
Comprehensive Leaf Cell Wall Analysis Using Carbohydrate ...
Upload
independentCategory
view
1download
0
ELSEVIER Journal of Sea Research 42 (1999) 131–146
Carbohydrate secretion by phototrophic communities in tidal sediments
B. de Winder a, N. Staats a, L.J. Stal b,Ł, D.M. Paterson c
a Laboratory for Microbiology, ARISE, University of Amsterdam, Nieuwe Achtergracht 127, 1018 WS Amsterdam, Netherlandsb Netherlands Institute of Ecology–Centre of Estuarine and Coastal Ecology, P.O. Box 140, 4400 AC Yerseke, Netherlands
c SERG, Gatty Marine Laboratory, University of St. Andrews, Fife KY16 8LB, Scotland, UK
Received 8 April 1998; accepted 23 March 1999
Abstract
Two different benthic phototrophic communities on tidal flats were investigated for their carbohydrate content anddistribution. Carbohydrates were analysed as two operationally defined fractions, related to the difficulty of extractionfrom the sediment matrix. Water-soluble (colloidal) and EDTA-extractable (capsular) carbohydrates were measured ina cyanobacterial mat and a diatom biofilm. The chlorophyll-specific carbohydrate content of the two communities wasvery different. The diatom biofilm contained up to 100 times more colloidal carbohydrate than the cyanobacterial mat.The concentrations of colloidal carbohydrates in the diatom biofilm correlated with biomass (chlorophyll-a), but thiswas not the case with the carbohydrate in the EDTA extract. It is proposed that the capsular carbohydrates wereprobably recalcitrant to mineralisation and therefore accumulated in the sediment. Neither colloidal nor EDTA-extractablecarbohydrate in the cyanobacterial mat correlated with chlorophyll-a. This was probably an artefact caused by the factthat approximately 50% of the chlorophyll-a in the mat was attributed to diatoms. The characteristics of extracellularpolysaccharides were investigated in laboratory cultures of the dominant organisms. Extracellular polysaccharides of thecyanobacterium Microcoleus chthonoplastes and of the diatom Navicula menisculus did not contain uronic acids. However,carboxylated sugars were found in large quantities in the capsular polysaccharides of the cyanobacterium and were presentin equal ratios in the extracellular and capsular carbohydrate of the diatom Cylindrotheca closterium. Both in laboratorymodel systems of diatom biofilms and in situ, enhanced colloidal carbohydrate production was observed in the light. Nolight-dependent increase in carbohydrate concentration was found for the cyanobacterial mat. The cyanobacteria formeda mat in which the filamentous organisms entangled sand grains and attached firmly to the substratum. The interparticlespaces were completely occluded by polymers, whereas in the diatom biofilm the organic matrix was less well developedand void spaces could still be discerned. It is conceived that the properties of extracellular polysaccharides influence thestability of the sediment bed. 1999 Elsevier Science B.V. All rights reserved.
Keywords: benthic diatoms; cyanobacteria; microbial mat; EPS; carbohydrate secretion
1. Introduction
Tidal sediment deposits often support abundantpopulations of phototrophic microorganisms (Stal et
Ł Corresponding author. Tel.: C31 113 577 497; Fax: C31 113573616; E-mail: [email protected]
al., 1985; Underwood and Smith, 1998). Such pop-ulations may produce macroscopically recognisablemicrobial mats, often dominated by the cyanobac-terium Microcoleus chthonoplastes or by visiblebiofilms of epipelic diatoms. On cohesive and non-cohesive sediments, biological structures are oftenembedded in a thick mucilaginous matrix of extracel-
1385-1101/99/$ – see front matter 1999 Elsevier Science B.V. All rights reserved.PII: S 1 3 8 5 - 1 1 0 1 ( 9 9 ) 0 0 0 2 1 - 0

132 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
lular polymeric substances (EPS), which may consistof 90% or more of polysaccharides (Hoagland etal., 1993). This EPS has been implicated in theadhesion of microorganisms to sediment particles,thereby influencing the rheology and stability of thesediment bed (Paterson, 1989; Dade et al., 1990; Pa-terson, 1997; Ruddy et al., 1998a). It has also beenshown that polysaccharides are more efficient agentsof sediment–particle aggregation than other organiccomponents of the sediment matrix, for instance hu-mic and fulvic acids (Greenland et al., 1961). In theprocess of matrix formation, the initial step involvesthe secretion of a polymer to bridge the gap betweenthe particles and the organism (Wetherbee et al.,1998). This allows the organism to adhere to the sub-stratum. Scanning electron microscopy has shownthat EPS occurs as a matrix or network of strandsin the interparticle voids of the sediment (Chenu andJaunet, 1992; Chenu, 1993; Defarge et al., 1996).The nature of the polymers visualised in sedimentsis strongly related to the SEM preparation techniqueand related sample dehydration (see Paterson, 1995).The polymers forming the matrix should be con-sidered a diffuse medium that permeates the voidspaces, and coats the particles. This process altersthe size and shape of the grains, changes their poten-tial to adhere to surfaces and affects the geometry ofthe sediment in such a way that organism–particle,particle–particle, and organism–organism aggregatesare formed. The properties of the polymers that de-termine their capacity to adhere to surfaces dependon the involvement of several types of chemicalbonding such as hydrogen bonding between neutralcompounds and metal-bridging between ionogeniccompounds (Stotzky, 1980; Decho, 1994).
Yallop et al. (1994) assumed that the distributionand nature of EPS in the diatom biofilm and thefilamentous nature of the cyanobacterial mat under-lie the differences in sediment stability and responseto physical forcing. Yet LTSEM images from thatstudy also indicate that the infilling of void spaces byorganic material was more extreme in the cyanobac-terial mat.
The present investigation aims to examine theEPS content of cyanobacterial mats in more detailand compare the EPS content of benthic assemblagesdominated by cyanobacteria and diatoms and to in-vestigate EPS production by selected representative
species. Special emphasis will be placed on (1) thesecretion of extracellular carbohydrate polymers, (2)the nature of the secreted EPS, and (3) the influenceof light on the secretion of the EPS by cyanobacteriaand diatoms. The implications that EPS secretionmight have for sediment stability will be discussed.
2. Material and methods
2.1. Study site
The study site was situated on a tidal sandybeach of the Wadden Sea area of the Island ofTexel (De Cocksdorp, 53º0904500N, 4º1203000E), theNetherlands (Fig. 1) (for details, see Yallop et al.,1994). The experimental station, characterised bymats of cyanobacteria, was situated at approximatelymean high-water level. The sediment had an averageparticle size of 257 µm, sorting of 1300 µm and
Fig. 1. Map showing the location of the study site on the islandof Texel, The Netherlands.

