
Galactomannan hydrolysis and mannose metabolism in Cellvibrio mixtus
10
Galactomannan hydrolysis and mannose metabolism in Cellvibrio mixtus Maria S.J. Centeno 1 , Catarina I.P.D. Guerreiro 1 , Fernando M.V. Dias 1 , Carl Morland 2 , Louise E. Tailford 2 , Arun Goyal 1 , Jos ´ e A.M. Prates 1 , Lu´ ıs M.A. Ferreira 1 , Rui M.H. Caldeira 1 , Emmanuel F. Mongodin 3 , Karen E. Nelson 3 , Harry J. Gilbert 2 & Carlos M.G.A. Fontes 1 1 CIISA – Faculdade de Medicina Veterin ´ aria, P ´ olo Universit ´ ario do Alto da Ajuda, Lisboa, Portugal; 2 Institute for Cell and Molecular Biosciences, University of Newcastle upon Tyne, The Medical School, Newcastle upon Tyne, UK; and 3 The Institute for Genomic Research, Rockville, MD, USA Correspondence: Carlos M.G.A Fontes, CIISA – Faculdade de Medicina Veterin ´ aria, P´ olo Universit ´ ario do Alto da Ajuda, Avenida da Universidade T ´ ecnica, 1300-477 Lisboa, Portugal. Tel.: 1351 213652876; fax: 1351 213652889; e-mail: [email protected] Present address: Arun Goyal, Department of Biotechnology, Indian Institute of Technology Guwahati, North Guwahati, Guwahati 781 039, Assam, India. Received 29 March 2006; revised 1 June 2006; accepted 1 June 2006. First published online June 2006. doi:10.1111/j.1574-6968.2006.00342.x Editor: Marco Moracci Keywords a-galactosidase; epimerase; galactomannan; glycoside hydrolase; Cellvibrio . Abstract Galactomannan hydrolysis results from the concerted action of microbial endo- mannanases, manosidases and a-galactosidases and is a mechanism of intrinsic biological importance. Here we report the identification of a gene cluster in the aerobic soil bacterium Cellvibrio mixtus encoding enzymes involved in the degradation of this polymeric substrate. The family 27 a-galactosidase, termed CmAga27A, preferentially hydrolyse galactose containing polysaccharides. In addition, we have characterized an enzyme with epimerase activity, which might be responsible for the conversion of mannose into glucose. The role of the identified enzymes in the hydrolysis of galactomannan by aerobic bacteria is discussed. Introduction The plant cell wall represents the most abundant reservoir of renewable energy and carbon within the biosphere (Gilbert et al., 2002). Mannan consists of a backbone of b-1,4-linked mannose residues while in glucomannans the backbone comprises both glucose and mannose residues randomly distributed (Brett & Waldren, 1996). Galactose monomers may decorate the mannose residues of both hemicelluloses through a-1,6 linkages and therefore these polysaccharides are usually referred to as galactomannans and galactoglucoman- nans, respectively. Galactomannans are also storage carbohy- drates in some plant seeds such as carob (Brett & Waldren, 1996). The hydrolysis of mannose containing polysaccharides into its monomeric components requires the action of endo- b-1,4-mannanases, exo-b-1,4-mannosidases and b-1,4-gluco- sidases (McCleary, 1988; Puls & Schuseil, 1996). In the sequence-based classification of glycoside hydro- lases (GH) a-galactosidases are located in families 4, 27 and 36 (Coutinho & Henrissat, 1999). Family 4 is exclusively prokaryotic, family 36 contains both bacterial and eukaryo- tic enzymes, while family 27 comprises predominantly eukaryotic GHs. While most eukaryotic a-galactosidases are in family 27, the majority of prokaryotic enzymes have been classified into families 4 or 36. Ultimately, the substrate specificity of these enzymes is not restricted to the de- branching activity of hemicelluloses, as a-galactosidases have been shown to be crucial in other biological systems. For example, removal of galactose from short chain galacto- oligosaccharides, such as melibiose, raffinose and stachyose that are carbohydrate reserves in leguminous plants, is catalysed by plant and microbial a-galactosidases usually located in families 4 and 36 (Fridjonsson et al., 1999). In eukaryotes, lysosomal a-galactosidases are involved in the catabolism of large macromolecules, such as glycoproteins and glycolipids, and these enzymes belong to family 27 (Garman & Garboczi, 2004). Interestingly, most a-galacto- sidases acting on branched hemicelluloses were shown to FEMS Microbiol Lett 261 (2006) 123–132 c 2006 Federation of European Microbiological Societies Published by Blackwell Publishing Ltd. All rights reserved
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Galactomannan hydrolysis and mannose metabolism in Cellvibrio mixtus

Galactomannan hydrolysis andmannosemetabolism inCellvibriomixtusMaria S.J. Centeno1, Catarina I.P.D. Guerreiro1, Fernando M.V. Dias1, Carl Morland2, Louise E. Tailford2,Arun Goyal1, Jose A.M. Prates1, Luıs M.A. Ferreira1, Rui M.H. Caldeira1, Emmanuel F. Mongodin3,Karen E. Nelson3, Harry J. Gilbert2 & Carlos M.G.A. Fontes1
1CIISA – Faculdade de Medicina Veterinaria, Polo Universitario do Alto da Ajuda, Lisboa, Portugal; 2Institute for Cell and Molecular Biosciences,
University of Newcastle upon Tyne, The Medical School, Newcastle upon Tyne, UK; and 3The Institute for Genomic Research, Rockville, MD, USA
Correspondence: Carlos M.G.A Fontes,
CIISA – Faculdade de Medicina Veterinaria,
Polo Universitario do Alto da Ajuda, Avenida
da Universidade Tecnica, 1300-477 Lisboa,
Portugal. Tel.: 1351 213652876; fax: 1351
213652889; e-mail: [email protected]
Present address: Arun Goyal, Department of
Biotechnology, Indian Institute of Technology
Guwahati, North Guwahati, Guwahati 781
039, Assam, India.
Received 29 March 2006; revised 1 June 2006;
accepted 1 June 2006.
First published online June 2006.
doi:10.1111/j.1574-6968.2006.00342.x
Editor: Marco Moracci
Keywords
a-galactosidase; epimerase; galactomannan;
glycoside hydrolase; Cellvibrio .
Abstract
Galactomannan hydrolysis results from the concerted action of microbial endo-
mannanases, manosidases and a-galactosidases and is a mechanism of intrinsic
biological importance. Here we report the identification of a gene cluster in the
aerobic soil bacterium Cellvibrio mixtus encoding enzymes involved in the
degradation of this polymeric substrate. The family 27 a-galactosidase, termed
CmAga27A, preferentially hydrolyse galactose containing polysaccharides. In
addition, we have characterized an enzyme with epimerase activity, which might
be responsible for the conversion of mannose into glucose. The role of the
identified enzymes in the hydrolysis of galactomannan by aerobic bacteria is
discussed.
Introduction
The plant cell wall represents the most abundant reservoir of
renewable energy and carbon within the biosphere (Gilbert
et al., 2002). Mannan consists of a backbone of b-1,4-linked
mannose residues while in glucomannans the backbone
comprises both glucose and mannose residues randomly
distributed (Brett & Waldren, 1996). Galactose monomers
may decorate the mannose residues of both hemicelluloses
through a-1,6 linkages and therefore these polysaccharides are
usually referred to as galactomannans and galactoglucoman-
nans, respectively. Galactomannans are also storage carbohy-
drates in some plant seeds such as carob (Brett & Waldren,
1996). The hydrolysis of mannose containing polysaccharides
into its monomeric components requires the action of endo-
b-1,4-mannanases, exo-b-1,4-mannosidases and b-1,4-gluco-
sidases (McCleary, 1988; Puls & Schuseil, 1996).
