
The human ALL-1/MLL/HRX antigen is predominantly localized in the nucleus of resting and...
8
1997;57:2035-2041. Published online May 1, 1997. Cancer Res Maria-Grazia Ennas, Claudio Sorio, Ralf Greim, et al. Blood Mononuclear Cells Peripheral Localized in the Nucleus of Resting and Proliferating The Human ALL-1/MLL/HRX Antigen Is Predominantly Updated Version http://cancerres.aacrjournals.org/content/57/10/2035 Access the most recent version of this article at: Citing Articles http://cancerres.aacrjournals.org/content/57/10/2035#related-urls This article has been cited by 8 HighWire-hosted articles. Access the articles at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications American Association for Cancer Research Copyright © 1997 on July 13, 2011 cancerres.aacrjournals.org Downloaded from
-
Upload
uni-frankfurt -
Category
Documents
-
view
0 -
download
0
Transcript of The human ALL-1/MLL/HRX antigen is predominantly localized in the nucleus of resting and...

1997;57:2035-2041. Published online May 1, 1997.Cancer Res Maria-Grazia Ennas, Claudio Sorio, Ralf Greim, et al. Blood Mononuclear Cells
PeripheralLocalized in the Nucleus of Resting and Proliferating The Human ALL-1/MLL/HRX Antigen Is Predominantly
Updated Version http://cancerres.aacrjournals.org/content/57/10/2035
Access the most recent version of this article at:
Citing Articles http://cancerres.aacrjournals.org/content/57/10/2035#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
[email protected] atTo order reprints of this article or to subscribe to the journal, contact the AACR Publications
To request permission to re-use all or part of this article, contact the AACR Publications
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

[CANCERRESEARCH57, 2035-2041. May 15. 19971
ABSTRACT
The ALL-I gene is an important regulator of embryonal and hematopoletic development, and structural variants of the human gene generatedby chromosomal translocations and other genomic alterations presumablyact as oncogenesin the pathogenesisof acuteleukemiasand otherhematological malignancies. Antisera against two different epitopes of thehuman ALL-i protein (anti.ALL1-N and anti-ALLi-C) were produced.Both sera revealed Indistinguishable patterns of antigen localization inhuman peripheral blood mononuclear cells (PBMCs). In resting PBMCs,the antigen was distributed in a speckled pattern across the nuclei, with anincreased density at the nuclear envelope and the nuclear indentation. Inmitotically stimulated PBMCs, the antigen surrounded the condensingchromosomes but did not colocalize with chromatin or the nuclear scaf.fold. The antigen is considered a marker for a novel nuclear subcompartmeat, a perichromosomal area termed the “chromosomalenvelope.―InWestern blot experiments, the anti-ALLi-N serum reacted with apolypeptide corresponding to the expected full-length 430-kDa ALL-i
protein. Recombinant proteins representing the AT-hook and zinc bindingsubdomains of the ALL-i protein interacted in vitro with a degeneratemixture of double-stranded oligodeoxynucleotides. Thus, the ALL-i protein probably is a DNA-binding protein with both a sequence-unspecific(AT-hook) and a sequence-specific (zinc binding subdomains) doublestranded DNA binding mode.
INTRODUCTION
Genetic alterations of the human ALL-]/MLUHRX (AU-]) gene,located at chromosomal position 11q23, are consistently associatedwith acute malignancies of hematopoietic cells. Among these areacute lymphoblastic and myeloid leukemias with extremely pooroutcome (1—6).The molecular alterations include chromosomal translocations, internal duplications, and specific internal deletions (7—11).
The ALL-] gene was first mapped to a 92-kb DNA fragment locatedbetween the CD3@ and Thy-i loci (12). The complete cDNA has beencloned and sequenced (13, 14), and the exon-intron structure of theALL-] gene has been determined recently (15—17).The gene consistsof at least 37 exons that encode an open reading frame of 12,018 bp(16, 17). The predicted protein is composed of 4,005 amino acids andshows homology to the Trithorax protein, a regulatory protein involved in Drosophila embryogenesis (18—20). The human ALL-lprotein shares similar length and domains along the protein axis withthe Trithorax protein. Knockout experiments of the mouse ALL-] generevealed a role of the ALL-l protein in normal hematopoietic development (21), a homeotically transformed phenotype during embryonaldevelopment, and hematopoietic dysfunctions in mice heterozygous
Received 1/22197;accepted 3/25/97.I Supported by Research Grants 5FB466-C4 (to R. M. and J. G.) and SFB466-C5 (to
G.H.F. andR.M.)fromtheDeutscheForschungsgemeinschaft;ResearchGrantMar/93from Johannes and Frieda Mambo Stiftung (to R. M. and J. G); a research grant from theAssociazioneItalianaRicercaCancroto theIstitutoAnatomiaPatologicaof Verona;anda seed grant from the Associazione Italiana Ricerca Cancro (to C. S.). R. M. wassupported by a Career Development Award from the Ria Freifrau von Fritsch Stiftung.M-G.E., C. S., and R. G. contributedequally to this work.
2 To whom requests for reprints should be addressed, at Department of Genetics,
University of Erlangen-Ntlrnberg, Staudtstrasse 5, D 91058 Erlangen, Germany. Phone:49-9131-858530; Fax: 49-9131-858626; E-mail: [email protected].
for the deletion, whereas a homozygous deletion led to embryoniclethality (22). These findings suggested a similar role in embryogenesis for the murine ALL-l and Drosophila Trithorax proteins.
Several domains of the human ALL-l protein have DNA-bindingand transcriptional activation activities (23, 24). At the amino-terminal part, AT-hooks and SPKK motifs have been identified (14),representing minor groove DNA binding motifs similar to thosepresent in HMG-Y proteins (25). For the AT-hook motif, a DNAbinding activity to presumably cruciform DNA molecules has alreadybeen demonstrated (23). Motifs with homology to cysteine methyltransferase (26) and a predicted bipartite zinc binding motif werelocated further downstream (13, 14, 27). The predicted bipartite zincbinding domain was categorized as a PHD motif (28), due to homologies with plant homeo domain proteins, with potential DNA-bindingand/or protein-proteininteractingactivities(I1).
