
Multipathways for transdifferentiation of human prostate cancer cells into neuroendocrine-like...
16
Multipathways for transdi¡erentiation of human prostate cancer cells into neuroendocrine-like phenotype Stanislav Zelivianski a , Michael Verni b , Carissa Moore a , Dmitriy Kondrikov a , Rodney Taylor b;c;d , Ming-Fong Lin a;b;c;d; * a Department of Biochemistry/Molecular Biology, University of Nebraska Medical Center, 984525 Nebraska Medical Center, Omaha, NE 68198, USA b Section of Urologic Surgery, College of Medicine, Omaha, NE 68198, USA c UNMC Eppley Cancer Center, University of Nebraska Medical Center, Omaha, NE 68198, USA d Omaha VA Hospital, Omaha, NE 68198, USA Received 5 October 2000; received in revised form 15 February 2001; accepted 21 March 2001 Abstract The neuroendocrine (NE) cell is a minor cell population in normal human prostate glands. The number of NE cells is increased in advanced hormone-refractory prostate carcinomas (PCA). The mechanism of increased NE cell population in these advanced tumors is poorly understood. We examined molecular mechanisms which may be involved in the regulation of the transdifferentiation process of human PCA cells leading to a NE phenotype. We compared PCA cell lines LNCaP and PC-3 in the following medium conditions : steroid-reduced (SR), interleukin-6 (IL-6)-supplemented, or dibutyrate cAMP (db- cAMP)-supplemented. We found that androgen-responsive C-33 LNCaP cells responded to all treatments, having a neuronal-like morphology. In contrast, C-81 LNCaP cells, having a decreased androgen responsiveness, had a less pronounced effect although followed a similar trend. Androgen-unresponsive PC-3 cells showed little change in their morphology. Grown in the SR condition, the level of neuron-specific enolase (NSE), a marker of neuronal cells, was upregulated in C-33 LNCaP cells, while to a lesser degree in the presence of IL-6. In the presence of db-cAMP, the NSE level in C-33 cells was decreased, lower than that in control cells. An opposite effect was observed for C-81 LNCaP cells. Nevertheless, the NSE level was only elevated in db-cAMP-treated PC-3 cells, but no change was found in PC-3 cells grown in the SR- or IL-6-supplemented medium. Thus, a similar gross phenotypic change may correlate with differential molecular expressions. We also analyzed the expression of protein tyrosine phosphatase K (RPTPK) since it plays a critical role in normal neuronal differentiation and signaling. Our results showed that the expression of RPTPK correlates with the NE phenotypic change of LNCaP cells in the SR condition. In summary, our data clearly show that the molecular process by which cultured human prostate cancer cells undergo a transdifferentiation process to a NE cell-like phenotype is accompanied by differential expressions of different markers, and a gross NE cell-like phenotype can occur by exposing PCA cells to different pharmacological agents. ß 2001 Elsevier Science B.V. All rights reserved. Keywords : Cellular transdi¡erentiation ; Prostate cancer ; Neuroendocrine cell ; Neuron-speci¢c enolase ; Receptor protein tyrosine phos- phatase K 1. Introduction Prostate cancer is the most common malignancy in 0167-4889 / 01 / $ ^ see front matter ß 2001 Elsevier Science B.V. All rights reserved. PII:S0167-4889(01)00087-8 * Corresponding author, at address a. Fax: 402-559-6650; E-mail : [email protected] Biochimica et Biophysica Acta 1539 (2001) 28^43 www.bba-direct.com
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Multipathways for transdifferentiation of human prostate cancer cells into neuroendocrine-like...

Multipathways for transdi¡erentiation of human prostate cancer cellsinto neuroendocrine-like phenotype
Stanislav Zelivianski a, Michael Verni b, Carissa Moore a, Dmitriy Kondrikov a,Rodney Taylor b;c;d, Ming-Fong Lin a;b;c;d;*
a Department of Biochemistry/Molecular Biology, University of Nebraska Medical Center, 984525 Nebraska Medical Center,Omaha, NE 68198, USA
b Section of Urologic Surgery, College of Medicine, Omaha, NE 68198, USAc UNMC Eppley Cancer Center, University of Nebraska Medical Center, Omaha, NE 68198, USA
d Omaha VA Hospital, Omaha, NE 68198, USA
Received 5 October 2000; received in revised form 15 February 2001; accepted 21 March 2001
Abstract
The neuroendocrine (NE) cell is a minor cell population in normal human prostate glands. The number of NE cells isincreased in advanced hormone-refractory prostate carcinomas (PCA). The mechanism of increased NE cell population inthese advanced tumors is poorly understood. We examined molecular mechanisms which may be involved in the regulationof the transdifferentiation process of human PCA cells leading to a NE phenotype. We compared PCA cell lines LNCaP andPC-3 in the following medium conditions: steroid-reduced (SR), interleukin-6 (IL-6)-supplemented, or dibutyrate cAMP (db-cAMP)-supplemented. We found that androgen-responsive C-33 LNCaP cells responded to all treatments, having aneuronal-like morphology. In contrast, C-81 LNCaP cells, having a decreased androgen responsiveness, had a lesspronounced effect although followed a similar trend. Androgen-unresponsive PC-3 cells showed little change in theirmorphology. Grown in the SR condition, the level of neuron-specific enolase (NSE), a marker of neuronal cells, wasupregulated in C-33 LNCaP cells, while to a lesser degree in the presence of IL-6. In the presence of db-cAMP, the NSE levelin C-33 cells was decreased, lower than that in control cells. An opposite effect was observed for C-81 LNCaP cells.Nevertheless, the NSE level was only elevated in db-cAMP-treated PC-3 cells, but no change was found in PC-3 cells grownin the SR- or IL-6-supplemented medium. Thus, a similar gross phenotypic change may correlate with differential molecularexpressions. We also analyzed the expression of protein tyrosine phosphatase K (RPTPK) since it plays a critical role innormal neuronal differentiation and signaling. Our results showed that the expression of RPTPK correlates with the NEphenotypic change of LNCaP cells in the SR condition. In summary, our data clearly show that the molecular process bywhich cultured human prostate cancer cells undergo a transdifferentiation process to a NE cell-like phenotype isaccompanied by differential expressions of different markers, and a gross NE cell-like phenotype can occur by exposing PCAcells to different pharmacological agents. ß 2001 Elsevier Science B.V. All rights reserved.
Keywords: Cellular transdi¡erentiation; Prostate cancer; Neuroendocrine cell ; Neuron-speci¢c enolase; Receptor protein tyrosine phos-phatase K
1. Introduction
Prostate cancer is the most common malignancy in
0167-4889 / 01 / $ ^ see front matter ß 2001 Elsevier Science B.V. All rights reserved.PII: S 0 1 6 7 - 4 8 8 9 ( 0 1 ) 0 0 0 8 7 - 8
* Corresponding author, at address a. Fax: 402-559-6650;E-mail : [email protected]
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
Biochimica et Biophysica Acta 1539 (2001) 28^43www.bba-direct.com

men, and the second leading cause of cancer-relatedmortality in US males [49]. Although androgen abla-tion is the most e¡ective therapy for patients withadvanced prostate cancer, progression of cancer cellsto androgen independence evidently occurs and re-mains the main obstacle to improve the survival andquality of life [21]. The tumor progression to andro-gen-refractory stage is a complex process that may beinvolving both clonal selection and adaptive mecha-nisms in heterogeneous tumors composed of cellsthat respond di¡erently to androgen ablation. Thepresent research is focused on understanding the mo-lecular mechanisms that allow human prostate can-cer cells to acquire androgen independence.