In the sequence-based classification of glycoside hydro-
lases (GH) a-galactosidases are located in families 4, 27 and
36 (Coutinho & Henrissat, 1999). Family 4 is exclusively
prokaryotic, family 36 contains both bacterial and eukaryo-
tic enzymes, while family 27 comprises predominantly
eukaryotic GHs. While most eukaryotic a-galactosidases
are in family 27, the majority of prokaryotic enzymes have
been classified into families 4 or 36. Ultimately, the substrate
specificity of these enzymes is not restricted to the de-
branching activity of hemicelluloses, as a-galactosidases
have been shown to be crucial in other biological systems.
For example, removal of galactose from short chain galacto-
oligosaccharides, such as melibiose, raffinose and stachyose
that are carbohydrate reserves in leguminous plants, is
catalysed by plant and microbial a-galactosidases usually
located in families 4 and 36 (Fridjonsson et al., 1999). In
eukaryotes, lysosomal a-galactosidases are involved in the
catabolism of large macromolecules, such as glycoproteins
and glycolipids, and these enzymes belong to family 27
(Garman & Garboczi, 2004). Interestingly, most a-galacto-
sidases acting on branched hemicelluloses were shown to
FEMS Microbiol Lett 261 (2006) 123–132 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

have broad substrate specificities, also hydrolysing low
molecular weight substrates (Dey et al., 1993). There is,
however, a paucity of information relating to the molecular
determinants of substrate specificity in enzymes acting on
the removal of side chains from hetero-polymeric mannans.
The metabolism of the hydrolysis products of galactose
containing mannans and glucomannans, by aerobic soil
bacteria such as Cellvibrio mixtus, is poorly understood.
This is surprising, considering the detailed information that
is available on the hydrolysis of plant cell wall polysacchar-
ides. It is generally established that mannose residues are
phosphorylated by a transmembrane permease to mannose
6-phosphate and then converted to fructose 6-phosphate by
an intracellular phosphomannose isomerase (Binet et al.,
1998). In addition, galactose might be transformed into
galactose 1-phosphate by an intracellular galactokinase and
then converted into glucose 1-phosphate through the Leloir
pathway (Holden et al., 2003). These possibilities remain,
however, to be confirmed in Cellvibrio.
Here we report the cloning and the biochemical charac-
terization of two enzymes involved in the degradation and
metabolism of galactomannan, from the prolific plant cell
wall degrader Cellvibrio mixtus. It is shown that the family
27 a-galactosidase of Cellvibrio mixtus is a polymer specific
enzyme with very limited activity on galacto-oligosacchar-
ides of the raffinose series. In addition, CmEpiA is respon-
sible for the interconversion of mannose into glucose by
Cellvibrio mixtus.
Materials and methods
Bacterial strains, plasmids and cultureconditions
Escherichia coli strains used in this study were BL21(DE3),
Origami (Novagen) and XL1-Blue (Stratagene). The bacter-
iophages and plasmids used in this work were lZAPII
(Stratagene), pBluescript SK� (Stratagene), pGEM-T (Pro-
mega) and pET21a (Novagen) and pET28a (Novagen).
Cellvibrio mixtus (NCIMB 8633) was cultured aerobically at
20 1C in Dubos mineral salts medium or on Dubos agar
plates overlaid with filter paper (Millward-Sadler et al.,
1995). Culture conditions were maintained as described
previously (Millward-Sadler et al., 1995). The recombinant
proteins, encoded in pET21a and pET28a derivatives, were
expressed in E. coli BL21 and Origami strains, as described
below.
General recombinant DNA procedures
Transformation of E. coli, digestion of DNA with restriction
endonucleases, ligation of DNA with T4 DNA ligase, agarose
gel electrophoresis, Southern hybridization, slot blot and
plaque hybridizations were carried out as described pre-
viously (Berns & Thomas, 1965; Sambrook et al., 1989). The
genomic library was constructed in lZAPII using the
approach described by Clarke et al. (1991). The library was
plated on NZYM top agar at a density of three plaques per
cm2 and screened for a-galactosidase activity using the
methylumbelliferyl a-D-galactosidase overlay technique.
DNA hybridizations were performed using the fluorescein
system from Amersham according to the manufacturer’s
protocol. Two inverse PCR amplifications were performed
to clone the full-length sequence of the genes located
upstream from aga27 (encodes CmAga27A). Briefly, for the
first inverse PCR experiment, Cellvibrio mixtus genomic
DNA was digested with the enzyme EcoRI and resulting
DNA fragments ranging from 4.5 to 5.5 kb were eluted from
agarose gels and religated. In the second experiment, the
enzyme used was PstI and the fragments ranging from 1.5
and 2.5 kb were ligated. The primer pairs used in the first
and second amplifications were as follows: first amplifica-
tion (EcoRI), 50-GCAGGTCCATAAACATACGC-30 and 50-
GGTTGGCCAATATTTTGCGC-30; second amplification
(PstI), 50-CAATGACAGATGAAGGAAGC-30 and 50-CTTAG
CGCTATTGCTGATTG-30. PCRs were performed using 1 U
of the thermostable DNA polymerase pFU turbo (Strata-
gene) and following the manufacturer’s instructions. PCR
products were cloned into pGEM-T (Promega) and the
resulting plasmids were named pZC3 and pZC4, respec-
tively. The nucleotide sequence of DNA was determined
with an ABI Prism Ready Reaction DyeDeoxy terminator
cycle sequencing kit and an Applied Biosystems 377A
sequencing system.
Expression and purification of recombinantproteins
To express CmAga27A, CmEpiA and CmUnkA in E. coli, the
DNA encoding the mature enzymes were amplified by PCR
from Cellvibrio mixtus genomic DNA using the thermo-
stable DNA polymerase pFU Turbo (Stratagene). The pri-
mers used for the amplifications incorporated designed
restriction sites (Table 1). The PCR products were cloned
into pGEM-T and sequenced to ensure that no mutations
had occurred during PCR. The recombinant derivatives of
pGEM-T containing the Cellvibrio mixtus genes were
digested with the required restriction enzymes (Table 1)
and the excised DNA fragments were cloned into the
similarly digested vector. The three recombinant proteins
contain N- or C-terminal His6-tag. The resulting plasmids
encoding recombinant CmAga27A, CmEpiA and CmUnkA,
were named pZC5, pZC6 and pZC7, respectively.
Escherichia coli BL21 harbouring pET derivatives encod-
ing CmEpiA and CmUnkA was cultured in LB contain-
ing 100 mg mL�1 ampicillin at 37 1C to mid-exponential
phase (A550 nm 0.6) at which point isopropyl-b-D-thiogalacto-
FEMS Microbiol Lett 261 (2006) 123–132c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
124 M.S.J. Centeno et al.
pyranoside (IPTG) was added to a final concentration of
1 mM and the cultures were incubated for a further 5 h.
Recombinant CmAga27A formed inclusion bodies when ex-
pressed under those conditions using E. coli BL21. Therefore,
the pET21a derivative, pZC5, encoding the Cellvibrio mixtus
a-galactosidase was transformed into E. coli Origami. Recom-
binant E. coli was cultured as described above but after
induction the cells were maintained at 16 1C for 16 h. Cells
were pelleted by these cultures by centrifugation and the
bacterial pellets were resuspended in 50 mM sodium HEPES
buffer, pH 7.5, containing 1 M NaCl and 10 mM imidazole.
