
Global analysis of carbohydrate utilization by Lactobacillus acidophilus using cDNA microarrays
Upload
nottinghamCategory
view
1download
0
Frontline:
Carbohydrate-independent recognition of collagensby the macrophage mannose receptor
Luisa Martinez-Pomares*1, Dirk Wienke2, Richard Stillion1, Emma J. McKenzie1, James N. Arnold3,James Harris*1, Eamon McGreal*1, Robert B. Sim3, Clare M. Isacke2 and Siamon Gordon1
1 Sir William Dunn School of Pathology, Oxford, UK2 Breakthrough Breast Cancer Research Centre, The Institute of Cancer Research, London, UK3 Medical Research Council Immunochemistry Unit, University of Oxford, Oxford, UK
Mannose receptor (MR) is the best characterised member of a family of four endocyticmolecules that share a common domain structure; a cysteine-rich (CR) domain, afibronectin-type II (FNII) domain and tandemly arranged C-type lectin-like domains(CTLD, eight in the case of MR). Two distinct lectin activities have been described forMR. The CR domain recognises sulphated carbohydrates while the CTLD mediatebinding to mannose, fucose or N-acetylglucosamine. FNII domains are known to beimportant for collagen binding and this has been studied in the context of two membersof the MR family, Endo180 and the phospholipase A2 receptor. Here, we haveinvestigated whether the broad and effective lectin activity mediated by the CR domainand CTLD of MR is favoured to the detriment of FNII-mediated interaction(s). We showthat MR is able to bind and internalise collagen in a carbohydrate-independent mannerand that MR deficient macrophages have a marked defect in collagen IV and gelatininternalisation. These data have major implications at the molecular level as there arenow three distinct ligand-binding sites described for MR. Furthermore our findingsextend the range of endogenous ligands recognised by MR, a molecule firmly placed atthe interface between homeostasis and immunity.
Introduction
Resolution of inflammation involves the coordinateeffort of cellular and soluble components of the immunesystem in cooperation with fibroblasts and endothelial
cells, leading to the clearance of the initial insult, theelimination of cellular debris and tissue remodelling.Macrophages (MU) are important players in theregulation of this process. During the acute phase of
Correspondence: Dr. Luisa Martinez-Pomares, School ofMolecular Medical Sciences, Floor A, West Block, Queen'sMedical Centre, NG7 2UH, Nottingham, UKFax: +44-115-823-0759e-mail: [email protected]
Received 5/11/05Revised 2/2/06
Accepted 8/3/06
[DOI 10.1002/eji.200535685]
Key words:CD206/208 � Endo180� Extracellular matrix
� Macrophages� Mannose receptor
Abbreviations: AP: alkaline-phosphatase � CR: cysteine-rich �CTLD: C-type lectin-like receptor � FNII: fibronectin type II �MR: mannose receptor
Luisa Martinez-Pomares et al. Eur. J. Immunol. 2006. 36: 1074–10821074
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
* Present address: L. Martinez-Pomares, School of MolecularMedical Sciences, Institute of Infection, Immunity andInflammation, Queen's Medical Centre, NG7 2UH, Notting-ham, UKJ. Harris, Department ofMolecular Genetics andMicrobiology,University of New Mexico, Health Sciences Center, Albu-querque NM87131, MexicoE. McGreal, Department of Child Health, Wales College ofMedicine, Cardiff University, Cardiff CF14 4XN, UK
the inflammatory response MU favour elimination offoreign material through microbial killing and phago-cytosis. During the resolution stage of inflammation MUmediate clearance through phagocytic and enzymaticactivity and influence extracellular matrix productionand angiogenesis, through the production of growthfactors. Furthermore, MU affect the deposition andorganisation of collagen through the secretion ofcollagenase and other neutral proteinases. Under someconditions MU facilitate fibrosis and scarring throughthe release of fibroblast growth factors and promotecollagen deposition through the up-regulation ofarginase activity [1–3]. These properties are particularlyassociated with alternatively activated MU generated inresponse to the Th2 T cell-derived cytokines IL-4 andIL-13 [1]. Another Th2 T cell-derived cytokine, IL-10,can also be produced following MU stimulation [4, 5].This cytokine induces a deactivated anti-inflammatorystate in the Mu and is a key player in the control ofinflammation in vivo [6]
One of the hallmarks of alternatively activated anddeactivated MU is increased expression of the mannose
receptor (MR, CD206) [1, 7]. The MR was originallydescribed as an endocytic receptor for lysosomalenzymes [8] and studies in MR-deficient mice supportthe idea that MR is primarily a homeostatic clearancesystem with an additional role as a pathogen receptor[9–12].