The aim of the present study was to analyze the intracellularlocalization of the human ALL-I protein in normal PBMCs.3 Fromthese experiments, we expected new insights into the functions of theALL-l protein in hematopoietic cells. Understanding the functions ofthe wild-type ALL-l protein may help to unravel the malignantprocess in hematopoietic cells that carry one of the known geneticalterations of the ALL-] gene.
MATERIALS AND METHODS
Cloning of Recombinant Proteins, Isolation and Immunization of Rabbits. Parts ofthe ALL-l cDNA [amino acids 192—503(AT-hook), amino acids1428—1513 (PHD-A), amino acids 1428—1576 (PHD-AB), amino acids 1513—
1663 (PHD-BC), amino acids 1428—1663(PHD-ABC), amino acids 2364—2764 (ALL1-C), and amino acids 3750—4005(SET)] were cloned into theprokaryotic expression vectors pETl5b (Invitrogen, NV Leek, Netherlands) or
pGEX-5T (Pharmacia, Freiburg, Germany). The former encodes a HHT, andthe latter encodes a HHT and a GST tag, respectively. Recombinant clones
were selected and verified by sequence analysis. Fusion proteins were inducedby isopropyl-thiogalactoside, and optimal expression was determined by meas
urement of the induction kinetics. Large-scale preparations were performed foreach construct under optimal conditions. Crude extracts were obtained afterlysis in 6 M guanidinium hydrochloride and fractionated on nickel-chelate
agarose columns (Quiagen, Hilden, Germany). All fractions containing recom
binant fusion proteins were pooled and rechromatographed over a second
affinity column. Successful purifications were monitored by SDS-PAGE, and
purified recombinant proteins were sequentially dialyzed against decreasingconcentrations of guanidinium hydrochloride in PBS and, finally, against PBS.A synthetic peptide including amino acids 446—456(ALL1-N) or a purifiedrecombinant protein (amino acids 2364—2764, ALLI-C) was used for immu
nizations of rabbits. The immunization was performed by s.c. injection of
lOO-p.g portions emulsified in complete Freund's adjuvant for the initialimmunization and emulsified in incomplete Freund's adjuvant for the booster
injections. Booster injections were performed every 14 days for a period of 3months. After 4 months, the rabbits were bled, and the serum was recovered.
3 The abbreviations used are: PBMC, peripheral blood mononuclear cell; HHT, hex
americ histidine tag; GST, glutathione S-transferase; PHA, phytohemagglutinin; PBT,PBS/0.1% Triton X-lOO; SWPD, South-Western protein dot blot; kDa, kilodalton(s); ds,double-stranded.
2035
The Human ALL-1IMLLIHRX Antigen Is Predominantly Localized in the Nucleusof Resting and Proliferating Peripheral Blood Mononuclear Cells1
Maria-Grazia Ennas, Claudio Sorio, Raif Greim, Mariella Nieddu, Aldo Scarpa, Simonetta Orlandini,Carlo M. Croce, Georg H. Fey, and Rolf Marschalek2
Department of Cytomorphology [M-G. E.J and Institute of Biology [M. N.J. Via Porcell 2. University of Cagliari, 09!OO Cagliari, Italy: Department of Genetics. Staudtstrasse 5,University ofErlangen-Nurnberg. 91058 Erlangen, Germany (R. G., G. H .F., R. M.J: Institute ofSurgical Pathology, Strada Le Grazie, University of Verona, 37!34 Verona, ItalyIC.S.,A. S.. S. 0.1; and KimmelCancerInstitute,JeffersonMedicalCollegeof ThomasJeffersonUniversity,Philadelphia,Pennsylvania19!07(C.M.C.J
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

NUCLEAR LOCALIZATION OF THE ALL-I ANTIGEN
The anti-ALL1-N immune serum was affinity-purified by absorption on affinity matrices carrying the recombinant antigen using standard procedures (29).
Cell Isolation and Cell Culture. Human mononuclear cells were isolatedfrom the peripheral blood of healthy donors after informed consent by centrifugation on Ficoll-Hypaque density gradients. Cells were cultured for 72 hin macrowells at 2 X 106 cells/mI in RPM! 1640 containing 10% FCS in the
presence of 1% v/v PHA (Life Technologies, Inc., Milan, Italy). PHA-stimulated cells were treated with 10 p@g/mlColcemid (Life Technologies, Inc.) for2 h. Mitotic preparations were obtained by hypotonic shock (0.07 M KC1 and
30 mM glycerol) for 15 mm at 37°C.Immunocytochemistry. Resting and PHA-stimulated PBMCs as well as
mitotic preparations were centrifuged for 4 mm at 700 rpm in a Shandon II
cytospin centrifuge, and the slides were fixed for 10 mm in methanol at —20°C
and immediately used for immunocytochemistry. The use of methanol fixationfor mitotic preparations, avoiding the classic Carnoy fixation after the hypotonic shock, was preferred to preserve the native structure and antigenicity of
cellular components.
Slides with resting or PHA-stimulated cells were rehydrated with PBS and
preincubated with phenylhydrazine (10 nmi) in PBS to block endogenousperoxidase activity. Slides were rinsed four times in PBS, with the addition of0.1% Triton X-lOO (PBT) in the last wash, and preincubated with normal goat
serum (1 : 10) for I S mm at room temperature. Preparations were incubated with
ALLI-C antiserum (I :200 in PBT/l% normal goat serum) or affinity-purifiedALL1-N antiserum (30 p@g/ml)for 2 h in a moist chamber. After three washsteps with PBT, the slides were incubated with biotinylated anti-rabbitt IgG(I :200; Vector Laboratories, Inc., Burlingame, CA) for 30 mm. After threemore washes, acetylated avidin-peroxidase was added (1 :250 in PBS; BlO,Milan, Italy). The reaction was developed with diaminobenzidine (1 mg/ml inPBS/0.03% H202). Negative controls were performed using preimmune sera orirrelevant rabbit IgG as appropriate.
Chromosome preparations were immunostained with the same procedure,except for using avidin DCS-FITC (1:1000 in PBS; Vector Laboratories, Inc.)after the secondary antibody. Preparations were finally stained with a fluorescence antifade solution containing propidium iodide (0.5 p@g/ml)and triethylenediamine (2% w/v; Sigma).