The buffer of CmAga27 also contained 5 mM dithiothreitol.
The three recombinant proteins were purified by immobilized
nickel affinity chromatography as described previously (Car-
valho et al., 2004) and exchanged into the buffers described
below.
Enzyme assays
The activity of CmAga27A was routinely determined with
10 mM p-nitrophenyl a-D-galactopyranoside (pNPaGAL),
at 37 1C, in 25 mM Tris-HCl buffer, pH 8.5 (pH optimum of
the enzyme), containing 5 mM dithiothreitol. To screen for
activity against 4-nitrophenyl-glycosides, substrate and en-
zyme concentrations were 10 mM and 1 mM, respectively,
and assays were carried out for up to 1 h. One unit of
enzyme for pNPaGAL was defined as the amount of enzyme
that liberates 1 mmol of p-nitrophenol per min from pNPa-
GAL. The pH profile of the a-galactosidase was determined
with 10 mM pNPaGAL using the following buffers: pH
6–8.5, 50 mM phosphate; pH 8.5–9.5, 50 mM Tris-HCl.
The resistance of the enzyme to proteinase and thermo-
stability were determined as described previously (Fontes
et al., 1995). Activity of CmAga27A against oligosaccharides
of the raffinose series and polysaccharides was determined
by incubating 5 mg of enzyme, in 25 mM Tris-HCl buffer, pH
8.5, containing 5 mM dithiothreitol, with the appropriate
substrate [1% (w/v)] in a volume of 100mL and released
reducing sugar was measured with the Somogyi–Nelson
assay (Nelson, 1944; Somogyi, 1952). To determine the
kinetic parameters of the enzyme against its target sub-
strates, an appropriate dilution of the enzyme was incubated
with concentrations of substrate ranging from 0.05 to
10 g L�1 for polysaccharides and 0.01–10 mM for pNPaGAL.
Activities were measured in the linear phase of the enzyme
reactions. All reported results are the mean of three
replicates.
Phosphomannose isomearse activity (converts mannose
6-phosphate to fructose 6-phosphate) was measured using a
coupled enzymatic assay utilizing phosphoglucose isomer-
ase (converts fructose 6-phosphate to glucose 6-phosphate)
and glucose 6-phosphate dehydrogenase (oxidizes glucose
6-phosphate and reduces NADP 1 to NADPH). Mannose
6-phosphate was used as the substrate and the progress of
the reaction was followed spectrophotometrically at 340 nm
by measuring the production of NADPH (Todd & Tague,
2001). To confirm the validity of the assay, the increase in
absorbance at 340 nm was monitored in a reaction contain-
ing 0.6 units of a commercial phosphomannose isomerase
(Sigma p-5153). The reaction mixture consisted of 27 mM
Tris-HCl, pH 7.5, containing 5 mM MgCl2, 1 mM b-NADP,
1 U mL�1 glucose 6-phosphate dehydrogenase (Sigma G-
8289) and 0.5 U mL�1 of phosphoglucose isomerase (Sigma
P-5381). The reaction was started by adding 3 mM of
mannose 6-phosphate (Sigma M-6876) and was monitored
at 37 1C in an Ultrospec III spectophotometer (Pharmacia)
using the enzyme kinetic module of Biochrom Ltd software
package v. 1.01 (Pharmacia). Mannokinase activity was
measured using, essentially, the assay system described
above but using mannose as the substrate and Sigma
phosphomannose isomerase to convert mannose 6-phos-
phate into fructose 6-phosphate. To measure the kinetic
parameters of CmEpiA, 25 mg of enzyme were incubated
with 5–500 mM mannose in a volume of 100mL at 37 1C in
the buffer conditions described above. Glucose production
was monitored at various time points (up to 30 min) with
the D-Glucose-HK kit from Megazyme, using a final ATP
concentration of 20 mM. Optimum pH and temperatures
were determined as described above.
Results
Cloning and sequencing of Cellvibrio genes
A Cellvibrio mixtus genomic library, prepared in lZAPIII,
was screened for a-galactosidase activity. Four a-galactosi-
dase positive clones were isolated from the library and
Table 1. Primers, vectors and restriction sites used to hyperexpress CmAga27A, CmEpiA and CmUnkA in Escherichia coli
Gene Restriction sites Vector Primers
aga27A NdeI pET21a 50CTCCATATGCAAAAATTTGAGCATCTC
XhoI 50CACCTCGAGCTGCGGTGTTAAACGCAA
epiA NheI pET28a 50CTCGCTAGCAACGCCGCCATGCTTG
XhoI 50CACCTCGAGTTAAGTTGTGTGTAC
unkA NheI pET21a 50CTCGCTAGCTTTAAAGAAAAAGCAAAA
XhoI 50CACCTCGAGAACCAAATCCTTCATTAC
Bold type represents restriction sites designed in primers.
FEMS Microbiol Lett 261 (2006) 123–132 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
125Galactomannan utilization by Cellvibrio mixtus

purified to homogeneity by replating on the host strain. The
corresponding recombinant plasmids were excised from the
isolated clones and rescued into E. coli cells which displayed
activity against pNPaGAL (not shown). Restriction map-
ping and DNA hybridization studies revealed that all iso-
lated DNA fragments comprise the same 4.7-kb Cellvibrio
mixtus genomic region (not shown) and this plasmid was
named pZC2. The isolated DNA fragment was sequenced
revealing the presence of an ORF of 1215 bp, which was
designated aga27A. The gene encodes a polypeptide of 405
residues, named CmAga27A, with a predicted Mr of 45819.
The region upstream aga27, which revealed the presence of
an incomplete ORF, was cloned using the inverse PCR
approach described previously (Fig. 1a). The derived se-
quence comprised a cluster of three genes of 1179, 1230 and
1368 bp, which were designated unkA, epiA and man5A,
respectively (Fig. 1b). The intercistronic regions separating
unk-epiA and epiA-man5A were of 3 and 12 bp, respectively.
The proteins encoded by these three genes were designated
CmUnkA, CmEpiA and CmMan5A, respectively, and dis-
played predicted Mrs of 44188, 46524 and 51620. The
nucleotide sequence of the 7-kb sequence characterized in
this report appears in GenBank with the accession number
AY526725. Southern hybridization using aga27A as the
probe showed that a single copy of the gene is present in
the bacterial genome (data not shown). Two putative
promoters were identified upstream of unkA, epiA, man5A
and aga27A, with � 10 and � 35 well conserved regions
(Fig. 1c). In addition, two regions of dyad symmetry were
identified 18 and 22 nucleotides downstream of the putative
translational stop codon of man5A and aga27A, respectively
(Fig. 1b). The two 26-bp regions have the potential to form a
substantial stem–loop structure, characteristic of a r-inde-
pendent transcription terminator sequences.
