
Kinetic studies of small molecule interactions with protein kinases using biosensor technology
Upload
independentCategory
view
1download
0
Molecular Microbiology (2000) 35(6), 1483±1492
The type III protein translocation system ofenteropathogenic Escherichia coli involvesEspA±EspB protein interactions
Elizabeth L. Hartland,1 Sarah J. Daniell,1
Robin M. Delahay,1 Bianca C. Neves,1 Tim Wallis,2
Robert K. Shaw,3 Christine Hale,1 Stuart Knutton3
and Gad Frankel1*1Department of Biochemistry, Imperial College of Science,
Technology and Medicine, London SW7 2AZ, UK.2Institute of Animal Health, Compton, Berkshire, UK.3Institute of Child Health, University of Birmingham,
Birmingham, UK.
Summary
Enteropathogenic Escherichia coli (EPEC), like many
bacterial pathogens, use a type III secretion system to
deliver effector proteins across the bacterial cell wall.
In EPEC, four proteins, EspA, EspB, EspD and Tir are
known to be exported by a type III secretion system
and to be essential for `attaching and effacing' (A/E)
lesion formation, the hallmark of EPEC pathogenicity.
EspA was recently shown to be a structural protein and
a major component of a large, transiently expressed,
filamentous surface organelle which forms a direct link
between the bacterium and the host cell. In contrast,
EspB is translocated into the host cell where it is
localized to both membrane and cytosolic cell frac-
tions. EspA and EspB are required for translocation of
Tir to the host cell membrane suggesting that they may
both be components of the translocation apparatus. In
this study, we show that EspB co-immunoprecipitates
with the EspA filaments and that, during EPEC infec-
tion of HEp-2 cells, EspB localizes closely with EspA.
Using a number of binding assays, we also show that
EspB can bind and be copurified with EspA. Never-
theless, binding of EspA filaments to the host cell
membranes occurred even in the absence of EspB.
These results suggest that following initial attachment
of the EspA filaments to the target cells, EspB is
delivered into the host cell membrane and that the
interaction between EspA and EspB may be important
for protein translocation.
Introduction
Most bacterial virulence determinants are surface located
or secreted outside the bacterium. Many pathogens of
animals (Yersinia, Salmonella, Shigella and Bordetella sp.)
and plants (Pseudomonas syringae, Ralstonia solanacearum,
Xanthomonas campestris) utilize a specialized mechanism of
secretion, termed type III secretion, to transport virulence
effector proteins across the bacterial cell envelope into the
host cell membrane or the cytosol of the target cell (Hueck,
1998). Generally the proteins secreted by type III secretion
systems are classified either as effector proteins, which act
to subvert host cell processes, or translocases, which
deliver the effector proteins into the eukaryotic cell. The
structural basis for protein translocation has not been fully
elucidated for any type III secretion system. Nevertheless,
data from studies of different type III secretion systems
suggest the presence of channel-forming proteins of bac-
terial origin, in both the bacterial outer membrane and in
the plasma membrane of the infected host cells (Hueck,
1998).
The components responsible for protein secretion are
broadly conserved in all type III secretion systems so that
one system can often export the proteins usually secreted
by another type III system (Rosqvist et al., 1995; Elliott
et al., 1999). Many of these type III components also
share homology with proteins involved in the export of
flagellar subunits (Hueck, 1998) and electron microscopy
studies of the Salmonella SPI1 and Shigella flexneri type III
secretion systems have shown the existence of a macro-
molecular complex which spans both bacterial membranes.
This complex consists of a basal-body-like structure, a
neck domain and an external needle (Kubori et al., 1998;
Blocker et al., 1999).
The enteric pathogens, enteropathogenic (EPEC) and
enterohaemorrhagic (EHEC) Escherichia coli, constitute a
significant risk to human health worldwide (reviewed by
Nataro and Kaper, 1998) and both depend on a type III
secretion system to colonize the intestinal mucosa (Jarvis
et al., 1995; Jarvis and Kaper, 1996). A hallmark of infec-
tion with either pathogen is the development of character-
istic `attaching and effacing' (A/E) lesions on the intestinal
mucosa (reviewed by Frankel et al., 1998). The genes
required for A/E lesion formation map to a chromosomal
pathogenicity island termed the LEE (Locus of Enterocyte
Q 2000 Blackwell Science Ltd
Received 25 August, 1999; revised 13 December, 1999; accepted 16December, 1999. *For correspondence. E-mail [email protected];Tel. (144) 171 594 5253; Fax (144) 171 594 5255.

1484 E. L. Hartland et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
Effacement) (McDaniel et al., 1995). The LEE-encoded
bacterial adhesion molecule intimin (Jerse et al., 1990;
Kelly et al., 1999), is essential for A/E lesion formation and
mediates intimate bacterium±host cell interaction through
binding to an intimin receptor (Tir), another LEE-encoded
protein that is delivered by the type III secretion system
into the host cell membrane (Kenny et al., 1997).
Three major E. coli secreted proteins (Esps) are known
to be exported by the LEE-encoded type III secretion
system (Elliott et al., 1998; Jarvis et al., 1995), EspA, EspB
and EspD (reviewed by Frankel et al., 1998). All three Esp
proteins are required for signal transduction and for the
formation of the A/E lesion, although until recently very
little was known about their biological function. EspA is
now known to be a structural protein and a major com-
ponent of a large (< 50 nM diameter) transiently
expressed filamentous surface organelle which, prior to
intimin-mediated intimate attachment, forms a direct link
between the bacterium and the host cell (Ebel et al., 1998;
Knutton et al., 1998). However, it is not known if EspA
binds host cells directly or via other proteins. EspB is not
thought to be a structural component of the filament
because antibodies to EspB do not stain EspA filaments
and intact EspA filaments can be observed on the surface
of an espB mutant of EPEC (Knutton et al., 1998). An
espD mutant, on the other hand, secretes only low levels of
EspA and produces barely detectable EspA filaments
(Knutton et al., 1998). At present, the reason for this is not
clear.
Following bacterial attachment, EspB is translocated
into the host cell where it is distributed in both membrane
and cytosol. This process requires the presence of EspA
filaments and is strongly enhanced by intimate bacterial
attachment (Wolff et al., 1998). EspB has weak homology
(21% identity) to YopB of Yersinia (Pallen et al., 1997), a
protein believed to form the translocation channel in the
host cell membrane through which other effectors are
delivered (Hakansson et al., 1996). While this similarity
suggests that EspB plays a similar role in EPEC/EHEC
pathogenesis, EspB also exhibits weak homology with
YopD (19% identity) and the structural organization of
EspB is more reminiscent of the YopD protein. Both pro-
teins have only one putative transmembrane region and
one predicted trimeric coiled±coil region (Pallen et al.,
1997). YopD, like EspB, is required for the translocation of
effector proteins, but is also itself translocated (Francis
and Wolf-Watz, 1998). Recently, EspB was expressed in
transfected HeLa cells where it induced changes in cell
morphology and actin stress fibres (Taylor et al., 1999). It is
possible therefore that EspB has a dual role in virulence.
