
Rat hepatic stellate cells contribute to the acute-phase response with increased expression of...
11
Rat Hepatic Stellate Cells Contribute to the Acute-Phase Response With Increased Expression of a1(I) and a1(IV) Collagens, Tissue Inhibitor of Metalloproteinase-1, and Matrix-Metalloproteinase-2 Messenger RNAs NATALIA NIETO, 1 JOSE A. DOMINGUEZ-ROSALES, 1 LUIS FONTANA, 1 ADRIANA SALAZAR, 3 JUAN ARMENDARIZ-BORUNDA, 3 PATRICIA GREENWEL, 4 AND MARCOS ROJKIND 1,2 The acute-phase response (APR) represents a systemic reaction of the organism to multiple nonspecific inflamma- tory stimuli. In general, it is protective for the host, and hepatocytes are the main cells responding with alterations in the expression of a set of liver-specific proteins named the acute-phase proteins. We have previously shown that al- though a turpentine-induced APR is not fibrogenic per se, it enhances collagen deposition in rats treated with CCl 4 and up-regulates expression of hepatic a1(I) collagen and tissue inhibitor of metalloproteinases 1 (TIMP-1) messenger RNAs (mRNAs). In this report we extended our studies and showed that turpentine induced, in a time-dependent man- ner, expression of a1(I) and a1(IV) collagens, TIMP-1, and matrix-metalloproteinase 2 (MMP-2) mRNAs. We further showed that expression of these mRNAs occurs in hepatic stellate cells, but not in hepatocytes obtained 6 hours after the induction of an APR episode. These changes were ac- companied by increased blood levels of tumor necrosis fac- tor a (TNF-a) and interleukin 6 (IL-6) without noticeable immediate changes in the expression of their respective mR- NAs in the liver. In contrast to CCl 4 -induced liver damage, turpentine alone, whether administered as a single dose or as a weekly dose for 3 weeks did not up-regulate expression of transforming growth factor b1 (TGF-b1) mRNA and did not result in excess collagen deposition. Overall, these find- ings suggest that collagen deposition in the livers of rats with repeated APR episodes may be enhanced only when given together with a fibrogenic stimulus that activates he- patic stellate cells (HSCs) and/or up-regulates TGF-b1 mRNA expression. (HEPATOLOGY 2001;33:597-607.) The acute-phase response (APR) is a systemic reaction of the organism to nonspecific inflammatory stimuli that is ac- companied by increased production of a cytokine cascade, which includes, among others, tumor necrosis factor a (TNF- a), interleukin 1 (IL-1), IL-6, and oncostatin M. 1-5 This cyto- kine cascade, which appears to be triggered by TNF-a, is responsible for inducing changes in the expression of a set of hepatocyte-specific proteins named the acute phase pro- teins. 1-5 Although the APR could be beneficial for the host, and some APR cytokines play a key role in liver cell regener- ation, 6-12 patients with alcohol-induced hepatitis that have several manifestations of the APR, including high blood levels of APR cytokines, develop cirrhosis. 13-15 These findings strongly suggest that in some instances, the APR and/or its cytokines may also have harmful effects and contribute to the development of liver fibrosis. 16 The molecular mechanisms that determine whether the actions exerted by the APR cyto- kines are beneficial (i.e., enhance hepatocyte proliferation) or harmful (i.e., induce liver fibrosis) to the patient have not been established and, therefore, need to be further investi- gated. Hepatic stellate cells (HSCs) are the main producers of ex- tracellular matrix components in normal and cirrhotic liver 17 and, thus, could play a key role in collagen gene expression induced during the APR. However, most molecular studies pertaining to the APR have been performed using either total liver and/or isolated hepatocytes. 1-6 Thus, the role of HSCs in the APR has not been investigated, except in some studies in which cultured cells were used. 18,19 We have previously shown that IL-6 up-regulates expres- sion of a1(I) collagen messenger RNA (mRNA) in HSCs by a transcriptional and protein synthesis– dependent mecha- nism. 16 Moreover, we showed that the increased expression of a1(I) collagen mRNA that occurs in rats with an APR can be prevented by the administration of a neutralizing antibody to IL-6, but not by a nonspecific immunoglobulin. 20 In addition, others have shown that another APR cytokine, oncostatin M, induces expression of a1(I) collagen mRNA in cultured hu- man HSCs. 21 Taken together, these data strongly suggest that Abbreviations: APR, acute phase response; TNF-a, tumor necrosis factor a; IL-6, interleukin-6; HSC, hepatic stellate cell; mRNA, messenger RNA; TIMP-1, tissue inhib- itor of metalloproteinases 1; MMP, matrix metalloproteinase; TGF-b1, transforming growth factor b1; cDNA, complementary DNA; GAPDH, glyceraldehyde-3-phosphate- dehydrogenase; RT-PCR, reverse transcription-polymerase chain reaction. From the 1 Marion Bessin Liver Research Center, Departments of Medicine and 2 Pa- thology, Albert Einstein College of Medicine, Bronx, NY; 3 Instituto de Biologı´a Molec- ular en Medicina, Centro Universitario de Ciencias de la Salud, Universidad de Guadala- jara, Jalisco, Me ´xico; and 4 Brookdale Center in the Department of Biochemistry and Molecular Biology, Mount Sinai School of Medicine, New York, NY. Received July 17, 2000; accepted December 18, 2000. Supported by USPHS Grants RO1 AA09231 and RO1 AA10541 (M.R.), RO1 AA12196 (P.G.), and by Grant 28832-M from Consejo Nacional de Ciencia y Tecnologı ´a, Me ´xico (J.A.B.). N.N.’s present address is the Department of Biochemistry and Molecular Biology, Mount Sinai School of Medicine, New York, NY. Address reprint requests to: Marcos Rojkind, M.D., Ph.D., Professor of Medicine and Pathology, Marion Bessin Liver Research Center, U-625, Albert Einstein College of Medicine, 1300 Morris Park Avenue, Bronx, NY 10461. E-mail: [email protected]; fax: 718-430-8975. Copyright © 2001 by the American Association for the Study of Liver Diseases. 0270-9139/01/3303-0016$35.00/0 doi:10.1053/jhep.2001.22520 597
Transcript of Rat hepatic stellate cells contribute to the acute-phase response with increased expression of...

