
Fibromodulin Interactions with Type I and II Collagens
9
This article was downloaded by:[University of Toronto] [University of Toronto] On: 25 May 2007 Access Details: [subscription number 769429897] Publisher: Informa Healthcare Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Connective Tissue Research Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713617769 Fibromodulin Interactions with Type I and II Collagens To cite this Article: Viola 1 , Manuela, Bartolini 1 , Barbara, Sonaggere, Myriam, Giudici, Camilla, Tenni, Ruggero and Tira, M. Enrica , 'Fibromodulin Interactions with Type I and II Collagens', Connective Tissue Research, 48:3, 141 - 148 To link to this article: DOI: 10.1080/03008200701276133 URL: http://dx.doi.org/10.1080/03008200701276133 PLEASE SCROLL DOWN FOR ARTICLE Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material. © Taylor and Francis 2007
-
Upload
geo-social -
Category
Documents
-
view
3 -
download
0
Transcript of Fibromodulin Interactions with Type I and II Collagens

This article was downloaded by:[University of Toronto][University of Toronto]
On: 25 May 2007Access Details: [subscription number 769429897]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Connective Tissue ResearchPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713617769
Fibromodulin Interactions with Type I and II Collagens
To cite this Article: Viola 1, Manuela, Bartolini 1, Barbara, Sonaggere, Myriam,Giudici, Camilla, Tenni, Ruggero and Tira, M. Enrica , 'Fibromodulin Interactionswith Type I and II Collagens', Connective Tissue Research, 48:3, 141 - 148To link to this article: DOI: 10.1080/03008200701276133URL: http://dx.doi.org/10.1080/03008200701276133
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.
© Taylor and Francis 2007

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
Connective Tissue Research, 48:141–148, 2007Copyright c© Informa HealthcareISSN: 0300-8207 print / 1521-0456 onlineDOI: 10.1080/03008200701276133
Fibromodulin Interactions with Type I and II Collagens
Manuela Viola1
Dipartimento di Scienze Biomediche Sperimentali e Cliniche, Universita dell’Insubria, Varese, Italy
Barbara Bartolini,1 Myriam Sonaggere, Camilla Giudici,Ruggero Tenni, and M. Enrica TiraDipartimento di Biochimica “A. Castellani,” Universita di Pavia, Pavia, Italy
Fibromodulin is a keratan-sulfate small leucine-rich proteogly-can (SLRP) regulating collagen I and II fibril formation. In vivostudies suggest that, alongside decorin, fibromodulin plays an im-portant role in the maintenance of mature tissues. To characterizefibromodulin/decorin differences in binding to type I and II colla-gen, we tested the collagen CNBr peptides in solid-phase assays.Only one peptide from collagen II and several peptides from col-lagen I interacted with fibromodulin, pointing to multiple bindingsites in the collagen I molecule. By Scatchard-type analysis, thefibromodulin molecule showed only one class of binding sites forcollagen I and both low and high affinity (classes of) binding sitesfor collagen II. Lys/Hyl residues in both collagens are essential forthe interaction. Fibril formation assays showed the concomitantpresence of fibromodulin and decorin in fibrils and a cumulativeinhibitory effect. In solid-phase assays decorin seems to inhibitfibromodulin binding, whereas the contrary does not occur. Wefound fibromodulin and decorin have similarities and differencesthat may represent the biochemical basis of redundancy in SLRPfunction with compensation between different (classes of) SLRPs.
Keywords Collagen, Decorin, Fibromodulin
INTRODUCTIONFibromodulin (FM) is a keratan-sulfate proteoglycan (KS-
PG) belonging to the extracellular matrix family of smallleucine-rich proteoglycans (SLRPs) that also includes lumican(a KS-PG), decorin and biglycan, both chondroitin/dermatansulfate (CS/DS) PGs. For most of these proteoglycans the firstfunction described was their ability to interact with collagensI and II and to control the fibril formation process of fibrillarcollagens (for FM, decorin, and lumican see [1], and literaturecited therein; for biglycan see [2]).
Received 27 November 2006; revised 17 January 2007; accepted10 February 2007.
1These authors contributed equally to the work.Address correspondence to Manuela Viola, PhD, Dipartimento di
Scienze Biomediche Sperimentali e Cliniche, Universita dell’Insubria,via J. H. Dunant 5, 21100 Varese, Italy. E-mail: [email protected]
The morphology and function of the extracellular matrix(ECM) of connective tissues are highly regulated and the pro-teoglycans populating the ECM influence both aspects. Decorin,biglycan, lumican, and fibromodulin are widely present in con-nective tissues, with a different pattern of distribution withineach tissue [3].