The cells in human prostate gland include basalepithelial, secretory epithelial, and neuroendocrine(NE) cells, in addition to stromal cells [7,23]. TheNE or endocrine^paracrine cells, although only rep-resenting a minor cell population in normal prostateglands, are known to produce and secrete some po-tent neuro-hormones, including bombesin/gastrin,calcitonin, and parathyroid hormone-related peptide,that likely regulate the activity and homeostasis ofthe adjacent basal and/or secretory epithelial cells[31]. NE cells may represent one potential pathwayof di¡erentiation of prostatic stem cells. Recently,results of several clinical studies suggested that NEcells may also have a role in the progression of pros-tate cancer cells to the hormone-independent state[22,24,25]. Histological studies reveal that NE cellsare signi¢cantly more abundant in cancerous tissuesthan in non-cancerous tissues [32]. In fact, the overallabundance of NE cells is increased in proportion tothe progression of prostate cancer from the earlystage to the advanced, hormone-resistant state [61].Results from analyses on sera from hormone-depen-dent and clinically advanced, hormone-refractoryprostate cancer patients show an increased level ofchromogranin A [1,37] and neuron-speci¢c enolase(NSE) [58], markers for NE cells, in advanced stagepatients. These observations have led to the hypoth-esis that NE cells function to provide paracrine stim-ulatory factors in an androgen-deprived condition,leading to an accelerated growth and progression ofsurrounding prostate cancer cells toward an andro-gen-independent state. This hypothesis is supportedby the observation of an increased proliferation in-dex of tumor cells in the close proximity to the NE
cell foci [9]. Thus, NE cells can apparently modulatethe proliferation of adjacent epithelial cells includingcancerous and non-cancerous cells.
Although the origin of the NE cell population inprostate cancer tissues remains uncertain, a numberof recent studies has shown that prostate cancer cellsthemselves have the ability to `transdi¡erentiate' (orconvert) into cells with a NE-like phenotype in cul-ture. This phenomenon can occur when culturedprostate cancer cells are exposed to pharmacologicalagents that increase the intracellular level of cyclicAMP (cAMP) [4,16], in the presence of interleukin-6 (IL-6) [47,51], or in a steroid-reduced medium[10,55]. These treatments induce a morphologicalchange in cell phenotype, resembling cultured neuro-nal cells with narrow cell bodies and elongated pro-cesses. However, no systematic study has been per-formed to analyze the susceptibility of each cell lineto di¡erent treatments that relate to clinical phenom-ena. The molecular mechanism of transdi¡erentia-tion also remains to be further delineated.
Protein tyrosine phosphorylation plays a criticalrole in regulating various cellular events, includingcell di¡erentiation and proliferation. Protein tyrosinekinases (PTKs) and protein tyrosine phosphatases(PTPs) function dynamically to maintain cellular ho-meostasis. Unbalanced tyrosine phosphorylation canalter signaling for cell growth and di¡erentiation,leading to neoplastic diseases or cell growth arrest.Protein tyrosine phosphatase K (RPTPK), alsoknown as leukocyte common antigen-related phos-phatase (LRP), is a transmembrane member of thePTP family [39,40,54]. It has been demonstrated thatRPTPK can be involved in GRB2-mediated signaling[19,57], and participate in activation of mitogen-acti-vated protein kinases (MAPKs) and c-Jun transcrip-tion [66]. Additionally, RPTPK can regulate the spe-ci¢c activity of Src kinase by dephosphorylating itscarboxyl-terminal tyrosine [20,67]. Interestingly,RPTPK has also been determined to play a criticalrole in normal neuronal di¡erentiation [60,62]. Thus,RPTPK is involved in multisignaling pathways. Wehave reported that RPTPK mRNA is expressed innormal and cancerous prostate cells [64]. However,its functional role in those cells remains to be clari-¢ed.
It has been established that modi¢cation of nucle-osome structure is an important regulatory mecha-
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 29

nism of gene expression. For example, butyrate is anon-competitive inhibitor of histone deacetylase, andthus provokes a hyperacetylation level of core his-tones [44]. The hyperacetylation of histones by buty-rate induces a more open state of chromatin, whichleads to the activation of expression of numerousgenes and cellular di¡erentiation [12], similar tothat by trichostatin A (TSA), a potent speci¢c inhib-itor of histone deacetylase [63]. Butyrate treatmentcan also increase the phosphorylation of MAPK,i.e., ERK1/2 [27], and induce di¡erentiation inmany cell types [15,29,53]. Interestingly, some tran-scriptional e¡ects by butyrate can be abolished byinhibitors of protein phosphatases [17]. Thus, themolecular mechanism of butyrate in cellular di¡er-entiation involving phosphorylation signaling re-mains to be delineated.
In an e¡ort to understand how NE cells progres-sively emerge in prostate malignancies and their re-lationship to hormone-refractory prostate carcino-mas, we studied the pharmacological e¡ect ontransdi¡erentiation of two human prostate cancercell lines, LNCaP and PC-3. Although LNCaP pa-rental cells, i.e. C-33 LNCaP cells, have been studiedwith regard to their growth patterns in response tovarious culture conditions, less is known about trans-di¡erentiation of PC-3 cells nor is their transdi¡er-entiation related to prostate cancer progression. Wealso examined the transdi¡erentiation process of C-81 LNCaP cells, a subline of LNCaP cells which lackandrogen responsiveness of growth stimulation, de-spite the expression of a functional AR. We investi-gated the biochemical parameters of transdi¡erentia-tion, and examined the e¡ect of butyrate ontransdi¡erentiation of LNCaP cells. Our results sug-gest that di¡erent transdi¡erentiation processes areinvolved in the accumulation of NE cells in advancedhormone-refractory human prostate cancers.
2. Materials and methods
2.1. Materials
Fetal bovine serum (FBS), cell culture mediumRPMI 1640, gentamicin, horseradish peroxidase-con-jugated anti-mouse and anti-rabbit immunoglobulins(IgGs) were purchased from Life Technologies
(Grand Island, NY). Charcoal/Dextran-treated, cer-ti¢ed FBS with testosterone concentration below3 ng/dl was from HyClone (Logan, UT). Proteinmolecular weight standard markers, acrylamide,and the protein assay kit were obtained from Bio-Rad (Hercules, CA). Sodium butyrate, trichostatinA, dibutyrate cAMP (db-cAMP), and L-actin-speci¢cantibody were from Sigma (St. Louis, MO). IL-6 andNSE-speci¢c antibodies were purchased from Up-state Biotechnology (Lake Placid, NY). RPTPK-spe-ci¢c antibody was obtained from TransductionLaboratories (San Diego, CA). Gastrin (GS), chro-mogranin A (ChA), and neurotensine (NT)-speci¢cantibody and ABC Staining System were purchasedfrom Santa Cruz Biotechnology (Santa Cruz, CA).All other reagents were obtained as described in pre-vious publications [30,42].
2.2. Methods
2.2.1. Cell cultureCommercially available human prostate carcinoma
cell lines, LNCaP and PC-3, were originally obtainedfrom the American Type Culture Collection, androutinely maintained in phenol red (PR)-containingRPMI 1640 medium supplemented with 5% FBS, 1%glutamine, and 0.5% gentamicin, as described in pre-vious publications [30,42]. LNCaP cells are andro-gen-responsive and express PSA, while PC-3 cellsare androgen-insensitive and lack the expression ofPSA [38,42]. For experiments, LNCaP cells that hadpassage numbers less than 33 were designated as C-33, passage numbers between 80 and 120 as C-81, asdescribed in our previous publication [42]. Despitethe expression of functional AR in C-81 cells, thedegree of androgen-stimulating cell growth is de-creased. Furthermore, the growth rate of C-81 cellsis more rapidly than that of C-33 cells with a dou-bling time half that of C-33 cells [42]. For experi-ments, cells were routinely seeded in PR-containingRPMI 1640 medium supplemented with 5% FBS for2 days prior to any treatment. The total cell numberwas counted using a Coulter Counter Z1 model[42,64].
2.2.2. E¡ect of steroid-depleted condition on prostatecancer cells
For these experiments, LNCaP and PC-3 cells
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4330

were plated in the regular medium at a cell density of0.5U106 cells per T-25 £ask for PC-3 and C-81LNCaP cells, and 1U106 cells per T-25 £ask for C-33 LNCaP cells, and allowed to attach for 48 h. Cellswere then fed with a steroid-reduced (SR) medium,i.e., PR-free RPMI 1640 medium containing heat-in-activated 5% charcoal-stripped fetal bovine serum(CS-FBS) and gentamicin, for 5 days as describedpreviously [64].
2.2.3. IL-6 and db-cAMP-induced neuroendocrinedi¡erentiation
Cell were seeded in a density of 0.5U106 cells perT-25 £ask for PC-3 and C-81 LNCaP cells, and1U106 cells per T-25 £ask for C-33 LNCaP cells inRPMI 1640 medium containing 5% FBS for 2 days,then 24 h in a SR medium, and followed by treat-ment with 50 ng/ml IL-6 or 1 mM db-cAMP for3 days as described previously [4,51].
