
Lamprey parasitism of sharks and teleosts: high capacity urea excretion in an extant vertebrate...
29
Review Copy 1 Lamprey Parasitism of Sharks and Teleosts: High Capacity Urea Excretion and Ornithine Urea Cycle Activity in an Extant Vertebrate Relic Michael P. Wilkie *1 , Steven Turnbull 2 , Jonathan Bird 3 , Yuxiang S. Wang 4 , Jaime F. Claude 1 , and John H. Youson 5 1. Department of Biology, Mount Allison University, Sackville, New Brunswick, E4L 1E4. 2. Department of Biology, University of New Brunswick Saint John, Saint John, New Brunswick, E2L 4L5. 3. Oceanic Research Group, 59 Old Andover Road, North Reading, Massachussets, 01864. 4. Department of Biology, Queen’s University, Kingston, Ontario, K7L 3N6. 5. Department of Life Sciences, University of Toronto at Scarborough, Toronto, Ontario, M1C 1A4. * To whom all correspondence should be addressed. Present address: Department of Biology, McMaster University, 1280 Main Street West, Hamilton, Ontario, L8S 4K1. Phone: 905 525-9140 ext. 27257. FAX: 905 522-6066 Email: [email protected] Running Title: Nitrogen Excretion by Parasitic Lampreys 1 of 29 Monday , February 23, 2004 Elsevier
Transcript of Lamprey parasitism of sharks and teleosts: high capacity urea excretion in an extant vertebrate...

Revie
w C
opy
1
Lamprey Parasitism of Sharks and Teleosts: High Capacity Urea Excretion and Ornithine Urea Cycle Activity in an Extant Vertebrate Relic
Michael P. Wilkie*1, Steven Turnbull2, Jonathan Bird3, Yuxiang S. Wang4, Jaime F. Claude1, and
John H. Youson5
1. Department of Biology, Mount Allison University, Sackville, New Brunswick, E4L 1E4.
2. Department of Biology, University of New Brunswick Saint John, Saint John, New
Brunswick, E2L 4L5.
3. Oceanic Research Group, 59 Old Andover Road, North Reading, Massachussets, 01864.
4. Department of Biology, Queen’s University, Kingston, Ontario, K7L 3N6.
5. Department of Life Sciences, University of Toronto at Scarborough, Toronto, Ontario,
M1C 1A4.
* To whom all correspondence should be addressed.
Present address: Department of Biology, McMaster University, 1280 Main Street West,
Hamilton, Ontario, L8S 4K1.
Phone: 905 525-9140 ext. 27257. FAX: 905 522-6066
Email: [email protected]
Running Title: Nitrogen Excretion by Parasitic Lampreys
1 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
2
ABSTRACT
Although phylogenetically ancient sea lampreys (Petromyzon marinus) have plagued Great
Lakes’ fisheries for many years, little is known about the diet of rarely seen anadromous sea
lampreys during their marine phase. We observed 10 sea lampreys parasitizing basking sharks
(Cetorhinus maximus), the world’s second largest fish, in the Bay of Fundy. Due to the high
concentrations of urea in the blood and tissues of ureosmotic elasmobranchs, we hypothesized
that sea lampreys would require a mechanism(s) to eliminate co-ingested urea while feeding on
basking sharks. Initial urea excretion rates (JUrea) in two lampreys removed from the sharks by
divers were approximately 450 (≈ 9,000 µmol⋅kg-1⋅h-1) and 75 times (≈ 1,500 µmol⋅kg-1⋅h-1)
greater than basal (non-feeding) rates (≈ 20 µmol⋅kg-1⋅h-1). In contrast, JUrea increased by 15-fold
after parasitic lampreys were removed from non-ureosmotic rainbow trout (Oncorhynchus
mykiss). Since activities of the ornithine urea cycle enzymes carbamoyl phosphate synthetase III
(CPSase III) and ornithine carbamoyl transferase (OCT) were relatively low in liver, and below
detection in intestine and muscle, the excreted urea unlikely arose from de novo urea synthesis.
High arginase activities suggested that hydrolysis of dietary arginine made a minor contribution
to JUrea.. Post-feeding ammonia excretion rates (JAmm) were 15-25 fold greater than basal rates in
lampreys removed from both basking sharks and rainbow trout, suggesting that parasitic
lampreys have a high capacity to deaminate amino acids. We conclude that the sea lamprey’s
ability to penetrate the dermal denticle armour of sharks, an ability to rapidly excrete large
volumes of urea, and a high capacity to deaminate amino acids, represent adaptations that have
contributed to the evolutionary success of these phylogenetically ancient vertebrates.
KEY WORDS: arginase, ammonia, basking shark, carbamoyl phosphate synthetase, dermal
denticle, facilitated urea transport, osmoregulation, sea lamprey, trimethylamine oxide, urea,
uricolysis
2 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
3
INTRODUCTION
Lampreys are extant representatives of the superclass Agnatha, an ancient group of
jawless vertebrates whose life style and body structure has been highly conserved for at least 350
million years (Forey and Janvier, 1993). After a filter-feeding larval phase lasting several years,
lampreys undergo a complex metamorphosis characterized by major changes in external and
internal body structure including a re-organization of the nervous, reproductive, respiratory,
digestive, hepatic and renal systems (reviewed by Youson, 1997). Following metamorphosis, the
sexually immature young of parasitic species begin feeding on the blood and/or flesh of fishes,
while non-parasitic species by-pass feeding and immediately begin sexual maturation. It is
generally accepted that these non-parasitic species arose from anadromous, parasitic ancestors
(Potter, 1980).
Although sea lampreys are anadromous, migrating to sea during their parasitic phase
(Scott and Scott, 1988), a landlocked population is also present in the Great Lakes. Indeed,
invading sea lampreys decimated commercial and recreational fisheries in this region in the early
20th century, and continue to threaten Great Lakes fisheries (Klar and Shleen, 2000). The sea
lamprey has several adaptations that make it a highly effective parasite, or a predator when it
ultimately kills its host. Through rhythmic rasping with its tongue, and by creating negative
pressure pulses with its sucker-like oral disc, blood is drawn into the lamprey’s oral cavity
(Rovainen, 1996). Blood flow is promoted by lamphredin secreted by paired buccal glands,
which has anticoagulant and cytolytic properties (Farmer, 1980). In addition, the gills are tidally
ventilated, a respiratory strategy likely selected due to the difficulty associated with drawing
water into the oral cavity while attached to a fish during feeding, or to the substrate (Randall,
1972; Rovainen, 1996).
Despite such adaptations, it is not clear if sea lampreys can parasitize sharks, which
would be ideal hosts due to their large size. Potential obstacles to parasitism could include the
3 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
4
shark’s sharp dermal denticles (scales), which could act as a barrier to sea lamprey attacks, and
the very high osmolarity of the shark’s blood, which is due in part to high concentrations of urea
(e.g. Smith, 1936; Goldstein et al. 1968; Schmidt-Nielsen et al., 1972; Wood et al., 1995). In
contrast to ureosmotic elasmobranchs, sea lampreys are hyposmotic and use mechanisms of
osmoregulation similar to marine teleosts, in which water is extracted from ingested sea water in
the gut, and the co-ingested Na+ and Cl- actively excreted across the gills (Beamish et al., 1978).
