
Import of hybrid forms of CYP11A1 into yeast mitochondria
10
Import of hybrid forms of CYP11A1 into yeast mitochondria A.N. Minenko, L.A. Novikova, V.N. Luzikov † , I.E. Kovaleva ⁎ Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, Moscow 119992, Russia abstract article info Article history: Received 14 February 2008 Received in revised form 5 June 2008 Accepted 16 June 2008 Available online 21 June 2008 Keywords: CYP11A1 Topogenic signal Yeast mitochondria Membrane insertion Heterologous expression in yeast of mCYP11A1 fusions with different topogenic signals of yeast mitochondrial proteins for artificial channeling to different translocases of the inner membrane was used to gain insight in the mechanism of its topogenesis in mitochondria. To ensure insertion of the CYP11A1 domain into the inner mitochondrial membrane during the process of translocation, topogenic sequences containing transmembrane segments of Bcs1p(1–83), DLD(1–72), and full-sized AAC protein were used when constructing modified forms of CYP11A1, and the Su9(1–112) addressing signal was included to stimulate membrane insertion of CYP11A1 after its translocation to the matrix. Alternatively, to promote slippage of the hybrid molecules into the matrix, the hybrid of mCYP11A1 with the precursor of steroidogenic mitochondria matrix protein adrenodoxin (preAd) was designed. The extra sequences used for intramitochondrial sorting of CYP11A1 apparently ensured predicted topology of hybrid molecules in yeast mitochondria. All of the addressing sequences, containing transmembrane domains, provided effective insertion of the hybrid proteins AAC-mCYP11A1, Bcs1p(1–83)-mCYP11A1, DLD(1–72)-mCYP11A1 and Su9(1–116)-mCYP11A1 into the inner membrane. preAd-mCYP11A1 hybrid molecules were shown to be translocated across the inner membrane and tightly associated with the membrane on its matrix side but not membrane inserted. Measuring specific activities of hybrid proteins in the mitochondrial fractions upon addition of Ad and AdR showed that the hybrids predetermined for cotranslocational insertion of CYP11A1 into the inner membrane were more active in the reaction of cholesterol side-chain cleavage than those destined for insertion on the matrix side of the IM, the Ad-mCYP11A1 hybrid demonstrating only residual enzyme activity. The data obtained reinforce the proposal that complete transfer of the polypeptide chain into the matrix is not a necessary stage in its topogenesis, but rather persistent interaction of the polypeptide chain with the membrane during the process of translocation is of importance for heme binding, folding and membrane insertion. © 2008 Elsevier B.V. All rights reserved. 1. Introduction The majority of mitochondrial proteins are encoded within the nuclear genome and synthesized on cytosolic ribosomes, so they must subsequently be imported into the organelle and sorted into one of the four mitochondrial subcompartments: outer membrane, intermem- brane space (IMS), inner membrane and matrix. Targeting of these proteins to mitochondria and sorting processes are mediated by import signals within the preproteins. Several membrane-embedded protein translocases (as well as numerous chaperones and cochaper- ones in the matrix and IMS) provide the import of mitochondrial precursors to different subcompartments [1,2]. Two translocases of the inner membrane are responsible for the import of mitochondrial proteins into the matrix or the inner membrane (TIM complexes — TIM23 and TIM22) [1–5].While multispanning carrier proteins of the inner membrane which possess internal targeting information are integrated into the lipid bilayer by the TIM22 complex, those proteins containing presequences are destined for the mitochondrial matrix or in some cases for the inner membrane and are transported across the inner membrane by the presequence translocase (TIM23 complex) [3– 6]. For some polytopic inner membrane proteins with N-terminal presequences a re-export mechanism was proposed for membrane insertion after the complete translocation into the matrix, with the participation of Oxa1p [7,8]. In the mitochondrial matrix the presequence is proteolytically removed from the precursor by the matrix processing peptidase (MPP) generating the mature form of the imported protein. While both TIM22 and TIM23 complexes use the membrane potential across the inner membrane as a driving force to translocate precursors into or across the inner membrane, complete translocation of presequence-containing precursors into the matrix by the TIM23 complex requires the ATP-powered mitochondrial import motor as a second driving force [1–5]. The main component of this “presequence translocase associated motor” complex (PAM) is the Biochimica et Biophysica Acta 1780 (2008) 1121–1130 Abbreviations: mCYP11A1, mature form of CYP11A1; Ad, adrenodoxin; AdR, adrenodoxin reductase; DLD, D-lactate dehydrogenase; PiC, inorganic phosphate carrier; AAC, adenine nucleotide translocase; PMSF, phenylmethylsulfonyl fluoride; SSQ1p, a thermal shock response protein of the mitochondrial matrix ⁎ Corresponding author. Fax: +74 95 939 3181. E-mail address: [email protected] (I.E. Kovaleva). † Passed away on July 18, 2007. 0304-4165/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbagen.2008.06.006 Contents lists available at ScienceDirect Biochimica et Biophysica Acta journal homepage: www.elsevier.com/locate/bbagen
-
Upload
moscowstate -
Category
Documents
-
view
1 -
download
0
Transcript of Import of hybrid forms of CYP11A1 into yeast mitochondria

Biochimica et Biophysica Acta 1780 (2008) 1121–1130
Contents lists available at ScienceDirect
Biochimica et Biophysica Acta
j ourna l homepage: www.e lsev ie r.com/ locate /bbagen
Import of hybrid forms of CYP11A1 into yeast mitochondria
A.N. Minenko, L.A. Novikova, V.N. Luzikov †, I.E. Kovaleva ⁎Belozersky Institute of Physico-Chemical Biology, Lomonosov Moscow State University, Moscow 119992, Russia
Abbreviations: mCYP11A1, mature form of CYP1adrenodoxin reductase; DLD, D-lactate dehydrogenascarrier; AAC, adenine nucleotide translocase; PMSF, pSSQ1p, a thermal shock response protein of the mitocho⁎ Corresponding author. Fax: +74 95 939 31 81.
E-mail address: [email protected] (I.E. Kova† Passed away on July 18, 2007.
0304-4165/$ – see front matter © 2008 Elsevier B.V. Aldoi:10.1016/j.bbagen.2008.06.006
a b s t r a c t
a r t i c l e i n f oArticle history:
Heterologous expression Received 14 February 2008Received in revised form 5 June 2008Accepted 16 June 2008Available online 21 June 2008Keywords:CYP11A1Topogenic signalYeast mitochondriaMembrane insertion
in yeast of mCYP11A1 fusions with different topogenic signals of yeastmitochondrial proteins for artificial channeling to different translocases of the inner membrane was usedto gain insight in the mechanism of its topogenesis in mitochondria. To ensure insertion of the CYP11A1domain into the inner mitochondrial membrane during the process of translocation, topogenic sequencescontaining transmembrane segments of Bcs1p(1–83), DLD(1–72), and full-sized AAC protein were used whenconstructing modified forms of CYP11A1, and the Su9(1–112) addressing signal was included to stimulatemembrane insertion of CYP11A1 after its translocation to the matrix. Alternatively, to promote slippage of thehybrid molecules into the matrix, the hybrid of mCYP11A1 with the precursor of steroidogenic mitochondriamatrix protein adrenodoxin (preAd) was designed. The extra sequences used for intramitochondrial sortingof CYP11A1 apparently ensured predicted topology of hybrid molecules in yeast mitochondria. All of theaddressing sequences, containing transmembrane domains, provided effective insertion of the hybridproteins AAC-mCYP11A1, Bcs1p(1–83)-mCYP11A1, DLD(1–72)-mCYP11A1 and Su9(1–116)-mCYP11A1 intothe inner membrane. preAd-mCYP11A1 hybrid molecules were shown to be translocated across the innermembrane and tightly associated with the membrane on its matrix side but not membrane inserted.Measuring specific activities of hybrid proteins in the mitochondrial fractions upon addition of Ad and AdRshowed that the hybrids predetermined for cotranslocational insertion of CYP11A1 into the inner membranewere more active in the reaction of cholesterol side-chain cleavage than those destined for insertion on thematrix side of the IM, the Ad-mCYP11A1 hybrid demonstrating only residual enzyme activity. The dataobtained reinforce the proposal that complete transfer of the polypeptide chain into the matrix is not anecessary stage in its topogenesis, but rather persistent interaction of the polypeptide chain with themembrane during the process of translocation is of importance for heme binding, folding and membraneinsertion.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
The majority of mitochondrial proteins are encoded within thenuclear genome and synthesized on cytosolic ribosomes, so theymustsubsequently be imported into the organelle and sorted into one of thefour mitochondrial subcompartments: outer membrane, intermem-brane space (IMS), inner membrane and matrix. Targeting of theseproteins to mitochondria and sorting processes are mediated byimport signals within the preproteins. Several membrane-embeddedprotein translocases (as well as numerous chaperones and cochaper-ones in the matrix and IMS) provide the import of mitochondrialprecursors to different subcompartments [1,2]. Two translocases ofthe inner membrane are responsible for the import of mitochondrial
1A1; Ad, adrenodoxin; AdR,e; PiC, inorganic phosphatehenylmethylsulfonyl fluoride;ndrial matrix
leva).