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 133
skewness of 995 µm. The water content of thestation was 20:7 š 0:1% (w=w) with a salinity of30 psu. The diatom-biofilm station was situated 150m seawards from the cyanobacterial station. At thislower site, the mean particle size was much smaller(154 µm, sorting of 486 µm, skewness of 1444 µm)and the water content was 23:8 š 0:9% (w=w) witha salinity of 30 psu. At the diatom station, over20% of the sediment particles were smaller than 63µm compared to less than 2% at the cyanobacterialstation. Both stations were situated in an area of agently sloping tidal flat.
2.2. Sampling methodology
2.2.1. Low-temperature scanning electronmicroscopy
Sediments were visualised following the low-temperature scanning electron microscopy (LTSEM)method of Paterson (1995). Sediments were sampledby inserting thin foil planchettes into the surface ofthe sediment. These were withdrawn and frozen insitu in liquid nitrogen (�196ºC). The samples werekept frozen under liquid nitrogen until after examina-tion on the specially adapted stage of a SEM (Oxfordcryo-systems with Joel 35Fc SEM). Excess surfacewater was sublimed into vacuum by heat-etching at atemperature of �90ºC. Sediments were prepared forLTSEM for surface visualisation and by freeze-frac-ture to examine the internal structure of the sedimentinterface region (Paterson, 1995).
2.2.2. Sediment samplingThe microbial mats at the site have been the
subject of investigation since 1991 (Krumbein et al.,1994; Yallop et al., 1994). Cells for the recent culturestudies were obtained in June and July 1995. Mate-rial for LTSEM analysis was obtained in 1991 and1992 (Krumbein et al., 1994; Yallop et al., 1994).Samples for chemical analyses were taken with astainless-steel corer (2.4 cm2). The top cm of thesediment was sampled and sub-sectioned into 1-mmslices. Five randomly taken cores were pooled foreach depth interval to minimise the effects of patch-iness. A time-series was initiated, the first samplesbeing taken when the ebb waters had fallen to a depthof approximately 50 cm above the sediment and fin-ished at tidal re-immersion of the station. Sample
cores were sliced in the field and immediately cooledon ice and subsequently transported to the labora-tory where they were stored in a freezer at �20ºC.Samples obtained during culture experiments in thelaboratory were lyophilised and stored desiccated insealed vials at �20ºC.
2.2.3. Mesocosm experimentsSmall mesocosm experiments were carried out by
incubating sediment cores taken in PVC tubes (innerdiameter 10 cm, height 15 cm) in thermostaticallycontrolled aquariums. Cores from both stations wereallowed to equilibrate overnight in the dark at 15ºCin a simulated natural tidal regime. Subsequently,when the sediment was exposed, randomly chosenparts of the surface of the two cores were kept inthe dark while an equal surface area was incubatedin the light (200 µmol m�2 s�1, provided by a 50-Whalogen lamp). After 6 h of incubation the coreswere sampled as described above.
2.3. Organisms and culture conditions
Axenic strains of the diatoms Navicula menis-culus and Cylindrotheca closterium were grown inbatch culture in Erlenmeyer flasks (250 cm3). Thebottom of each flask was covered with a layer ofacid-washed sea sand (Merck), which provided asubstratum for the benthic diatoms. The cultureswere grown in Kester medium (Kester et al., 1967) at20ºC and under continuous light (white fluorescentlamps, Philips TLE 32=33) at a photon flux densityof 30 µmol m�2 s�1. C. closterium was obtainedfrom H. Peletier (RIKZ, Haren, the Netherlands).This diatom was originally isolated from intertidalsediment of the Wadden Sea. N. menisculus wasisolated from the experimental site. The cyanobac-terium Microcoleus chthonoplastes strain SAG 3192(Collection of Algae of the University of Gottingen,Germany) was also grown in Erlenmeyer flasks (250cm3) as axenic batch cultures in ASN-III medium(Rippka et al., 1979). This cyanobacterium was orig-inally isolated from a Wadden Sea microbial mat.Cultures of M. chthonoplastes were incubated at20ºC and under continuous light (white fluorescentlamps, Philips TLE 32=33) at a photon flux densityof 30 µmol m�2 s�1 in a Gallenkamp (Sanyo-Gal-lenkamp, UK) illuminated orbital shaking incubator.

134 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
2.4. Isolation of carbohydrate fractions
2.4.1. Field materialThe extraction of carbohydrate from intertidal
sediments was performed as follows. Weighedlyophilised sediment samples were incubated witha known amount of distilled H2O (typically 1 cm3
H2O per 500 mg sediment) for 60 min at 30ºC ina water bath. Carbohydrate remaining in the super-natant after centrifugation for 10 min at 2800 g wasreferred to as the sediment colloidal fraction (SCF),and is identical to colloidal carbohydrate obtainedby the method of Underwood et al. (1995). Subse-quently, the remaining pellet was treated with 100mM EDTA (final concentration), pH 4.0 at 20ºC forat least 4 h. The carbohydrates present in the super-natant after further centrifugation for 10 min at 2800g were referred to as the sediment capsular fraction(SCAF).
2.4.2. Culture materialCells and culture medium were analysed after 4
to 6 days when the cells entered the late exponentialphase of growth. Polymeric carbohydrates found inthe supernatant after incubation at 30ºC for 60 minand subsequent centrifugation at 20,000 g at 4ºCfor 15 min are referred to as the culture colloidalfraction (CCF).
Depending on the organisms the remaining pel-lets were treated differently. The pellet of the diatomculture was suspended and incubated in water at70ºC for 30 min. After centrifugation at 20,000 gfor 15 min the polysaccharides remaining in the su-pernatant were referred to as the culture capsularfraction (CCAF). The remaining pellet of Micro-coleus chthonoplastes trichomes was suspended ina 0.1 M potassium phosphate buffer, pH 7.2. Thissuspension was passed three times through a French
Table 1Summary of carbohydrate fraction terminology
Source Operational separation Terminology Indent.
Culture Extracted in 30ºC water Colloidal fraction CCFExtracted in 70ºC water or using French Press a Capsular fraction CCAF
Sediment Extracted in water Colloidal fraction SCFExtracted with EDTA Capsular fraction SCAF
a CCAF from diatoms were extracted with water at 70ºC and from M. chthonoplastes by French Press (see Section 2).
Press (4ºC, 80 MPa) and, subsequently, centrifugedat 20,000 g for 15 min. The supernatant was concen-trated to a volume of about 10 cm3 by evaporationand subsequently treated with α-amylase and amy-loglucosidase to assure the removal of glycogen (DeWinder et al., 1990).