Consumption of shark blood would therefore represent an additional osmotic challenge, but
toxicity could also result from urea’s perturbing effects on macromolecules such as proteins
(Yancey, 2001). A sea lamprey feeding on an elasmobranch therefore requires a mechanism(s)
to prevent internal urea accumulation, and/or an ability to eliminate co-ingested urea from their
blood. Accordingly, two objectives of the present study were to (i) confirm that sea lampreys
were able to penetrate the dermal denticle armour of sharks, and (ii) test the hypothesis that sea
lampreys have mechanisms to eliminate co-ingested urea from their blood while feeding.
The presence of basking sharks (Cetorhinus maximus) in the Bay of Fundy in late
summer and early autumn made it possible to determine if they were attacked and parasitized by
sea lampreys that are known to inhabit this area (Scott and Scott, 1988). Basking sharks, which
may attain lengths exceeding 10 m (Scott and Scott, 1988), migrate into this region in search of
zooplankton, the main food item of these giant filter-feeders (Sims and Quayle, 1998). In the
present study, lamprey parasitism of basking sharks was photographically documented. This was
followed by measurements of post-feeding urea and ammonia excretion rates in lampreys after
their removal from the ureosmotic basking sharks, or from freshwater acclimated rainbow trout,
which served as non-ureosmotic controls. To establish if endogenous urea production via
uricolysis or the ornithine urea cycle (OUC) contributed to post-feeding urea excretion patterns,
key enzymes of enzymes of these pathways were also measured in tissues of parasitic sea
4 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
5
lampreys. These latter measurements also enabled us to examine the long-standing hypothesis
that the OUC was present in ancestral agnathans (Griffith, 1991; Walsh, 1997).
5 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
6
MATERIALS AND METHODS
Experimental Animals and Protocol
All experiments were approved by the Mount Allison University, University of New
Brunswick at Saint John, and the University of Toronto Animal Care Committees, and followed
Canadian Council on Animal Care guidelines. Two parasitic sea lampreys (Petromyzon
marinus) were removed from basking sharks (Cetorhinus maximus) and were identified based on
the dentition of their oral disc and body structure (Scott and Scott, 1988). Following removal, the
lampreys were rapidly transported to the University of New Brunswick’s research vessel,
weighed, and transferred to separate, aerated, darkened flux chambers for immediate
determination of urea excretion (JUrea) and ammonia excretion (JAmm) rates (Wilkie and Wood,
1991). Water samples (15 mL) were taken at regular intervals (e.g. 0, 0.5, 1.0, 1.5, 2.0, 3.0 h)
using a 20 mL plastic syringe. In one lamprey, JUrea and JAmm were also monitored between 16
and 20 h post-capture. The lampreys were then transported in a 90 L cooler containing aerated
sea-water to Mount Allison University, where they were held for 2 weeks in a 500 L Living
Stream (Frigid Units Inc), containing sea water at 12ºC, without access to food. Basal JUrea and
JAmm were then determined and the animals were then killed with buffered anaesthetic (3 g
buffered tricaine methanesulfonate⋅L-1; Syndel laboratories), and blood samples (5 mL) taken
from the heart using 5 mL plastic syringes, coated with Na+-heparin (Sigma), for later
determination of ammonia and urea.
A separate series of experiments were conducted at the University of Toronto at
Scarborough, on landlocked, parasitic sea lampreys, provided courtesy of the Hammond Bay
Biological Station, U.S. Fish and Wildlife Service, Millersburg, Michigan. These animals were
held in freshwater at 15ºC for a minimum of 1 week before JUrea and JAmm were determined under
basal (non-feeding) conditions or after feeding on rainbow trout (Oncorhynchus mykiss). One
objective was to determine if similar post-feeding increases in JUrea took place after the ingestion
6 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
7
of blood from a non-ureosmotic fish. A second goal was to determine if parasitic lampreys could
produce urea via the ornithine urea cycle (OUC) or uricolysis.
The rainbow trout, weighing approximately 300 g, were held in a 500 L Living Stream
receiving fresh-water at a rate of 1-2 L⋅min-1, and were fed three times per week with
commercial trout pellets. Parasitic sea lampreys (N = 8) were then introduced. Three days after
initial attachment of lamprey was noted, the trout, with the lamprey still attached, were removed
from the water, and placed in a 5 L bath containing anaesthetic (tricaine methanesulfonate; 1:10
000 dilution) for 1 minute, which rendered the trout unconscious, but not the sea lamprey. The
sea lamprey were then removed and placed in individual flux chambers for measurement of JUrea
and JAmm over 1 h time intervals for 8 h to 72 h. After 24 h or 72 h, the lampreys were killed
with anaesthetic, and blood, liver, intestine and muscle samples were collected. Tissues were
snap-frozen using liquid nitrogen cooled aluminum tongs. Non-feeding lampreys ( N = 10)
served as controls.
Analytical Techniques and Calculations
Measurements of JAmm and JUrea for both anadromous and landlocked, parasitic lampreys
were based on changes in the concentration of each waste product appearing in the water over 30
minutes to 2 hours, after correcting for the animal’s body weight and the volume of water in the
chamber (Wilkie and Wood, 1991). Water and plasma urea concentrations were determined
using a modification of the diacetyl monoxime method (Crocker, 1967) and commercial reagents
(Sigma). Water ammonia was measured using the salicylate-hypochlorite assay (Verdouw et al.,
1978), while plasma ammonia concentration was determined enzymatically (glutamate
dehydrogenase) using a commercial kit (Sigma).
Determination of OUC enzyme activities were optimized and analyzed as previously
described (Mommsen and Walsh, 1989; Barber and Walsh, 1993; Walsh, 1996; Wilkie et al.,
1999). Briefly, chunks of frozen tissues (liver, muscle, intestine) were transferred to a chilled 1.5
7 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
8
mL centrifuge tube and ground for 30 seconds in 4 volumes of homogenization buffer (pH 7.5;
20 mmol.L-1 KH2PO4, 10 mmol.L-1 HEPES, 0.5 mmol.L-1 EDTA, 1 mmol.L-1 dithiothreitol; 50
percent glycerol) using a PowerGen model 125 tissue homogenizer. Following
homogenization, the samples were centrifuged for 3 minutes at 13 000 G and 4oC, afterwhich
enzyme activities were determined on the supernatant using a 96 well plate spectrophotometer at
the appropriate wavelengths (Molecular Devices, Spectramax340). Carbamoyl phosphate
synthetase III (CPSase III) determinations were based on the appearance of citrulline in the
reaction buffer following a 60 minute incubation of homogenate in a cocktail containing N-
acetyl glutamate, ornithine, glutamine and ornithine carbamoyl transferase. To ensure potential
CPSase II activity did not interfere with CPSase III determinations, uridine triphosphate (UTP)
was included in the cocktail to inhibit the former enzyme (Anderson, 1995). Ornithine
carbamoyl transferase activity, also based on citrulline generation, was determined in the
presence of saturating concentrations of ornithine and carbamoyl phosphate. Arginase activity
was based on observed increases in urea concentration following incubation of the liver
homogenate with arginine. Glutamine synthetase activity was based on the rate of appearance of
γ-glutamylhydroxamate, in the presence of glutamine, hydroxylamine and ferric chloride (Walsh,
1996). All enzyme activities are expressed as µmol product formed per gram of wet tissue per
minute at 20°C (µmol⋅g-1⋅min-1).
