
Mitochondria as targets for detection and treatment of cancer

Research ArticleOn the Mechanism(s) of MembranePermeability Transition in Liver Mitochondria of Lamprey,Lampetra fluviatilis L.: Insights from Cadmium
Elena A. Belyaeva, Larisa V. Emelyanova, Sergey M. Korotkov,Irina V. Brailovskaya, and Margarita V. Savina
I.M. Sechenov Institute of Evolutionary Physiology and Biochemistry, Russian Academy of Sciences, Thorez pr. 44,Saint Petersburg 194223, Russia
Correspondence should be addressed to Elena A. Belyaeva; [email protected]
Received 28 February 2014; Revised 29 April 2014; Accepted 1 May 2014; Published 3 June 2014
Academic Editor: Hartmut Jaeschke
Copyright © 2014 Elena A. Belyaeva et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Previously we have shown that opening of the mitochondrial permeability transition pore in its low conductance state is the casein hepatocytes of the Baltic lamprey (Lampetra fluviatilis L.) during reversible metabolic depression taking place in the period ofits prespawning migration when the exogenous feeding is switched off. The depression is observed in the last year of the lampreylife cycle and is conditioned by reversible mitochondrial dysfunction (mitochondrial uncoupling in winter and coupling in spring).To further elucidate the mechanism(s) of induction of the mitochondrial permeability transition pore in the lamprey liver, weused Cd2+ and Ca2+ plus Pi as the pore inducers. We found that Ca2+ plus Pi induced the high-amplitude swelling of the isolated“winter” mitochondria both in isotonic sucrose and ammonium nitrate mediumwhile both low and high Cd2+ did not produce themitochondrial swelling in these media. Low Cd2+ enhanced the inhibition of basal respiration rate of the “winter” mitochondriaenergized by NAD-dependent substrates whereas the same concentrations of the heavy metal evoked its partial stimulation onFAD-dependent substrates. The above changes produced by Cd2+ or Ca2+ plus Pi in the “winter” mitochondria were only weakly(if so) sensitive to cyclosporine A (a potent pharmacological desensitizer of the nonselective pore) added alone and they were notsensitive to dithiothreitol (a dithiol reducing agent). Under monitoring of the transmembrane potential of the “spring” lampreyliver mitochondria, we revealed that Cd2+ produced its decrease on both types of the respiratory substrates used that was stronglyhampered by cyclosporineA, and themembrane potential was partially restored by dithiothreitol.The effects of differentmembranepermeability modulators on the lamprey liver mitochondria function and the seasonal changes in their action are discussed.
1. Introduction
In autumn lamprey Lampetra fluviatilis L.migrates from theGulf of Finland to the Neva River (Northwestern Russia) andswitches off the exogenous feeding during all the period ofits prespawning migration. After the years of investigations,it became clear that a pronounced and reversible metabolicdepression (for comprehensive reviews, see [1–7]) occurs inliver of the Baltic lamprey during the winter prespawningperiodwhich is characterized by substantial decrease of tissueand cellular endogenous respiration, an oxidative phosphory-lation reduction, and both ATP content and mitochondrialmembrane potential drop [8–13]. Besides, one of the most
important peculiarities of hepatocyte energy metabolism ofthe “winter” lamprey is the absence of significant contributionof glycolysis in ATP production as well as its regulation viathe availability of fatty acids for oxidation, which are foundto be the main respiratory substrates [9–12, 14]. In spring theenergymetabolism activation is observed in the lamprey liverfollowing the appearance of the secondary sex characters [15–17].The sharp activation of oxidation and phosphorylation inthe lamprey liver mitochondria is followed by spawning anddeath of the animal in the end of spring—in the beginning ofsummer. As suggested, a strict and genetically programmedhormonal regulation of seasonal variations in metabolicrate to preserve energy resources for spawning takes place.
Hindawi Publishing CorporationBioMed Research InternationalVolume 2014, Article ID 691724, 14 pageshttp://dx.doi.org/10.1155/2014/691724

2 BioMed Research International
However,molecularmechanism(s) of the phenomenon is stillelusive.
Previously using isolated mitochondria as a model, wehave revealed clear-cut seasonal variations of themain bioen-ergetics parameters of the lamprey liver [12, 14]. The foundchanges indicate that the metabolic depression observedduring the last winter of the lamprey’s life cycle is medi-ated by prolonged reversible mitochondrial dysfunction.Thedysfunction is shown to manifest itself in the following:(1) the very low activity of mitochondrial respiratory chain,especially of its complex I, (2) low oxidative phospho-rylation, (3) decreased content of mitochondrial adeninenucleotides, (4) high level of reduced pyridine nucleotides,and (5) leaky mitochondrial membranes [12, 14]. In par-ticular, it has been found that the “winter” lamprey livermitochondria (LLM) passively swell in an isotonic NH
4NO3
solution that indicates the increased proton permeability ofthe inner membranes of these mitochondria. Besides, thenonenergized LLM have been found to be permeable toK+ and Cl− as well [12, 14, 18]. The obtained data havebrought us to the assumption that the enhanced membranepermeability demonstrated by the LLM in respect to protonsand monovalent cations, but not to sucrose molecules, maybe due to the opening of the mitochondrial permeabilitytransition (MPT) pore [19–25] in its low-conductance state(this type of the pore conformation, as well-known, allowssmall solutes, like H+, K+, Ca2+, and so forth, to diffusein and out of the mitochondrial matrix). Moreover, on theisolated mitochondria we have demonstrated the closing ofthe pore in the presence of its classical inhibitors, such asMg2+, ADP (in the presence and absence of cyclosporine A,CsA), and [ethylenebis(oxyethylenenitrilo)]tetraacetic acid(EGTA), as well as after energization of the LLM [12, 14].Since we observed the whole range of energetic states of themitochondria (from uncoupling in winter till coupling inspring) during the prespawning migration of the lamprey, wehave supposed that in vivo the MPT pore of the animal isstrictly regulated [12–14, 18, 26–29].
It should be mentioned also that in our early work [29]we have got the evidence that the LLM function is sensitiveto toxic action of such heavy metal, as cadmium (Cd2+).Before on isolated rat liver mitochondria (RLM), that is, onthe conventional model to study the MPT pore involvement,we conducted a thorough investigation on mechanism(s) ofCd2+ toxicity and found that Cd2+ induced RLM dysfunctionmediated by the disturbance of mitochondrial respiratorychain (mtETC) and the opening of the nonselective poreboth in its low- and high-conductance states [30–37]. In thepresent study a one more attempt to puzzle out the mysteryof life and decease of the lamprey was made. In particular,with the aim to better understand the mechanism(s) of theMPT pore induction in the lamprey liver, we used Cd2+ andfor comparison, Ca2+ plus Pi (in some cases also a few fattyacids) as the pore inducers and the isolated mitochondriaof both “winter” and “spring” animals. The seasonal changesin action of different permeability transition modulators onthe lamprey liver mitochondria function were revealed and
discussed. Portion of this study was presented previously inan abstract form [38].
2. Materials and Methods
2.1. Chemicals. ADP, CsA, EGTA, dithiothreitol (DTT),bovine serum albumin (BSA), nupercaine (dibucaine),carbonyl cyanide p-trifluoromethoxyphenylhydrazone(FCCP), carbonyl cyanide 3-chlorophenylhydrazone(CCCP), rotenone, oligomycin, valinomycin (Val),succinate, glutamate, pyruvate, laurate, acetate, ascorbate,tetramethyl-p-phenylenediamine (TMPD), Tris-OH,safranine, ruthenium red (RR), CdCl
2, CaCl
2, MgCl
2,
H3PO4(Pi), and so forth were purchased from Sigma (USA)
or Serva (Germany); sucrose was refined with the help ofa cationite KU-2-8 column; KCl was recrystallized twice;heparin, 2, 4-dinitrophenol (DNP), KSCN, KNO
3, NH4NO3
were of analytical grade. 137Cs+ was purchased from Isotop(Russia).
2.2. Isolation ofMitochondria. Lampreys (Lampetra fluviatilisL.) of 50–70 g were caught in estuary of the Neva Riverin October-November during their prespawning migrationfrom the Baltic Sea and maintained until May in holdingtanks with cooled (2–6∘C) and permanently aerated water.Eight to ten adult animals, if other not indicated, weretaken for each experiment.Themitochondria from liver wereisolated by the method of differential centrifugation in themedium of the following composition (mM): 250 sucrose,3 Tris-HCl (pH 7.3), and 0.5 EGTA. Mitochondrial pelletwas washed twice with the medium containing 250mMsucrose, 3mM Tris-HCl (pH 7.3). At the first washing, BSA(1mg/mL of medium) was introduced into the medium forremoving free fatty acids. Suspending of the mitochondriawas conducted usually in the medium without BSA. Theprotein concentration was determined by Bradford or Biuretmethods with BSA as standard as before [12, 14]. The mito-chondrial suspension contained 25–30mg of protein/mL.In some cases (see explanations in the text) heparin (25or 50 units/mL) and different concentrations of nupercaine(Nup) were added in an isolation and/or assay medium.