Rat Hepatic Stellate Cells Contribute to the Acute-Phase ResponseWith Increased Expression of a1(I) and a1(IV) Collagens, TissueInhibitor of Metalloproteinase-1, and Matrix-Metalloproteinase-2
Messenger RNAs
NATALIA NIETO,1 JOSE A. DOMINGUEZ-ROSALES,1 LUIS FONTANA,1 ADRIANA SALAZAR,3 JUAN ARMENDARIZ-BORUNDA,3
PATRICIA GREENWEL,4 AND MARCOS ROJKIND1,2
The acute-phase response (APR) represents a systemicreaction of the organism to multiple nonspecific inflamma-tory stimuli. In general, it is protective for the host, andhepatocytes are the main cells responding with alterationsin the expression of a set of liver-specific proteins named theacute-phase proteins. We have previously shown that al-though a turpentine-induced APR is not fibrogenic per se, itenhances collagen deposition in rats treated with CCl4 andup-regulates expression of hepatic a1(I) collagen and tissueinhibitor of metalloproteinases 1 (TIMP-1) messenger RNAs(mRNAs). In this report we extended our studies andshowed that turpentine induced, in a time-dependent man-ner, expression of a1(I) and a1(IV) collagens, TIMP-1, andmatrix-metalloproteinase 2 (MMP-2) mRNAs. We furthershowed that expression of these mRNAs occurs in hepaticstellate cells, but not in hepatocytes obtained 6 hours afterthe induction of an APR episode. These changes were ac-companied by increased blood levels of tumor necrosis fac-tor a (TNF-a) and interleukin 6 (IL-6) without noticeableimmediate changes in the expression of their respective mR-NAs in the liver. In contrast to CCl4-induced liver damage,turpentine alone, whether administered as a single dose oras a weekly dose for 3 weeks did not up-regulate expressionof transforming growth factor b1 (TGF-b1) mRNA and didnot result in excess collagen deposition. Overall, these find-
ings suggest that collagen deposition in the livers of ratswith repeated APR episodes may be enhanced only whengiven together with a fibrogenic stimulus that activates he-patic stellate cells (HSCs) and/or up-regulates TGF-b1mRNA expression. (HEPATOLOGY 2001;33:597-607.)
The acute-phase response (APR) is a systemic reaction ofthe organism to nonspecific inflammatory stimuli that is ac-companied by increased production of a cytokine cascade,which includes, among others, tumor necrosis factor a (TNF-a), interleukin 1 (IL-1), IL-6, and oncostatin M.1-5 This cyto-kine cascade, which appears to be triggered by TNF-a, isresponsible for inducing changes in the expression of a set ofhepatocyte-specific proteins named the acute phase pro-teins.1-5 Although the APR could be beneficial for the host,and some APR cytokines play a key role in liver cell regener-ation,6-12 patients with alcohol-induced hepatitis that haveseveral manifestations of the APR, including high blood levelsof APR cytokines, develop cirrhosis.13-15 These findingsstrongly suggest that in some instances, the APR and/or itscytokines may also have harmful effects and contribute to thedevelopment of liver fibrosis.16 The molecular mechanismsthat determine whether the actions exerted by the APR cyto-kines are beneficial (i.e., enhance hepatocyte proliferation) orharmful (i.e., induce liver fibrosis) to the patient have notbeen established and, therefore, need to be further investi-gated.
Hepatic stellate cells (HSCs) are the main producers of ex-tracellular matrix components in normal and cirrhotic liver17
and, thus, could play a key role in collagen gene expressioninduced during the APR. However, most molecular studiespertaining to the APR have been performed using either totalliver and/or isolated hepatocytes.1-6 Thus, the role of HSCs inthe APR has not been investigated, except in some studies inwhich cultured cells were used.18,19
We have previously shown that IL-6 up-regulates expres-sion of a1(I) collagen messenger RNA (mRNA) in HSCs by atranscriptional and protein synthesis–dependent mecha-nism.16 Moreover, we showed that the increased expression ofa1(I) collagen mRNA that occurs in rats with an APR can beprevented by the administration of a neutralizing antibody toIL-6, but not by a nonspecific immunoglobulin.20 In addition,others have shown that another APR cytokine, oncostatin M,induces expression of a1(I) collagen mRNA in cultured hu-man HSCs.21 Taken together, these data strongly suggest that
Abbreviations: APR, acute phase response; TNF-a, tumor necrosis factor a; IL-6,interleukin-6; HSC, hepatic stellate cell; mRNA, messenger RNA; TIMP-1, tissue inhib-itor of metalloproteinases 1; MMP, matrix metalloproteinase; TGF-b1, transforminggrowth factor b1; cDNA, complementary DNA; GAPDH, glyceraldehyde-3-phosphate-dehydrogenase; RT-PCR, reverse transcription-polymerase chain reaction.
From the 1Marion Bessin Liver Research Center, Departments of Medicine and 2Pa-thology, Albert Einstein College of Medicine, Bronx, NY; 3Instituto de Biologıa Molec-ular en Medicina, Centro Universitario de Ciencias de la Salud, Universidad de Guadala-jara, Jalisco, Mexico; and 4Brookdale Center in the Department of Biochemistry andMolecular Biology, Mount Sinai School of Medicine, New York, NY.
Received July 17, 2000; accepted December 18, 2000.Supported by USPHS Grants RO1 AA09231 and RO1 AA10541 (M.R.), RO1 AA12196
(P.G.), and by Grant 28832-M from Consejo Nacional de Ciencia y Tecnologıa, Mexico(J.A.B.).
N.N.’s present address is the Department of Biochemistry and Molecular Biology,Mount Sinai School of Medicine, New York, NY.
Address reprint requests to: Marcos Rojkind, M.D., Ph.D., Professor of Medicine andPathology, Marion Bessin Liver Research Center, U-625, Albert Einstein College ofMedicine, 1300 Morris Park Avenue, Bronx, NY 10461. E-mail: [email protected];fax: 718-430-8975.
Copyright © 2001 by the American Association for the Study of Liver Diseases.0270-9139/01/3303-0016$35.00/0doi:10.1053/jhep.2001.22520
597

the APR could be fibrogenic and that members of the IL-6super family are fibrogenic both in vitro and in vivo. Indeed,the superimposition of weekly APR episodes to rats receivingCCl4 to produce cirrhosis resulted in an enhancement of thefibrogenic process.16 This was manifested by increased livercollagen content, as well as by enhanced expression of mRNAsencoding for a1(I) collagen and tissue inhibitor of metallo-proteinases (TIMP-1).16 However, in the studies describedabove, the temporal relationships between blood and liverlevels of the APR cytokines TNF-a and IL-6, or between thesecytokines and expression of a1(I) collagen mRNA was notinvestigated. Moreover, the possible role of HSCs in vivo inexpressing other extracellular matrix components such astype IV collagen, or in proteins involved in its remodeling, i.e.,matrix-metalloproteinase-2 (MMP-2), TIMP-1, or transform-ing growth factor b1 (TGF-b1) during the APR was not ana-lyzed. Thus, to further study the role of the APR and its cyto-kines in enhancing expression of extracellular matrixcomponents by HSCs in vivo, we induced an APR in rats anddetermined levels of expression of a1(I) and a1(IV) collag-ens, TGF-b1, MMP-2, and TIMP-1 mRNAs in freshly isolatedHSCs obtained 6 hours after the induction of an APR. Ourresults show that a single APR episode suffices to induce inHSCs, but not in hepatocytes, a transient expression of a1(I)and a1(IV) collagen, MMP-2, and TIMP-1 mRNAs in a time-dependent manner. Interestingly, up-regulation of thesemRNAs follows a rapid and sharp raise in blood TNF-a levelsand were accompanied by increased blood levels of IL-6. Onthe other hand, we also showed that in contrast to what hap-pens in fibrotic liver, during the APR, expression of TGF-b1mRNA was not induced either in HSCs or hepatocytes.
MATERIALS AND METHODS
Animals. Male Fisher or Sprague-Dawley rats (100 6 10 g)(Charles River Breeding Laboratories, Wilmington, MA) were usedfor all experiments and, except for albumin mRNA whose expressionis down-regulated during the APR in Sprague-Dawley but not inFisher rats, all findings were similar. All animals used in the presentstudy received humane care in compliance with the guidelines of theNIH and the Animal Institute of Albert Einstein College of Medicine.Rats had free access to water and were fed ad libitum with a rodentchow diet. All animals were food deprived the night prior to perform-ing the experiments.