The physiological importance of the interactions betweencollagen and SLRPs was demonstrated in mice knocked outfor FM and other SLRPs [4]. Null mice do not show any grossanatomical defects, grow to normal size, and have a normal lifespan. Collagen fibril formation and morphology are affected bythe absence of SLRPs, different tissues of the same animalsshowing different alterations [5, 6]. All these effects depend onthe developmental stage of the tissues, because different behav-iors have been reported for early differentiation stage or ma-ture tissues [7]. The absence of SLRP leads to an increasedexpression or content of another SLRP of the same subclass(e.g., decorin [DCN] and biglycan or fibromodulin and lumi-can) [5, 7]. It is noteworthy that the most severe phenotypesfor collagen organization and mice fitness arise in animals dou-bly deficient for FM-lumican (both KS-PGs) or biglycan-DCN(both CS/DS-PGs) with respect to single PG knock-out mice.Several authors concluded that SLRP function is redundant dur-ing development, and there is a sort of compensation betweendifferent SLRPs or classes of SLRPs (for FM, see [4, 5, 7, 8]).
Early morphological studies on the interactions of FM andDCN with collagen molecules estimated that KS-PGs (such asFM) associate with the a and c bands of the D-period pattern,whereas CS/DS-PGs (like DCN) bind specifically to the d ande bands [9]. Immunolocalization studies showed FM at the sur-face of collagen fibrils in tendon and cartilage, i.e., associatedwith collagen type I and II fibrils, respectively, with a prefer-ence for the gap regions of both collagens [10]. So far, no dataare available either on the correct localization of the FM bind-ing region on collagen molecules or on the relation with otherCS/DS-PGs.
FM structure has been extensively studied in terms of gly-cosaminoglycan (GAG) chain synthesis, protein folding, andbiochemical characteristics [11–14]. For example, glycosylation
141

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
142 M. VIOLA ET AL.
appears to be tissue-specific, developmentally regulated, andage-dependent [15, 16].
About 70% of the total amino acids of FM comprise theleucine-rich repeats, sequences of 20–26 residues with as-paragine and leucine residues in conserved positions, that givethe SLRPs their peculiar shape [17] necessary for correct foldingand collagen interaction. FM binds collagens I and II and slowsthe collagen fibril formation rate in vitro [18], and the binding totype I collagen is mediated by multiple domains [1]. Notwith-standing the similar distribution, FM and DCN do not competefor binding to collagen I and II during fibril forming experiments[1, 19], demonstrating distinct binding sites on these collagens.
Our research aimed at determining similarities and differ-ences between FM and DCN for their binding to collagens I andII and their CNBr peptides to define their own binding speci-ficity; also we wanted to emphasize the effective cooperationand competition between FM and DCN through fibrillogenesisand solid-phase assays.
MATERIALS AND METHODSAcid-soluble type I collagen from bovine skin, pepsinized
type II collagen from bovine cartilage of nasal septum, andtheir CNBr peptides were available and characterized in ourlaboratory [20, 21]. p-nitrophenyl phosphate, avidin conjugatedwith alkaline phosphatase, and sulfosuccinimidobiotin camefrom Pierce; chondroitinase ABC (EC 4.2.2.4) and endo-β-galactosidase (EC 3.2.1.103) were from Seikagaku; DEAE-Sephacel from Pharmacia, microtiter plates from Nunc and Vi-vaspin concentrators came from Vivascience Ltd; avidin conju-gated with horseradish peroxidase was from Sigma. All otherchemicals were of analytical grade.
Purification and Characterization of Fibromodulinand Decorin
FM was purified from bovine tendon with minor modificationof the method described by Font et al. [22]. Proteoglycans wereextracted from bovine tendon with 4 M sodium chloride, pH 5.6,in a 50 mM sodium acetate buffer containing protease inhibitors(5 mM benzamidine, 0.1 M ε-aminocaproic acid, 10 mM EDTA,1 mM PMSF). After centrifugation, the supernatant was loadedon a DEAE Sephacel column (1.5 × 25 cm) equilibrated with0.3 M NaCl, 10 mM Tris-HCl, 10 mM EDTA, 0.05 mM PMSF,pH 7.0; elution was performed with a linear gradient (0.3–0.8 MNaCl) in the same buffer. Fractions collected were extensivelydialyzed against the same buffer without NaCl and lyophilized.Decorin was purified as described elsewhere [20]. Both purifiedPGs were desalted with Vivaspin concentrators. Preparations ofboth PGs were analyzed by the Bradford method for their proteincontent [23] and by SDS-PAGE followed by gel staining withcoomassie brilliant blue R250.
The purified PGs were then labelled with sulfosuccinimidobi-otin [20]. Briefly, the samples (2 mg/ml) in PBS were incubatedwith a 20-fold molar excess of sulfosuccinimidobiotin for 2 hrat room temperature. Concentrated Tris-HCl buffer, pH 7.5, was
then added to 50 mM final concentration and the samples wereincubated for 1 hr, extensively dialyzed against PBS and storedat −20◦C.
To characterize the glycosidic moiety, FM was treated withtwo enzymes: endo-β-galactosidase specific for KS and chon-droitinase ABC (ABCase) active selectively toward CS/DS. Forendo-β-galactosidase hydrolysis, 5 µg of PGs were incubatedfor 18 hr with 4 mU of enzyme at 50◦C in 0.1 M sodium acetate,pH 5.8 [24]. ABCase digestion was performed using a ratio of10 µg of protein per 1.5 mU of enzyme, in a 20 mM Tris-HCl,150 mM NaCl buffer, pH 8.0, for 16 hr at 37◦C [25]. After bothtreatments, aliquots of FM were analyzed by SDS-PAGE.