The JUrea and JAmm are presented separately for each lamprey removed from the basking
sharks. All other data are expressed as the mean ± 1 standard error of the mean (SEM).
Unpaired and paired data were analyzed using analysis of variance (ANOVA). When significant
variability was noted, statistical significance was determined using the Tukey-Kramer post-test at
the P < 0.05 level.
8 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
9
RESULTS
Five basking sharks were observed south of the Wolve’s Islands, Bay of Fundy (Water T
=12°C), with a total of 10 lampreys attached to 4 of the sharks. The lampreys were generally
positioned behind the dorsal fin in groups of 1 to 4 (Fig. 1), and numerous lamprey scars and
fresh wounds in the vicinity of the dorsal fin indicated active feeding (Fig. 1). Divers
subsequently removed 2 lampreys from the basking sharks (Fig. 1 inset) and returned them to the
research vessel, where on-board measurements of urea excretion (JUrea) and ammonia excretion
(JAmm) rates were immediately initiated.
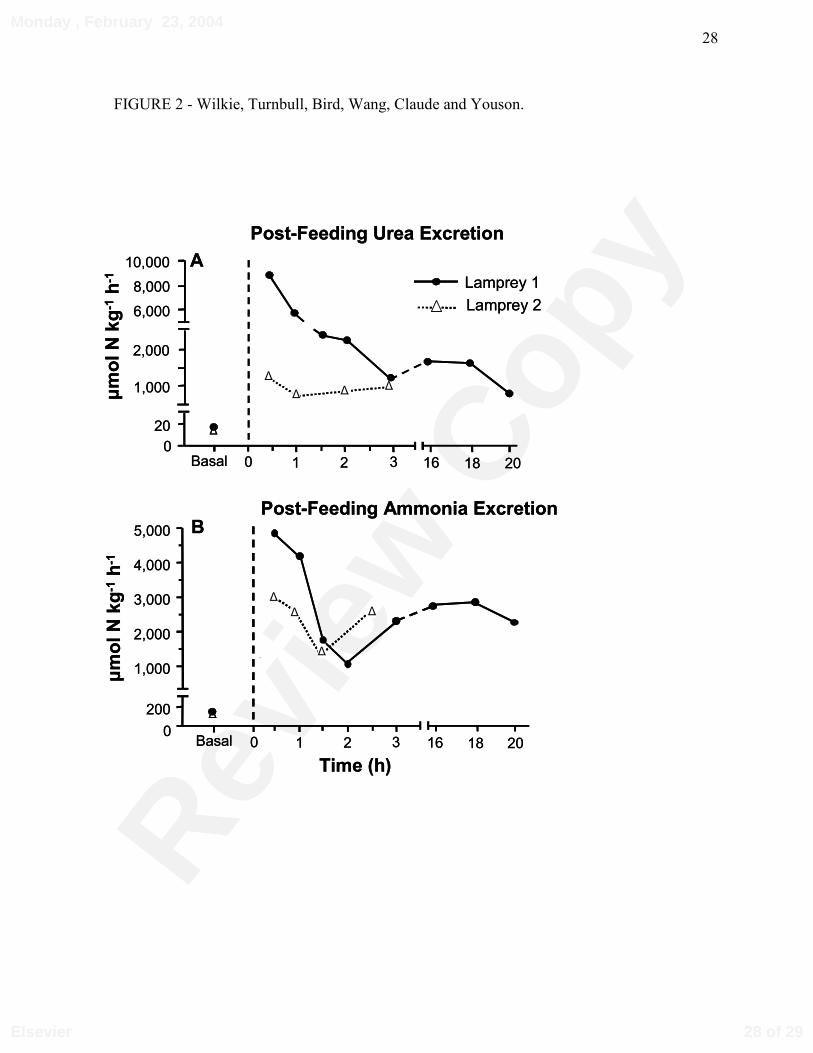
In the first lamprey, post-removal JUrea approached 9,000 µmol⋅kg-1⋅h-1 (Fig. 2A) over the
first 30 minutes, which was approximately 450 times greater than basal rates measured in the
laboratory after a two week fasting period, under identical saltwater conditions (full strength sea
water, 12°C). JUrea rapidly decreased over the next 2 h, and stabilized at 1,000-1,500 µmol⋅kg-1⋅h-
1 (Fig. 2A). Compared to basal rates, post-removal JAmm was 20-25 times greater during the first
hour. However, JAmm was 25 to 50 % lower than the corresponding measurements of JUrea during
the same period (Compare Fig. 2A to Fig. 2B). In the second lamprey, JUrea fluctuated between
500 and 1,000 µmol⋅kg-1⋅h-1 over 3 h, but JAmm was 2-3 fold greater (Fig. 2A, 2B). Due to rough
seas, it was not possible to monitor post-removal JUrea and JAmm further. Plasma ammonia and
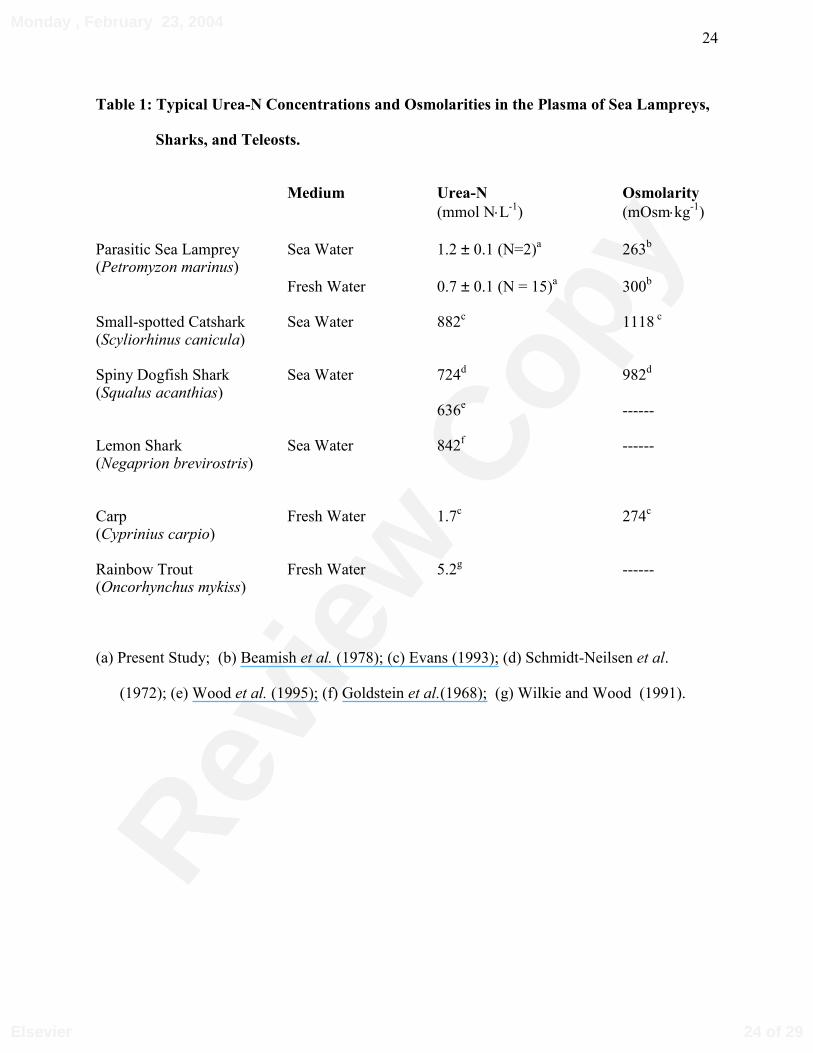
urea concentrations in the two lampreys were similar after 2 weeks, approximately 1.2 mmol⋅L-1
(Table 1) and 130 µmol⋅L-1 (data not shown), respectively.