Induction of Liver APR. Rats were divided into 7 groups (n 5 4each). Animals in the control group did not receive any treatmentand those in the 6 remaining groups received a single intradermalinjection of 0.25 mL of turpentine (EM Science, Cherry Hill, NJ) per100 g of body weight. Animals were sacrificed under ethyl etheranesthesia at 0, 3, 6, 12, 24, and 48 hours post-turpentine adminis-tration; the blood was collected by aortic puncture and transferredinto tubes containing 8 mmol/L Na2-EDTA. For determination ofserum IL-6 levels, blood was collected as described above and cen-trifuged at 1,000g for 15 minutes. Livers were immediately removed,frozen in liquid nitrogen and stored at 270°C.
Induction of Acute-Phase Episodes in CCl4-Treated Rats. Rats were di-vided into 4 groups (n 5 4 each). The control group did not receiveany treatment. The acute-phase group received once a week an in-tradermal injection of 0.25 mL of turpentine per 100 g of bodyweight. Liver fibrosis was induced in the third group by intraperito-neal injection 3 times a week of 0.15 mL CCl4 (Fisher Scientific,Pittsburgh, PA) in mineral oil (Sigma, St. Louis, MO) (1:5) as previ-ously described.16 The fourth group received the combined treat-ment of CCl4 to produce fibrosis and turpentine to induce acutephase episodes at the same doses as described above. Rats were sac-rificed under ethyl ether anesthesia after 9 doses of CCl4 and/or 3APR episodes. All specimens were obtained 7 days after the last CCl4
dose and/or 72 hours after the last turpentine administration. Bloodwas obtained as described above, and livers were removed, frozen inliquid nitrogen, and stored at 270°C.
Isolation of HSCs and Hepatocytes. Three control and 3 turpentine-treated rats were used for these experiments. Six hours after theadministration of turpentine, hepatocytes and HSCs were isolated byliver perfusion with bacterial collagenase according to previouslypublished protocols.18,19 Cell viability was assessed by the trypanblue exclusion test and was found to be 98%. Purity of each fractionwas determined as described previously and shown to be 95% forhepatocytes and 92% for HSCs.
Northern Analysis. Total RNA was extracted from either 50 mg ofliver or from freshly isolated cells.22 Ten micrograms of RNA fromliver or 3 mg from freshly isolated cells were electrophoresed underdenaturing conditions in 1% agarose/formaldehyde gels and trans-ferred onto Gene-screen nylon membranes (Dupont, Boston, MA).The following 32P-labeled complementary DNA (cDNA) probes wereused: rat a1(I)23 and a1(IV) collagens,24 fibronectin,25 MMP-13,26
and MMP-2,27 fibrinogen,28 albumin,29 TIMP-1,30 and human glyc-eraldehyde-3-phosphate dehydrogenase (GAPDH) (ATCC, Rock-ville, MD). All blots were washed under stringent conditions, exceptwhen hybridized to human GAPDH cDNA when washes were per-formed under nonstringent conditions.31 Quantitative analysis ofsignals generated by X-ray film exposure was performed by photo-densitometry. Values were corrected using the GAPDH signal as acontrol and refer to mean 6 SE (n 5 4).
Plasma levels of TNF-a were measured using a rat TNF-a ultrasensitive cytoscreen US ELISA Kit (Biosource, Camarillo, CA) ac-cording to the instructions provided by the manufacturer. Values areexpressed as pg/mL of plasma and refer to mean 6 SE (n 5 4).
Serum Levels of IL-6. IL-6 levels were determined measuring theIL-6–dependent proliferation of the B9 murine plasmacytoma cellline, as previously described32-34 using serial dilutions of serum. Oneunit of activity is defined as the dilution that gives half-maximal B9cell growth in 72 hours and corresponds to approximately 2 ng/LrhIL-6.
Reverse Transcription-Polymerase Chain Reaction. First-strand cDNAwas synthesized using 2 mg of total RNA, 1 mL of placental RNaseinhibitor (Ambion, Austin, TX), 1 mL of reverse transcription-poly-merase chain reaction (RT-PCR) buffer (Roche Diagnostics, India-napolis, IN), 2.5 mmol/L dNTPs (Roche), 500 ng of random primers,and 1 U of Moloney Murine leukemia virus reverse transcriptase(Roche) in a final volume of 20 mL. The reaction was carried out for60 minutes at 42°C.
The PCR reaction was performed as previously described35 usingthe following primer pairs: 59 CGA GTG ACA AGC CCG TAG CC 39(TNF-a forward); 59 GGA TGA ACA CGC CAG TCG CC 39 (TNF-areverse); 59ATG GAT GAC GAT ATC GCT 39 (b actin forward); 59TGG ACT GTC TGA TGG AGTA 39 (b actin reverse). For TGF-b1and 18S, the cDNA was amplified using commercially available setsof primers (Ambion). Each PCR reaction contained 0.4 mmol/L ofspecific primer pairs, 200 mmol/L dNTPs, 1 U of platinum Taq highfidelity polymerase (Gibco, Grand Island, NY), 1 mL high fidelityPCR buffer (Gibco), and 2 mCi of radiolabeled dCTP (Amersham,Piscataway, NJ) in a total volume of 50 mL. Amplification reactionswere cycled as follows: initial denaturation at 95°C for 5 minutes; 30cycles: 95°C for 1 minute, 60°C for 1 minute, and 72°C for 1 minute;final extension was carried out at 72°C for 5 minutes. The PCRproducts were resolved on a 6% nondenaturating acrylamide gel,dried, and quantified using a Phosphorimager and the Image Quantsoftware (Molecular Dynamics, Sunnyvale, CA). Under these exper-imental conditions, results obtained were within the linear range.35
Statistical Analysis. Statistical analysis was performed with the Stu-dent’s t test.
RESULTS
To induce an APR and avoid producing overt liver injury weused half the recommended dose of turpentine.36,37 There-
598 NIETO ET AL. HEPATOLOGY March 2001

fore, we first determined whether the rats had indeed an APRand measured blood levels of TNF-a and IL-6 and liversteady-state levels of TNF-a, IL-6, and fibrinogen mRNAs. Asillustrated in Fig. 1A, within the first 5 minutes after turpen-tine administration to rats, there was a sharp increment inplasma TNF-a levels from 13.9 to 53.7 pg/mL (P , .001). Thisincrease was transient, since TNF-a values decreased to 24.5pg/mL by 90 minutes (P , .005) and reached normal values
by 24 hours. Semi-quantitative RT-PCR studies using totalliver RNA revealed a significant increase in TNF-a mRNAexpression by 12 hours (3.7-fold above controls) (P , .001)(Fig. 1B). However, mRNA levels did not correlate with bloodcytokine concentrations (Fig. 1A). Although TNF-a mRNAvalues decreased to 2.5-fold above control levels by 24 hours(P , .05), they remained elevated by 48 hours (3.2-fold; P ,.05). As shown in Fig. 1C, serum IL-6 levels, as determined bythe IL-6–dependent proliferation of the B9 murine plasmocy-toma cell line, in control untreated animals were undetect-able. In contrast with these findings, there was a significantincrease in IL-6 levels in serum obtained from rats treatedwith turpentine for 6 hours (4.5 ng/L 3 103), the same timepoint selected to analyze mRNA expression in HSCs isolatedfrom turpentine-treated rats (see below) (Fig. 1C). These lev-els of serum IL-6 are in agreement with previous publicationsin which turpentine was used to induce an APR in rats.38 IL-6mRNA levels in total liver, analyzed by PCR studies were verylow and similar in all samples analyzed (data not shown).Increased levels of blood TNF-a and IL-6 were accompaniedby up-regulation of fibrinogen mRNA, a bona fide positive APRmRNA. As shown in Fig. 2A, fibrinogen mRNA reached itsmaximum value at 12 hours (9.3-fold above control; P ,.001) and decreased thereafter to 3-fold above controls by 48hours (P , .005).