l rights reserved.
proteins into the matrix or the inner membrane (TIM complexes —
TIM23 and TIM22) [1–5].While multispanning carrier proteins of theinner membrane which possess internal targeting information areintegrated into the lipid bilayer by the TIM22 complex, those proteinscontaining presequences are destined for the mitochondrial matrix orin some cases for the inner membrane and are transported across theinnermembrane by the presequence translocase (TIM23 complex) [3–6]. For some polytopic inner membrane proteins with N-terminalpresequences a re-export mechanism was proposed for membraneinsertion after the complete translocation into the matrix, with theparticipation of Oxa1p [7,8]. In the mitochondrial matrix thepresequence is proteolytically removed from the precursor by thematrix processing peptidase (MPP) generating the mature form ofthe imported protein. While both TIM22 and TIM23 complexes usethe membrane potential across the inner membrane as a driving forceto translocate precursors into or across the innermembrane, completetranslocation of presequence-containing precursors into thematrix bythe TIM23 complex requires the ATP-powered mitochondrial importmotor as a second driving force [1–5]. The main component of this“presequence translocase associated motor” complex (PAM) is the

1122 A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
mtHsp70-a mitochondrial member of the Hsp70 chaperone family.According to current views on the presequence translocase of theinner membrane, distinct forms of TIM23 complex catalyze a matriximport or inner membrane sorting of presequence-containingmitochondrial proteins — a TIM23/PAM supercomplex or a PAM freeform of the TIM23 complex — TIM23SORT [1,2,9], respectively. In thelatter case a hydrophobic stop-transfer sequence usually follows inclose proximity to the presequence.
Cytochrome P450scc (CYP11A1) is a key enzyme of themammaliansteroidogenesis system that transforms cholesterol into pregneno-lone. This protein is located in the inner membrane of adrenocorticalmitochondria with its active center exposed to the matrix [10,11].Similarly to other mitochondrial cytochromes P450, CYP11A1 issynthesized as a precursor with an N-terminal cleavable presequence,and has no predictable transmembrane domains which can play a roleof a stop-transfer sequence [12]. So it can be supposed to be asubstrate for joint action of TIM23 and PAM complexes in a process oftranslocation across the inner membrane to the matrix. But, accordingto the pioneer works on its topology in adrenocortical mitochondria, itis an integral protein of the inner membrane, as it is not extractablefrom the membrane by sodium carbonate treatment [13]. Anotherdistinctive feature of CYP11A1 as a substrate for mitochondrial importconsists in the fact that it is a cytochrome, i.e. a hemoprotein, andtherefore it must bind heme during the process of its topogenesis.Theoretically CYP11A1 may be inserted into the inner membrane inthe process of translocation by the PAM free TIM23SORT complex ormay be translocated into the matrix by the TIM23/PAM supercomplexand subsequently become membrane inserted. In both cases, the“nature” of the sorting signal is unclear.
Previously it was shown that when imported into yeast mitochon-dria CYP11A1 was predominantly mislocalized to the matrix while asmall portion of molecules inserted into membrane became properlyfolded and active [14,15,16]. An attempt to stimulate membranebinding by substitution of the N-terminal fragment of the subunit 9(Su9) of the Fo-ATP synthase (from Neurospora crassa) which mustensure membrane insertion of a polypeptide chain by a re-exportmechanism, after translocation to the matrix [8], for the naturalpresequence, led to membrane insertion of the modified form ofCYP11A1, but its enzymatic activity was low [16]. It was proposed thatrather than translocation to matrix constant binding of the polypep-tide chain to themembrane is of basic importance for holocytochromeformation. To further discriminate between the two possible modes ofCYP11A1 topogenesis, we constructed a set of plasmids, codinghybrids of mCYP11A1 with the targeting sequences of some integralinner membrane proteins of yeast mitochondria, which are cotran-slocationally inserted to the membrane without their transfer to thematrix and utilize different translocases of the inner membrane formembrane insertion. Namely, AAC-mCYP11A1 — a hybrid with thefull-size ATP/ADP carrier of the inner membrane, including 6transmembrane domains that is inserted by TIM22 [1,2,5]; DLD(1–72)-mCYP11A1 [17] — a hybrid with an addressing signal of theintermembrane space protein DLD, consisting of a cleavable prese-quence followed by a transmembrane domain, which arreststranslocation in the N-in–C-out orientation [18] (TIM23SORT [1]); andBcs1p(1–83)-mCYP11A1with an addressing signal of Bcs1p, an innermembrane protein exposed to the matrix, consisting of a transmem-brane domain and an intrinsic signal sequence, leading to the N-out–C-in orientation of the imported protein in the inner membrane [19](the mechanism of translocation is unknown). Alternatively, tostimulate polypeptide chain slippage into the matrix to large extent,a hybrid of mCYP11A1 with the precursor of the matrix protein ofsteroidogenic mitochondria – adrenodoxin – was designed — preAD-mCYP11A1. It was previously shown that the adrenodoxin precursorwith the unusually long (60 amino acids) and highly positivelycharged presequence [20] was successfully imported and processed inmitochondria of nonsteroidogenic tissues [21]. Based on the adreno-
doxin precursor features, it may be suggested that the TIM23/PAMsupercomplex performs its matrix translocation.
The proficiency of holocytochrome formation in hybrid moleculeswith different addressing sequences specific towards different innermembrane translocases was studied. For this aim, the hybrid proteinswere heterologously expressed in yeast Saccharomyces serevisiae andcharacterized in terms of mitochondrial import, processing, mem-brane binding, intramitochondrial localization and cholesterol side-chain activity. The previously described hybrid with the addressingsequence of Su9–Su9(1–116)-mCYP11A1 for realization of a re-exportmechanism of insertion [16] was also included in all the experimentsfor comparison. Since it is rather complicated to interpret the resultswith the CYP11A1 having its own presequence for which multipleprocessing was shown in yeast [16] the hybrid of mCYP11A1 with atypical cleavable matrix presequence of the IV subunit of yeastcytochrome c oxidase COXIV-mCYP11A1 was used as its analog for acontrol. On the whole the behaviour of the native and hybrid proteinswas shown to be similar in yeast mitochondria [16].
Although P450scc is a resident protein ofmammalianmitochondriawe think it reasonable to use yeast heterologous system for studying itstopogenesis for several reasons. Most of our knowledge regarding theprocess of protein import into mitochondria has come from researchemploying fungal systems. Many of the molecular components, firstidentified in fungal species, have nowbeen found in animal species andthe composition of the core translocase components and majorreceptors is essentially the same [22]. And finally one of the mainreasons to choose the yeast system for our experiments was moreextensive knowledge of the import signals (within well-characterizedproteins of yeastmitochondria)mediating the sorting process. Besides,studying of TIM/PAM complexes involvement is available only in yeastat the moment due to the existence of appropriate antibodies andmutants. One of such mutants with defective functions of PAM(pam17Δ) [23] was used in our work to investigate the influence ofthe mutation on the topogenesis of the COXIV-mCYP11A1 hybrid inyeast mitochondria. We propose also that studying of “anomalous”behaviour of the mammalian mitochondrial protein in the mitochon-dria of yeast may help to gain insight in the differences between theyeast and mammalian systems of protein import.
2. Materials and methods
2.1. Chemicals
22R-Hydroxycholesterol, secondary antibodies–horseradish per-oxidase conjugate, glucose-6-phosphate dehydrogenase, cholesteroloxidase, and NADPH were from Sigma (USA); restriction endonu-cleases, the Klenow fragment, T4-DNA ligase, and a reagent kit forelution of DNA were from Fermentas (Latvia); zymolyase 20T wasfrom ICN Biomedicals (USA). Antibodies to adenine nucleotidetranslocase (AAC), a component of the outer membrane translocaseTom40p, inorganic phosphate carrier (PiC), a thermal shock responseprotein of the mitochondrial matrix (SSQ1p), D-lactate dehydrogenase(DLD), and the soluble domain of DLD were generous gift from Prof. V.Neupert (Institute of Physiological Chemistry, Munich, Germany).Cytochrome P450scc, adrenodoxin reductase (AdR), and adrenodoxin(Ad) from bovine adrenal cortex and antibodies to P450scc were givenby V.M. Shkumatov (Institute of Physico-Chemical Problems, Byelor-ussian State University, Minsk, Belarus). Progesteronewas determinedby enzyme immunoassay (ELISA) using a kit presented by A.G.Pryadko (Institute of Bioorganic Chemistry, Academy of Sciences ofBelarus, Minsk, Belarus).
2.2. Cell strains and plasmids
Escherichia coli cells of the JM109 strain (Promega, USA) and theyeast Saccharomyces cerevisiae strain 2805 [MATa pep4::HIS3, prbl-δ,

1123A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
can1, GAL2, his3δ, ura3-52] from S_K. Ri (Research Institute of GeneEngineering, South Korea) were used. Yeast were cultured at 30 °C inselective SD media (0.67% yeast nitrogen bases, 2% α-D-glucose, 0.1%casamino acids) or in SG media (0.67% yeast nitrogen bases, 2% α-D-galactose, 0.1% casamino acids). Yeast strains YPH499 and theappropriate mutant strain pam17Δ were obtained from Prof. N.Pfanner [23].