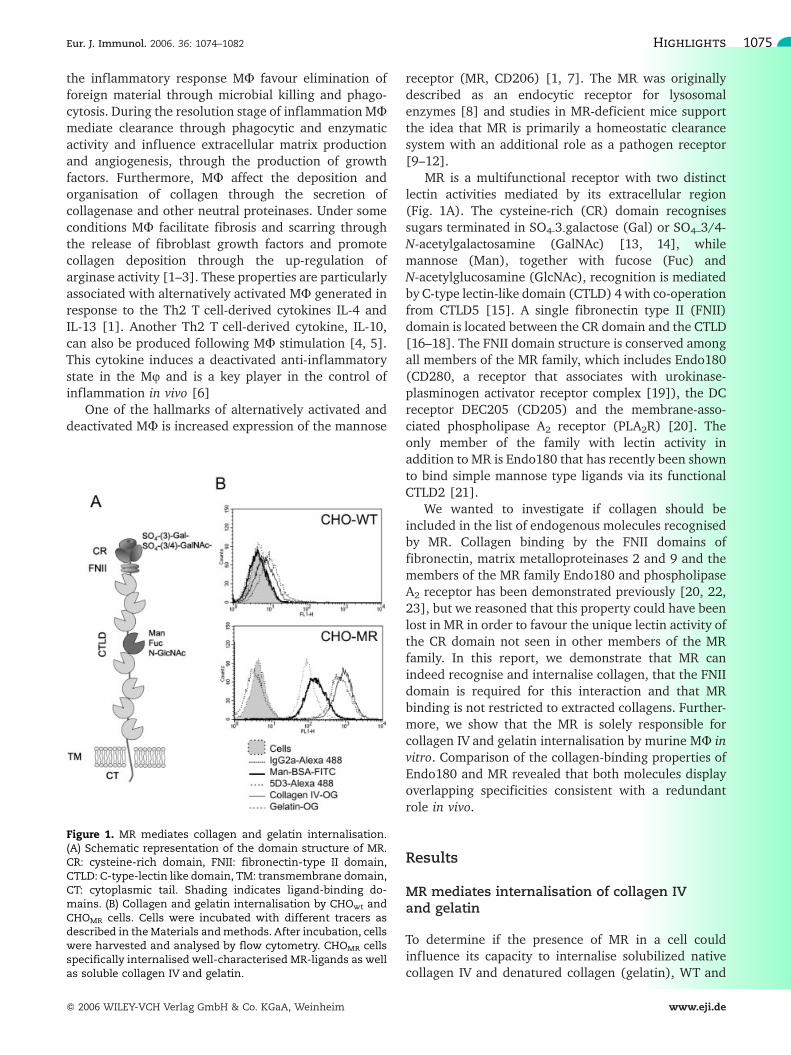
MR is a multifunctional receptor with two distinctlectin activities mediated by its extracellular region(Fig. 1A). The cysteine-rich (CR) domain recognisessugars terminated in SO4-3-galactose (Gal) or SO4–3/4-N-acetylgalactosamine (GalNAc) [13, 14], whilemannose (Man), together with fucose (Fuc) andN-acetylglucosamine (GlcNAc), recognition is mediatedby C-type lectin-like domain (CTLD) 4 with co-operationfrom CTLD5 [15]. A single fibronectin type II (FNII)domain is located between the CR domain and the CTLD[16–18]. The FNII domain structure is conserved amongall members of the MR family, which includes Endo180(CD280, a receptor that associates with urokinase-plasminogen activator receptor complex [19]), the DCreceptor DEC205 (CD205) and the membrane-asso-ciated phospholipase A2 receptor (PLA2R) [20]. Theonly member of the family with lectin activity inaddition to MR is Endo180 that has recently been shownto bind simple mannose type ligands via its functionalCTLD2 [21].
We wanted to investigate if collagen should beincluded in the list of endogenous molecules recognisedby MR. Collagen binding by the FNII domains offibronectin, matrix metalloproteinases 2 and 9 and themembers of the MR family Endo180 and phospholipaseA2 receptor has been demonstrated previously [20, 22,23], but we reasoned that this property could have beenlost in MR in order to favour the unique lectin activity ofthe CR domain not seen in other members of the MRfamily. In this report, we demonstrate that MR canindeed recognise and internalise collagen, that the FNIIdomain is required for this interaction and that MRbinding is not restricted to extracted collagens. Further-more, we show that the MR is solely responsible forcollagen IV and gelatin internalisation by murine MU invitro. Comparison of the collagen-binding properties ofEndo180 and MR revealed that both molecules displayoverlapping specificities consistent with a redundantrole in vivo.
Results
MR mediates internalisation of collagen IVand gelatin
To determine if the presence of MR in a cell couldinfluence its capacity to internalise solubilized nativecollagen IV and denatured collagen (gelatin), WT and
Figure 1. MR mediates collagen and gelatin internalisation.(A) Schematic representation of the domain structure of MR.CR: cysteine-rich domain, FNII: fibronectin-type II domain,CTLD: C-type-lectin like domain, TM: transmembrane domain,CT: cytoplasmic tail. Shading indicates ligand-binding do-mains. (B) Collagen and gelatin internalisation by CHOwt andCHOMR cells. Cells were incubated with different tracers asdescribed in theMaterials andmethods. After incubation, cellswere harvested and analysed by flow cytometry. CHOMR cellsspecifically internalised well-characterised MR-ligands as wellas soluble collagen IV and gelatin.
Eur. J. Immunol. 2006. 36: 1074–1082 Highlights 1075
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

MR expressing cells (CHOwt and CHOMR, respectively[24]) were incubated with OregonGreen-gelatin (gela-tin-OG) and OregonGreen-collagen IV (collagen IV-OG)and processed for flow cytometry as described in theMaterials and methods. Strikingly, collagen IV andgelatin behaved as the MR-specific ligands Man-BSA-FITC and anti-MR mAb MR5D3-Alexa 488 in that theywere endocytosed by CHOMR but not by CHOwt (Fig.1B).Similar results were obtained with NIH-3T3wt and NIH-3T3MR cells (data not shown). The method used toharvest the cells for analysis (trypsin-EDTA) ensuredthat no material associated with MR at the plasmamembrane was being analysed as MR is a trypsinsensitive molecule. Confocal microscopy analysis con-firmed the selectivity of the uptake and demonstratedthat collagen was internalized into intracellular com-partments in the CHOMR cells (data not shown).