Western Blot Analysis. Mononuclear cells were harvested, washed inPBS, and lysed (20 X 106cells/mI) by adding radioimmunoprecipitation assaybuffer [150 mM NaCl; 20 mM HEPES; 1% Triton X-lOO; 0.5% sodiumdeoxycholate; 0.1% SDS; 1 m@iEDTA; I mr@iphenylmethylsulfonyl fluoride;5 @xg/m1leupeptin, pepstatin, chymostatin, and aprotinin; and I mM DTT (pH7.5)]. After rotating for 20 mm at 4°C, the lysate was centrifuged for 10 mm
at 14,000 x g, and the supernatant was collected. Protein concentrations weremeasured with the Bradford assay (Bio-Rad). Fifty @xgIlanewere subjected toSDS-PAGE in a 7% gel, which was then blotted onto nitrocellulose membraneusing the Bio-Rad Trans-Blot electrophoretic transfer cell (900 mA, 6 h). For
immunodetection, the membrane was blocked with 3% BSA in PBS for 1 h and
incubated for 12 h at 4°C with affinity-purified ALL1-N antiserum at aconcentration of 3 @xg/ml in PBT/l % BSA. The blot was washed three times
in PBT for 5 mm at room temperature. Biotinylated goat anti-rabbit immuno
globulin antibody (Sigma) was diluted 1:10,000 in PBT/l% BSA and incubated for 1 h at room temperature. After three more washes in PBT, it wasincubated for 45 mm at room temperature with peroxidase-streptavidin complex diluted I:2,000 in PBT/1% BSA. After three additional washes in PBT,the enhanced chemiluminescence Western detection system (Amersham, MiIan, Italy) was used. The blot was finally exposed to X-ray film for 1—10mm.As a control for the specificity of the reaction, the primary antiserum was
preincubated with a molar excess of the immunizing peptide for 30 mm at
room temperature.DNA Binding Studies Using Recombinant ALL-i Proteins in SWPD
Experiments. The following oligonucleotides were used for the DNA bindingassay: BWI (5'-AGACGGATCCATrGCA(N)18CTGTAGGAA1'TCGGAG
3'); BWAS (5'-CTCCGAATFCCTACAG-3'); and BWS (5'-AGACGGATCCATFGCA-3'). Labeling reactions were performed by annealing BWAS (240pmol) to BW1 (120 pmol) at 96°Cfor 5 mm and subsequent slow cooling toroom temperature. DNA synthesis was carried out in the presence of 10 mMTris-HCI (pH 7.5), 10 mM MgC1,, 100 ng/ml gelatin, 60 mr@isodium chloride,
1 m@iDTT, 0.6 mM dGTP and dTFP, 2 @xlof each of the radionucleotidesdATP and dCTP (3000 Ci/mmol each; Amersham Buchler), and 5 units ofKlenow polymerase I. The reaction was incubated for 20 mm at 37°C.After
chasing with 0.6 mr@idATP and dCTP, the reaction was incubated for an
additional 20 mm at 37°C. Radiolabeled ds DNA was size-separated by
electrophoresis in a 12% polyacrylamide gel, and ds oligonucleotides werelocated by exposure to X-ray film and subsequently excised from the gel. Gelslices were covered with 500 @zlof elution buffer [0.3 M NaCI and 10 mM
Tris-HC1 (pH 7.5)] overnight at 42°C,and the eluted DNA was precipitatedusing 2 volumes of ethanol before determining the specific activity by counting
in a scintillation counter (Beckmann Instruments, Munich, Germany). For
hybridization, 5,000,000 cpm (120 pmol) were diluted in 10 ml of hybridiza[ion buffer [10 mM Tris-HCI (pH 8.0), 50 mrvi sodium chloride, and 0.5 mM
zinc chloride].
SWPD Experiments. Recombinant proteins containing the AT-hook motif, the zinc binding motif PHD-ABC, or parts of the zinc binding motif
(PHD-A, PHD-AB, PHD-BC), ALL1-C, or the SET domain were expressed asrecombinant fusion proteins in Escherichia coli. The HHT-GST protein encoded by the empty pGEX-5T plasmid vector was used as a negative control.Diluted recombinant proteins (2, 1,and 0.5 @xg)were applied to a nitrocellulosemembrane (BA85; Schleicher & Schuell) using a dot blot machine. Aftertransfer, the membrane was blocked for I h in hybridization buffer [10 mMTris-HC1 (pH 8.0), 50 mM sodium chloride, and 0.5 mM zinc chloride)
containing 3% BSA. The membrane was then incubated with a mixture of
radiolabeled randomized ds oligonucleotides (500,000 cpmlml) in hybridiza
tion buffer for 16 h at 4°C.To eliminate unspecific binding of radiolabeled dsDNA to immobilized proteins, three wash steps were performed (30 mm at
4°C)with a buffer containing 10 m@iTris-HCI (pH 8.0), 100 mr@isodiumchloride, and 0.5 mM zinc chloride. Subsequently, the membrane was exposedto X-ray film for 2—3days.