Comparison of the primary sequence of CmAga27A with
biological databanks, searched by BLAST (www.ncbi.nlm.
nih.gov/BLAST), revealed that the protein display consider-
able sequence similarity with GH27 enzymes. Sequence
alignments (Fig. 2a), show that CmAga27A displays the
highest identities (4 40%) with GH27 putative enzymes
from Microbulbifer degradans (ZP_066516), Saccharopoly-
spora erythraea (AAC99325), Streptomyces avermitilis
(NP_822650) and a-galactosidases CjAga27A and Aga27A
from Cellvibrio japonicus and Clostridium josui, respectively
(BAB83765). Interestingly, GH27 enzymes presenting lower
identity scores with CmAga27A (below 47%) are all from
eukaryotic origin. It has been shown that in GH27 substrate
specificity is mediated by the nature of the b5–a5 loop,
which was recently called the substrate recognition loop
(Fujimoto et al., 2003). In the six GH27 bacterial enzymes
the presence of cysteine and tryptophan residues in the loop,
which coordinate the 2-hydroxyl of galactose in plant and
fungi a-galactosidases, suggest that all the bacterial enzymes
are a-galactose specific (Fig. 2a). A phylogeny tree of the 36
GH27 enzymes with demonstrated a-galactosidase activity
(see CAZy database), reveals that prokaryotic and eukaryotic
proteins have evolved from a common precursor, although
mammalian, plant, fungi and bacterial sequences segregate
into distinct clusters (Fig. 2b). In common with most a-
galactosidases, CmAga27A displays no evidence of modular
architecture or delineating linker sequences. In addition,
CmAga27A contains a typical signal peptide in which two
N-terminal basic residues are followed by 20 small hydro-
phobic amino acids capable of forming a a-helical structure,
with cleavage predicted to occur between Ala-23 and Gln-24.
In a previous report, it was demonstrated that CmMan5A
belongs to GH5 and displays exo-b-mannosidase activity,
releasing mannose from the nonreducing end of manno-
oligosaccharides and polysaccharides (Dias et al., 2004).
Analysis of the primary structures of CmEpiA and CmUnkA,
suggests that both proteins are nonmodular and intracellular.
Comparison of the sequence of the two deduced polypep-
tides with biological databanks showed that the protein
encoded by unkA has various protein homologues with no
demonstrated enzymatic function, while CmEpiA exhibits
55% sequence identity with a protein from M. degradans
(ZP_0315883) and lower identity scores (below 25%) with
bacterial N-acyl-D-glucosamine-2-epimerases and phospho-
mannose isomerases. In this work we were unable to assign a
biochemical role for CmUnkA.
Biochemical properties of CmAga27A
To investigate the biochemical properties of CmAga27A, the
protein was expressed in E. coli Origami and purified to
unkA man5AepiA aga27A
T TP P
−35 −10 Start
P TTAACT <21 bp> TATAAG <4 bp> A
P TTGACG <25 bp> TATAAT <5 bp> A
1 kb
Pst I Pst IEcoR I EcoR I
pZC2pZC3
pZC4
(a)
(b)
(c)
Fig. 1. Organization of the Cellvibrio gene cluster encoding galacto-
mannan degrading enzymes. Plasmids containing DNA fragments that
allowed the assembly of the 7-kb sequence containing the four genes
identified in this work are displayed in (a). Putative terminator and
promoter sequences identified in the sequence and the position and
orientation of the ORFs are displayed in (b). (c) depicts the sequences of
the putative promoters. P, promoter; T, terminator.
FEMS Microbiol Lett 261 (2006) 123–132c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
126 M.S.J. Centeno et al.

SaNP822650 TPPMGFNNWNSTGCRPEFNED--------MVKGTADIFVEKGLKDAGYQYVNLDDCWAL- 51SeAAC99325 TPPMGWNSWNSFGC--DIDER--------LIRDTADALVGSGMRDAGYQYVVVDDCWFD- 49CjBAB83765 TPPMGWNSWNIFGG--DINED--------KIKEIADAMVTTGMKDAGYEYVNLDDNWMAN 50CmAga27A TPQLGWNSWNTFAC--DVNEK--------MIREMADAMVASGMKDAGYEYINIDDCWHG- 49CjAga27A TPQMGWNSWNTFGC--NVDEK--------MIRAMADAMVTSGMKAAGYEYINIDDCWHG- 49MdZP00066516 TPPMGWNSWNNFGC--DVDEK--------LIKETADYMVSSGMKDAGYEYVNIDDCWHG- 49HsAAH02689 TPTMGWLHWERFMCNLDCQEEPDSCISEKLFMEMAELMVSEGWKDAGYEYLCIDDCWMA- 59