2.3. Monitoring of Cs+ (Analog of K+) Uptake by IsotopeTechniques. 137Cs+ accumulation by isolated LLMwas deter-mined by the Millipore filtration method, the details ofwhich were described earlier [31, 39]. In particular, samplesof 0.1mL were taken at intervals and mixed with ice-coldmedium free of the isotope, and the mitochondria wereimmediately filtered through filters of 0.45mM pore size,washed once with 2mL ice-cold medium, and assayed fortheir activity, with correction of filter backgrounds obtainedby filtration of mitochondria-free incubation medium andwashing. The uptake experiment results were expressed asthe distribution ratio of the tracer (𝑟
𝑡) between matrix
and medium. As mentioned elsewhere [31, 39], the kineticsof the process of uptake of 137Cs+ by valinomycin-treatedand substrate-energized mitochondria is well characterizedby coefficients of distribution (𝑟
𝑡) of the tracer between
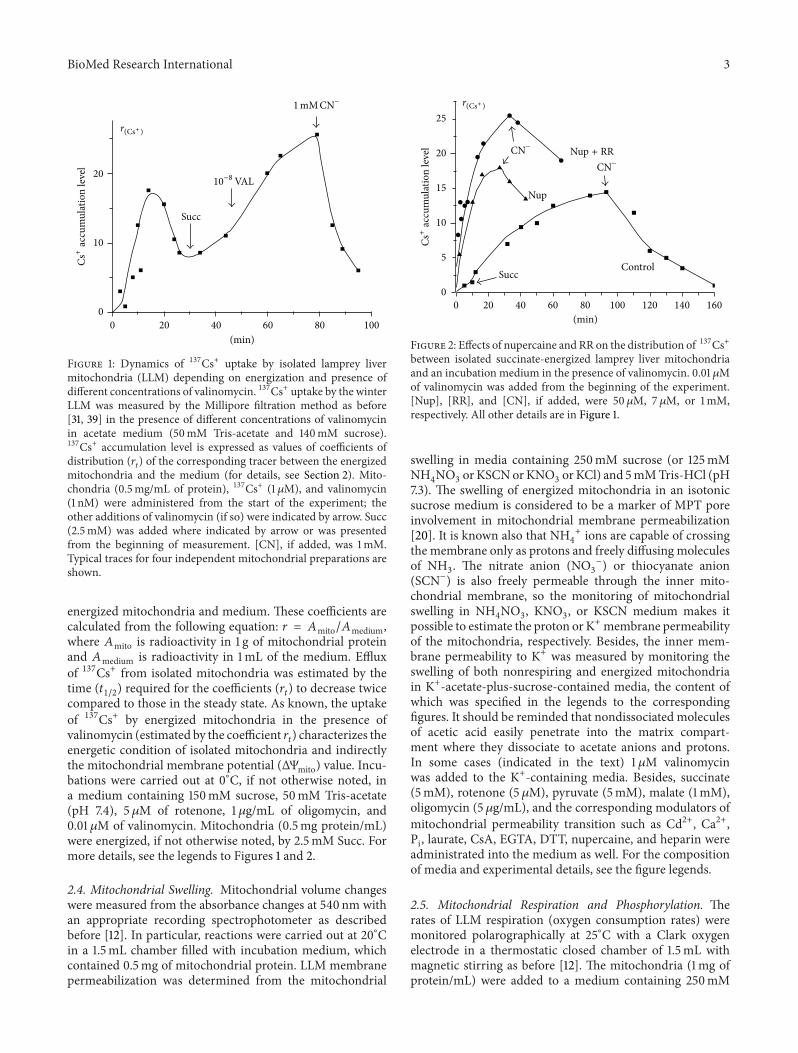
BioMed Research International 3Cs
+ac
cum
ulat
ion
leve
l
20
20
10
0
0 40 60 80 100
(min)
r(Cs+)
Succ
10−8 VAL
1mM CN−
Figure 1: Dynamics of 137Cs+ uptake by isolated lamprey livermitochondria (LLM) depending on energization and presence ofdifferent concentrations of valinomycin. 137Cs+ uptake by the winterLLM was measured by the Millipore filtration method as before[31, 39] in the presence of different concentrations of valinomycinin acetate medium (50mM Tris-acetate and 140mM sucrose).137Cs+ accumulation level is expressed as values of coefficients ofdistribution (𝑟
𝑡) of the corresponding tracer between the energized
mitochondria and the medium (for details, see Section 2). Mito-chondria (0.5mg/mL of protein), 137Cs+ (1 𝜇M), and valinomycin(1 nM) were administered from the start of the experiment; theother additions of valinomycin (if so) were indicated by arrow. Succ(2.5mM) was added where indicated by arrow or was presentedfrom the beginning of measurement. [CN], if added, was 1mM.Typical traces for four independent mitochondrial preparations areshown.
energized mitochondria and medium. These coefficients arecalculated from the following equation: 𝑟 = 𝐴mito/𝐴medium,where 𝐴mito is radioactivity in 1 g of mitochondrial proteinand 𝐴medium is radioactivity in 1mL of the medium. Effluxof 137Cs+ from isolated mitochondria was estimated by thetime (𝑡
1/2) required for the coefficients (𝑟
𝑡) to decrease twice
compared to those in the steady state. As known, the uptakeof 137Cs+ by energized mitochondria in the presence ofvalinomycin (estimated by the coefficient 𝑟
𝑡) characterizes the
energetic condition of isolated mitochondria and indirectlythe mitochondrial membrane potential (ΔΨmito) value. Incu-bations were carried out at 0∘C, if not otherwise noted, ina medium containing 150mM sucrose, 50mM Tris-acetate(pH 7.4), 5 𝜇M of rotenone, 1 𝜇g/mL of oligomycin, and0.01 𝜇M of valinomycin. Mitochondria (0.5mg protein/mL)were energized, if not otherwise noted, by 2.5mM Succ. Formore details, see the legends to Figures 1 and 2.
2.4. Mitochondrial Swelling. Mitochondrial volume changeswere measured from the absorbance changes at 540 nm withan appropriate recording spectrophotometer as describedbefore [12]. In particular, reactions were carried out at 20∘Cin a 1.5mL chamber filled with incubation medium, whichcontained 0.5mg of mitochondrial protein. LLM membranepermeabilization was determined from the mitochondrial
25
20
15
10
5
0
0 20 40 60 80 100 120 140 160
Nup
Control
Cs+
accu
mul
atio
n le
vel
(min)
r(Cs+)
Succ
CN−
CN−
Nup + RR
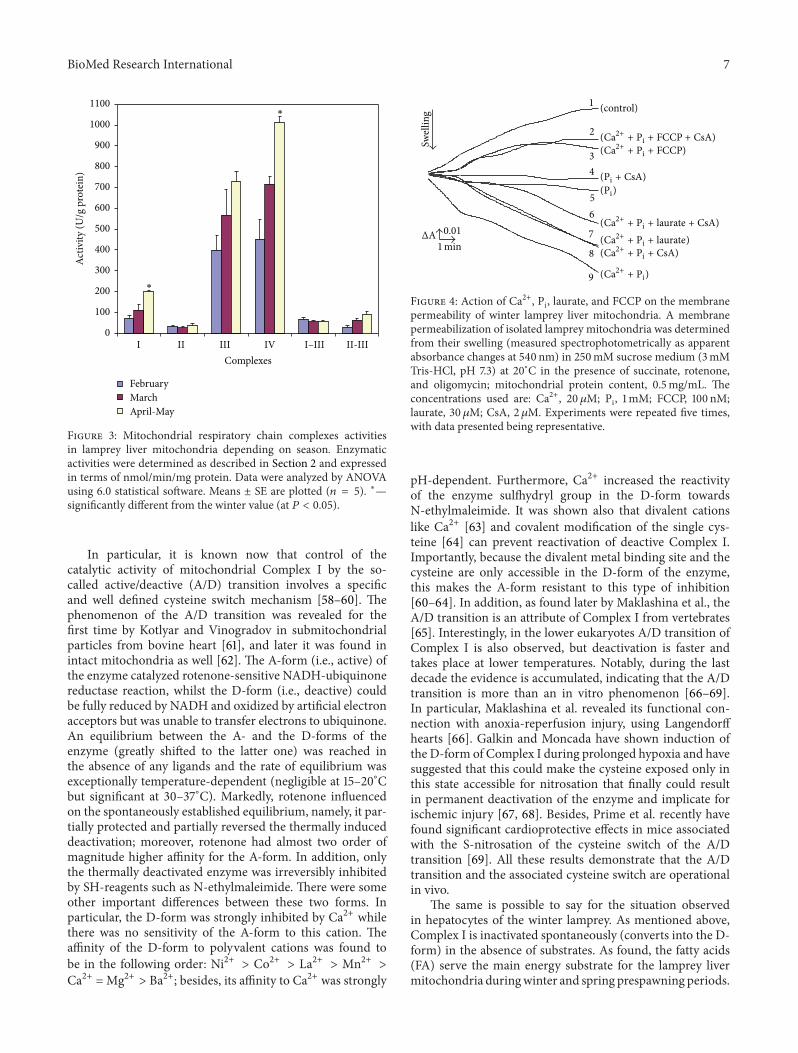
Figure 2: Effects of nupercaine and RR on the distribution of 137Cs+between isolated succinate-energized lamprey liver mitochondriaand an incubation medium in the presence of valinomycin. 0.01 𝜇Mof valinomycin was added from the beginning of the experiment.[Nup], [RR], and [CN], if added, were 50 𝜇M, 7 𝜇M, or 1mM,respectively. All other details are in Figure 1.
swelling in media containing 250mM sucrose (or 125mMNH4NO3orKSCNorKNO
3orKCl) and 5mMTris-HCl (pH
7.3). The swelling of energized mitochondria in an isotonicsucrose medium is considered to be a marker of MPT poreinvolvement in mitochondrial membrane permeabilization[20]. It is known also that NH
4
+ ions are capable of crossingthe membrane only as protons and freely diffusing moleculesof NH
3. The nitrate anion (NO
3
−) or thiocyanate anion(SCN−) is also freely permeable through the inner mito-chondrial membrane, so the monitoring of mitochondrialswelling in NH
4NO3, KNO
3, or KSCN medium makes it
possible to estimate the proton or K+membrane permeabilityof the mitochondria, respectively. Besides, the inner mem-brane permeability to K+ was measured by monitoring theswelling of both nonrespiring and energized mitochondriain K+-acetate-plus-sucrose-contained media, the content ofwhich was specified in the legends to the correspondingfigures. It should be reminded that nondissociated moleculesof acetic acid easily penetrate into the matrix compart-ment where they dissociate to acetate anions and protons.In some cases (indicated in the text) 1 𝜇M valinomycinwas added to the K+-containing media. Besides, succinate(5mM), rotenone (5 𝜇M), pyruvate (5mM), malate (1mM),oligomycin (5𝜇g/mL), and the corresponding modulators ofmitochondrial permeability transition such as Cd2+, Ca2+,Pi, laurate, CsA, EGTA, DTT, nupercaine, and heparin wereadministrated into the medium as well. For the compositionof media and experimental details, see the figure legends.
2.5. Mitochondrial Respiration and Phosphorylation. Therates of LLM respiration (oxygen consumption rates) weremonitored polarographically at 25∘C with a Clark oxygenelectrode in a thermostatic closed chamber of 1.5mL withmagnetic stirring as before [12]. The mitochondria (1mg ofprotein/mL) were added to a medium containing 250mM

4 BioMed Research International
sucrose, 3mM MgCl2, 3mM KH
2PO4, and 3mM Tris-HCl
(pH 7.3). In some cases the respiratory buffer contained100mM sucrose, 40mM KCl, 3mMMgCl
2, 3mM KH
2PO4,
and 20mM Tris-HCl, pH 7.3 (±50𝜇M of EGTA and/or1mg/mL BSA). Besides, 5mM pyruvate and 1mM malateor 5mM succinate (±5 𝜇M of rotenone) were added to theincubation medium for energization of the mitochondria; ina few sets of experiments 5mM glutamate and 5mM malatewere used as respiratory substrates as well.Therewere studiedvarious metabolic states of LLM: basal respiration, in thepresence of substrates, state 3 respiration, in the presence ofsubstrates and ADP, state 4 respiration, after exhausting ofexogenous ADP, and uncoupled respiration, in the presenceof 2, 4-DNP, FCCP, or CCCP (i.e., the maximal rate ofrespiration, limited only by capacity of the mitochondrialrespiratory chain, state 3u [40, 41]). In some cases, oligomycin(1 or 5𝜇g/mL) was supplemented in the assay medium. Forfurther details of the experimental procedure, see text andlegends to the corresponding figures. Oxygen consumptionrate values are presented in nmol O
2/min/mg of mito-
chondrial protein (in some cases, in ngatom O/min/mg ofmitochondrial protein). Respiration control ratio (RCR, state3/state 4 or state 3u/state 4), ADP/O, and phosphorylationrate were calculated from polarographic curves.