RESULTS
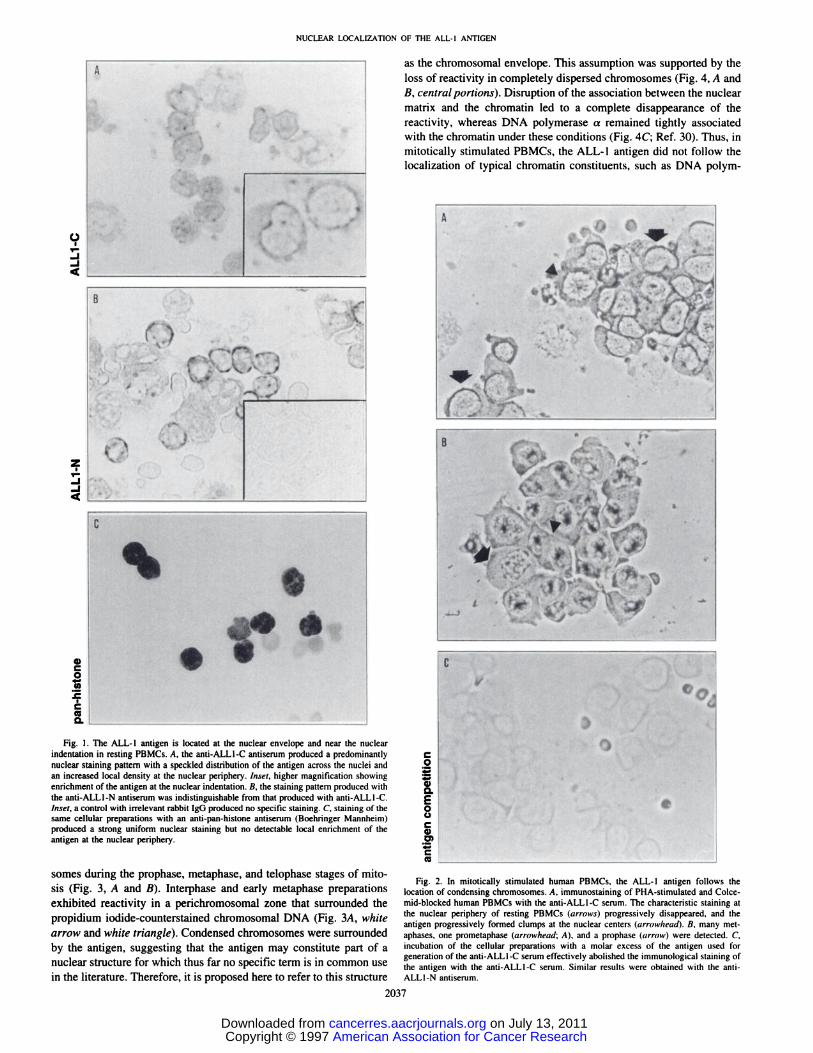
The ALL-i Antigen Is Located Predominantly in the Nucleus ofResting PBMCs. In resting human PBMCs, both the anti-ALL1-Cand anti-ALLI-N sera generated overlapping pauerns of antigen localization (Fig. 1). The antigen was predominantly located in thenuclei, with a speckled distribution across the nuclei and an increasedstaining intensity near the nuclear envelope. Particularly strong staining was consistently detected near the nuclear indentation (Fig. 1A,inset). Neither irrelevant rabbit IgG nor preimmune sera reacted withthese cellular structures (Fig. lB. inset), whereas an anti-pan-histoneantiserum generated a pattern of intense nuclear staining with uniformdistribution across the nuclei (Fig. IC). Thus, the ALL-l antigen
showed a pattern of nuclear localization that was clearly distinct fromthe distribution of histones.
In Mitogen-stimulated Human PBMCs, the ALL-i Antigen IsAssociated with the Chromosomal Envelope. The localization ofthe ALL-! antigen in mitogen-stimulated human PBMCs showedremarkable differences from the pattern observed in resting PBMCs.In PHA/Colcemid-treated cells, the anti-ALL1-C serum revealed adistribution of the antigen that was closely associated with chromatinin various stages of mitosis (Fig. 2, A and B). Early in the mitotic
cycle, the antigen was still associated predominantly with the nuclearenvelope (Fig. 2A, black arrows). In later stages of mitosis, theantigen was associated more with nuclear chromatin and less stronglywith the nuclear envelope (Fig. 2, A and B, black triangles). Thereactivity of this antiserum was completely abolished by incubatingthe cellular preparations with a molar excess of the same bacteriallyexpressed purified protein fragment that represented the core portionof the ALL-l protein and that was used as the immunogen for thegeneration of this antiserum (Fig. 2C).
To analyze the location of the ALL-! antigen with greater precisionand to address the question of whether the ALL-i antigen is solelyattached to the chromosomal envelope or possibly also attached tochromosomal DNA, preparations of mitotic PBMCs were stained byimmunofluorescent techniques (Fig. 3). Using these techniques, theantigen was visualized as a halo surrounding the condensing chromo
2036
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from
NUCLEAR LOCALIZATION OF ThE ALL-1 ANTIGEN
I asthechromosomalenvelope.Thisassumptionwassupportedbythe@ loss of reactivity in completely dispersed chromosomes (Fig. 4, A and
B, centralportions). Disruption of the association between the nuclearmatrix and the chromatin led to a complete disappearance of thereactivity, whereas DNA polymerase a remained tightly associated
with the chromatin under these conditions (Fig. 4C; Ref. 30). Thus, in@ mitotically stimulated PBMCs, the ALL-! antigen did not follow thej localizationoftypicalchromatinconstituents,suchasDNApolym
A
/
‘°-@.@
@ . @y@j
; •-@@:@@
.+.@‘I
:@@
,. @°\ -tj.
.‘@-
B@ . .@
) I.::@:,I ;@@:;c@'c::@
@ , . .@@ - _______
@ @.
B
C
S
CV
)
0.@
‘C
(Ua.
Fig. I . The ALL-l antigen is located at the nuclear envelope and near the nuclearindentation in resting PBMCs. A, the anti-ALL1-C antiserum produced a predominantlynuclear staining pattern with a speckled distribution of the antigen across the nuclei and .2an increased local density at the nuclear periphery. Inset, higher magnification showing _
(Uenrichment of the antigen at the nuclear indentation. B, the staining pattern produced withthe anti-ALLI-N antiserum was indistinguishable from that produced with anti-ALLI-C. EInset, a control with irrelevantrabbit IgG produced no specific staining. C, staining of the 0same cellular preparations with an anti-pan-histone antiserum (Boehringer Mannheim)
Cproduced a strong uniform nuclear staining but no detectable local enrichment of the@antigen at the nuclear periphery. .2'
.@ ________________________(U __________________
somes during the prophase, metaphase, and telophase stages of mitoFig. 2. In mitotically stimulated human PBMCs, the ALL-l antigen follows the
sis (Fig. 3, A and B). Interphase and early metaphase preparations locationof condensingchromosomes.A, immunostainingof PHA-stimulatedandColceexhibited reactivity in a perichromosomal zone that surrounded the mid-blockedhumanPBMCswiththe anti-ALL1-Cserum.Thecharacteristicstainingatpropidium iodide-counterstained chromosomal DNA (Fig. 3A, white the nuclearperipheryof restingPBMCs(arrows)progressivelydisappeared.and the
antigen progressively formed clumps at the nuclear centers (arrowhead). B, many metarrow and white triangle). Condensed chromosomes were surrounded aphases,one prometaphase(arrowhead;A), anda prophase(arrow)weredetected.C,by the antigen, suggesting that the antigen may constitute part of a incubationof the cellularpreparationswith a molar excessof the antigenused for
generation of the anti-ALLI-C serum effectively abolished the immunological staining ofnuclear structure for which thus far no specific term is in common use the antigenwith the anti-ALLI-Cserum.Similarresultswereobtainedwith the antiin the literature. Therefore, it is proposed here to refer to this structure ALLI-Nantiserum.
