
Mannose Receptor Expression and Function Define a New Population of Murine Dendritic Cells
10
of July 12, 2015. This information is current as Cells Dendritic Define a New Population of Murine Mannose Receptor Expression and Function Martinez-Pomares Andrew D. Lucas, James Harris, Siamon Gordon and Luisa Emma J. McKenzie, Philip R. Taylor, Richard J. Stillion, http://www.jimmunol.org/content/178/8/4975 doi: 10.4049/jimmunol.178.8.4975 2007; 178:4975-4983; ; J Immunol References http://www.jimmunol.org/content/178/8/4975.full#ref-list-1 , 38 of which you can access for free at: cites 54 articles This article Subscriptions http://jimmunol.org/subscriptions is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/ji/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/cgi/alerts/etoc Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved. Copyright © 2007 by The American Association of 9650 Rockville Pike, Bethesda, MD 20814-3994. The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on July 12, 2015 http://www.jimmunol.org/ Downloaded from by guest on July 12, 2015 http://www.jimmunol.org/ Downloaded from
-
Upload
nottingham -
Category
Documents
-
view
1 -
download
0
Transcript of Mannose Receptor Expression and Function Define a New Population of Murine Dendritic Cells

of July 12, 2015.This information is current as
CellsDendriticDefine a New Population of Murine
Mannose Receptor Expression and Function
Martinez-PomaresAndrew D. Lucas, James Harris, Siamon Gordon and Luisa Emma J. McKenzie, Philip R. Taylor, Richard J. Stillion,
http://www.jimmunol.org/content/178/8/4975doi: 10.4049/jimmunol.178.8.4975
2007; 178:4975-4983; ;J Immunol
Referenceshttp://www.jimmunol.org/content/178/8/4975.full#ref-list-1
, 38 of which you can access for free at: cites 54 articlesThis article
Subscriptionshttp://jimmunol.org/subscriptions
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/ji/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/cgi/alerts/etocReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of9650 Rockville Pike, Bethesda, MD 20814-3994.The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 12, 2015
http://ww
w.jim
munol.org/
Dow
nloaded from

Mannose Receptor Expression and Function Define a NewPopulation of Murine Dendritic Cells1
Emma J. McKenzie,2 Philip R. Taylor, Richard J. Stillion, Andrew D. Lucas,3 James Harris,4
Siamon Gordon,5 and Luisa Martinez-Pomares5
In vitro the mannose receptor (MR) mediates Ag internalization by dendritic cells (DC) and favors the presentation of manno-sylated ligands to T cells. However, in vivo MR seems to play a role not in Ag presentation but in the homeostatic clearance ofendogenous ligands, which could have the secondary benefit of reducing the levels of endogenous Ag available for presentation tothe adaptive immune system. We have now observed that while MR� cells are consistently absent from T cell areas of spleen andmesenteric lymph nodes (LN), peripheral LN of untreated adult mice contain a minor population of MR�MHCII� in the para-cortex. This novel MR� cell population can be readily identified by flow cytometry and express markers characteristic of DC.Furthermore, these MR� DC-like cells located in T cell areas can be targeted with MR ligands (anti-MR mAb). Numbers ofMR�MHCII� cells in the paracortex are increased upon stimulation of the innate immune system and, accordingly, the amountof anti-MR mAb reaching MR�MHCII� cells in T cell areas is dramatically enhanced under these conditions. Our results indicatethat the MR can act as an Ag-acquisition system in a DC subpopulation restricted to lymphoid organs draining the periphery.Moreover, the effect of TLR agonists on the numbers of these MR� DC suggests that the immunogenicity of MR ligands could beunder the control of innate stimulation. In accordance with these observations, ligands highly specific for the MR elicit enhancedhumoral responses in vivo only when administered in combination with endotoxin. The Journal of Immunology, 2007, 178:4975–4983.
D endritic cells (DCs)6 are professional APCs with crucialroles in the induction and control of tolerance to self-Ags and immunity to pathogen-derived Ags. These
unique cells sample Ag constitutively and migrate from the pe-riphery to secondary lymphoid organs (1, 2) where they presentprocessed Ags to T cells. In the absence of infection Ag presen-tation will result in tolerance, whereas in the presence of microbialsignals DC maturation will occur, facilitating the induction of ef-fector responses. The induction of tolerance to self and innocuous
foreign Ags may also be influenced by the efficient clearance ofself-Ags by macrophages (M�s), restricting exposure of DCsto Ags.
In vivo DCs are a rare but heterogeneous collection of cellsexpressing a wide range of germline-encoded pattern recognitionreceptors. These receptors encompass several families of mole-cules including TLRs, C-type lectins, and C-type lectin-like recep-tors expressed both at the cell surface and intracellularly. DCs areable to sense the presence of foreign microbes via these receptors.Multiple signals provided by a pathogen are transduced upon li-gand recognition and ultimately govern the course of the effectorresponse toward the invader.
The mannose receptor (MR) is a C-type lectin that provides anefficient cellular internalization system for both endogenous andmicrobe-derived molecules and has a well-established role in themaintenance of tissue homeostasis as exemplified in studies ofMR-deficient mice generated by Lee et al. (3). These mice exhib-ited defective clearance of neoglycoconjugates and elevated serumlevels of multiple lysosomal hydrolases, indicating impaired clear-ance (3). The MR recognizes sulfated carbohydrates through itscysteine-rich (CR) domain (4, 5), native and denatured collagensthrough its fibronectin type II domain (6), and oligosaccharidesterminating in mannose, fucose, or N-acetyl glucosamine throughits C-type lectin-like carbohydrate recognition domains (recentlyreviewed in Ref. 7).
No expression of the MR has been documented on murine DCpopulations in vivo; in agreement with its major role in clearance,the MR is present in most tissue M�s and in hepatic and lymphaticendothelia (8). In humans, the MR has been detected in cells lo-cated within the dermis, lamina propria, and T cell areas of thetonsil (9), in inflammatory epidermal DCs from patients withatopic dermatitis (10), and in cells lining venous sinuses in thespleen (11). Evidence for the involvement of the MR in Ag pre-sentation to the acquired immune system is limited and in some
Sir William Dunn School of Pathology, University of Oxford, Oxford, United Kingdom
Received for publication September 7, 2006. Accepted for publication January22, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was funded by the Arthritis Research Campaign, the Medical ResearchCouncil (U.K.), the Edward P. Abraham Research Fund, and the University ofNottingham.2 Current address: Department of Medicine and Therapeutics, Institute of MedicalSciences University of Aberdeen, Foresterhill, Aberdeen, Scotland AB25 2ZD, U.K.3 Current address: Centre for Clinical Immunology and Biomedical Statistics andMurdoch University, Department of Clinical Immunology, Level 2, North Block,Royal Perth Hospital, Wellington Street, Perth 6001, Australia.4 Current address: Department of Clinical Medicine, Trinity Centre for Health Sci-ences, St. James Hospital, Dublin 8, Ireland.5 Address correspondence and reprint requests to Dr. Siamon Gordon, Sir WilliamDunn School of Pathology, South Parks Road, Oxford OX1 3RE, U.K.; E-mail ad-dress: [email protected] or Dr. Luisa Martinez-Pomares at the current ad-dress: Institute of Infection, Immunity and Inflammation, School of Molecular Med-ical Sciences, University of Nottingham, Queen’s Medical Centre, Floor A, WestBlock NG7 2UH, U.K.; E-mail address: [email protected] Abbreviations used in this paper: DC, dendritic cell; AP, alkaline phosphatase; CRcysteine-rich; LN, lymph node; M�, macrophage; mLN, mesenteric LN; moDC,monocyte-derived DC; MR, mannose receptor; Pam3Cys4, (S)-]2,3-bis(palmitoyloxy)-(2-RS)-propyl]-N-palmitoyl-(R)-Cys-(S)-Ser-(S)-Lys4-OH,3HCl;pLN, peripheral LN; poly(I:C), polyinosinic-polycytidylic acid; WT, wild type.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

cases contradictory. The MR is expressed by human and murineDCs generated in vitro, human monocyte-derived DCs (moDCs)and mouse bone marrow-derived DCs. Uptake of mannosylatedligands by moDCs leads to the delivery of Ag to MHCII� (12) andCD1b� (13) compartments and enhanced presentation to T cells(14–16). Delivery of the melanoma Ag, pmel17 through the MR inhuman moDCs using an anti-MrmAb-pmel17 fusion protein led toAg presentation via both HLA I and HLA II molecules (17), in-dicating that in human DCs the MR could provide an efficientmechanism for Ag acquisition and delivery into Ag processingpathways. In mice, bone marrow-derived DCs were shown to in-ternalize Ag through the MR for presentation to T cells, althoughMR ligands were not presented as efficiently as ligands for DEC-205, another member of the MR family of proteins (18), and MRexpression is required for cross-presentation of the soluble modelAg OVA (19). In contrast, Napper and Taylor recently reportedthat fibroblasts cotransfected with the MR and MHCII were notable to enhance the presentation of glycosylated Ag to T cells (20),and Ags engineered in fungi to enhance mannosylation elicit T cellresponses independently of the MR (21).
Several of the endogenous molecules recognized by the MR aretargeted by the immune system in autoimmune diseases such asthyroiditis (thyroglobulin) (22, 23), antineutrophil cytoplasm Ab-associated vasculitis (myeloperoxidase) (Ref. 24 and our own un-published data), rheumatoid arthritis (collagen II, a major compo-nent of cartilage) (25), and Goodpasture’s disease (collagen IV)(26). This correlation led us to consider that if the MR contributedto Ag presentation in vivo it could mediate the inappropriate pre-sentation of its endogenous ligands to the acquired immune sys-tem. The aim of this work was to investigate whether the MR couldmediate Ag acquisition for presentation to the adaptive immunesystem under any circumstance in vivo. For this purpose we haveanalyzed MR expression in DC, determined the fate of MR ligandsupon in vivo administration and quantified the humoral responsesagainst MR ligands in naive and stimulated animals.