Yeast shuttle-vector pYeDP/1–8/2 [24], including a galactose-inducible chimeric promoter GAL10-CYC1 and an autotrophic markerURA3, was used for expression of cDNAs encoding hybrid proteins.cDNAs for hybrid proteins were designed in the plasmid pGEM-4Z orpTrc99(A) and then recloned into the pYeDP/1–8/2 vector.
2.3. Construction of plasmids
Construction of the expression plasmid for hybrid of mCYP11A1with the N-terminal fragment of subunit 9 (Su9) of the Fo-ATPsynthase (from Neurospora crassa) (pYeDP/Su9(1–112)-mCYP11A1)was described previously [16]. The expression plasmid for DLD(1–72)-mCYP11A1 hybrid was constructed on the basis of the intermediateplasmid pGEM-4Z/Su9(1–112)-mCYP11A1 by replacement of the Su9(1–112) coding fragment by the sequence coding DLD(1–72) fragment[17]. cDNA for the AAC-mCYP11A1 hybrid was constructed in a similarway. DNA fragment encoding the sequence for full-sized AAC wasexcised by restriction sites EcoRI and BamHI from the plasmidpGEM4Z/AAC-DHFR (which was a gift from Prof. N. Pfanner) andinserted in frame into the plasmid pGEM4Z/Su9(1–112)-mCYP11A1after detachment of the EcoRI–BamHI fragment encoding the se-quence Su9(1–112). The resulting plasmid pGEM4Z/AAC-mCYP11A1was treated successively with the restrictase EcoRI, Klenow fragment,and KpnI to isolate cDNA for the hybrid protein AAC-mCYP11A1. Thehybrid protein cDNA was ligated with the vector pYeDP, which wassuccessively cleaved with the restrictase BamHI, treated with Klenowfragment, and cleaved with KpnI. For construction of cDNA for Bcs1p(1–83)-mCYP11A1, a DNA fragment coding the first 83 amino acids ofBcs1pwith the site for EcoRI at the 5′-end and the BamHI site at the 3′-end was obtained by PCR using pGEM4Z/DHFR-Bcs1p(1–83) [19] as atemplate with the following primers: 5′-CGGAATTCATGTCGGATAA-GCCGATT-3′ and 5′-GCGGATCCCATTTGTCGGTATAG-3′. The AAC cod-ing region was excised from the pGEM4Z/AAC-mCYP11A1 plasmidand replaced by the PCR product after restriction with EcoRI andBamHI. Recloning of cDNA for Bcs1p(1–83)-mCYP11A1 into pYeDPwas performed by the same procedure as for AAC-mCYP11A1.Plasmids pGEM4Z/DHFR-Bcs1p(1–83) and pGEM4Z/DLD(1–72)-DHFR[18] were given by Prof. W. Neupert. Construction of cDNA for preAd-mCYP11A1hybridwas performed on the basis of the plasmid pTrc99(A)/Ad-mCYP11A1 [25] inwhich the NcoI–SpeI fragment coding for matureform of Ad was replaced by NcoI–SpeI fragment coding for theprecursor form of Ad. The latter was obtained by PCR using thepreAdx-pEcE [26] plasmid as a template and the primers: 5′-CATGCC-ATGGTACCATGGCTGCCGCTGGGG-3′ and 5′-GGACTAGTGCCGTCGGT-GGAGGTCTTGCCCACA-3′ containing restriction sites for NcoI and SpeI,respectively. Nucleotide sequence for preAd-mCYP11A1 was isolatedfrom the pTrc99(A)/preAd-mCYP11A1 by restrictases NcoI and SalIfollowed by treatment with the Klenow fragment and ligated then intothe pYeDP vector after its cleavage in the BamHI site in the polylinkerand treatmentwith the Klenow fragment. To ensure that no errorswereintroduced during the amplification process, the appropriate plasmidconstructs were verified by DNA sequencing.
2.4. Isolation and fractionation of yeast mitochondria
Yeast mitochondria were isolated as described in [27]. For alkalineextraction of mitochondrial proteins, mitochondria were reprecipi-tated at 12,000 rpm for 12 min with an Eppendorf (Germany)centrifuge, suspended in a fresh solution of 100 mM Na2CO3, and
incubated on ice for 30 min. The membrane fraction was precipitatedby centrifugation at 125,000 g for 30 min (SW50; Beckman, USA).Proteins of the supernatant were precipitated with 10% trichloroaceticacid (TCA). Sonication of mitochondria was performed in 10 mM Tris–HCl buffer (pH 7.2) in the presence of 500 mM KCl. Mitoplasts wereprepared as described in [28]. Mitochondria were reprecipitated bycentrifugation and resuspended in buffer SHKCl containing 0.6 Msorbitol, 50 mM HEPES-KOH (pH 7.4), and 80 mM KCl. An aliquot ofmitochondria was subjected to osmotic shock with nine volumes of20 mM HEPES-KOH (pH 7.4) followed by incubation on ice for 20 min.Then a fivefold excess volume of buffer SHKCl was added, themitoplasts were centrifuged at 14,000 rpm for 12 min with anEppendorf centrifuge, and the resulting precipitate was resuspendedin SHKCl buffer. The mitochondria and mitoplasts were treated on icewith proteinase K (50 μg/ml) in SHKCl buffer for 20 min. Proteinasewas inactivated by incubation of the preparations with 1mMPMSF for10 min. Sonication of mitochondria was performed in 10 mM Tris–HClbuffer (pH 7.2) in the presence of 500 mM KCl.
Trypsin (100 μg/ml) was added to mitoplasts suspended in SHKLbuffer and incubationwas carried out at 15 °C for 60 min. The reactionwas terminated by addition of soybean trypsin inhibitor.
2.5. The experiments with E. coli cells
Homogenates of the E. coli cells transformed with the plas-mid pTrc99(A)/Ad-mCYP11A1 were prepared as it was describedpreviously [25].
2.6. Enzyme assay
Cholesterol side-chain cleavage activities of hybrid variants ofCYP11A1 were assayed in the reconstituted system as describedpreviously [16] with minor modifications. Mitoplasts were preincu-bated in SHKL buffer supplemented with 0.3% Emulgen 913 during30 min on ice, then ninefold excess of SHKL was added. The reactionmixture for determination of the cholesterol side-chain cleavageactivity contained 200–300 μg mitochondrial protein, purified Ad(0.5 nmol), AdR (0.5 nmol), and 22R-hydroxycholesterol (25 nmol).The reactionwas started by addition of a NADPH-regenerating systemand conducted for 10 min at 37 °C and then the procedure wasperformed according to the protocol presented in [16]. Pregnenolonewas quantitatively converted to progesterone by cholesterol oxidase.The content of progesterone was determined with an ELISA testsystem based on antiprogesterone antibodies.
2.7. Other methods
SDS-PAGE and immunoblotting were performed using standardprocedures. To determine the contents of recombinant CYP11A1versions in mitochondria, 30 μg/lane of mitochondrial fractions wasseparated by 10% PAGE, blotted onto nitrocellulose membrane anddecorated with polyclonal antibodies to CYP11A1 and secondaryhorseradish peroxidase-conjugated anti-rabbit IgG. The densities ofbands after immunodetection were estimated by programs ScnImageand GelScan V5.1. Purified bovine CYP11A1 was used as a calibrationstandard. To control that the samples were evenly loaded blots weresubsequently reprobed with anti-PiC antibodies.