Collagen Type I and II Fibril FormationFreeze-dried type I and II collagens were redissolved in 0.1 M
acetic acid at a final concentration of 2 mg/ml on the basis of thehydroxyproline content [26]. Aliquots of 0.15 mg of collagenwere lyophilized and redissolved in 0.15 ml of 0.05 M aceticacid. Then 30 µl of this solution (30 µg of collagen, 0.11 nmol)were added to 250 µl of PBS 2× and 220 µl of an aqueous solu-tion with 12µg (0.25 nmol) of PGs at a temperature of 4◦C. Fibrilformation was monitored by following the turbidity change inabsorbance at 313 nm in a Beckman DU-62 spectrophotometerthermostatted at 32◦C. Absorbance data were recorded everyminute for 120 min. At the end of the incubation, the sampleswere centrifuged in a vial coated with 1% BSA, and the PGbound to the fibrils was revealed in the pellet after SDS-PAGEand Western blotting using avidin conjugated with horseradishperoxidase.
Chemical Modifications of Collagens and CNBr PeptidesChemical derivatizations were performed in mild conditions
with two different methods, both involving the primary ε-aminogroup of lysine and hydroxylysine side chains. N-methylation
FIG. 1. Analysis of fibromodulin. SDS-PAGE (10% polyacrylamide) of fibro-modulin and effects of enzymatic digestions: (A) FM before and after digestionwith ABCase; (B) FM before and after digestion with endo-β-galactosidase.

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
FIBROMODULIN INTERACTIONS WITH COLLAGEN 143
was performed with formaldehyde in the presence of NaBH3CNas reducing agent, whereas N-acetylation was performed withsulfosuccinimidyl acetate. Derivatives were characterized aspreviously reported; in particular, CNBr peptides used in thiswork are able to form trimers having triple helical conforma-tion, and the above chemical modification did not significantlyalter the thermal stability of trimeric collagen I and II and oftrimers of CNBr peptides [20, 21].
Binding AssayCollagenous samples were dissolved in 0.1 M acetic acid at
1–1.5 mg/ml and maintained at 4◦C for ≥7 days, with occa-sional vortexing. The actual concentration was determined bya hydroxyproline assay after acidic hydrolysis [26]. After clar-ification by centrifugation, working solutions were prepared bydilution with PBS, at 25 µg/ml for collagens or equimolecularamounts of CNBr peptides.
First, 96-well microtiter plates were coated overnight at4◦C with solutions containing 5 µg (18 pmol) of collagens
FIG. 2. Fibril forming assays. (A) Acid soluble type I collagen. Incubation at 32◦C of collagen at 60 µg/ml (0.22 µM) and biotinylated fibromodulin (bFM)and/or decorin (bDCN), in a total volume of 0.5 ml, was followed by turbidimetry monitored at 313 nm. The incubation mixture contained: (◦) = collagen only,no PG; (�) = 24 µg/ml of bFM (0.5 µM); (�) = 24 µg/ml of bDCN (0.5 µM); (×) = 24 µg/ml of each biotinylated PG. Each kinetic profile is representative ofat least three assays. (B) Western blots of pellets recovered by centrifugation at the end of each fibrillogenesis of type I collagen. Lane 1 = 5 µg of bDCN usedas control; lane 2 = 5 µg of bFM used as control; lane 3 = collagen and both bDCN and bFM; lane 4 = collagen and bFM; lane 5 = collagen and bDCN. (C)Pepsinized type II collagen. The assays conditions were the same as described in (A), the only difference being the use of collagen II instead of collagen I. Symbolsas in (A). (D) Western blots of pellets recovered by centrifugation at the end of the fibrillogenesis of type II collagen. Lane 1 = 5 µg of bDCN used as control; lane2 = 5 µg of bFM used as control; lane 3 = collagen and both bDCN and bFM; lane 4 = collagen and bFM; lane 5 = collagen and bDCN. White bands visible at∼120 kDa overlapping bDCN signal are due to collagen α-chains (Figures 2B and 2D, lanes 3 and 5). In western blots biotinylated PGs bound to the fibrils weredetected by means of avidin conjugated with horseradish peroxidase.
or equimolecular amount of their CNBr peptides in PBS(200 µl/well); control wells were coated with 200 µl containing5 µg of BSA in PBS. All analyses were done at least in tripli-cate. Binding efficiency of collagens and CNBr peptides to themultiwell plates was found to be 9.4% ± 3.0 (mean ± standarddeviation) and 15.1% ± 3.0, respectively [20].
After rinsing with 0.15 M NaCl, 0.05% Tween-20, the wellswere incubated with 200 µl of 1% BSA in PBS for 1 hr at roomtemperature. After rinsing as above, the coated wells were incu-bated for 2 hr at room temperature with 24 pmol of biotinylatedFM (bFM) (when using type II collagen and its peptides) or48 pmol of bFM (when using type I collagen and its peptides) in200 µl of PBS, 0.05% Tween-20. The different amounts of FMused were related to the different saturation levels for the dif-ferent collagens. For Scatchard-type analysis, the same amountof collagens or peptides were used for coating and incubatedwith increasing concentrations of biotinylated FM; affinity con-stants were then obtained according to Hedbom and Heinegard[18].