Once we established that the rats were indeed in an APR, weanalyzed expression of several genes related to extracellularmatrix formation. As shown in Fig. 2B, a1(I) collagen mRNAexpression was induced 7.6-fold after 6 hours and remained3.3-fold above control values at 48 hours (P , .001). Changesin expression of a1(I) collagen were not accompanied by adetectable increase in either MMP-13 or TIMP-1 mRNAs(data not shown). Fibronectin mRNA levels showed the samepattern of induction as fibrinogen mRNA with an 11.2-foldincrease above controls at 12 hours (P , .001) (Fig. 2C).
Type IV collagen is a key component of the space of Disse.Since collagenization of this space and formation of a base-ment membrane are important events in liver fibrogenesis, weinvestigated whether a1(IV) collagen and MMP-2 mRNA ex-pression was also induced during the APR. Since a1(IV) col-lagen and MMP-2 mRNA expression was not detected in thecontrol group, results were expressed in arbitrary units. Asshown in Fig. 3A, rats with an APR episode showed a signifi-cant induction of a1(IV) collagen mRNA with maximal valuesat 12 hours (P , .001). Although levels of expression of thismRNA decreased thereafter, they were still detectable at 48
4™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™™FIG. 1. (A) Plasma levels of TNF-a in rats with an APR episode. Plasma
TNF-a concentration was measured by ELISA at 0, 5 minutes, and 1.5, 3, 6,12, 24, and 48 hours after the intradermal injection of a single dose of tur-pentine. A sharp, but transient increase in plasma TNF-a levels was observedwithin the first 5 minutes, decreasing at 90 minutes and returning to normalvalues by 24 hours. Results are expressed as pg/mL, and refer to mean 6 SE(n 5 4). *P , .05; **P , .005; ***P , .001 when compared to values at 0hours. (B) TNF-a semiquantitative RT-PCR of RNA from rats with an APRepisode. Liver TNF-a expression showed a maximum value of 3.7-fold abovecontrol at 12 hours and remained elevated above control values by 48 hours.All values were corrected for the b-actin signal as described in the Materialsand Methods. Results are expressed as fold increase above control, and referto mean 6 SE (n 5 4). *P , .05; **P , .005; ***P , .001 when comparedwith the control group. (C) Serum levels of IL-6 in rats with APR. Serum IL-6concentration was determined 6 hours after the injection of turpentine. Al-though no levels of IL-6 were detected in control rats, 4.5 ng/L 3 103 werefound 6 hours after turpentine administration (n 5 4).
HEPATOLOGY Vol. 33, No. 3, 2001 NIETO ET AL. 599

FIG. 2. Expression of fibrinogen, a1(I) collagen, and fibronectin mRNAs in livers of rats with an APR episode. Left panel: (A) The histogram depicts asummary of Northern assays performed with total RNA obtained from 4 livers; a 9.3-fold increase in fibrinogen mRNA above control values was observed at12 hours and a decrease to 3-fold at 48 hours. (B) The histogram shows a 7.6-fold increase in a1(I) collagen mRNA after 6 hours and the subsequent decreaseto 3.3-fold above the control by 48 hours. (C) Maximum expression of fibronectin mRNA was observed at 12 hours. Right panel: Representative Northern blotsof the results shown on the left. All values were corrected for differences in loading using the GAPDH signal as described in the Materials and Methods. Resultsare expressed as fold increase above control for fibrinogen and fibronectin with control assigned a value of 1 and in arbitrary units for a1(I) collagen and referto mean 6 SE. *P , .05; **P , .005; ***P , .001.
600 NIETO ET AL. HEPATOLOGY March 2001

hours post-turpentine administration (P , .005). As shown inFig. 3B, a parallel elevation in the expression of MMP-2mRNA, a gelatinase involved in type IV collagen degradation,was observed after 3 hours (P , .005). Its expression was alsotransient and significantly decreased to control levels by 48hours. No detectable levels of TIMP-1 mRNA were found insamples of total liver RNA at all time points investigated (datanot shown).
The results described above suggest that expression ofa1(I) and a1(IV) collagens, fibronectin, and MMP-2 mRNAsincrease in livers of rats with an APR. However, these exper-iments did not reveal which liver cell type is responsible forthe induction of such mRNAs. Hence, to study the role ofhepatocytes and HSCs in the expression of these mRNAs, weisolated total RNA from hepatocytes and HSCs obtained fromrats treated with or without turpentine for 6 hours. This timepoint was selected because, as shown in Figs. 2 and 3, steady-state levels of the mRNAs investigated were already elevatedseveral fold above control values. As shown in Fig. 4A and B,
fibrinogen mRNA was detected in hepatocytes of control ratsand its expression was significantly increased after turpentineadministration. As expected, fibrinogen mRNA was not ex-pressed in HSCs from control rats. Surprisingly, however, lowbut nonetheless measurable levels of fibrinogen mRNA weredetected in HSCs obtained from turpentine-treated rats. Al-though we have no explanation for these findings, it is possi-ble that the HSC fraction was contaminated with small hepa-tocytes that do not sediment at low speed centrifugation andare resistant to pronase digestion.
As illustrated in Fig. 4A, no signal for a1(I) collagen,MMP-2, or TIMP-1 mRNAs was detected in hepatocytes fromcontrol or turpentine-treated rats. In contrast to these find-ings, HSCs expressed variable levels of a1(I) and a1(IV) col-lagen mRNAs, and these were significantly up-regulated incells obtained from livers of rats with an APR episode (Fig. 4Cand D). On the other hand, although a weak signal for a1(IV)collagen mRNA was detected in hepatocytes from control ratsand its expression was slightly increased after turpentine ad-
FIG. 3. a1(IV) collagen and MMP-2 mRNA expression in livers of rats with an APR episode. Left panel: (A) Summary histogram of 4 experiments showsa maximum increase in a1(IV) collagen mRNA expression at 12 hours post-turpentine administration. (B) Histogram showing an increase in the expressionof MMP-2 mRNA at 3 hours after turpentine treatment, with maximum values at 6 hours. Right panel: Representative Northern blots are shown. All values werecorrected for differences in loading and are expressed in arbitrary units and refer to mean 6 SE. *P , .05; **P , .005; ***P , .001.