Structural requirements for MR-collageninteraction
To define the domain(s) involved in collagen uptake, weprepared the Fc chimeric proteins CR-Fc [25], CR-FNII-CTLD1-Fc, CR-FNII-CTLD1-3-Fc and CTLD4-7-Fc [26](Fig. 2A). All four constructs were readily secreted bytransfected cells and appeared as a single entity by SDS-PAGE (Fig. 2B). Furthermore, gel filtration demon-strated that the vast majority of each protein in thepreparations was, as expected, dimeric (Fig. 2C).CR-FNII-CTLD1-Fc and CR-FNII-CTLD1-3-Fc, togetherwith CR-Fc and CTLD4-7-Fc, were used as inhibitors inendocytosis assays. As expected, Man-BSA uptake couldbe specifically blocked with CRD4-7-Fc and mannan butnot by CR-Fc, CR-FNII-CTLD1-Fc (data not shown), orCR-FNII-CTLD1-3-Fc (Fig. 3A). No inhibition of collageninternalisation could be observed in the presence ofCR-FNII-CTLD1-Fc (data not shown) or CR-FNII-CTLD1-3-Fc, indicating that multimerisation could play animportant role in MR FNII-mediated recognition. Toassess the requirement for protein cross-linking in thecase of FNII domain-mediated recognition we analysedthe binding of the chimeric proteins CR-Fc, CR-FNII-CTLD1-Fc, CR-FNII-CTLD1-3-Fc or CTLD4-7-Fc tocollagen IV-coated plates using pre-complexed or non-complexed proteins. Pre-complexed proteins wereprepared by preincubating the Fc proteins with anti-human Fc Ab. As shown in Fig. 3B, no binding of CR-Fcor CTLD4-7-Fc to collagen IV-coated plates was observedunder any conditions and binding of CR-FNII-CTLD1-Fcand CR-FNII-CTLD1-3-Fc was only detected whenproteins were pre-incubated with an anti-human Fcreagent. Data shown in Fig. 3B also demonstrate thatcollagen binding is calcium independent. The CR-FNII-Fc protein [25] was not included in these studiesbecause questions regarding the conformation of FNII
domain in this construct were raised by its poorproduction and the aggregated material observed afterSDS-PAGE analysis. Our results indicate that thepresence of CTLD1 could facilitate the correct foldingof FNII. No differences between the binding propertiesof CR-FNII-CTLD1-Fc and CR-FNII-CTLD1-3-Fc wereobserved indicating that CTLD2 and CTLD3 do notcontribute substantially to FNII domain-mediatedbinding.
To ascertain if MR could preferentially recognise aparticular type of collagen, we investigated the bindingof cross-linked Fc chimeric proteins to native collagens I,II, III and V and denatured collagen (gelatin). Forcomparison and as a positive control, an Fc chimericprotein containing the CR, FNII and CTLD1–4 domains
Figure 2. Analysis of MR Fc chimeric proteins containing FNII.(A) Schematic representation of Fc proteins used in this study.Fc: Fc region of human IgG1. (B) MR Fc chimeric proteins havethe predicted relative molecular weight. Purified CR-Fc, CR-FNII-CTLD1-Fc CR-FNII-CTLD1-3-Fc and CTLD4-7-Fc wereelectrophoresed under reducing conditions and visualisedusing Coomassie blue. Molecular weight markers are shown.(C) MR Fc chimeric proteins did not formaggregates. Purified Fcchimeric proteins were analysed by gel filtration on aSepharose 12 column and the OD280 of each fraction wasmeasured. In all cases, single peaks were observed elutingaccording to predicted molecular weight; 20 lL of serum wasrun as a standard.
Luisa Martinez-Pomares et al. Eur. J. Immunol. 2006. 36: 1074–10821076
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

of MR family member, Endo180, were included in thesestudies [23]. Data shown in Fig. 4 demonstrate that MRdisplays a broad specificity as it is able to interact with alltypes of collagen tested, including denatured collagen(gelatin) and that there are no major differencesconcerning collagen recognition between MR andEndo180.
MR FNII ligands are broadly distributed in situ
As all the binding studies described earlier wereperformed using purified material extracted frombiological samples, we decided to investigate the bindingproperties of CR-FNII-CTLD1-3-Fc in situ to determine iforganised native collagen embedded in extracellularmatrix would be a suitable ligand for MR. For thispurpose we developed a binding assay based on thatpreviously employed to detect CR domain ligands inlymphoid organs using CR-Fc [25, 27]. Data shown inFig. 5 demonstrate that CR-FNII-CTLD1-3-Fc binds toligands inmouse tissues in situ and that these ligands arewidely distributed. In agreement with previous bindingassays, FNII-mediated binding could only be observedupon cross-linking. In lymphoid organs the presence ofthe FNII domain in the Fc constructs enabled detectionof ligands in addition to those detected by CR-Fc alone;these were distributed as predicted for components ofextracellular matrix. Furthermore, CR-FNII-CTLD1-3-FcFNII also recognised structures in non-lymphoid organssuch as pancreas, adrenal glands (Fig. 5), heart, testisand skin dermis (data not shown) that correlate withrecognition of basement membranes. Interestingly,binding of the CR-Fc construct to lymphoid tissues[25, 27] was substantially enhanced when cross-linked.In particular, the white pulp of spleen, in addition to themarginal zone, was now readily recognised by this probe(Fig. 5). Binding of the CR-FNII-CTLD1-Fc construct wasidentical to that observed with the CR-FNII-CTLD1-3-Fcconstruct (data not shown).
MR is solely responsible for collagen and gelatininternalisation by bone marrow-derived MU
To investigate the contribution of endogenous MR tocollagen internalisation, BM-derived MU from WT andMR–/– animals were assessed for their ability tointernalise collagen IV and gelatin. As shown inFig. 6, MU from MR–/– mice were not only unable tointernalise the MR-specific ligands tested (Man-BSA andanti-MR mAb MR5D3) but also could not internalisecollagen IV or gelatin. The same results were obtainedwhen internalisation assays were performed for 20 min(data not shown) and 1 h, indicating that the differencesobserved were not due to altered kinetics of uptake ordegradation in the MR–/– cells. To demonstrate thatMR–/– MU did not have a general defect in receptor-mediated endocytosis we used the endocytic tracerspecific for scavenger receptors (acetylated low-densitylipoprotein) and determined that its internalisation wasnormal in MR-deficient BM-MU (Fig. 6, insets).