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
144 M. VIOLA ET AL.
FIG. 3. Binding of biotinylated fibromodulin to collagenous samples.Polystyrene wells were coated with type I or its CNBr peptides (A) and typeII collagen and its CNBr peptides (B). First 5 µg (18 pmol) of collagens orequimolecar amounts of their CNBr peptides were dissolved in 200 µl of PBSand used for each well. Next, 48 pmol of biotinylated fibromodulin were addedto collagen type I samples (A) whereas 24 pmol were added to collagen type IIsamples (B). Bound fibromodulin was determined by the avidin-alkaline phos-phatase system. The absorbance is the mean of analyses performed in triplicate.CNBr peptides: their names and position along the triple helical domain arereported below each histogram.
Bound FM was detected using avidin conjugated with al-kaline phosphatase diluted 1:1000 in 1% BSA in PBS, 0.05%Tween-20 (200 µl/well), followed by a rinse and by 200 µlof the substrate solution (p-nitrophenyl phosphate at 1 mg/ml in0.9 M diethanolamine-HCl buffer, 0.5 mM MgCl2, 3 mM NaN3,pH 9.5). The absorbance was measured at 405 nm before andafter color development. Dissociation constants (Kd) were deter-mined with GraphPad Prism4 software (GraphPad, San Diego,CA, USA) [27].
Another series of solid-phase assays was done to test com-petition between FM and DCN for their binding to collagens.Wells were coated with 5 µg (18 pmol) of collagens I orII and incubated with various amount of FM or DCN withbiotinylated-DCN (bDCN) or bFM, respectively. The concen-trations were chosen on the basis of their saturation curves, atsaturable concentration or below this point. The amount of eachPG in competition experiments is specified in the appropriatefigures.
RESULTS
Analysis of FibromodulinSDS-PAGE of FM showed a dispersed band centered at
66 kDa (Figure 1). After digestion with chondroitinase ABC(ABCase) (Figure 1A), there was no change in the migration pat-tern of the PG, whereas endo-β-galactosidase treatment causeda shift to 47 kDa (Figure 1B). The difference between the molec-ular mass found by SDS-PAGE and that deduced by sequenceanalysis is the result of carbohydrate substitution [18].
Fibril Forming AssaysPurified FM and DCN were tested in collagen type I and
II fibril forming assays (Figure 2). The presence of 24 µg/ml(0.5 µM) biotinylated FM with 60 µg/ml (0.22 µM) collageninhibited the formation of fibrils, as shown by the smaller in-crease in absorbance (50 and 75% inhibition for type I and IIcollagens, respectively). Comparable results were obtained withDCN (where the inhibition was 50 and 10% for type I or II col-lagen, respectively) (Figures 2A and 2C). After centrifugation,FM and DCN were detected by Western blotting in the pellet inassociation with collagen fibrils (Figures 2B and 2D). A largereffect on collagen fibril formation was observed when both FMand DCN were present in the assay: the inhibition was 90% forcollagen I and 75% for collagen II and apparently cumulative,suggesting that the two PGs act independently. Furthermore,both FM and DCN were found in the pellet of collagen fibrils(Figures 2B and 2D, lanes 3).
Binding AssayThe interaction of FM with type I and II collagen and their
CNBr peptides was tested by solid-phase assays using biotiny-lated FM and equimolar amounts of collagenous samples. FMbound collagen type I/II and the peptides α1(I) CB6, CB7, CB8,α2(I) CB4, and α1(II) CB11 (Figure 3). The binding required thenative triple helical conformation of collagens/peptides, sincethermal denaturation of the collagenous samples dramaticallydecreased the binding to FM (data not shown). The binding wasionic, as increasing the ionic strength (up to 250 mM NaCl) dur-ing the incubation with FM decreased the interaction, whereasthe presence of the nonionic detergent Triton X-100 (1% v/v)did not affect the binding (data not shown).
The affinities between FM and collagens/peptides were de-termined in solid-phase assays using increasing amounts of thebiotinylated proteoglycan and equimolar amounts of collage-nous samples (Figure 4 for collagen I and II). The Scatchard-type plots, drawn according to Hedbom and Heinegard [18],indicated a unimodal behavior of FM for binding to collagenI (inset of Figure 4A), and a bimodal behavior for both colla-gen II (inset in Figure 4B) and its peptide α1(II) CB11 (datanot shown). These data suggest that FM possesses high and lowaffinity classes of binding sites for collagen II and its peptide

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
FIBROMODULIN INTERACTIONS WITH COLLAGEN 145
FIG. 4. Scatchard-type analysis of fibromodulin binding to type I and II colla-gens. Increasing amounts of biotinylated fibromodulin were added to polystyrenewells coated with 5 µg (18 pmol) of collagens or equimolecular amounts of theirCNBr peptides. Binding was measured using the avidin-alkaline phosphatasesystem. (A) Saturation curve of type I collagen and related Scatchard plot in theinset. All CNBr peptides from type I collagen able to bind FM (Figure 3) showa similar behavior (data not shown). (B) Saturation curve of type II collagenand related Scatchard plot in the inset. CB11 from type II collagen shows asimilar behavior (data not shown). All analyses were done at least in triplicate.We could not use peptide α1(II) CB9,7 because its purification was not possible.Dissociation constants (Kd) were determined with GraphPad Prism4 software(GraphPad, San Diego, CA, USA).
and only one class of binding sites for collagen I. All the cal-culated dissociation constants are reported in Table 1 and are inthe nanomolar range, as for DCN [20].