3. Results
3.1. Expression of hybrid CYP11A1 variants in yeast cells
To functionally characterize the CYP11A1 variants imported intomitochondria with different addressing sequences, hybrid proteinswere expressed in yeast Saccharomyces cerevisiae. Yeast cells of S.cerevisiae 2805 strain were transformed with the plasmids designed
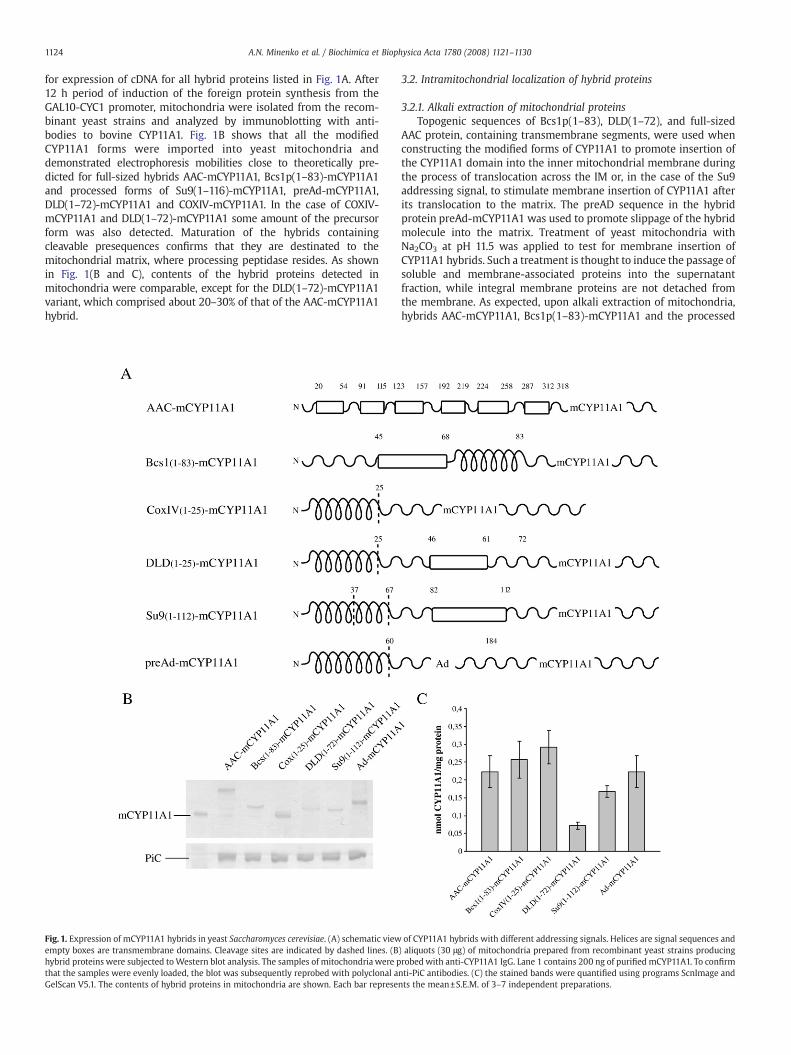
1124 A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
for expression of cDNA for all hybrid proteins listed in Fig. 1A. After12 h period of induction of the foreign protein synthesis from theGAL10-CYC1 promoter, mitochondria were isolated from the recom-binant yeast strains and analyzed by immunoblotting with anti-bodies to bovine CYP11A1. Fig. 1B shows that all the modifiedCYP11A1 forms were imported into yeast mitochondria anddemonstrated electrophoresis mobilities close to theoretically pre-dicted for full-sized hybrids AAC-mCYP11A1, Bcs1p(1–83)-mCYP11A1and processed forms of Su9(1–116)-mCYP11A1, preAd-mCYP11A1,DLD(1–72)-mCYP11A1 and COXIV-mCYP11A1. In the case of COXIV-mCYP11A1 and DLD(1–72)-mCYP11A1 some amount of the precursorform was also detected. Maturation of the hybrids containingcleavable presequences confirms that they are destinated to themitochondrial matrix, where processing peptidase resides. As shownin Fig. 1(B and C), contents of the hybrid proteins detected inmitochondria were comparable, except for the DLD(1–72)-mCYP11A1variant, which comprised about 20–30% of that of the AAC-mCYP11A1hybrid.
Fig. 1. Expression of mCYP11A1 hybrids in yeast Saccharomyces cerevisiae. (A) schematic viewempty boxes are transmembrane domains. Cleavage sites are indicated by dashed lines. (Bhybrid proteins were subjected toWestern blot analysis. The samples of mitochondria were pthat the samples were evenly loaded, the blot was subsequently reprobed with polyclonal aGelScan V5.1. The contents of hybrid proteins in mitochondria are shown. Each bar represe
3.2. Intramitochondrial localization of hybrid proteins
3.2.1. Alkali extraction of mitochondrial proteinsTopogenic sequences of Bcs1p(1–83), DLD(1–72), and full-sized
AAC protein, containing transmembrane segments, were used whenconstructing the modified forms of CYP11A1 to promote insertion ofthe CYP11A1 domain into the inner mitochondrial membrane duringthe process of translocation across the IM or, in the case of the Su9addressing signal, to stimulate membrane insertion of CYP11A1 afterits translocation to the matrix. The preAD sequence in the hybridprotein preAd-mCYP11A1 was used to promote slippage of the hybridmolecule into the matrix. Treatment of yeast mitochondria withNa2CO3 at pH 11.5 was applied to test for membrane insertion ofCYP11A1 hybrids. Such a treatment is thought to induce the passage ofsoluble and membrane-associated proteins into the supernatantfraction, while integral membrane proteins are not detached fromthe membrane. As expected, upon alkali extraction of mitochondria,hybrids AAC-mCYP11A1, Bcs1p(1–83)-mCYP11A1 and the processed
of CYP11A1 hybrids with different addressing signals. Helices are signal sequences and) aliquots (30 μg) of mitochondria prepared from recombinant yeast strains producingrobed with anti-CYP11A1 IgG. Lane 1 contains 200 ng of purified mCYP11A1. To confirmnti-PiC antibodies. (C) the stained bands were quantified using programs ScnImage andnts the mean±S.E.M. of 3–7 independent preparations.

1125A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
forms of DLD(1–72)-mCYP11A1 and Su9(1–116)-mCYP1A1 werefound in the membrane pellet like the integral protein PiC, whichindicates that they became integral membrane proteins after importinto mitochondria (Fig. 2A). In the case of CoxIV(1–25)-mCYP11A1 andpreAd-mCYP11A1, topogenic sequences used for import into mito-chondria did not contain targeting information for membraneinsertion (i.e. transmembrane domains). So membrane insertion ofthese proteins could be achieved only by means of an endogenoustopogenic signal to stop-transfer or of a signal of reinsertion from thematrix in the interior of the mCYP11A1 domain moiety. The datapointing to the aggregation of CYP11A1 in the matrix after processingof CoxIV-mCYP11A1 hybrid [16] make the method of alkali extractionto be noninformative for this protein (Fig. 2A).
But when alkali extraction procedure was applied to Ad-mCYP11A1containing mitochondria, this hybrid protein was found exclusively inthe fraction of the supernatant (Fig. 2B). But at the same time it wasunextractable from membranes by high salt (Fig. 2B). So the Ad-mCYP11A1 hybrid appears not to be an integral membrane protein,but rather to be tightly associated with the inner membrane in a
Fig. 2. Fractionation of mitochondria containing the hybrid proteins by alkalineextraction. (A) after carbonate treatment of mitochondria proteins the precipitate andsoluble fraction were analyzed by SDS-PAGE followed by immunoblotting. The filterswere stained with polyclonal antibodies to CYP11A1 and with polyclonal antibodies toyeast mitochondria proteins: SSQ1 (matrix), phosphate carrier PiC (an integral proteinof the inner membrane). Lanes: T — lysate of mitochondria; P — precipitate; S —
supernatant. (B) study of the interaction of Ad-mCYP11A1 with inner mitochondriamembrane. Mitochondria with Ad-mCYP11A1 were subjected to carbonate extraction(left panel) or were sonicated in 10mM Tris–HCl buffer (pH 7.2) containing 500mMKCl.The sample was separated into the pellet (P) and the supernatant (S) by centrifugationat 125,000 g for 30 min. The filter was stained with antiserum to CYP11A1 or withantibodies to SSQ1and PiC (right panel).
peripheral manner, resembling in such experiments the behavior ofTIM44, which is held to be peripherally associated protein of the innermembrane [29]. These data are in contradiction with those obtainedfor a hybrid of the mature form of Ad with mCYP11A1 expressed in E.coli cells [25], where a substantial portion of hybrid molecules wasshown to be carbonate unextractable. According to our data, it hasappeared that either CYP11A1 does not have an endogenous stop-transfer signal, or in the artificial hybrid molecule it is too far from thesignal sequence to be effectively functioning [30]. The data alsoindicate that CYP11A1 cannot be inserted properly into the membraneafter translocation to the matrix.
3.2.2. Limited proteolysis of mitochondrial fractions with proteinase KProtease protection experiments on swollen mitochondria (mito-
plasts) were performed to determine the intramitochondrial topologyof hybrid CYP11A1 variants after import into mitochondria withdifferent topogenic signals. The behavior of the intermembrane spacemarker protein—DLD and thematrix marker protein—Hep1was alsofollowed in these experiments. Under the experimental conditions oflimited proteolysis when the inner membrane remains still intact, theCYP11A1 domain in modified variants appeared to have limitedsensitivity to protease action, which could be due to its autonomousfolding in hybrid proteins after heme attachment. Limited proteolysiswith proteinase K of mitoplasts containing the Bcs1p-mCYP11A1hybrid resulted in a degradation of a fragment, consisting of about 45amino acids, which correlated in size with the part of the Bcs1ptopogenic signal exposed to the intermembrane space [19]. The bulkof the polypeptide chain remained to be protected in mitoplasts butwas degraded after membrane lysis with detergent, when someamount of CYP11A1 domain also appeared (Fig. 3A). Principally Bcs1p-mCYP11A1 behavior in protease protection experiments largelyresembled that for native Bcs1p where the bulk of the proteinfollowing the internal signal sequence is proposed to be exposed tothe matrix [19]. The processed form of the preAd-mCYP11A1 hybrid(Ad-mCYP11A1) remained completely intact after the proteasetreatment of mitoplasts and underwent proteolytic degradationwhen the detergent was added (Fig. 3B). Taking into accountprocessing of preAd-mCYP11A1 in mitochondria by matrix processingpeptidase and the fact that adrenodoxin is a soluble matrix protein[31], preAd-mCYP11A1 is most likely to be translocated across theinner membrane and is associated with the inner membrane from itsmatrix side.