Carbohydrate polymers in all extracted fractionswere precipitated overnight at 4ºC in ice-cold ethanolat a final concentration of 75% (v=v). The precipi-tate was recovered by centrifugation (15 min, 20,000g, 4ºC). Chrysolaminaran, the storage carbohydrateof diatoms, does not precipitate in ethanol. Table 1provides a summary of carbohydrate fraction termi-nology.
2.5. Analyses
2.5.1. Monosaccharide analysisFreeze-dried extracellular polysaccharide (EPS)
fractions of the cultures (CCF and CCAF) were hy-drolysed and methylated in 0.6 M methanol=HCl, andtrimethylsilylate derivatives were prepared (Chaplin,1986). Mannitol was used as an internal standard.Samples were injected in split mode on a fused silicaWCOT column (25 m by 0.32 mm) coated with CP Sil5CB mounted in a Chrompack CP9000 gas chromato-graph (Chrompack, The Netherlands). Oven temper-ature was programmed as follows: 10 min at 140ºC,then increasing, at 3ºC min�1, to a final temperatureof 220ºC for 5 min.
2.5.2. PigmentsFor the characterisation of the phototrophic com-
munity, freeze-dried sediment samples were assayedby HPLC (Van der Staay et al., 1992). All other fieldsamples and culture material were extracted overnightin the dark with dimethylformamide (DMFA). Ex-tinction was measured before and after acidifica-

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 135
tion. Calculations were performed after Lorenzenand Newton Downs (1986). The values for K andR for DMFA were experimentally derived using pu-rified chlorophyll-a and pheophytin from cyanobac-teria and were estimated to be 2.26 and 1.80, respec-tively.
Total carbohydrate was assayed by the phenol-sulphuric acid method using glucose as a reference(Chaplin, 1986). Uronic acids were estimated by themeta-dihydroydiphenyl method (Blumenkrantz andAsboe-Hansen, 1973) using galacturonic acid as areference.
3. Results
3.1. Visualisation of the sediments by LTSEM
Low-temperature Scanning Electron Microscopy(LTSEM) was performed on samples from the sitesto demonstrate the physical nature of the assemblagemicrostructure of the diatom and cyanobacterial mats(Yallop et al., 1994). While culture isolates wereobtained in a different year, these perennial matshave been noted for many seasons and the structureseems stable.
The Microcoleus chthonoplastes mat had clearlydeveloped into a thick matrix at the sediment surface.The surface of the sediment was generally infilledwith few particles visible (Fig. 2A) forming a co-herent layer when the surface was viewed obliquely(Fig. 2B). In areas where the cyanobacterial matwas less developed, particles were more obvious atthe surface (Fig. 2C). A fracture face through thecyanobacterial area confirms the observations of Yal-lop et al. (1994) that the upper interparticle spacesare completely filled and no separate sediment par-ticle could be discerned (Fig. 2D). Lower in themat, particles could be clearly seen and less organicmaterial was evident (Fig. 2E). The bundle sheathsof Microcoleus filaments were arranged in twistedfibres often comprised of >50 individual trichomes.Freeze-fracture analysis of these bundles showedthat the contents were mainly cellular and that EPScoatings were relatively thin (Fig. 2F). Analysis ofthe surface preparation gave little further informationsince the polymer layer was continuous, obscuringdetail of the surface.
The diatom biofilm developed a much less dis-tinct matrix than the cyanobacterial mat assemblages(Fig. 3). Surface views showed dense assemblagesof mainly naviculoid diatoms (Fig. 3A–B), whilefreeze-fracture of the surface indicated that the di-atom biofilm was highly concentrated in the upper100 mm of the sediment. Polymeric material wasclearly visible, but unlike the cyanobacterial assem-blage, the sediments were not fully saturated withpolymer and void spaces were visible (Fig. 3C–D). Some evidence of other autotrophes was found(Fig. 3E–F) comprising isolated filaments of greenalgae (cf. Enteromorpha=Monostroma) and isolatedbundles of cyanobacterial trichomes located beneaththe diatom biofilm (Fig. 3E).
3.2. Depth distribution of pigments
In both communities, the highest concentration ofpigment was found in the layer between 0 and 6mm (Fig. 4). In the diatom biofilm, the maximumpigment concentration was at the sediment surface,decreasing gradually with depth (Fig. 4, bottom).In this community, only pigments characteristic ofdiatoms were found, indicating that they were theonly significant biomass of phototrophic organismspresent. This was confirmed by microscopic exam-ination. In the cyanobacterial mat, pigments wereconcentrated in the top 2 mm (Fig. 4, top). However,this station was shown to contain a mixed assem-blage, since a considerable amount of fucoxanthinand other diatom-specific pigments were found. As-suming a ratio of fucoxanthin to chlorophyll-a of1 : 2, a similar value to that found in the top layersof the diatom biofilm, then 50% of the chlorophyll-apresent in the mat of cyanobacteria originates fromthe diatoms.
In both communities, significant amounts of pig-ment were found at depths below the photic zone.The photic zone is defined as the depth at whichlight intensity is reduced to 1% of incident intensity,which was reached at a depth of 1.2 mm at bothstations, as was measured by fibre-optic light probes(data not shown).
Microscopic examination of the phototrophiccommunity in the diatom biofilm showed abundantpopulations of Navicula spp. (Fig. 3B) and Cylin-drotheca closterium. Both epipelic (mostly navicu-

136 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
loids) and epipsammic diatoms were present. Mats ofcyanobacteria were dominated by Microcoleus chtho-noplastes. It was estimated that this species accounted
for up to 80% of the total biovolume of the cyanobac-teria. Other species that were present belonged to thegenera Oscillatoria, Lyngbya, and Phormidium.

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 137
When the sediment was exposed, the diatoms mi-grated to the surface, which was evidenced by thebrown coloration. In contrast, no such migration orchange in coloration was observed in the mat ofcyanobacteria.
3.3. Extracellular carbohydrates
The diatom-dominated sediment contained con-siderably higher amounts of colloidal carbohydratesthan the sediment from the cyanobacterial station(Table 2). On a sediment dry-weight basis, the di-atom community contained up to 100 times morecolloidal carbohydrate than the cyanobacterial as-semblage. However, when normalised to chloro-phyll-a, as a phototrophic biomass parameter, thedifference was considerably smaller. In this case,the diatom community contained about 10 times asmuch biomass-specific colloidal carbohydrate as thecyanobacteria. The differences in EDTA-extractablecarbohydrates were not so large. The diatom com-munity contained 2 to 3 times as much of thesepolysaccharides. This material was therefore rela-tively more important in the mat of cyanobacteria.