To investigate the relationship between MR andcollagen in vivo we determined how MR expressionrelated to collagen IV localisation in situ. A highly
Figure 3. FNII-mediated recognition of collagen requiresprotein multimerisation. (A) MR Fc protein containing FNIIdo not inhibit collagen internalisation by CHOMR. CHOMR andCHOwt (data not shown) were incubated with different tracers(5 lg/mL) in the presence or absence of Fc chimeric proteins(100 lg/mL) or mannan (2 mg/mL). After incubation, cells wereharvested and analysed by flow cytometry. Man-BSA-FITCinternalisation was effectively inhibited by CTLD4-7-Fc andmannan but not by CR-FNII-CTLD1-3-Fc. Collagen IV inter-nalisationwas not inhibitedunder any of the conditions tested.(B) CR-FNII-CTLD1-Fc and CR-FNII-CTLD1-3-Fc recognise coll-agen IV in vitro. Cross-linked (+) and non-cross-linked (-) CR-Fc,CR-FNII-CTLD1-Fc, CR-FNII-CTLD1-3-Fc, and CTLD4-7-Fc wereincubated in collagen IV-coated wells in the presence (10 mMTris-HCl, pH 7.4, 150 mM NaCl, 10 mM CaCl2) or absence (PBS)of calcium. Specific binding ofmultimerised CR-FNII-CTLD1-Fcor CR-FNII-CTLD1-3-Fc was observed in a calcium-indepen-dent manner.
Eur. J. Immunol. 2006. 36: 1074–1082 Highlights 1077
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

suggestive pattern was obtained in mouse skin (Fig. 7);in this anatomical location MR-positive cells (mostlyMU, E. McKenzie et al, manuscript in preparation) werepresent in dermis and were arranged as rows liningcollagen IV-containing basement membranes. Theseresults are indicative of a contribution of MR to basalmembrane biology.
Discussion
MR contains three functional binding sites
MR is member of a family of proteins comprising threeadditional endocytic receptors, Endo180, PLA2R andDEC-205 [20]. A distinct ligand-binding profile has beendescribed for each of these molecules illustrating howsimilar domain arrangements can be exploited to fulfildiverse physiological roles. While MR and Endo180share the ability to bind similar monosaccharides in vitrothrough selected CTLD (CTLD4 in the case of MR andCTLD2 in the case of Endo180), no natural glycosylatedligands have been characterised for Endo180. MR is theonly family member in which the CR domain containsthe residues required for the formation of the neutralbinding pocket mediating recognition of sulphatedsugars. Therefore, recognition of glycoproteins bearingthese acidic sugars is likely to be mediated exclusively byMR [20, 28].
The ability of MR to bind collagens adds a newdimension to the study of MR biology. The requirement
for extensive cross-linking to visualise this interaction isindicative of a low affinity interaction requiring domainmultimerisation to increase avidity. This property hasbeen observed, to a lesser extent, in the case of CRrecognition of sulphated sugars; we have observed that(i) targeting of CR containing Fc proteins to cells bearingCR ligands in vivo is enhanced by cross-linking [29], (ii)presence of CR domain containing Fc proteins does notblock uptake of SO4-3-Gal byMR transductants (data notshown) and (iii) monomeric soluble MR displays verypoor binding to sulphated sugars [30]. Our observationspredict that monomeric soluble MR would bind poorlyto surfaces rich in sulphated sugars and would be unableto interact with collagen-rich surfaces. Multimerisation,either through interactions with polymeric mannos-ylated sugars [31] or through lack of terminal sialic acidin its N-linked carbohydrates [30] would favour bothinteractions (Fig. 8). On the other hand cell-associatedMR, likely to be clustered at the plasma membrane,would be endowed with the ability to interact with threedistinct ligands. This raises questions regarding theeffect of the extracellular matrix on the sugar-recogni-tion capacities of MR as steric hindrance could hamperrecognition of sulphated sugars by collagen-engagedMR.
MR and self recognition
Our results increase the range of endogenous ligandsidentified for MR. Some of MR endogenous ligands aretargeted by the immune system in the context of
Figure 4. MR recognises collagens I to V and gelatin. Cross-linked MR-derived Fc constructs CR-Fc (data not shown), CR-FNII-CTLD1-Fc andCR-FNII-CTLD1-3-Fc, andCTLD4-7-Fc and Endo180-derived Fc construct E-CR-FNII-CTLD1-4-Fc, were used in in vitrobinding assays as described in the Materials and methods. All proteins containing an FNII domain displayed a similar ability tobind collagens I to Vand gelatin. IgG: human IgG1. No binding of Fc proteins to BSA or fibronectin was observed (data not shown).
Luisa Martinez-Pomares et al. Eur. J. Immunol. 2006. 36: 1074–10821078
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

autoimmune diseases such as thyroglobulin in the caseof thyroditis [26, 32] and myeloperoxidase in the case ofglomerulonephritis [33]. We previously hypothesisedthat inappropriate presentation of these ligands to theacquired immune system could be facilitated throughtheir interaction with MR expressed by subpopulationsof DC [12]. It is intriguing that collagens II and IV canalso be targeted by the acquired system since collagen II,a major component of cartilage, is an importantautoantigen in rheumatoid arthritis patients [34], andcollagen IV contains the antigen recognised by auto-antibodies from Goodpasture syndrome patients [35].