Our results demonstrate that stimulation of the innate immuneresponse has a profound effect on the involvement of MR in theinduction of adaptive immune responses. We have identified anovel DC population expressing a functional MR. These MR�
DCs are restricted to peripheral (p) lymph nodes (LNs) and theirnumbers are controlled by the presence of selected TLR agonists.In agreement with these data, the induction of humoral responsesagainst MR ligands in vivo takes place only in the presence ofendotoxin. The relevance of these results in regard to DC hetero-geneity and autoimmunity will be discussed.
Materials and MethodsAnimals
Mice used in this study (BALB/c, C57BL/6, and MR�/�, which were onthe C57BL/6 genetic background) were bred within our own institutionalcolonies, sex matched, and between the ages of 7 and 16 wk at the time ofstudy. Animals were kept and handled in accordance with institutionalguidelines. MR�/� mice were provided by Prof. M. Nussenzweig, (Rock-efeller University, New York).
Reagents
The TLR agonists used in these studies are LPS purified from the Hae-mophilus influenzae type b strain Eagen (a gift from S. Zamze, EdwardJenner Institute for Vaccine Research, Compton, U.K.), flagellin purifiedfrom Salmonella typhimurium (InvivoGen), polyinosinic-polycytidylicacid (poly(I:C); Amersham Biosciences) and (S)-]2,3-bis(palmitoyloxy)-(2-RS)-propyl]-N-palmitoyl-(R)-Cys-(S)-Ser-(S)-Lys4-OH,3HCl (Pam3Cys4;Calbiochem).
Tissue digestion
Peripheral (cervical, brachial, axillary, inguinal, and popliteal) and mesen-teric LNs were digested with 1.5 mg/ml collagenase D (Roche) and 1mg/ml DNase I (Roche) in RPMI 1640 for 25 min at 37°C with gentleshaking. Tissues were further broken down with gentle pipetting. Cell sus-pensions were washed twice in PBS containing 0.5% BSA and 5 mMEDTA. In some experiments cell suspensions were enriched in CD11c�
cells using anti-CD11c MACS beads (Miltenyi Biotec) following the man-ufacturer’s instructions, and in other experiments cells were used directlyfor FACS staining.
Flow cytometry
Single cell suspensions were blocked for 45 min at 4°C in 5% (v/v) heat-inactivated rabbit serum, 0.5% BSA, 5 mM EDTA, 2 mM NaN3, and 4�g/ml Fc�RII/III blocking mAb (clone 2.4G2) to reduce nonspecific Fcreceptor binding. Blocked cells were incubated with primary mAbs dilutedin the above-described blocking solution for 60 min in the dark at 4°C,washed three times with washing buffer containing 0.5% BSA, 5 mMEDTA, and 2 mM NaN3, and fixed with 1% formaldehyde. If biotinylatedmAbs were used, cells were incubated for a further 30 min in the dark at4°C with streptavidin-allophycocyanin (BD Pharmingen), washed threetimes in washing buffer, and fixed as above. Ag-specific staining obtainedwith mAbs was compared with that obtained with isotype-matched controlAbs. The primary mAbs used in this study were MR5D3- and MR6C3-Alexa Fluor 488/biotin (MR, rat IgG2a; produced in house), HL3-PE(CD11c, hamster IgG; BD Pharmingen), M5114-biotin (MHCII, rat IgG2b;produced in house), 5C6-biotin (CD11b, rat IgG2b; produced in house),KT15-PE (CD8�, ratIgG2a; Serotec), PO3-biotin (CD86, rat IgG2b; BDPharmingen), and NLDC-145-biotin (DEC-205, rat IgG2a; Cedarlane Lab-oratories). Analysis was conducted using a FACSCalibur flow cytometerand CellQuest 3.1 software (both BD Biosciences).
Immunohistochemistry
Slides were fixed for 10 min on ice with 2% paraformaldehyde, perme-abilized with 0.1% Triton X-100 in PBS, and then blocked with 5% (v/v)normal goat serum (Invitrogen Life Technologies) in PBS for 30 min toblock irrelevant binding sites. Further blocking of endogenous biotin wasachieved using an avidin/biotin blocking kit (Vector Laboratories) as perthe manufacturer’s recommendations. Staining Abs were prepared to ap-propriate concentrations in 5% (v/v) normal goat serum in PBS and incu-bated with slides for 60 min. Goat anti-rat IgG-Alexa Fluor 488 secondaryAb (Molecular Probes) diluted in PBS was applied for a further 30 min. Inthe case of double labeling with another Ab also raised in rat, an additional30-min blocking step was conducted with 100 �g/ml rat IgG (Sigma-Aldrich) before incubation with the second biotinylated primary reagent for60 min. This was followed by 30 min of incubation with a streptavidin-Cy3/-Cy5 (Jackson ImmunoResearch) or Alexa Fluor 488 (MolecularProbes) secondary reagent. Slides were counterstained with 400 ng/ml4�,6�-diamidino-2-phenylindole (Sigma-Aldrich) before mounting. Slideswere washed between each step with PBS.
Ear skin explant
Ears from BALB/c mice were removed at the base and split into dorsal andventral sides. Each half was placed dermal side down into a well of a24-well tissue culture plate containing 2 ml of medium and incubated for24 h in a 5% CO2, 95% humidity incubator at 37°C. Migrated cells werethen collected and washed in medium and cytospins were prepared.
In vivo targeting of MR ligands
Anti-MR mAbs or isotype controls were injected s.c. into the forelimb justabove the wrist or in some experiments in the leg just above the ankleunder anesthesia with 3% isoflurane in air. At various times after injection,the spleen and the peripheral and mesenteric LNs were collected and freshfrozen for immunohistochemical analysis. Injected mAb could be detectedin tissue sections by fluorescently labeled goat anti-rat IgG reagents (Mo-lecular Probes/Invitrogen Life Technologies).
Immunizations
Animals were anesthetized as previously indicated before the s.c. injectionof Ag into forelimbs just above the wrist. MR6C3, MR5D3, and controlIgG2a (clone GLIII/10; provided by Dr. R. Hodes, National Institutes ofHealth, Bethesda, MD) were used in these studies in the presence or ab-sence of LPS. After 7–14 days, animals were sacrificed and bled and serawere prepared and stored at �20°C. The mAbs were purified from hybrid-oma supernatants prepared in Iscove’s medium containing low endotoxin
4976 ROLE OF THE MANNOSE RECEPTOR IN ANTIGEN PRESENTATION IN VIVO
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
and IgG-depleted FCS (Invitrogen Life Technologies) using a GammaBindPlus Sepharose column (Invitrogen Life Technologies). All preparationswere quantified using a BCA assay (Perbio), analyzed for purity by Coo-massie staining, and tested for endotoxin contamination using the Limulusamebocyte Lysate assay (Cambrex/BioWhittaker). All proteins were ali-quoted and stored at �20°C until required.
ELISA
Total mouse anti-rat IgG produced by each animal was determined byELISA. Flat-bottom, 96-well, high-binding enzyme immunoassay/radio-immunoassay (EIA/RIA) plates (Corning) were coated with 10 �g/ml ratIgG (Sigma-Aldrich) at 50 �l/well in PBS overnight at 4°C. Plates wereblocked with 3% BSA (w/v) in PBS for 60 min at 37°C before the additionof appropriate dilutions of serum in duplicate for 1 h at room temperature.Wells were then incubated for 1 h with 50 �l/well anti-mouse IgG-alkalinephosphatase (AP) (Sigma-Aldrich) to detect mouse IgG bound to anti-ratAbs or with anti-mouse IgG1-AP or anti-mouse IgG2a-AP (both BDPharmingen) to detect specific subclasses. All AP conjugates were used ata 1/1000 dilution in PBS. Ab levels were visualized using p-nitrophenolphosphate (Sigma-Aldrich). Absorbance was measured at a wavelength of405 nm using a microplate reader. Plates were washed three times betweenincubations in PBS supplemented with 0.1% Tween 20 (Sigma-Aldrich).IgG titers were ascertained by calculating the dilution of serum required toachieve an absorbance value of 0.2. Animals that did not make a detectableanti-rat IgG response were assigned an arbitrary value, the minimum di-lution level of serum used and, thus, the level of detection.
Statistical analysis
Statistical analysis was performed using ANOVA and the Bonferroni testwith GraphPad Prism software version 3.02. Where appropriate, p valuesare indicated within the figures.
ResultsMR� cells are found in the outer paracortical areas of selectedsecondary lymphoid tissues
The expression of the MR in secondary lymphoid tissues was in-vestigated by immunofluorescent staining. In line with our previ-ous studies, MR is abundantly expressed in the medullary regionsof LN and the red pulp of the spleen, and small numbers of MR�
cells were observed in the outer paracortex of LNs close to B cellareas (8). When different LNs were compared we found that theparacortical MR� cells were restricted to pLNs (Fig. 1A, leftpanel), which drain the skin, and absent from mesenteric (m)LNs(Fig. 1A, right panel), which drain the gut. As reported, MR isabsent from the white pulp of the spleen (data not shown and Fig.
4A). Further analysis confirmed that paracortical MR� cells alsoexpress MHCII (Fig. 1B), indicating that the MR could be ex-pressed by a subset of DCs.