The AAC-mCYP11A1 hybrid was accessible for proteinase action inintact mitoplasts, which led to the appearance of a band correspond-ing to mCYP11A1 in size and crossreacting with anti-CYP11A1antibodies (Fig. 3C). These data obviously mean that the mCYP11A1domainwas freed fromAAC as a result of a proteolytic “nick” at the siteof the AAC and mCYP11A1 junction or as a result of AAC moietyfragmentation with proteinase K. mCYP11A1 remained to be asso-ciated with membrane as it was co-sedimented with mitoplasts afterprotease treatment. Protease treatment of intact mitoplasts also led tosome decrease in mCYP11A1 content, which points to its predominantexposure to the intermembrane space.
The protease treatment of DLD(1–72)-mCYP11A1 containingmitoplasts led to the release of the mCYP11A1 domain, which wasanalogous to the release of the soluble DLD domain in native DLD [18]but in contrast to the DLD-soluble domain, themCYP11A1 domainwasassociated with the membrane (Fig. 3D). Thus the hybrid DLD(1–72)-mCYP11A1 is inserted into the inner membrane in such away that it isaccessible for proteinase K from the side of the intermembrane space,similarly to DLD.
Based on the fact that all the above characterized hybridsunderwent proteolysis under proteinase K action, it can be concludedthat they are not aggregated in the mitochondrial milieu.
Less obvious are the results obtained for CoxIV(1–25)-mCYP11A1and Su9(1–116)-mCYP11A1 hybrids. These hybrid proteins were
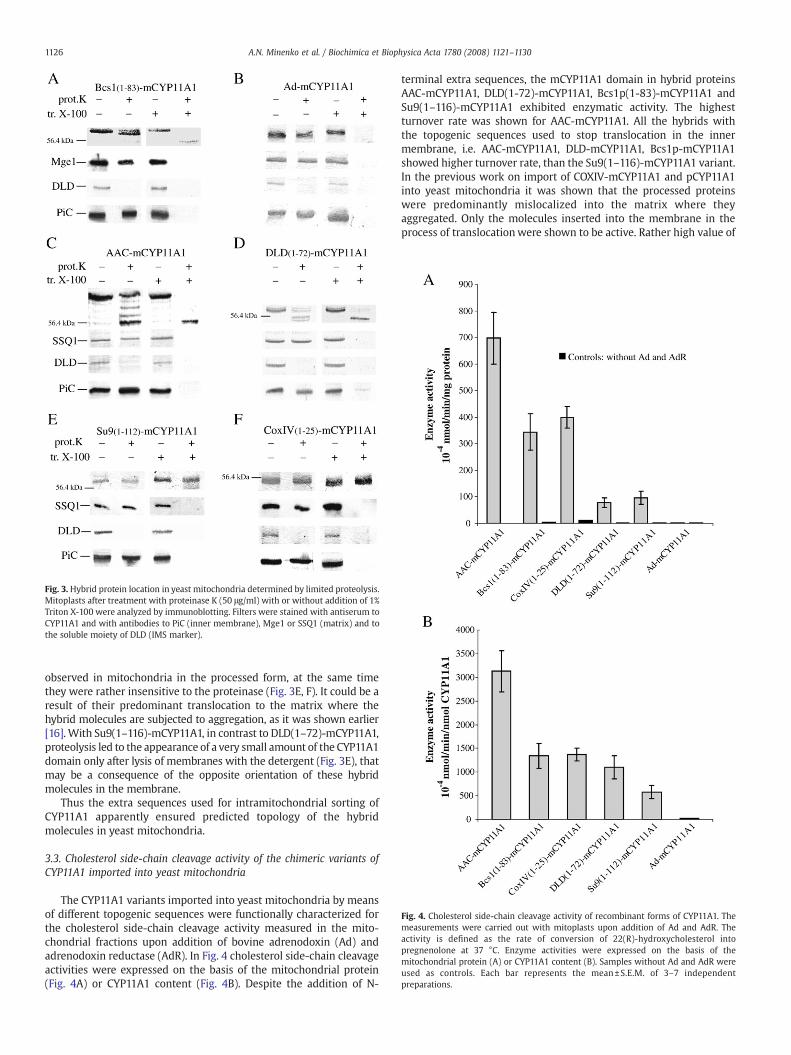
Fig. 4. Cholesterol side-chain cleavage activity of recombinant forms of CYP11A1. Themeasurements were carried out with mitoplasts upon addition of Ad and AdR. Theactivity is defined as the rate of conversion of 22(R)-hydroxycholesterol intopregnenolone at 37 °C. Enzyme activities were expressed on the basis of themitochondrial protein (A) or CYP11A1 content (B). Samples without Ad and AdR wereused as controls. Each bar represents the mean±S.E.M. of 3–7 independentpreparations.
Fig. 3. Hybrid protein location in yeast mitochondria determined by limited proteolysis.Mitoplasts after treatment with proteinase K (50 μg/ml) with or without addition of 1%Triton X-100 were analyzed by immunoblotting. Filters were stained with antiserum toCYP11A1 and with antibodies to PiC (inner membrane), Mge1 or SSQ1 (matrix) and tothe soluble moiety of DLD (IMS marker).
1126 A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
observed in mitochondria in the processed form, at the same timethey were rather insensitive to the proteinase (Fig. 3E, F). It could be aresult of their predominant translocation to the matrix where thehybrid molecules are subjected to aggregation, as it was shown earlier[16]. With Su9(1–116)-mCYP11A1, in contrast to DLD(1–72)-mCYP11A1,proteolysis led to the appearance of a very small amount of the CYP11A1domain only after lysis of membranes with the detergent (Fig. 3E), thatmay be a consequence of the opposite orientation of these hybridmolecules in the membrane.
Thus the extra sequences used for intramitochondrial sorting ofCYP11A1 apparently ensured predicted topology of the hybridmolecules in yeast mitochondria.
3.3. Cholesterol side-chain cleavage activity of the chimeric variants ofCYP11A1 imported into yeast mitochondria
The CYP11A1 variants imported into yeast mitochondria by meansof different topogenic sequences were functionally characterized forthe cholesterol side-chain cleavage activity measured in the mito-chondrial fractions upon addition of bovine adrenodoxin (Ad) andadrenodoxin reductase (AdR). In Fig. 4 cholesterol side-chain cleavageactivities were expressed on the basis of the mitochondrial protein(Fig. 4A) or CYP11A1 content (Fig. 4B). Despite the addition of N-
terminal extra sequences, the mCYP11A1 domain in hybrid proteinsAAC-mCYP11A1, DLD(1-72)-mCYP11A1, Bcs1p(1-83)-mCYP11A1 andSu9(1–116)-mCYP11A1 exhibited enzymatic activity. The highestturnover rate was shown for AAC-mCYP11A1. All the hybrids withthe topogenic sequences used to stop translocation in the innermembrane, i.e. AAC-mCYP11A1, DLD-mCYP11A1, Bcs1p-mCYP11A1showed higher turnover rate, than the Su9(1–116)-mCYP11A1 variant.In the previous work on import of COXIV-mCYP11A1 and pCYP11A1into yeast mitochondria it was shown that the processed proteinswere predominantly mislocalized into the matrix where theyaggregated. Only the molecules inserted into the membrane in theprocess of translocationwere shown to be active. Rather high value of
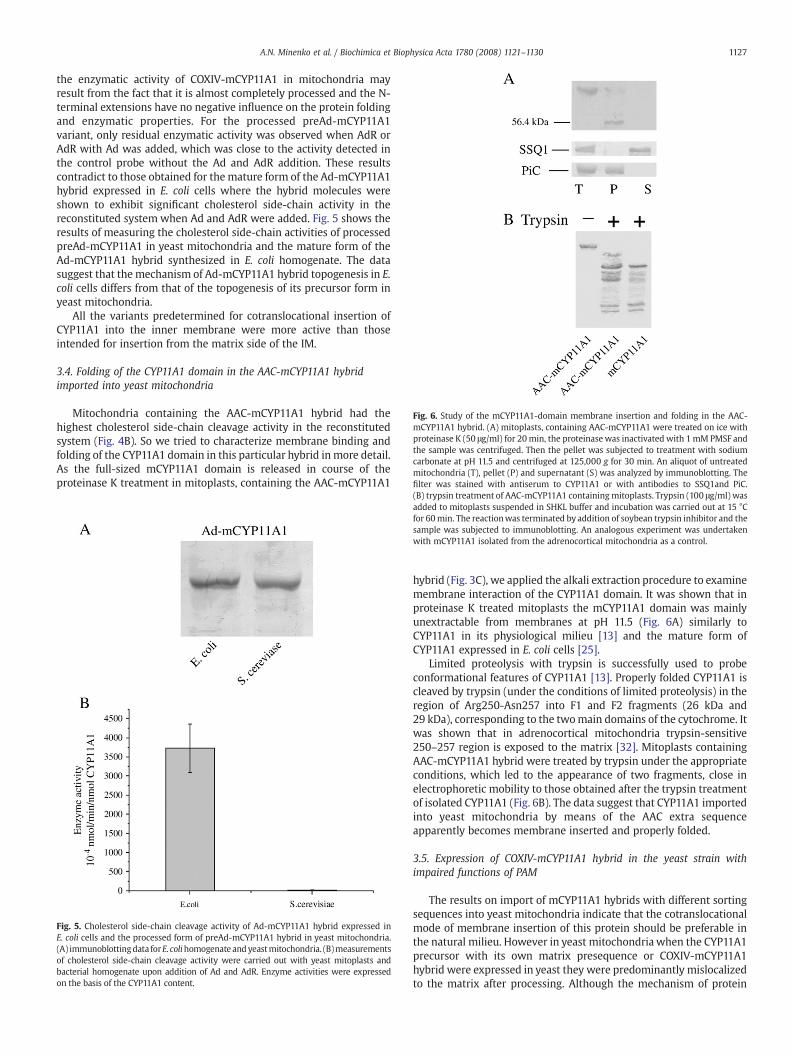
Fig. 6. Study of the mCYP11A1-domain membrane insertion and folding in the AAC-mCYP11A1 hybrid. (A) mitoplasts, containing AAC-mCYP11A1 were treated on ice withproteinase K (50 μg/ml) for 20min, the proteinase was inactivated with 1 mM PMSF andthe sample was centrifuged. Then the pellet was subjected to treatment with sodiumcarbonate at pH 11.5 and centrifuged at 125,000 g for 30 min. An aliquot of untreatedmitochondria (T), pellet (P) and supernatant (S) was analyzed by immunoblotting. Thefilter was stained with antiserum to CYP11A1 or with antibodies to SSQ1and PiC.