2.6. Assessment ofMitochondrial RespiratoryChainComplexesActivities. The measurement of the specific activity of theindividual complexes of the LLM respiratory chain was per-formed spectrophotometrically as described by Barrientos[42]withmodifications. Activitieswere expressed in units permilligramofmitochondrial protein. LLMwere freeze-thawedthree times to obtain maximum activities.
NADH-UbiquinoneOxidoreductase (Complex I) activitywas assayed as a decrease in absorbance at 340 nm byfollowing the oxidation of NADH. Mitochondria (40 𝜇g ofprotein/mL) were incubated for 2min in 800𝜇L of Milliporewater. 200𝜇L of reaction medium (50mM Tris (pH 8.0),5mg/mLBSA, 4mMNa azide, 4𝜇MantimycinA, and 80 𝜇M2,3-dimethoxy-5-methyl-6-n-decyl-1,4-benzoquinone, DB)was added and the reaction was initiated by the addition of0.8mM NADH. The activity of Complex I was determinedusing the rotenone-sensitive rate. The enzymatic activity wasquantified using an extinction coefficient of 6.22mM−1 cm−1.
For determination of NADH-Cytochrome c Oxidoreduc-tase (Complex I–III) activity the assay was performed at340 nm following the increase in absorbance resulting fromreduction of cytochrome c. At first, LLM were incubated inMillipore water (like for the Complex I activity determina-tion; see above).The reactionmedium containing 50mMTrisand 5mg/mL BSA was supplemented with 40𝜇M oxidizedcytochrome c and 4mM Na azide. The specific rotenone-sensitive rate was calculated with an extinction coefficient of6.22mM−1 cm−1.
Succinate-Ubiquinone Oxidoreductase (ComplexII) activity was determined by following the secondaryreduction of 2,6-dichlorphenolindophenol (DCPIP) by DBat 600 nm. The reaction mixture contained 10mM KH
2PO4
(pH 7.8), 2mM EDTA, and 1mg/mL BSA and 80 𝜇MDCPIP,
4 𝜇M rotenone, 4 𝜇M antimycin A, 0.2mM ATP, 10mMsuccinate, and 40 𝜇g of mitochondrial protein. The reactionwas initiated by the addition of 80𝜇M DB. The activity wasexpressed by using 1mM thenoyltrifluoroacetone (TTFA).
Measurement of Succinate-Cytochrome c Oxidoreduc-tase (Complex II-III activity) is performed at 550 nm fol-lowing the increase in absorbance resulting from reduc-tion of cytochrome c. The reaction medium contained10mM KH
2PO4(pH 7.8), 2mM EDTA, 1mg/mL BSA, 4 𝜇M
rotenone, 4mMNa azide, 0.2mMATP, 10mM succinate, andthemitochondria (40𝜇g of protein).The reaction was startedby addition of 40 𝜇M oxidized cytochrome c. The activityof Complex II was measured by using antimycin A-sensitiverate.
The activity of Ubiquinol-Cytochrome c Oxidoreductase(Complex III) was assayed at 550 nm by monitoring the rateof reduction of cytochrome c by reduced DB. The reactionwas initiated by addition of oxidized cytochrome c in thebuffer containing 10mM KH
2PO4(pH 7.8), 2mM EDTA,
1mg/mL BSA, 80𝜇MDBH2, 4 𝜇M rotenone, 4mMNa azide,
0.2mM ATP, and the mitochondria (40 𝜇g of protein). Thespecific activity of Complex III was calculated by subtractingthe nonenzymatic rate.
The activity of Cytochrome c-O2Oxidoreductase (Com-
plex IV) was measured by following the oxidation of reducedcytochrome c as a decrease in absorbance at 550 nm. Thereaction buffer contained 50mM H
2PO4(pH 7.0), 1mg/mL
BSA, 0.2% Tween, and the mitochondria (40 𝜇g of protein).The reaction was started by adding 20𝜇M of reduced cyto-chrome c.
The activities of Complexes II, III, IV, I–III, and II-IIIwere calculated by using 19.1mM−1 cm−1 extinction coeffi-cient.
In addition, quantitative determination of cytochromes inthe LLMwas also performed using the method of differentialspectrophotometry as before [14]. In particular, for oxidation-reduction of the cytochromes medium of incubation andcorresponding additions analogous to those described in[43] was used. The recording of spectra was performedon a Specord M-40 spectrophotometer (Germany) and theconcentrations of cytochromes c, c1, and aa3 were calculatedaccording to [43].
2.7. Determination of Mitochondrial Membrane Potential(ΔΨ𝑚𝑖𝑡𝑜
). An estimation of LLM membrane potential(ΔΨmito) was conducted according to [44], with thehelp of fluorescent lipophilic cation-safranine O, usinga Shimadzu RF-1501 spectrofluorometer (Japan) at theexcitation wavelength of 485 nm and emission wavelengthof 590 nm as before [27, 28]. The dynamics of ΔΨmitodissipation was determined also by the Millipore filtrationmethod (see Section 2.3). For more details see legendsto Figures 1, 2, and 6.
2.8. Data Analysis. Data were expressed as mean values ±SE. The results shown are mean or representative of a seriesof at least three independent experiments, unless otherwiseindicated. The statistics were analyzed with ANOVA and

BioMed Research International 5
Student’s t-tests, with 𝑃 < 0.05 assumed as the significancethreshold.Themitochondrial respiration and swelling curveswere plotted with the help of the program Origin 6.0.
3. Results and Discussion
3.1. Seasonal Changes in Action of Different Membrane Per-meability Modulators on the LLM Function. With the aimto obtain more tightly coupled mitochondria from lampreytissues, namely, from lamprey liver, for working during awhole year not depending on a season, in our early works[8, 9, 18] we modified isolation and incubation conditions;in particular, we found that in December–February LLMwith the highest RCR (see Section 2.5) could be obtainedwhen the isolation and/or incubation medium was supple-mented by nupercaine, Nup (also known under the nameof dibucaine), and/or heparin and/or BSA. Contrary, inour late works [12, 14] with the aid to study the seasonchanges in bioenergetics parameters of LLM, we removed theabove mentioned substances from the isolation/incubationprocedure. To better understand the mechanism(s) of theimproving action of these compounds, in this work weinvestigated more thoroughly the effects of these reagents onthe in vitro mitochondrial function of the lamprey liver.
Using the Millipore filtration method (see Section 2.3and [31, 39]), we studied an uptake of 137Cs+ by isolatedwinter LLM depending on energization in the absence andin the presence of different concentrations of Val in acetatemedium with or without Nup and/or RR (Figures 1 and 2).As known, the uptake of 137Cs+ by energized mitochondriain the presence of Val (estimated by the coefficient 𝑟
𝑡) charac-
terizes the energetic condition of isolated mitochondria andindirectly theΔΨmito value.Thismethod ofΔΨmito monitoringis especially convenient for determination of the dynamicsof ΔΨmito dissipation. We found herein (see also [18, 45])that at 0∘C the nonenergized winter LLM were practicallynot able to accumulate 137Cs+ in the absence of Val in theassay medium. As seen from Figure 1, an addition of 10−9Mof Val into the medium resulted in the significant 137Cs+uptake by the mitochondria, which further was followed bydecrease of the distribution ratio of 137Cs+ (𝑟Cs) betweenmatrix andmedium due to the developing diffusion potentialdissipated with time. An energization of the mitochondria bySucc and especially an additional supplement of Val (10−8M)into the medium produced the sharp rise of 𝑟Cs that, inturn, was declined by addition of 1mM NaCN (Figure 1).As seen from Figure 2, an addition of Nup (50𝜇M) intothe assay medium increased the rate of 137Cs+ uptake bywinter LLM, and the simultaneous addition of RR (7 𝜇M)still more enhanced this effect. It should be stressed thatthe values of 𝑟Cs found herein for winter LLM were 15–20 times lower than those of RLM measured by us beforeunder the same conditions [30–34].The lower rates of 137Cs+uptake by the LLM and the lower values of 𝑟Cs (comparedwith RLM) are in a good accordance with the increasedion membrane permeability of winter LLM found in ourprevious works [12, 14]. A one more marked distinction wehave found betweenwinter LLMandRLM relates to the effect
of Nup on the functioning of these mitochondria. Previouslyon isolated RLM [30–34] we showed that Nup (i.e., localanesthetics and an amphiphilic cation which is known toaffect membrane lipid-protein interactions and is consideredto be effective MPT pore inhibitor [46]) did not improve thefunction of untreated (control) RLM that is in contrast tothe situation found herein for the winter LLM the energeticcondition ofwhich becamebetter after addition of the effectorunder test (see Figure 2). Moreover, as we revealed earlier[33], Nup per se (taken in concentration of 50𝜇M) producedsignificant decrease in 137Cs+ uptake level of Succ-energizedRLM incubated in isotonic (sucrose plus acetate)-containedmedium supplemented by Val, oligomycin, and rotenone,that is, under the same experimental conditions which areused in this study and presented on Figure 2. Likewise, wehave found that Nup does not improve the functioning of thespring LLM and sometimes even made them worse (data notshown).
Practically the same situation we observed under study-ing the action of heparin on the in vitro LLM function hereinand before [12, 14]. As known, heparin is a sulfated anionicglycosaminoglycanwidely used as an anticoagulant in clinicalpractice for injections and to form an inner anticoagulantsurface on various experimental and medical devices such astest tubes and renal dialysis machines [47, 48]. Heparin andheparin-like glycosaminoglycans can modulate the cationicenvironment in different physiological systems, interactingwith such vital metal cations as iron, copper, and calcium[49–52]. It is known also that heparin suppresses lipid raft-mediated signaling in various biological systems [53]. In ourhands, the mitochondrial bioenergetics parameters of thewinter lamprey liver such as basal respiration rate, ADP-stimulated (phosphorylating) respiration rate, and RCR (seeSection 2.5) were found to be significantly affected by heparinand someMPT inhibitors (ADP,Mg2+, CsA, EGTA, BSA, RR,etc.); namely, they were improved in some extent by supple-ment of these reagents, while an influence of these chemicalson the uncoupler-stimulated mitochondrial respiration rate(the maximal respiratory rate, limited only by capacity of themitochondrial respiratory chain [40, 41]) depressed stronglyduring synchonia was marginal. Besides, heparin added tothe isolation and/or incubation medium decreased partiallythe enhanced ion permeability of the nonenergized winterLLM (see also [12, 14]). Opposite to this, heparin did notproduce any improving action on the function of the springLLM.
In this connection, it should be reminded about the veryinteresting data which was found by one of us more than30 years ago, concerning the action of heparin on the mito-chondrial reverse electron transport in the lamprey liver (i.e.,from succinate to NAD+ via Complex I of the mitochondrialrespiratory chain) [9]. In particular, Savina and Derkachevrevealed that in thewinter LLM the reverse electron transportwhich registered through changes in the pyridine nucleotidefluorescence according to [54] was disturbed. Remarkably,in the winter LLM isolated in the presence of heparin (50units/mL) the reverse electron transport was completely

6 BioMed Research International
restored. The occurrence of the mitochondrial reverse elec-tron transport in the LLM was used by us as an additionaltest for coupling of the mitochondria. It was suggested thatthe unknown factor, which uncoupled LLM andwas opposedby heparin, existed only at lower temperatures (4-5∘C) andvanished after adaptation of winter lampreys to elevatingtemperatures (from 1 to 18 days at 17∘C), because the livermitochondria of the adapted winter lamprey were tightlycoupled, including the restored reverse electron transport viaComplex I.Moreover, thesemitochondria resembled stronglyliver mitochondria of mammals, such as rat and mouse.Importantly, after placing the adapted lampreys to cold water,the mitochondria again became uncoupled that points tothe existence of a low-temperature hibernation period in thelamprey’s life cycle during synchonia [9].