2037
S
.‘
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

.@:t@°
@ ‘@c@@°4-J.@@
.‘-. :-@ r@@@ .‘..@ -‘
...@,‘
NUCLEAR LOCALIZATION OF THE ALL. I ANTIGEN
Fig. 3. In mitotically stimulated human PBMCs, the ALL-I antigen colocalizes withthe chromosomal envelope. a nuclear subcompartment surrounding mitotic chromosomes.Mitotic chromosomes were prepared by hypotonic shock of PHA-stimulated and Colcemid-blocked human PBMCs. A, preparations from cells in prophase (arrow) and metaphase (arrowhead). B, preparations from cells in telophase. Immunofluorescent stainingin A and B with the anti-ALLI-C antiserum, counterstaining of DNA with propidiumiodide. The green-yellow fluorescence indicates the presence of the antigen in thechromosomal envelope (definition in the text). C, control with preimmune serum from thesame rabbits that were later used to produce the hyperimmune serum. Magnification,x 100.
calculated to be in the 430,000 range (Fig. 5, lefifour lanes). Irrelevast rabbit IgG did not react with this molecular species. The reactivity of the anti-ALL1-N serum with this 430-kDa molecule wascompletely abolished when the primary antibody was preincubated for30 mm with a molar excess of the same synthetic peptide that wasused as the immunogen for the generation of this antiserum (Fig. 5,
A
A , A@. “.4
‘:@I@
‘@@
:;,@ .
.@@
:.; *° - .
fi ‘@:@.. / @‘@‘:s
-!..@@@
--@@
V@
C
. .
IE
----@--@@__Fig. 4. The ALL-l antigen is detached from completely dispersed chromosomes and
does not colocalize with the chromosomal scaffold. As an accident of chromosomalpreparations, the chromosomes occasionally are detached or completely dispersed fromthe nuclear matrix. A, a preparation containing several partially disrupted nuclei withpartially dispersed chromosomes in which the nuclear matrix remained partially intact.Immunostaining with the anti-ALL1-C antiserum. B, in cells with completely detachedchromosomes, such as the one seen in the center of this field, the immunological reactivitydisappeared, indicating a loss of the antigen from the chromosomal envelope. C, bycontrast, DNA polymerase a remained associated with the chromosomes even in nuclei
with completely dispersed chromosomes. Immunological staining with an antiserumagainst DNA polymerase a, an established marker of chromatin and, in particular, of thechromosomal scaffold.
erase a, but was predominantly located in the non-chromatin compartment of the chromosomal envelope.
The ALL-i Protein Is Present as a 430-kDa Molecule in HumanPBMC Extracts. Both the anti-ALL1-N and anti-ALL1-C antiserawere used in Western blot experiments with whole-cell extracts fromresting and PHA-stimulated human PBMCs. The anti-ALL1-N serumbut not the anti-ALL1-C serum reacted with a molecular species of anelectrophoretic mobility corresponding to a Mr greater than 360,000,
2038
4@y@
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

AT PHDABC
NUCLEAR LOCALIZATION OF ThE ALL.I ANTIGEN
this pattern was generated by a DNA-binding mode that is not highlysequence-specific.
DISCUSSION
The ALL-I protein has attracted the interest of investigators for twomain reasons: (a) structural variants of this protein generated bychromosomal translocations and other alterations of the gene areprime candidates for novel oncoproteins that mediate the pathogenesisof hematological malignancies consistently associated with thesechromosomal alterations; and (b) the wild-type ALL-I protein apparently plays an important role in both normal embryogenesis andnormal hematopoiesis. Disruption of the murine ALL-I gene by homologous recombination generated mice with homeotic abnormalitiesand perturbations of hematopoiesis in the heterozygous state and ledto embryonic lethality in the homozygous state (22).
The ALL-I protein carries sequence domains that are expected toendow it with DNA-binding capacity and transcriptional transactivator potential. In fact, recombinant fragments of the protein have beenshown by other authors to have DNA-binding properties in vitro (23,31) and, in transfection experiments into mammalian cells, to havetranscriptional transactivator potential (23, 24). Therefore, it is reasonable to expect a nuclear location and a function as regulators ofgene expression for both the intact ALL-! protein and its presumablyoncogenic derivatives. This expectation was reinforced by the observation that the ALL-i protein has a high degree of sequence similaritywith the Drosophila Trithorax protein, a confirmed regulator of chromatin structure and of the maintenance of expression of homeoticgenes in Drosophila (32). However, although these observations werecompelling motivations to study the ALL-i protein and its structuralderivatives, thus far, surprisingly little was known about these proteins. The reason for this scarcity of information probably included alow abundance of these proteins in the relevant cells, the difficulty of
A B A B@ -- ++ ++Peptide
- + + + - +PH@A
rightfour lanes). A 140-kDa molecule was observed in all lanes of thegel, presumably caused by nonspecific reactivity of the second antibody.
Recombinant, Bacterially Expressed Proteins Representing theAT-Hook Protein and Zinc Finger Domains of the ALL-i ProteinReact in Vitro with ds DNA. Recombinant proteins representing theAT-hook motif, the entire zinc binding domain (PHD-ABC), or threeseparated overlapping portions ofthis domain (PHD-A, PHD-AB, andPHD-BC), the ALL1-C domain, and the SET domain were separately
expressed in E. coli and chromatographically purified from bacterialextracts. The HHT-GST protein encoded by the empty pGEX-5Tplasmid vector was used as negative control (see “MaterialandMethods―). All recombinant proteins were applied to nitrocellulosemembranes in a grid pattern, and the membrane was then incubatedwith 120 pmol of a degenerate mixture of radiolabeled ds syntheticoligonucleotides. These oligonucleotides were 50 bp in length, with acore of 18 nucleotides of random sequence flanked by 16 nucleotidesof specific sequences on either side to facilitate cloning and detectionby PCR (primer binding sites, restriction enzyme cleavage sites; see“Materialand Methods―). The mixture contained approximately7 X 1013ds DNA molecules with approximately 7 X lO'°differentmolecules, each present in more than iO@copies (see “MaterialandMethods―for details). The recombinant protein containing the AThook domain displayed a strong DNA-binding activity, and weakerDNA binding was also observed for the PHD-A, PHD-AB, andPHD-BC proteins, whereas the GST control, ALL1-C, and the com
plete PHD-ABC and SET proteins loaded in comparable molar quanrifles to the filter showed no detectable DNA binding (Fig. 6). Thus,the recombinant AT-hook domain showed detectable DNA bindingeven to an unselected mixture of ds DNA molecules, suggesting that
kO.