We propose that in EPEC and EHEC, the EspA fila-
ments form a structural link between a bacterial mem-
brane pore (suggested by homology to be EscC, a putative
outer membrane component of the EPEC type III secretion
system) and the host cell translocation channel (EspD/
EspB), a model which implies interaction between EspA,
and the predicted pore forming proteins, EspB and EspD
(Frankel et al., 1998). Here we investigated protein±
protein interactions between EspA and EspB. The results
show that although EspA filaments can interact with host
cells when expressed in an espB deficient strain, EspB
binds EspA, is copurified with EspA filaments and local-
izes closely with EspA in tissue culture cells infected with
EPEC.
Results
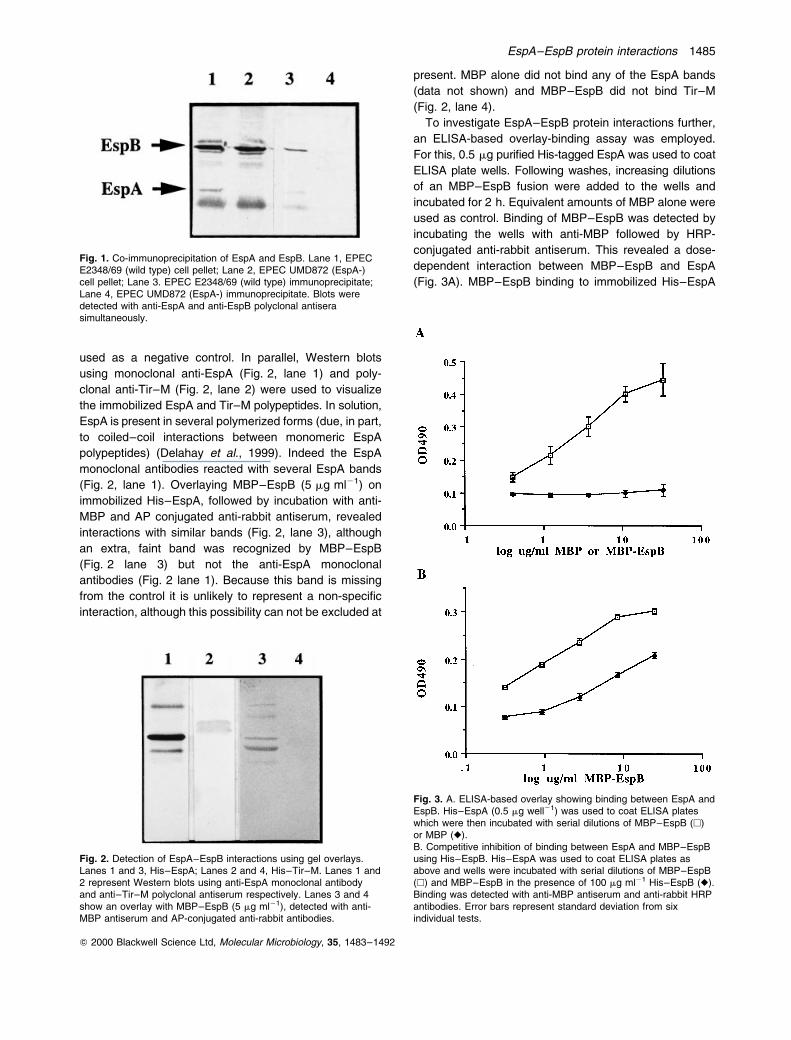
Co-immunoprecipitation of EspB and EspA with EspA
antibodies
EspA filaments are a crucial factor in EPEC pathogenesis
and are believed to act in concert with other Esps to make
up the type III protein translocation apparatus (trans-
locon). In this study, we investigated whether other Esps
associate directly with the EspA filaments during bacterial
growth by immunoprecipitating the filaments with mono-
clonal antibodies to EspA. The prototype wild-type EPEC
strain (E2348/69) was grown under conditions that favour
EspA filament production. An espA mutant derivative of
E2348/69 (UMD872) (Kenny et al., 1996) was used as a
negative control. Following a 3 h incubation, the bacteria
were washed, incubated with the mouse monoclonal
EspA antibodies overnight and then immunoprecipitated
with protein-G agarose beads. The immunoprecipitated
material and bacterial pellets were subjected to Western
blots using rabbit polyclonal EspA and mouse polyclonal
EspB antisera. EspA was specifically detected in both the
immunoprecipitated material and bacterial cell pellet from
E2348/69. EspA could not be detected in either sample
obtained from UMD872. As expected, EspB was detected
in the bacterial cell pellets of both E2348/69 and UMD872
whereas EspB was detected in the immuno-precipitated
material from E2348/69 only (Fig. 1). EspB was not
detected in the bacterial cell pellet or immunoprecipitated
material of an espB mutant, strain UMD864 (Donnenberg
et al., 1993) (data not shown). These results suggest that
EspB is associated with EspA filaments during bacterial
growth and in the absence of host cells.
Binding of EspB and EspA on solid phase
To verify the specificity of the association, EspA and EspB
were subjected to a number of in vitro binding assays to
examine protein±protein interactions. Purified His±EspA
and MBP±EspB fusion proteins were used in gel
overlays to probe the ability of EspB to bind EspA.