SaNP822650 PARDSNGKLVPDPARFPGGIKAVADYVHSKGLKLGIYTSAGTKTCNEAG--FP---GALG 106SeAAC99325 PQRDPQGNLRANPERFPSGIRALADYVHSRGLKFGIYQVPTEKTCAQRGGTYPGATGSLG 109CjBAB83765 PARDANGKLIPDPKRFPSGMKALADYIHSKGLKFGIYGDRGVTTCCNIP-----QSGSQG 105CmAga27A -ERDKQGFIQVDKKSFPSGMKALADYVHSKGLKLGIYSDAGNTTCAGRP-------GSRG 101CjAga27A -ERDKNGFIQADKKHFPSGMKALADYVHAKGLKLGIYSDAGNTTCAGRP-------GSRG 101MdZP00066516 -ERDANGFIQADPERFPSGIKALADYVHSKGLKFGIYSDAGWTTCGGKP-------GSRG 101HsAAH02689 PQRDSEGRLQADPQRFPHGIRQLANYVHSKGLKLGIYADVGNKTCAGFP-------GSFG 112
SaNP822650 HEYSDAQQFADWGVDYLKYDNCNNQG--VDAKLRYTTMRDALKATGRPIVYSLCEWG--- 161SeAAC99325 HEEQDARTFAEWGVDYLKYDWCSPEGTLEDQIAAFTKMRDALAATGRPIVYSINSNSYHP 169CjBAB83765 YEEQDAKTFAEWGLDYLKYDNCASDS---NLQAGYEKMRDALLKTGRPIFYSICCWY--- 159CmAga27A HEYQDAVTYASWGIDYVKYDWCDTKD--INPKAAYATMRDAIHKAGRPMLFSICEWG--- 156CjAga27A HEYQDALTYASWGIDYVKYDWCDTQD--INPKSAYATMRDAIHKAGRPMLFSICEWG--- 156MdZP00066516 YEFQDAQMYAKWGVDYLKYDWCATDG--LKAEGAYQTMREAIHKAGRPMVFSICEWG--- 156HsAAH02689 YYDIDAQTFADWGVDLLKFDGCYCDS-LENLADGYKHMSLALNRTGRSIVYS-CEWPLYM 170
SaNP822650 ---ENKPWEWASDVGQLWRTTGDISDSWGS-----------MLSILKQ----NLPLAPYA 203SeAAC99325 D--KNGATHDWSPVANMWRTTEDIKPVWDSGNEN--EYPMGVVNIIDV----NRGLAAQA 221CjBAB83765 -----FAGPWMVDCGNSWRTTGDISDSWGS-----------IIRNIDE----NSKSAAYA 199CmAga27A ---DNKPWEWATDVGHSWRTTGDIYPCWNCEHNHGSWSSWGVLPILDK----QAGLRKYA 209CjAga27A ---DNQPWEWAQDVGHSWRTTGDIYPCSNCEHNHGSWSSFGVLPILDK----QAGLRKYA 209MdZP00066516 ---DNQPWEWAKPIGHLWRTTGDIYNCFDCEYDHGTWSSWGVLQILDM----QDDLRQYA 209HsAAH02689 WPFQKPNYTEIRQYCNHWRNFADIDDSWKS-----------IKSILDWTSFNQERIVDVA 219
SaNP822650 GPGHWNDPDMLEVGN------SGMTDTEYRTHFSMWSIMAAPLLIGSDLRKASAATFDIL 257SeAAC99325 RPGHWNDPDMLEVGVYDVEGFKGLTDTEARAHLSMWALMASPLIAGNNVTRMPDGIRDIL 281CjBAB83765 GPGHWNDPDMLEVGN------GNMTDTEYKAHFSMWCMMAAPLIAGNDLRNMTPATKEIL 253CmAga27A GPGHWNDMDMMEVGN-------GMNEDEDRAHFSLWAMMASPLIAGNDLRKMSEATKKIL 262CjAga27A GPGHWNDMDMMECFN-------GMTEEEDRAHFSLWAFMASPLIAGNDLRNMSDTTRAIL 262MdZP00066516 GPGHWNDPDMMEVGN-------GMTEAEDRSHFSMWAMLAAPLIAGNDIRNMSEATRKIL 262HsAAH02689 GPGGWNDPDMLVIGN------FGLSWNQQVTQMALWAIMAAPLFMSNDLRHISPQAKALL 273
SaNP822650 DNKEVIAVDQDPLGKQGTVLSSEGGRWAVTKEMK-DGS-RAVALFNETDSAQRITTTAQA 315SeAAC99325 TNREVVAVDQDPAGAQGVPVRDHGDREVWVKNMA-DGS-RVVALFNRGERPAGIRTTARE 339CjBAB83765 TNKEVIAIDQDAAGVQGTKVSSSGELEVWCKPLGTDGTTKAVALLNRGATSADITVNWSD 313CmAga27A TNKDMLAINQDKLGIQAMKWIDEGDIEIYVKPLE-KGD-YAVLFLNRADTTVNYSLDWGF 320CjAga27A THKETIAINQDKLGIQAMKWIDEGDLEIYIKPLE-KGH-YAVLFLNRADDAMDYRFDWSF 320MdZP00066516 TNKAVIAVDQDELGVQGFKYSSKNGVEVWFKPLA-NDE-WAMAVLNRNKGEVKFEFKWRN 320HsAAH02689 QDKDVIAINQDPLGKQGYQLRQGDNFEVWERPLS--GLAWAVAMINRQEIGGPRSYTIAV 331
SaNP822650 VGLPKAH----------AYTLRDLWQHRSYNTAGT-ISATVPAHGTVLVRVSA------- 357SeAAC99325 AGLPKAS----------EYQVRDLWAHETSTTDGE-IRAEVPAHGVVLLRVSAQG----- 383CjBAB83765 IQLADG-----------PVTVRDLWEHKDCGTFNTGYTANVPSHGVVVLKV--------- 353CmAga27A HYMKDDISKHEIFFDKKKFNWRDIWN-GGKGSTAEKLNLTMAAHSVAVLRLTPQ------ 373CjAga27A HYMKDDISKHEIFFDKQAFNWRNIWN-GETGSTKEVLNIKVPAHGVVVLRLSPR------ 373MdZP00066516 EVVKDELTHRTITFNEQKFDWQDLWNKSNKGHTKKFLKTKIAGHDTLMFRLTPAKN---- 376HsAAH02689 ASLGKGVACN---PACFITQLLPVKRKLGFYEWTSRLRSHINPTGTVLLQLENTMQMSLK 388
(a)
Fig. 2. Alignment of bacterial GH27 enzymes (a) and unrooted phylogenetic tree of functional a-galactosidases from GH27 (b). In (a), GH27 bacterial
a-galactosidases are aligned with the Homo sapiens homologue from which the 3D structure is known (Garman & Garboczi, 2004). Catalytic residues
(D152 and D207) and the tryptophan that stacks galactose at the active site (W40) are depicted in gray. The substrate recognition loop is boxed (see
text). Secondary structure elements are depicted using arrows (b-strands) and cylinders (a-helix). In (b), the scale bar indicates the number of
substitutions per position following alignment with CLUSTALW (Thompson et al., 1994) and bootstrap analysis using the same software. The tree was
displayed with TreeView (Page, 1996). Branches not circled contain sequences from fungi. Two letters that indicate the genus and species, respectively,
of the endogenous host precede the protein accession numbers (GenBankTM or Swiss-Prot).
FEMS Microbiol Lett 261 (2006) 123–132 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
127Galactomannan utilization by Cellvibrio mixtus

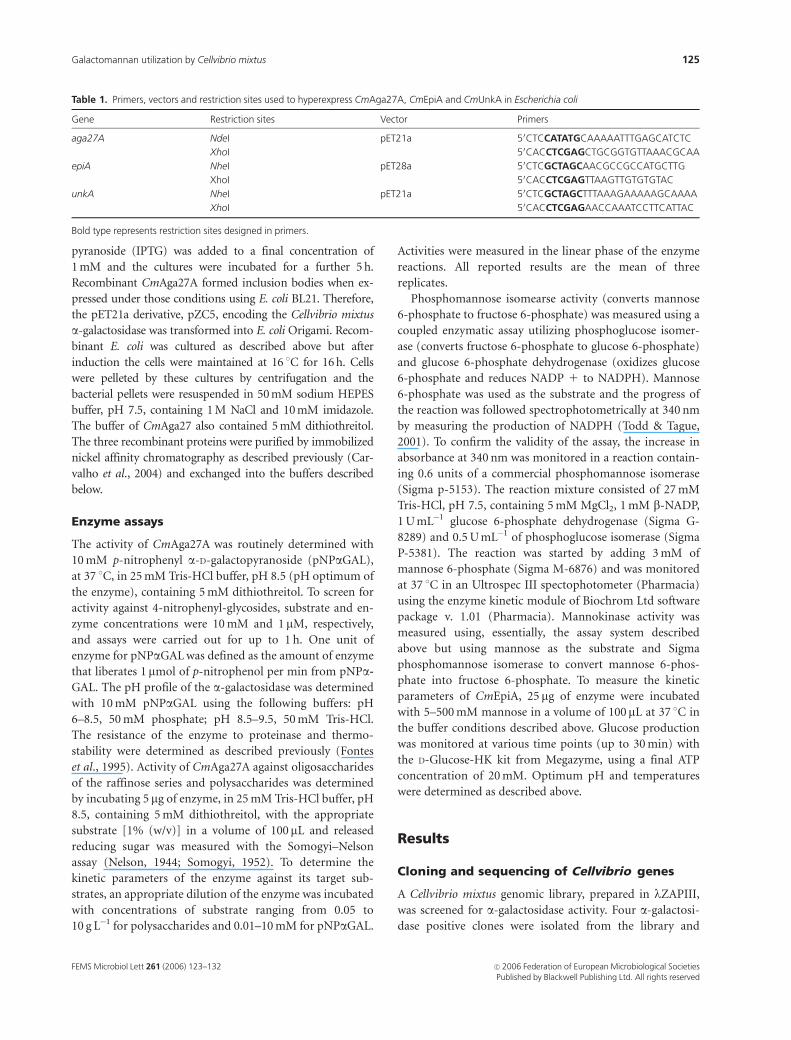
homogeneity by metal affinity chromatography (Fig. 3a).