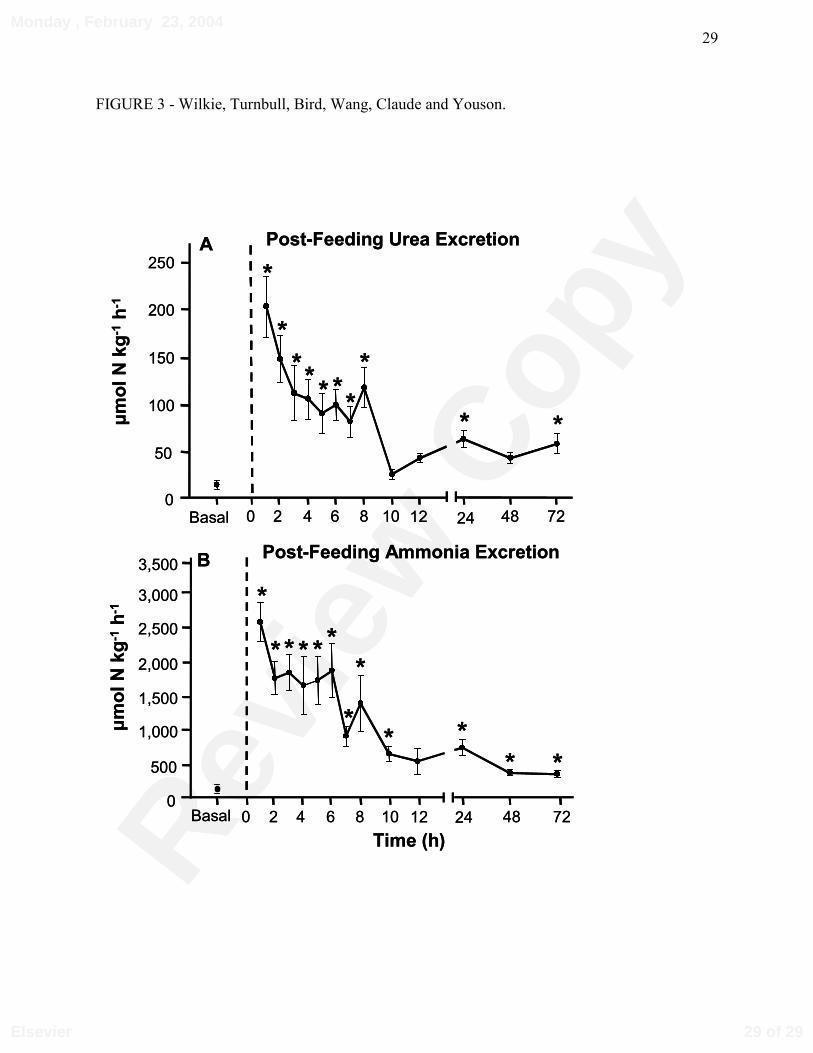
When JUrea and JAmm were measured in landlocked, laboratory held parasitic lampreys,
under control conditions, in which they were denied access to food for 7-10 days, basal JUrea was
approximately 15 µmol⋅kg-1⋅h-1 (Fig. 3A), while JAmm fluctuated around 100 µmol⋅kg-1⋅h-1 (Fig.
3B). After parasitizing rainbow trout, JUrea was stimulated 15-fold (Fig. 3A), and remained
elevated by 5- to 10-fold over 8 h before returning to pre-feeding levels. Similarly, post-feeding
9 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
10
JAmm increased by approximately 25-fold, before returning towards basal levels after 8 h (Fig.
3B). Basal (non-feeding) plasma urea and ammonia concentrations averaged 0.7 ± 0.1 mmol⋅L-1
(N = 15; Table 1), and 685 ± 18 µmol⋅L-1 (N = 15; data not shown), respectively.
Key enzymes of the OUC and uricolytic pathways were monitored in the liver, intestine
and muscle of landlocked parasitic sea lampreys. Analysis of four key OUC enzymes revealed
that each was present in the liver (Table 2). However, CPSase III, and ornithine carbamoyl
transferase were below levels of detection in the intestine and muscle. Arginase, which catalyzes
the hydrolysis of arginine to urea in the final step of the OUC, exhibited 10-fold greater activity
in the liver compared to the intestine, but was below levels of detection in the muscle (Table 2).
Glutamine synthetase activity was detected in all three tissues, but was greatest in the intestine.
The two terminal enzymes in the uricolytic pathway, allantoinase and allantoicase, were also
detected in the liver of parasitic lampreys (Table 2).
10 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
11
DISCUSSION
The very high JUrea of anadromous sea lampreys provides convincing evidence that these
parasites were feeding sometime prior to their removal from the basking sharks. Earlier studies
and anecdotal observations suggested possible lamprey parasitism of basking sharks (Bigelow
and Shroeder, 1948), and more recently it has been reported that sea lampreys also attack
sandbar (Carcharhinus plumbeus) and dusky sharks (Carcharhinus obscurus; Jensen and
Schwartz, 1994), as well as tiger sharks (Galeocerdo cuvier; Jensen et al., 1998). It is not clear if
lampreys were feeding in these other instances, due to the difficulty and danger associated with
removing lampreys from these more active, predacious shark species. However, open wounds
were reported on the bodies of the sandbar and dusky sharks (Jensen and Schwartz, 1994).
Parasitism of elasmobranchs could be one reason why anadromous sea lampreys are
approximately twice as large as their landlocked counterparts, whose diet is restricted to much
smaller teleosts (Farmer, 1980). Indeed, anadromous sea lampreys weigh up to 1 kg and attain
lengths of more than 70 cm (Beamish and Potter, 1975).
The observed JUrea indicate that parasitic sea lampreys can exhibit facultative ureotelism,
in which urea becomes the dominate nitrogenous waste product excreted, following the ingestion
of urea-rich blood. The rapid decrease in JUrea following the first lamprey’s removal from the
basking shark likely reflected the cessation of feeding. During actual feeding, JUrea would need to
be sustained at the high levels initially measured following removal from the shark. The initially
lower JUrea of the second lamprey may because feeding ceased sometime prior to its removal
from the basking shark.