Thus, all this agree well with our recent findings pointingto the opening of the MPT pore in its low-conductancestate in the winter LLM as well as with the suggestion aboutpossible involvement of Complex I of the mitochondrialrespiratory chain in this process ([12, 14], see also [55] andreferences therein). It should be stressed also that in ourfurther experiments, the results of which are presented inthis work, we used both heparin-treated and nontreated LLMdepending on the task and the season.
3.2. Seasonal Changes in Respiratory Chain Complexes Activ-ities in the LLM. In our previous works ([12, 14], see alsoFigure 5 herein) we have shown that the respiration rates ofthe LLM in various metabolic states (see Section 2.5) displaystrong seasonal differences depending on substrate used foroxidation. In particular, in winter (December–February) therates of oxidation of NAD-dependent substrates, pyruvate,and malate (i.e., Complex I substrates) by the LLM weredepressed significantly in all metabolic states under studywhile in spring (April) the rates of oxidation of the NAD-dependent substrates in state 4 increased 3 times, in thestate 3, 6 times and in the state 3u, 7-8 times; besides,the respiratory controls increased 2 times and the rates ofphosphorylation, 7–10 times. At the same time, the rates ofoxidation of FAD-dependent substrates, namely, succinate(i.e., Complex II substrates), both in the presence and inthe absence of rotenone (Complex I inhibitor) did not havesuch pronounced seasonal changes. In spring, their valuesincreased 1.3–2 times (depending on the metabolic state ofthe mitochondria) as compared to that in winter.
As well-known, three mitochondrial respiratory chaincomponents are involved in oxidation of NAD-dependentsubstrates, such as pyruvate, malate, and glutamate, namely,Complex I (EC 1.6.99.3), Complex III (EC 1.6.5.3), andComplex IV (EC 1.9.3.1). In oxidation of FAD-dependent sub-strates, such as succinate, Complex II (EC 1.3.99.1) participatein combination with Complexes III and IV. We have foundthat the content of cytochromes c, c1, and aa3 does not changethroughout the whole prespawning period (November–May): 0.15–0.17, 0.11–0.13, and 0.19–0.21 nmol/mg of mito-chondrial protein, respectively. We proposed that the samewas likely true for Complex II, as the rate of succinate
oxidation in the presence of an uncoupler (i.e., the max-imal rate of respiration, limited only by capacity of themitochondrial respiratory chain, state 3u [40, 41]), did notdiffer significantly during the indicated seasons (e.g., inthe case of 2, 4-DNP it was 18.7 ± 0.2 nmol O
2/min⋅mg
protein in winter and 25.1 ± 6.0 nmol O2/min⋅mg protein
in spring). We suggested also that Complexes III and IVremained equally active at the winter and spring periods ofprespawning migration. In contrast, the rates of oxidationof NAD-dependent substrates (pyruvate and malate) in thepresence of 2, 4-DNP substantially decreased at the winterperiod (4.4±0.6 nmol O
2/min⋅mg protein) as compared with
spring (31.4±8.3 nmolO2/min⋅mgprotein). Since the activity
of Complexes III and IV participating in theNAD-dependentsubstrates oxidation did not change during the prespawningperiod, one had to accept that during winter the respiratoryComplex I was inactivated. Importantly, the data obtainedwith heparin and under using of the thermal adaptation (seeSection 3.1 and [9]) also indicated that some substantial andreversible changes in functioning of the LLM respiratoryComplex I (in its activity, and/or in its conformation, and/orin its redox regulation) took place during the prespawningperiod.
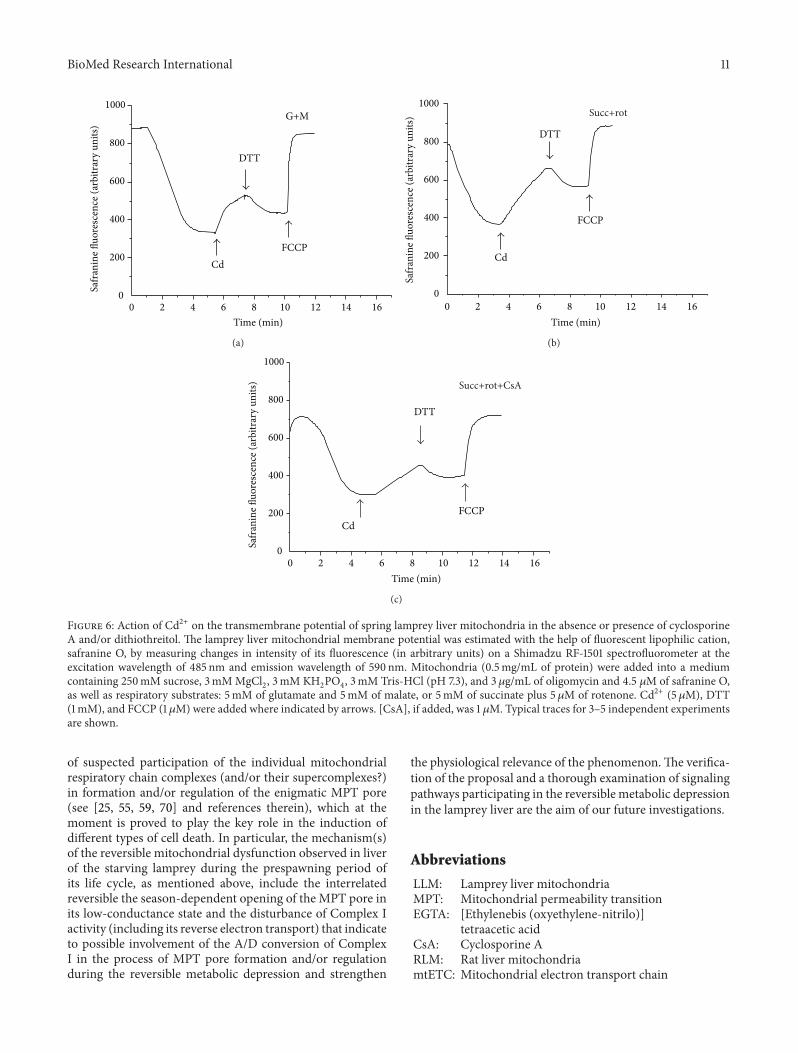
In the present study we decided to measure seasonalchanges (if so) in respiratory chain complexes activitiesin the LLM. In Figure 3 the activities of respiratory chaincomplexes in LLM in different periods of synchonia aredemonstrated (in units/g protein). It is seen that there areno significant seasonal differences in Complex II, ComplexIII, Complex I–III, and Complex II-III activities. ComplexI activity increased in March (110.5 ± 24.5) and April (199.1± 6.0), as compared with that in February (71.9 ± 9.9).Complex IV activity increased more than in one and ahalf—two times in spring (from 451.7 ± 95.1 in February to714.1 ± 40.4 and 1011.2 ± 27.2 in March and April, resp.).The differences between the activities in February and inApril were significant for both Complex I and Complex IV(Figure 3).
The shown data, namely, the substantially decreasedactivity of Complex I and the nonchanged activity of Com-plex II in the winter LLM (see Figure 3), agree well withthe data obtained from respiratory experiments conductedby us before [12, 14] and herein; in particular, as seen fromFigure 5(a), in winter the FAD-linked LLM respiration (trace5) is only partially suppressed in contrast to the NAD-linkedone, which is practically switched off (trace 4). This wasespecially evident, when the mitochondria were isolated inthe absence of heparin (data not shown). Notably, in springthe LLM respire well on both types of respiratory substratesused not depending on the presence of heparin, BSA, and soforth in the isolation/incubation medium (Figure 5(c)).
In this connection, it should be stressed that in differenttypes of cells mitochondrial respiratory Complex I is foundto be a subject of reversible covalent regulation by phospho-rylation and dephosphorylation [56, 57] and by thiol-basedredox modifications (for the recent comprehensive reviews,see [58, 59]). It seems quite possible that something likethat may relate to mechanism(s) of seasonal regulation ofComplex I in the lamprey liver as well.
BioMed Research International 7
0
100
200
300
400
500
600
700
800
900
1000
1100
I II III IV I–III II-IIIComplexes
Activ
ity (U
/g p
rote
in)
FebruaryMarchApril-May
∗
∗
Figure 3: Mitochondrial respiratory chain complexes activitiesin lamprey liver mitochondria depending on season. Enzymaticactivities were determined as described in Section 2 and expressedin terms of nmol/min/mg protein. Data were analyzed by ANOVAusing 6.0 statistical software. Means ± SE are plotted (𝑛 = 5). ∗—significantly different from the winter value (at 𝑃 < 0.05).
In particular, it is known now that control of thecatalytic activity of mitochondrial Complex I by the so-called active/deactive (A/D) transition involves a specificand well defined cysteine switch mechanism [58–60]. Thephenomenon of the A/D transition was revealed for thefirst time by Kotlyar and Vinogradov in submitochondrialparticles from bovine heart [61], and later it was found inintact mitochondria as well [62]. The A-form (i.e., active) ofthe enzyme catalyzed rotenone-sensitive NADH-ubiquinonereductase reaction, whilst the D-form (i.e., deactive) couldbe fully reduced by NADH and oxidized by artificial electronacceptors but was unable to transfer electrons to ubiquinone.An equilibrium between the A- and the D-forms of theenzyme (greatly shifted to the latter one) was reached inthe absence of any ligands and the rate of equilibrium wasexceptionally temperature-dependent (negligible at 15–20∘Cbut significant at 30–37∘C). Markedly, rotenone influencedon the spontaneously established equilibrium, namely, it par-tially protected and partially reversed the thermally induceddeactivation; moreover, rotenone had almost two order ofmagnitude higher affinity for the A-form. In addition, onlythe thermally deactivated enzyme was irreversibly inhibitedby SH-reagents such as N-ethylmaleimide. There were someother important differences between these two forms. Inparticular, the D-form was strongly inhibited by Ca2+ whilethere was no sensitivity of the A-form to this cation. Theaffinity of the D-form to polyvalent cations was found tobe in the following order: Ni2+ > Co2+ > La2+ > Mn2+ >Ca2+ =Mg2+ > Ba2+; besides, its affinity to Ca2+ was strongly
Swel
ling (control)
9
8
7
65
43
2
1
0.01ΔA
1 min
(Ca2++ Pi + FCCP + CsA)
(Ca2++ Pi + FCCP)
(Pi + CsA)(Pi)
(Ca2++ Pi + laurate + CsA)
(Ca2++ Pi + laurate)
(Ca2++ Pi + CsA)
(Ca2++ Pi)
Figure 4: Action of Ca2+, Pi, laurate, and FCCP on the membranepermeability of winter lamprey liver mitochondria. A membranepermeabilization of isolated lamprey mitochondria was determinedfrom their swelling (measured spectrophotometrically as apparentabsorbance changes at 540 nm) in 250mM sucrose medium (3mMTris-HCl, pH 7.3) at 20∘C in the presence of succinate, rotenone,and oligomycin; mitochondrial protein content, 0.5mg/mL. Theconcentrations used are: Ca2+, 20 𝜇M; Pi, 1 mM; FCCP, 100 nM;laurate, 30𝜇M; CsA, 2 𝜇M. Experiments were repeated five times,with data presented being representative.