Figure 5. MR FNII ligands can be detected in situ. Tissues fromadultmicewere collected, processed and incubatedwith cross-linkedMR-derivedFcconstructsCR-FcandCR-FNII-CTLD1-3-Fcas described in theMaterials andmethods. Proteins containingFNII bound to structures resembling basementmembrane in allorgans tested. In lymphoid organs, CR-FNII-CTLD1-3-Fc dis-played a dual binding activity with recognition of sulphatedglycans in the marginal zone and B cell follicles of spleen,subcapsular sinus of lymph nodes and medulla of the thymus,in addition to basal membrane components in the red pulp ofspleen, T cell region and medulla of lymph nodes and thymiccortex. Magnification 10x. Areas in insets have been electro-nically magnified.
Figure 6.MR has a non-redundant role in collagen-internalisa-tion by MU. BM-MU from WT and MR-deficient mice weretested for their ability to internalise MR-specific (Man-BSA-FITC and MR5D3-Alexa 488), scavenger receptor-specific (DI-labelled acetylated LDL) tracers and collagen IV-OG andgelatin-OG as described in the Materials and methods. Alltracers were used at 5 lg/mL and cells were collected at 1 h.
Figure 7.MR+ cells in dermis are localised alongside collagen IVfibres. Mouse ears were collected, processed and labelled forMR and collagen IV using MR5D3 (red) and anti-collagen IVantibody (green) as described in the Materials and methods.Top panels show labelling of a cross-section, bottom panelsshow labelling of transverse section. Insets show control singlelabelling. Areas in dashed box are shown at higher magnifica-tion.
Eur. J. Immunol. 2006. 36: 1074–1082 Highlights 1079
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

The involvement of MR to Ag presentation is still unclearsince no major contribution of MR to immunity againstpathogens has been evident in experimental models ofinfection but rather a non-redundant role in homeostaticclearance of lysosomal enzymes has been demonstrated[9–11]. We have identified a novel MR-positive DCpopulation in selected lymphoid organs that can bindMR ligands in vivo (E. McKenzie et al. manuscript inpreparation). These cells are controlled by innatestimulation, leading us to propose that MR-mediatedantigen presentation is favoured upon microbial infec-tion. We hypothesise that in this environment enhancedpresentation of endogenous ligands will also take placethat could trigger activation of self-reactive T cells thatescape central and peripheral mechanisms of tolerance.
MU and collagen
Even though MU produce a wide range of moleculesrelevant to tissue remodelling, the existence of com-pensatory mechanisms is illustrated by the normal foetaldevelopment and wound healing observed in mice
lacking the transcription factor PU.1 which are deficientinMU [36]. With this inmind it is not surprising that MRdeficiency does not seem to have a major effect onorganogenesis under steady-state conditions. Indeed,our preliminary results indicate that the distribution ofligands for the CR and FNII domains and of several MUpopulations is not altered in tissues from MR-/- animals(data not shown). Endo180 expressed by fibroblasts,among others, could compensate for lack of MR.Nevertheless, we have observed that MR plays a non-redundant role in collagen internalisation by MU. Basedon this observation and together with the closerelationship between MR+ cells and collagen IVobserved in mouse skin, we predict that under MU-dominated pathologies such as granulomata, lack of MRwould have a substantial effect.
In summary, we have demonstrated that murine MRrecognises collagens, that this function is mediated byFNII domain and requires protein cross-linking. Ourresults are in agreement with a recent report by Napperet al. [37] demonstrating collagen recognition by theMRFNII domain. We have also shown that MR is responsiblefor collagen internalisation by MU and that there is aclose association between collagen fibres and MR+ cellsin skin. Our studies extend the list of endogenous ligandsrecognised by MR and raise further questions regardingMR involvement in induction of autoimmunity andtissue homeostasis.
Materials and methods
Animals
WTBALB/c andWTandMR–/– C57BL/6mice (kindly providedby Dr. M. Nussenzweig, Rockefeller University) were main-tained under specific pathogen-free conditions and used at8–12 weeks.
Cells
Parental and MR expressing NIH 3T3 and CHO cells (NIH-3T3wt, CHOwt, NIH-3T3MR and CHOMR) have been describedpreviously [24]. Preparation of BM-MU was performed asdescribed [4].
Fc chimeric proteins
cDNA encoding CR-FNII-CTLD-1-Fc and CR-FNII-CTLD1-3-Fcwere amplified using HF-2 polymerase (Clontech, CA, USA)from a plasmid containing the full length mannose receptorcDNA [24] using reverse primer 50-GGAGAT CTA CTT ACC TGTCACATGCTTGCTGAGGGAATGATAAATG(CR-FNII-CTLD-1-Fc)and 50-GGAGAT CTACTTACC TGT TGG ACATTT GGG TTC AGGAGT TGT TGT GG (CR-FNII-CTLD1-3-Fc) and a commonforward primer 50-GGG ATA TCG ACC TTG GAC TGA GCA AAGGGG CAA CCT GG. EcoRV/BglII digested PCR products were
Figure 8. Correlation between state of oligomerisation andbinding properties in MR. Schematic representation of thebinding properties proposed for different forms of MR detectedin vitro and in vivo: (a) monomeric soluble MR, (b) oligomericsoluble MR and (c) cell associated MR. Unlike most C-type-likelectins, tandemly arranged CTLD facilitate binding of mono-meric MR to mannosylated carbohydrates [30]. On the otherhand, efficient recognition of sulphated glycans and collagens(through the CR and FNII domains, respectively) requiresoligomerisation. The interaction of sMR with appropriatemultivalent mannosylated glycoconjugates would lead toenhanced recognition of sulphated ligands [31] (and probablycollagens) as avidity would increase through the display ofseveral binding sites in close proximity. These sMR-ligandcomplexes could bind to cell-associated ligands for the CRdomain [25, 29] or exposed collagens. Cell associated MR hasthe capacity to interact with three distinct types of ligands.