Characterization of MR� cells in lymph nodes by flow cytometry
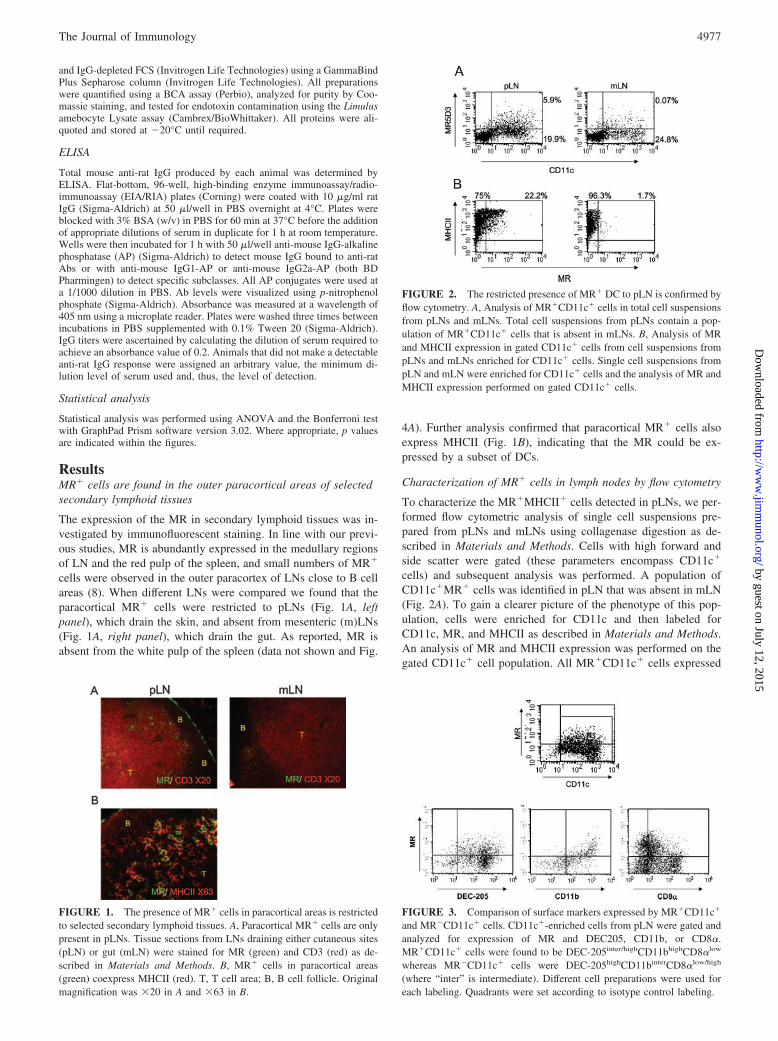
To characterize the MR�MHCII� cells detected in pLNs, we per-formed flow cytometric analysis of single cell suspensions pre-pared from pLNs and mLNs using collagenase digestion as de-scribed in Materials and Methods. Cells with high forward andside scatter were gated (these parameters encompass CD11c�
cells) and subsequent analysis was performed. A population ofCD11c�MR� cells was identified in pLN that was absent in mLN(Fig. 2A). To gain a clearer picture of the phenotype of this pop-ulation, cells were enriched for CD11c and then labeled forCD11c, MR, and MHCII as described in Materials and Methods.An analysis of MR and MHCII expression was performed on thegated CD11c� cell population. All MR�CD11c� cells expressed
FIGURE 2. The restricted presence of MR� DC to pLN is confirmed byflow cytometry. A, Analysis of MR�CD11c� cells in total cell suspensionsfrom pLNs and mLNs. Total cell suspensions from pLNs contain a pop-ulation of MR�CD11c� cells that is absent in mLNs. B, Analysis of MRand MHCII expression in gated CD11c� cells from cell suspensions frompLNs and mLNs enriched for CD11c� cells. Single cell suspensions frompLN and mLN were enriched for CD11c� cells and the analysis of MR andMHCII expression performed on gated CD11c� cells.
FIGURE 3. Comparison of surface markers expressed by MR�CD11c�
and MR�CD11c� cells. CD11c�-enriched cells from pLN were gated andanalyzed for expression of MR and DEC205, CD11b, or CD8�.MR�CD11c� cells were found to be DEC-205inter/highCD11bhighCD8�low
whereas MR�CD11c� cells were DEC-205highCD11binterCD8�low/high
(where “inter” is intermediate). Different cell preparations were used foreach labeling. Quadrants were set according to isotype control labeling.
FIGURE 1. The presence of MR� cells in paracortical areas is restrictedto selected secondary lymphoid tissues. A, Paracortical MR� cells are onlypresent in pLNs. Tissue sections from LNs draining either cutaneous sites(pLN) or gut (mLN) were stained for MR (green) and CD3 (red) as de-scribed in Materials and Methods. B, MR� cells in paracortical areas(green) coexpress MHCII (red). T, T cell area; B, B cell follicle. Originalmagnification was �20 in A and �63 in B.
4977The Journal of Immunology
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
MHCII, with the majority of the cells expressing high levels ofMHCII (Fig. 2B). In accordance with previous results,CD11c�MR�MHCII� cells were absent in mLNs. MR�CD11c�
and MR�CD11c� cells from pLN were compared for the expres-sion of several DC-associated Ags (Fig. 3). MR�CD11c� cellswere found to be DEC-205inter/highCD11bhighCD8�low, whereasMR�CD11c� cells were DEC-205highCD11binterCD8�low/high
(where “inter” is intermediate). Based on their levels of DEC-205and CD11b expression, MR�MHCII�CD11c� cells seemed tocorrespond to the interstitial DC population described by Henri etal. (27). Further studies showed that MR�CD11c� cells expressedthe costimulatory molecules CD40 (data not shown) and CD86(see Fig. 4B).
Numbers of MR� DC are under the control of innatestimulation
To address the possibility of MR� DC being influenced by innatestimulation in a similar way to that described for cells expressingligands for the CR domain of the MR (28–30), we analyzed theeffect of systemic and local stimulation with microbial products
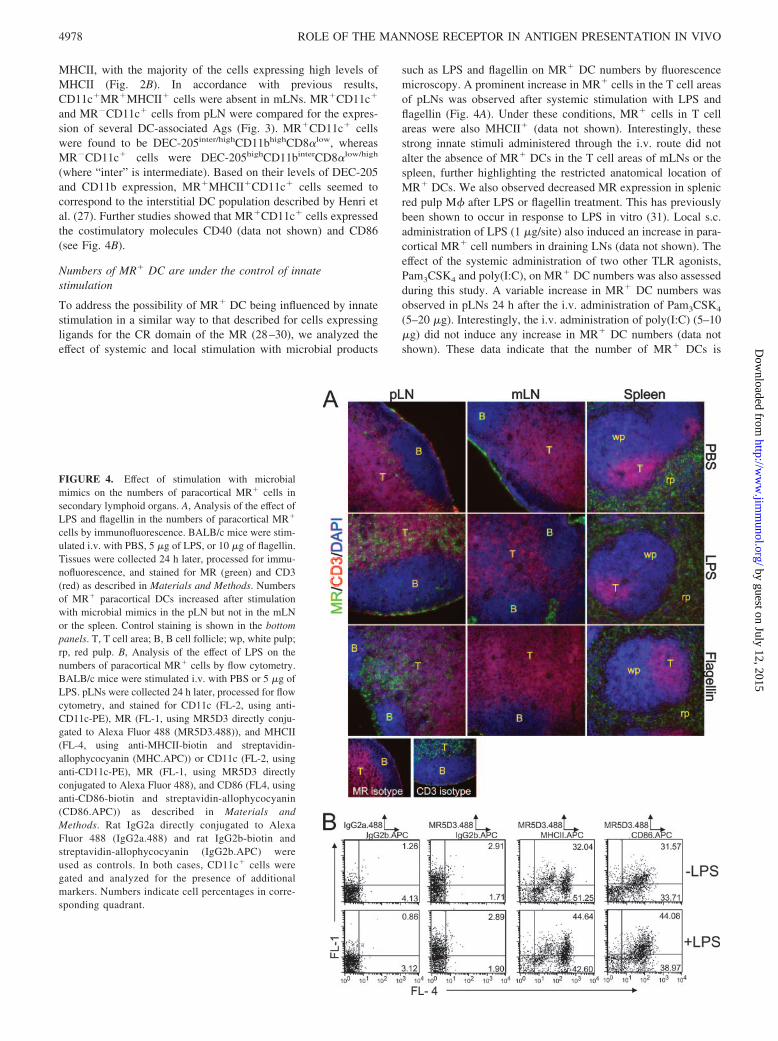
such as LPS and flagellin on MR� DC numbers by fluorescencemicroscopy. A prominent increase in MR� cells in the T cell areasof pLNs was observed after systemic stimulation with LPS andflagellin (Fig. 4A). Under these conditions, MR� cells in T cellareas were also MHCII� (data not shown). Interestingly, thesestrong innate stimuli administered through the i.v. route did notalter the absence of MR� DCs in the T cell areas of mLNs or thespleen, further highlighting the restricted anatomical location ofMR� DCs. We also observed decreased MR expression in splenicred pulp M� after LPS or flagellin treatment. This has previouslybeen shown to occur in response to LPS in vitro (31). Local s.c.administration of LPS (1 �g/site) also induced an increase in para-cortical MR� cell numbers in draining LNs (data not shown). Theeffect of the systemic administration of two other TLR agonists,Pam3CSK4 and poly(I:C), on MR� DC numbers was also assessedduring this study. A variable increase in MR� DC numbers wasobserved in pLNs 24 h after the i.v. administration of Pam3CSK4
(5–20 �g). Interestingly, the i.v. administration of poly(I:C) (5–10�g) did not induce any increase in MR� DC numbers (data notshown). These data indicate that the number of MR� DCs is
FIGURE 4. Effect of stimulation with microbialmimics on the numbers of paracortical MR� cells insecondary lymphoid organs. A, Analysis of the effect ofLPS and flagellin in the numbers of paracortical MR�
cells by immunofluorescence. BALB/c mice were stim-ulated i.v. with PBS, 5 �g of LPS, or 10 �g of flagellin.Tissues were collected 24 h later, processed for immu-nofluorescence, and stained for MR (green) and CD3(red) as described in Materials and Methods. Numbersof MR� paracortical DCs increased after stimulationwith microbial mimics in the pLN but not in the mLNor the spleen. Control staining is shown in the bottompanels. T, T cell area; B, B cell follicle; wp, white pulp;rp, red pulp. B, Analysis of the effect of LPS on thenumbers of paracortical MR� cells by flow cytometry.BALB/c mice were stimulated i.v. with PBS or 5 �g ofLPS. pLNs were collected 24 h later, processed for flowcytometry, and stained for CD11c (FL-2, using anti-CD11c-PE), MR (FL-1, using MR5D3 directly conju-gated to Alexa Fluor 488 (MR5D3.488)), and MHCII(FL-4, using anti-MHCII-biotin and streptavidin-allophycocyanin (MHC.APC)) or CD11c (FL-2, usinganti-CD11c-PE), MR (FL-1, using MR5D3 directlyconjugated to Alexa Fluor 488), and CD86 (FL4, usinganti-CD86-biotin and streptavidin-allophycocyanin(CD86.APC)) as described in Materials andMethods. Rat IgG2a directly conjugated to AlexaFluor 488 (IgG2a.488) and rat IgG2b-biotin andstreptavidin-allophycocyanin (IgG2b.APC) wereused as controls. In both cases, CD11c� cells weregated and analyzed for the presence of additionalmarkers. Numbers indicate cell percentages in corre-sponding quadrant.