pH-dependent. Furthermore, Ca2+ increased the reactivityof the enzyme sulfhydryl group in the D-form towardsN-ethylmaleimide. It was shown also that divalent cationslike Ca2+ [63] and covalent modification of the single cys-teine [64] can prevent reactivation of deactive Complex I.Importantly, because the divalent metal binding site and thecysteine are only accessible in the D-form of the enzyme,this makes the A-form resistant to this type of inhibition[60–64]. In addition, as found later by Maklashina et al., theA/D transition is an attribute of Complex I from vertebrates[65]. Interestingly, in the lower eukaryotes A/D transition ofComplex I is also observed, but deactivation is faster andtakes place at lower temperatures. Notably, during the lastdecade the evidence is accumulated, indicating that the A/Dtransition is more than an in vitro phenomenon [66–69].In particular, Maklashina et al. revealed its functional con-nection with anoxia-reperfusion injury, using Langendorffhearts [66]. Galkin and Moncada have shown induction ofthe D-form of Complex I during prolonged hypoxia and havesuggested that this could make the cysteine exposed only inthis state accessible for nitrosation that finally could resultin permanent deactivation of the enzyme and implicate forischemic injury [67, 68]. Besides, Prime et al. recently havefound significant cardioprotective effects in mice associatedwith the S-nitrosation of the cysteine switch of the A/Dtransition [69]. All these results demonstrate that the A/Dtransition and the associated cysteine switch are operationalin vivo.
The same is possible to say for the situation observedin hepatocytes of the winter lamprey. As mentioned above,Complex I is inactivated spontaneously (converts into the D-form) in the absence of substrates. As found, the fatty acids(FA) serve the main energy substrate for the lamprey livermitochondria duringwinter and spring prespawning periods.

8 BioMed Research International
6
5
4
3
2
1
5 min
S 26.157.4
41.8
ADP
DNP
60.9
S + RT
Py + M
S
S + RT
Py + M Cd
DNP
DNPDNP
DNPDNP
DNP
DNPDNP
DNP
ADP
ADP
ADP
Cd
Cd
ADP18.436.7
24.527.9
37.732.8
11.327.813.9 14.8 13.9 8.7
DNP
DNPDNP 36.8
39.139.139.6
48.148.1
22.4 529.2
25.0
520.725.3
39.127.6
512.0
12.012.3
15.5 DNP10.3
10.3DNP
300ngatom O
(a)
5 min
300 ngatom O 6
5
4
3
2
1
Cd5
Cd5
Cd5
Cd5
Cd5
Cd5 DNPDNPDNP
DNP DNPDNP
DNP
DNPDNP
DNP
DNPDNP
DNP
DNPDNP
S + RT
Py + M
S + RT + CsA
Py + M + CsA
S + CsA
S
38.438.4
49.949.0 42.2
38.3
45.4
51.144.0
51.1
38.4
40.342.238.4
46.146.1
46.140.3
40.3
42.2
12.2 6.1 4.4 3.5 2.6
12.28.7
8.7 8.7 4.4
(b)
DNPDNP
DNP
ADP
DNPDNP
DNPDNP
DNPDNP
DNP
ADP
ADP
G + M
G + M
Succ + rot
4.526.8
9.0
28.7
28.06.7
27.711.5
28.727.1
10.6
42.016.2
39.535.7
5 min
300ngatom O
RCR = 2.98 + BSA
RCR = 2.41− BSA
RCR = 2.59 + BSA
(c)
Figure 5: The lamprey liver mitochondrial respiration depending on the season, the substrate used and the presence of Cd2+ and/orcyclosporine A. The rates of LLM respiration (oxygen consumption rates) were monitored polarographically at 25∘C with a Clark oxygenelectrode in a thermostatic closed chamber of 1.5mL with magnetic stirring. The mitochondria (1mg of protein/mL) were added into amedium containing 250mM sucrose, 3mM MgCl
2, 3mM KH
2PO4, 3mM Tris-HCl, pH 7.3 ((a) and (b), winter LLM) or 100mM sucrose,
40mM KCl, 3mM MgCl2, 3mM KH
2PO4, and 20mM Tris-HCl, pH 7.3 (±50𝜇M of EGTA and/or 1mg/mL BSA) ((c), spring LLM). 5mM
pyruvate or glutamate and 1 or 5mMmalate or 5mM succinate (±5𝜇M of rotenone) were added to the incubation medium for energizationof the mitochondria. The concentration of oligomycin, if added, was 1–5 𝜇g/mL. The basal respiration rate (in the presence of substrates),in state 3 (substrates and 50 𝜇M ADP), in state 4 (after exhausting of ADP) and uncoupled respiration rate (2, 4-DNP was added in state4) were estimated. The respiration rates (ngatom O/min/mg of protein), respiratory control ratios [VO
2(3)/VO
2(4)], ADP/O
2, and the
phosphorylation rate were calculated from polarographic curves. Typical traces for three independentmitochondrial preparations are shown.

BioMed Research International 9
NADH generated in the process of 𝛽-oxidation of FA is thenoxidized by the respiratory chain Complex I. It seems thatsuppression of the energy metabolism in the lamprey livercells in winter starts from an inactivation of the Complex Iof the mitochondrial respiratory chain, one of the causes ofwhich, the most likely, is inaccessibility at this period of FA—themain energy substrate for the hepatocytes. It is known alsothat the D-form of Complex I slowly turns back into the A-form in the presence of NADH (orNADPH) and ubiquinone.So, in spring the transition of Complex I from inactive toactive forms likely to occur, when under effect of estrogenthe swift lipolysis of fats stored in the hepatocytes in the formof lipid droplets begins and their energy substrates, FA startcoming to the mitochondria [12, 14]. Besides, the data con-cerning the action of heparin and the thermal adaptation onthe mitochondrial reverse electron transport in the lampreyliver strongly support the idea that mechanism(s) of seasonalregulation of Complex I in the lamprey liver can involve theA/D transition of the complex. Moreover, the transition fromone form to another is associated with the conformationalalterations of the enzymatic complex by means of which, inopinion of several authors including us, the mitochondrialrespiratory chain components (individual complexes and/orsupercomplexes) can participate in regulation (and even information) of the Ca2+-dependent nonselective pore ([23, 25,33, 55, 70] and references therein).
3.3. Effects of Cd2+, Ca2+, Pi, and Fatty Acids on the LLMFunction andTheir Season Dependence
3.3.1. Action on the LLM Membrane Permeability. As men-tioned above, in our previous works [12, 14] we have foundthat the reversiblemitochondrial dysfunction (mitochondrialuncoupling in winter and coupling in spring) is observed inthe lamprey liver during the metabolic depression in the lastyear of the lamprey life cycle. Importantly, the LLM dysfunc-tion is found to manifest itself in the significant mitochon-drial membrane permeabilization. It should be reminded,however, that the winter LLM are highly permeable onlyto different ions, including protons, but not permeable tosucrose. Besides, proton membrane permeability of the LLMdepends on season and energization. In particular, the winterLLM are highly permeable to protons and swell extensivelyin 125mM NH
4NO3medium both in the absence and in
the presence of the oxidized substrates. In winter the passiveswelling intensity in NH
4NO3medium is about 30% higher
than that in April, and the permeabilization is completelyinsensitive to CsA. As reported elsewhere, CsA (i.e., a potentpharmacological desensitizer of the classical nonselectivepore in the inner mitochondrial membrane) is incapable toclose the pore in mitochondria depleted by ATP and ADP[20–25]. Before we have found that the content of ATPand ADP in the LLM, which is low per se, decreases inwinter by more than 20–30% [10, 11], so we have speculatedthat the loss of matrix adenine nucleotides could be oneof possible explanations why CsA does not have any effecton the winter nonenergized mitochondria. In spring thepassive proton permeability of the LLM membrane becomes
lower, while the intramitochondrial levels of ATP and ADPget higher. As a result, the inhibitory (although moderate)influence of CsA on the passive mitochondrial swelling inNH4NO3medium is observed. As to the energized LLM,
we have found that they are highly permeable to protonsin winter whilst in spring they lose this capability. Besides,the energized winter mitochondria are susceptible, althoughin a different extent, to several MPT pore inhibitors takenseparately or in combination, such as Mg2+, ADP, EGTA,RR, BSA, heparin, nupercaine, and CsA. However, bothnonenergized and energized winter LLM were not sensitiveto a dithiol reducing agent, dithiothreitol, DTT ([12, 14] andherein).
To better understand the mechanism(s) of inductionof the MPT pore in the lamprey liver and especially itsapparent seasonal dependence and physiological relevance,in the present work we continued studies in this direction.For this purpose, we used heavy metal ion Cd2+ (a potentdithiol reagent and Ca2+ agonist) and for comparison, Ca2+plus Pi (in some cases also several fatty acids, mainlylaurate) as potential MPT pore inducers and the isolatedmitochondria of both “winter” and “spring” animals. Wefound that Ca2+ plus Pi induced the high-amplitude swellingof the isolated winter LLM both in isotonic sucrose andammonium nitrate medium while both low and high Cd2+(1–10 𝜇M and up to 1mM, resp.) per se did not producethe mitochondrial swelling in these media. In particular, asseen from Figure 4, Ca2+ (20𝜇M), Pi (1mM), and laurate(30 𝜇M) applied separately or in combination produce high-amplitude swelling of the winter LLM energized by succinate(Succ) in the sucrose medium and it is only partially sensitiveto CsA. Moreover, both natural (laurate) and artificial (p-trifluoromethyl carbonyl cyanide phenylhydrazone, FCCP)uncouplers depressed markedly the Ca2+ plus Pi-promotedswelling of the lamprey mitochondria oxidizing Succ insucrose medium (Figure 4). It seems also that the Succ-energized untreated (control) winter LLM shrink in thismedium (Figure 4, trace 1). The addition of Pi per se to themedium (Figure 4, trace 5) stops this apparent shrinkage thatcorrelates well with recently revealed inhibitory potency of Pi[25]. In opposite, Cd2+ did not induce any swelling of thewin-ter LLM in the sucrose medium at all concentrations understudy and all experimental conditions used (including thepresence of heparin in isolation and/or incubation media).This sharply differ them from RLMwhere Cd2+ was found tobe the very effectiveMPT inducer [30–36].The close situationwas found by us for other heavy metals, namely, for Hg2+and Cu2+ (data not shown). Not long ago it was shown alsothat isolated hepatocytes of the winter lamprey were highlytolerant to exposure with different metabolic inhibitors, suchas rotenone, antimycin A, and oligomycin, and their viabilityin the presence of these stressors are temperature- and pH-dependent [13]. All these findings are in a good accordancewith data existed in literature in the field which obtained ondifferent species, such as oysters, shrimps, birds, bats, andnaked mole rats [4, 6, 7, 71–75].