1127A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
the enzymatic activity of COXIV-mCYP11A1 in mitochondria mayresult from the fact that it is almost completely processed and the N-terminal extensions have no negative influence on the protein foldingand enzymatic properties. For the processed preAd-mCYP11A1variant, only residual enzymatic activity was observed when AdR orAdR with Ad was added, which was close to the activity detected inthe control probe without the Ad and AdR addition. These resultscontradict to those obtained for the mature form of the Ad-mCYP11A1hybrid expressed in E. coli cells where the hybrid molecules wereshown to exhibit significant cholesterol side-chain activity in thereconstituted system when Ad and AdR were added. Fig. 5 shows theresults of measuring the cholesterol side-chain activities of processedpreAd-mCYP11A1 in yeast mitochondria and the mature form of theAd-mCYP11A1 hybrid synthesized in E. coli homogenate. The datasuggest that the mechanism of Ad-mCYP11A1 hybrid topogenesis in E.coli cells differs from that of the topogenesis of its precursor form inyeast mitochondria.
All the variants predetermined for cotranslocational insertion ofCYP11A1 into the inner membrane were more active than thoseintended for insertion from the matrix side of the IM.
3.4. Folding of the CYP11A1 domain in the AAC-mCYP11A1 hybridimported into yeast mitochondria
Mitochondria containing the AAC-mCYP11A1 hybrid had thehighest cholesterol side-chain cleavage activity in the reconstitutedsystem (Fig. 4B). So we tried to characterize membrane binding andfolding of the CYP11A1 domain in this particular hybrid in more detail.As the full-sized mCYP11A1 domain is released in course of theproteinase K treatment in mitoplasts, containing the AAC-mCYP11A1
Fig. 5. Cholesterol side-chain cleavage activity of Ad-mCYP11A1 hybrid expressed inE. coli cells and the processed form of preAd-mCYP11A1 hybrid in yeast mitochondria.(A) immunoblottingdata forE. colihomogenate andyeastmitochondria. (B)measurementsof cholesterol side-chain cleavage activity were carried out with yeast mitoplasts andbacterial homogenate upon addition of Ad and AdR. Enzyme activities were expressedon the basis of the CYP11A1 content.
(B) trypsin treatment of AAC-mCYP11A1 containing mitoplasts. Trypsin (100 μg/ml) wasadded to mitoplasts suspended in SHKL buffer and incubation was carried out at 15 °Cfor 60min. The reactionwas terminated by addition of soybean trypsin inhibitor and thesample was subjected to immunoblotting. An analogous experiment was undertakenwith mCYP11A1 isolated from the adrenocortical mitochondria as a control.
hybrid (Fig. 3C), we applied the alkali extraction procedure to examinemembrane interaction of the CYP11A1 domain. It was shown that inproteinase K treated mitoplasts the mCYP11A1 domain was mainlyunextractable from membranes at pH 11.5 (Fig. 6A) similarly toCYP11A1 in its physiological milieu [13] and the mature form ofCYP11A1 expressed in E. coli cells [25].
Limited proteolysis with trypsin is successfully used to probeconformational features of CYP11A1 [13]. Properly folded CYP11A1 iscleaved by trypsin (under the conditions of limited proteolysis) in theregion of Arg250-Asn257 into F1 and F2 fragments (26 kDa and29 kDa), corresponding to the twomain domains of the cytochrome. Itwas shown that in adrenocortical mitochondria trypsin-sensitive250–257 region is exposed to the matrix [32]. Mitoplasts containingAAC-mCYP11A1 hybrid were treated by trypsin under the appropriateconditions, which led to the appearance of two fragments, close inelectrophoretic mobility to those obtained after the trypsin treatmentof isolated CYP11A1 (Fig. 6B). The data suggest that CYP11A1 importedinto yeast mitochondria by means of the AAC extra sequenceapparently becomes membrane inserted and properly folded.
3.5. Expression of COXIV-mCYP11A1 hybrid in the yeast strain withimpaired functions of PAM
The results on import of mCYP11A1 hybrids with different sortingsequences into yeast mitochondria indicate that the cotranslocationalmode of membrane insertion of this protein should be preferable inthe natural milieu. However in yeast mitochondria when the CYP11A1precursor with its own matrix presequence or COXIV-mCYP11A1hybrid were expressed in yeast they were predominantly mislocalizedto the matrix after processing. Although the mechanism of protein
1128 A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
import into mitochondria was shown to be highly conserved acrossphylogenetic boundaries some data on differences between yeast andmammalian systems are also available at the moment. Of particularimportance is that while fungal Tim44 (component of PAM) is tightlyassociated with the matrix-face of the inner membrane, mammalianTim44 is largely found in the soluble matrix fraction [22]. In fungi, ithas been proposed that Tim44 and three other membrane proteins,Pam16, Pam17, and Pam18, are required to coordinate the action ofmatrix Hsp70 at the TIM23 complex, i.e., at the exit site for preproteinsat the inner membrane, so that this chaperone can bind and assist inthe translocation of precursor proteins [1–3]. ATP hydrolysis isthought to result in structural rearrangements of Hsp70, leading tothe generation of force and pulling of precursors into thematrix [1–3].Since mammalian Tim44 is largely found in the matrix fraction ofmitochondria, it suggests that mtHsp70 in this systemmay not be ableto perform a pulling function. In the recent time yeast mutants withdefects in PAM function became available. We used one of thedescribed mutants [23] lacking the open reading frame of PAM17, inwhich mitochondria, lacking Pam17, show a characteristic defect ofPAM, i.e., inhibition of matrix import while inner membrane sortingcan take place. It was of interest to express the COXIV-mCYP11A1
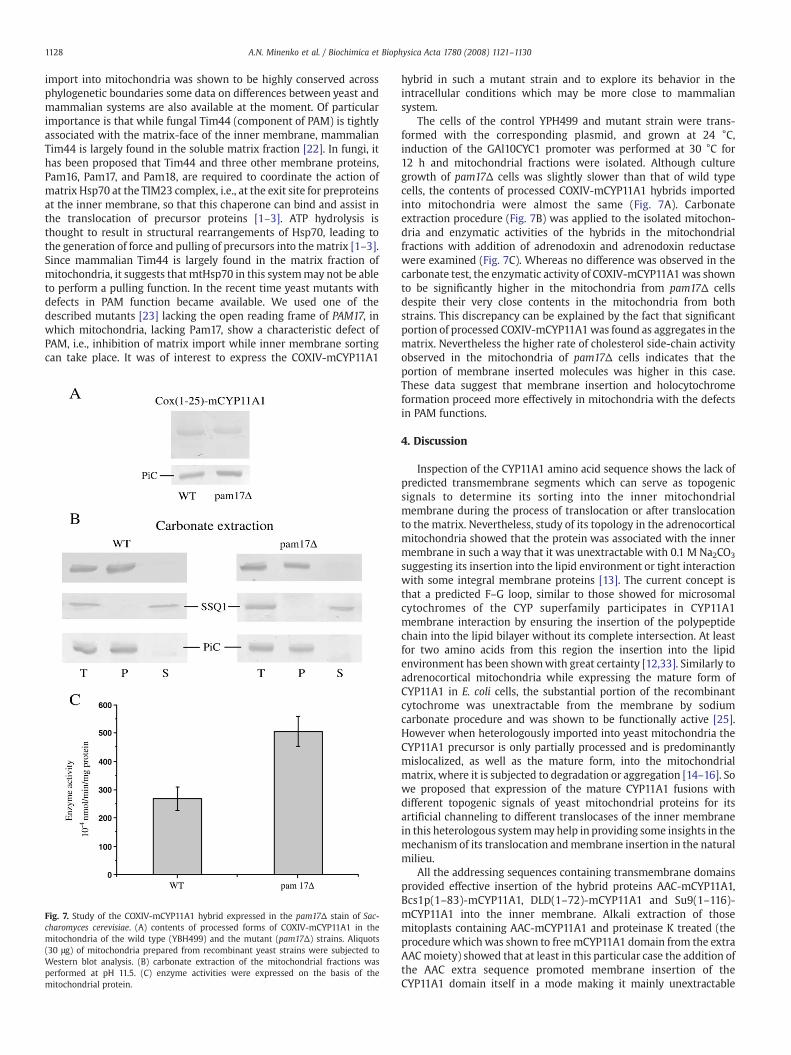
Fig. 7. Study of the COXIV-mCYP11A1 hybrid expressed in the pam17Δ stain of Sac-charomyces cerevisiae. (A) contents of processed forms of COXIV-mCYP11A1 in themitochondria of the wild type (YBH499) and the mutant (pam17Δ) strains. Aliquots(30 μg) of mitochondria prepared from recombinant yeast strains were subjected toWestern blot analysis. (B) carbonate extraction of the mitochondrial fractions wasperformed at pH 11.5. (C) enzyme activities were expressed on the basis of themitochondrial protein.