4978 ROLE OF THE MANNOSE RECEPTOR IN ANTIGEN PRESENTATION IN VIVO
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
regulated by selective TLR agonists. These results were supportedby the flow cytometric analysis of cells suspensions from the pLNsof LPS-treated or untreated animals, which demonstrated that LPStreatment led to the presence of an increased percentage ofCD11c� cells in pLNs (from 2.15 to 3.3% and from 2.69 to 3.55%in two separate experiments); a higher proportion of MR�MHCII�
and MR�CD86� cells were detected within the CD11c� cellpopulation from treated animals (Fig. 4B).
MR� cells in the skin as potential precursors of MR� DC in LNs
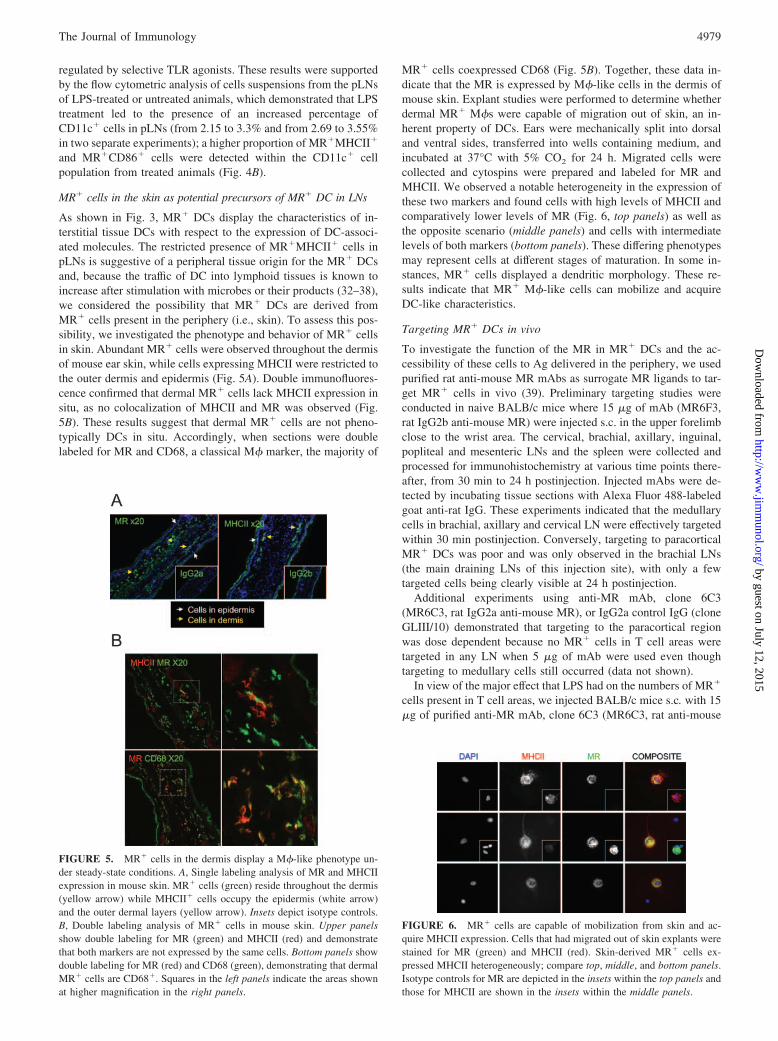
As shown in Fig. 3, MR� DCs display the characteristics of in-terstitial tissue DCs with respect to the expression of DC-associ-ated molecules. The restricted presence of MR�MHCII� cells inpLNs is suggestive of a peripheral tissue origin for the MR� DCsand, because the traffic of DC into lymphoid tissues is known toincrease after stimulation with microbes or their products (32–38),we considered the possibility that MR� DCs are derived fromMR� cells present in the periphery (i.e., skin). To assess this pos-sibility, we investigated the phenotype and behavior of MR� cellsin skin. Abundant MR� cells were observed throughout the dermisof mouse ear skin, while cells expressing MHCII were restricted tothe outer dermis and epidermis (Fig. 5A). Double immunofluores-cence confirmed that dermal MR� cells lack MHCII expression insitu, as no colocalization of MHCII and MR was observed (Fig.5B). These results suggest that dermal MR� cells are not pheno-typically DCs in situ. Accordingly, when sections were doublelabeled for MR and CD68, a classical M� marker, the majority of
MR� cells coexpressed CD68 (Fig. 5B). Together, these data in-dicate that the MR is expressed by M�-like cells in the dermis ofmouse skin. Explant studies were performed to determine whetherdermal MR� M�s were capable of migration out of skin, an in-herent property of DCs. Ears were mechanically split into dorsaland ventral sides, transferred into wells containing medium, andincubated at 37°C with 5% CO2 for 24 h. Migrated cells werecollected and cytospins were prepared and labeled for MR andMHCII. We observed a notable heterogeneity in the expression ofthese two markers and found cells with high levels of MHCII andcomparatively lower levels of MR (Fig. 6, top panels) as well asthe opposite scenario (middle panels) and cells with intermediatelevels of both markers (bottom panels). These differing phenotypesmay represent cells at different stages of maturation. In some in-stances, MR� cells displayed a dendritic morphology. These re-sults indicate that MR� M�-like cells can mobilize and acquireDC-like characteristics.
Targeting MR� DCs in vivo
To investigate the function of the MR in MR� DCs and the ac-cessibility of these cells to Ag delivered in the periphery, we usedpurified rat anti-mouse MR mAbs as surrogate MR ligands to tar-get MR� cells in vivo (39). Preliminary targeting studies wereconducted in naive BALB/c mice where 15 �g of mAb (MR6F3,rat IgG2b anti-mouse MR) were injected s.c. in the upper forelimbclose to the wrist area. The cervical, brachial, axillary, inguinal,popliteal and mesenteric LNs and the spleen were collected andprocessed for immunohistochemistry at various time points there-after, from 30 min to 24 h postinjection. Injected mAbs were de-tected by incubating tissue sections with Alexa Fluor 488-labeledgoat anti-rat IgG. These experiments indicated that the medullarycells in brachial, axillary and cervical LN were effectively targetedwithin 30 min postinjection. Conversely, targeting to paracorticalMR� DCs was poor and was only observed in the brachial LNs(the main draining LNs of this injection site), with only a fewtargeted cells being clearly visible at 24 h postinjection.
Additional experiments using anti-MR mAb, clone 6C3(MR6C3, rat IgG2a anti-mouse MR), or IgG2a control IgG (cloneGLIII/10) demonstrated that targeting to the paracortical regionwas dose dependent because no MR� cells in T cell areas weretargeted in any LN when 5 �g of mAb were used even thoughtargeting to medullary cells still occurred (data not shown).
In view of the major effect that LPS had on the numbers of MR�
cells present in T cell areas, we injected BALB/c mice s.c. with 15�g of purified anti-MR mAb, clone 6C3 (MR6C3, rat anti-mouse
FIGURE 5. MR� cells in the dermis display a M�-like phenotype un-der steady-state conditions. A, Single labeling analysis of MR and MHCIIexpression in mouse skin. MR� cells (green) reside throughout the dermis(yellow arrow) while MHCII� cells occupy the epidermis (white arrow)and the outer dermal layers (yellow arrow). Insets depict isotype controls.B, Double labeling analysis of MR� cells in mouse skin. Upper panelsshow double labeling for MR (green) and MHCII (red) and demonstratethat both markers are not expressed by the same cells. Bottom panels showdouble labeling for MR (red) and CD68 (green), demonstrating that dermalMR� cells are CD68�. Squares in the left panels indicate the areas shownat higher magnification in the right panels.
FIGURE 6. MR� cells are capable of mobilization from skin and ac-quire MHCII expression. Cells that had migrated out of skin explants werestained for MR (green) and MHCII (red). Skin-derived MR� cells ex-pressed MHCII heterogeneously; compare top, middle, and bottom panels.Isotype controls for MR are depicted in the insets within the top panels andthose for MHCII are shown in the insets within the middle panels.