4304
22O@@
144@@
Fig. 5. The anti-ALLI-N antiserum detects a protein in whole-cell extracts from bothresting and mitotically stimulated human PBMCs with a size in excess of 360 kDa andconsistent with a size of 430 Wa. A, the anti-ALLI-N antiserum detected a protein of amobility in excess of 360 kDa (the size marker used in this gel was fatty acid synthetaseisolated from green algae with a M1 of 360,000 and mouse ALL-l protein with acalculatedMrof454,000)andcompatiblewitha sizeof430 Wa, thepredictedsizeof thefull-lengthALL-i protein from cDNA in whole-cellextractsfrom resting (—)andPHA-stimulated (+) PBMCs (lefifour lanes). Western blot. B, an independent preparation.Thereactivitydisappearedwhentheblotwaspreincubatedwitha molarexcessofthesame peptide that was used as the antigen to produce the anti-ALL1-N serum (right fourlanes).
@:.@ 1j@tg
O.2p.g
ALLI-C SETjCOOH
4005Fig. 6. A recombinant protein containing the AT-hook domain of the ALL-I protein
binds to degenerate mixture of ds synthetic oligonucleotides. SWPD experiment. Increasing amounts of recombinant proteins (0.2, 1. and 2 @zgdeposited per dot; three verticalcolumns), representing different functional domains of the human ALL-I protein, wereproduced in E. coli, chromatographically purified, and deposited onto nitrocellulose in theform of dots. The proteins contained the following domains: GST, GST fragment, noALL-I sequences, negative control; AT-hook, AT-hook domain; SE!', SET domain;ALLI-C, core domain; PHD-ABC, entire zinc binding domain, consisting of three domains(A, B, and C), each resembling the zinc binding domains of known zinc finger proteins.Bottom, the approximate locations of these domains in the ALL-I protein are shownschematically. The filters were incubated with a radiolabeled mixture of degenerate dsoligonucleotides with an 18-bp random-core sequence. After the reaction, the filter waswashed and exposed for autoradiography.
2039
0.@ 0m mu
0 •
U)@w@j 11110 <U) 4 a. a. a. a.
. 2j.tg
NH2II I1
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

NUCLEAR LOCALIZATION OF THE ALL-I ANTIGEN
ALL-l protein, it would be premature to speculate about thefunction of this newly recognized nuclear compartment. That thiscompartment was not identical with chromatin was established bythe different staining patterns generated with anti-ALLi and antiDNA polymerase a antisera, in which DNA polymerase a is aconfirmed marker for chromatin, which is closely associated withthe chromosomal scaffold (30).
Although binding of recombinant fragments of the ALL-l proteincontaining the AT-hook domain to DNA had previously been reportedby other authors (23), the result presented here represents a stepforward in our understanding of this DNA-binding property. Theseother authors had reported that the AT-hook domain exclusively bindsto DNA containing sequences capable of forming cruciform structures. When the same sequences were contained in an arrangementthat was capable of forming only linear DNA double strands, nobinding was reported (23). Due to the strong binding to a degeneratemixture of ds oligonucleotides observed here (Fig. 6), we concludedthat this binding mode probably lacked the high degree of sequencespecificity generally observed for proteins, which bind to the majorgroove. By contrast, AT-hook proteins and others that bind DNA withlittle sequence specificity generally attach to the minor groove ofAT-rich DNA. Thus, the new information presented here is that theAT-hook domain is also capable of binding to ds DNA that does notform cruciform structures, because our sequences were random andnot specifically designed to build cruciform structures for which an18-bp sequence would be too short.
Interestingly, the recombinant proteins representing parts of theso-called zinc binding domain (a domain with sequence homology tozinc finger proteins, as predicted from the cDNA sequence) of ALL-Ialso showed evidence of DNA binding in this assay (Fig. 6). However,the binding was much weaker than that observed with AT-hookcontaining recombinant fragments. The exact explanation of thisfinding is unknown. Possible explanations include, among others,both a DNA-binding mode with a higher degree of sequence specificity, as expected from the known binding specificity of other zincfinger proteins, and the presence in this experiment of different molaramounts of the actively DNA-binding species on the filter. To distinguish between these possibilities and other possible explanations willrequire future isolation of individual DNA-binding target sequencesfor both the AT-hook and zinc binding domains and analysis of theiraffinities and specificities. Preliminary data4 showed that the zincbinding domain of the ALL-! protein indeed possesses preferentialbinding properties to specific DNA target sequences. It will also beimportant in the future to isolate genes containing such binding sitesas potential target genes for the regulation of their expression byALL-] gene-derived polypeptides.
The current picture does not yet allow us to draw conclusions aboutthe function of the normal ALL-i protein and its presumably oncogenic derivatives, but the availability of the antisera presented hereand the discovery of new DNA-binding abilities will allow futureinvestigators to make progress in providing at least partial answers tothese important questions.
ACKNOWLEDGMENTS
We thank Dr. E. Geyer (University of Marburg) for the immunization ofrabbits and the production of the anti-ALL1-C antisera. We also thank Dr. J.
Hofmann (University of Erlangen-NUrnberg) for providing the purified fatty
acid synthetase protein from green algae, used as a molecular weight marker
for the Western blot experiments.
REFERENCES
1. Chen, C. S., Hilden, J. M., Frestedt, J., Domer, P. H., Moore, R., Korsmeyer, S. J., andKersey, J. H. The chromosome 4q21 gene (AF-4/FEL) is widely expressed in normal
their detection due to their large sizes (above 200 kDa for the derivatives, above 400 kDa for the intact protein), and the nonavailabilityof antisera of sufficient quality.
The main new results presented in this study are: (a) the generationof two new antisera of high quality and sufficiently high titer to beuseful for both immunolocalization and Western blot studies; (b) theobservation that the ALL-l antigen was indeed predominantly locatedin the nucleus of human leukocytes, as anticipated from the knownprotein sequence and its DNA-binding domains; and (c) the ability ofpartial domains of this protein to bind ds DNA even if this was notarranged in potentially cruciform sequences.