These JUrea’s are amongst the highest ever recorded in fishes; only the Lake Magadi
tilapia (Alcolapia grahami; Randall et al., 1989) and gulf toadfish (Opsanus beta; Mommsen and
Walsh, 1989) have had JUrea in the range reported here. However, the Lake Magadi tilapia, gulf
toadfish, and other ureogenic fishes such as the Indian catfishes (Heteropneustes fossilis; Saha
11 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
12
and Ratha, 1989), have a fully functional ornithine urea cycle (OUC), which produces urea at
high rates to detoxify ammonia under conditions (e.g. alkaline water, air exposure, crowding)
that oppose ammonia excretion. Moreover, the urea generated by the Magadi tilapia and the gulf
toadfish is excreted across the gills via facilitated urea transport (Walsh et al., 2000, 2001), while
elasmobranchs appear to use similar transport mechanisms to retain urea for osmoregulation (e.g.
Schmidt-Nielsen et al., 1972; Wood et al., 1995; Smith and Wright, 1999; Fines et al., 2001).
Although urea has a low molecular mass (60 Da) and high water solubility, its very low lipid
partition co-efficient precludes urea movement across lipid bilayers, necessitating the use of
facilitated and active urea transporters (reviewed by Walsh and Smith, 2001). Indeed, facilitated
and active urea transporters have been well characterized in mammalian renal tubules, where
they concentrate urea in the inner renal medulla to maximize water reabsorption (Knepper and
Chou, 1995; You et al., 1993; Sands, 1999). The high JUrea following feeding on basking sharks,
as well as the 15-fold increase following feeding on rainbow trout, therefore suggests that similar
urea transport mechanisms are probably present in the gills, kidneys, and/or the gut of sea
lampreys. However, further investigation is required to confirm this hypothesis and to identify
the route(s) of urea excretion in these animals.
It is unlikely that post-feeding JUrea by parasitic lampreys arose from de novo urea
synthesis. For instance, the plasma urea concentrations were relatively low in both seawater and
freshwater adapted parasitic sea lampreys that were not feeding (Table 1). Although OUC
enzymes were present in the livers of the landlocked sea lampreys, the activities of key enzymes
such as CPSase III and OCT were relatively low, and below detection in intestine and muscle
(Table 2). Arginase activities were relatively high (Table 2) compared to other fishes, which
suggests that hydrolysis of dietary arginine contributed to the high post-feeding JUrea observed in
lampreys following their removal from both basking sharks and trout. Given the low blood
arginine concentrations in both sharks and salmonid fishes, it seems unlikely that arginine
12 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
13
hydrolysis accounted for more than a small fraction of the observed JUrea. For instance, plasma
arginine concentrations are approximately 0.2 mmol·L-1 in elasmobranchs (Gutiérrez et al.,
1987), while urea concentrations range from approximately 600-900 mmol·L-1 (Table 1).
Nevertheless, it would be useful to more precisely quantify the role that dietary arginine plays in
post-feeding urea excretion by parasitic lampreys.
Since ureosmotic elasmobranchs retain urea in their body fluids at concentrations that are
2-3 orders of magnitude greater than in lampreys and teleosts (Table 1), a more likely
explanation for the high post-feeding JUrea is that the sea lampreys were simply clearing urea that
was co-ingested as they parasitized their hosts. For instance, lampreys parasitizing salmonid
fishes consume blood at rates ranging from 3-30 % of their body weight per day (Farmer et al.,
1975). Thus, if a 100 g sea lamprey were to ingest the blood of a typical elasmobranch
(assuming a blood urea concentration of 636 mmol⋅L-1; Wood et al., 1995) at similar rates, a urea
load of 19-190 mmol urea-N·kg-1 body weight per day would result! Although typical blood
urea concentrations are much lower in salmonids, at most 5 mmol⋅L-1 (Wilkie and Wood, 1991),
a lamprey feeding on a trout would experience a urea load ranging from approximately 0.16-1.60
mmol urea-N·kg-1 body weight per day. Such a urea load from a trout would not likely have any
detrimental effects on a parasitic lamprey. However, the 100-fold greater urea load that may
arise from the parasitism of an elasmobranch such as a basking shark could be toxic due to urea’s
perturbing effects on protein structure. Indeed, urea is known to perturb enzyme function,
microtubule assembly, and the structural integrity of membranes (reviewed by Yancey, 2001).
It might be argued that that the high concentrations of counteracting solutes such as
trimethylamine oxide (TMAO) in elasmobranch blood and tissues would confer some degree of
protection to a lamprey parasitizing a shark. Indeed, TMAO is thought to protect elasmobranchs
against urea by stabilizing protein structure (Yancey, 2001), and would likely have similar
benefits if it were co-ingested with urea by a parasitic sea lamprey. However, it should also be
13 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
14
pointed out that elasmobranchs have additional adaptations that may provide further protection
against urea’s destabilizing effects such as urea tolerant proteins that include actomyosin
ATPases and hemoglobin (Yancey, 2001). The higher percentage of saturated fatty acids in the
mitochondrial membranes of elasmobranchs are also thought to increase their resistance to urea’s
perturbing effects (Glemet and Ballantyne, 1996). Further studies are required to quantify the
urea and TMAO load in the blood and tissues of lampreys parasitizing elasmobranchs.
Additional experiments should also evaluate the possibility that co-ingested TMAO ameliorates
urea’s perturbing effects, as well as probing for lamprey-specific adaptations that might allow
them to withstand large, transient elevations of internal urea.
Although urea synthesis does not substantially contribute to greater post-feeding JUrea,
these findings may shed some light on the origin of the OUC in vertebrates. Most fishes are
ammoniotelic, and readily excrete most of their nitrogenous wastes as highly soluble ammonia
across the gills (Wilkie, 2002). Because it is less toxic than ammonia, urea synthesis is
considered one of the key adaptations that enabled vertebrates to invade terrestrial habitats
(Mommsen and Walsh, 1989; Griffith, 1991). A central hypothesis is that the OUC was present
in ancestral agnathans, but that its expression was subsequently silenced in fishes because it was
energetically less expensive to produce and excrete ammonia (Brown and Brown, 1985; Griffith,
1991; Wright et al.,1995; Walsh, 1997). The presence of key OUC enzymes in many teleosts,
along with elasmobranchs, coelacanths, lungfishes, amphibians and mammals supports the
hypothesis that the OUC is a monophyletic trait in the vertebrate lineage (Mommsen and Walsh,
1989, 1991). However, this hypothesis has not yet been substantiated by measurements of OUC
enzymes in extant agnathans such as the hagfishes or lampreys.