Immobilized His±Tir±M (a 107-amino-acid Tir fragment
involved in intimin binding) (Hartland et al., 1999) was
EspA±EspB protein interactions 1485
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
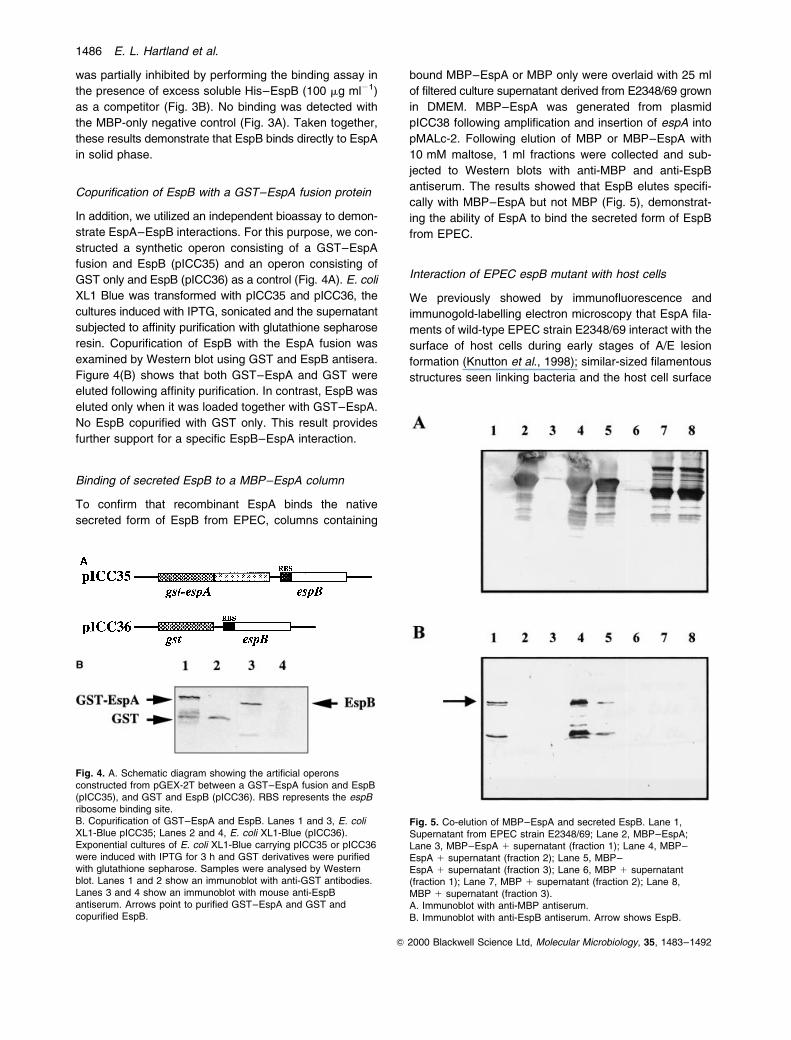
used as a negative control. In parallel, Western blots
using monoclonal anti-EspA (Fig. 2, lane 1) and poly-
clonal anti-Tir±M (Fig. 2, lane 2) were used to visualize
the immobilized EspA and Tir±M polypeptides. In solution,
EspA is present in several polymerized forms (due, in part,
to coiled±coil interactions between monomeric EspA
polypeptides) (Delahay et al., 1999). Indeed the EspA
monoclonal antibodies reacted with several EspA bands
(Fig. 2, lane 1). Overlaying MBP±EspB (5 mg ml21) on
immobilized His±EspA, followed by incubation with anti-
MBP and AP conjugated anti-rabbit antiserum, revealed
interactions with similar bands (Fig. 2, lane 3), although
an extra, faint band was recognized by MBP±EspB
(Fig. 2 lane 3) but not the anti-EspA monoclonal
antibodies (Fig. 2 lane 1). Because this band is missing
from the control it is unlikely to represent a non-specific
interaction, although this possibility can not be excluded at
present. MBP alone did not bind any of the EspA bands
(data not shown) and MBP±EspB did not bind Tir±M
(Fig. 2, lane 4).
To investigate EspA±EspB protein interactions further,
an ELISA-based overlay-binding assay was employed.
For this, 0.5 mg purified His-tagged EspA was used to coat
ELISA plate wells. Following washes, increasing dilutions
of an MBP±EspB fusion were added to the wells and
incubated for 2 h. Equivalent amounts of MBP alone were
used as control. Binding of MBP±EspB was detected by
incubating the wells with anti-MBP followed by HRP-
conjugated anti-rabbit antiserum. This revealed a dose-
dependent interaction between MBP±EspB and EspA
(Fig. 3A). MBP±EspB binding to immobilized His±EspA
Fig. 2. Detection of EspA±EspB interactions using gel overlays.Lanes 1 and 3, His±EspA; Lanes 2 and 4, His±Tir±M. Lanes 1 and2 represent Western blots using anti-EspA monoclonal antibodyand anti±Tir±M polyclonal antiserum respectively. Lanes 3 and 4show an overlay with MBP±EspB (5 mg ml21), detected with anti-MBP antiserum and AP-conjugated anti-rabbit antibodies.
Fig. 1. Co-immunoprecipitation of EspA and EspB. Lane 1, EPECE2348/69 (wild type) cell pellet; Lane 2, EPEC UMD872 (EspA-)cell pellet; Lane 3. EPEC E2348/69 (wild type) immunoprecipitate;Lane 4, EPEC UMD872 (EspA-) immunoprecipitate. Blots weredetected with anti-EspA and anti-EspB polyclonal antiserasimultaneously.
Fig. 3. A. ELISA-based overlay showing binding between EspA andEspB. His±EspA (0.5 mg well21) was used to coat ELISA plateswhich were then incubated with serial dilutions of MBP±EspB (A)or MBP (V).B. Competitive inhibition of binding between EspA and MBP±EspBusing His±EspB. His±EspA was used to coat ELISA plates asabove and wells were incubated with serial dilutions of MBP±EspB(A) and MBP±EspB in the presence of 100 mg ml21 His±EspB (V).Binding was detected with anti-MBP antiserum and anti-rabbit HRPantibodies. Error bars represent standard deviation from sixindividual tests.
1486 E. L. Hartland et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
was partially inhibited by performing the binding assay in
the presence of excess soluble His±EspB (100 mg ml21)
as a competitor (Fig. 3B). No binding was detected with
the MBP-only negative control (Fig. 3A). Taken together,
these results demonstrate that EspB binds directly to EspA
in solid phase.
Copurification of EspB with a GST±EspA fusion protein
In addition, we utilized an independent bioassay to demon-
strate EspA±EspB interactions. For this purpose, we con-
structed a synthetic operon consisting of a GST±EspA
fusion and EspB (pICC35) and an operon consisting of
GST only and EspB (pICC36) as a control (Fig. 4A). E. coli
XL1 Blue was transformed with pICC35 and pICC36, the
cultures induced with IPTG, sonicated and the supernatant
subjected to affinity purification with glutathione sepharose
resin. Copurification of EspB with the EspA fusion was
examined by Western blot using GST and EspB antisera.
Figure 4(B) shows that both GST±EspA and GST were
eluted following affinity purification. In contrast, EspB was
eluted only when it was loaded together with GST±EspA.
No EspB copurified with GST only. This result provides
further support for a specific EspB±EspA interaction.
Binding of secreted EspB to a MBP±EspA column
To confirm that recombinant EspA binds the native
secreted form of EspB from EPEC, columns containing
bound MBP±EspA or MBP only were overlaid with 25 ml
of filtered culture supernatant derived from E2348/69 grown
in DMEM. MBP±EspA was generated from plasmid
pICC38 following amplification and insertion of espA into
pMALc-2. Following elution of MBP or MBP±EspA with
10 mM maltose, 1 ml fractions were collected and sub-
jected to Western blots with anti-MBP and anti-EspB
antiserum. The results showed that EspB elutes specifi-
cally with MBP±EspA but not MBP (Fig. 5), demonstrat-
ing the ability of EspA to bind the secreted form of EspB
from EPEC.