2.2.4. Sodium butyrate e¡ectA cell density of 1U106 cells per T-25 £ask for C-
33 LNCaP cells and 0.5U106 cells per T-25 £ask forC-81 cells were plated in RPMI 1640 medium sup-plemented with 5% FBS for 2 days. Cells were thenfed with a SR medium with or without sodium bu-tyrate as described previously [64]. After 72 h, cellswere harvested for Western blot analyses.
2.2.5. Trichostatin A e¡ectA cell density of 1U106 cells per T-75 £ask for C-
33 LNCaP cells was plated and maintained in RPMI1640 medium supplemented with 5% FBS for 2 days.Cells were then fed with a SR medium with or with-out TSA. After 48 h, cells were harvested for West-ern blot analysis.
2.2.6. Immunocytochemical analysisFor immunocytochemical analysis, C-33 LNCaP
cells were plated at 2U104 cells per well in four-well glass slides (Nalge Nunc). Immunostaining wasperformed after treatments with a modi¢ed avidin^biotin complex technique [35]. Cells were washedbrie£y with PBS, ¢xed in 310³C methanol for5 min and air-dried. The nonspeci¢c immunoglobulinbinding was blocked by incubation with 1.5% normaldonkey serum (Santa Cruz Biotechnology) for GS,NT, ChA antibodies or goat serum (Santa Cruz Bio-
technology) for NSE antibody for 1 h. The primaryantibodies were applied to cells and incubated for 30min at room temperature. The slides were then incu-bated with biotinylated IgG secondary antibody(goat anti-mouse or donkey anti-goat depending onthe primary antibody host; Santa Cruz Biotechnol-ogy) in PBS with 1.5% normal blocking serum atroom temperature for 30 min. Avidin^biotin com-plexes were made according to the manufacturer'sinstructions, applied to the section, and incubatedat room temperature for 30 min. Immunopositivitywas visualized with the chromogen diaminobenzidine(0.025%) in substrate bu¡er and 0.1% hydrogen per-oxide. Immunostaining with the negative controls(no primary antibody) was proceeded as describedfor the primary antibody.
2.2.7. Western blotting of cellular protein and mediumCells were harvested and lysed as described previ-
ously [46]. Brie£y, cells were scraped, pelleted andrinsed with ice-cold HEPES-bu¡ered saline, pH 7.0,then lysed in an ice-cold cell lysis bu¡er containing abattery of protease and phosphatase inhibitors. Thecellular lysates were spun, the supernatants were re-covered and the protein concentration was deter-mined using the Bio-Rad protein assay kit. The me-dium was harvested and spun at 4³C for 10 min toremove any cell debris before usage. For immuno-blotting, an aliquot of total lysate protein (30 Wg) or15 Wl of culture medium in a 2USDS^PAGE samplebu¡er (1:1, v/v) was electrophoresed and transferredto nitrocellulose ¢lters. Filters were incubated withappropriate primary antibody in Tris-bu¡ered saline,pH 7.5, containing 0.1% Tween 20 (TBST) and 5%skim milk for 3 h, subsequently with secondary anti-body in TBST bu¡er for 1 h. The protein bandswere visualized by an ECL detection system. Forreprobing, ¢lters were agitated with the strippingbu¡er for 30 min at 50³C [42,45]. After two washeswith TBST bu¡er, the ¢lters were reacted with spe-ci¢c antibodies and the signal was detected by anECL method.
2.3. Data analysis
The intensities of hybridization bands were semi-quanti¢ed by densitometric analyses of autoradio-grams with di¡erent exposure time periods utilizing
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 31

a Molecular Dynamics Computing Densitometer andits software program.
3. Results
3.1. E¡ect of steroid-depleted medium, IL-6 anddb-cAMP on transdi¡erentiation of prostatecancer cells
Since several pharmacological reagents have beenreported individually to induce transdi¡erentiation ofLNCaP or PC-3 human prostate cancer cells to aNE-like phenotype, we examined if the same reagentcan have a similar e¡ect on these two prostate cancercells. C-81 LNCaP cells which exhibit androgen-un-
responsive cell growth despite the expression of func-tional AR were also included for analysis. Due to theandrogenic activity of PR [41], all cells were main-tained in a SR condition, i.e., PR-free, steroid-re-duced medium, in the presence or absence of IL-6,or db-cAMP. Control cells were maintained in theroutine growth condition, i.e. PR-positive mediumcontaining 5% FBS.
In the SR condition alone or with the addition ofdb-cAMP or IL-6, PC-3 cells showed little change intheir morphology in comparison with control cellsgrown in the PR-positive medium containing 5%FBS (data not shown and [4]). C-33 LNCaP controlcells had an epithelial morphology (Fig. 1A), andtended to grow in clusters with a somewhat acinarappearance (data not shown). In contrast, in the SR
Fig. 1. Morphological changes of C-33 LNCaP cells induced by di¡erent pharmacological reagents. (A) Control LNCaP cells main-tained in a PR-positive medium supplemented with 5% FBS; (B) LNCaP cells maintained in a PR-negative medium supplementedwith 5% SR-FBS; (C) LNCaP cells maintained in a PR-negative medium supplemented with 5% SR-FBS and 50 ng/ml IL-6; (D)LNCaP cells maintained in a PR-negative medium supplemented with 5% SR-FBS and 1 mM db-cAMP. The photo of the morpho-logical alternation was made at day 3 (A,C,D) and day 5 (B) of treatment (100U magni¢cation). Similar results were observed for Cand D with a 5 day treatment (data not shown).
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4332
condition, C-33 LNCaP cells had a neuronal appear-ance characterized by multipolar cells with small cellbodies and long, irregular processes (Fig. 1B). Sim-ilar morphologic e¡ects were seen for these cellsgrown in the IL-6-containing medium although lesspronounced (Fig. 1C), while db-cAMP had a morepotent e¡ect on cellular morphology (Fig. 1D). Incomparison, C-81 LNCaP cells followed a similartrend to C-33 LNCaP cells in the phenotype alterna-
tion although the e¡ect was less pronounced (datanot shown).