Wilkie et al. (1999) demonstrated that larval sea lampreys excreted physiological relevant
amounts of urea, but were unable to detect the key OUC enzyme, CPSase III, in the liver of these
animals. We have no explanation for the lack of CPSase III activity in larval lamprey livers, but
14 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
15
it could be related to their much lower metabolic rate, slow rate of digestion (Potter and Rogers,
1972; Sutton and Bowen, 1994; Wilkie et al., 2001) and vastly different diet, which mainly
comprises detritus (Sutton and Bowen, 1994). Although Read (1968) detected significant
CPSase activity in the liver of adult Pacific lamprey (Entosphenus tridentatus), he was unable to
identify other key OUC enzymes such as OCT. Further, it was not clear if he detected CPSase
III, because he did not inhibit CPSase II using uridine triphosphate (Anderson, 1995), as was
done in the present study. The low activity of CPSase III and OCT in the liver of parasitic sea
lampreys suggests that the components of this key urea production pathway were probably
present in an ancestral jawless vertebrate. However, these low activities also suggest that the
OUC produces little, if any, physiological relevant amounts of urea in parasitic sea lampreys.
More likely, the small amounts of urea excreted under starved conditions represent endogenous
production arising from uricolysis, as was suggested for larval lampreys (Wilkie et al., 1999).
Indeed, the activities of allantoicase and allantoinase in the present study are comparable to
measurements made in the livers of larval lampreys.
Several-fold increases in JAmm are common in fishes following feeding, due to increased
proteolysis and amino acid catabolism (Wood, 2001). However, the 25-fold greater JAmm
observed in parasitic sea lampreys after feeding on both sharks and rainbow trout is unusually
high compared to other fishes, suggesting that parasitic lampreys have a high capacity to
deaminate excess amino acids arising from their protein rich blood diet and high feeding rates.
Further support for this hypothesis is provided by measurements of hepatic glutamate
dehydrogenase activity and expression, which is several-fold higher in the parasitic stage of the
sea lamprey’s life cycle (Wilkie, Wang, Claude, and Youson, unpublished observations). Such a
strategy would be beneficial during the more active, free swimming parasitic stage of the
lamprey life cycle, by providing the raw-materials (α-keto acids) required for ATP production
via oxidative pathways or glycogen synthesis (Mommsen et al., 1980). Muscle glycogen stores
15 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
16
could be critical for providing energy during episodes of burst exercise during subsequent
upstream migration and spawning by the sea lamprey (Boutilier et al., 1993). Indeed, the muscle
glycogen concentrations of migrating sea lampreys are comparable to stores measured in the
white muscle of salmonids (Milligan, 1996), which undergo similar extensive migrations.
Our observations reveal that anadromous sea lampreys parasitize basking sharks, and that
they rapidly unload co-ingested urea, which likely prevents deleterious increases in internal urea
and additional osmotic stress. Although the life style and body structure of lampreys has been
highly conserved for at least the last 350 million years (Forey and Janvier, 1993), we have little
evidence of their behaviour or physiology to explain their success. The ability of sea lampreys to
penetrate the dermal denticle armor of an elasmobranch, an ability to rapidly excrete potentially
toxic urea, and a high capacity to deaminate amino acids, may represent additional adaptations
that have contributed to the lamprey’s evolutionary success. The presence of key OUC enzymes
such as CPSase III and OCT, also supports the hypothesis that the components of this pathway of
urea production were likely present in an ancestral jawless fish(es).
16 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
17
ACKNOWLEDGEMENTS
We are grateful to Bob Bozene, Captain of the University of New Brunswick research vessel the
“Mary O”, and to Wayne Anderson who provided invaluable technical assistance. Ed Woods and
Tim Geers are acknowledged for their assistance locating and photographing the basking sharks
and lampreys observed in this study. We thank Dr. P.J. Walsh for his helpful comments. This
research was funded by Natural Sciences and Engineering Research Council (NSERC) of
Canada research grants to MPW and JHY; ST was supported by the University of New
Brunswick at Saint John; YSW held an NSERC postdoctoral fellowship.
17 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
18
REFERENCES
Anderson, P.M. 1995. Urea cycle in fish: molecular and mitochondrial studies. In: C.M. Wood;
T.J. Shuttleworth (Eds.), Fish Physiology. vol. 14. Cellular and molecular approaches to
fish ionic regulation. Academic Press, New York. pp. 57–83.
Barber, M.L., Walsh, P.J. 1993. Interactions and acid-base status and nitrogen excretion and
metabolism in the ureogenic teleost, Opsanus beta. J. Exp. Zool. 185, 87-105.
Beamish, F.W.H., Potter, I.C. 1975. The biology of the anadromous sea lamprey (Petromyzon
marinus) in New Brunswick. J. Zool., Lond. 177, 57-72.
Beamish, F.W.H., Strachan, P.D., Thomas, E. 1978. Osmotic and ionic performance of the
anadromous sea lamprey, Petromyzon marinus. Comp. Biochem. Physiol. 60A, 435-443.
Bigelow, H.B., Schroeder, W.C. 1948. Cyclostomes. In: Fishes of the Western North Atlantic.
Mem. Sears. Found. Mar. Res. 1(1): Yale Univ., New Haven, Conn. pp. 29-58.
Boutilier R.G., Ferguson, R.A., Henry, R.P., Tufts, B.L. 1993. Exhaustive exercise in the sea
lamprey (Petromyzon marinus): relationships between anaerobic metabolism and
intracellular acid-base balance. J. Exp. Biol. 178, 71–88.
Brown, G.W., Brown, S.G. 1985. On urea formation in primitive fishes. In: Foreman, R.E.,
Gorbman, A., Dodd, J.M., Olsson, R. (Eds.), Evolutionary Biology of Primitive Fishes.
NATO Advanced Research Workshop. Plenum Press, London and New York. pp. 321–
337.
Crocker, C.L. 1967. Rapid determination of urea nitrogen in serum or plasma without
deproteinization. Am. J. Med. Technol. 33, 361–365.
Evans, D.H. 1993. Osmotic and ion regulation. In: Evans, D.H. (Ed.). The Physiology of Fishes.
CRC Press, Boca Raton, pp. 315-341.
Farmer, G.J. 1980. Biology and physiology of feeding in adult lampreys. Can. J. Fish. Aquat.
Sci. 37, 1751-1761.
18 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
19
Farmer, G.J., Beamish, F.W.H., Robinson, G.A. 1975. Food consumption of the adult
landlocked sea lamprey, Petromyzon marinus. Comp. Biochem. Physiol. 50A, 753-757.
Fines, G.A., Ballantyne, J.S., Wright, P.A. 2001. Active urea transport and an unusual
basolateral membrane composition in the gills of a marine elasmobranch. Am. J. Physiol.
280, R16-R24.
Forey, P., Janvier, P. 1993. Agnathans and the origin of jawed vertebrates. Nature 361, 129-134.
Glemet, H.C., Ballantyne, J.S. 1996. A comparison of liver mitochondrial membranes from an
agnathan (Myxine glutinosa), an elasmobranch (Raja erinacea) and a teleost fish
(Pleuronectes americanus). Mar.Biol. 124, 509-518.
Goldstein. L., Oppelt, W.W., Maren, T.W. 1968. Osmotic regulation and urea metabolism in the
lemon shark Negaprion brevirostris. Am. J. Physiol. 215, 1493-1497.
Griffith, R.W. 1991. Guppies, toadfish, lungfish, coelacanths and frogs: a scenario for the
evolution of urea retention in fishes. Environ. Biol. Fishes 32, 199-218.