As further characterization, FM binding was tested withN-methylated and N-acetylated collagens and peptides. Bothchemical derivatizations modify the amino group of Lys/Hylside chains (see Methods section), the positive charge at physi-ological pH of Lys/Hyl being preserved only by N-methylation.No denaturation and no significant alteration of the thermal sta-bility of trimeric species of collagen I and II and of trimersof CNBr peptides occurred on derivatization [20, 21]. Fibro-modulin binding to N-methylated type I and II collagens and topeptides α1(I) CB8 and α1(II) CB11 was comparable to the un-derivatized counterparts, whereas some decrease was found forthe other peptides (Figure 5). Conversely, FM completely lostits binding ability with N-acetylated samples (Figure 5). Inter-estingly, peptides α1(I) CB8 and α1(II) CB11 were in a homol-ogous region in the N-terminal half of their respective collagen
TABLE 1Affinity constants (Kd) calculated from Scatchard-type plots
for each collagenous sample
Samples FM Kd (nM) DCN Kd (nM)a
Collagen type I 87 41b
α1(I) CB8 18 44α1(I) CB7 61 13α1(I) CB6 177 (c)α2(I) CB4 22 16Collagen type II 8 and 583 42b
α1(II) CB11 3 and 582 22
aThe data reported are from Tenni et al. [20].bThe value reported was obtained from the linear interpolation of alldata points.cIt was impossible to calculate the dissociation constant for this peptidebecause a saturation level was not clearly identifiable.
molecules (see CNBr peptides location, bottom of Figures 3Aand 3B). Finally, as control of the chemical derivatization ef-fect, we tested α1(I) CB3 and α1(II) CB8: these peptides didnot interact with FM before (Figure 3) and after N-methylationor N-acetylation (data not shown).
Competitions in solid-phase assays were performed to es-tablish whether DCN and FM simultaneously bind collagens atdifferent sites, as indicated by the fibrillogenesis assays. To per-form these experiments we chose PG concentrations above orbelow saturation concentration [28]. In the case of FM-collagentype II, a saturation and two lower concentrations were used, onthe basis of its bimodal behavior.
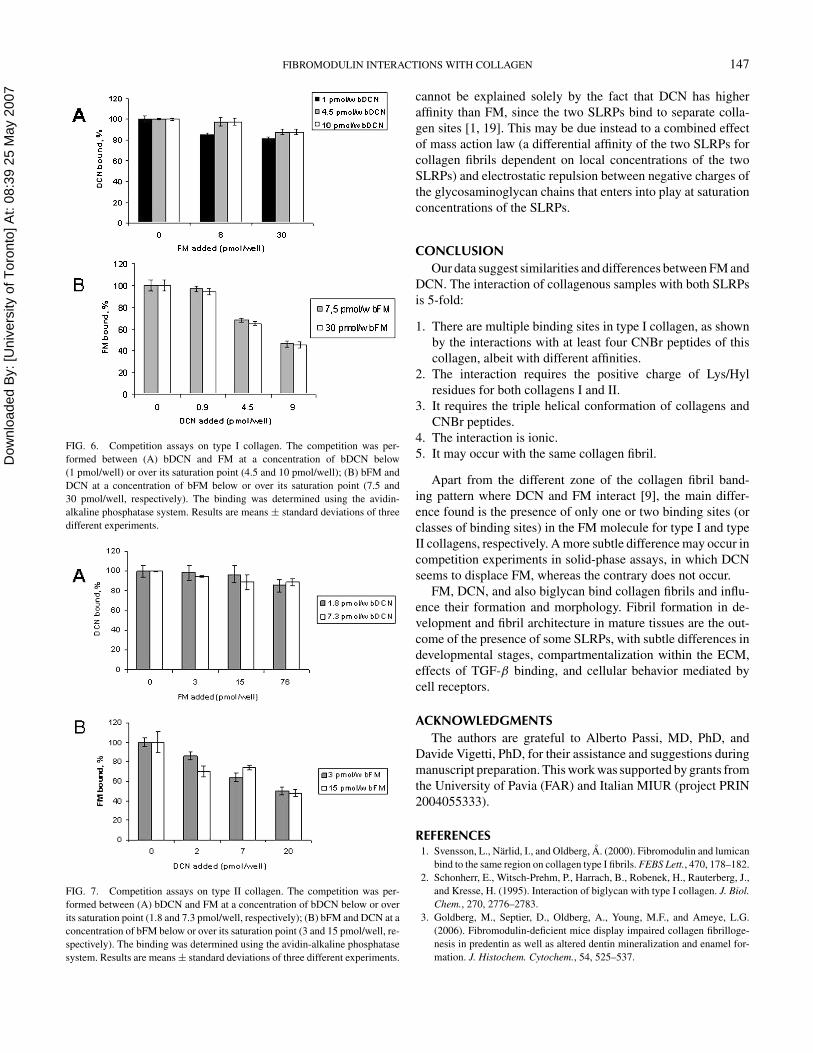
Concerning type I collagen (Figure 6A), the presence of FMhad no major effect on bDCN binding at any of the concen-trations used (maximum decrease of 11% of the control). Onthe contrary, in the bFM/DCN competition (Figure 6B), bindingdropped by up to 50% with 9 pmol/well of DCN or less withlower amounts. Similarly, for the competition on type II colla-gen FM did not affect the displacement of bDCN (Figure 7A),whereas DCN decreased the binding of bFM in percentagesranging from 50% to 10% of the control (Figure 7B).