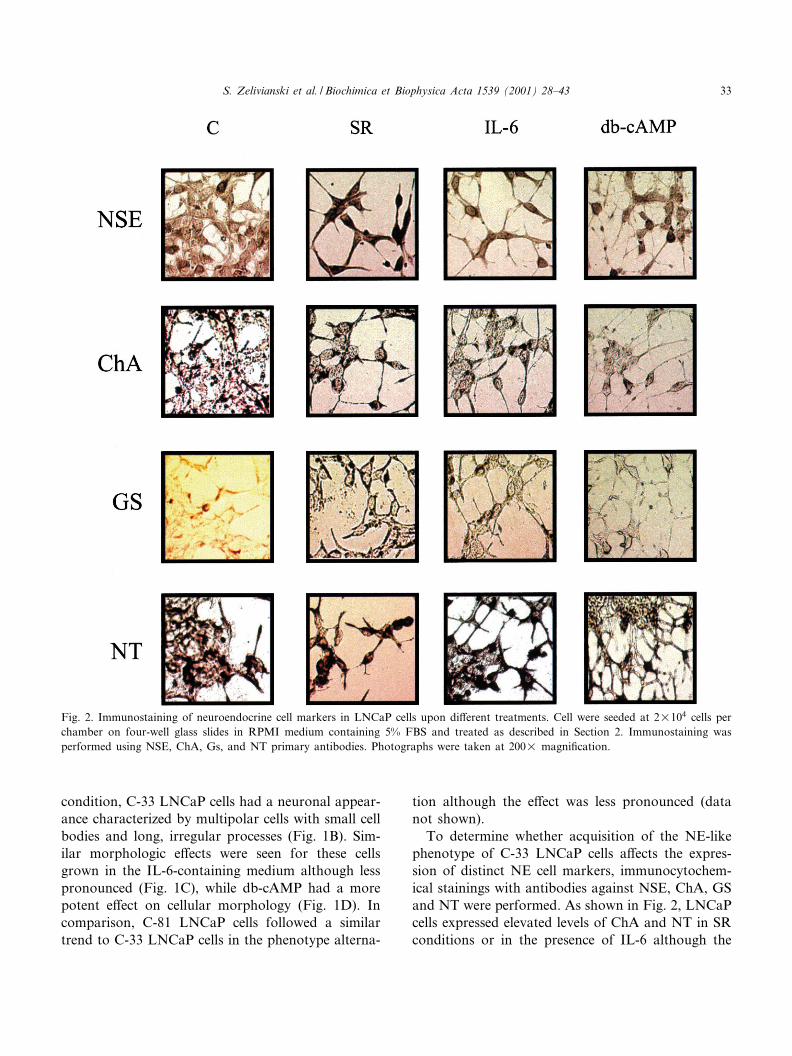
To determine whether acquisition of the NE-likephenotype of C-33 LNCaP cells a¡ects the expres-sion of distinct NE cell markers, immunocytochem-ical stainings with antibodies against NSE, ChA, GSand NT were performed. As shown in Fig. 2, LNCaPcells expressed elevated levels of ChA and NT in SRconditions or in the presence of IL-6 although the
Fig. 2. Immunostaining of neuroendocrine cell markers in LNCaP cells upon di¡erent treatments. Cell were seeded at 2U104 cells perchamber on four-well glass slides in RPMI medium containing 5% FBS and treated as described in Section 2. Immunostaining wasperformed using NSE, ChA, Gs, and NT primary antibodies. Photographs were taken at 200U magni¢cation.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 33

degree of elevation is low. These data showed thatLNCaP cells exhibit the ability to express ChA andNT prior to induced transdi¡erentiation by exoge-nous stimuli. In contrast, the immunoreactivitywith NSE and GS antibodies was found to be highlyincreased in cells maintained in the SR condition andin the presence of IL-6. Interestingly, while db-cAMPhad a potent e¡ect on cellular morphology (Fig. 1D),it did not signi¢cantly a¡ect the levels of NSE, ChA,GS, and NT. Nevertheless, the immunostainingclearly showed increasing NE characteristics of C-33 LNCaP cells in SR conditions including manifes-tation of the cytoplasmic NE granules in ChA stain-ing and the cytosolic staining of NSE. Thus, di¡erentgrowth conditions including pharmacological re-agents can induce NE-like phenotypic changes inprostate cancer cells, while they may have di¡erentiale¡ects on the expression of biomarkers. Based onthese data, we used NSE as the major marker inour further studies to monitor the transdi¡erentia-tion process of LNCaP cells into NE-like cells.
Western blot analyses were performed to semi-quantify the expression of NSE in cells grown indi¡erent conditions. As shown in Fig. 3 in C-33LNCaP cells grown in the SR condition, the NSElevel was elevated by approx. 3-fold, while to a lesserdegree with an addition of IL-6, as observed in Fig.2. Unexpectedly, in the presence of db-cAMP, theexpression of NSE was decreased, even lower thanthat in control cells grown in the regular growthcondition. An opposite e¡ect was observed in C-81LNCaP cells in which the NSE level was decreased inthe SR condition, while elevated in db-cAMP-supple-mented condition (Fig. 3). The level of NSE was notsigni¢cantly altered by adding IL-6 to those cells.Interestingly, as shown in Fig. 3, PC-3 control cellsexpressed a high basal level of NSE in comparisonwith LNCaP cells. The NSE level was only elevatedin the db-cAMP-treated cells, but did not change inthe SR medium or IL-6-supplemented medium.Thus, despite a similar e¡ect on the morphology ofC-33 and C-81 LNCaP cells by the steroid-reducedcondition and the db-cAMP supplement, thesegrowth conditions have an opposite e¡ect on theNSE expression. Thus, di¡erential molecular expres-sions can correlate with a similar gross phenotypicchange.
PC-3 cells are poorly di¡erentiated prostate cancer
cells, and do not express any known biomarkers in-cluding PSA. Similar to the observations by Bang etal. [4], we found that PC-3 cells only exhibited aminor change in their phenotype and a marginal el-evation of NSE level upon db-cAMP treatment. Wetherefore focused our e¡orts on characterizing bio-markers during transdi¡erentiation utilizing geneti-cally lineaged C-33 and C-81 LNCaP cells sincethey are derived from the same cell origin, andmay represent di¡erent stages of tumor progressionregarding androgen responsiveness [42], but not PC-3cells. We utilized these two cells as our model systemfor further studies.
3.2. E¡ect of steroid-depleted medium, IL-6, anddb-cAMP on PSA and AR expression inLNCaP cells
To determine molecular changes that are associ-ated with NE di¡erentiation of prostate cells, we
Fig. 3. Expression of NSE in human prostate cancer cells indi¡erent growth conditions. Cells were maintained in a PR-pos-itive medium supplemented with 5% FBS (C), a PR-negativemedium supplemented with 5% SR-FBS (SR), PR-negative me-dium supplemented with 5% SR-FBS and 50 ng/ml IL-6 (IL-6),or 5% SR-FBS and 1 mM db-cAMP (db-cAMP). Total cell ly-sates were electrophoresed and transferred to nitrocellulosemembranes for Western blotting with anti-NSE antibody. Aportion of membrane was dissected for Coomassie blue stainingto show a similar amount of loaded protein in each lane (datanot shown). Similar results were obtained from three sets of in-dependent experiments.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4334

analyzed the expression of PSA and AR in LNCaPcells grown in di¡erent growth conditions. As shownin Fig. 4A, in the SR medium in the presence orabsence of IL-6, the cellular level of PSA was down-
regulated in both C-33 and C-81 cells, although alesser e¡ect was obtained in C-33 cells. Db-cAMPtreatments resulted in an increase in the intracellularlevel of PSA in both LNCaP cells (Fig. 4A). Thesedata were consistent with the secreted level of PSA in
Fig. 4. E¡ect of the steroid-depleted condition, IL-6, or db-cAMP on PSA and AR expression in LNCaP cells. C-33 and C-81 LNCaPcells were plated and treated with 50 ng/ml IL-6 (IL-6) or 1 mM db-cAMP (db-cAMP) as described in Fig. 3. The same set of sam-ples was used for Western blot analyses to detect the expression level of PSA (A) and AR (B). 30 Wg total lysate proteins and 25 Wlmedium were used for Western blot analyses on PSA level in LNCaP cell lysates and in their conditioned medium. The ratio of PSAexpression in cell lysates was similar to that in conditioned medium from LNCaP cells. Similar results were obtained from three setsof independent experiments.
C
Fig. 5. Western blot analyses of RPTPK expression in LNCaPcells. Cells were cultured at growth conditions in a PR-positivemedium supplemented with 5% FBS (C), PR-negative mediumsupplemented with 5% SR-FBS (SR), PR-negative medium sup-plemented with 5% SR-FBS and 50 ng/ml IL-6 (IL-6), or 5%SR-FBS and 1 mM db-cAMP (db-cAMP). 30 Wg total lysateproteins were electrophoresed and blotted on a nitrocellulosemembrane, and the membrane was subsequently hybridizedwith anti-RPTPK antibody. It should be noted that a prolongedexposure showed a low level of RPTPK expression in C-33 cellsgrown in the IL-6-containing medium (data not shown). A por-tion of membrane was dissected for Coomassie blue staining toshow a similar amount of loaded protein in each lane (data notshown). Similar results were obtained from three sets of inde-pendent experiments.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 35
the conditioned medium from those cells (Fig. 4A).In the SR condition, the level of AR was elevated inC-33 LNCaP cells, while markedly decreased in C-81cells (Fig. 4B). IL-6 also caused an increase of AR inC-33 cells, but a decrease in C-81 cells (Fig. 4B). Db-cAMP exhibited a negative e¡ect on the AR level inC-81 cells, but not in C-33 cells. Thus, PSA, an ep-ithelial cell di¡erentiation marker, is regulated di¡er-ently from NSE, a NE cell marker.