Gutiérrez, J., Fernandez, J., Geese, J.M., Blasco, J. 1987. Amino acid levels in whole blood and
plasma of Scylorhinus canicula. Comp. Biochem. Physiol. 87A, 57-61.
Jensen, C., Schwartz, F.J. 1994. Atlantic Ocean occurrence of the sea lamprey, Petromyzon
marinus, (Petromyzontiformes,Petromyzontidae), parasitizing sandbar, Carcharinus
plumbeus, and dusky, C. obscurus (Carcharhiniformes, Carcharhinidae), sharks off North
Carolina and South Carolina. Brimleyana 21, 69-72.
Jensen, C., Schwartz, F.J., Hopkins, G. 1998. A sea lamprey (Petromyzon marinus)-tiger shark
(Galeocerdo cuvier) parasitic relationship off North Carolina. Journal of the Elisha
Mitchell Scientific Society. 114, 72.
Klar, G.T., Shleen, L.P. 2000. Integrated management of the sea lampreys in the Great Lakes
1999. Annual report to the Great Lakes Fishery Commission.
19 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
20
Knepper MA, Chou C-L. 1995. Urea and ammonium transport in the mammalian kidney. In:
Walsh, P.J., Wright, P.A. (Eds.), Nitrogen Metabolism and Excretion. CRC Press, Boca
Raton, Florida, pp. 205-227.
Milligan, C.L. 1996. Metabolic recovery from exhaustive exercise in rainbow trout. Comp.
Biochem. Physiol. 113A, 51–60.
Mommsen, T.P., Walsh, P.J. 1989. Evolution of urea synthesis in vertebrates: the piscine
connection. Science 243, 72-75.
Mommsen, T.P., Walsh, P.J. 1991. Urea synthesis in fishes: evolutionary and biochemical
perspectives. In: Hochacka, P., Mommsen, T.P. (Eds.), Biochemistry and Molecular
Biology of Fishes, Volume 1. Elsevier Science Publications, New York, pp. 137-163.
Potter, I.C. 1980. The Petromyzontiformes with particular reference to paired species. Can. J.
Fish. Aquat. Sci. 37, 1595-1615.
Potter I.C., Rogers, M..J. 1972. Oxygen consumption in burrowed and unburrowed ammocoetes
of Lampetra planeri (Bloch). Comp. Biochem. Physiol. 41A, 427–432.
Randall, D.J. 1972. Respiration. In: Hardisty, M.W. , Potter, I.C. (Eds.), The Biology of
Lampreys, vol. 2. Academic Press, New York, pp. 287-306 .
Randall, D.J., Wood, C.M., Perry, S.F., Bergman, H., Maloiy, G.M.O., Mommsen, T.P., Wright
P.A. 1989. Urea excretion as a strategy for survival in a fish living in a very alkaline
environment. Nature 337,165-166.
Read, L.J. 1968. A study of ammonia and urea production and excretion in the fresh-water-
adapted form of the Pacific lamprey (Entosphenus tridentatus). Comp. Biochem. Physiol.
26, 455–466.
Rovainen, C.M. Feeding and breathing in lampreys. 1996. Brain Behav. Evol. 48, 297-305.
Saha N., Ratha B.K. 1989. Comparative study of ureagenesis in freshwater, air-breathing
teleosts. J. Exp. Zool. 252, 1-8.
20 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
21
Sands, J.M. 1999. Regulation of renal urea transporters. J. Am. Soc. Nephrol. 10, 635-646.
Schmidt-Nielsen, B., Truniger, B., Rabinowitz, L. 1972. Sodium-linked urea transport by the
renal tubule of the spiny dogfish Squalus acanthias. Comp. Biochem. Physiol. 42A, 13-25.
Scott, W.B., Scott, M.G. 1988. Atlantic Fishes of Canada. Can. Bull. Fish. Aquat. Sci. 219.
Sims, D.W., Quayle, V.A. 1998. Selective foraging behaviour of basking sharks on zooplankton
in a small-scale front. Nature 393, 460-464.
Smith, C.P., Wright P.A. 1999. Molecular characterization of an elasmobranch urea transporter.
Am. J. Physiol. 276, R622-R626.
Smith, H.W. 1936. The retention and physiological role of urea in the Elasmobranchii. Biol.
Bull.11,49-82.
Sutton, T.M., Bowen, S.H. 1994. Significance of organic detritus in the diet of larval lampreys in
the Great Lakes basin. Can. J. Fish. Aquat. Sci. 51, 2380–2387.
Verdouw, H., van Echted, C.J.A., Dekkers, E.M.J. 1978. Ammonia determination based on
indophenol formation with sodium salicylate. Water Res. 12, 399-402.
Walsh, P.J. 1996. Purification and properties of hepatic glutamine synthetases from the ureotelic
Gulf toadfish, Opsanus beta. Comp. Biochem. Physiol. B, 115, 523–532.
Walsh, P.J. 1997. Evolution and regulation of urea synthesis and ureotely in (Batrachoidid)
fishes. Annu. Rev. Physiol. 59, 299-323.
Walsh PJ, Smith CP. 2001. Urea transport. In: Wright, P.A., Anderson P.M. (Eds.), Fish
Physiology Vol. 20: Nitrogen Excretion. Academic Press, New York, pp. 279-307.
Walsh P.J., Walsh PJ, Grosell M., Goss, G.G., Bergman, H.L., Bergman, A.N., Wilson, P.,
Laurent, P., Alper, S.L., Smith, C.P., Kamunde, C., Wood, C.M. 2001. Physiological and
molecular characterization of urea transport by the gills of the Lake Magadi tilapia
(Alcolapia grahami). J. Exp. Biol. 204, 509-520.
21 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
22
Walsh, P.J., Heitz, M.J., Campbell, C.E., Cooper, G.J., Medina, M., Wang, Y.S., Goss, G.G.,
Vincek, V., Wood, C.M., Smith, C.P. 2000. Molecular characterization of a urea
transporter in the gill of the gulf toadfish (Opsanus beta). J. Exp. Biol. 203, 2357-2364.
Wilkie, M.P., Wood, C.M. 1991. Nitrogenous waste excretion, acid-base regulation and
ionoregulation in rainbow trout (Oncorhynchus mykiss) exposed to extremely alkaline
water. Physiol. Zool. 64,1069-1086.
Wilkie, M.P., Wang, Y, Walsh, P.J., Youson, J.Y. 1999. Nitrogenous waste excretion by the
larvae of a phylogenetically ancient vertebrate: the sea lamprey (Petromyzon marinus).
Can. J. Zool. 77, 707-715.
Wilkie, M.P. 2002. Ammonia excretion and urea handling by fish gills: present understanding
and future research challenges. J. Exp. Zool. 293, 284-301.
Wood, C.M. 1993. Ammonia and urea metabolism and excretion. In: Evans, D.H. (Ed.), The
Physiology of Fishes. CRC Press, Boca Raton, Florida. pp. 379-425.
Wood, C.M. 2001. Influence of feeding, exercise, and temperature on nitrogen metabolism and
excretion. In: Wright, P.A, Anderson, P. (Eds.), Fish Physiology: Nitrogen Excretion, vol.