4979The Journal of Immunology
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
IgG2a), or IgG2a control IgG (clone GLIII/10) in the upper andlower forelimbs of mice treated i.v. with of 5 �g of LPS or PBS 10min earlier. We detected MR6C3 in the medullary regions of thecervical, brachial, axillary, and inguinal LNs of LPS- and PBS-treated mice. In PBS-treated animals few targeted cells were de-tected in the T cell areas of brachial or inguinal LNs (Fig. 7, A andB show representative inguinal LN). In the presence of LPS, theanti-MR mAb targeted numerous cells within the paracortical ar-eas of brachial, axillary and inguinal LN (Fig. 7, C and D; a rep-resentative inguinal LN is shown). In all cases, targeted paracor-tical cells were MR� and MHCII�, indicating specific delivery toMR� DCs (Fig. 7, E–G). Delivery of anti-MR mAb was exquis-itely restricted to local draining LNs because no Ag could be de-tected in other nondraining lymphoid tissues such as, for example,the spleen (Fig. 7, H and I). No targeting of the rat IgG2a controlAb to LNs (shown in Fig. 7, A and C, insets) or spleen (Fig. 7, Hand I, insets) was observed, indicating that no targeting systemselective for rat IgG2a is present in secondary lymphoid organs.Similar results were obtained when the anti-MR mAb was injecteds.c. in combination with LPS (1 �g/site), data not shown. Thus,MR� DCs can acquire MR ligands delivered in the periphery withnumbers of MR� DCs in T cell areas containing MR ligands beingincreased in the presence of LPS.
Generation of an anti-rat IgG Ab response after immunizationwith anti-MR mAbs
Because we were able to target MR� DCs specifically in vivousing rat anti-mouse MR mAbs, we sought to determine whetherthe delivery of Ag via the MR results in presentation to the adap-tive immune system in an immunogenic fashion by assessing the
generation of anti-rat IgG Abs in sera from s.c. immunized ani-mals. Preliminary studies using 15 �g of MR6C3, MR5D3, andIgG2a indicated that no detectable response could be obtained inthe absence of LPS and that 1 �g was better than 0.1 �g of LPSin promoting a humoral response (data not shown). These exper-iments also indicated that the mAb clone MR5D3 could elicit amore robust response than MR6C3.
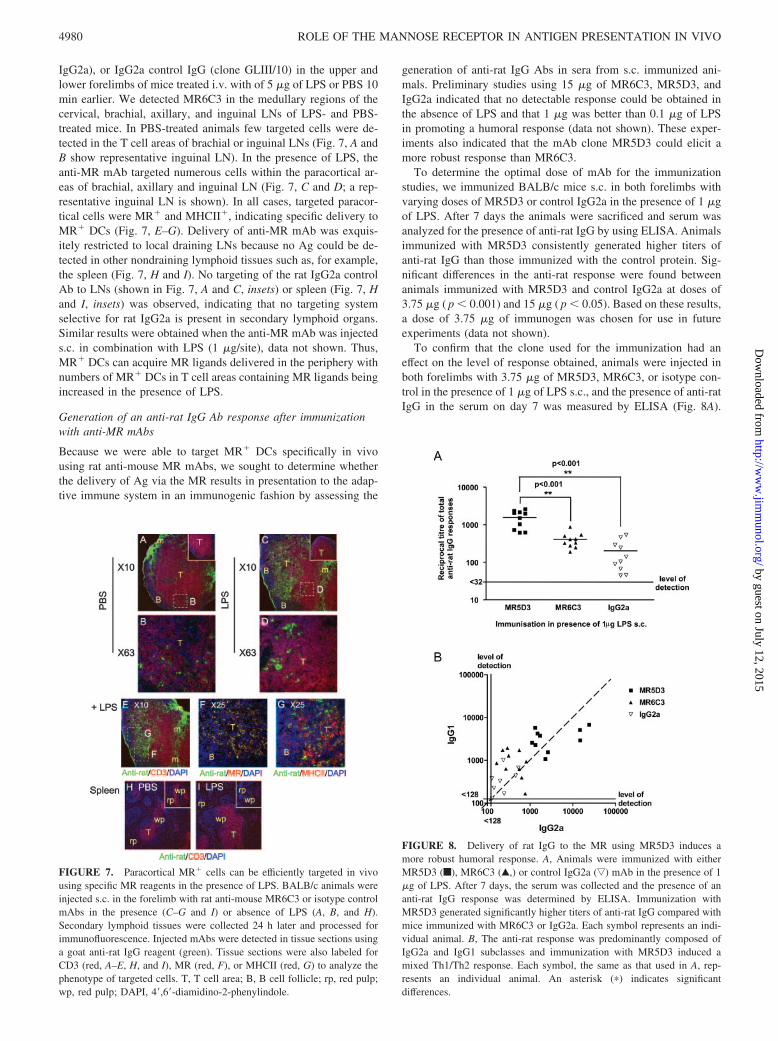
To determine the optimal dose of mAb for the immunizationstudies, we immunized BALB/c mice s.c. in both forelimbs withvarying doses of MR5D3 or control IgG2a in the presence of 1 �gof LPS. After 7 days the animals were sacrificed and serum wasanalyzed for the presence of anti-rat IgG by using ELISA. Animalsimmunized with MR5D3 consistently generated higher titers ofanti-rat IgG than those immunized with the control protein. Sig-nificant differences in the anti-rat response were found betweenanimals immunized with MR5D3 and control IgG2a at doses of3.75 �g ( p � 0.001) and 15 �g ( p � 0.05). Based on these results,a dose of 3.75 �g of immunogen was chosen for use in futureexperiments (data not shown).
To confirm that the clone used for the immunization had aneffect on the level of response obtained, animals were injected inboth forelimbs with 3.75 �g of MR5D3, MR6C3, or isotype con-trol in the presence of 1 �g of LPS s.c., and the presence of anti-ratIgG in the serum on day 7 was measured by ELISA (Fig. 8A).
FIGURE 7. Paracortical MR� cells can be efficiently targeted in vivousing specific MR reagents in the presence of LPS. BALB/c animals wereinjected s.c. in the forelimb with rat anti-mouse MR6C3 or isotype controlmAbs in the presence (C–G and I) or absence of LPS (A, B, and H).Secondary lymphoid tissues were collected 24 h later and processed forimmunofluorescence. Injected mAbs were detected in tissue sections usinga goat anti-rat IgG reagent (green). Tissue sections were also labeled forCD3 (red, A–E, H, and I), MR (red, F), or MHCII (red, G) to analyze thephenotype of targeted cells. T, T cell area; B, B cell follicle; rp, red pulp;wp, red pulp; DAPI, 4�,6�-diamidino-2-phenylindole.
FIGURE 8. Delivery of rat IgG to the MR using MR5D3 induces amore robust humoral response. A, Animals were immunized with eitherMR5D3 (f), MR6C3 (Œ,) or control IgG2a (ƒ) mAb in the presence of 1�g of LPS. After 7 days, the serum was collected and the presence of ananti-rat IgG response was determined by ELISA. Immunization withMR5D3 generated significantly higher titers of anti-rat IgG compared withmice immunized with MR6C3 or IgG2a. Each symbol represents an indi-vidual animal. B, The anti-rat response was predominantly composed ofIgG2a and IgG1 subclasses and immunization with MR5D3 induced amixed Th1/Th2 response. Each symbol, the same as that used in A, rep-resents an individual animal. An asterisk (�) indicates significantdifferences.
4980 ROLE OF THE MANNOSE RECEPTOR IN ANTIGEN PRESENTATION IN VIVO
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from
Differences between the responses induced by MR5D3 and IgG2aand by MR5D3 and MR6C3 were found to be highly significant( p � 0.001). These data indicate that Ag delivery through the MR,achieved by immunization with MR5D3 in the presence of LPS,induced improved Ab production compared with the control. In-terestingly, the efficiency of anti-rat responses also appears to bedependent on the clone of mAb used.
Analysis of the presence of the IgG1 and IgG2a subclasses gen-erated in immunized mice (Fig. 8B) showed that the animals im-munized with MR5D3 generated stronger IgG2a and IgG1 re-sponses compared with animals immunized with MR6C3 or theIgG2a control. The production of both IgG2a and IgG1 indicatedthat a mixed Th1/Th2 response was generated.