Representatives of the dominant species fromeach phototrophic community were cultured to char-acterise the carbohydrates secreted (Table 3). Thethree organisms that were studied yielded two op-erationally defined types of carbohydrate. The twospecies of diatoms secreted most carbohydrate asthe CCF. This fraction differed in composition be-tween the two species investigated. In Navicula,glucose was quantitatively the most important withsome small amounts of galactose and rhamnose. Thecolloidal carbohydrate of Cylindrotheca was morecomplex in composition. Another difference betweenthe two species was the absence of uronic acids inNavicula. A small amount (5% of the total amount
Fig. 2. Low-temperature scanning electron micrographs of the intertidal sediment dominated by a mixed cyanobacterial=diatomassemblage. (A) Surface view of infilling between surface sediment grains (bar marker 50 µm). (B) An oblique view of the surfaceshowing the confluent nature of the matrix and the lack of protruding grains (bar marker 100 µm). (C) Surface view of an area ofsediment where cyanobacteria were less prevalent. The surface grains are more apparent and the infilling is less complete (bar marker250 µm). (D) A freeze fracture face through the surface region of a cyanobacterial mat. The upper (surface) area is occluded bypolymeric material which becomes less evident with depth (bar marker 250 µm). (E) Deeper in the sediment (>2.5 µm), the organicmaterial produced by the biofilm is no longer present and individual grains are clear (bar marker 250 µm). (F) A fracture face througha Microcoleus bundle. The filaments are closely associated with a thin EPS layer both around the individuals and around the sheath (barmarker 2.5 µm).
of EPS) was more firmly associated with the celland was considered to be capsular (CCAF). In thetwo species, this capsular material was composedof different monomeric sugars. The most remarkabledifference was the absence of uronic acids from theN. menisculus CCAF.
The cyanobacterium also secreted EPS into themedium, but most of the extracellular polysaccha-ride was capsular (CCAF). This material, whichaccounted for 65% of the total EPS, contained avariety of different monomeric sugars. Uronic acidswere found only in the capsular polysaccharides.The colloidal EPS of M. chthonoplastes (CCF) wascomposed virtually exclusively of glucose with smallamounts of galactose.
3.4. Depth distribution of carbohydrates
Fig. 5 depicts the typical depth distributionof colloidal carbohydrate and chlorophyll-a in thecyanobacterial mat. This graph shows that both col-loidal carbohydrate and chlorophyll decreased withdepth but this decrease was more pronounced in thecase of carbohydrate, suggesting that the parameterswere not directly correlated.
The variation of the ratio of colloidal andEDTA-extracted carbohydrates over chlorophyll-awith depth is depicted in Fig. 6. If the ratio isconstant over depth, a close coupling between theproduction of carbohydrates and the biomass of thephototrophic microorganisms may be assumed. Thishypothesis was tested for all data (F test, P < 0:05),Table 4). In the diatom community the colloidal car-bohydrate fraction correlated well with chlorophyll-aat all depths (Fig. 6B). This relationship was shownin four independent samples (Table 4) (Fig. 6B). Inall samples of the diatom community, the EDTA-extractable carbohydrates increased with depth (pos-
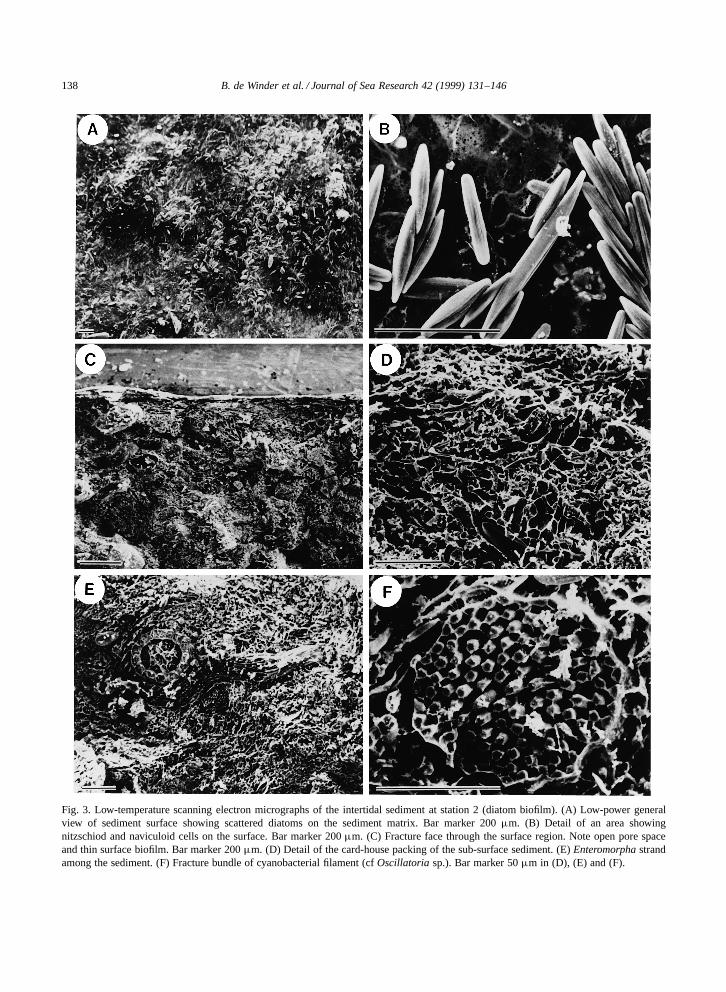
138 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
Fig. 3. Low-temperature scanning electron micrographs of the intertidal sediment at station 2 (diatom biofilm). (A) Low-power generalview of sediment surface showing scattered diatoms on the sediment matrix. Bar marker 200 µm. (B) Detail of an area showingnitzschiod and naviculoid cells on the surface. Bar marker 200 µm. (C) Fracture face through the surface region. Note open pore spaceand thin surface biofilm. Bar marker 200 µm. (D) Detail of the card-house packing of the sub-surface sediment. (E) Enteromorpha strandamong the sediment. (F) Fracture bundle of cyanobacterial filament (cf Oscillatoria sp.). Bar marker 50 µm in (D), (E) and (F).