HEPATOLOGY Vol. 33, No. 3, 2001 NIETO ET AL. 601
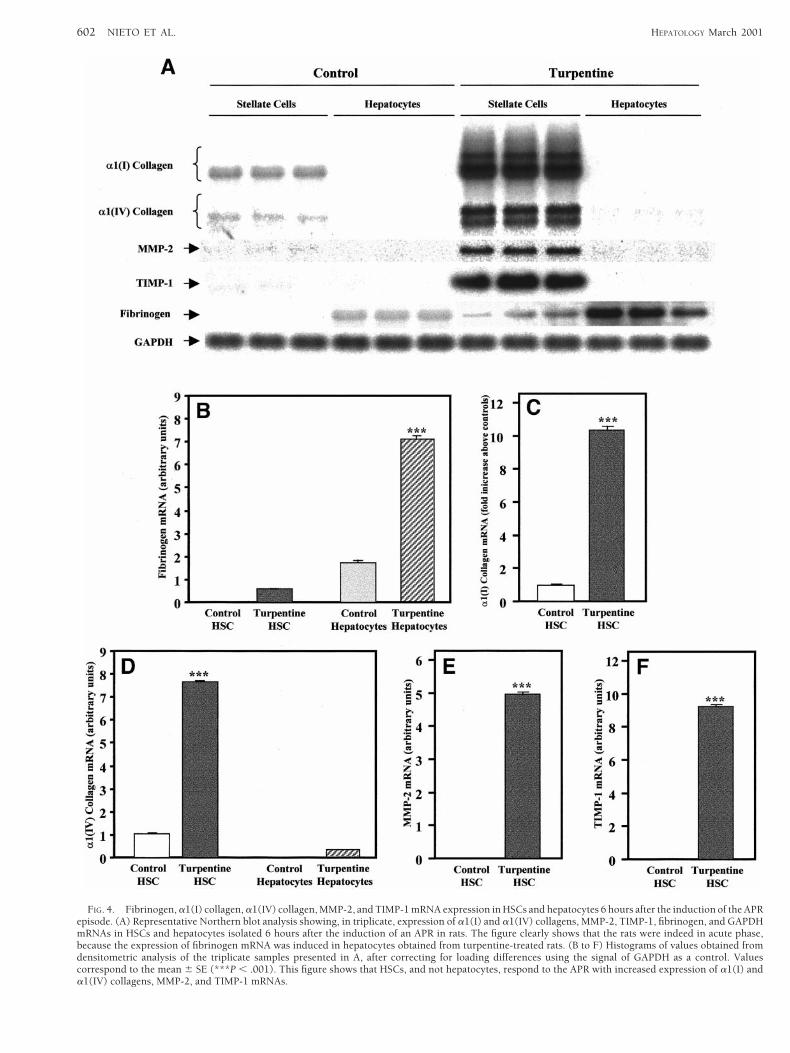
FIG. 4. Fibrinogen, a1(I) collagen, a1(IV) collagen, MMP-2, and TIMP-1 mRNA expression in HSCs and hepatocytes 6 hours after the induction of the APRepisode. (A) Representative Northern blot analysis showing, in triplicate, expression of a1(I) and a1(IV) collagens, MMP-2, TIMP-1, fibrinogen, and GAPDHmRNAs in HSCs and hepatocytes isolated 6 hours after the induction of an APR in rats. The figure clearly shows that the rats were indeed in acute phase,because the expression of fibrinogen mRNA was induced in hepatocytes obtained from turpentine-treated rats. (B to F) Histograms of values obtained fromdensitometric analysis of the triplicate samples presented in A, after correcting for loading differences using the signal of GAPDH as a control. Valuescorrespond to the mean 6 SE (***P , .001). This figure shows that HSCs, and not hepatocytes, respond to the APR with increased expression of a1(I) anda1(IV) collagens, MMP-2, and TIMP-1 mRNAs.
602 NIETO ET AL. HEPATOLOGY March 2001

ministration, the differences observed were marginal andwere not statistically significant (Fig. 4D). No signal forMMP-2 or TGF-b1 mRNA was detected in HSCs from controlrats. However, while the former mRNA was significantly up-regulated in HSCs from turpentine-treated rats (Fig. 4E), thelatter remained undetectable (data not shown). Interestingly,although TIMP-1 mRNA levels were not detectable in totalRNA from control or turpentine-treated rats (data notshown), very low levels were detected in RNA obtained fromcontrol HSCs. Moreover, this mRNA was significantly up-regulated in HSCs from turpentine-treated rats (Fig. 4F).
After establishing the changes in blood TNF-a and serumIL-6 and liver mRNA levels, expression of a1(I) and a1(IV)collagens, MMP-2, TIMP-1, and TGF-b1 mRNAs induced by asingle APR episode, we analyzed these parameters in ratstreated once a week for 3 weeks with turpentine, rats treatedwith CCl4 3 times a week for 3 weeks to produce liver fibrosis,and in rats receiving the combined treatment. Based on ourfindings and those of others,39,40 namely the transient expres-sion of the various mRNAs investigated after turpentine oracute CCl4 administration, in the chronic experiments ratswere sacrificed 7 days after the last turpentine administrationand/or 3 days after the final dose of CCl4. Accordingly, mRNAvalues reflect alterations induced by chronic treatment andnot those induced by the last dose of turpentine or CCl4. Asshown in Fig. 5, plasma TNF-a levels increased from 13.8 60.63 pg/mL in untreated control animals to 37.1 6 2.23 pg/mLin those receiving turpentine (P , .05). It is noteworthy tomention that this value, which was obtained 7 days post-turpentine administration, is significantly higher (;2- to3-fold) than that found by 48 hours in rats receiving a singleAPR episode, thus suggesting a sustained effect in animalsreceiving multiple APR stimuli. TNF-a values in CCl4-treatedrats were normal (14.1 6 0.84 pg/mL) and were similar tocontrol values. The combined treatment of CCl4 to producecirrhosis plus a weekly induction of APR episodes increasedTNF-a plasma levels to 55.7 6 3.1 pg/mL, a value signifi-cantly higher than that found in rats with APR (P , .005). Onthe other hand, IL-6 mRNA levels, determined by RT-PCRrevealed that this mRNA was increased in all the experimentalgroups. However, differences among them were not signifi-cant (Fig. 5B).
The data regarding the expression of extracellular matrix–related mRNAs confirmed our previous observations.16 Asshown in Fig. 6A and B, we confirmed that a1(I) collagen andTIMP-1 mRNAs are up-regulated in rats treated for 3 weekswith CCl4, and that these mRNAs are further up-regulated inrats receiving turpentine plus CCl4 (P , .001; P , .001, re-spectively). In addition, we showed that expression of a1(IV)collagen and MMP-2 mRNAs is also up-regulated in CCl4-treated rats and that their levels are significantly higher in ratsreceiving the combined treatment (P , .001, P , .001, respec-tively) (Fig. 6C and D).
To investigate whether TGF-b1 plays a role, if any, in theAPR-mediated up-regulation of the collagen genes investi-gated, TGF-b1 mRNA levels were measured in livers of ratschronically treated with turpentine, CCl4, and turpentine plusCCl4. As shown in Fig. 7, TGF-b1 mRNA was only up-regu-lated in livers of rats receiving CCl4 alone or in combinationwith turpentine (P , .001). However, TGF-b1 mRNA levelswere not statistically different in livers receiving the combinedtreatment as compared with those treated only with CCl4.