Humoral responses to anti-MR mAbs are abrogated in MR�/�
animals
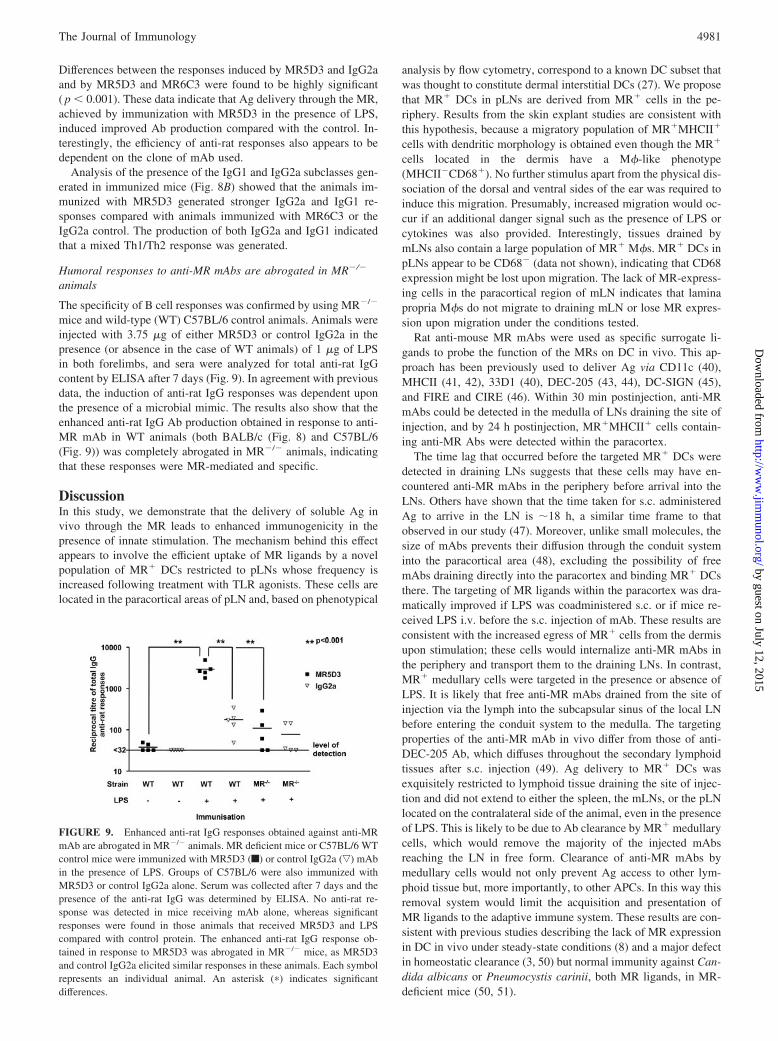
The specificity of B cell responses was confirmed by using MR�/�
mice and wild-type (WT) C57BL/6 control animals. Animals wereinjected with 3.75 �g of either MR5D3 or control IgG2a in thepresence (or absence in the case of WT animals) of 1 �g of LPSin both forelimbs, and sera were analyzed for total anti-rat IgGcontent by ELISA after 7 days (Fig. 9). In agreement with previousdata, the induction of anti-rat IgG responses was dependent uponthe presence of a microbial mimic. The results also show that theenhanced anti-rat IgG Ab production obtained in response to anti-MR mAb in WT animals (both BALB/c (Fig. 8) and C57BL/6(Fig. 9)) was completely abrogated in MR�/� animals, indicatingthat these responses were MR-mediated and specific.
DiscussionIn this study, we demonstrate that the delivery of soluble Ag invivo through the MR leads to enhanced immunogenicity in thepresence of innate stimulation. The mechanism behind this effectappears to involve the efficient uptake of MR ligands by a novelpopulation of MR� DCs restricted to pLNs whose frequency isincreased following treatment with TLR agonists. These cells arelocated in the paracortical areas of pLN and, based on phenotypical
analysis by flow cytometry, correspond to a known DC subset thatwas thought to constitute dermal interstitial DCs (27). We proposethat MR� DCs in pLNs are derived from MR� cells in the pe-riphery. Results from the skin explant studies are consistent withthis hypothesis, because a migratory population of MR�MHCII�
cells with dendritic morphology is obtained even though the MR�
cells located in the dermis have a M�-like phenotype(MHCII�CD68�). No further stimulus apart from the physical dis-sociation of the dorsal and ventral sides of the ear was required toinduce this migration. Presumably, increased migration would oc-cur if an additional danger signal such as the presence of LPS orcytokines was also provided. Interestingly, tissues drained bymLNs also contain a large population of MR� M�s. MR� DCs inpLNs appear to be CD68� (data not shown), indicating that CD68expression might be lost upon migration. The lack of MR-express-ing cells in the paracortical region of mLN indicates that laminapropria M�s do not migrate to draining mLN or lose MR expres-sion upon migration under the conditions tested.
Rat anti-mouse MR mAbs were used as specific surrogate li-gands to probe the function of the MRs on DC in vivo. This ap-proach has been previously used to deliver Ag via CD11c (40),MHCII (41, 42), 33D1 (40), DEC-205 (43, 44), DC-SIGN (45),and FIRE and CIRE (46). Within 30 min postinjection, anti-MRmAbs could be detected in the medulla of LNs draining the site ofinjection, and by 24 h postinjection, MR�MHCII� cells contain-ing anti-MR Abs were detected within the paracortex.
The time lag that occurred before the targeted MR� DCs weredetected in draining LNs suggests that these cells may have en-countered anti-MR mAbs in the periphery before arrival into theLNs. Others have shown that the time taken for s.c. administeredAg to arrive in the LN is �18 h, a similar time frame to thatobserved in our study (47). Moreover, unlike small molecules, thesize of mAbs prevents their diffusion through the conduit systeminto the paracortical area (48), excluding the possibility of freemAbs draining directly into the paracortex and binding MR� DCsthere. The targeting of MR ligands within the paracortex was dra-matically improved if LPS was coadministered s.c. or if mice re-ceived LPS i.v. before the s.c. injection of mAb. These results areconsistent with the increased egress of MR� cells from the dermisupon stimulation; these cells would internalize anti-MR mAbs inthe periphery and transport them to the draining LNs. In contrast,MR� medullary cells were targeted in the presence or absence ofLPS. It is likely that free anti-MR mAbs drained from the site ofinjection via the lymph into the subcapsular sinus of the local LNbefore entering the conduit system to the medulla. The targetingproperties of the anti-MR mAb in vivo differ from those of anti-DEC-205 Ab, which diffuses throughout the secondary lymphoidtissues after s.c. injection (49). Ag delivery to MR� DCs wasexquisitely restricted to lymphoid tissue draining the site of injec-tion and did not extend to either the spleen, the mLNs, or the pLNlocated on the contralateral side of the animal, even in the presenceof LPS. This is likely to be due to Ab clearance by MR� medullarycells, which would remove the majority of the injected mAbsreaching the LN in free form. Clearance of anti-MR mAbs bymedullary cells would not only prevent Ag access to other lym-phoid tissue but, more importantly, to other APCs. In this way thisremoval system would limit the acquisition and presentation ofMR ligands to the adaptive immune system. These results are con-sistent with previous studies describing the lack of MR expressionin DC in vivo under steady-state conditions (8) and a major defectin homeostatic clearance (3, 50) but normal immunity against Can-dida albicans or Pneumocystis carinii, both MR ligands, in MR-deficient mice (50, 51).
FIGURE 9. Enhanced anti-rat IgG responses obtained against anti-MRmAb are abrogated in MR�/� animals. MR deficient mice or C57BL/6 WTcontrol mice were immunized with MR5D3 (f) or control IgG2a (ƒ) mAbin the presence of LPS. Groups of C57BL/6 were also immunized withMR5D3 or control IgG2a alone. Serum was collected after 7 days and thepresence of the anti-rat IgG was determined by ELISA. No anti-rat re-sponse was detected in mice receiving mAb alone, whereas significantresponses were found in those animals that received MR5D3 and LPScompared with control protein. The enhanced anti-rat IgG response ob-tained in response to MR5D3 was abrogated in MR�/� mice, as MR5D3and control IgG2a elicited similar responses in these animals. Each symbolrepresents an individual animal. An asterisk (�) indicates significantdifferences.
4981The Journal of Immunology
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

After identifying the conditions under which MR could be ex-ploited by professional APCs to internalize Ag for presentation tothe adaptive immune system in vivo, anti-MR mAb were used todetermine whether the enhanced targeting of MR ligands to DCs inT cell areas correlated with the generation of enhanced humoralresponses. This strategy involved the immunization of animalswith purified rat IgG preparations and analysis of anti-rat IgG re-sponses in the serum from injected animals.
In the presence of LPS, a single s.c. dose of rat anti-mouse MRmAb induced significant anti-rat IgG production compared withthe isotype control mAb. The enhanced anti-rat response was com-pletely abrogated in MR�/� mice, illustrating the capacity of MR�
cells to promote B cell responses and the specificity of the system.These results provide the first conclusive in vivo evidence of a rolefor the MR in the induction of adaptive immune responses. Inter-estingly, differential humoral responses were induced by immuni-zation with the MR5D3 and MR6C3 clones. Both of these mAbsrecognize the C-type lectin-like carbohydrate recognition domain4-7 region of the MR, but it is not known whether the bindingaffinities of these clones differ and/or whether their intracellularhandling is different. This requires further investigation but mayreflect results in the human system the where differential engage-ment of the MR on moDCs by anti-human MR mAbs inducesdifferential programs of activation. This was also shown to occurfor some natural ligands (52).
It will also be important to determine the exact contribution ofMR� DCs in the generation of humoral responses, given that M�sbearing ligands for the CR domain have been shown to bind in-jected CR domain-bearing fusion proteins in the subcapsular sinus,potentially mimicking the delivery of Ags by soluble MRs (53). Inaddition to direct targeting to MR� cells in the draining LNs, it ispossible that the mAbs injected in this study also bound to freesoluble MR and were delivered to CR domain-ligand� cells lo-cated in the subcapsular sinus. In the presence of stimulation, tar-geted CR domain-ligand� cells would migrate into B cell follicles(28, 30) and present native Ags in complex with soluble MR todifferentiating B cells in the germinal center. However, the patternof anti-MR mAb targeting in vivo is not consistent with the de-livery of mAbs to CR domain-ligand� cells via soluble MR. Fur-thermore, the soluble MR Ag delivery pathway would not be fa-vored under the conditions tested here because CR domainmultimerization is required for optimal targeting to CR domain-ligand� cells in vivo (53) and, given that the anti-MR mAb isprobably in a monomeric form, it would be an unsuitable ligand forinducing CR domain multimerization.
The induction of CD4� or CD8� T cell responses via Ag de-livery through the MR was not addressed in this study and is thefocus of future work. However, indirect evidence from the datapresented here suggests that T cells can become activated and as-sist in the process of Ig isotype switching in response to MR li-gands. During the course of these studies a role for MR in thecross-presentation of soluble OVA in vitro and in vivo has beensuggested by others (19). Intriguingly, in this work the authorsdemonstrate a defect in the uptake by CD11c� cells and the cross-presentation of soluble OVA in the spleen and bone marrow ofMR-deficient animals. These results are not in agreement with ourstudies demonstrating the lack of MR expression in splenic DC byboth immunofluorescence and flow cytometric analysis of splenicCD11c� cells (data not shown). Future studies using chimeric anti-MR mAbs bearing CD4 and CD8 epitopes will clarify this issue,because the contribution of other putative receptors or a defect inDC function in the absence of MR can be ruled out by using thissystem.