DISCUSSIONThe SLRP family comprises different PGs, among which both
fibromodulin and decorin (a KS and a CS/DS PG, respectively)are reported to bind collagen type I and II and slow the fibrilformation rate in vitro [18, 29]. Our work addressed the sites incollagen types I and II involved in the interaction with FM byanalyzing the ability of FM to bind CNBr peptides from type Iand II collagens. CNBr peptides were in the native triple helicalconformation that is maintained after chemical modifications (asdemonstrated in the past [20, 21]). We also aimed to elucidatethe mutual role of DCN and FM in cooperation/competitionfor these collagen types, since literature data reported that FM

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
146 M. VIOLA ET AL.
FIG. 5. Effect of chemical modifications on collagen-FM interaction. Polystyrene wells were coated with 5 µg (18 pmol) of collagen type I (A) or II (B) orequimolecular amounts of their CNBr peptides, unmodified or after chemical modifications. Next, 48 pmol of biotinylated FM were added to type I collagen andits peptides, whereas 24 pmol of FM were added to type II collagen and its peptides. The bound fibromodulin was determined by means of the avidin-alkalinephosphatase system. The absorbance is the mean of analyses performed in triplicate.
and DCN do not compete in fibril forming experiments withcollagens I and II [1, 19].
With our protocols and samples, the FM interaction with typeI collagen and its CNBr peptides was unimodal; the FM in-teraction was bimodal with collagen II and its peptide CB11with affinity constants differing by about two orders of magni-tude. The full-length molecule of type I collagen has less affinity(87 nM, Table 1) than that reported in the literature, where theinteraction of recombinant FM with type I collagen shows Kd’sof 0.2 and 1.6 nM [1], and FM from bovine tendon has one bind-ing site with a Kd of 9.9 nM [19] or 35 nM [18]. A hint of twobinding sites for type II collagen was reported for the bovineFM molecule [19].
Using CNBr peptides, both collagen types tested had a bind-ing region for FM located at the N-terminus of the molecule, anda region in the C-terminal half for collagen type I. Comparisonof the affinities of FM for CNBr peptides with those obtained us-ing DCN (Table 1) showed differential affinities of some CNBrpeptides for the two SLRPs.
The fact that at least four CNBr peptides interacted with bothFM and DCN does not appear to be an artefact since all peptidescontain regions falling in the d and e bands of the D-period ofcollagen fibrils (where DCN binds) and also regions falling in thea and c bands (where FM binds) [9]. Therefore, we deduced thattype I collagen may contain multiple binding sites for both FM(this work) and DCN [20]. A recent publication on the microfib-rillar structure of type I collagen in rat tail tendon reported two
shallow holes at 0.6D and 0.8D in the gap zone of each D-periodof the fibrils, postulated to provide a protein-to-protein contactarea and potential association sites for DCN [30]. This implies aD-periodicity of SLRP binding sites and probably also that eachSLRP molecule is in contact with more than one collagen triplehelix.
The involvement of collagen Lys/Hyl residues in FM bindingis a further novel aspect of our findings. In fact, the consequencesof N-acetylation of the primary amine of collagenous sampleson the interaction with FM clearly indicate that the side chainof some Lys/Hyl residues in collagens belong to, or are near,the binding site(s) for FM. The positive charge of Lys/Hyl istherefore essential for the interaction. The same was found forDCN [20].
The presence of low and high affinity classes of binding sitesfor collagen II found in the FM molecule (by Scatchard-typeanalysis) and the multiple domains in collagen I for FM mayconnect the functional redundancy in SLRPs with their specificcharacteristics, and with FM’s ability to bind collagens also in thepresence of DCN. Our experiments involving the co-presenceof FM and DCN showed two features. Both PGs were asso-ciated with type I and type II collagen fibrils formed in vitroin fibril-forming assays and had a cumulative inhibitory effecton collagen I and II fibril formation (Figure 2). In solid-phaseassays where FM and DCN compete for binding to a limitedamount of collagens I or II, DCN seems to inhibit FM bind-ing, whereas the contrary does not occur (Figures 6 and 7). This
Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
FIBROMODULIN INTERACTIONS WITH COLLAGEN 147
FIG. 6. Competition assays on type I collagen. The competition was per-formed between (A) bDCN and FM at a concentration of bDCN below(1 pmol/well) or over its saturation point (4.5 and 10 pmol/well); (B) bFM andDCN at a concentration of bFM below or over its saturation point (7.5 and30 pmol/well, respectively). The binding was determined using the avidin-alkaline phosphatase system. Results are means ± standard deviations of threedifferent experiments.