Luisa Martinez-Pomares et al. Eur. J. Immunol. 2006. 36: 1074–10821080
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

introduced in-frame into the EcoRV and BamHI restriction sitesof the expression vector pIg generating vectors encodingMR-Fc chimeric proteins under the control of the CMVpromoter. CR-Fc, CRD4-7-Fc and E-CR-FNII-CTLD1-4-Fc havebeen previously described [23, 25, 26].
Plasmids encoding the Fc-chimeric proteins were intro-duced into the 293T cell line by transient transfection withGene-Juice reagent (Novagen) following manufacturer'sguidelines. After transfection cell media were replaced withmedia containing 4% IgG-depleted foetal bovine serum andmediawere conditioned for 7 days. After this time, media wereharvested and Fc-chimeric proteins were recovered by affinitypurification on Protein A-Sepharose (Amersham Biosciences)and elution with 0.1 M glycine, pH2.9. After neutralisationwith 0.1 volumes of 1 M Tris, pH 9.5, proteins were dialysedagainst PBS. Of each reduced chimeric Fc protein, 4-10 lg wasconcentrated using the Strataclean method and run on 4–15%Bris-Tris gel (Invitrogen), alongside 7 lL of MultiMark proteinladder (Invitrogen).
Gel filtration analysis
Gel filtration was performed on a Superose 12 column(Pharmacia) using 10 mM Tris, 140 mM NaCl, 10 mM CaCl2,pH7.4 as running buffer; 20 lL of serumwas run as a standard.
Solid phase collagen-binding assay
The 96-well high-binding plates (Costar) were coated withappropriate substratum diluted at 0.1 mg/mL, (100 lL/well)and incubated overnight at 37�C. Plates were washed threetimes with 150 lL PBS/0.3% BSA and blocked with 0.3% BSAin PBS for 1 h at room temperature (RT). Fc chimeric proteins(diluted at 2–10 lg/mL) were preincubated or not preincu-bated with anti-human Fc-alkaline-phosphatase (AP) conju-gate (1:100–1:200, Sigma) for 1 h at RT in PBS/0.3% BSA.These were added to the plate and incubated for 1 h at RT.After three washes in PBS/0.3% BSA, plates were incubatedwith anti-human Fc-AP (1:500–1:1000) for 1 h at RT in PBS/0.3% BSA. After washing three times with PBS/0.3% BSA,wells were washed twice with AP substrate buffer (100 mMTris-HCl, 100 mM NaCl, 1 mM MgCl2, pH 9.5) and 50 lL ofsubstrate solution were added to each well (1 mg/mLp-nitrophenyl phosphate in substrate buffer) and incubatedfor 10–30 min at RT. Reactions were stopped by adding 50 lLof stop solution (100 mM EDTA). Absorbance was read at405 nm. In some instances 10 mM Tris-HCl, pH 7.4, 150 mMNaCl, 10 mM CaCl2 was used as buffer in the assay.
Internalisation assays
CHOwt, CHOMR, and BM-MU from WT C57BL/6 and MR–/–
mice were washed in serum-free medium and maintained inthis medium for 1 h at 37oC. Different tracers (diluted at 5 lg/mL in serum-free media) were then added. The differenttracers used were: Man-BSA-FITC (EY laboratories), CollagenIV-Oregon Green (OG) (Molecular Probes), gelatin-OG(Molecular Probes), anti-MR mAb MR5D3 and isotype controlIgG2a both prepared in house and labelled with Alexa 488(Molecular Probes) following the manufacturer's instructions.
In some experiments CR-Fc, CR-FNII-CTLD-1-Fc, CR-FNII-CTLD1-3-Fc or CTLD 4-7-Fc diluted at 100 lg/mL or mannan(2 mg/mL) were present during the incubation. After incuba-tion cells were washed in PBS, harvested using trypsin/EDTA(CHO cells) or lidocaine/EDTA (BM-MU) and fixed in 2%formaldehyde solution in PBS. Uptake was analysed using a BDFACScalibur and CellQuest software.
In situ detection of FNII ligands
Mouse tissues were collected, embedded in OCT compoundand frozen in dry ice-cooled isopentane. The 5-lm sectionswere cut, placed on charged slides and kept frozen until use. Todetect ligands for CR and FNII domains in situ, sections werefixed in 2% paraformaldehyde (10 min, 4oC), permeabilisedusing 1% Triton X-100 in PBS (1 h, RT) and blocked with 5%normal goat serum (NGS) in PBS (30 min, RT). Preparationswere incubated with CR-Fc, CR-FNII-CTLD-1-Fc, CR-FNII-CTLD1-3-Fc pre-complexed for 1 h using goat-anti-human APconjugate (Chemicon), diluted 1:100 in PBS (overnight, 4oC)in the presence of 5% NGS. Protein binding was detected usinggoat-anti-human AP conjugate diluted 1:100 in 5% NGS. Theslides were developed using the BCIP/NBT alkaline phospha-tase substrate kit IV (Vector Laboratories).