In addition, we revealed the clear-cut season depend-ence in action of laurate on the LLM function that will

10 BioMed Research International
be discussed elsewhere. Briefly, the action of laurate asthe pore inducer depended on energetic condition of themitochondria, namely, in winter in the low-energetic LLMlaurate decreased the inducing action of Ca2+ plus Pi (seeFigure 4, trace 7), whereas in spring it enhanced the actionof these effectors. It is worthy of note also that in springmore than two times higher concentrations of Ca2+, Pi, andlaurate for the pore induction in the LLM were needed (datanot shown).
3.3.2. Action on the LLM Respiration. It is well known nowthat the respiratory function of the lamprey liver is a subject ofseason regulation; besides, the respiration of isolated winterlamprey mitochondria is substrate-dependent ([12, 14] andFigure 5 herein). Substantial differences in the action ofdifferent MPT pore modulators in the presence of NAD- orFAD-dependent respiratory substrates depending on seasonwere revealed as well. In the present work we studied theeffects of different concentrations of Cd2+ on the LLMrespiration in different metabolic states in the presence andin the absence of CsA. As seen from Figure 5(a), in the winterLLM low Cd2+ (5 𝜇M) decreased the ADP-stimulated (phos-phorylating) respiration rate (i.e., in state 3) and the DNP-uncoupled respiration rate (i.e., the maximal respiratory rate,limited only by the capacity of the mitochondrial respiratorychain [40, 41]) as well as RCR (respiratory controls) on bothtypes of respiratory substrates used; however, all the Cd2+effects were less pronounced as compared with that in RLM[30–36]. Notably, Cd2+ action on the basal respiration andon the resting state respiration (i.e., in state 4) was stronglydependent on the substrate used. In particular, we foundthat low Cd2+ enhanced the inhibition of the basal and state4 respiration rates of the lamprey mitochondria oxidizingNAD-dependent substrates, namely, Glu plusMal or Pyr plusMal (Figure 5(a), traces 1 and 4), whereas it evoked theirpartial stimulation on FAD-dependent substrates, namely, onSucc in the presence (Figure 5(a), traces 2 and 5) and in theabsence of rotenone (Figure 5(a), traces 3 and 6). Besides, allthe aforementioned changes in the respiration rates of thewinter LLM promoted by the heavy metal were only weakly(if so) sensitive to CsA (the MPT pore desensitizer) addedalone and in combination with oligomycin (Figure 5(b)).TheCd2+ effects were revealed in the LLM isolated both in thepresence and in the absence of heparin.
3.3.3. Action on the LLMMembrane Potential (ΔΨ𝑚𝑖𝑡𝑜
). Aswefound before under measuring the dynamics of the lampreyliver mitochondrial membrane potential (during the wholeprespawning migration period of the animal) estimated byaccumulation of fluorescent lipophilic cations, safranine Oby isolatedmitochondria [27, 28] or TMRM(tetramethylrho-daminemethyl ester) by isolated cells [13], theΔΨmito changeswere strongly dependent on season and types of substratesused for energization (the latter—for isolated LLM). Inparticular, it was found on the isolated hepatocytes that theΔΨmito values were maximal in October and March-April,while in winter its values were substantially declined that pro-voked likely by prolonged starvation and profoundmetabolic
arrest [13]. On the isolated winter LLM it was shown alsothat the intensity of safranine O fluorescence reflecting themembrane potential was lower at the pyruvate and malateoxidation during the period of metabolic depression anddissipated much more rapidly than at oxidation of succinate[27]. Remarkably, the addition of ADP or/and EGTA (i.e.,the potent MPT effectors) into incubation medium led tothe significant delay of the membrane potential dissipation ofthe winter LLM energized by NAD-depended substrates thatstrongly supported the suggestion about the involvement ofthe Complex I in the process of the pore formation and/orregulation.
In the present work we investigated the action of Cd2+ onthe ΔΨmito changes of the isolated LLM energized by differenttypes of respiratory substrates in the presence and in theabsence of CsA (the strong MPT pore desensitizer) and/orDTT (the potent dithiol reductant).
As seen fromFigure 6, in spring the LLMwere found to behighly sensitive to Cd2+ when energized by both NAD- andFAD-depended substrates. In particular, under monitoringof the transmembrane potential of the “spring” lamprey livermitochondria with the help of safranine O, we revealed thatCd2+ produced the significant decrease of the ΔΨmito on bothtypes of the respiratory substrates used that was stronglyhampered by CsA, and the membrane potential was partiallyrestored by DTT (Figure 6).
So, the data obtained by us herein and before indicatethat in winter the pore in the LLM opened in its low-conductance state is Ca2+-dependent, sensitive to Mg2+,ADP, CsA, heparin, nupercaine, and so forth (although ina different extent) but not sensitive to DTT. Moreover, thewinter LLM were highly tolerant (in some aspects) to suchpotent thiol reagents and inducers of oxidative stress andMPT, as heavy metals; this sharply distinct them from RLM.On the contrary, in spring the LLM resemble RLM verymuch, including the sensitivity to Cd2+ and DTT (Figure 6).These facts combined with the observations of high level ofreduced PN in the LLM point to the absence of the oxidativestress in the winter lamprey hepatocytes. At present it isclear that the energetic condition of the lamprey hepatocytesis strictly regulated; however, signaling pathways involvedin this regulation are so far unknown. In our opinion,one of these pathways probably connect with creation ofinaccessibility of some critical thiol(s) most likely related tothe A/D transition of Complex I and involved in regulationor/and formation of the MPT pore (see Section 3.2 and [58,59, 64–69]). Importantly, the absence of the severe oxidativestress may be one of the main causes of reversibility of themetabolic depression observed in the lamprey hepatocytesduring prespawning migration of the animal in contrast tothe irreversible metabolism suppression and mitochondrialdysfunction revealed in various types of cells under differentdiseases and pathological conditions [76–78].
3.4. Concluding Remarks. In conclusion, it should be stressedthat liver mitochondria and hepatocytes of the Baltic lampreyand the season changes observed in their functioning arean excellent “natural” model for elucidating mechanism(s)
BioMed Research International 11
FCCPCd
DTT
Safr
anin
e fluo
resc
ence
(arb
itrar
y un
its)
Time (min)0 2 4 6 8 10 12 14 16
1000
800
600
400
200
0
G+M
(a)
FCCP
Cd
DTT
Safr
anin
e fluo
resc
ence
(arb
itrar
y un
its)
Time (min)0 2 4 6 8 10 12 14 16
1000
800
600
400
200
0
Succ+rot
(b)
FCCPCd
DTT
Safr
anin
e flu
ores
cenc
e (a
rbitr
ary
units
)
Time (min)0 2 4 6 8 10 12 14 16
1000
800
600
400
200
0
Succ+rot+CsA
(c)
Figure 6: Action of Cd2+ on the transmembrane potential of spring lamprey liver mitochondria in the absence or presence of cyclosporineA and/or dithiothreitol. The lamprey liver mitochondrial membrane potential was estimated with the help of fluorescent lipophilic cation,safranine O, by measuring changes in intensity of its fluorescence (in arbitrary units) on a Shimadzu RF-1501 spectrofluorometer at theexcitation wavelength of 485 nm and emission wavelength of 590 nm. Mitochondria (0.5mg/mL of protein) were added into a mediumcontaining 250mM sucrose, 3mMMgCl
2, 3mM KH
2PO4, 3mM Tris-HCl (pH 7.3), and 3𝜇g/mL of oligomycin and 4.5 𝜇M of safranine O,
as well as respiratory substrates: 5mM of glutamate and 5mM of malate, or 5mM of succinate plus 5 𝜇M of rotenone. Cd2+ (5 𝜇M), DTT(1mM), and FCCP (1𝜇M) were added where indicated by arrows. [CsA], if added, was 1 𝜇M. Typical traces for 3–5 independent experimentsare shown.
of suspected participation of the individual mitochondrialrespiratory chain complexes (and/or their supercomplexes?)in formation and/or regulation of the enigmatic MPT pore(see [25, 55, 59, 70] and references therein), which at themoment is proved to play the key role in the induction ofdifferent types of cell death. In particular, the mechanism(s)of the reversible mitochondrial dysfunction observed in liverof the starving lamprey during the prespawning period ofits life cycle, as mentioned above, include the interrelatedreversible the season-dependent opening of the MPT pore inits low-conductance state and the disturbance of Complex Iactivity (including its reverse electron transport) that indicateto possible involvement of the A/D conversion of ComplexI in the process of MPT pore formation and/or regulationduring the reversible metabolic depression and strengthen
the physiological relevance of the phenomenon.The verifica-tion of the proposal and a thorough examination of signalingpathways participating in the reversible metabolic depressionin the lamprey liver are the aim of our future investigations.
Abbreviations
LLM: Lamprey liver mitochondriaMPT: Mitochondrial permeability transitionEGTA: [Ethylenebis (oxyethylene-nitrilo)]
tetraacetic acidCsA: Cyclosporine ARLM: Rat liver mitochondriamtETC: Mitochondrial electron transport chain

12 BioMed Research International
Pi: Inorganic phosphateDTT: DithiothreitolBSA: Bovine serum albuminFCCP: Carbonyl cyanide
p-trifluoromethoxyphenylhydrazoneCCCP: Carbonyl cyanide
3-chlorophenylhydrazoneTMPD: Tetramethyl-p-phenylenediamineVal: ValinomycinDNP: 2, 4-DinitrophenolNup: Nupercaine (dibucaine)RR: Ruthenium RedRCR: Respiratory control ratioΔΨmito: Mitochondrial membrane potentialPN: Pyridine nucleotidesROS: Reactive oxygen speciesDB: 2, 3-Dimethoxy-5-methyl-6-n-decyl-
1,4-benzoquinoneDCPIP: 2, 6-DichlorphenolindophenolTTFA: 2-ThenoytrifluoroacetoneFA: Fatty acidsSucc: SuccinatePy: Pyruvate.
Conflict of Interests
The authors do not have a direct financial relation with thecommercial identity mentioned in the paper that might leadto conflict of interests for any of the authors.
Acknowledgments
This work has been supported partially by grants from theRussian Foundation for Basic Research (no. 07-04-00722, no.08-04-00564, and no. 13-04-00011). The authors thank Dr V.V. Glazunov (isotope technique) andDrA.D.Nadeev (ΔΨmitomonitoring) for their fruitful assistance.
References
[1] P.W. Hochachka and P. L. Lutz, “Mechanism, origin, and evolu-tion of anoxia tolerance in animals,” Comparative Biochemistryand Physiology: B Biochemistry and Molecular Biology, vol. 130,no. 4, pp. 435–459, 2001.