Interaction of EPEC espB mutant with host cells
We previously showed by immunofluorescence and
immunogold-labelling electron microscopy that EspA fila-
ments of wild-type EPEC strain E2348/69 interact with the
surface of host cells during early stages of A/E lesion
formation (Knutton et al., 1998); similar-sized filamentous
structures seen linking bacteria and the host cell surface
Fig. 4. A. Schematic diagram showing the artificial operonsconstructed from pGEX-2T between a GST±EspA fusion and EspB(pICC35), and GST and EspB (pICC36). RBS represents the espBribosome binding site.B. Copurification of GST±EspA and EspB. Lanes 1 and 3, E. coliXL1-Blue pICC35; Lanes 2 and 4, E. coli XL1-Blue (pICC36).Exponential cultures of E. coli XL1-Blue carrying pICC35 or pICC36were induced with IPTG for 3 h and GST derivatives were purifiedwith glutathione sepharose. Samples were analysed by Westernblot. Lanes 1 and 2 show an immunoblot with anti-GST antibodies.Lanes 3 and 4 show an immunoblot with mouse anti-EspBantiserum. Arrows point to purified GST±EspA and GST andcopurified EspB.
Fig. 5. Co-elution of MBP±EspA and secreted EspB. Lane 1,Supernatant from EPEC strain E2348/69; Lane 2, MBP±EspA;Lane 3, MBP±EspA 1 supernatant (fraction 1); Lane 4, MBP±EspA 1 supernatant (fraction 2); Lane 5, MBP±EspA 1 supernatant (fraction 3); Lane 6, MBP 1 supernatant(fraction 1); Lane 7, MBP 1 supernatant (fraction 2); Lane 8,MBP 1 supernatant (fraction 3).A. Immunoblot with anti-MBP antiserum.B. Immunoblot with anti-EspB antiserum. Arrow shows EspB.
EspA±EspB protein interactions 1487
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
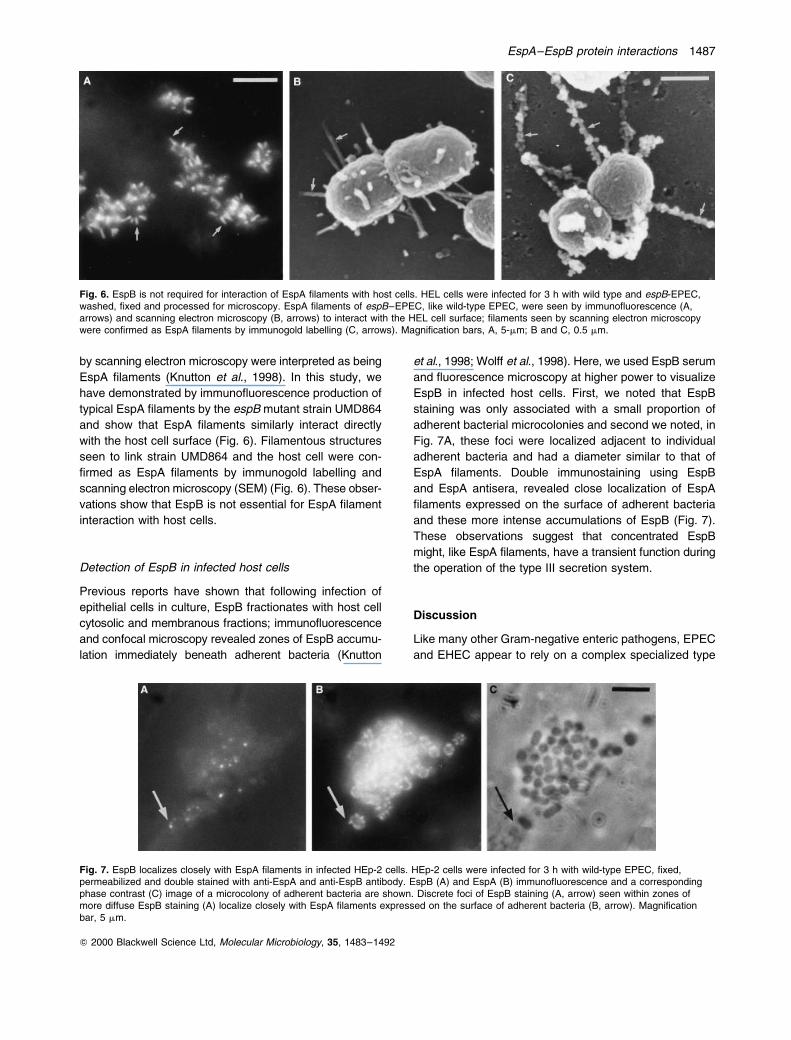
by scanning electron microscopy were interpreted as being
EspA filaments (Knutton et al., 1998). In this study, we
have demonstrated by immunofluorescence production of
typical EspA filaments by the espB mutant strain UMD864
and show that EspA filaments similarly interact directly
with the host cell surface (Fig. 6). Filamentous structures
seen to link strain UMD864 and the host cell were con-
firmed as EspA filaments by immunogold labelling and
scanning electron microscopy (SEM) (Fig. 6). These obser-
vations show that EspB is not essential for EspA filament
interaction with host cells.
Detection of EspB in infected host cells
Previous reports have shown that following infection of
epithelial cells in culture, EspB fractionates with host cell
cytosolic and membranous fractions; immunofluorescence
and confocal microscopy revealed zones of EspB accumu-
lation immediately beneath adherent bacteria (Knutton
et al., 1998; Wolff et al., 1998). Here, we used EspB serum
and fluorescence microscopy at higher power to visualize
EspB in infected host cells. First, we noted that EspB
staining was only associated with a small proportion of
adherent bacterial microcolonies and second we noted, in
Fig. 7A, these foci were localized adjacent to individual
adherent bacteria and had a diameter similar to that of
EspA filaments. Double immunostaining using EspB
and EspA antisera, revealed close localization of EspA
filaments expressed on the surface of adherent bacteria
and these more intense accumulations of EspB (Fig. 7).
These observations suggest that concentrated EspB
might, like EspA filaments, have a transient function during
the operation of the type III secretion system.
Discussion
Like many other Gram-negative enteric pathogens, EPEC
and EHEC appear to rely on a complex specialized type
Fig. 6. EspB is not required for interaction of EspA filaments with host cells. HEL cells were infected for 3 h with wild type and espB-EPEC,washed, fixed and processed for microscopy. EspA filaments of espB±EPEC, like wild-type EPEC, were seen by immunofluorescence (A,arrows) and scanning electron microscopy (B, arrows) to interact with the HEL cell surface; filaments seen by scanning electron microscopywere confirmed as EspA filaments by immunogold labelling (C, arrows). Magnification bars, A, 5-mm; B and C, 0.5 mm.