Antisera directed against various recombinant cDNA-derivedALL-i protein fragments had previously been produced by severallaboratories including ours. However, in our hands, these sera hadfailed to detect the intact ALL-l protein in Western blot experimentswith extracts from normal human leukocytes and failed to detect thepresumed oncogenic derivatives in extracts from leukemia-derivedcell lines with translocations to the ALL-] gene.4 The main progressreported here is that a polyclonal rabbit antipeptide antiserum directedagainst amino acids 446—456of the ALL-I protein sequence (13, 14)detected a protein in Western blot experiments with extracts fromnormal human PBMCs that corresponded in size to the expected430-kDa intact protein. This presumed full-length protein was at leastsufficiently stable in these cells to be detected as the major species inthis experiment. It is interesting that the antiserum directed against apurified recombinant protein representing the ALL-! core (antiALL! -C) did not detect this high molecular weight species in theWestern blots, although it generated an indistinguishable pattern ofantigenic reactivity in immunocytochemical experiments as the antiALL1-N antiserum (Fig. 1). The reason for this observation is unknown but could simply reflect insufficient affinity of this antiserum.
Other interesting findings reported here for the first time include:(a) the nuclear localization of the wild-type ALL-i antigen in humanPBMCs; and (b) the difference in nuclear staining observed betweenresting and mitotically stimulated PBMCs, with a localization predominantly near the nuclear envelope in resting cells and a localization predominantly associated with the chromosomal envelope inmitotically stimulated cells.
The molecular basis for this difference in distribution is unknown.It could be explained by a redistribution of the antigen between thesetwo compartments induced by the mitotic stimulation or by thegeneration of a different set of antigenic products, such as a differentset of proteolytic products, with different nuclear sublocalizationproperties. To distinguish between these and other possible explanations will require the biochemical extraction of the antigen fromnuclei in different stages of proliferation and its molecular characterization by protein chemical methods.
The association of the antigen with the nuclear structure termedthe chomosomal envelope in mitotically stimulated PBMCs is ofparticular interest, because this structure had not previously beenrecognized as a distinct entity and thus had not been given aspecific name. If this structure had been a well-known nuclearcompartment, then it would have been mandatory to search for anantibody directed against a known marker for this compartmentand to demonstrate colocalization of the ALL-i antigen with thismarker. However, no such markers were available; therefore, theALL-i antigen as recognized by the anti-ALL1-N and antiALL!-C sera becomes itself the prototypical marker for this novelsubcompartment. Given the absence of other known components ofthis structure and the scarcity of functional knowledge about the
4 R. Greim, unpublished observations.
2040
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from

NUCLEAR LOCALIZATION OF THE ALL-I ANTIGEN
tissues and shows breakpoint diversity in t(4;1 l)(q2l;q23) acute leukemia. Blood, 82:1080—1085,1993.
2. Kaneko, Y., Maseki, N., Takasaki, N., Sakurai, M., Hayashi, Y., Nakazawa, S., Mori,T., Sakurai, M., Takeda, T., Shikano, 1., and Hiyoshi, Y. Clinical and hematologiccharacteristics in acute leukemia with 11q23 translocations. Blood, 67: 484—491,1986.
3. Katz, F., Malcolm, S., Gibbons, B., Tilly, R., Lam, G., Robertson, M. E.,Czepulkowski, B., and Chessels, J. Cellular and molecular studies of infant null acutelymphoblastic leukemia. Blood, 71: 1438—1447, 1988.
4. Pui, C. H., Raimondi, S. C., Murphy, S. B., Ribeiro, R. C., Kalwinsky, D. K., Dahl,G. V., Crist, W. M., and Williams, D. L. An analysis of leukemic cell chromosomalfeatures in infants. Blood, 69: 1289—1293, 1987.
5. Pui, C. H., Crist, W. M., and Look, A. T. Biology and clinical significance ofcytogenetic abnormalities in childhood acute lymphoblastic leukemia. Blood, 76:1449—1463,1990.
6. Pui, C. H., Frankel, L. S., Carroll, A. J., Raimondi, S. C., Shuster, J. i., Head, D. R.,Crist, W. M., Land, V. J., Pullen, D. J., Steuber, C. P., Behm, F. G., and Borowitz,M.J. Clinicalcharacteristicsandtreatmentoutcomeofchildhoodacutelymphoblasticleukemia with t(4;l l)(q21;q23): a collaborative study of 40 cases. Blood, 77: 440—447, 1991.
7. Bernard, 0. A., and Berger, R. Molecular basis of I 1q23 rearrangements in hematopoietic malignant proliferations. Genes Chromosomes & Cancer, 13: 75—85,1995.
8. Schichman, S. A., Caligiuri, M. A., Strout, M. P., Carter, S. L., Gu, Y., Canaani, E.,Bloomfield, C. D., and Croce, C. M. ALL-l tandem duplication in acute myeloidleukemia with a normal karyotype involves homologous recombination between Aluelements. Cancer Res., 54: 4277—4280, 1994.
9. Schichman, S. A., Caligiuri, M. A., Gu, Y., Strout, M. P., Canaani, E., Bloomfield,C. D., and Croce,C. M. ALL-l partialduplicationin acuteleukemia.Proc.Natl.Aced. Sci. USA, 9!: 6236—6239,1994.
10. Caligiuri, M. A., Strom, M. P., Schichman, S. A., MrOzek, K., Arthur, D. C., Herzig,G.P.,Baer,M.R.,Schiffer,C.A.,Heinonen,K.,Knuutila,S.,Nousiainen,T.,Ruutu,T., Block, A. W., Schulman, P., Pedersen-Bjergaard, J., Croce, C. M., andBloomfield, C. D. Partial tandem duplication of ALL! as recurrent molecular defectin acute myeloid leukemia with trisomy II. Cancer Res., 56: 1418—1425, 1996.
11. Lochner, K., Siegler, G., Führer,M., Greil, J., Beck, J. D., Fey, G. H., andMarschalek,R.Aspecificdeletionin thebreakpointclusterregionof theAU-i geneis associated with acute lymphoblastic T-cell leukemias. Cancer Res., 56: 2 171—2177,1996.