[2] R. G. Boutilier and J. St-Pierre, “Adaptive plasticity of skeletalmuscle energetics in hibernating frogs: mitochondrial protonleak during metabolic depression,” Journal of ExperimentalBiology, vol. 205, no. 15, pp. 2287–2296, 2002.
[3] K. B. Storey and J. M. Storey, “Tribute to P. L. Lutz: Putting lifeon ‘pause’—molecular regulation of hypometabolism,” Journalof Experimental Biology, vol. 210, no. 10, pp. 1700–1714, 2007.
[4] S. C. Hand and M. A. Menze, “Mitochondria in energy-limitedstates: mechanisms that blunt the signaling of cell death,”Journal of Experimental Biology, vol. 211, no. 12, pp. 1829–1840,2008.
[5] M. V. Savina, L. V. Emelyanova, and I. V. Brailovskaya, “Bioen-ergetics of the lower vertebrates. Molecular mechanisms ofadaptations to anoxia and hypoxia,” Journal of EvolutionaryBiochemistry and Physiology, vol. 45, no. 2, pp. 197–210, 2009.
[6] M. A. Menze, G. Fortner, S. Nag, and S. C. Hand, “Mechanismsof apoptosis in Crustacea: what conditions induce versussuppress cell death?” Apoptosis, vol. 15, no. 3, pp. 293–312, 2010.
[7] F. Van Breukelen, G. Krumschnabel, and J. E. Podrabsky,“Vertebrate cell death in energy-limited conditions and how toavoid it: what wemight learn frommammalian hibernators andother stress-tolerant vertebrates,” Apoptosis, vol. 15, no. 3, pp.386–399, 2010.
[8] M.V. Savina,A.Wroniszewska, andL.Wojtczak, “Mitochondriafrom the lamprey (Lampetra fluviatilis). Oxidative phosphory-lation and related processes,” Acta Biochimica Polonica, vol. 22,no. 3, pp. 229–238, 1975.
[9] M. V. Savina and E. F. Derkachev, “Switch on and switch off phe-nomenon of liver gluconeogenic function in lamprey (Lampetrafluviatilis L.) under the influence of season and temperature,”Comparative Biochemistry and Physiology B: Biochemistry andMolecular Biology, vol. 75, no. 3, pp. 531–539, 1983.
[10] M. V. Savina and N. L. Gamper, “Respiration and adeninenucleotides of Baltic lamprey (Lampetra fluviatilis L.) hepato-cytes during spawning migration,” Comparative Biochemistryand Physiology B: Biochemistry and Molecular Biology, vol. 120,no. 2, pp. 375–383, 1998.
[11] N. L. Gamper and M. V. Savina, “Reversible metabolic depres-sion in hepatocytes of lamprey (Lampetra fluviatilis) duringpre-spawning: regulation by substrate availability,”ComparativeBiochemistry and Physiology B: Biochemistry and MolecularBiology, vol. 127, no. 2, pp. 147–154, 2000.
[12] M. V. Savina, L. V. Emelyanova, and E. A. Belyaeva, “Bioener-getic parameters of lamprey and frog liver mitochondria duringmetabolic depression and activity,” Comparative Biochemistryand Physiology B: Biochemistry and Molecular Biology, vol. 145,no. 3-4, pp. 296–305, 2006.
[13] M. V. Savina, S. A. Konovalova, I. S. Zubatkina, and A. A. Niki-forov, “Reversible metabolic depression in lamprey hepatocytesduring prespawning migration: dynamics of mitochondrialmembrane potential,”Comparative Biochemistry and PhysiologyB: Biochemistry and Molecular Biology, vol. 160, no. 4, pp. 194–200, 2011.
[14] L. V. Emel'yanova, M. V. Savina, E. A. Belyaeva, and I. V.Brailovskaya, “Peculiarities of functioning of livermitochondriaof the river lamprey Lampetra fluviatilis and the common frogRana temporaria at periods of suppression and activation ofenergy metabolism,” Journal of Evolutionary Biochemistry andPhysiology, vol. 43, no. 6, pp. 564–572, 2007.
[15] L. O. Larsen, “Physiology of adult lampreys, with special regardto natural starvation, reproduction and death after spawning,”Canadian Journal of Fisheries and Aquatic Sciences, vol. 37, no.11, pp. 1762–1779, 1980.
[16] T. P. Mommsen and P. J. Walsh, “Vitellogenesis and oocyteassembly,” Fish Physiology, vol. 11, pp. 347–406, 1988.
[17] K. R. Mewes, M. Latz, H. Golla, and A. Fischer, “Vitellogeninfrom female and estradiol-stimulated male river lampreys(Lampetra fluviatilis L.),” Journal of Experimental Zoology, vol.292, no. 1, pp. 52–72, 2002.
[18] M. V. Savina, I. A. Skul’skii, V. V. Glazunov, T. I. Ivanova, andS. M. Korotkov, “Functional peculiarities of liver mitochondriafrom the lamprey Lampetra fluviatilis,” Journal of EvolutionaryBiochemistry and Physiology, vol. 25, no. 6, pp. 710–717, 1989.
[19] D. R. Hunter, R. A. Haworth, and J. H. Southard, “Relationshipbetween configuration, function, and permeability in calciumtreated mitochondria,”The Journal of Biological Chemistry, vol.251, no. 16, pp. 5069–5077, 1976.

BioMed Research International 13
[20] M. Zoratti and I. Szabo, “The mitochondrial permeabilitytransition,” Biochimica et Biophysica Acta, Reviews on Biomem-branes, vol. 1241, no. 2, pp. 139–176, 1995.
[21] F. Ichas and J.-P. Mazat, “From calcium signaling to cell death:two conformations for the mitochondrial permeability tran-sition pore. Switching from low- to high-conductance state,”Biochimica et Biophysica Acta, Bioenergetics, vol. 1366, no. 1-2,pp. 33–50, 1998.
[22] A. P. Halestrap, “What is the mitochondrial permeability tran-sition pore?” Journal of Molecular and Cellular Cardiology, vol.46, no. 6, pp. 821–831, 2009.
[23] L. Azzolin, S. von Stockum, E. Basso, V. Petronilli, M. A. Forte,and P. Bernardi, “The mitochondrial permeability transitionfromyeast tomammals,”FEBS Letters, vol. 584, no. 12, pp. 2504–2509, 2010.
[24] S. Uribe-Carvajal, L. A. Luevano-Martınez, S. Guerrero-Castillo, A. Cabrera-Orefice, N. A. Corona-de-la-Pena, andM. Gutierrez-Aguilar, “Mitochondrial Unselective Channelsthroughout the eukaryotic domain,”Mitochondrion, vol. 11, no.3, pp. 382–390, 2011.
[25] P. Bernardi, “The mitochondrial permeability transition pore: amystery solved?” Frontiers in Physiology, vol. 4, no. 95, 2013.
[26] L. V. Emel’yanova, “Disturbance of themitochondrial functionsin hepatocytes of the lamprey Lampetra fluviatilis L. at thewinter period of prespawning migration,” Vestnyk MolodykhUchenykh, vol. 2, pp. 50–57, 2004.
[27] I. V. Brailovskaya, L. V. Emel'yanova, S. M. Korotkov, M. V.Savina, and V. V. Furaev, “Role of electron transport chainin liver mitochondria of the lamprey Lampetra fluviatilis dur-ing a decrease and activation of energy metabolism at theprespawning period,” Journal of Evolutionary Biochemistry andPhysiology, vol. 43, no. 6, pp. 593–595, 2007.
[28] M. V. Savina, L. V. Emel'yanova, S. M. Korotkov, I. V.Brailovskaya, andA.D.Nadeev, “Bioenergetics ofmitochondriaof the liver with biliary atresia during prolonged starvation,”Doklady Biochemistry and Biophysics, vol. 425, no. 1, pp. 80–83,2009.
[29] I. A. Skul'skiı, S. M. Korotkov, V. V. Glazunov, T. I. Ivanova, andM. V. Savina, “Effect of cadmium ions on the respiration, cationtransport and morphology of the liver mitochondria in the ratand the lamprey,” Tsitologiya, vol. 30, no. 8, pp. 956–962, 1988.
[30] E. A. Belyaeva, V. V. Glazunov, E. R. Nikitina, and S. M.Korotkov, “Bivalent metal ions modulate Cd2+ effects onisolated rat liver mitochondria,” Journal of Bioenergetics andBiomembranes, vol. 33, no. 4, pp. 303–318, 2001.
[31] E. A. Belyaeva, V. V. Glazunov, and S. M. Korotkov,“Cyclosporin A-sensitive permeability transition pore isinvolved in Cd2+-induced dysfunction of isolated rat livermitochondria: doubts no more,” Archives of Biochemistry andBiophysics, vol. 405, no. 2, pp. 252–264, 2002.
[32] E. A. Belyaeva and S. M. Korotkov, “Mechanism of pri-mary Cd2+-induced rat liver mitochondria dysfunction: dis-crete modes of Cd2+ action on calcium and thiol-dependentdomains,” Toxicology and Applied Pharmacology, vol. 192, no. 1,pp. 56–68, 2003.
[33] E. A. Belyaeva, V. V. Glazunov, and S.M. Korotkov, “Cd2+ versusCa2+-produced mitochondrial membrane permeabilization: aproposed direct participation of respiratory complexes I andIII,”Chemico-Biological Interactions, vol. 150, no. 3, pp. 253–270,2004.
[34] E. A. Belyaeva, V. V. Glazunov, and S. M. Korotkov, “Cd2+-promotedmitochondrial permeability transition: a comparison
with other heavy metals,” Acta Biochimica Polonica, vol. 51, no.2, pp. 545–551, 2004.
[35] E. A. Belyaeva, “Mitochondrial respiratory chain inhibitorsmodulate the metal-induced inner mitochondrial membranepermeabilization,” Acta Biochimica Polonica, vol. 57, no. 4, pp.435–441, 2010.
[36] E. A. Belyaeva, S. M. Korotkov, and N.-E. Saris, “In vitromodulation of heavymetal-induced rat liver mitochondria dys-function: a comparison of copper andmercury with cadmium,”Journal of Trace Elements in Medicine and Biology, vol. 25, no. 1,pp. S63–S73, 2011.
[37] E. A. Belyaeva, D. Dymkowska, M. R. Wieckowski, and L.Wojtczak, “Mitochondria as an important target in heavy metaltoxicity in rat hepatoma AS-30D cells,” Toxicology and AppliedPharmacology, vol. 231, no. 1, pp. 34–42, 2008.
[38] E.A. Belyaeva, L. V. Emel’yanova, S.M.Korotkov, andM.V. Sav-ina, “Mechanism(s) of liver mitochondria dysfunction inducedby heavy metal ions in vitro: a comparison of homothermic(rat) and poikilothermic (lamprey) vertebrates,” in Proceedingsof the All-Russian conference ‘Structural and functional bases ofevolution of functions and physiology of extreme conditions’, pp.18–19, IEPhB RAS, Sankt-Petersburg, Russia, 2008.