In this study, we demonstrate the presence of a previously un-known murine MR� DC subpopulation whose numbers are con-trolled by innate stimulation. These cells are most likely derivedfrom myeloid skin leukocytes that can mobilize and acquire a DCphenotype under appropriate stimulation. Efficient targeting of MRligands to MR� DC takes place when LPS is present, and thiscorrelates with an enhanced induction of humoral responsesagainst these ligands. These data provide the first in vivo evidenceof a role for MR in Ag presentation to the acquired immune systemand reveal potential pathways available for endogenous moleculesrecognized by the MR to be presented in an immunogenic form tothe acquired immune system. Moreover, the correlation betweenthe immunogenicity of MR ligands and the presence of surrogatesigns of infection (e.g., endotoxin) observed in our studies parallelsthe triggering effect that infection can have in the induction ofautoimmune diseases and thus place the MR in a pivotal positionin the induction of autoimmunity.
The benefits obtained from exploiting a homeostatic receptor asan Ag acquisition system by immunogenic DC would be derivedfrom its usefulness in increasing the sampling ability of APCs.Because the MR has a well established ability to bind pathogen-derived products and, when expressed on DCs, is able to target Agfor presentation (see Ref. 54 for review), the existence of a highlyregulated and restricted MR-mediated Ag presentation pathway inthe context of infection would ensure the recognition of microbialproducts that could otherwise escape presentation due to efficientclearance. In this way, the presentation of endogenous moleculeswould be minimized and this, together with effective induction ofcentral and peripheral tolerance, will limit the generation of patho-logical immune responses.
AcknowledgmentsWe thank Prof. M. Nussenzweig for providing the MR-deficient mice andElizabeth Darley for technical support.
DisclosuresThe authors have no financial conflict of interest.
References1. Pugh, C. W., G. G. MacPherson, and H. W. Steer. 1983. Characterization of
nonlymphoid cells derived from rat peripheral lymph. J. Exp. Med. 157:1758–1779.
2. Huang, F.-P., N. Platt, M. Wykes, J. R. Major, T. J. Powell, C. D. Jenkins, andG. G. MacPherson. 2000. A discrete subpopulation of dendritic cells transportsapoptotic intestinal epithelial cells to T cell areas of mesenteric lymph nodes.J. Exp. Med. 191: 435–444.
3. Lee, S. J., S. Evers, D. Roeder, A. F. Parlow, J. Risteli, L. Risteli, Y. C. Lee,T. Feizi, H. Langen, and M. C. Nussenzweig. 2002. Mannose receptor-mediatedregulation of serum glycoprotein homeostasis. Science 295: 1898–1901.
4. Leteux, C., W. Chai, R. W. Loveless, C. T. Yuen, L. Uhlin-Hansen,Y. Combarnous, M. Jankovic, S. C. Maric, Z. Misulovin, M. C. Nussenzweig,and T. Feizi. 2000. The cysteine-rich domain of the macrophage mannose recep-tor is a multispecific lectin that recognizes chondroitin sulfates A and B andsulfated oligosaccharides of blood group Lewis(a) and Lewis(x) types in additionto the sulfated N-glycans of lutropin. J. Exp. Med. 191: 1117–1126.
5. Liu, Y., A. J. Chirino, Z. Misulovin, C. Leteux, T. Feizi, M. C. Nussenzweig, andP. J. Bjorkman. 2000. Crystal structure of the cysteine-rich domain of mannosereceptor complexed with a sulfated carbohydrate ligand. J. Exp. Med. 191:1105–1116.
6. Martinez-Pomares, L., D. Wienke, R. Stillion, E. J. McKenzie, J. N. Arnold,J. Harris, E. McGreal, R. B. Sim, C. M. Isacke, and S. Gordon. 2006. Carbohy-drate-independent recognition of collagens by the macrophage mannose receptor.Eur. J. Immunol. 36: 1074–1082.
7. Taylor, P. R., L. Martinez-Pomares, M. Stacey, H. H. Lin, G. D. Brown, andS. Gordon. 2005. Macrophage receptors and immune recognition. Annu. Rev.Immunol. 23: 901–944.
8. Linehan, S. A., L. Martinez-Pomares, P. D. Stahl, and S. Gordon. 1999. Mannosereceptor and its putative ligands in normal murine lymphoid and nonlymphoidorgans: in situ expression of mannose receptor by selected macrophages, endo-thelial cells, perivascular microglia, and mesangial cells, but not dendritic cells.J. Exp. Med. 189: 1961–1972.
9. Engering, A., T. B. Geijtenbeek, S. J. van Vliet, M. Wijers, E. van Liempt,N. Demaurex, A. Lanzavecchia, J. Fransen, C. G. Figdor, V. Piguet, and
4982 ROLE OF THE MANNOSE RECEPTOR IN ANTIGEN PRESENTATION IN VIVO
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from

Y. van Kooyk. 2002. The dendritic cell-specific adhesion receptor DC-SIGNinternalizes antigen for presentation to T cells. J. Immunol. 168: 2118–2126.
10. Wollenberg, A., M. Mommaas, T. Oppel, E. M. Schottdorf, S. Gunther, andM. Moderer. 2002. Expression and function of the mannose receptor CD206 onepidermal dendritic cells in inflammatory skin diseases. J. Invest. Dermatol. 118:327–334.
11. Martinez-Pomares, L., L. G. Hanitsch, R. Stillion, S. Keshav, and S. Gordon.2005. Expression of mannose receptor and ligands for its cysteine-rich domain invenous sinuses of human spleen. Lab. Invest. 85: 1238–1249.
12. Sallusto, F., M. Cella, C. Danieli, and A. Lanzavecchia. 1995. Dendritic cells usemacropinocytosis and the mannose receptor to concentrate macromolecules in themajor histocompatibility complex class II compartment: downregulation by cy-tokines and bacterial products. J. Exp. Med. 182: 389–400.
13. Prigozy, T. I., P. A. Sieling, D. Clemens, P. L. Stewart, S. M. Behar,S. A. Porcelli, M. B. Brenner, R. L. Modlin, and M. Kronenberg. 1997. Themannose receptor delivers lipoglycan antigens to endosomes for presentation toT cells by CD1b molecules. Immunity 6: 187–197.
14. Engering, A. J., M. Cella, D. Fluitsma, M. Brockhaus, E. C. Hoefsmit,A. Lanzavecchia, and J. Pieters. 1997. The mannose receptor functions as a highcapacity and broad specificity antigen receptor in human dendritic cells. Eur.J. Immunol. 27: 2417–2425.
15. Tan, M. C., A. M. Mommaas, J. W. Drijfhout, R. Jordens, J. J. Onderwater,D. Verwoerd, A. A. Mulder, A. N. van der Heiden, D. Scheidegger, L. C. Oomen,et al. 1997. Mannose receptor-mediated uptake of antigens strongly enhancesHLA class II-restricted antigen presentation by cultured dendritic cells. Eur.J. Immunol. 27: 2426–2435.
16. Mommaas, A. M., A. A. Mulder, R. Jordens, C. Out, M. C. Tan, P. Cresswell,P. M. Kluin, and F. Koning. 1999. Human epidermal Langerhans cells lack func-tional mannose receptors and a fully developed endosomal/lysosomal compart-ment for loading of HLA class II molecules. Eur. J. Immunol. 29: 571–580.
17. Ramakrishna, V., J. F. Treml, L. Vitale, J. E. Connolly, T. O’Neill, P. A. Smith,C. L. Jones, L. Z. He, J. Goldstein, P. K. Wallace, et al. 2004. Mannose receptortargeting of tumor antigen pmel17 to human dendritic cells directs anti-melanomaT cell responses via multiple HLA molecules. J. Immunol. 172: 2845–2852.
18. Mahnke, K., M. Guo, S. Lee, H. Sepulveda, S. L. Swain, M. Nussenzweig, andR. M. Steinman. 2000. The dendritic cell receptor for endocytosis, DEC-205, canrecycle and enhance antigen presentation via major histocompatibility complexclass II-positive lysosomal compartments. J. Cell Biol. 151: 673–684.
19. Burgdorf, S., V. Lukacs-Kornek, and C. Kurts. 2006. The mannose receptormediates uptake of soluble but not of cell-associated antigen for cross-presenta-tion. J. Immunol. 176: 6770–6776.
20. Napper, C. E., and M. E. Taylor. 2004 The mannose receptor fails to enhanceprocessing and presentation of a glycoprotein antigen in transfected fibroblasts.Glycobiology 14: 7C–12C.
21. Lam, J. S., M. K. Mansour, C. A. Specht, and S. M. Levitz. 2005. A modelvaccine exploiting fungal mannosylation to increase antigen immunogenicity.J. Immunol. 175: 7496–7503.
22. Linehan, S. A., L. Martinez-Pomares, R. P. da Silva, and S. Gordon. 2001. En-dogenous ligands of carbohydrate recognition domains of the mannose receptorin murine macrophages, endothelial cells and secretory cells; potential relevanceto inflammation and immunity. Eur. J. Immunol. 31: 1857–1866.
23. Roitt, I., D. Doniach, R. Campbell, and R. Hudson. 1956. Autoantibodies inHashimoto’s disease (lymphadenoid goitre). Lancet 2: 820–821.
