Soc Grassl Sci 2005;25:191e8.
[40] Gillham DJ, Dodge AD. Hydrogen peroxide scavengingsystems in pea chloroplasts. Planta 1986;167:246e51.
[41] Marschner H, Possingham JV. Effects of K and Na on thegrowth of leaf discs of sugar beet and spinach. ZPflanzenphysiol 1975;75:6e16.
[42] Flohe L, Gunzler WA. Assays of glutathione peroxidase.Methods Enzymol 1984;105:141e8.
[43] Calbert I, Mannervik B. Glutathionereductase. MethodsEnzymol 1985;113:484e90.
Copyright © 2022 FDOKUMEN




