DISCUSSION
TNF-a, IL-6, and IL-1 are 3 of the main cytokines involvedin the APR,1-5 which also play a key role in liver regenera-tion,6-12 and influence the expression and turnover of variousextracellular matrix components.16,20,21,41 It has been postu-lated that the APR, its cytokines and/or the proteins whoseexpression is modified by them, play an important role inmaintaining homeostasis.42,43 However, recent evidence sug-gests that in some instances, the APR and/or its cytokines mayenhance and/or accelerate pathologic processes.16,44-47 In-deed, patients with alcohol-induced hepatitis have clinical
FIG. 5. Expression of TNF-a in CCl4-treated rats with and without re-peated APR episodes. Rats were treated with either turpentine once a week for3 weeks, an intraperitoneal CCl4 injection 3 times a week for 3 weeks, or both.Blood TNF-a values, determined by ELISA, increased from 13.8 6 0.63pg/mL in the control group to 37.1 6 2.23 pg/mL in those receiving turpen-tine (P , .05). Cytokine blood levels in CCl4-treated rats remained un-changed, whereas in those receiving the combined treatment, TNF-a levelsincreased to 55.7 6 3.1 pg/mL. Results are expressed in pg/mL and refer tomean 6 SE (n 5 4). *P , .05; **P , .005. (B) IL-6 mRNA values, asdetermined by RT-PCR, show that although there is increased expression ofthis mRNA in livers of rats treated with CCl4 and CCl4 plus turpentine, therewere no significant differences among these 2 groups.
HEPATOLOGY Vol. 33, No. 3, 2001 NIETO ET AL. 603

manifestations of the APR and their blood APR cytokine levelsare increased.13-15 Moreover, although not all alcoholic pa-tients develop cirrhosis, a relatively large percent of thosewith alcohol-induced hepatitis develop it.13-15 In a rat modelof alcohol-induced liver fibrosis, levels of APR cytokines in-crease prior to the development of overt liver fibrosis.48 Inaddition, we have previously shown that the superimpositionof weekly APR episodes to rats receiving chronic treatmentwith CCl4 accelerates the deposition of collagen and, thus,enhances the development of liver fibrosis.16 Because rats re-ceiving weekly injections of turpentine for 3 weeks did notdevelop a sustained elevation in a1(I) collagen and TIMP-1mRNAs and there was no increase in total liver collagen, weconcluded that the APR, per se, is not fibrogenic.16 Con-versely, because those rats receiving the combined CCl4/tur-pentine had more liver fibrosis, contained more liver collagen,
and expressed higher levels of a1(I) collagen and TIMP-1mRNAs than those treated with CCl4 alone, we proposed thatthe APR enhances fibrosis in rats only when they are under-going liver injury.16 Because HSCs are already activated andproduce more collagen when isolated from CCl4-treated rats,and these cells respond to IL-6 administration with a signifi-cant elevation in a1(I) collagen and TGF-b1 mRNAs,20 wespeculated that the APR could be fibrogenic only when HSCsare already activated.
Based on the aforementioned data, the aim of the presentstudy was to establish the contribution of HSCs to the APR ingeneral, and to start elucidating some of the mechanismswhereby these cells increase their expression of extracellularmatrix components during the APR. Thus, based on theknown fact that TNF-a is one of the APR cytokines that up-regulates expression of IL-6,1-3 and IL-6 is one of the cytokines
FIG. 6. Expression of a1(I) col-lagen, a1(IV) collagen, MMP-2, andTIMP-1 mRNA in CCl4-treated ratswith and without repeated APR epi-sodes. Rats were treated with eitherturpentine once a week for 3 weeks,or an intraperitoneal CCl4 injection3 times a week for 3 weeks, or both,and total RNA was isolated and ana-lyzed by Northern blot. a1(I) colla-gen (A) and TIMP-1 (B) mRNAs areup-regulated in rats treated for 3weeks with CCl4 and these mRNAsare further up-regulated in rats re-ceiving turpentine plus CCl4. Theexpression of a1(IV) collagen (C)and MMP-2 (D) mRNAs is also up-regulated in CCl4-treated rats andtheir levels are significantly higher inrats receiving the combined treat-ment (P , .001). All values were cor-rected for loading differences usingGAPDH cDNA as a control. Resultsare expressed as arbitrary units andare means 6 SE of triplicate experi-ments (***P , .001).
604 NIETO ET AL. HEPATOLOGY March 2001

that induces a1(I) collagen gene expression in cultured ratHSCs,20 we first analyzed the temporal expression of TNF-aand IL-6 in blood and livers of rats with an APR episode. Inaddition, we performed experiments to investigate the contri-bution of HSCs to the expression of genes encoding someextracellular matrix components during the APR. Finally, be-cause we have previously shown that TGF-b1 is induced byIL-6 only in HSCs derived from cirrhotic livers, we deter-mined whether expression of this mRNA was elevated in liversof turpentine-treated rats, and whether it played a role inup-regulating a1(I) and a1(IV) collagen, MMP-2, andTIMP-1 mRNAs in livers of turpentine-treated rats.
Our studies revealed that rats treated acutely with turpen-tine were indeed in acute phase as manifested by the increasedexpression of fibrinogen mRNA in total RNA extracted fromlivers, as well as from hepatocytes. Moreover, because bloodTNF-a levels increased sharply prior to detecting a significantincrease in the expression of its messenger in the liver, thedata suggested the possibility that this cytokine may be re-leased from a preexisting pool. Indeed, it has been previouslyshown that a precursor form of TNF-a is present in cells andthat the active cytokine can be released by limited proteolysisby a specific TNF-a–converting enzyme named TACE.49
However, the cells that express this cytokine and/or TACEwere not analyzed, and thus this needs to be further investi-gated. On the other hand, our data clearly showed that there isan increase in serum IL-6 levels in rats treated with turpentinefor 6 hours. Although the changes in IL-6 were relativelymodest, they were, nonetheless, significant and were only de-tected in turpentine-treated rats and not in controls. Thesefindings are in agreement with data in the literature showingthat in contrast to endotoxin, turpentine induces a more mod-est cytokine response.38 However, in both cases, namely tur-pentine or endotoxin administration, cytokine levels are suf-ficient to induce the typical changes observed in the APR.
With regard to the role of HSCs in the APR, we showed thata single injection of turpentine sufficed to induce expressionof a1(I) and a1(IV) collagens, TIMP-1, and MMP-2 mRNAsby HSCs isolated 6 hours after turpentine administration. Be-
cause these cells were isolated directly from rats treated withturpentine and RNA extracted immediately upon isolation,the steady-state levels of a1(I) and a1(IV) collagens, MMP-2,TIMP-1, and TGF-b1 mRNAs found reflect the level of theirexpression in vivo. These findings show that HSCs are impor-tant mediators of the APR and contribute, together with hepa-tocytes, to the overall response of the host to nonspecific in-flammatory stimuli.