hybrid in such a mutant strain and to explore its behavior in theintracellular conditions which may be more close to mammaliansystem.
The cells of the control YPH499 and mutant strain were trans-formed with the corresponding plasmid, and grown at 24 °C,induction of the GAl10CYC1 promoter was performed at 30 °C for12 h and mitochondrial fractions were isolated. Although culturegrowth of pam17Δ cells was slightly slower than that of wild typecells, the contents of processed COXIV-mCYP11A1 hybrids importedinto mitochondria were almost the same (Fig. 7A). Carbonateextraction procedure (Fig. 7B) was applied to the isolated mitochon-dria and enzymatic activities of the hybrids in the mitochondrialfractions with addition of adrenodoxin and adrenodoxin reductasewere examined (Fig. 7C). Whereas no difference was observed in thecarbonate test, the enzymatic activity of COXIV-mCYP11A1was shownto be significantly higher in the mitochondria from pam17Δ cellsdespite their very close contents in the mitochondria from bothstrains. This discrepancy can be explained by the fact that significantportion of processed COXIV-mCYP11A1was found as aggregates in thematrix. Nevertheless the higher rate of cholesterol side-chain activityobserved in the mitochondria of pam17Δ cells indicates that theportion of membrane inserted molecules was higher in this case.These data suggest that membrane insertion and holocytochromeformation proceed more effectively in mitochondria with the defectsin PAM functions.
4. Discussion
Inspection of the CYP11A1 amino acid sequence shows the lack ofpredicted transmembrane segments which can serve as topogenicsignals to determine its sorting into the inner mitochondrialmembrane during the process of translocation or after translocationto the matrix. Nevertheless, study of its topology in the adrenocorticalmitochondria showed that the protein was associated with the innermembrane in such a way that it was unextractable with 0.1 M Na2CO3
suggesting its insertion into the lipid environment or tight interactionwith some integral membrane proteins [13]. The current concept isthat a predicted F–G loop, similar to those showed for microsomalcytochromes of the CYP superfamily participates in CYP11A1membrane interaction by ensuring the insertion of the polypeptidechain into the lipid bilayer without its complete intersection. At leastfor two amino acids from this region the insertion into the lipidenvironment has been shownwith great certainty [12,33]. Similarly toadrenocortical mitochondria while expressing the mature form ofCYP11A1 in E. coli cells, the substantial portion of the recombinantcytochrome was unextractable from the membrane by sodiumcarbonate procedure and was shown to be functionally active [25].However when heterologously imported into yeast mitochondria theCYP11A1 precursor is only partially processed and is predominantlymislocalized, as well as the mature form, into the mitochondrialmatrix, where it is subjected to degradation or aggregation [14–16]. Sowe proposed that expression of the mature CYP11A1 fusions withdifferent topogenic signals of yeast mitochondrial proteins for itsartificial channeling to different translocases of the inner membranein this heterologous systemmay help in providing some insights in themechanism of its translocation andmembrane insertion in the naturalmilieu.
All the addressing sequences containing transmembrane domainsprovided effective insertion of the hybrid proteins AAC-mCYP11A1,Bcs1p(1–83)-mCYP11A1, DLD(1–72)-mCYP11A1 and Su9(1–116)-mCYP11A1 into the inner membrane. Alkali extraction of thosemitoplasts containing AAC-mCYP11A1 and proteinase K treated (theprocedure which was shown to free mCYP11A1 domain from the extraAACmoiety) showed that at least in this particular case the addition ofthe AAC extra sequence promoted membrane insertion of theCYP11A1 domain itself in a mode making it mainly unextractable

1129A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
with Na2CO3 (Fig. 5A), as it was shown for CYP11A1 in its physiologicalmilieu [13]. By contrast, when a precursor of a soluble matrix proteinwas used for import and intramitochondrial sorting, the hybridprotein preAd-mCYP11A1, after import and processing, was recoveredexclusively in the supernatant fraction after alkali treatment ofmitochondria and subsequent high speed centrifugation (Fig. 2B),while the binding of the hybrid protein (Ad-mCYP11A1) to mem-branes could not be reversed by high salt (Fig. 2B) indicating tightinteraction with the membrane offered most likely by the CYP11A1domain [34]. Thus the behaviour of Ad-mCYP11A1 in the fractionationexperiments closely resembles those of TIM44, which is held to beperipherally associated protein of the inner membrane. Taking intoaccount processing of preAd-mCYP11A1 in mitochondria by thematrix processing peptidase together with the results of proteaseprotection experiments (Fig. 3B), preAd-mCYP11A1 is most likely to betranslocated across the innermembrane and associatedwith the innermembrane from its matrix side. That means that the CYP11A1 matureform does not contain its own endogenous signal capable to stoptranslocation across the inner membrane, or if such a signal exists, it islocated too far from the presequence in the hybrid molecule toeffectively function [28]. It can also be concluded that aftertranslocation to the matrix, the CYP11A1 domain cannot be properlyinserted into the inner membrane.
Thus artificial “confinement” of CYP11A1 in the membrane media-ted its membrane insertion, whereas when CYP11A1 is imported intomitochondria by means of the matrix protein precursor protein(preAd-mCYP11A1 hybrid) which is obviously a substrate of theTIM23/PAM supercomplex, the proper membrane insertion of theCYP11A1 domain could not be achieved.
The comparing of the values of specific activities of hybrid proteinsmeasured in the mitochondrial fractions upon addition of Ad and AdRshowed that the hybrids containing signals for arrest of translocation,i.e. AAC-mCYP11A1, Bcs1p(1–83)-mCYP11A1, DLD(1–72)-mCYP11A1,were more active in the reaction of cholesterol side-chain cleavagethan Su9(1–116)-mCYP11A1 which is proposed to be inserted into theinner mitochondrial membrane by the re-export mechanism. Ad-mCYP11A1 hybrid demonstrated only residual enzymatic activitywhen AdR or AdR with Ad was added (Fig. 4B). The data obtainedreinforce the proposal that complete transfer of the polypeptide chaininto the matrix is not a necessary stage in its topogenesis, but ratherpersistent interaction of the polypeptide chain with the membrane iscrucial for heme binding, folding and membrane insertion. In thisconnection it should be noted that upon expression of Ad-mCYP11A1(hybrid of mature adrenodoxin with CYP11A1) in E. coli cells asignificant portion of hybrid molecules became membrane insertedand exhibited cholesterol side-chain cleavage activity [25]. Thediscrepancy between the results obtained in the two expressionsystems shows that the process of Ad-mCYP11A1 topogenesis in E. colidiffers from those for its precursor form in yeast mitochondria.
Constant interaction with membrane during the process ofholocytochrome formation may be essential for heme binding, thelast stage of which is known to be performed at the matrix side of theinner mitochondrial membrane [35]. It could be proposed that uponimport into steroidigenic tissues mitochondria CYP11A1 is recognizedas a substrate for the PAM free TIM23SORT complex, thus facilitating itsmembrane insertion. An endogenous signal for TIM23SORT channelingcould be heme attachment to CYP11A1 in the membrane. Alterna-tively, the current views on the monotopic binding of CYP11A1 to themembrane implying that the bulk of the protein globule is exposed tothe matrix [36] allow us to speculate that the PAM motor participatesin polypeptide chain protruding across the inner membrane.
However since mammalian Tim44 is largely found in the matrixfraction of mitochondria, mtHsp70 in this system may not be able toperform a pulling function. So a more passive model of Hsp70 actionsuggesting its binding to the precursor and thus preventing itsretrograde movement may be realized in more extent in mammalian
mitochondria. Our results with the COXIV-mCYP11A1 hybridexpressed in pam17Δ mitochondria are in agreement with such aproposal.