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 139
Fig. 4. Depth distribution of the most abundant pigments in themat of cyanobacteria (top) and in the diatom biofilm (bottom).The amounts of pigments are expressed in µg per g dry weightof sediment.
itive value in Table 4). In the upper 4 mm of the matof cyanobacteria, the opposite was found (Fig. 6A).The chlorophyll-a-specific amounts of colloidal car-bohydrates and the EDTA-extractable carbohydrateswere both higher in the top layers of the sediment(Fig. 6A) and decreased with depth. This is partic-
Table 2A comparison of carbohydrate content in two fractions (SCF Dsediment colloidal fraction; SCAF D sediment capsular fraction)of tidal benthic phototrophic communities
SCF SCAF SCF=chl-a(mg g�1) (mg g�1)
Cyanobacteria mat 0.1–0.5 0.5–1.2 1–20Diatom film 0.5–50 1.5–2.5 2–200
Numbers indicate the range of values for the top 5 mm .n > 20/.
ularly the case for colloidal carbohydrates (negativevalue in Table 4). For EDTA-extractable carbohy-drate this is less clear.
The colloidal carbohydrate content in the diatombiofilm was found to be highly dynamic (Fig. 7). Dur-ing exposure of the sediment (low tide) to the light,the amount of colloidal carbohydrates increased dra-matically, particularly in the top layer of the sediment.However, the increase of carbohydrates was not onlyrestricted to the euphotic zone. This was evidentlydue to EPS synthesis and=or secretion by the diatomsthat migrate to the surface during exposure. Duringthe subsequent period of inundation most of this car-bohydrate disappeared and the concentration returnedto its initial level (not shown).
3.5. Effect of light on carbohydrate production
The increase in the concentration of SCF carbohy-drate in the diatom biofilm was dependent on light.When a sediment core with a biofilm of diatomswas transferred from the dark to light, a dramaticincrease of colloidal carbohydrate was observed af-ter 6 h of incubation. In the control experiment,kept in the darkness, this increase was not observed(Fig. 8). The same experiment was carried out witha mat of cyanobacteria and no increase in colloidalcarbohydrate was found (Fig. 9) in the light or indarkness.
4. Discussion
4.1. Assemblage structure and composition
Low-temperature scanning electron microscopy isthe technique chosen to preserve the structure ofhydrated samples (Jeffree and Read, 1991). LTSEMvisualisation of the sediment retains structural waterand therefore preserves the natural fabric of the ma-trix, including the EPS polymers of interest in thepresent study (Defarge et al., 1996; Defarge, 1997).The composition of the two phototrophic communi-ties differed markedly. The biofilm of diatoms oc-curred on relatively fine-grained sediment, whereasthe mat of cyanobacteria developed on much coarsersediment. It has been suggested that cyanobacteriacannot cope with the more mobile and dynamic co-

140 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
Table 3Composition of the carbohydrates isolated from cultured organisms
Total a Uronic acid b Monomer composition c
gl ga ar ma rh xy g.a.
CyanobacteriaM. chthonoplastes CCF 35 0 9 1
CCAF 65 100 4 8 1 1 1 6
DiatomsC. closterium CCF 95 50 4 2 1 2 4
CCAF 5 50 4 8 1 1 1 6N. menisculus CCF 95 0 8 1 1
CCAF 5 0 3 2 2 2 1 2
The numbers indicate the percentage of carbohydrate. The sum of colloidal and capsular carbohydrates set as 100%. The monomercomposition is expressed as a relative contribution of the specific compound.a Assayed with the phenol-sulphuric acid method using glucose as a reference (Chaplin, 1986).b Assayed with the meta-dihydroydiphenyl method (Blumenkrantz and Asboe-Hansen, 1973) using galacturonic acid as a reference.c Assayed as trimethylsilylate derivatives using gas chromatography (Chaplin, 1986): gl D glucose; ga D galactose; ar D arabinose; maD mannose; rh D rhamnose; xy D xylose; g.a. D galacturonic acid.
Table 4Correlation of colloidal- (SCF) and EDTA-extractable (SCAF) carbohydrate with chlorophyll-a in vertical profiles in the diatom biofilmand the mat of cyanobacteria
Diatom biofilm Cyanobacteria mat
samples: 1 2 3 4 1 2 3 4 5 6
SCF ns ns ns ns �1.44 �1.64 �1.48 ns �3.34 �1.86SCAF 2.21 4.41 2.44 3.32 ns ns ns ns �3.00 �2.83
The test was carried out for 4 and 6 independent samples in the diatom biofilm and the mat of cyanobacteria, respectively. A positivevalue indicates an increase of carbohydrate with respect to chlorophyll-a with depth and a negative value indicates a decrease. The valueindicates the slope of the line from linear regression carbohydrate over chlorophyll-a with depth. ns indicates a non-significant differencefrom a constant ratio carbohydrate-over-chlorophyll-a at all depths (F test, P < 0:05).
hesive sediments and develop only in the translucent,fine-grained, non-cohesive quartz sediments (Stal,1994). However, it should be noted that the microor-ganisms themselves alter the sediment, including itsgrain size and porosity (Wachendorfer et al., 1994).A critical point is the relative infilling of the voidspaces depicted by LTSEM. The matrix was far morecontinuous and developed in the cyanobacterial matthan in the diatom biofilm where it was much morerestricted and patchy (Figs. 2 and 3). On the basisof the LTSEM images and carbohydrate analysis,it can be concluded that the nature of the mate-rial occluding the void can be identified as colloidalcarbohydrate for the diatom assemblages (SCF) andas capsular carbohydrate for the cyanobacterial as-semblages (SCAF). Examination of fracture surfaces
(e.g. Wachendorfer et al., 1994; Yallop et al., 1994)suggests that the cyanobacterial infilling is moreextensive (deeper) and more complete than that pro-duced by diatom assemblages on their own. Thisconclusion is also supported by the analytical studiesconducted. The diatom biofilm contained much morecolloidal than capsular carbohydrate, while the op-posite was found in the cyanobacterial mat (Table 2).Both microphytobenthic communities formed clearlyrecognisable, self-sustaining structures that are theresult of growth processes rather than of sedimenta-tion. They have been considered analogues to animalor plant tissues by Wachendorfer et al. (1994), whosuggests the term ‘parahistology’ as the appropri-ate technique to study microbial mats and biofilms.Our results confirm the view of Wachendorfer et al.

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 141
Fig. 5. Depth distribution of colloidal carbohydrate (–�–) andchlorophyll-a (–ž–) concentrations in the cyanobacterial mat.Colloidal carbohydrate is the fraction in the supernatant afterwater extraction at 30ºC for 60 min. Amounts are expressed inµg per g dry weight of sediment.
(1994) that EPS polymers produced by the microor-ganisms play a key role for the formation of a matrix,influence the porosity of the sediment, and enhancethe deposition of fresh sediments (Underwood andPaterson, 1993).