Detection of MR and collagen IV in tissue sections
Mouse ears were collected, embedded in OCT compound andfrozen in dry ice-cooled isopentane. The 5-lm sections werecut and kept frozen until use. Sections were thawed at roomtemperature, fixed in 2% paraformaldehyde (10 min, 4oC),permeabilised using 0.1% Triton X100 in PBS (1 h), blockedwith 5% NGS in PBS (30 min, RT) followed by blocking ofendogenous biotin using a avidin/biotin kit (Vector Labora-tories). Sections were incubated with rabbit anti-mouseCollagen IV (Immunologicals Direct) in 5% NGS in PBS(1 h) followed by Alexa488-conjugated goat anti-rabbit IgG(Molecular Probes) in PBS, to detect collagen, and then withbiotinylated MR5D3 (10 lg/mL) in 5% NGS in PBS (1 h) andstreptavidin Cy5 (Jackson Immunoresearch Laboratories) inPBS (30 min). Sections were counterstained with DAPI(Sigma) at 400 ng/mL prepared in H2O for 5 min, mountedusing DAKO fluorescent mounting media (Dakocytomation)and analysed using a Zeiss Axioplan 2e microscope fitted witha 16-bit 1024 � 1024 pixels CCD camera. Images wereprocessed using Metamorph software.
Acknowledgements: Funding sources: Medical Re-search Council, The Edward P. Abraham Research Fundand Breakthrough Breast Cancer.
References
1 Gordon, S.,Alternative activation ofmacrophages.Nat. Rev. Immunol. 2003.3: 23–35.
2 Munder, M., Eichmann, K., Moran, J. M., Centeno, F., Soler, G. andModolell, M., Th1/Th2-regulated expression of arginase isoforms in murinemacrophages and dendritic cells. J. Immunol. 1999. 163: 3771–3777.
Eur. J. Immunol. 2006. 36: 1074–1082 Highlights 1081
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de

3 Munder, M., Mallo, M., Eichmann, K. and Modolell, M., Murinemacrophages secrete interferon gamma upon combined stimulation withinterleukin (IL)-12 and IL-18: A novel pathway of autocrine macrophageactivation. J. Exp. Med. 1998. 187: 2103–2108.
4 Peiser, L., De Winther, M. P., Makepeace, K., Hollinshead, M., Coull, P.,Plested, J., Kodama, T. et al., The class A macrophage scavenger receptor isa major pattern recognition receptor for Neisseria meningitidis which isindependent of lipopolysaccharide and not required for secretory responses.Infect. Immun. 2002. 70: 5346–5354.
5 Sutterwala, F. S., Noel, G. J., Salgame, P. and Mosser, D. M., Reversal ofproinflammatory responses by ligating the macrophage Fcgamma receptortype I. J. Exp. Med. 1998. 188: 217–222.
6 Pestka, S., Krause, C. D., Sarkar, D., Walter, M. R., Shi, Y. and Fisher, P.B., Interleukin-10 and related cytokines and receptors. Annu. Rev. Immunol.2004. 22: 929–979.
7 Stein, M., Keshav, S., Harris, N. and Gordon, S., Interleukin 4 potentlyenhances murine macrophage mannose receptor activity: a marker ofalternative immunologic macrophage activation. J. Exp. Med. 1992. 176:287–292.
8 Pontow, S. E., Kery, V. and Stahl, P. D., Mannose receptor. Int. Rev. Cytol.1992. 137B: 221–244.
9 Lee, S. J., Evers, S., Roeder, D., Parlow, A. F., Risteli, J., Risteli, L., Lee, Y.C. et al., Mannose receptor-mediated regulation of serum glycoproteinhomeostasis. Science 2002. 295: 1898–1901.
10 Lee, S. J., Zheng, N. Y., Clavijo, M. and Nussenzweig, M. C., Normal hostdefense during systemic candidiasis in mannose receptor-deficient mice.Infect. Immun. 2003. 71: 437–445.
11 Swain, S. D., Lee, S. J., Nussenzweig, M. C. and Harmsen, A. G., Absenceof the macrophagemannose receptor inmice does not increase susceptibilityto Pneumocystis carinii infection in vivo. Infect. Immun. 2003. 71: 6213–6221.
12 Taylor, P. R., Gordon, S. and Martinez-Pomares, L., The mannosereceptor: linking homeostasis and immunity through sugar recognition.Trends Immunol. 2005. 26: 104–110.
13 Fiete, D. J., Beranek, M. C. and Baenziger, J. U., A cysteine-rich domain ofthe “mannose” receptor mediates GalNAc-4-SO4 binding. Proc. Natl. Acad.Sci. USA 1998. 95: 2089–2093.
14 Leteux, C., Chai, W., Loveless, R. W., Yuen, C. T., Uhlin-Hansen, L.,Combarnous, Y., Jankovic, M. et al., The cysteine-rich domain of themacrophage mannose receptor is a multispecific lectin that recognizeschondroitin sulfates A and B and sulfated oligosaccharides of blood groupLewis(a) and Lewis(x) types in addition to the sulfated N-glycans of lutropin.J. Exp. Med. 2000. 191: 1117–1126.
15 Taylor, M. E., Bezouska, K. and Drickamer, K., Contribution to ligandbinding by multiple carbohydrate-recognition domains in the macrophagemannose receptor. J. Biol. Chem. 1992. 267: 1719–1726.
16 Ezekowitz, R. A., Sastry, K., Bailly, P. and Warner, A., Molecularcharacterization of the human macrophage mannose receptor: demonstra-tion of multiple carbohydrate recognition-like domains and phagocytosis ofyeasts in Cos-1 cells. J. Exp. Med. 1990. 172: 1785–1794.
17 Harris, N., Super, M., Rits, M., Chang, G. and Ezekowitz, R. A.,Characterization of the murine macrophage mannose receptor: demonstra-tion that the downregulation of receptor expression mediated by interferon-gamma occurs at the level of transcription. Blood 1992. 80: 2363–2373.
18 Taylor, M. E., Conary, J. T., Lennartz, M. R., Stahl, P. D. and Drickamer,K., Primary structure of the mannose receptor contains multiple motifsresembling carbohydrate-recognition domains. J. Biol. Chem. 1990. 265:12156–12162.