Fig. 7. EspB localizes closely with EspA filaments in infected HEp-2 cells. HEp-2 cells were infected for 3 h with wild-type EPEC, fixed,permeabilized and double stained with anti-EspA and anti-EspB antibody. EspB (A) and EspA (B) immunofluorescence and a correspondingphase contrast (C) image of a microcolony of adherent bacteria are shown. Discrete foci of EspB staining (A, arrow) seen within zones ofmore diffuse EspB staining (A) localize closely with EspA filaments expressed on the surface of adherent bacteria (B, arrow). Magnificationbar, 5 mm.

1488 E. L. Hartland et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
III secretion system for their exploitation of host cell
machinery (Hueck, 1998). In EPEC, three major Esp
proteins are secreted by the type III secretion system and
their precise function is the subject of intense investiga-
tion. Because Esps are known to be essential for signal
transduction and A/E lesion formation, elucidating their
role in virulence is crucial for the understanding of EPEC
pathogenesis. Using a molecular approach to clone, purify
and raise antiserum to EspA, we and others showed that
EspA is a structural protein and a major component of a
large filamentous surface organelle (Ebel et al., 1998;
Knutton et al., 1998), the precise structure and composi-
tion of which we are now investigating. Recently, we
demonstrated that the EspA filaments are made up of
multimeric EspA and that intermolecular EspA coiled±coil
interactions are essential for the structural integrity of the
filaments (Delahay et al., 1999). In this study, we have
investigated protein±protein interactions between the
EspA filaments and EspB. We have shown that EspB is
copurified with the EspA filaments in the absence of host
cells and that EspB binds EspA in solid and liquid phase.
We also demonstrate that EspB associates closely with
EspA filaments on infected host cells but that EspB is not
required for attachment of the filaments to the host cell
surface.
Given that EspB copurifies with EspA filaments in the
absence of host cells, it is possible that EspB is located
inside the filament prior to infection where it is inacces-
sible to anti-EspB antibodies or folded into a form that is
not recognized and is then released only upon contact
with the host cell surface. So far we have been unable to
demonstrate co-immunoprecipitation of EspA filaments with
EspD or Tir, suggesting that these Esps are not located in
the filament prior to host cell attachment (unpublished
data).
EspB is a translocated protein that requires both the
EspA filaments and functional EspB to be transported into
the host cell where it is found in both the cell membrane
and the cytosol (Knutton et al., 1998; Wolff et al., 1998).
Initial studies failed to show staining of the EspA filaments
with polyclonal EspB antiserum, suggesting that EspB is
not a structural component of the filament. Here we show
that EspA filaments can interact with host cells in the
absence of EspB, demonstrating that EspB is not the
initial receptor for the filament. EspB [and EspD, which
was also shown to be translocated to the host cell mem-
brane (Wachter et al., 1999)] has weak structural similarity
to YopB and YopD, two proteins secreted by Yersinia that
form a translocation channel through which cytotoxins are
delivered to the host cell (Hakansson et al., 1996; Tardy
et al., 1999). However, to date no EspA filament-like struc-
tures have been observed in Yersinia. Importantly, the
results presented here show that attachment of EspA
filaments to host cells can be distinguished from protein
translocation. Based on similarity of EspB/D with YopD/B
(Neyt and Cornelis, 1999a) and IpB/C (Menard et al., 1996;
Blocker et al., 1999), if a pore is produced by a complex of
these Esp proteins, then EspA filaments do not need a
functional pore to bind to the host cell surface; the associ-
ation of EspB/D in the membrane and their presumed
interaction with EspA filaments must be a subsequent
event.
Evidence supporting the formation of membrane pores
was demonstrated using contact haemolytic activity (Hakans-
son et al., 1996) and we (unpublished observations) and
others (Warawa et al., 1999) have recently shown that
EPEC induce haemolysis of red blood cells. In support of
EspB generated pores in the red blood cell membrane is
the observation that, unlike wild-type EPEC, strain UMD864
(espB±) was unable to lyse red blood cells. In this paper,
high power fluorescence microscopy of translocated EspB
revealed discrete foci of fluorescent staining closely
adjacent to adherent bacteria and with a diameter similar
to that of the EspA filaments. Although, at the present
time, we are unable to precisely define the localization of
such EspB staining, the discrete nature of the fluores-
cence suggests that localized EspB could be concen-
trated in the host cell membrane. Previous studies have
shown that in the presence of host cells EspB is protease
resistant (Kenny and Finlay, 1995), suggesting this pro-
tein does not have a surface location. However, it is likely
that EspB is protected from proteolytic degradation by its
interaction with EspA and insertion into the host membrane.
As mentioned above, EspB also has similarity with
YopD, which is translocated into the host cell cytosol and
membrane by Yersinia (Francis and Wolf-Watz, 1998).
Although the precise mechanism of action of YopD remains
unclear, this protein appears to have direct roles in viru-
lence gene regulation and translocation, as well as pos-
sible functions in the host cell. Thus, although EspB
appears to form part of the type III protein translocation
apparatus with EspA and possibly with EspD, it may also
have a cytosolic function.
EscC (formerly SepC) is a major component of the
EPEC type III secretion system and is predicted to be
exposed at the bacterial surface (Elliott et al., 1998). YscC,
a Yersinia homologue of EscC, has recently been shown to
form a ring-shaped oligomeric complex in the outer mem-
brane with an < 20 nM diameter central pore (Koster et al.,
1997). Thus, EscC may also represent this type of novel
channel-forming protein in the bacterial outer membrane.
Our working hypothesis is that proteins are carried from
pathogenic bacteria to the host cell through a pore in
hollow EspA-like filaments which bridge the bacterial (EscC)
and host cell membrane (EspB and EspD) channels.
Although it remains to be shown if the separate com-
ponents of the proposed EPEC translocon are function-
ally integrated as suggested, such an apparatus could

EspA±EspB protein interactions 1489
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
function as a `molecular syringe' which, powered by the
type III secretion system ATPase, EscN, injects proteins
directly from the bacterium into the host cell cytosol
(Frankel et al., 1999).
In conclusion, our observations now suggest that the
EspA filaments might have a dual function: they may func-
tion initially to sense host target cells and subsequently,
following pore formation, function as an integral part of the
type III translocon. The data presented in this study show,
for the first time, protein±protein interactions between
different Esps. We believe that this provides an important
step forward in our understanding of EPEC and EHEC
pathogenesis and the function of type III secretion
systems in general.
Experimental procedures
Bacterial strains and plasmids
Bacterial strains used in this study include EPEC strainsE2348/69 (wild type), UMD872 (espA-) (Kenny et al., 1996)and UMD864 (espB-) (Donnenberg et al., 1993), E. coli XL-1Blue and BL21. Strains were grown in Luria broth or on Luriaagar supplemented with kanamycin (100 mg ml21) or ampi-cillin (100 mg ml21) where appropriate. The plasmids usedare listed in Table 1.