4.2. Distribution of carbohydrate in the sediment
The measurement of carbohydrate fractions show-ed variations in their distribution and concentration inthe two assemblages. In the light, epipelic diatoms mi-grate to the surface as soon as the sediment is exposedat low tide (Pinckney and Zingmark, 1991). This wasalso observed in the diatom biofilm investigated in thepresent study as judged from a change in the colourof the sediment surface from grey to brown. Motilityof diatoms is associated with the secretion of polysac-charides (Edgar and Picket-Heaps, 1984). Accumula-tion of polysaccharides in the diatomaceous sedimentwas observed 3 h after exposure (Fig. 7). The major
Fig. 6. Depth distribution of the ratio of colloidal (squares) andEDTA-extractable (circles) carbohydrates to chlorophyll-a in acyanobacterial mat (A) and a diatom biofilm (B).
increase in SCF was in the surface layers althoughnot exclusively. SFC may be transported below theeuphotic depth by diffusion or micro-bioturbation ofthe surface layers by meiofauna such as harpactecoidcopepods and nematodes. Because most of the mi-gration occurred during the first couple of hours afterexposure of the sediment (Paterson, 1989), the accu-mulation of EPS cannot be ascribed exclusively tomigration of the diatoms although cell motility maycontinue on the sediment surface.
The rate of accumulation of colloidal EPS in thesediment samples cannot be due solely to growth ofthe diatoms. In cultures, a maximum growth rate ofthe diatoms of 1 d�1 was measured (Staats, unpubl.observ.). Hence, this would contribute only littleto the five-fold increase in carbohydrate that wasobserved during the 6-h period of exposure (Fig. 7).
It is likely that other factors were responsiblefor the increase in EPS. For example, the massiveaccumulation of diatoms at the sediment surface at

142 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
Fig. 7. Depth distribution and time series of contents (mg perg sediment) of colloidal carbohydrates in the top 1 cm of thesediment with a diatom biofilm during a 7-h period of low tide.The first measurement was done at time 14.00 h, which was 30min before the sediment was exposed. The light period lasted till23.00 h.
low tide would quickly deplete nutrients and growthwould become nutrient-limited (Kromkamp et al.,1995). It has been shown that CO2 can be limitingunder high light intensity for dense diatom biofilms(Lorenzen, 1996). Nutrient limitation in phototrophicorganisms often results in the over-production ofcarbohydrate caused by unbalanced growth (Ortega-Calvo and Stal, 1994). Ruddy et al. (1998a,b) reportnutrient mass balances from field data of benthicdiatoms that also support this hypothesis, and theyhave developed a model that demonstrates that nu-trient limitation in diatom biofilms is very likelyto happen. In fact, the energy dissipation that isachieved by secretion of carbohydrate may servean important function, for instance to protect thediatoms from photo-oxidative damage.
The cyanobacterial mat contained much less car-bohydrate and did not show such dynamic changes.
An explanation for this could be that nutrient limita-tion is less likely, because these mats are known tofix nitrogen (Stal et al., 1984).
The depth variations of the ratios of carbohy-drates to chlorophyll-a in the diatom biofilm and thecyanobacterial mat were notably different (Fig. 6). Inthe diatom biofilm the ratio of colloidal carbohydrateto chlorophyll-a was constant over depth. This is inagreement with the observations of Underwood andSmith (1998), who demonstrate that EPS concentra-tions in intertidal mudflats can be predicted from sed-iment chlorophyll-a. The ratio of EDTA-extractablecarbohydrate to chlorophyll-a in the diatom biofilmincreased with depth. It is possible that this carbo-hydrate is more recalcitrant to mineralisation and ac-cumulates to some extent. In the upper 4 mm of thecyanobacterial mat, a strong decrease of colloidal andEDTA-extractable carbohydrate to chlorophyll-a ratiooccurred. Below 4 mm these carbohydrate fractionsbehaved as in the diatom biofilm. This complex be-haviour of carbohydrate in the cyanobacterial mat canbe explained by the high diatom biomass. On the basisof pigment ratios it can be calculated that about 50%of the chlorophyll-a could be attributed to diatoms.However, the diatoms were relatively more abundantin the surface layers (Fig. 4). This and the much higherchlorophyll-specific carbohydrate content of diatombiofilm (Table 2) would explain the pattern observed.Below 4 mm depth in the cyanobacterial mat the ratioof carbohydrate to chlorophyll-a is largely attributedto cyanobacteria. Hence, as in diatoms, the ratio ofcolloidal carbohydrate to chlorophyll-a in cyanobac-teria seems to be constant, although considerablylower (Table 2). A contributory factor in EPS distri-bution may be the changes in sediment structure withdepth (Taylor and Paterson, 1998). As the bulk den-sity of the sediment increases with depth there is lesspore water and hence less colloidal material can accu-mulate. In addition, bulk density of the sediment alsocontrols light availability, which is particularly impor-tant for EPS dynamics in diatom-dominated systems(Figs. 7 and 8).
4.3. Consequences of carbohydrate dynamics forsediment stability
The importance of biology in determining the re-sponse of sediments to physical forcing is being in-

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 143
Fig. 8. Vertical profiles of the changes in the content of colloidal carbohydrates in the diatom biofilm (A) after incubation for 6 h inthe light (200 µmol m�2 s�1), and (B) in the dark (control), compared to the initial situation obtained after one night in the dark. Thesediments were subject to an artificial tidal cycle in a thermostat aquarium at 15ºC. During the 6-h light period the sediment surface wasexposed (low tide).
Fig. 9. Vertical profiles of the changes in the content of colloidal carbohydrates in the mat of cyanobacteria (A) after incubation for 6 hin the light (200 µmol m�2 s�1), and (B) in the dark. Further details as in Fig. 8.
creasingly recognised (Wachendorfer et al., 1994; Pa-terson, 1997). The presence of benthic communitiesof phototrophic microorganisms is known to cause
an increase in the mechanical stability of the sedi-ment (Neuman et al., 1970; Grant and Gust, 1987). Ithas been suggested that extracellular polysaccharides

144 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
are responsible for adhesion and aggregate formationand for the increase in the critical threshold for ero-sion (Paterson, 1989; Dade et al., 1990; Underwoodand Paterson, 1993). Also factors that influence bio-logical processes have been shown to alter sedimentstability. For example it has been shown that light in-creases the erosion threshold value for tidal sediments(Paulic et al., 1986; Madsen et al., 1993). These ob-servations strongly suggest that benthic diatoms havea significant impact on the resistance to erosion andthe stability of tidal sediments. In other studies, it hasbeen shown that the colloidal fraction of EPS corre-lates well with the biovolume or biomass of epipelicdiatoms (Underwood and Paterson, 1993; Underwoodand Smith, 1998; Taylor and Paterson, 1998).