In response to a single APR stimulus, HSCs express in-creased levels of mRNAs encoding extracellular matrix com-ponents that are found elevated in cirrhotic livers. Indeed,type I collagen is one of the main components of scar tissueand type IV collagen is a key protein involved in the capillar-ization of the space of Disse.50 Nonetheless, no expression ofTGF-b1 mRNA was detected and no overt deposition of col-lagen was found as determined by histologic and/or histo-chemical analysis of the liver (data not shown). These dataconfirmed our previous findings suggesting that the APR byitself is not fibrogenic and that additional events, which mayoccur after CCl4 administration, are required for the APR toenhance the fibrogenic process. Because TGF-b1 is the classicfibrogenic cytokine and its expression is up-regulated in liversof cirrhotic animals and patients, we further investigated thiscytokine.51,52 Thus, we analyzed expression of TGF-b1mRNA in total liver RNA from rats treated with a single doseof turpentine to induce the APR, in rats treated for 3 weekswith CCl4 to induce liver fibrosis, and in rats treated with CCl4
for 3 weeks plus a weekly induction of APR episodes. Asshown in this report, a single dose of turpentine up-regulatedexpression of a1(I) and a1(IV) collagens, TIMP-1, andMMP-2 in HSCs, while no TGF-b1 mRNA was detected.These findings suggest that TGF-b1 does not play a role in theup-regulation of the expression of this mRNA in turpentine-treated rats. However, further studies measuring TGF-b1 pro-tein levels may be required to confirm this hypothesis.
On the other hand, our result showing increased produc-tion of IL-6, together with our previous findings showing thata neutralizing antibody to IL-6 prevented a1(I) collagenmRNA up-regulation induced by turpentine, suggest that al-
FIG. 7. Expression of TGF-b in CCl4-treated rats with and without repeated APR episodes. Summary histograms of 4 experiments showing expression ofTGF-b1 mRNA in livers of control rats, rats treated with turpentine, CCl4, or turpentine plus CCl4. TGF-b1 expression increased only in livers of rats treatedwith CCl4 alone or in combination with turpentine. Results are expressed in arbitrary units and were corrected for loading differences using an 18S ribosomalprotein cDNA as a control. Values are means 6 SE (***P , .001).
HEPATOLOGY Vol. 33, No. 3, 2001 NIETO ET AL. 605

terations in collagen gene expression in HSCs observed afterturpentine administration are due to the production of IL-6.However, we cannot rule that other cytokines, including on-costatin M may play a role in this process.21 In the chronicstudies we confirmed our previous observations showing thatthe combined treatment of CCl4 plus turpentine is more fibro-genic that CCl4 treatment alone.16 In addition we furthershowed that the acute or chronic treatment with turpentineresults in up-regulation of a1(I) collagen and MMP-2 mRNAs.While the former is involved in basement membrane forma-tion, the latter plays a role in its remodeling.53 In this chronicmodel, levels of expression of TGF-b1 mRNA are elevatedonly in rats receiving CCl4 or the combined treatment of CCl4
plus a weekly APR episode, but not in control animals receiv-ing only a weekly induction of an APR episode. Interestingly,although TGF-b1 mRNA levels were higher in animals withthe combined treatment, they were not significantly differentfrom values obtained with rats treated only with CCl4. Thus,although the data support the notion that the APR enhancesfibrogenesis in CCl4-treated rats, the molecular eventswhereby this occurs remain to be fully elucidated. BecauseHSCs in rats treated with CCl4 are already activated in vivo, itis conceivable to suggest that the fibrogenic actions of the APRtake place only in livers in which HSCs are activated andcommitted to produce collagen. Consequently, it could bespeculated that in patients with chronic liver disease, in whichHSCs are already activated and/or already have various de-grees of liver fibrosis, repeated APR episodes occurring duringacute liver injury (i.e., alcohol-induced hepatitis) or second-ary to nonspecific inflammatory events, may exacerbate liverfibrogenesis. This possibility is currently being explored inour laboratory.
Acknowledgment: The authors express their appreciationto Dr. Gary Lebretton for helping in the determination ofserum IL-6 levels.
REFERENCES
1. Heinrich PC, Castell JV, Andus T. Interleukin-6 and the acute phaseresponse. Biochem J 1990;265:621-636.
2. Baumann H, Gauldie J. The acute phase response. Immunol Today 1994;15:74-80.
3. Gauldie J, Richards C, Baumann H. IL6 and the acute phase reaction. ResImmunol 1992;143:755-759.
4. Richards CD, Brown TJ, Shovab M, Baumann H, Gauldie J. Recombinantoncostatin M stimulates the production of acute phase proteins in HepG2cells and rat primary hepatocytes in vitro. J Immunol 1992;148:1731-1736.
5. Gabay C, Kushner I. Acute-phase proteins and other systemic responsesto inflammation. N Engl J Med 1999;340:448-454.
6. Cressman DE, Greenbaum LE, DeAngelis RA, Ciliberto G, Furth EE,Poli V, Taub R. Liver failure and defective hepatocyte regeneration ininterleukin-6–deficient mice. Science 1996;274:1379-1383.
7. Webber EM, Bruix J, Pierce RH, Fausto N. Tumor necrosis factor primeshepatocytes for DNA replication in the rat. HEPATOLOGY 1998;28:1226-1234.
8. Yamada Y, Webber EM, Kirillova I, Peschon JJ, Fausto N. Analysis of liverregeneration in mice lacking type 1 or type 2 tumor necrosis factor re-ceptor: requirement for type 1 but not type 2 receptor. HEPATOLOGY 1998;28:959-970.
9. Kirillova I, Chaisson M, Fausto N. Tumor necrosis factor induces DNAreplication in hepatic cells through nuclear factor kappaB activation. CellGrowth Differ 1999;10:819-828.
10. Diehl AM. Effect of ethanol on tumor necrosis factor signaling duringliver regeneration. Clin Biochem 1999;32:571-578.
11. Diehl AM. Cytokine regulation of liver injury and repair. Immunol Rev2000;174:160-171.
12. Fausto N. Liver regeneration. J Hepatol 2000;32:19-31.
13. McClain CJ, Barve S, Barve S, Deaciuc I, Hill DB. Tumor necrosis factorand alcoholic liver disease. Alcohol Clin Exp Res 1998;22:248S-252S.
14. McClain CJ, Barve S, Deaciuc I, Kugelmas M, Hill DB. Cytokines inalcoholic liver disease. Semin Liver Dis 1999;19:205-219.
15. Diehl AM. Cytokines and the molecular mechanisms of alcoholic liverdisease. Alcohol Clin Exp Res 1999;23:1419-1424.
16. Greenwel P, Rojkind M. Accelerated development of liver fibrosis inCCl4-treated rats by the weekly induction of acute phase response epi-sodes: upregulation of a1(I) procollagen and tissue inhibitor of metallo-proteinase-1 mRNAs. Biochim Biophys Acta 1997;1361:177-184.
17. Friedman SL. Cytokines and fibrogenesis. Semin Liver Dis 1999;19:129-140.
18. Greenwel P, Schwartz M, Rosas M, Peyrol S, Grimaud JA, Rojkind M.Characterization of fat-storing cell lines derived from normal and CCl4-cirrhotic livers. Differences in the production of interleukin-6. Lab Invest1991;65:644-653.
19. Greenwel P, Rubin J, Schwartz M, Hertzberg EL, Rojkind M. Liver fat-storing cell clones obtained from a CCl4-cirrhotic rat are heterogeneouswith regard to proliferation, expression of extracellular matrix compo-nents, interleukin-6, and connexin 43. Lab Invest 1993;69:210-216.
20. Greenwel P, Iraburu MJ, Reyes-Romero M, Meraz-Cruz N, Casado E,Solıs-Herruzo JA, Rojkind M. Induction of an acute phase response inrats stimulates the expression of alpha 1(I) procollagen messenger ribo-nucleic acid in their livers. Possible role of interleukin-6. Lab Invest1995;72:83-91.