24. Savige, J., M. Gallicchio, T. Georgiou, and D. Davies. 1990. Diverse target an-tigens recognized by circulating antibodies in anti-neutrophil cytoplasm anti-body-associated renal vasculitides. Clin. Exp. Immunol. 82: 238–243.
25. Corrigall, V. M., and G. S. Panayi. 2002. Autoantigens and immune pathways inrheumatoid arthritis. Crit. Rev. Immunol. 22: 281–293.
26. Wang, X.-P., A. B. Fogo, S. Colon, G. Giannico, S. R. Abul-Ezz, J. H. Miner, andD.-B. Borza. 2005. Distinct epitopes for anti-glomerular basement membraneAlport alloantibodies and Goodpasture autoantibodies within the noncollagenousdomain of �3(IV) collagen: a Janus-faced antigen. J. Am. Soc. Nephrol. 16:3563–3571.
27. Henri, S., D. Vremec, A. Kamath, J. Waithman, S. Williams, C. Benoist,K. Burnham, S. Saeland, E. Handman, and K. Shortman. 2001. The dendritic cellpopulations of mouse lymph nodes. J. Immunol. 167: 741–748.
28. Berney, C., S. Herren, C. A. Power, S. Gordon, L. Martinez-Pomares, andM. H. Kosco-Vilbois. 1999. A member of the dendritic cell family that enters Bcell follicles and stimulates primary antibody responses identified by a mannosereceptor fusion protein. J. Exp. Med. 190: 851–860.
29. Mueller, C. G., I. Cremer, P. E. Paulet, S. Niida, N. Maeda, S. Lebeque,W. H. Fridman, and C. Sautes-Fridman. 2001. Mannose receptor ligand-positivecells express the metalloprotease decysin in the B cell follicle. J. Immunol. 167:5052–5060.
30. Yu, P., Y. Wang, R. K. Chin, L. Martinez-Pomares, S. Gordon,M. H. Kosco-Vibois, J. Cyster, and Y. X. Fu. 2002. B cells control the migrationof a subset of dendritic cells into B cell follicles via CXC chemokine ligand 13in a lymphotoxin-dependent fashion. J. Immunol. 168: 5117–5123.
31. Martinez-Pomares, L., J. A. Mahoney, R. Kaposzta, S. A. Linehan, P. D. Stahl,and S. Gordon. 1998. A functional soluble form of the murine mannose receptoris produced by macrophages in vitro and is present in mouse serum. J. Biol.Chem. 273: 23376–23380.
32. MacPherson, G. G., C. D. Jenkins, M. J. Stein, and C. Edwards. 1995. Endotoxin-mediated dendritic cell release from the intestine. Characterization of releaseddendritic cells and TNF dependence. J. Immunol. 154: 1317–1322.
33. Roake, J. A., A. S. Rao, P. J. Morris, C. P. Larsen, D. F. Hankins, andJ. M. Austyn. 1995. Dendritic cell loss from nonlymphoid tissues after systemicadministration of lipopolysaccharide, tumor necrosis factor, and interleukin 1.J. Exp. Med. 181: 2237–2247.
34. De Smedt, T., B. Pajak, E. Muraille, L. Lespagnard, E. Heinen, P. De Baetselier,J. Urbain, O. Leo, and M. Moser. 1996. Regulation of dendritic cell numbers andmaturation by lipopolysaccharide in vivo. J. Exp. Med. 184: 1413–1424.
35. Reis e Sousa, C., S. Hieny, T. Scharton-Kersten, D. Jankovic, H. Charest,R. N. Germain, and A. Sher. 1997. In vivo microbial stimulation induces rapidCD40 ligand-independent production of interleukin 12 by dendritic cells and theirredistribution to T cell areas. J. Exp. Med. 186: 1819–1829.
36. Suzuki, H., B. Wang, G. M. Shivji, P. Toto, P. Amerio, M. A. Tomai,R. L. Miller, and D. N. Sauder. 2000. Imiquimod, a topical immune responsemodifier, induces migration of Langerhans cells. J. Invest. Dermatol. 114:135–141.
37. Doxsee, C. L., T. R. Riter, M. J. Reiter, S. J. Gibson, J. P. Vasilakos, andR. M. Kedl. 2003. The immune response modifier and Toll-like receptor 7 agonistS-27609 selectively induces IL-12 and TNF-� production inCD11c�CD11b�CD8� dendritic cells. J. Immunol. 171: 1156–1163.
38. Turnbull, E. L., U. Yrlid, C. D. Jenkins, and G. G. Macpherson. 2005. Intestinaldendritic cell subsets: differential effects of systemic TLR4 stimulation on mi-gratory fate and activation in vivo. J. Immunol. 174: 1374–1384.
39. Martinez-Pomares, L., D. M. Reid, G. D. Brown, P. R. Taylor, R. J. Stillion,S. A. Linehan, S. Zamze, S. Gordon, and S. Y. Wong. 2003. Analysis of mannosereceptor regulation by IL-4, IL-10, and proteolytic processing using novel mono-clonal antibodies. J. Leukocyte Biol. 73: 604–613.
40. Finkelman, F. D., A. Lees, R. Birnbaum, W. C. Gause, and S. C. Morris. 1996.Dendritic cells can present antigen in vivo in a tolerogenic or immunogenicfashion. J. Immunol. 157: 1406–1414.
41. Carayanniotis, G., and B. H. Barber. 1987. Adjuvant-free IgG responses inducedwith antigen coupled to antibodies against class II MHC. Nature 327: 59–61.
42. Lunde, E., K. H. Western, I. B. Rasmussen, I. Sandlie, and B. Bogen. 2002.Efficient delivery of T cell epitopes to APC by use of MHC class II-specificTroybodies. J. Immunol. 168: 2154–2162.
43. Hawiger, D., K. Inaba, Y. Dorsett, M. Guo, K. Mahnke, M. Rivera, J. V. Ravetch,R. M. Steinman, and M. C. Nussenzweig. 2001. Dendritic cells induce peripheralT cell unresponsiveness under steady state conditions in vivo. J. Exp. Med. 194:769–779.
44. Bonifaz, L., D. Bonnyay, K. Mahnke, M. Rivera, M. C. Nussenzweig, andR. M. Steinman. 2002. Efficient targeting of protein antigen to the dendritic cellreceptor DEC-205 in the steady state leads to antigen presentation on majorhistocompatibility complex class I products and peripheral CD8� T cell toler-ance. J. Exp. Med. 196: 1627–1638.
45. Tacken, P. J., I. J. de Vries, K. Gijzen, B. Joosten, D. Wu, R. P. Rother, S. J. Faas,C. J. Punt, R. Torensma, G. J. Adema, and C. G. Figdor. 2005. Effective inductionof naive and recall T-cell responses by targeting antigen to human dendritic cellsvia a humanized anti-DC-SIGN antibody. Blood 106: 1278–1285.
46. Corbett, A. J., I. Caminschi, B. S. McKenzie, J. L. Brady, M. D. Wright,P. L. Mottram, P. M. Hogarth, A. N. Hodder, Y. Zhan, D. M. Tarlinton, et al.2005. Antigen delivery via two molecules on the CD8� dendritic cell subsetinduces humoral immunity in the absence of conventional “danger”. Eur. J. Im-munol. 35: 2815–2825.
47. Ingulli, E., D. R. Ulman, M. M. Lucido, and M. K. Jenkins. 2002. In situ analysisreveals physical interactions between CD11b� dendritic cells and antigen-spe-cific CD4 T cells after subcutaneous injection of antigen. J. Immunol. 169:2247–2252.
48. Gretz, J. E., C. C. Norbury, A. O. Anderson, A. E. Proudfoot, and S. Shaw. 2000.Lymph-borne chemokines and other low molecular weight molecules reach highendothelial venules via specialized conduits while a functional barrier limits ac-cess to the lymphocyte microenvironments in lymph node cortex. J. Exp. Med.192: 1425–1440.
49. Bonifaz, L. C., D. P. Bonnyay, A. Charalambous, D. I. Darguste, S.-I. Fujii,H. Soares, M. K. Brimnes, B. Moltedo, T. M. Moran, and R. M. Steinman. 2004.In vivo targeting of antigens to maturing dendritic cells via the DEC-205 receptorimproves T cell vaccination. J. Exp. Med. 199: 815–824.
50. Swain, S. D., S. J. Lee, M. C. Nussenzweig, and A. G. Harmsen. 2003. Absenceof the macrophage mannose receptor in mice does not increase susceptibility toPneumocystis carinii infection in vivo. Infect. Immun. 71: 6213–6221.
51. Lee, S. J., N. Y. Zheng, M. Clavijo, and M. C. Nussenzweig. 2003. Normal hostdefense during systemic candidiasis in mannose receptor-deficient mice. Infect.Immun. 71: 437–445.
52. Chieppa, M., G. Bianchi, A. Doni, A. Del Prete, M. Sironi, G. Laskarin, P. Monti,L. Piemonti, A. Biondi, A. Mantovani, et al. 2003. Cross-linking of the mannosereceptor on monocyte-derived dendritic cells activates an anti-inflammatory im-munosuppressive program. J. Immunol. 171: 4552–4560.
53. Taylor, P. R., S. Zamze, R. J. Stillion, S. Y. Wong, S. Gordon, andL. Martinez-Pomares. 2004. Development of a specific system for targeting pro-tein to metallophilic macrophages. Proc. Natl. Acad. Sci. USA 101: 1963–1968.
54. Taylor, P. R., S. Gordon, and L. Martinez-Pomares. 2005. The mannose receptor:linking homeostasis and immunity through sugar recognition. Trends Immunol.26: 104–110.
4983The Journal of Immunology
by guest on July 12, 2015http://w
ww
.jimm
unol.org/D
ownloaded from




