As shown in this study, a considerable proportionof the colloidal carbohydrates produced by the di-atoms (SCF) are soluble in water and therefore maydisappear as soon as the tide comes in. The colloidalcarbohydrates may therefore contribute only to a lim-ited extent to sediment stability. However, in shallowwater the maximum tidal-flow-induced stress experi-enced by a mudflat bed occurs shortly after immersionin the flood phase of the cycle (Christie and Dyer,1998). Thus under non-wave-dominated conditions,the sediment may still be protected by colloidal EPS.In addition to the colloidal carbohydrates (SCF), theEDTA-extractable fraction (SCAF) may have an im-portant role that has not been fully recognised. Thesecarbohydrates contain negatively charged groups andalthough uronic acids were apparently absent in Nav-icula menisculus, preliminary evidence suggests thatthis EPS contains sulphated sugars (Staats, unpubl.observ.). It seems reasonable to assume that the EDTAfractions play a prominent role in the formation ofsediment–sediment and sediment–organism interac-tions. Sutherland (1980) and Geddie and Sutherland(1993) have demonstrated the co-precipitation andgel formation of seawater polysaccharides with metalions, and a small amount of this carbohydrate gaverise to very strong bonding, especially when they be-came irreversibly denatured and dehydrated (Foster,1981). The operational separation between SCF andSCAF has its limitations. The change of polymers be-tween the gel and sol states may be governed by theavailability of metal ions and=or desiccation history(Decho, 1994) and therefore have a relatively largeinfluence on sediment stability.
Acknowledgements
This investigation was supported by the Nether-lands Organisation for Scientific Research (NWO)(BOA research theme ‘The morpho-dynamic andbio-dynamic behaviour of mud in tidal areas’) and bythe European Commission Environment and ClimateProgramme contract EV5V-CT94-0411 (Pro-Mat)and Marine Science and Technology Programme,contract MAS3-CT95-0022 (INTRMUD). The au-thors would also like to thank Mrs. N. Dijkman foranalytical help, Dr. J. Huisman for help on statistics,and Dr. C. George for LTSEM analysis (Fig. 3). Thisis Publication 2517 of the Centre of Estuarine andCoastal Research, Yerseke, The Netherlands.
References
Blumenkrantz, N., Asboe-Hansen, G., 1973. New method forquantitative determination of uronic acids. Anal. Biochem. 54,484–489.
Chaplin, M.F., 1986. Monosaccharides. In: Chaplin, M.F.,Kennedy, J.F. (Eds.), Carbohydrate Analysis, a Practical Ap-proach. IRL Press, Oxford, pp. 1–36.
Chenu, C., 1993. Clay- or sand-polysaccharide associations asmodels for the interface between micro-organisms and soil:water related properties and microstructure. Geoderma 56,143–156.
Chenu, C., Jaunet, M., 1992. Cryoscanning electron microscopyof microbial extracellular polysaccharides and their associationwith minerals. Scanning 14, 360–364.
Christie, M.C., Dyer, K., 1998. Measurements of the turbid tidaledge over the Skeffling mudflats. In: Black, K.S., Paterson,D.M., Cramp, A. (Eds.), Sedimentary Processes in the Inter-tidal Zone. Geol. Soc. London, Spec. Publ. 139, 45–55.
Dade, W.B., Davis, J.D., Nichols, P.D., Nowell, A.R.M., Thistle,D., Trexler, M.B., White, D.C., 1990. Effects of bacterial ex-opolymer adhesion on the entrapment of sand. Geomicrobiol.J. 8, 1–16.
Decho, A.W., 1994. Molecular-scale events influencing themacro-scale cohesiveness of exopolymers. In: Krumbein,W.E., Paterson, D.M., Stal, L.J. (Eds.), Biostabilization ofSediment. BIS Verlag, Oldenburg, pp. 135–148.
Defarge, C., 1997. Apports du cryo-microscope electronique abalayage et du microscope electronique a balayage haute re-solution a l’etude des matieres organiques et des relationsorgano-minerales naturelles. Exemple des sediments microbi-ens actuels. C.R. Acad. Sci. Paris 324, Ser. IIa, 553–561.
Defarge, C., Trichet, J., Jaunet, A., Robert, M., Tribble, J.,Sansone, F.J., 1996. Texture of microbial sediments revealedby cryo-scanning electron microscopy. J. Sediment. Res. 66,935–947.
De Winder, B., Stal, L.J., Mur, L.R., 1990. Crinalium epipsam-

B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146 145
mum sp. nov.: a filamentous cyanobacterium with trichomescomposed of elliptical cells and containing poly-β-(1,4) glucan(cellulose). J. Gen. Microbiol. 136, 1645–1653.
Edgar, L.A., Picket-Heaps, J.D., 1984. Diatom locomotion.Progr. Phycol. Res. 3, 47–88.
Foster, R.C., 1981. Polysaccharides in soil fabrics. Science 214,665–667.
Geddie, J.L., Sutherland, I.W., 1993. Uptake of metals by bacte-rial polysaccharides. J. Appl. Bacteriol. 74, 467–472.
Grant, J., Gust, G., 1987. Prediction of coastal sediment stabilityfrom photopigment content of mats of purple sulphur bacteria.Nature 330, 244–246.
Greenland, D.J., Lindstrom, G.R., Quirk, J.P., 1961. Role ofpolysaccharides in stabilisation of natural soil aggregates. Na-ture 191, 1283–1284.
Hoagland, K.D., Rosowski, J.R., Gretz, M.R., Roemer, S.C.,1993. Diatom extracellular polymeric substances: function,fine structure, chemistry, and physiology. J. Phycol. 29, 537–566.
Jeffree, C.E., Read, N.D., 1991. Ambient- and low-temperaturescanning electron microscopy. In: Hall, J.L., Hawes, C. (Eds.),Electron Microscopy of Plant Cells. Academic Press, London,pp. 313–413.
Kester, D.R., Duedall, I.W., Connors, N., Pytkowics, R.M., 1967.Preparation of artificial seawater. Limnol. Oceanogr 12, 176–179.
Kromkamp, J., Peene, J., Van Rijswijk, P., Sandee, A., Goosen,N., 1995. Nutrients, light and primary production by phy-toplankton and microphytobenthos in the eutrophic, turbidWesterschelde estuary (The Netherlands). Hydrobiologia 311,9–19.
Krumbein, W.E., Paterson, D.M., Stal, L.J. (Eds.), 1994. Biosta-bilization of Sediment. BIS Verlag, Oldenburg.