FIG. 7. Competition assays on type II collagen. The competition was per-formed between (A) bDCN and FM at a concentration of bDCN below or overits saturation point (1.8 and 7.3 pmol/well, respectively); (B) bFM and DCN at aconcentration of bFM below or over its saturation point (3 and 15 pmol/well, re-spectively). The binding was determined using the avidin-alkaline phosphatasesystem. Results are means ± standard deviations of three different experiments.
cannot be explained solely by the fact that DCN has higheraffinity than FM, since the two SLRPs bind to separate colla-gen sites [1, 19]. This may be due instead to a combined effectof mass action law (a differential affinity of the two SLRPs forcollagen fibrils dependent on local concentrations of the twoSLRPs) and electrostatic repulsion between negative charges ofthe glycosaminoglycan chains that enters into play at saturationconcentrations of the SLRPs.
CONCLUSIONOur data suggest similarities and differences between FM and
DCN. The interaction of collagenous samples with both SLRPsis 5-fold:
1. There are multiple binding sites in type I collagen, as shownby the interactions with at least four CNBr peptides of thiscollagen, albeit with different affinities.
2. The interaction requires the positive charge of Lys/Hylresidues for both collagens I and II.
3. It requires the triple helical conformation of collagens andCNBr peptides.
4. The interaction is ionic.5. It may occur with the same collagen fibril.
Apart from the different zone of the collagen fibril band-ing pattern where DCN and FM interact [9], the main differ-ence found is the presence of only one or two binding sites (orclasses of binding sites) in the FM molecule for type I and typeII collagens, respectively. A more subtle difference may occur incompetition experiments in solid-phase assays, in which DCNseems to displace FM, whereas the contrary does not occur.
FM, DCN, and also biglycan bind collagen fibrils and influ-ence their formation and morphology. Fibril formation in de-velopment and fibril architecture in mature tissues are the out-come of the presence of some SLRPs, with subtle differences indevelopmental stages, compartmentalization within the ECM,effects of TGF-β binding, and cellular behavior mediated bycell receptors.
ACKNOWLEDGMENTSThe authors are grateful to Alberto Passi, MD, PhD, and
Davide Vigetti, PhD, for their assistance and suggestions duringmanuscript preparation. This work was supported by grants fromthe University of Pavia (FAR) and Italian MIUR (project PRIN2004055333).
REFERENCES1. Svensson, L., Narlid, I., and Oldberg, A. (2000). Fibromodulin and lumican
bind to the same region on collagen type I fibrils. FEBS Lett., 470, 178–182.2. Schonherr, E., Witsch-Prehm, P., Harrach, B., Robenek, H., Rauterberg, J.,
and Kresse, H. (1995). Interaction of biglycan with type I collagen. J. Biol.Chem., 270, 2776–2783.
3. Goldberg, M., Septier, D., Oldberg, A., Young, M.F., and Ameye, L.G.(2006). Fibromodulin-deficient mice display impaired collagen fibrilloge-nesis in predentin as well as altered dentin mineralization and enamel for-mation. J. Histochem. Cytochem., 54, 525–537.

Dow
nloa
ded
By: [
Uni
vers
ity o
f Tor
onto
] At:
08:3
9 25
May
200
7
148 M. VIOLA ET AL.
4. Ameye, L., and Young, M.F. (2002). Mice deficient in small leucine-richproteoglycans: novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome. Glycobiology, 12, 107R–116R.
5. Svensson, L., Aszodi, A., Reinholt, F.P., Fassler, R., Heinegard, D., andOldberg, A. (1999). Fibromodulin-null mice have abnormal collagen fibrils,tissue organization, and altered lumican deposition in tendon. J. Biol. Chem.,274, 9636–9647.
6. Danielson, K.G., Baribault, H., Holmes, D.F., Graham, H., Kadler, K.E.,and Iozzo, R.V. (1997). Targeted disruption of decorin leads to abnormalcollagen fibril morphology and skin fragility. J. Cell Biol., 136, 729–743.
7. Zhang, G., Ezura, Y., Chervoneva, I., Robinson, P.S., Beason, D.P., Carine,E.T., Soslowsky, L.J., Iozzo, R.V., and Birk, D.E. (2006). Decorin regulatesassembly of collagen fibrils and acquisition of biomechanical propertiesduring tendon development. J. Cell Biochem., 98, 1436–1439.
8. Young, M.F., Bi, Y., Ameye, L., and Chen, X.D. (2002). Biglycan knockoutmice: new models for musculoskeletal diseases. Glycoconjugate J., 19, 257–262.