Preparation of monoclonal antibodies to EspA
For the production of monoclonal antibodies, BALB/c micewere immunized by two subcutaneous injections with 90 mgof purified His±EspA (Knutton et al., 1998) in incompleteFreund's adjuvant. A final intraperitoneal injection of antigen(90 mg) without adjuvant was performed 4 days before themouse was killed and the spleen removed. Splenocytes wereprepared and fused with SP2/0 mouse myeloma cells usingpolyethylene glycol 1500 (Boehringer) following standardprocedures (Harlow and Lane, 1988). The fusion mix wasdiluted in RPMI/HAT medium with 10% foetal bovine serumand plated into 96-well plates seeded 48 h previously withmouse peritoneal macrophages. The plates were incubatedat 378C in a 5% CO2 atmosphere for 10 days when samples
of the supernatant were taken to test for antibody. Positivewells were subcultured and the hybridoma cells cloned twiceby limiting dilution. Four lines, HF7, HA9, AC2 and EG4 wereconsidered to be monoclonal. All four lines were isotyped asIgG1, and were able to detect secreted EspA by ELISA andWestern blot. Culture supernatant (50 ml) containing anti-body secreted from clone HA9 was selected for purificationusing a Protein G Sepharose column (Pharmacia) followingstandard techniques.
Preparation of mouse polyclonal antiserum to EspB
His±EspB was purified as described previously (Frankelet al., 1996). Six- to eight-week-old TO mice were immunizedsubcutaneously with < 20 mg purified His-EspB and wereboosted twice with the same antigen in incomplete His±EspBin complete Freund's adjuvant. Animals Freund's adjuvantwith 2 week intervals before exsanguination. Antiserum wastested against purified EspB by Western blotting.
Co-immunoprecipitation of EspA and EspB
EPEC strains E2348/69 and UMD872 (EspA-) were grownin 5 ml Luria±Bertani (LB) standing at 378C overnight. Theovernight cultures were used to inoculate 5 mls of DMEMcontaining 10% fetal calf serum at a 1:100 dilution. Thecultures were incubated at 378C without agitation for 3 h andcentrifuged at 4000 r.p.m. for 10 min at 48C to pellet the cells.The cell pellets were resuspended gently in 500 ml PBS andre-centrifuged. This was repeated twice. Cell pellets were theneither used for immunoprecipitation, or resuspended in100 ml of Laemmli buffer for SDS±PAGE and Westernblotting. For immunoprecipitation, the cell pellets were resus-pended in 50 ml of PBS and 5 ml of monoclonal anti-EspAantibody was added. The samples were incubated at 48Covernight. EspA filaments were then gently dislodged fromthe bacteria and the samples centrifuged at 15 000 r.p.m. ina microfuge for 5 min to pellet the bacterial cells. The super-natant was transferred to a fresh tube and the filamentsprecipitated with 10 ml of a 50% protein G-agarose beadslurry. The samples were centrifuged (as above) to pellet theprotein G-beads. The beads were washed in 50 ml of PBSand resuspended in 100 ml of Laemmli buffer for analysis bySDS±PAGE and Western blotting.
Table 1. Plasmids used in this study.
Plasmid Relevant features Reference
pETespA pET28a expressing His-tagged EspA Knutton et al. (1998)pETespB pET28a expressing His-tagged EspB Frankel et al. (1996)pMALc-2 Cloning vector generating
translational fusions with MBPNEB
pGEX-2T Cloning vector generatingtranslational fusions with GST
Pharmacia
pICC35 pGEX-2T expressing GST±EspAfusion protein and EspB
This study
pICC36 pGEX-2T expressing GST and EspB This studypICC37 pMALc-2 expressing MBP±EspB
fusion proteinThis study
pICC38 pMALc-2 expressing MBP±EspAfusion protein
This study

1490 E. L. Hartland et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
EspA/EspB gel overlays
His±EspA was purified as described previously (Knuttonet al., 1998). EspB was amplified by PCR as describedpreviously (Frankel et al., 1996) and cloned into pMALc-2(NEB) to generate a translational fusion with maltose bindingprotein (MBP) (pICC37). The recombinant plasmids weretransformed into E. coli TG1 and log phase cultures wereinduced with 1 mM IPTG followed by incubation at 378C for3 h with shaking. MBP±EspB was purified with amylose resinas described (Frankel et al., 1994).
His±EspA and His±Tir±M were separated by SDS±PAGE,blotted onto nitrocellulose and blocked with 10% skimmedmilk in PBS, 0.1% Tween-20 for 1 h. Membranes were incu-bated with MBP±EspB or MBP (5 mg ml21) overnight at 48C,washed and detected with anti-MBP antiserum and AP-conjugated anti-rabbit antibodies as described previously(Hartland et al., 1999).
EspA/EspB ELISA overlays
Purified His±EspA (5 mg ml21) in carbonate/bicarbonate bufferwas used to coat an ELISA plate (100 ml well21). Wells wereincubated with serial dilutions of MBP±EspB or MBP alone(100 ml well21) for 2 h and then detected with anti-MBPantiserum (1/2000 dilution, 1.5 h) followed by HRP-conjugatedanti-rabbit antiserum (1/2000, 1.5 h). The colour reaction wasdeveloped with o-phenylenediamine(OPD)/H2O2 (Sigma) andstopped with 3 M sulphuric acid. The final OD was measuredat 490 nM.
Construction of gst±espA, espB synthetic operons
espA was amplified by PCR from EPEC strain 2348/69 usingthe primer pair, 5 0-CGGGATCCATGGATACATCAACTAC-3 0
and 5 0-CGGGATCCTTATTTACCAAGGGATATTCC-3 0. ThePCR product was cloned into the BamHI site of pGEX-2T(Pharmacia) to generate a translational fusion with gluta-thione S-transferase (GST) (Neyt and Cornelis, 1999b). espBwas amplified by PCR using the primers 5 0-CGGAATTCT-TATTAAAGAGGCGTTTATTATG-3 0 and 5 0-CGGAATTCTT-ACCCAGCTAAGCGAGC-3 0, which were designed to includethe native ribosome binding site of espB. This fragment wascloned downstream of the GST±EspA fusion into the EcoRIsite of pGEX-2T to generate a transcriptional fusion with gst±espA. The resulting plasmid was called pICC35. A negativecontrol plasmid (gst, espB) was constructed by redigestingpICC35 with BamHI to remove espA. 5 0 overhangs were thenrepaired with Klenow enzyme to generate blunt ends andintroduce a stop codon after gst. This plasmid was religatedand called pICC36. PICC35 and pICC36 were transformedinto E. coli XL1-Blue.