Lorenzen, C.J., Newton Downs, J., 1986. The specific absorptioncoefficients of chlorophyllide a and pheophorbide a in 90%acetone and comments on the fluorometric determinations ofchlorophyll and pheopigments. Limnol. Oceanogr. 31, 449–456.
Lorenzen, J., 1996. Begrænsende Faktorer for MikrofytobentiskFotosyntese. Unpublished PhD thesis, University of Aarhus.
Madsen, K.N., Nilsson, P., Sundback, K., 1993. The influence ofbenthic microalgae on the stability of a subtidal sediment. J.Exp. Mar. Biol. Ecol. 170, 159–177.
Neuman, A.C., Gebelein, C.D., Scoffin, T.P., 1970. The com-position, structure and erodibility of subtidal mats, Abaco,Bahamas. J. Sediment. Petrol. 40, 274–297.
Ortega-Calvo, J.J., Stal, L.J., 1994. Sulphate-limited growth inthe N2-fixing unicellular cyanobacterium Gloeothece (Nageli)sp PCC 6909. New Phytol. 128, 273–281.
Paterson, D.M., 1989. Short-term changes in the erodibility of in-tertidal cohesive sediments related to the migratory behaviourof epipelic diatoms. Limnol. Oceanogr. 34, 223–234.
Paterson, D.M., 1995. The biogenic structure of early sedimentfabric visualised by low-temperature scanning electron mi-croscopy. J. Geol. Soc. 152, 131–140.
Paterson, D.M., 1997. Biological mediation of sediment erodibil-ity: ecology and physical dynamics. In: Burt, N., Parker, R.,
Watts, J. (Eds.), Cohesive Sediments. Wiley, Chichester, pp.215–229.
Paulic, M., Montague, C.L., Mehta, A.J., 1986. Influence of lighton sediment erodibility. In: Shen, H.W. (Ed.), Third Int. Symp.River Sedimentation, Mississippi, pp. 1758–1764.
Pinckney, J., Zingmark, R.G., 1991. Effects of tidal stage andsun angles on intertidal benthic microalgal productivity. Mar.Ecol. Prog. Ser. 76, 81–89.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M., Stanier,R.Y., 1979. Generic assignments strain histories and propertiesof pure cultures of cyanobacteria. J. Gen. Microbiol. 111, 1–61.
Ruddy, G., Turley, C.M., Jones, T.E.R., 1998a. Ecological inter-action and sediment transport on an intertidal mudflat, I. Ev-idence for a biologically mediated sediment–water interface.In: Black, K.S., Paterson, D.M., Cramp, A. (Eds.), Sedimen-tary Processes in the Intertidal Zone. Geol. Soc. Spec. Publ.139, 135–148.
Ruddy, G., Turley, C.M., Jones, T.E.R., 1998b. Ecological inter-action and sediment transport on an intertidal mudflat, II. Anexperimental dynamic model of the sediment–water interface.In: Black, K.S., Paterson, D.M., Cramp, A. (Eds.), Sedimen-tary Processes in the Intertidal Zone. Geol. Soc. Spec. Publ.139, 149–166.
Stal, L.J., 1994. Microbial mats in coastal environments. In:Stal, L.J., Caumette, P. (Eds.), Microbial Mats. Structure,Development and Environmental Significance. NATO ASI Ser.G35, Springer, Heidelberg, pp. 21–32.
Stal, L.J., Grossberger, S., Krumbein, W.E., 1984. Nitrogen fix-ation associated with the cyanobacterial mat of a marinelaminated microbial ecosystem. Mar. Biol. 82, 217–224.
Stal, L.J., Van Gemerden, H., Krumbein, W.E., 1985. Structureand development of a benthic marine microbial mat. FEMSMicrobiol. Ecol. 31, 111–125.
Stotzky, G., 1980. Surface interactions between clay mineralsand microbes, viruses and soluble organics, and the probableimportance of these interactions to the ecology of microbesin soil. In: Berkeley, R.C.W., Lynch, J.M., Melling, J., Rutter,P.R., Vincent, B. (Eds.), Microbial Adhesion to Surfaces. EllisHorwood, Chichester, pp. 231–247.
Sutherland, I.W., 1980. Polysaccharides in the adhesion of ma-rine and freshwater bacteria. In: Berkeley, R.C.W., Lynch,J.M., Melling, J., Rutter, P.R., Vincent, B. (Eds.), MicrobialAdhesion to Surfaces. Ellis Horwood, Chichester, pp. 329–338.
Taylor, I.S., Paterson, D.M., 1998. Microspatial variation in car-bohydrate concentrations with depth in the upper millimetresof intertidal cohesive sediments. Estuarine Coastal Shelf Sci.46, 359–370.
Underwood, G.J.C., Paterson, D.M., 1993. Seasonal changes indiatom biomass, sediment stability and biogenic stabilisationin the Severn estuary. J. Mar. Biol. Assoc. UK 73, 871–887.
Underwood, G.J.C., Smith, D.J., 1998. Predicting epipelic di-atom exopolymer concentrations in intertidal sediments fromsediment chlorophyll a. Microbiol. Ecol. 35, 116–125.
Underwood, G.J.C., Paterson, D.M., Parkes, R.J., 1995. The

146 B. de Winder et al. / Journal of Sea Research 42 (1999) 131–146
measurement of microbial carbohydrate exopolymers fromintertidal sediments. Limnol. Oceanogr. 40, 1243–1253.
Van der Staay, G.W.M., Brouwer, A., Baard, R.L., Van Mourik,F., Matthijs, H.C.P., 1992. Separation of photosystems I and IIfrom the oxychlorobacterium (prochlorophyte) Prochlorotrixhollandica and association of chlorophyll b binding antennaewith photosystem II. Biochim. Biophys. Acta 1102, 220–228.
Wachendorfer, V., Krumbein, W.E., Schellnhuber, H.J., 1994.Bacteriogenic porosity of marine sediments — a case ofbiomorphogenesis of sedimentary rocks. In: Krumbein, W.E.,
Paterson, D.M., Stal, L.J. (Eds.), Biostabilization of Sediment.BIS Verlag, Oldenburg, pp. 203–220.
Wetherbee, R., Lind, J.L., Burke, J., Quatrano, R.S., 1998. Thefirst kiss: establishment and control of initial cohesion byraphid diatoms. J. Phycol. 34, 9–15.
Yallop, M.L., De Winder, B., Paterson, D.M., Stal, L.J., 1994.Comparative structure, primary production and biogenic sta-bilisation of cohesive and non-cohesive marine sediments in-habited by microphytobenthos. Estuarine Coastal Shelf Sci.39, 565–582.
Copyright © 2022 FDOKUMEN




