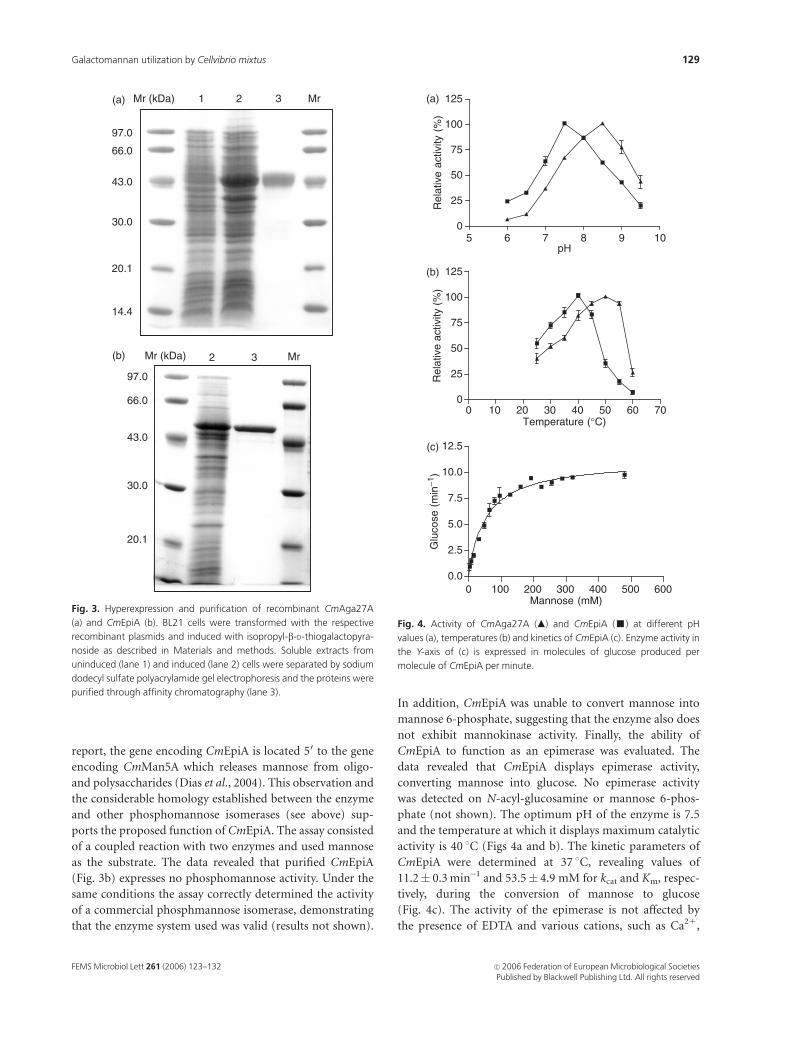
The pH and temperature profiles of CmAga27A were
determined with pNPaGAL (Figs 4a and b). The enzyme
was found to display maximum catalytic activity at 50 1C
although it was rapidly inactivated at temperatures above
55 1C (not shown). The pH optimum of CmAga27A was
found to be of 8.5 and the enzyme, in common with the
majority of the extracellular plant cell wall hydrolases, was
completely resistant to proteolytic inactivation. Purified
CmAga27A display high levels of activity against pNPaGAL
although substrate inhibition was detected at substrate
concentrations above 20 mM. Enzyme kinetics were ob-
served over the range 0.01–10 mM pNPaGAL, allowing for
the estimation of the apparent kinetic parameters that gave
values of 184.9� 4.8 min�1 and 0.23� 0.02 mM for kcat and
Km, respectively. In contrast, the enzyme does not hydrolyse
aryl-b-mannosides, 4-nitrophenyl-b-glucopyranoside or
aryl-b-arabinosides or the polysaccharides xylan, hydro-
xyethyl and carboxymethyl cellulose, mannan, undecorated
glucomannan, lichenan and laminarin. Interestingly, CmA-
ga27A exhibited activity against carob, locust bean and guar
gum galactomannans. The kinetic constants displayed by the
enzyme against these substrates are presented in Table 2. The
capacity of CmAga27A to hydrolyse the a-1,6-galactoside
linkage in oligosaccharides of the raffinose family was
evaluated. The data demonstrated that the Cellvibrio mixtus
a-galactosidase display a very marginal capacity to hydrolyse
raffinose (0.01 U mg�1) and estaquiose (0.006 U mg�1).
Taken together the results suggest that CmAga27A is an
enzyme that Cellvibrio mixtus uses to remove galactose side
chains from galactomannans.
Biochemical properties of CmEpiA
Initially, the possibility that CmEpiA was the phosphoman-
nose isomerase responsible for the interconversion of man-
nose 6-phosphate into fructose 6-phosphate in Cellvibrio
mixtus, was evaluated. In the genetic locus identified in this
0.1
HjCAA93244
PpBAA22992
AnCAB46229
PsCAA08915
MvBAA33931
MvAAB35252
PcAAG24510PcAAG24511ZcAAA35280
ZmBAA99555TdBAA86883
SpCAA64759
ScAAA34770ScCAA85739
SpAAA34769
SmCAA64760
HsAAH02689
MmAAA74453
MdZP00066516
CmAga27A
CjAga27A
CjBAB83765
SaNP822650
SeAAC99325
PsCAF34023
LeAAF04591
OsBAB12570
HaBAC66445
CsABC88435
CtCAA32772
GmAAA73963PvAAA73964
CaCAI47559CaAAA33022
CcCAI47560
Bacteria
Mammals
Plants
Fungi
(b)
Fig. 2. Continued.
FEMS Microbiol Lett 261 (2006) 123–132c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
128 M.S.J. Centeno et al.
report, the gene encoding CmEpiA is located 50 to the gene
encoding CmMan5A which releases mannose from oligo-
and polysaccharides (Dias et al., 2004). This observation and
the considerable homology established between the enzyme
and other phosphomannose isomerases (see above) sup-
ports the proposed function of CmEpiA. The assay consisted
of a coupled reaction with two enzymes and used mannose
as the substrate. The data revealed that purified CmEpiA
(Fig. 3b) expresses no phosphomannose activity. Under the
same conditions the assay correctly determined the activity
of a commercial phosphmannose isomerase, demonstrating
that the enzyme system used was valid (results not shown).
In addition, CmEpiA was unable to convert mannose into
mannose 6-phosphate, suggesting that the enzyme also does
not exhibit mannokinase activity. Finally, the ability of
CmEpiA to function as an epimerase was evaluated. The
data revealed that CmEpiA displays epimerase activity,
converting mannose into glucose. No epimerase activity
was detected on N-acyl-glucosamine or mannose 6-phos-
phate (not shown). The optimum pH of the enzyme is 7.5
and the temperature at which it displays maximum catalytic
activity is 40 1C (Figs 4a and b). The kinetic parameters of
CmEpiA were determined at 37 1C, revealing values of
11.2� 0.3 min�1 and 53.5� 4.9 mM for kcat and Km, respec-
tively, during the conversion of mannose to glucose
(Fig. 4c). The activity of the epimerase is not affected by
the presence of EDTA and various cations, such as Ca21,
Mr (kDa)
97.0
66.0
43.0
30.0
20.1
14.4
1 2 3 Mr
Mr (kDa) Mr
97.0
66.0
43.0
30.0
20.1
2 3
(a)
(b)
Fig. 3. Hyperexpression and purification of recombinant CmAga27A
(a) and CmEpiA (b). BL21 cells were transformed with the respective
recombinant plasmids and induced with isopropyl-b-D-thiogalactopyra-
noside as described in Materials and methods. Soluble extracts from
uninduced (lane 1) and induced (lane 2) cells were separated by sodium
dodecyl sulfate polyacrylamide gel electrophoresis and the proteins were
purified through affinity chromatography (lane 3).