Copurification of GST±EspA and EspB
Production of GST or GST±EspA was induced after theaddition of 1 mM IPTG to log phase cultures and incubationat 378C for 3 h with shaking. For purification of GST proteins,induced cultures were sonicated and 10 ml clarified sonicatewas mixed with 100 ml (50% slurry) glutathione sepharose 4Bbeads (Pharmacia). After 30 min, beads were washed three
times with 1 ml PBS and resuspended directly into 50 ml ofLaemmli buffer for analysis by SDS±PAGE and Westernblotting.
MBP±EspA/EspB affinity column
espA was amplified by PCR from EPEC strain 2348/69 usingthe primer pair, 5 0-GGAATTCATGGATACATCAACTACAG-CATCA-3 0 and 5 0-TATCTGCAGTTATTTACCAAGGGATATTCCTG-3 0. The PCR product was cloned into the site of pMALc-2(NEB) to generate a translational fusion with MBP. Therecombinant plasmid was transformed into E. coli TG1 andlog phase cultures were induced with 1 mM IPTG followed byincubation at 378C for 3 h with shaking. MBP±EspA waspurified with amylose resin as described (Frankel et al.,1994). For affinity columns, MBP or MBP±EspA was boundto amylose resin according to the standard purificationprocedure (Frankel et al., 1994). The column was thenoverlaid with 25 ml of filtered culture supernatant from EPECstrain 2348/69 grown overnight in DMEM. Following washeswith 10 volumes of column buffer (50 mM Tris-Cl pH 7.4,200 mM NaCl, 1 mM EDTA), MBP or MBP±EspA andassociated proteins were eluted with 10 mM maltosedissolved in column buffer. Fractions were collected in 1 mlvolumes and 15 ml of each fraction was subjected to SDS±PAGE and immunoblotting with anti-MBP and anti-EspB andAP-conjugated anti-rabbit antibodies.
Immunofluorescence and scanning electron microscopy
Epithelial cells [HEp-2, HEL (human embryonic lung)] wereseeded and grown overnight on glass coverslips. After a 3 hEPEC infection, cells were washed with PBS and fixed ineither 4% formalin (immunofluorescence) or 0.1% glutaralde-hyde (SEM).
For immunofluorescence, fixed cells were washed threetimes with PBS, permeabilized with 0.1% Triton X-100 in PBSfor 4 min and washed a further three times with PBS. Cellswere incubated with 20 ml of primary (anti-EspA, anti-EspB)antibodies for 45 min, washed and incubated with 20 ml ofsecondary Alexa488 conjugated anti-rabbit IgG antibody(Molecular Probes). For double staining of EspA and EspB, aprimary mouse anti-EspA and secondary anti-mouse Alexa594conjugated IgG antibody was used. Samples were examinedand photographed with a Leitz Dialux microscope.
For scanning electron microscopy, glutaraldehyde fixedHEL cell monolayers were washed, incubated with EspAantiserum for 2 h, washed again and fixed in 3% bufferedglutaraldehyde. For immunogold labelling, samples wereincubated for 2 h with 30 nM gold conjugated anti-rabbit IgG(British BioCell) prior to fixation in 3% glutaraldehyde. Mono-layers were post-fixed in 1% osmium tetroxide, dehydratedthrough graded acetone solutions and critical point dried.Mounted specimens were sputter coated with platinum(Polaron) and examined in a Jeol 1200EX scanning trans-mission electron microscope operated at 40 kV.
Acknowledgements
E.L.H. is the recipient of a Royal Society/NHMRC Howard Florey
Fellowship. This work was supported by the BBSRC and the
Leverhulme and Wellcome Trusts.

EspA±EspB protein interactions 1491
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
References
Blocker, A., Gounon, P., Larquet, E., Niebuhr, K., Cabiaux, V.,
Parsot, C., et al. (1999) The tripartite type III secreton of
Shigella flexneri inserts IpaB and IpaC into host membranes. JCell Biol 147: 683±693.
Delahay, R.M., Knutton, S., Shaw, R.K., Hartland, E.L., Pallen,
M.J., and Frankel, G. (1999) The coiled-coil domain of EspA is
essential for the assembly of the type III secretion transloconon the surface of enteropathogenic E coli. J Biol Chem 274:
35969±35974.
Donnenberg, M.S., Yu, J., and Kaper, J.B. (1993) A secondchromosomal gene necessary for intimate attachment of
enteropathogenic Escherichia coli to epithelial cells. J Bacteriol
75: 4670±4680.
Ebel, F., Podzadel, T., Rohde, M., Kresse, A.U., Kramer, S.,Deibel, C., et al. (1998) Initial binding of Shiga toxin-producing
Escherichia coli to host cells and subsequent induction of actin
rearrangements depend on filamentous EspA-containing sur-face appendages. Mol Microbiol 30: 147±161.
Elliott, S.J., Dubois, M.S., Hutcheson, S.W., Wainwright, L.A.,
Batchelor, M., Frankel, G., et al. (1999) Identification of CesT, a
chaperone for the type III secretion of Tir in EnteropathogenicEscherichia coli. Mol Microbiol 33: 1176±1189.
Elliott, S.J., Wainwright, L.A., McDaniel, T.K., Jarvis, K.G., Deng,
Y.K., Lai, L.C., et al. (1998) The complete sequence of thelocus of enterocyte effacement (LEE) from enteropathogenic
Escherichia coli E2348/69. Mol Microbiol 28: 1±4.
Francis, M.S., and Wolf-Watz, H. (1998) YopD of Yersinia
pseudotuberculosis is translocated into the cytosol of HeLaepithelial cells: evidence of a structural domain necessary for
translocation. Mol Microbiol 29: 799±813.
Frankel, G., Candy, D.C., Everest, P., and Dougan, G. (1994)Characterization of the C-terminal domains of intimin-like pro-
teins of enteropathogenic and enterohemorrhagic Escherichia
coli, Citrobacter freundii, and Hafnia alvei. Infect Immun 62:
1835±1842.
Frankel, G., Phillips, A.D., Novakova, M., Field, H., Candy, D.C.,
Schauer, D.B., et al. (1996) Intimin from enteropathogenic
Escherichia coli restores murine virulence to a Citrobacterrodentium eaeA mutant: induction of an immunoglobulin A
response to intimin and EspB. Infect Immun 64: 5315±5325.
Frankel, G., Phillips, A.D., Rosenshine, I., Dougan, G., Kaper,J.B., and Knutton, S. (1998) Enteropathogenic and entero-
haemorrhagic Escherichia coli: More Subversive Elements.
Mol Microbiol 30: 911±921.
Frankel, G., Wren, B., Dougan, G., Delahay, R.M., Batchelor, M.,Hale, C., et al. (1999) Host/pathogen interactions during
infection by enteropathogenic Escherichia coli: a role for
signaling. In Microbial Signaling and Communication. England,
R., Hobbs, G., Bainton, N., and Roberts, D. McL. (eds). Societyfor General Microbiology Symposium 57. Cambridge, UK:
Cambridge University Press, 197±213.