3.3. Expression of RPTPK in LNCaP cells duringneuroendocrine transdi¡erentiation
It has been shown that RPTPK plays a critical role
in neuronal di¡erentiation and signaling [60,62]. Weinvestigated if RPTPK may also be involved in pros-tate NE di¡erentiation. As shown in Fig. 5, in cellscultured in a SR medium, the level of RPTPK ex-pression was increased in both LNCaP cells, corre-lating with their morphological changes. However, incells grown in the presence of IL-6 or db-cAMP, theexpression of RPTPK was markedly decreased (Fig.5). Thus, in the androgen-depleted condition, the ex-pression of RPTPK correlates with phenotypicchanges of LNCaP cells, independent of their andro-gen responsiveness. Furthermore, the expression ofRPTPK in these two cells has the same response indi¡erent growth conditions.
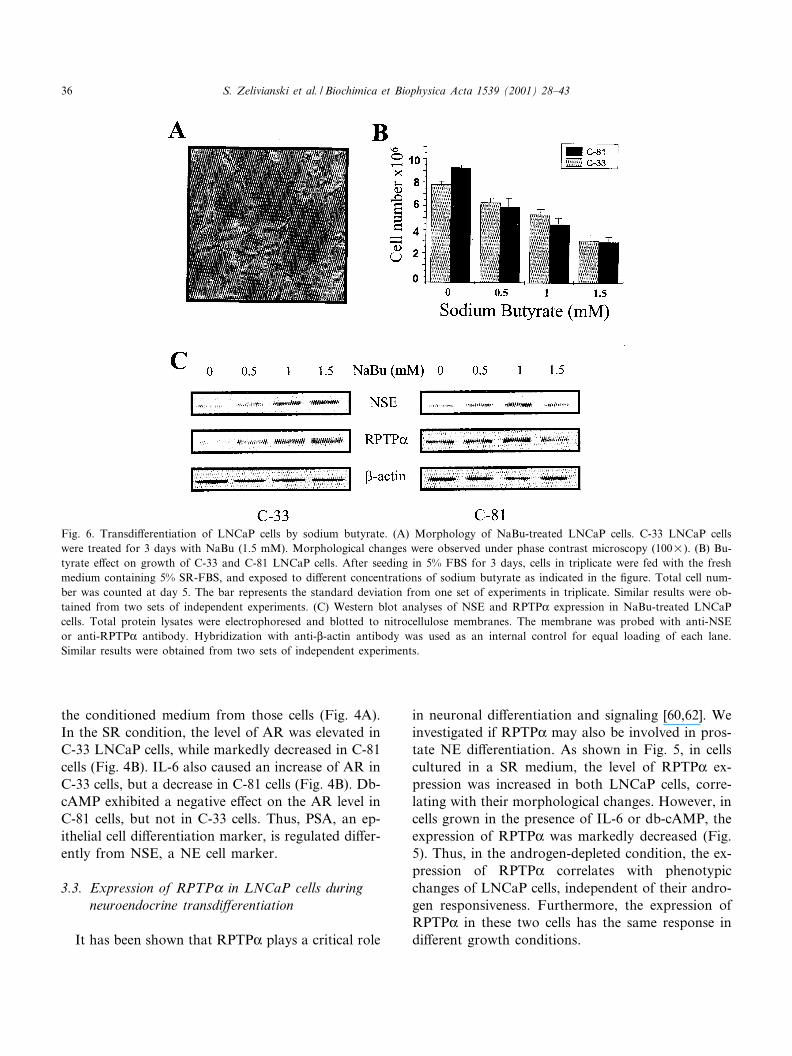
Fig. 6. Transdi¡erentiation of LNCaP cells by sodium butyrate. (A) Morphology of NaBu-treated LNCaP cells. C-33 LNCaP cellswere treated for 3 days with NaBu (1.5 mM). Morphological changes were observed under phase contrast microscopy (100U). (B) Bu-tyrate e¡ect on growth of C-33 and C-81 LNCaP cells. After seeding in 5% FBS for 3 days, cells in triplicate were fed with the freshmedium containing 5% SR-FBS, and exposed to di¡erent concentrations of sodium butyrate as indicated in the ¢gure. Total cell num-ber was counted at day 5. The bar represents the standard deviation from one set of experiments in triplicate. Similar results were ob-tained from two sets of independent experiments. (C) Western blot analyses of NSE and RPTPK expression in NaBu-treated LNCaPcells. Total protein lysates were electrophoresed and blotted to nitrocellulose membranes. The membrane was probed with anti-NSEor anti-RPTPK antibody. Hybridization with anti-L-actin antibody was used as an internal control for equal loading of each lane.Similar results were obtained from two sets of independent experiments.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4336

3.4. Sodium butyrate e¡ect on NSE and RPTPKexpression in LNCaP cells
To further examine the correlation of NSE andRPTPK expression with NE di¡erentiation of pros-tate carcinoma cells, sodium butyrate (NaBu) wasutilized to induce cell di¡erentiation [2,33]. NaBuinduced a NE-di¡erentiation phenotype (Fig. 6A),similar to the IL-6 e¡ect (Fig. 1C) that correlatedwith a diminished cell growth (Fig. 6B). Interestingly,NaBu treatment resulted in an elevated level of NSEand RPTPK protein in both LNCaP cells although ithad a lesser e¡ect with di¡erent dose^response pat-terns in C-81 cells than in C-33 cells (Fig. 6C). Thus,the treatment of LNCaP cells with NaBu leads to amarked increase of NSE and RPTPK levels, whichcorrelates with a NE-like phenotype of LNCaP cells,independent of androgen responsiveness of growthregulation.
3.5. Trichostatin A e¡ect on NSE and RPTPKexpression in C-33 LNCaP cells
NaBu exhibits several biological e¡ects includingthat it can function as an inhibitor of histone deace-tylases (HDAC) and cause a hyperacetylation of corehistones. To investigate if histone hyperacetylationhas the same e¡ect on the expression of NSE andRPTPK during NE di¡erentiation by NaBu, C-33
cells were treated with trichostatin A (TSA), a spe-ci¢c HDAC inhibitor. As shown in Fig. 7, TSA in-duced NSE expression in a dose-dependent manner,while it inhibited the expression level of RPTPK. Theresults thus indicated that the activation of RPTPKexpression by NaBu is mediated by the activation ofcellular pathways other than hyperacetylation of corehistones.
4. Discussion
Clinical results indicate that NE cells may play acritical role in androgen-refractory growth of pros-tate cancer cells [9]. While the origin of these NEcells is not clearly de¢ned in clinical studies, recentexperimental studies have shown that prostate cancercells can directly give rise to cells with a NE-likephenotype. This process is referred to as transdi¡er-entiation [4,51,55]. Although di¡erent prostate can-cer cells can be transdi¡erentiated into a similar NEcell-like phenotype by di¡erent treatments [4,13,26,51,55], there is no direct comparison among thosetreatments in di¡erent cells. Thus, it is not clear ifdi¡erent cells can follow the same pathway of trans-di¡erentiation. The experimental work in this studyis to investigate molecular mechanisms of transdi¡er-entiation in di¡erent prostate cancer cells includingdi¡erent LNCaP cells. LNCaP cells are partiallydi¡erentiated prostate cancer cells and expressPSA, a prostate epithelium-speci¢c di¡erentiationmarker. In di¡erent LNCaP cells, despite a similarexpression level of functional AR and PSA, theresponsiveness of androgen stimulation of cell prolif-eration is altered, imitating prostate tumor progres-sion [42]. Thus, di¡erent LNCaP cells represent aninteresting, unique cell model in studying the trans-di¡erentiation of prostate cancer cells during tumorprogression.
In our experiments, the end product of prolongedmaintenance of C-33 LNCaP cells in the SR condi-tion is a cell that resembles the phenotype of NE cellsmuch more than the morphology of prostate epithe-lial cells. By growth in this condition, the alteredphenotype of NE transdi¡erentiated C-33 LNCaPcells is dramatically di¡erent from that of controlcells with di¡erentiated characteristics of prostate ep-ithelial cells (Fig. 1A vs. B). We found that andro-
Fig. 7. E¡ect of TSA on NSE and RPTPK expression inLNCaP cells. C-33 LNCaP cells were plated at a density of1U106 per T-75 £ask. 48 h after seeding, cells were treated for48 h with di¡erent concentrations of TSA as indicated in the¢gure. Total lysate proteins were separated on polyacrylamidegels, transferred to nitrocellulose membrane, and probed withan anti-NSE and anti-RPTPK antibody, respectively. Hybridiza-tion with anti-L-actin antibody was used as a control for dem-onstrating an equal loading of each lane. Similar results wereobserved in two sets of independent experiments.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 37

gen-responsive C-33 LNCaP cells grown in steroid-reduced condition express higher levels of di¡erentmarkers of NE cells including NSE, NT, ChA andGS than control cells (Figs. 2 and 3). Thus our re-sults clearly showed that C-33 LNCaP cells canundergo transdi¡erentiation into a cell type withstrong characteristics of NE cells, suggesting the pos-sibility that such an event might underline the appar-ent accumulation of NE cells in hormone-refractorytumors. This phenomenon may also re£ect clinicalobservations that, during androgen-ablation therapy,the androgen-responsive prostate cancer cells cantransdi¡erentiate into NE-like cells, and the popula-tion of NE cells is thus increased.