21. Levy MT, Trojanovska M, Reuben A. Oncostatin M: a cytokine up-regu-lated in human cirrhosis increases collagen production by human hepaticstellate cells. J Hepatol 2000;32:218-226.
22. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem1987;162:156-159.
23. Genovese C, Rowe D, Kream B. Construction of DNA sequences comple-mentary to rat a(1) and a(2) collagens mRNA and their use in studyingthe regulation of type I collagen synthesis by 1,25-dihydroxyvitamin D.Biochemistry 1984;23:6210-6216.
24. Oberbaumer I, Laurent M, Schwarz U, Sakurai Y, Yamada Y, Vogeli G,Voss T, et al. Amino acid sequence of the non-collagenous globulardomain (NC1) of the alpha 1 (IV) chain of basement membrane collagenas derived from complementary DNA. Eur J Biochem 1985;147:217.
25. Schwarzbauer JE, Tamkun JW, Lemischka IR, Hynes RO. Three differentfibronectin mRNAs arise by alternative splicing within the coding region.Cell 1983;35:421-431.
26. Henriet P, Rousseau GG, Eeckhout Y. Cloning and sequencing of mousecollagenase cDNA. Divergence of mouse and rat collagenases from theother mammalian collagenases. FEBS Lett 1992;310:175-178.
27. Reponen P, Sahlberg C, Huhtala P, Hurskainen T, Thesleff I, TryggvasonK. Molecular cloning of murine 72-kDa type IV collagenase and its ex-pression during mouse development. J Biol Chem 1992;267:7856-7862.
28. Crabtree GR, Kant JA. Molecular cloning of cDNA for the alpha, beta, andgamma chains of rat fibrinogen. A family of coordinately regulated genes.J Biol Chem 1981;256:9718.
29. Zern MA, Chakraborty PR, Ruiz-Opazo N, Yap SH, Shafritz DA. Devel-opment and use of a rat albumin cDNA clone to evaluate the effect ofchronic ethanol administration on hepatic protein synthesis. HEPATOL-OGY 1983 3:317-322.
30. Coulombe B, Skup D. In vitro synthesis of the active tissue inhibitor ofmetalloproteinases encoded by a complementary DNA from virus-in-fected murine fibroblasts. J Biol Chem 1988;263:1439-1443.
31. Nieto N, Friedman SL, Greenwel P, Cederbaum AI. CYP2E1-mediatedoxidative stress induces collagen type I expression in hepatic stellatecells. HEPATOLOGY 1999;30:987-996.
32. Mosmann T. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J Immunol Methods1983;65:55-63.
33. Aarden LA, De Groot ER, Schaap OL, Lansdorp PM. Production of hy-bridoma growth factor by human monocytes. Eur J Immunol 1987;17:1411-1416.
34. Helle M, Boeije L, Aarden LA. Functional discrimination between inter-leukin 6 and interleukin 1. Eur J Immunol 1988;18:1535-1540.
35. Delgado-Rizo V, Salazar A, Panduro A, Armendariz-Borunda J. Treat-ment with anti-transforming growth factor beta antibodies influences analtered pattern of cytokines gene expression in injured rat liver. BiochimBiophys Acta 1998;1442:20-27.
36. Baumann H, Latimer JJ, Glibetic MD. Mouse alpha 1-protease inhibitor isnot an acute phase reactant. Arch Biochem Biophys 1986;246:488-493.
606 NIETO ET AL. HEPATOLOGY March 2001

37. Hochepied T, Van Molle W, Berger FG, Baumann H, Libert C. Involve-ment of the acute phase protein alpha 1-acid glycoprotein in nonspecificresistance to a lethal gram-negative infection. J Biol Chem 2000;275:14903-14909.
38. Billiar TR, Curran RD, Williams DL, Kispert PH. Liver nonparenchymalcells are stimulated to provide interleukin 6 for induction of hepaticacute-phase response in endotoxemia but not in remote localized inflam-mation. Arch Surg 1992;127:36-37.
39. Panduro A, Shalaby F, Shafritz DA. Liver-specific gene expression invarious pathophysiologic states. HEPATOLOGY 1987;7(Suppl):10S-18S.
40. Panduro A, Shalaby F, Weiner FR, Biempica L, Zern MA, Shafritz DA.Transcriptional switch from albumin to alpha-fetoprotein and changes intranscription of other genes during carbon tetrachloride induced liverregeneration. Biochemistry 1986;25:1414-1420.
41. Armendariz-Borunda J, Katayama K, Seyer JM. Transcriptional mecha-nisms of type I collagen gene expression are differentially regulated byinterleukin-1 beta, tumor necrosis factor alpha, and transforming growthfactor beta in Ito cells. J Biol Chem 1992;267:14316-14321.
42. Milland J, Tsykin A, Thomas T, Aldred AR, Cole T, Schreiber G. Geneexpression in regenerating and acute-phase rat liver. Am J Physiol 1990;259:G340-G347.
43. Moshage H. Cytokines and the hepatic acute phase response. J Pathol1997;181:257-266.
44. van Gool J, van Vugt H, de Nie I. Acute phase reactants enhanceCCl4 induced liver cirrhosis in the rat. Exp Mol Pathol 1986;44:157-168.
45. Van Gool J, de Nie I, Smit J, Zuyderhoudt FM. Mechanisms by whichacute phase proteins enhance development of liver fibrosis: effects oncollagenase and prolyl-4-hydroxylase activity in the rat liver. Exp MolPathol 1986;45:160-170.
46. Mihara M, Ohsugi Y. Possible role of IL-6 in pathogenesis of immunecomplex-mediated glomerulonephritis in NZB/W F1 mice: induction ofIgG class anti-DNA autoantibody production. Int Arch Allergy Appl Im-munol 1990;93:89-92.
47. Van Gool J, Van Tiel D, Doenhoff MJ, Van Vugt H. Effect of acute phaseproteins, especially alpha 2-macroglobulin, on granuloma formation aroundSchistosoma mansoni eggs in the rat. Parasitology 1991;102:49-56.
48. Kamimura S, Tsukamoto H. Cytokine gene expression by Kupffer cells inexperimental alcoholic liver disease. HEPATOLOGY 1995;22:1304-1309.
49. Killar L, White J, Black R, Peschon J. Adamalysins. A family of metzincinsincluding TNF-alpha converting enzyme (TACE). Ann N Y Acad Sci1999;878:442-452.
50. Greenwel P, Geerts A, Ogata I, Geerts A. Liver fibrosis. In: Arias IM, BoyerJL, Fausto N, Jakoby WB, Schachter DA, Shafritz DA, eds. The Liver:Biology and Pathobiology. New York: Raven Press, 1994;1367-1381.
51. Santos RM, Norton P, Degli Esposti S, Zern MA. TGF-beta isoforms inalcoholic liver disease. J Gastroenterol 1998;33:383-389.
52. De Bleser PJ, Niki T, Rogiers V, Geerts A. Transforming growth factor-beta gene expression in normal and fibrotic rat liver. J Hepatol 1997;26:886-893.
53. Arthur MJ. Degradation of matrix proteins in liver fibrosis. Pathol ResPract 1994;190:825-833.
HEPATOLOGY Vol. 33, No. 3, 2001 NIETO ET AL. 607




