20. Academic Press, New York, pp. 201-238.
Wood C.M., Pärt P., Wright P.A. 1995. Ammonia and urea metabolism in relation to gill
function and acid-base balance in a marine elasmobranch, the spiny dogfish (Squalus
acanthias). J. Exp. Biol. 198,1545-1558.
Wright, P.A., Felskie, A., Anderson, P.M. 1995. Induction of ornithine-urea cycle enzymes and
nitrogen metabolism and excretion in rainbow trout (Oncorhynchus mykiss) during early
life stages. J. Exp. Biol. 198, 127-135.
Yancey, P.H. Nitrogen compounds as osmolytes. 2001. In: Wright, P.A, Anderson, P. (Eds.),
Fish Physiology: Nitrogen Excretion, vol. 20. Academic Press, New York, pp. 309-341.
22 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
23
You, G., Smith, C.P., Kanai, Y., Lee, W-S., Stelzner, M., Hediger, M.A. 1993. Cloning and
characterization of the vasopressin-regulated urea transporter. Nature 365, 844-847.
Youson, J.H. 1997. Is lamprey metamorphosis regulated by thyroid hormones? Am. Zool. 37,
441-460.
23 of 29
Monday , February 23, 2004
Elsevier
Revie
w C
opy
24
Table 1: Typical Urea-N Concentrations and Osmolarities in the Plasma of Sea Lampreys,
Sharks, and Teleosts.
Medium Urea-N Osmolarity
(mmol N⋅L-1) (mOsm⋅kg-1)
Parasitic Sea Lamprey Sea Water 1.2 ± 0.1 (N=2)a 263b
(Petromyzon marinus)Fresh Water 0.7 ± 0.1 (N = 15)a 300b
Small-spotted Catshark Sea Water 882c 1118 c (Scyliorhinus canicula)
Spiny Dogfish Shark Sea Water 724d 982d
(Squalus acanthias)636e ------
Lemon Shark Sea Water 842f ------ (Negaprion brevirostris)
Carp Fresh Water 1.7c 274c
(Cyprinius carpio)
Rainbow Trout Fresh Water 5.2g ------ (Oncorhynchus mykiss)
(a) Present Study; (b) Beamish et al. (1978); (c) Evans (1993); (d) Schmidt-Neilsen et al.
(1972); (e) Wood et al. (1995); (f) Goldstein et al.(1968); (g) Wilkie and Wood (1991).
24 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
25
Table 2: Activities of Key Ornithine Urea Cycle and Uricolytic Enzymes in the Liver, Muscle and Intestine of Parasitic Sea Lampreys.
Liver Muscle Intestine Ornithine Urea Cycle Enyzmes Carbamoyl Phosphate Synthetase III 0.0065 ± 0.001 (7) BLD (6) BLD (6)
Ornithine Carbamoyl Transferase 0.13 ± 0.03 (7) BLD (6) BLD (6) Arginase 9.38 ± 2.57 (7) BLD (6) 0.83 ± 0.38 (6) Glutamine Synthetase 3.06 ± 0.59 (7) 0.33 ± 0.01 (7) 8.43 ± 1.77 (6)
Uricolytic Enzymes Allantoicase 0.03 ± 0.00 (4) ------ ------ Allantoinase 0.50 ± 0.02 (7) ------ ------
All enzyme activities are expressed as µmol product formed per gram of wet tissue per minute at
20°C (µmol⋅g-1⋅min-1). Number of animals sampled in parentheses. BLD; below level of
detection.
25 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
26
FIGURE LEGENDS
Figure 1: Parasitic sea lampreys attached to the dorsal body surface of basking sharks off of the
Wolves Islands, Bay of Fundy, Canada. Inset: Removal of a sea lamprey from the dorsal
body surface of a basking shark.
Figure 2: Individual (A) urea and (B) ammonia excretion rates of two anadromous, parasitic sea
lampreys following their removal from basking sharks, and their basal rates after a two week
fast. Solid circles represent the first lamprey captured (Mass = 80 g), and open triangles the
second (Mass 292 g).
Figure 3: Post-feeding (A) urea and (B) ammonia excretion rates of landlocked, parasitic sea
lampreys (N=8; Mass = 25.2 ± 7.7 g) after removal from rainbow trout. Basal rates were
determined on lampreys (N = 10; Mass = 22.3 ± 2.6 g) that had been fasted for at least 1
week. Asterisks (*) denote statistically significant differences between post-feeding and basal
(non-feeding) excretion rates (P<0.05).
26 of 29
Monday , February 23, 2004
Elsevier

Revie
w C
opy
27
FIGURE 1 - Wilkie, Turnbull, Bird, Wang, Claude and Youson.
27 of 29
Monday , February 23, 2004
Elsevier
Revie
w C
opy
28
FIGURE 2 - Wilkie, Turnbull, Bird, Wang, Claude and Youson.
020
1,000
2,000
6,0008,000
10,000
∆
0 1 2 3 16 18 20Basal
∆
∆
Post-Feeding Urea Excretion
Lamprey 1Lamprey 2
A
0200
1,000
2,000
3,000
4,000
5,000
0 1 2 3 16 18 20Time (h)
∆∆
∆
∆
∆
Post-Feeding Ammonia ExcretionB
Basal
µmol
Nkg
-1h-
1µm
olN
kg-1
h-1
∆ ∆
020
1,000
2,000
6,0008,000
10,000
∆
0 1 2 3 16 18 20Basal
∆
∆
Post-Feeding Urea Excretion
Lamprey 1Lamprey 2
A
0200
1,000
2,000
3,000
4,000
5,000
0 1 2 3 16 18 20Time (h)
∆∆
∆
∆
∆
Post-Feeding Ammonia ExcretionB
Basal
µmol
Nkg
-1h-
1µm
olN
kg-1
h-1
∆ ∆
28 of 29
Monday , February 23, 2004
Elsevier
Revie
w C
opy
29
FIGURE 3 - Wilkie, Turnbull, Bird, Wang, Claude and Youson.
-2 0 2 4 6 8 10 120
50
100
150
200
250
24 48 72
*
*** * **
*
* *
2 4 8 106 120 24 48 72Basal
A
-2 0 2 4 6 8 10 120
500
1,000
1,500
2,000
2,500
3,000
3,500
24 48 72Basal 2 4 8 106 120 24 48 72
*
* * * * *
*
*
**
**
B
Time (h)
µmol
Nkg
-1h-
1µm
olN
kg-1
h-1
Post-Feeding Urea Excretion
Post-Feeding Ammonia Excretion
-2 0 2 4 6 8 10 120
50
100
150
200
250
24 48 72
*
*** * **
*
* *
2 4 8 106 120 24 48 72Basal
A
-2 0 2 4 6 8 10 120
500
1,000
1,500
2,000
2,500
3,000
3,500
24 48 72Basal 2 4 8 106 120 24 48 72
*
* * * * *
*
*
**
**
B
Time (h)
µmol
Nkg
-1h-
1µm
olN
kg-1
h-1
Post-Feeding Urea Excretion
Post-Feeding Ammonia Excretion
29 of 29
Monday , February 23, 2004
Elsevier




