In archival specimens of normal, hyperplastic andneoplastic human prostate, NE cells lack AR and areconsidered to be androgen independent [8,36]. Incontrast to NE cells which are histologically consid-ered nonproliferative [23] and represent a terminallydi¡erentiated cell population [6], we found that NE-like cells derived from LNCaP cells still express AR(Fig. 4B) and could proliferate in steroid-restrictedconditions. Furthermore, a recent report shows thereversibility of the NE phenotype of LNCaP cells[16], suggesting that the acquisition of NE character-istics may be an epigenetic phenomenon that canoccur gradually, depending on the phenotypic poten-tial of a cell and the in£uence of its environment.These ¢ndings together indicate that there are appar-ent di¡erences between NE-like cells and NE cells ofthe prostate.
Interestingly, in C-81 cells grown in steroid-re-duced condition, the NSE level was not increased,while RPTPK had an increased expression as in C-33 cells (Fig. 3 vs. Fig. 5). C-81 LNCaP cells are anandrogen-unresponsive subline of LNCaP cells andexpress PSA (Fig. 4A and [42]). Thus, these cellsexhibit di¡erent biochemical properties from paren-tal C-33 LNCaP cells. Although these cells express afunctional AR (Fig. 4B), their growth is androgen-unresponsive [42] and exhibits a rapid growth rate insteroid restricted condition, similar to androgen-in-dependent PC-3 cells. In this condition, RPTPK ex-pression, but not NSE, was increased in steroid-re-stricted medium as parental C-33 cells. This could bedue to the fact that RPTPK is a di¡erent markerfrom NSE, and involved in di¡erent cellular func-tions in neuronal di¡erentiation. RPTPK may be in-
volved in the early stage of di¡erentiation, whileNSE is a di¡erentiated marker. The regulations oftheir expressions are therefore via di¡erent molecularmechanisms. The detailed molecular mechanism isunder investigation.
It has been reported that the IL-6 level in circula-tion is elevated in a subset of patients with advancedprostate carcinomas who have failed androgen-abla-tion therapy [59]. Upregulation of IL-6 in these pa-tients may be due to the negative regulation of theIL-6 gene by androgenic hormones [5] since the ex-pression of IL-6 receptor is increased after with-drawal of androgens [43]. These ¢ndings togetherpoint to the importance of cross-talks between IL-6and androgen signaling pathways in prostatic carci-nomas [45,50]. We have observed that IL-6-treatedLNCaP cells acquire a neuron-like morphology (Fig.1C). To determine whether these IL-6-treatedLNCaP cells exhibit some characteristics of NE cells,we analyzed the expression of NSE (Figs. 2 and 3).We also compared these cells with poorly di¡erenti-ated androgen-unresponsive PC-3 cells which expressa high basal level of NSE (Fig. 3). IL-6 has no e¡ecton the NSE level in PC-3 cells, correlating with alimited morphological change of these cells. How-ever, IL-6 has an intriguing e¡ect in LNCaP cells.In androgen responsive C-33 cells, although theNSE level is higher in IL-6-treated cells than in con-trol cells grown in the regular growth condition, theNSE level is decreased, lower than in the SR alone.On the contrary, in C-81 cells, the NSE level is high-er in the IL-6-containing medium than in the SRalone, indicating IL-6 counteracts the e¡ect by theSR medium. The results clearly show that the intra-cellular signaling of IL-6 is di¡erent in androgen-re-sponsive and -unresponsive prostate cancer cells. Onepossibility is that di¡erent signal transduction path-ways are activated in those cells, due to di¡erences inthe expression of signaling molecules. For example, ithas been reported that PC-3 cells may not expressfunctional STAT3 [56], a transducer of IL-6 signal-ing. Of course, there are other explanations. Furtherstudies are required to determine the IL-6 signalingin prostate cancer cells.
It has been shown that cAMP can reverse thetransformed phenotype of some cancer cells, possiblyby restoring the normal regulatory pathway [3]. Thecritical genes regulated by cAMP in prostate carci-
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4338

noma cells may include genes that induce NE di¡er-entiation as well as downregulate cell cycle progres-sion. Our data show that an elevation of cAMP in-duces transdi¡erentiation primarily in androgen-unresponsive C-81 LNCaP and PC-3 cells, indicatedby NSE expression and morphology change.
Our data thus reveal that androgen-unresponsivehuman prostate cancer cells respond di¡erently fromandrogen-responsive cells to pharmacological treat-ments. This phenomenon thus resembles the hetero-geneity of prostate cancer cells. Furthermore,although cAMP induces a dramatic change in thephenotype of C-33 LNCaP cells (Fig. 1D), cAMPcaused a decrease of NSE in those cells (Figs. 2and 3). The di¡erential responses of NSE expressionin di¡erent prostate cells is also similar to the re-sponse of c-myc gene expression to protein kinaseC (PKC) action on those same cells [34]. Thus, asimilar change in gross phenotype may accompanydi¡erential gene expressions.
As described above, IL-6 and db-cAMP have dif-ferential e¡ects on di¡erent prostate cancer cells. Inprostate cancer cells, results of several studies haveshown that IL-6 and db-cAMP can stimulate or in-hibit cell growth, depending on the targeted cells. Ithas been proposed that IL-6 and db-cAMP mayshare the same MAPK signaling pathway througha cross-talk with ErbB2 which activates the MAPKpathway in LNCaP cells, while it inhibits it in PC-3cells [11,50,56]. Taken together all these ¢ndings in-dicate that PC-3 may respond di¡erently fromLNCaP cells in the presence of di¡erent pharmaco-logical agents. The detailed molecular mechanismsremain to be clari¢ed.
To further examine the regulatory mechanism oftransdi¡erentiation of human prostate cells, we uti-lized di¡erent LNCaP cells, a genetically lineaged cellmodel system. PSA is expressed only in di¡erentiatedsecretory prostatic epithelial cells, and serves as amarker of prostate di¡erentiation [14]. Both LNCaPcells express a high level of PSA (Fig. 4A and [42]).In our studies, IL-6 induces NE-like morphologychanges and has an inhibition of PSA expression inboth LNCaP cells. Contrarily, db-cAMP upregulatesthe PSA level. It has been shown that human AR canbe activated through an alternate signaling pathwayinvolving db-cAMP in a ligand-independent mannerin prostate cancer cells [18,48]. The activation of AR
by db-cAMP can thus enhance the speci¢c activity ofAR, but not its protein level, which increases PSAexpression.