[39] I. A. Skulskii, N. E. L. Saris, andV. V. Glasunov, “The effect of theenergy state of mitochondria on the kinetics of unidirectionalcation fluxes,” Archives of Biochemistry and Biophysics, vol. 226,no. 1, pp. 337–346, 1983.
[40] B. Chance and G. R. Williams, “The respiratory chain andoxidative phosphorylation,”Advances in enzymology and relatedsubjects of biochemistry, vol. 17, pp. 65–134, 1956.
[41] M. D. Brand and D. G. Nicholls, “Assessing mitochondrialdysfunction in cells,” Biochemical Journal, vol. 435, no. 2, pp.297–312, 2011.
[42] A. Barrientos, “In vivo and in organello assessment of OXPHOSactivities,”Methods, vol. 26, no. 4, pp. 307–316, 2002.
[43] D. A. Knorre, V. I. Dedukhova, M. Y. Vyssokikh, and E.N. Mokhova, “Cyclosporin A-sensitive cytochrome c releaseand activation of external pathway of NADH oxidation inliver mitochondria due to pore opening by acidification ofphosphate-containing incubation medium,” Bioscience Reports,vol. 23, no. 2-3, pp. 67–75, 2003.
[44] K. O. Akerman andM. K. F. Wikstrom, “Safranine as a probe ofthe mitochondrial membrane potential,” FEBS Letters, vol. 68,no. 2, pp. 191–197, 1976.
[45] S. M. Korotkov, I. A. Skulskii, and V. V. Glazunov, “Cd2+effects on respiration and swelling of rat liver mitochondriawere modified by monovalent cations,” Journal of InorganicBiochemistry, vol. 70, no. 1, pp. 17–23, 1998.
[46] A. Szewczyk and L.Wojtczak, “Mitochondria as a pharmacolog-ical target,” Pharmacological Reviews, vol. 54, no. 1, pp. 101–127,2002.
[47] M. M. Cox and D. L. Nelson, Lehninger Principles of Biochem-istry, WH Freeman, Gordonsville, VA, USA, 2004.
[48] K. Mohanty, S. Mishra, J. Pani et al., “Heparin or EDTA:anticoagulant of choice in free radical estimation,”Oxidants andAntioxidants in Medical Science, vol. 1, no. 1, pp. 21–24, 2012.
[49] M. A. Ross, W. F. Long, and F. B. Williamson, “Inhibitionby heparin of Fe(II)-catalysed free-radical peroxidation oflinolenic acid,” Biochemical Journal, vol. 286, no. 3, pp. 717–720,1992.
[50] S. D. Grant, W. F. Long, and F. B. Williamson, “Similarityand dissimilarity in aspects of the binding to heparin of Ca2+

14 BioMed Research International
and Zn2+ as revealed by potentiometric titration,” BiochemicalSociety Transactions, vol. 24, no. 2, p. 203S, 1996.
[51] R. Albertini, S. Rindi, A. Passi, G. Pallavicini, and G. De Luca,“Heparin protection against Fe2+-and Cu2+-mediated oxidationof liposomes,” FEBS Letters, vol. 383, no. 3, pp. 155–158, 1996.
[52] T. Miralem and D. M. Templeton, “Heparin inhibitsCa2+/calmodulin-dependent kinase II activation and c-fosinduction in mesangial cells,” Biochemical Journal, vol. 330, no.2, pp. 651–657, 1998.
[53] Y.-T. Liu, L. Song, and D. M. Templeton, “Heparin suppresseslipid raft-mediated signaling and ligand-independent EGFreceptor activation,” Journal of Cellular Physiology, vol. 211, no.1, pp. 205–212, 2007.
[54] B. Chance and G. Hollunger, “Succinate-linked pyridinenucleotide reduction in mitochondria,” Federation proceedings,vol. 16, p. 163, 1957.
[55] E. A. Belyaeva, T. V. Sokolova, L. V. Emelyanova, and I. O.Zakharova, “Mitochondrial electron transport chain in heavymetal-induced neurotoxicity: effects of cadmium, mercury, andcopper,” Scientific World Journal, vol. 2012, Article ID 136063,2012.
[56] S. Scacco, R. Vergari, R. C. Scarpulla et al., “cAMP-dependentphosphorylation of the nuclear encoded 18-kDa (IP) subunit ofrespiratory complex I and activation of the complex in serum-starved mouse fibroblast cultures,” The Journal of BiologicalChemistry, vol. 275, no. 23, pp. 17578–17582, 2000.
[57] S. Papa, A. M. Sardanelli, S. Scacco et al., “The NADH:ubiquinone oxidoreductase (Complex I) of themammalian res-piratory chain and the cAMP cascade,” Journal of Bioenergeticsand Biomembranes, vol. 34, no. 1, pp. 1–10, 2002.
[58] M. P. Murphy, “Mitochondrial thiols in antioxidant protectionand redox signaling: distinct roles for glutathionylation andother thiol modifications,” Antioxidants and Redox Signaling,vol. 16, no. 6, pp. 476–495, 2012.
[59] S. Drose, U. Brandt, and I. Wittig, “Mitochondrial respiratorychain complexes as sources and targets of thiol-based redox-regulation,” Biochimica et Biophysica Acta, Proteins and Pro-teomics, 2014.
[60] A. D. Vinogradov, “Catalytic properties of the mitochon-drial NADH-ubiquinone oxidoreductase (Complex I) and thepseudo-reversible active/inactive enzyme transition,” Biochim-ica et Biophysica Acta, Bioenergetics, vol. 1364, no. 2, pp. 169–185,1998.
[61] A. B. Kotlyar and A. D. Vinogradov, “Slow active/inactivetransition of the mitochondrial NADH-ubiquinone reductase,”Biochimica et Biophysica Acta, Bioenergetics, vol. 1019, no. 2, pp.151–158, 1990.
[62] V. G. Grivennikova, A. N. Kapustin, and A. D. Vino-gradov, “Catalytic activity of NADH-ubiquinone oxidoreduc-tase (Complex I) in intact mitochondria: evidence for the slowactive/inactive transition,” The Journal of Biological Chemistry,vol. 276, no. 12, pp. 9038–9044, 2001.
[63] A. B. Kotlyar, V. D. Sled, and A. D. Vinogradov, “Effect of Ca2+ions on the slow active/inactive transition of the mitochondrialNADH-ubiquinone reductase,” Biochimica et Biophysica Acta,Bioenergetics, vol. 1098, no. 2, pp. 144–150, 1992.
[64] E. V. Gavrikova and A. D. Vinogradov, “Active/de-active statetransition of themitochondrial complex I as revealed by specificsulfhydryl group labeling,”FEBS Letters, vol. 455, no. 1-2, pp. 36–40, 1999.
[65] E. Maklashina, A. B. Kotlyar, and G. Cecchini, “Active/de-active transition of respiratory complex I in bacteria, fungi, and
animals,” Biochimica et Biophysica Acta, Bioenergetics, vol. 1606,no. 1–3, pp. 95–103, 2003.
[66] E. Maklashina, Y. Sher, H.-Z. Zhou, M. O. Gray, J. S. Karliner,and G. Cecchini, “Effect of anoxia/reperfusion on the reversibleactive/de-active transition of NADH-ubiquinone oxidoreduc-tase (complex I) in rat heart,” Biochimica et Biophysica Acta,Bioenergetics, vol. 1556, no. 1, pp. 6–12, 2002.
[67] A. Galkin and S. Moncada, “S-nitrosation of mitochondrialcomplex I depends on its structural conformation,”The Journalof Biological Chemistry, vol. 282, no. 52, pp. 37448–37453, 2007.
[68] A. Galkin, A. Y. Abramov, N. Frakich, M. R. Duchen, and S.Moncada, “Lack of oxygen deactivates mitochondrial complexI: implications for ischemic injury?” The Journal of BiologicalChemistry, vol. 284, no. 52, pp. 36055–36061, 2009.
[69] T. A. Prime, F. H. Blaikie, C. Evans et al., “A mitochondria-targeted S-nitrosothiol modulates respiration, nitrosates thiols,and protects against ischemia-reperfusion injury,” Proceedingsof the National Academy of Sciences of the United States ofAmerica, vol. 106, no. 26, pp. 10764–10769, 2009.
[70] G. Lenaz, A. Baracca, G. Barbero et al., “Mitochondrial respi-ratory chain super-complex I-III in physiology and pathology,”Biochimica et Biophysica Acta, Bioenergetics, vol. 1797, no. 6-7,pp. 633–640, 2010.
[71] I. M. Sokolova, “Cadmium effects on mitochondrial functionare enhanced by elevated temperatures in a marine poik-ilotherm, Crassostrea virginica Gmelin (Bivalvia: Ostreidae),”Journal of Experimental Biology, vol. 207, no. 15, pp. 2639–2648,2004.
[72] I. M. Sokolova, S. Evans, and F.M.Hughes, “Cadmium-inducedapoptosis in oyster hemocytes involves disturbance of cellularenergy balance but no mitochondrial permeability transition,”Journal of Experimental Biology, vol. 207, no. 19, pp. 3369–3380,2004.
[73] M. A. Menze, K. Hutchinson, S. M. Laborde, and S. C.Hand, “Mitochondrial permeability transition in the crustaceanArtemia franciscana: absence of a calcium-regulated pore inthe face of profound calcium storage,” American Journal ofPhysiology: Regulatory Integrative and Comparative Physiology,vol. 289, no. 1, pp. R68–R76, 2005.
[74] J. M. Harper, A. B. Salmon, S. F. Leiser, A. T. Galecki, andR. A. Miller, “Skin-derived fibroblasts from long-lived speciesare resistant to some, but not all, lethal stresses and to themitochondrial inhibitor rotenone,” Aging Cell, vol. 6, no. 1, pp.1–13, 2007.
[75] J.M.Harper,M.Wang, A. T. Galecki, J. Ro, J. B.Williams, and R.A. Miller, “Fibroblasts from long-lived bird species are resistanttomultiple forms of stress,” Journal of Experimental Biology, vol.214, no. 11, pp. 1902–1910, 2011.
[76] A. J. Kowaltowski, R. F. Castilho, and A. E. Vercesi, “Mitochon-drial permeability transition and oxidative stress,” FEBS Letters,vol. 495, no. 1-2, pp. 12–15, 2001.
[77] F. N. Gellerich, S. Trumbeckaite, T. Muller et al., “Energeticdepression caused by mitochondrial dysfunction,” Molecularand Cellular Biochemistry, vol. 256-257, no. 1-2, pp. 391–405,2004.
[78] A. Krivoruchko and K. B. Storey, “Forever young: mechanismsof natural anoxia tolerance and potential links to longevity,”OxidativeMedicine and Cellular Longevity, vol. 3, no. 3, pp. 186–198, 2010.
Submit your manuscripts athttp://www.hindawi.com
Stem CellsInternational
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Disease Markers
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation http://www.hindawi.com Volume 2014
Immunology ResearchHindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttp://www.hindawi.com Volume 2014
Parkinson’s Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttp://www.hindawi.com
Copyright © 2022 FDOKUMEN




