9. Scott, J.E., and Haigh, M. (1988). Identification of specific binding sitesfor keratan sulphate proteoglycans and chondroitin-dermatan sulphate pro-teoglycans on collagen fibrils in cornea by the use of cupromeronic bluein ‘critical-electrolyte-concentration’ techniques. Biochem. J., 253, 607–610.
10. Hedlund, H., Mengarelli-Widholm, S., Heinegard, D., Reinholt, F.P., andSvensson, O. (1994). Fibromodulin distribution and association with colla-gen. Matrix Biol., 14, 227–232.
11. Plaas, A.H.K., and Wong-Palm, S. (1993). Biosynthetic mechanisms for theaddition of polylactosamine to chondrocyte fibromodulin. J. Biol. Chem.,268, 26634–26644.
12. Plaas, A.H.K., Neame, P.J., Nivens, C.M., and Reiss, L. (1990). Identifica-tion of the keratan sulfate attachment sites on bovine fibromodulin. J. Biol.Chem., 265, 20634–20640.
13. Heathfield, T.F., Onnerfjord, P., Dahlberg, L., and Heinegard, D. (2004).Cleavage of fibromodulin in cartilage explants involves removal of the N-terminal tyrosine sulfate-rich region by proteolysis at a site that is sensitiveto matrix metalloproteinase-13. J. Biol. Chem., 279, 6286–6295.
14. Antonsson, P., Heinegard, D., and Oldberg, A. (1991). Posttranslationalmodifications of fibromodulin. J. Biol. Chem., 266, 16859–16861.
15. Cornuet, P.K., Blochberger, T.C., and Hassel, J.R. (1994). Molecular poly-morphism of lumican during corneal development. Invest. Ophthalmol. Vis.Sci., 35, 870–877.
16. Roughley, P.J., White, R.J., Cs-Szabo, G., and Mort, J.S. (1996). Changeswith age in the structure of fibromodulin in human cartilage. Osteoarth.Cartil., 4, 153–161.
17. Scott, P.G., McEwan, P.A., Dodd, C.M., Bergmann, E.M., Bishop, P.N., andBella, J. (2004). Crystal structure of the dimeric protein core of decorin, the
archetypal small leucine-rich repeat proteoglycan. Proc. Natl. Acad. Sci.USA, 101, 15633–15638.
18. Hedbom, E., and Heinegard, D. (1989). Interaction of a 59-kDa connectivetissue matrix protein with collagen I and collagen II. J. Biol. Chem., 264,6898–6905.
19. Hedbom, E., and Heinegard, D. (1993). Binding of fibromodulin anddecorin to separate sites on fibrillar collagens. J. Biol. Chem., 268, 27307–27312.
20. Tenni, R., Viola, M., Welser, F., Sini, P., Giudici, C., Rossi, A., and Tira,M.E. (2002). Interaction of decorin with CNBr peptides from collagens Iand II. Evidence for multiple binding sites and essential lysyl residues incollagen. Eur. J. Biochem., 269, 1428–1437.
21. Giudici, C., Viola, M., Tira, M.E., Forlino, A., and Tenni, R. (2003). Molec-ular stability of chemically modified collagen triple helices. FEBS Lett., 547,170–176.
22. Font, B., Eichenberger, D., Goldschmidt, D., Boutillon, M.M., and Hulmes,D.J.S. (1998). Structural requirements for fibromodulin binding to colla-gen and the control of type I collagen fibrillogenesis. Critical roles fordisulphide bonding and the C-terminal region. Eur. J. Biochem., 254, 580–587.
23. Bradford, M.M. (1976). A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of protein-dyebinding. Anal. Biochem., 72, 248–254.
24. Scott, P.G., Nakano, T., and Dodd, C.M. (1997). Isolation and character-ization of small proteoglycans from different zones of the porcine kneemeniscus. Biochim. Biophys. Acta, 1336, 254–262.
25. Yamagata, T., Saito, H., Habuchi, O., and Suzuki, S. (1968). Purificationand properties of bacterial chondroitinases and chondrosulfatases. J. Biol.Chem., 243, 1523–1535.
26. Huszar, G., Maiocco, J., and Naftolin, F. (1980). Monitoring of collagen andcollagen fragments in chromatography of protein mixtures. Anal. Biochem.,105, 424–429.
27. Schonherr, E., Sunderkotter, C., Iozzo, R.V., and Schaefer, L. (2005).Decorin, a novel player in the insulin-like growth factor system. J. Biol.Chem., 280, 15767–15772.
28. Thomson, C.A., Tenni, R., and Ananthanarayanan, V.S. (2003). MappingHsp47 binding site(s) using CNBr peptides derived from type I and type IIcollagen. Protein Sci., 12, 1792–1800.
29. Oldberg, A., Antonsson, P., Lindblom, K., and Heinegard, D. (1989). Acollagen-binding 59-kDa protein (fibromodulin) is structurally related tothe small interstitial proteoglycans PG-S1 and PG-S2 (decorin). EMBO J.,8, 2601–2604.
30. Orgel, J.P., Irving, T.C., Miller, A., and Wess, T.J. (2006). Microfibrillarstructure of type I collagen in situ. Proc. Natl. Acad. Sci. USA, 103, 9001–9005.




