5 6 7 8 9 100
25
50
75
100
125
pH
Rel
ativ
e ac
tivity
(%
)
0 10 20 30 40 50 60 700
25
50
75
100
125
Temperature (°C)R
elat
ive
activ
ity (
%)
0 100 200 300 400 500 6000.0
2.5
5.0
7.5
10.0
12.5
Mannose (mM)
Glu
cose
(m
in−1
)
(a)
(b)
(c)
Fig. 4. Activity of CmAga27A (m) and CmEpiA (’) at different pH
values (a), temperatures (b) and kinetics of CmEpiA (c). Enzyme activity in
the Y-axis of (c) is expressed in molecules of glucose produced per
molecule of CmEpiA per minute.
FEMS Microbiol Lett 261 (2006) 123–132 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
129Galactomannan utilization by Cellvibrio mixtus

Zn21, Na1, Mg21, K1 and Mn21, suggesting that the
enzyme does not have any metal ion requirement.
Discussion
This report describes the cloning and characterization of a
Cellvibrio mixtus gene cluster encoding enzymes involved in
the degradation of galactomannan and in the metabolism of
its hydrolysis products. It is interesting to note that three of
the four genes identified are very tightly linked and encode,
respectively, a protein of unknown function, a mannose
epimerase (CmEpiA) and an exo-b-mannosidase (CmMa-
n5A). Recently we showed that CmMan5A is a cell
bound family 5 GH that releases mannose from the non-
reducing end of both oligo- and polysaccharides (Dias et al.,
2004). The fourth gene identified here encodes a family
27 a-galactosidase, termed CmAga27A, that is located at the
30 end of, and in the same orientation to, the other three
genes.
Based on substrate specificities, a-galactosidases have
been classified into two groups (Dey et al., 1993). One
group primarily removes galactose from oligosaccharides
such as melibiose, raffinose and staquiose. These enzymes
have potential biotechnological application in reducing the
detrimental effects associated with the ingestion of galactose
containing oligosaccharides by monogastric animals (Jin-
dou et al., 2002). These oligosaccharide specific enzymes
are located in GH families 4 and 36. The second group of
a-galactosidases acts preferentially on large macromole-
cules, such as galactomannans, glycoproteins and glycoli-
pids, while retaining the capacity to hydrolyse low-Mr
substrates. These enzymes are exclusive to GH family
27 (Garman & Garboczi, 2004). The biochemical properties
of CmAga27A, a family 27 GH highly homologous to the
eukaryotic enzymes, is unusual; the enzyme attacks galacto-
mannan but not oligosaccharides containing terminal
a-D-galactose residues. Similarly, the homologous a-galacto-
sidase from Clostridium josui, Aga27A, was also shown
to prefer galactomannan rather than small substrates (Jindou
et al., 2002). It is unknown if this property is shared by the
other enzymes of the bacterial cluster of GH27, although
Cellvibrio japonicus a-galactosidase CjAga27A displayed
some activity against raffinose and stachyose (Halstead
et al., 2000).
The substrate specificity of CmAga27A might reflect the
scarcity of oligosaccharides of the raffinose family on the
milieu of Cellvibrio mixtus. The presence of a typical signal
peptide in the primary sequence of CmAga27A suggests that
this enzyme is extracellular. This is consistent with the
complete resistance of the enzyme to proteolytic inactiva-
tion, a property typical of most extra-cellular plant cell wall
polysaccharidases that must retain stability within an highly
proteolytic environment (Fontes et al., 1995). Taken to-
gether the data suggest that CmAga27A acts cooperatively
with the extracellular repertoire of endo-mannanses releas-
ing galactose and manno-oligosaccharides from galacto-
mannans, which are then absorbed by Cellvibrio mixtus.
Interestingly, CmAga27A does not possess an associated
carbohydrate binding module typical of GH acting on
complex polysaccharidases. Analysis of the molecular archi-
tectures of family 27 enzymes demonstrates that none of the
enzymes display a modular architecture, suggesting that
removal of galactose from complex macromolecules does
not require the presence of carbohydrate binding modules.
Ultimately, this might reflect the high solubility presented
by the decorated galactomannans and/or the relative acces-
sibility of the galactose side chains in the complex poly-
saccharide.
In contrast to CmAga27A, the lack of a signal peptide in
the primary sequence of CmEpiA might reflect its intracel-
lular location. It is generally assumed that mannose meta-
bolism in bacteria follows its phosphorylation and transport
into the cytoplasm by a transmembrane sugar kinase/
transporter with the subsequent isomerization to fructose
6-phosphate through the action of a phosphomanose iso-
merase. The data presented here suggest that an alternative
metabolic pathway for the utilization of mannose might
exist in Cellvibrio mixtus. Intracellular nonphosphorylated
mannose molecules can be directly converted into glucose
through the action of CmEpiA. It remains to be established
Table 2. Kinetic parameters of CmAga27A during the hydrolysis of pNPaGAL and various galactomannans
Substrate kcat (min�1)� Km (g L�1) kcat/Kmw
pNPaGAL 184.9� 4.8 0.23� 0.02z 803.91
Guar gum galactomannan 5.26� 0.5 0.82� 0.06 6.42
Locust bean galactomannan 8.27� 0.8 1.10� 0.07 7.52
Carob galactomannan (hv) 8.81� 0.4 0.62� 0.03 14.21
Carob galactomannan (lv) 31.3� 0.9 2.21� 0.04 14.16
�kcat are in molecules of product per molecule of enzyme per minute.wkcat/Km values are in (g L)�1 min�1.zSubstrate concentration in mM.
pNPaGAL, p-nitrophenyl a-D-galactopyranoside; hv, high viscosity; lv, low viscosity.
FEMS Microbiol Lett 261 (2006) 123–132c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
130 M.S.J. Centeno et al.

whether both metabolic pathways coexist in Cellvibrio
mixtus and, if so, which one predominates. As genes encod-
ing CmEpiA and CmMan5A are tightly linked, it is tempting
to speculate that the product generated by CmMan5A,
mannose, serves as substrate for CmEpiA. This observation
suggests that CmEpiA might be a fundamental enzyme in
the metabolism of mannose in aerobic soil bacteria. Indeed,
support for this view is provided by the genome sequence of
Cellvibrio japonicus, which reveals an identical gene arrange-
ment to Cellvibrio mixtus within the manA locus. Cellvibrio
japonicus contains a similar tight cluster of three genes
encoding CjUnkA, CjEpiA and CjMan5A, which display
86%, 73% and 54% sequence identity with their Cellvibrio
mixtus counterparts. Downstream of this locus is the gene
encoding CjAga27A, which exhibits 83% sequence identity
with the Cellvibrio mixtus a-galactosidase. Although it
might be tempting to speculate that CjUnkA and CmUnkA
mediate transport of mannose across the inner membrane,
this appears unlikely as the proteins display no membrane-
spanning motifs. Interestingly, however, immediately up-
stream of the Cellvibrio japonicus man5A-containing genetic
cluster, is a gene whose product displays significant sequence
identity to sodium-dependent sugar transporters, and thus
may mediate the trafficking of mannose across the inner
membrane. Cellvibrio japonicus also contains genes encod-
ing the three key enzymes of the Leloir pathway, galacto-
kinase (galK), galactose 1-phosphate uridylyltransferase
(galT) and UDP-galactose 4-epimerase (galE), which cata-
lyse the conversion of galactose to the more metabolically
useful sugar glucose 1-phosphate. Galactokinase and galac-
tose 1-phosphate uridylyltransferase are encoded by a two-
gene operon, while galE is not linked to galK or galT. The
genes encoding the Leloir pathway are not linked to the
man5A locus. As Cellvibrio mixtus and Cellvibrio japonicus
are able to release galactose and mannose from galactoman-
nan through the same complement of enzyme activities, it is
highly likely that both Cellvibrio strains will also contain the
genes encoding the Leloir pathway.