Hakansson, S., Schesser, K., Persson, C., Galyov, E.E.,Rosqvist, R., Homble, F., et al. (1996) The YopB protein of
Yersinia pseudotuberculosis is essential for the translocation of
Yop effector proteins across the target cell plasma membrane
and displays a contact-dependent membrane disruptingactivity. EMBO J 15: 5812±5823.
Harlow, E., and Lane, D., (eds)(1988) Antibodies, A Laboratory
Manual. Cold Spring Harbour, NY: Cold Spring HarbourLaboratory Press, p. 211.
Hartland, E.L., Batchelor, M., Delahay, R.M., Hale, C., Matthews,
S., Dougan, G., et al. (1999) Binding of intimin from
enteropathogenic Escherichia coli to Tir and to host cells.Mol Microbiol 32: 151±158.
Hueck, C.J. (1998) Type III protein secretion systems in bacterial
pathogens of animals and plants. Microbiol Mol Biol Rev 62:
379±433.
Jarvis, K.G., Giron, J.A., Jerse, A.E., McDaniel, T.K., Donnenberg,M.S., and Kaper, J.B. (1995) Enteropathogenic Escherichia coli
contains a putative type III secretion system necessary for the
export of proteins involved in attaching and effacing lesion
formation. Proc Natl Acad Sci USA 92: 7996±8000.
Jarvis, K.G., and Kaper, J.B. (1996) Secretion of extracellularproteins by enterohemorrhagic Escherichia coli via a putative
type III secretion system. Infect Immun 64: 4826±4829.
Jerse, A.E., Yu, J., Tall, B.D., and Kaper, J.B. (1990) A genetic
locus of enteropathogenic Escherichia coli necessary for theproduction of attaching and effacing lesions on tissue culture
cells. Proc Natl Acad Sci USA 87: 7839±7843.
Kelly, G., Prasannan, S., Daniell, S., Fleming, K., Frankel, G.,
Dougan, G., et al. (1999) Structure of the cell-adhesion
fragment of intimin from enteropathogenic Escherichia coli.Nature Struct Biol 6: 313±318.
Kenny, B., DeVinney, R., Stein, M., Reinscheid, D.J., Frey, E.A.,
and Finlay, B.B. (1997) Enteropathogenic E. coli (EPEC)
transfers its receptor for intimate adherence into mammalian
cells. Cell 91: 511±520.
Kenny, B., and Finlay, B.B. (1995) Protein secretion by entero-
pathogenic Escherichia coli is essential for transducing signals
to epithelial cells. Proc Natl Acad Sci USA 92: 7991±7995.
Kenny, B., Lai, L.C., Finlay, B.B., and Donnenberg, M.S. (1996)
EspA, a protein secreted by enteropathogenic Escherichia coli,is required to induce signals in epithelial cells. Mol Microbiol 20:
313±323.
Knutton, S., Rosenshine, I., Pallen, M.J., Nisan, I., Neves, B.C.,
Bain, C., et al. (1998) A novel EspA-associated surface organ-elle of enteropathogenic Escherichia coli involved in protein
translocation into epithelial cells. EMBO J 17: 2166±2176.
Koster, M., Bitter, W., de Cock, H., Allaoui, A., Cornelis, G.R.,
and Tommassen, J. (1997) The outer membrane component,
YscC, of the Yop secretion machinery of Yersinia enterocoliticaforms a ring-shaped multimeric complex. Mol Microbiol 26:
789±797.
Kubori, T., Matsushima, Y., Nakamura, D., Uralil, J., Lara-Tejero,
M., Sukhan, A., et al. (1998) Supramolecular structure of the
Salmonella typhimurium type III protein secretion system.Science 280: 602±605.
McDaniel, T.K., Jarvis, K.G., Donnenberg, M.S., and Kaper, J.B.
(1995) A genetic locus of enterocyte effacement conserved
among diverse enterobacterial pathogens. Proc Natl Acad SciUSA 92: 1664±1668.
Menard, R., Prevost, M.C., Gounon, P., Sansonetti, P., and
Dehio, C. (1996) The secreted Ipa complex of Shigella flexneri
promotes entry into mammalian cells. Proc Natl Acad Sci USA
93: 1254±1258.
Nataro, J.P., and Kaper, J.B. (1998) Diarrheagenic Escherichia
coli. Clin Microbiol Rev 11: 142±201.
Neyt, C., and Cornelis, G.R. (1999a) Insertion of a Yop
translocation pore into the macrophage plasma membrane by
Yersinia enterocolitica: requirement for translocators YopB andYopD, but not LcrG. Mol Microbiol 33: 971±981.
Neyt, C., and Cornelis, G.R. (1999b) Role of SycD, the chaper-
one of the Yersinia Yop translocators YopB and YopD. Mol
Microbiol 31: 143±156.
Pallen, M.J., Dougan, G., and Frankel, G. (1997) Coiled-coil

1492 E. L. Hartland et al.
Q 2000 Blackwell Science Ltd, Molecular Microbiology, 35, 1483±1492
domains in proteins secreted by type III secretion systems. MolMicrobiol 25: 423±425.
Rosqvist, R., Hakansson, S., Forsberg, A., and Wolf-Watz, H.(1995) Functional conservation of the secretion and transloca-
tion machinery for virulence proteins of yersiniae, salmonellae
and shigellae. EMBO J 14: 4187±4195.
Tardy, F., HombleÂ, F., Neyt, C., Wattiez, R., Cornelis, G.R.,
Ruysschaert, J.-M., et al. (1999) Yersinia enterocolitica type IIIsecretion-translocation system: channel formation by secreted
Yops. EMBO J 18: 6793±6799.
Taylor, K.A., Luther, P.W., and Donnenberg, M.S. (1999) Expres-
sion of the EspB protein of enteropathogenic Escherichia coli
within HeLa cells affects stress fibers and cellular morphology.Infect Immun 67: 120±125.
Wachter, C., Beinke, C., Mattes, M., and Schmidt, M.A. (1999)
Insertion of EspD into epithelial target cell membranes by
infecting enteropathogenic Escherichia coli. Mol Microbiol 31:1695±1707.
Warawa, J., Finlay, B.B., and Kenny, B. (1999) Type III secretion-
dependent hemolytic activity of enteropathogenic Escherichiacoli. Infect Immun 67: 5538±5540.
Wolff, C., Nisan, I., Hanski, E., Frankel, G., and Rosenshine, I.
(1998) Protein translocation into HeLa cells by infecting
enteropathogenic Escherichia coli. Mol Microbiol 28: 143±155.
Copyright © 2022 FDOKUMEN




