The transmembrane nature of the receptor-likePTPs indicates that these PTPases can be involvedin transducing extracellular signals. RPTPK has ahigh level of expression in brain [28,54], and canassociate with contactin, a neuronal cell adhesionmolecule, through the extracellular region [65]. Sev-eral lines of evidence have demonstrated that RPTPKplays a critical role in normal neuronal di¡erentia-tion [20,60,67]. We have shown that in the steroid-reduced condition, RPTPK expression in bothLNCaP cells is upregulated (Fig. 5), correlatingwith the phenotype change (Fig. 1B). To understandthe possible functional role of RPTPK in prostateNE transdi¡erentiation, NaBu is utilized since thisreagent can induce partial di¡erentiation of PC-3cells [33]. This reagent is currently under clinical con-sideration as a potential agent for the managementof advance prostate cancer [52]. However, the molec-ular mechanism in this process is not completelyknown. In this study, NSE and RPTPK are used asmarkers to investigate the signaling pathway byNaBu in cell di¡erentiation. Our data clearly showthat NaBu treatment results in morphologicalchanges and elevated NSE levels (Fig. 6), and thusresembles a transdi¡erentiation pathway of LNCaPcells. NaBu treatment results in an upregulation ofRPTPK expression in C-33 and C-81 LNCaP cells(Fig. 6C). Since TSA is a speci¢c inhibitor of histonedeacetylases [63], it is a useful tool to study the roleof histone acetylation in the regulation of NSE andRPTPK expression during NE di¡erentiation by bu-tyrate. Unexpectedly, TSA induces a dose-dependentactivation of NSE, while it inhibits RPTPK expres-sion in C-33 LNCaP cells (Fig. 7). Taken collectivelythe data suggest that NaBu activation of NSE andRPTPK expression is mediated by di¡erent signalingpathways although both NaBu and TSA have a sim-ilar e¡ect on histone acetylation. For the ¢rst time,our data show that RPTPK may act as a molecularmediator in the signaling pathway initiated by NaBualthough the link between sodium butyrate andRPTPK remains to be identi¢ed. Since cell growthand di¡erentiation can be regulated by protein phos-phorylation and dephosphorylation, RPTPK mayplay an important role in the modulation of gene
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 39
expression for transdi¡erentiation of prostate carci-noma cells during androgen-ablation therapy, andmay also serve as an additional marker for this pro-cess.
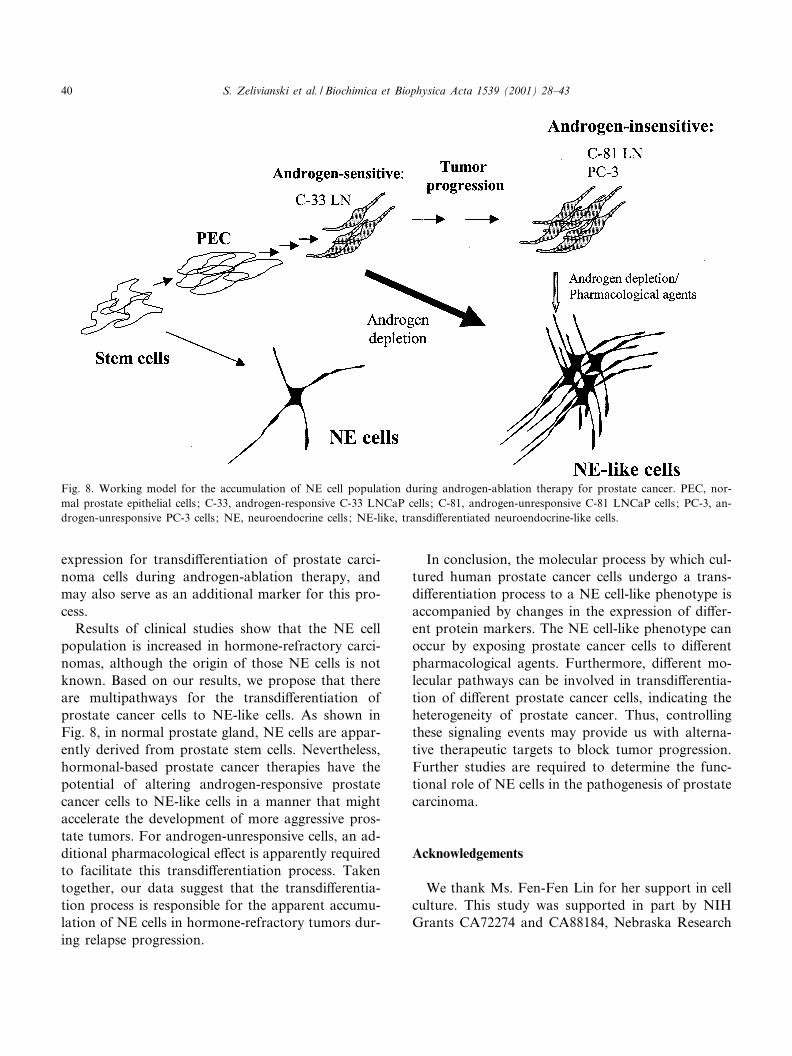
Results of clinical studies show that the NE cellpopulation is increased in hormone-refractory carci-nomas, although the origin of those NE cells is notknown. Based on our results, we propose that thereare multipathways for the transdi¡erentiation ofprostate cancer cells to NE-like cells. As shown inFig. 8, in normal prostate gland, NE cells are appar-ently derived from prostate stem cells. Nevertheless,hormonal-based prostate cancer therapies have thepotential of altering androgen-responsive prostatecancer cells to NE-like cells in a manner that mightaccelerate the development of more aggressive pros-tate tumors. For androgen-unresponsive cells, an ad-ditional pharmacological e¡ect is apparently requiredto facilitate this transdi¡erentiation process. Takentogether, our data suggest that the transdi¡erentia-tion process is responsible for the apparent accumu-lation of NE cells in hormone-refractory tumors dur-ing relapse progression.
In conclusion, the molecular process by which cul-tured human prostate cancer cells undergo a trans-di¡erentiation process to a NE cell-like phenotype isaccompanied by changes in the expression of di¡er-ent protein markers. The NE cell-like phenotype canoccur by exposing prostate cancer cells to di¡erentpharmacological agents. Furthermore, di¡erent mo-lecular pathways can be involved in transdi¡erentia-tion of di¡erent prostate cancer cells, indicating theheterogeneity of prostate cancer. Thus, controllingthese signaling events may provide us with alterna-tive therapeutic targets to block tumor progression.Further studies are required to determine the func-tional role of NE cells in the pathogenesis of prostatecarcinoma.
Acknowledgements
We thank Ms. Fen-Fen Lin for her support in cellculture. This study was supported in part by NIHGrants CA72274 and CA88184, Nebraska Research
Fig. 8. Working model for the accumulation of NE cell population during androgen-ablation therapy for prostate cancer. PEC, nor-mal prostate epithelial cells ; C-33, androgen-responsive C-33 LNCaP cells ; C-81, androgen-unresponsive C-81 LNCaP cells ; PC-3, an-drogen-unresponsive PC-3 cells ; NE, neuroendocrine cells ; NE-like, transdi¡erentiated neuroendocrine-like cells.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4340

Initiative, Nebraska Department of Health LB506,LB 595.
References
[1] A. Angelsen, U. Syversen, O.A. Haugen, M. Stridsberg,O.K. Mjolnerod, H.L. Waldum, Neuroendocrine di¡erentia-tion in carcinomas of the prostate: do neuroendocrine serummarkers re£ect immunohistochemical ¢ndings?, Prostate 30(1997) 1^6.
[2] S. Archer, S. Meng, J. Wu, J. Johnson, R. Tang, R. Hodin,Butyrate inhibits colon carcinoma cell growth through twodistinct pathways, Surgery 124 (1998) 248^253.
[3] F. Ashall, N. Sullivan, T.T. Puck, Speci¢city of the cAMP-induced gene exposure reaction in CHO cells, Proc. Natl.Acad. Sci. USA 85 (1988) 3908^3912.
[4] Y.J. Bang, F. Pirnia, W.G. Fang, W.K. Kang, O. Sartor, L.Whitesell, M.J. Ha, M. Tsokos, M.D. Sheahan, P. Nguyen,W.T. Niklinski, C.E. Myers, J.B. Trepel, Terminal neuroen-docrine di¡erentiation of human prostate carcinoma cells inresponse to increased intracellular cyclic AMP, Proc. Natl.Acad. Sci. USA 91 (1994) 5330^5334.
[5] T. Bellido, R.L. Jilka, B.F. Boyce, G. Girasole, H. Brox-meyer, S.A. Dalrymple, R. Murray, S.C. Manolagas, Regu-lation of interleukin-6, osteoclastogenesis, and bone mass byandrogens. The role of the androgen receptor, J. Clin. Invest.95 (1995) 2886^2895.
[6] H. Bonkho¡, Neuroendocrine cells in benign and malignantprostate tissue: morphogenesis, proliferation, and androgenreceptor status, Prostate 8 (Suppl.) (1998) 18^22.