Collectively, the data presented here and in other reports
suggest that galactomannan hydrolysis by Cellvibrio results
from the synergistic interaction of a large repertoire of
enzymes. Family 5 mannanases and CmAga27A are secreted
into the extracellular milieu where the mannose-containing
hemicelluloses are located (Hogg et al., 2003). The action of
CmAga27A exposes the mannan backbone to modular
family 5 mannanases. Membrane bound family 26 man-
nanses and CmMan5A further degrade the generated
manno-oligosaccharides and soluble galactomannans into
mannose and mannobiose (Halstead et al., 2000). These
hydrolysis products, which are generated at the surface of
the organism, can be readily taken up by Cellvibrio mixtus.
The absorbed mannose can be converted to glucose in the
cytoplasm through the action of CmEpiA.
Acknowledgements
This work was supported by grants SFRH/BD/16731/2004
(to C.I.P.D.G.), PraxisXXI/BD/21250/1999 (to F.M.V.D),
and POCI/CVT/61162/2004 from the Fundacao para a
Ciencia e a Tecnologia, Portugal.
References
Berns KI & Thomas CA Jr (1965) Isolation of high molecular
weight DNA from Hemophilus influenzae. J Mol Biol 11:
476–490.
Binet MRB, Rager MN & Bouvet OMM (1998) Fructose and
mannose metabolism in Aeromonas hydrophila: identification
of transport systems and catabolic pathways. Microbiology 144:
1113–1121.
Brett CT & Waldren K (1996) Physiology and Biochemistry of
Plant Cell Walls, Vol. 1. Chapman & Hall, London.
Carvalho AL, Goyal A, Prates JAM, Bolam DM, Gilbert HJ, Pires
VMR, Ferreira LMA, Romao MJ & Fontes CMGA (2004)
Crystal structure and functional properties of the family 11
carbohydrate-binding module of cellulosomal cellulase
Lic26A-Cel5E of Clostridium thermocellum. J Biol Chem 279:
34785–34793.
Clarke JH, Laurie JI, Gilbert HJ & Hazlewood GP (1991) Multiple
xylanases of Cellulomonas fimi are encoded by distinct genes.
FEMS Microbiol Lett 67: 305–309.
Coutinho PM & Henrissat B (1999) The modular structure of
carbohydrate-active enzymes: an integrated database
approach. Recent Advances in Carbohydrate Bioengineering
(Gilbert HJ, Davies G, Henrissat B & Svensson B, eds), pp.
3–12. The Royal Society of Chemistry, Cambridge.
Dey PM, Patel S & Brouwnleader MD (1993) Induction of
a-galactosidase in Penicellium ochrochloron by guar gum.
Biotechnol Appl Biochem 17: 361–371.
Dias FMV, Vincent F, Pell G, Prates JAM, Centeno MSJ, Tailford
LE, Ferreira LMA, Fontes CMGA, Davies GJ & Gilbert HJ
(2004) Insights into the molecular determinants of substrate
specificity in glycoside hydrolase family 5 revealed by the
crystal structure and kinetics of Cellvibrio mixtus mannosidase
5A. J Biol Chem 279: 25517–25526.
Fontes CMGA, Hall J, Hirst BH, Hazelwood GP & Gilbert HJ
(1995) The resistence of cellulases and xylanases to proteolytic
inactivation. Appl Microbiol Biotechnol 43: 52–57.
Fridjonsson O, Watzlawick H, Gehweiler A, Rohrhirsch T &
Mattes R (1999) Cloning of the gene encoding a novel
thermostable alpha-galactosidase from Thermus brockianus
ITI360. Appl Environ Microbiol 65: 3955–3963.
Fujimoto Z, Kaneko S, Momma M, Kobayashi H & Mizuno H
(2003) Crystal structure of rice a-galactosidase complexed
with D-galactose. J Biol Chem 278: 20313–20318.
Garman SC & Garboczi DN (2004) The molecular defect leading
to Fabry disease: structure of human a-galactosidase. J Mol
Biol 337: 319–335.
FEMS Microbiol Lett 261 (2006) 123–132 c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
131Galactomannan utilization by Cellvibrio mixtus

Gilbert HJ, Bolam DN & Szabo L (2002) Recent Advances in
Carbohydrate Bioengineering (Teeri TT, Gilbert HJ & Feizi T,
eds), pp. 89–98. The Royal Society of Chemistry, Cambridge.
Halstead JR, Fransen MP, Eberhart RY, Park AJ, Gilbert HJ &
Hazlewood GP (2000) a-galactosidase from Pseudomonas
fluorescens subsp. cellulosa: cloning, high level expression and
its role in galactomannan hydrolysis. FEMS Microbiol Lett 192:
197–203.
Hogg D, Pell G, Dupree P, Goubet F, Martin-Orue SM, Armand S
& Gilbert HJ (2003) The modular architecture of Cellvibrio
japonicus mannanases in glycoside hydrolase families 5 and 26
points to differences in their role in mannan degradation.
Biochem J 371: 1027–1043.
Holden HM, Rayment I & Thoden JB (2003) Structure and
function of enzymes of the Leloir pathway for galactose
metabolism. J Biol Chem 278: 43885–43888.
Jindou S, Karita S, Fujino E, Fujino T, Hayashi H, Kimura T,
Sakka K & Ohmiya K (2002) a-Galactosidase Aga27A, an
enzymatic component of the Clostridium josui cellulosome.
J Bacteriol 184: 600–604.
McCleary BV (1988) Carob and guar galactomannans. Methods
Enzymol 160: 523–527.
Millward-Sadler SJ, Davidson K, Hazlewood GP, Black GW,
Gilbert HJ & Clarke JH (1995) Novel cellulose-binding
domains, NodB homologues and conserved modular
architecture in xylanases from the aerobic soil bacteria
Pseudomonas fluorescens subsp. cellulosa and Cellvibrio mixtus.
Biochem J 312: 39–48.
Nelson N (1944) A photometric adaptation of the Somogyi
method for the determination of glucose. J Biol Chem 153:
375–380.
Page RDM (1996) TreeView: an application to display
phylogenetic trees on personal computers. Comput Appl Biosci
12: 357–358.
Puls J & Schuseil J (1996) Chemistry of hemicelluloses.
Hemicelluloses and Hemicellulases (Coughlan MP &
Hazlewood GP, eds), pp. 1–28. Portland Press, London, UK.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning: A
Laboratory Manual, 2nd edn. Cold Spring Harbor Laboratory,
Cold Spring Harbor, NY.
Somogyi M (1952) Notes on sugar determination. J Biol Chem
195: 19–23.
Thompson JD, Higgins DG & Gibson TJ (1994) CLUSTAL W:
improving the sensitivity of progressive multiple sequence
alignment through sequence weighting, position-specific gap
penalties and weight matrix choice. Nucleic Acids Res 22:
4673–4680.
Todd R & Tague BW (2001) Phosphomannose isomerase: a
versatile selectable marker for Arabidopsis thaliana germ-line
transformation. Plant Mol Biol Reporter 19: 307–319.
FEMS Microbiol Lett 261 (2006) 123–132c� 2006 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
132 M.S.J. Centeno et al.




