12. Ziemin van der Poel, S., McCabe, N. R., Gill, H. J., Espinosa, R., III, Patel, Y.,Harden, A., Rubinelli, P., Smith, S. D., LeBeau, M. M., Rowley, J. D., and Diaz,M.0. Identificationofa gene,MU, thatspansthebreakpointin 11q23translocationsassociated with human leukemias. Proc. Ned. Acad. Sci. USA, 88: 10735—10739,1991.
13. Gu, Y., Nakamura, 1., Alder, H., Prasad, R., Canaani, 0., Cimino, G., Croce, C. M.,and Canaani, E. The t(4;l 1) chromosome translocation of human acute leukemiasfuses the AU-i gene, related to Drosophila trithorax, to the AF-4 gene. Cell, 7i:701—708,1992.
14. Tkachuk, D. C., Kohler, S., and Cleary, M. L. Involvement of a homolog ofDrosophila trithorax by I 1q23chromosomal translocationsin acute leukemias. Cell,71: 691—700,1992.
15. Rasio, D., Schichman, S. A., Negrini, M., Canaani, E., and Croce, C. M. Completestructure of the ALLI gene. Cancer Res., 56: 1766—1769, 1996.
16. Nilson, I., LOchner, K., Siegler, G., Greil, J., Beck, J. D., Fey, 0. H., and Marschalek,R. Exon/intron structure of the human AU-I (MU) gene involved in translocations
to chromosomal region 11q23 and acute leukemias. Br. J. Haematol., 93: 966—972,1996.
17. Marschalek, R., Nilson, I., Ldchner, K., Greim, R., Siegler, G., Greil, J., Beck, J. D.,and Fey, G. H. The structure of the human AU-i/MLUHRX gene. Leuk. Lymphoma.in press, 1997.
18. Mazo, A. M., Huang, D. H., Mozer, B. A., and Dawid, I. B. The trithorax gene, atrans-acting regulator of the bithorax complex in Drosophila. encodes a protein withzinc-binding domains. Proc. NatI. Acad. Sci. USA, 87: 21 12—2116, 1990.
19. Mozer, B. A., and Dawid, I. B. Cloning and molecular characterization of thetrithorax locus of Drosophila melangaster. Proc. Natl. Acad. Sci. USA, 86: 3738—3742, 1989.
20. Sedkov, Y., Tillib, S., Mizrokhi, L., and Mazo, A. The bithorax complex is regulatedby trithorax earlier during Drosophila embryogenesis than is the Antennapediacomplex, correlating with a bithorax-like expression pattern of distinct early trithoraxtranscripts. Development (Carob.), i20: 1907—1917,1994.
21 . Fidanza, V., Melotti, P., Yano, T., Nakamura, T., Bradley, A., Canaani, E., Calabretta,B., and Croce, C. M. Double knockout of the AU-i gene blocks hematopoieticdifferentiation in vitro. Cancer Res., 56: 1179—1183, 1996.
22. Yu, B. D., Hess, J. L., Horning, S. E., Brown, G. A. J., and Korsmeyer, S. Altered Hoxexpression and segmental identity in MLL-mutant mice. Nature (Lond.), 378: 505—508, 1995.
23. Zeleznik-Le, N. J., Harden, A. M., and Rowley, J. D. I 1q23 translocations split the“AT-hook―cruciform DNA-binding region and the transcriptional repression domainfrom the activation domain of the mixed- lineage leukemia (MU) gene. Proc. Natl.Acad. Sci. USA, 9i: 10610—10614,1994.
24. Prasad, R.. Yano, T., Sorio, C., Nakamura, T., Rallapalli, R., Gu, Y., Leshkowitz, D.,Croce, C. M., and Canaani, E. Domains with transcriptional regulatory activity withinthe ALL1 and AF4 proteins involved in acute leukemia. Proc. Natl. Acad. Sci. USA,92: 12160—12164,1995.
25. Reeves, R., and Nissen, M. S. The kT-DNA-binding domain of mammalian HighMobility Group I chromosomal proteins. J. BioI. Chem., 265: 8573—8582, 1990.
26. Ma, Q., Alder, H., Nelson, K. K., Chatteijee, D., Gu, Y., Nakamura. T., Canaani, E.,Croce, C. M., Siracusa, L. D., and Buchberg, A. M. Analysis of the murine All-I genereveals conserved domains with human ALL- I and identifies a motif shared withDNA methyltransferases. Proc. NatI. Acad. Sci. USA, 90: 6350—6354, 1993.
27. Marschalek, R., Greil, J., Lochner, K., Nilson, I., Siegler, G., Zweckbronner, I., Beck,J. D., and Fey, 0. H. Molecular analysis of the chromosomal breakpoint and fusiontranscripts in the acute lymphoblastic SEM cell line with chromosomal translocationt(4:1 I). Br. J. Haematol., 90: 308—320,1995.
28. Aasland, R., Gibson, T. J., and Stewart, A. F. The PHD finger: implications forchromatin-mediated transcriptional regulation. Trends Biol. Sci., 20: 56—59,1995.
29. Harlow, E., and Lane, D. Antibodies: A Laboratory Manual, pp. 283—318.ColdSpring Harbor, NY: Cold Spring Harbor Laboratory. 1988.
30. Ennas, M. 0., Suzuki, H., Menegazzi, M., Carcereri, A., Hanoaka, F., Gremo, F.,Nieddu, M., and Mezzanotte, R. On the presence of DNA-polymerase a in humanlymphocyte nuclei and chromosomes. Heredity, 77: 186—191,1996.
31. Strissel-Broeker, P. L., Gill-Super, H., Thirman, M. J., Pomykala, H., Yonebayashi,Y., Tanabe, S., Zeleznik-Le, N., and Rowley, J. D. Distribution of I 1q23 breakpointswithin the MLL breakpoint cluster region in de novo acute leukemia and in treatmentrelated acute myeloid leukemia: correlation with scaffold attachment regions andtopoisomerase II consensus binding sites. Blood, 87: 1912—1922,1996.
32. Kennison, J. A. Transcriptional activation of Drosophila homeotic genes from distantregulatory elements. Trends Genet., 9: 75—79,1993.
2041
American Association for Cancer Research Copyright © 1997 on July 13, 2011cancerres.aacrjournals.orgDownloaded from




