19 Behrendt, N., Jensen, O. N., Engelholm, L. H., Mortz, E., Mann, M. andDano, K., A urokinase receptor-associated protein with specific collagenbinding properties. J. Biol. Chem. 2000. 275: 1993–2002.
20 East, L. and Isacke, C. M., The mannose receptor family. Biochim. Biophys.Acta 2002. 1572: 364–386.
21 East, L., Rushton, S., Taylor, M. E. and Isacke, C. M., Characterization ofsugar binding by the mannose receptor family member, Endo180. J. Biol.Chem. 2002. 277: 50469–50475.
22 Kjoller, L., Engelholm, L. H., Hoyer-Hansen, M., Dano, K., Bugge, T. H.and Behrendt, N., uPARAP/endo180 directs lysosomal delivery anddegradation of collagen IV. Exp. Cell Res. 2004. 293: 106–116.
23 Wienke, D., MacFadyen, J. R. and Isacke, C. M., Identification andcharacterization of the endocytic transmembrane glycoprotein Endo180 as anovel collagen receptor. Mol. Biol. Cell 2003. 14: 3592–3604.
24 Martinez-Pomares, L., Reid, D. M., Brown, G. D., Taylor, P. R., Stillion, R.J., Linehan, S. A., Zamze, S. et al., Analysis of mannose receptor regulationby IL-4, IL-10, and proteolytic processing using novel monoclonalantibodies. J. Leukoc. Biol. 2003. 73: 604–613.
25 Martinez-Pomares, L., Kosco-Vilbois, M., Darley, E., Tree, P., Herren, S.,Bonnefoy, J. Y. andGordon, S., Fc chimeric protein containing the cysteine-rich domain of the murine mannose receptor binds to macrophages fromsplenic marginal zone and lymph node subcapsular sinus and to germinalcenters. J. Exp. Med. 1996. 184: 1927–1937.
26 Linehan, S. A., Martinez-Pomares, L., da Silva, R. P. and Gordon, S.,Endogenous ligands of carbohydrate recognition domains of the mannosereceptor in murine macrophages, endothelial cells and secretory cells;potential relevance to inflammation and immunity. Eur. J. Immunol. 2001.31: 1857–1866.
27 Linehan, S. A., Martinez-Pomares, L., Stahl, P. D. and Gordon, S.,Mannose receptor and its putative ligands in normal murine lymphoid andnonlymphoid organs: In situ expression of mannose receptor by selectedmacrophages, endothelial cells, perivascular microglia, and mesangial cells,but not dendritic cells. J. Exp. Med. 1999. 189: 1961–1972.
28 Liu, Y., Chirino, A. J., Misulovin, Z., Leteux, C., Feizi, T., Nussenzweig, M.C. and Bjorkman, P. J., Crystal structure of the cysteine-rich domain ofmannose receptor complexed with a sulfated carbohydrate ligand. J. Exp.Med. 2000. 191: 1105–1116.
29 Taylor, P. R., Zamze, S., Stillion, R. J., Wong, S. Y., Gordon, S. andMartinez-Pomares, L., Development of a specific system for targetingprotein to metallophilic macrophages. Proc. Natl. Acad. Sci. USA 2004. 101:1963–1968.
30 Su, Y., Bakker, T., Harris, J., Tsang, C., Brown, G. D., Wormald, M. R.,Gordon, S. et al., Glycosylation influences the lectin activities of themacrophage mannose receptor. J. Biol. Chem. 2005. 280: 32811–32820.
31 Zamze, S., Martinez-Pomares, L., Jones, H., Taylor, P. R., Stillion, R. J.,Gordon, S. and Wong, S. Y., Recognition of bacterial capsular polysacchar-ides and lipopolysaccharides by the macrophage mannose receptor. J. Biol.Chem. 2002. 277: 41613–41623.
32 Chazenbalk, G. D., Pichurin, P. N., Guo, J., Rapoport, B. and McLachlan,S. M., Interactions between the mannose receptor and thyroid autoantigens.Clin. Exp. Immunol. 2005. 139: 216–224.
33 Xiao, H., Heeringa, P., Hu, P., Liu, Z., Zhao, M., Aratani, Y., Maeda, N. etal., Antineutrophil cytoplasmic autoantibodies specific for myeloperoxidasecause glomerulonephritis and vasculitis in mice. J. Clin. Invest. 2002. 110:955–963.
34 Corrigall, V. M. and Panayi, G. S., Autoantigens and immune pathways inrheumatoid arthritis. Crit. Rev. Immunol. 2002. 22: 281–293.
35 Netzer, K. O., Leinonen, A., Boutaud, A., Borza, D. B., Todd, P., Gunwar,S., Langeveld, J. P. and Hudson, B. G., The goodpasture autoantigen.Mapping the major conformational epitope(s) of alpha3(IV) collagen toresidues 17–31 and 127–141 of the NC1 domain. J. Biol. Chem. 1999. 274:11267–11274.
36 Martin, P., D'Souza, D., Martin, J., Grose, R., Cooper, L., Maki, R. andMcKercher, S. R.,Wound healing in the PU.1 null mouse–tissue repair is notdependent on inflammatory cells. Curr. Biol. 2003. 13: 1122–1128.
37 Napper, C. E., Drickamer, K. and Taylor, M. E., Collagen binding by themannose receptor mediated through the fibronectin type II domain.Biochem. J. 2006. DOI: 10.1042/BJ20052027.
Luisa Martinez-Pomares et al. Eur. J. Immunol. 2006. 36: 1074–10821082
f 2006 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji.de
Copyright © 2022 FDOKUMEN




