Acknowledgements
The authors are grateful to Dr. E.A. Kotova for correcting themanuscript and to A.A. Vinogradova for the helpful advices andassistance in some experiments. This workwas supported by the RFBRgrant 02-04-48204 and INTAS 03-1-4366.
References
[1] M. Bohnert, N. Pfanner, M. van der Laan, A dynamic machinery for import ofmitochondrial precursor protein. FEBS Letters 581 (2007) 2802–2810.
[2] M. van der Laan, M. Rissier, P. Rehling, Mitochondrial preprotein translocases asdynamic molecular machines, FEMS Yeast Res. 6 (2006) 849–861.
[3] R.E. Jensen, C.D. Dunn, Protein import into and across the mitochondrial innermembrane: role of the TIM23 and TIM22 translocons, Biochim. Biophys. Acta 1592(2002) 25–34.
[4] P. Rehling, K. Brandner, N. Pfanner, Mitochondrial import and the twin-poretranslocase, Nat. Rev. Mol. Cell Biol. 5 (2004) 519–530.
[5] W. Neupert, J.M. Herrmann, Translocation of proteins into mitochondria, Annu.Rev. Biochem. 76 (2007) 723–749.
[6] T. Endo, H. Yamamoto, M. Esaki, Functional cooperation and separation oftranslocators in protein import into mitochondria, the double-membranebounded organelles, J. Cell Sci. 116 (2003) 3259–3267.
[7] J.M. Herrmann, W. Neupert, R.A. Stuart, Insertion into the mitochondrial innermembrane of a polytopic protein, the nuclear encoded Oxa1p, EMBO J. 16 (1997)2217–2226.
[8] E.E. Rojo, B. Guiard, W. Neupert, R.A. Stuart, N-terminal tail export from themitochondrial matrix, J. Biol. Chem. 274 (1999) 19617–19622.
[9] A. Chacinska, M. Lind, A.E. Frazier, Mitochondrial presequence translocase:switching between TOM tethering and motor recruitment involves Tim21 andTim17, Cell 120 (2005) 817–829.
[10] P.F. Churchill, L.R. deAlvero, T. Kimura, Topological studies of steroid hydroxylasecomplexes in bovine adrenocortical mitochondria, J. Biol. Chem. 251 (1978)4924–4929.
[11] P.F. Churchill, T. Kimura, Topological studies of cytochromes P-450scc and P-45011beta in bovine adrenocortical inner mitochondrial membranes. Effects ofcontrolled tryptic digestion, J. Biochem. 254 (1979) 10443–10448.
[12] M.J. Headlam, M.C. Wilce, R.C. Tuckey, The F–G loop region of cytochrome P450scc(CYP11A1) interacts with the phospholipid membrane, Biochim. Biophys. Acta1617 (2003) 96–108.
[13] W. Ou, A. Ito, K. Morohashi, Y. Fujii_Kuriyama, T. Omura, Processing-independentin vitro translocation of cytochrome P-450(SCC) precursor across mitochondrialmembranes.
[14] A.S. Savel'ev, L.A. Novikova, V.L. Drutsa, L.V. Isaeva, A.A. Chernogolov, S.A. Usanov,V.N. Luzikov, Synthesis and some aspects of topogenesis of bovine cytochromeP450scc in yeast, Biochemistry (Moscow) 62 (1997) 779–786.
[15] A.S. Savel'ev, L.A. Novikova, I.E. Kovaleva, V.N. Luzikov, W. Neupert, T. Langer,P-dependent proteolysis in mitochondria. m-AAA protease and PIM1 proteaseexert overlapping substrate specificities and cooperate with the mtHsp70 system,J. Biol. Chem. 273 (1998) 20596–20602.
[16] I.E. Kovaleva, L.A. Novikova, P.A. Nazarov, S.I. Grivennikov, V.N. Luzikov, Effects ofvarious N-terminal addressing signals on sorting and folding of mammalianCYP11A1 in yeast mitochondria, Eur. J. Biochem. 270 (2003) 222–229.
[17] A.N. Minenko, V.N. Luzikov, I.E. Kovaleva, Use of the addressing sequence of yeastD-lactate dehydrogenase for insertion of CYP11A1 into the inner membrane ofyeast mitochondria, Biochemistry (Moscow) 71 (2006) 32–38.
[18] E.E. Rojo, B. Guiard, W. Neupert, R.A. Stuart, Sorting of D-lactate dehydrogenase tothe inner membrane of mitochondria. Analysis of topogenic signal and energeticrequirements, J. Biol. Chem. 273 (1998) 8040–8047.
[19] H. Folsch, B. Guiard, W. Neupert, R.A. Stuart, Internal targeting signal of the Bcs1protein: a novel mechanism of import into mitochondria, J. Mol. Biol. 15 (1996)479–487.
[20] T. Okamura, M.E. John, M.X. Zuber, E.R. Simson, M.R. Waterman, Molecular cloningand amino acid sequence of the precursor form of bovine adrenodoxin: evidencefor a previously unidentified COOH-terminal peptide, Proc. Natl. Acad. Sci. 82(1985) 5705–5709.
[21] T. Kumamoto, A. Ito, T. Omura, Characterization of a mitochondrial matrixprotease catalyzing the processing of adrenodoxin precursor, J. Biochem. 100(1986) 247–254.
[22] N.J. Hoogenraad, L.A. Ward, M.T. Ryan, Import and assembly of proteins intomitochondria of mammalian cells, Biochim. Biophys. Acta 1592 (2002) 97–105.
[23] M. van der Laan, A. Chacinska, M. Lind, I. Perschil, A. Sickmann, H.E. Meyer, B.Guiard, C. Meisinger, N. Pfanner, P. Rehling, Pam17 is required for architecture andtranslocation activity of the mitochondrial protein import motor, Mol. Cell Biol. 25(2005) 7449–7458.
[24] C. Cullin, D. Pompon, Synthesis of functional mouse cytochromes P-450 P1 andchimeric P-450 P3-1 in the yeast Saccharomyces cerevisiae, Gene 65 (1988)203–217.

1130 A.N. Minenko et al. / Biochimica et Biophysica Acta 1780 (2008) 1121–1130
[25] A.A. Vinogradova, V.N. Luzikov, L.A. Novikova, Comparative study of topogenesisof cytochrome P450scc (CYP11A1) and its hybrids with adrenodoxin expressed inEscherichia coli cells, Biochemistry (Moscow) 72 (2007) 247–254.
[26] J. Picado-Leonard, R. Voutilainen, L.C. Kao, B.C. Chung, J.F. Strauss III, W.L. Miller,Human adrenodoxin: cloning of three cDNAs and cycloheximide enhancement inJEG-3 cells, J. Biol. Chem. 263 (1988) 240–3244.
[27] G. Daum, P.C. Buhni, G. Schatz, Import of proteins into mitochondria. Cytochromeb2 and cytochrome c peroxidase are located in the intermembrane space of yeastmitochondria, J. Biol. Chem. 257 (1982) 13028–13033.
[28] A. Geissler, T. Krimmer, U. Bumer, B. Guiard, J. Rassow, N. Pfanner, Membranepotential-driven protein import into mitochondria. The sorting sequence ofcytochrome b(2) modulates the deltapsi-dependence of translocation of thematrix-targeting sequence, Mol. Biol. Cell 11 (2000) 3977–3991.
[29] D. Milenkovic, V. Kozjak, N. Wiedemann, C. Lohaus, H.E. Meyer, B. Guiard, N.Pfanner, C. Meisinger, Sam35 of the mitochondrial protein sorting and assemblymachinery is a peripheral outer membrane protein essential for cell viability,J. Biol. Chem. 279 (2004) 22781.
[30] H.M. McBride, I.S. Goping, G.C. Shore, The human mitochondrial import receptor,hTom20p, prevents a cryptic matrix targeting sequence from gaining access to theprotein translocation machinery, J. Cell. Biol. 134 (1996) 307–313.
[31] N.R. Orme-Johnson, Distinctive properties of adrenal cortex mitochondria,Biochim. Biophys. Acta 1020 (1990) 213–231.
[32] S.A. Usanov, A.A. Chernogolov, V.L. Chashchin, Immunochemical studyof cholesterol-hydroxylating cytochrome P450. Topology of the peptide chain of the heme proteinin the phospholipid membrane, Biochemistry (Moscow) 54 (1989) 916–925.
[33] I. Pikuleva, Putative F–G loop is involved in association with the membrane inP450scc (P450 11A1), Mol. Cell. Endocrinol. 215 (2004) 161–164.
[34] D. Schwarz, V. Kruger, A.A. Chernogolov, S.A. Usanov, A. Stier, Rotation ofcytochrome P450SCC (CYP11A1) in proteoliposomes studied by delayed fluores-cence depolarization, Biochem. Biophys. Res. Commun. 195 (1993) 889–896.
[35] H.A. Dailey, Terminal steps of haem biosynthesis, Biochem. Soc. Trans. 30 (2002)590–595.
[36] K.H. Storbeck, P. Swart, A.C. Swart, Cytochrome P450 side-chain cleavage: insightsgained from homology modeling, Mol. Cell Endocrinol. 265–266 (2007) 65–70.




