[7] H. Bonkho¡, K. Remberger, Di¡erentiation pathways andhistogenetic aspects of normal and abnormal prostaticgrowth: a stem cell model, Prostate 28 (1996) 98^106.
[8] H. Bonkho¡, U. Stein, K. Remberger, Multidirectional dif-ferentiation in the normal, hyperplastic, and neoplastic hu-man prostate: simultaneous demonstration of cell-speci¢cepithelial markers, Hum. Pathol. 25 (1994) 42^46.
[9] H. Bonkho¡, N. Wernert, G. Dhom, K. Remberger, Rela-tion of endocrine-paracrine cells to cell proliferation in nor-mal, hyperplastic, and neoplastic human prostate, Prostate19 (1991) 91^98.
[10] T. Burchardt, M. Burchardt, M.W. Chen, Y. Cao, A. de laTaille, A. Shabsigh, O. Hayek, T. Dorai, R. Buttyan, Trans-di¡erentiation of prostate cancer cells to a neuroendocrinecell phenotype in vitro and in vivo, J. Urol. 162 (1999) 1800^1805.
[11] T. Chen, R.W. Cho, P.J. Stork, M.J. Weber, Elevation ofcyclic adenosine 3P,5P-monophosphate potentiates activationof mitogen-activated protein kinase by growth factors inLNCaP prostate cancer cells, Cancer Res. 59 (1999) 213^218.
[12] W.L. Cheung, S.D. Briggs, C.D. Allis, Acetylation and chro-mosomal functions, Curr. Opin. Cell Biol. 12 (2000) 326^333.
[13] J.W. Chiao, T.C. Hsieh, W. Xu, R.J. Sklarew, R. Kancherla,Development of human prostate cancer cells to neuroendo-crine-like cells by interleukin-1, Int. J. Oncol. 15 (1999)1033^1037.
[14] T.M. Chu, M.F. Lin, PSA and acid phosphatase in the di-agnosis of prostate cancer, J. Clin. Ligand Assay 21 (1998)24^34.
[15] P. Constantoulakis, G. Knitter, G. Stamatoyannopoulos, Onthe induction of fetal hemoglobin by butyrates: in vivo andin vitro studies with sodium butyrate and comparison ofcombination treatments with 5-AzaC and AraC, Blood 74(1989) 1963^1971.
[16] M.E. Cox, P.D. Deeble, S. Lakhani, S.J. Parsons, Acquisi-tion of neuroendocrine characteristics by prostate tumorcells is reversible: implications for prostate cancer progres-sion, Cancer Res. 59 (1999) 3821^3830.
[17] L. Cuisset, L. Tichonicky, P. Ja¡ray, M. Delpech, The ef-fects of sodium butyrate on transcription are mediatedthrough activation of a protein phosphatase, J. Biol.Chem. 272 (1997) 24148^24153.
[18] Z. Culig, A. Hobisch, M.V. Cronauer, A.C. Cato, A. Hitt-mair, C. Radmayr, J. Eberle, G. Bartsch, H. Klocker, Mu-tant androgen receptor detected in an advanced-stage pro-static carcinoma is activated by adrenal androgens andprogesterone, Mol. Endocrinol. 7 (1993) 1541^1550.
[19] J. den Hertog, T. Hunter, Tight association of GRB2 withreceptor protein-tyrosine phosphatase alpha is mediated bythe SH2 and C-terminal SH3 domains, EMBO J. 15 (1996)3016^3027.
[20] J. den Hertog, C.E. Pals, M.P. Peppelenbosch, L.G. Tertoo-len, S.W. de Laat, W. Kruijer, Receptor protein tyrosinephosphatase alpha activates pp60c-src and is involved inneuronal di¡erentiation, EMBO J. 12 (1993) 3789^3798.
[21] L. Denis, G.P. Murphy, Overview of phase III trials oncombined androgen treatment in patients with metastaticprostate cancer, Cancer 72 (1993) 3888^3895.
[22] P.A. di Sant'Agnese, Neuroendocrine di¡erentiation in hu-man prostatic carcinoma, Hum. Pathol. 23 (1992) 287^296.
[23] P.A. di Sant'Agnese, Neuroendocrine di¡erentiation in car-cinoma of the prostate. Diagnostic, prognostic, and thera-peutic implications, Cancer 70 (1992) 254^268.
[24] P.A. di Sant'Agnese, A.T. Cockett, The prostatic endocrine-paracrine (neuroendocrine) regulatory system and neuroen-docrine di¡erentiation in prostatic carcinoma: a review andfuture directions in basic research, J. Urol. 152 (1994) 1927^1931.
[25] P.A. di Sant'Agnese, K.L. de Mesy Jensen, Neuroendocrinedi¡erentiation in prostatic carcinoma, Hum. Pathol. 18(1987) 849^856.
[26] M. Diaz, M. Abdul, N. Hoosein, Modulation of neuroendo-crine di¡erentiation in prostate cancer by interleukin-1 and-2, Prostate 8 (Suppl.) (1998) 32^36.
[27] E. Espinos, M.J. Weber, Activation of the MAP kinase cas-cade by histone deacetylase inhibitors is required for thestimulation of choline acetyltransferase gene promoter.,Brain Res. Mol. Brain Res. 56 (1998) 118^124.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 41

[28] K.S. Fang, M. Martins-Green, L.T. Williams, H. Hanafusa,Characterization of chicken protein tyrosine phosphatase al-pha and its expression in the central nervous system, BrainRes. Mol. Brain Res. 37 (1996) 1^14.
[29] W.D. Figg, R.G. Walls, M.R. Cooper, A. Thibault, O. Sar-tor, N.A. McCall, C.E. Myers, D. Samid, In vitro antitumore¡ect of hydroxyurea on hormone-refractory prostate cancercells and its potentiation by phenylbutyrate, AnticancerDrugs 5 (1994) 336^342.
[30] R. Garcia-Arenas, F.F. Lin, D. Lin, L.P. Jin, C.C. Shih, C.Chang, M.F. Lin, The expression of prostatic acid phospha-tase is transcriptionally regulated in human prostate carcino-ma cells, Mol. Cell. Endocrinol. 111 (1995) 29^37.
[31] P.J. Gkonos, A. Krongrad, B.A. Roos, Neuroendocrine pep-tides in the prostate, Urol. Res. 23 (1995) 81^87.
[32] J.L. Guate, S. Escaf, C.L. Menendez, M. del Valle, J.A.Vega, Neuroendocrine cells in benign prostatic hyperplasiaand prostatic carcinoma: e¡ect of hormonal treatment,Urol. Int. 59 (1997) 149^153.
[33] J. Halgunset, T. Lamvik, T. Espevik, Butyrate e¡ects ongrowth, morphology, and ¢bronectin production in PC-3prostatic carcinoma cells, Prostate 12 (1988) 65^77.
[34] P. Henttu, P. Vihko, The protein kinase C activator, phorbolester, elicits disparate functional responses in androgen-sen-sitive and androgen-independent human prostatic cancercells, Biochem. Biophys. Res. Commun. 244 (1998) 167^171.
[35] S.M. Hsu, L. Raine, H. Fanger, Use of avidin-biotin-peroxi-dase complex (ABC) in immunoperoxidase techniques: acomparison between ABC and unlabeled antibody (PAP)procedures, J. Histochem. Cytochem. 29 (1981) 577^580.
[36] T. Jiborn, A. Bjartell, P.A. Abrahamsson, Neuroendocrinedi¡erentiation in prostatic carcinoma during hormonal treat-ment, Urology 51 (1998) 585^589.
[37] D. Kadmon, T.C. Thompson, G.R. Lynch, P.T. Scardino,Elevated plasma chromogranin-A concentrations in prostaticcarcinoma, J. Urol. 146 (1991) 358^361.
[38] M.E. Kaighn, D. Kirk, M. Szalay, J.F. Lechner, Growthcontrol of prostatic carcinoma cells in serum-free media:interrelationship of hormone response, cell density, and nu-trient media, Proc. Natl. Acad. Sci. USA 78 (1981) 5673^5676.
[39] R. Kaplan, B. Morse, K. Huebner, C. Croce, R. Howk, M.Ravera, G. Ricca, M. Jaye, J. Schlessinger, Cloning of threehuman tyrosine phosphatases reveals a multigene family ofreceptor-linked protein-tyrosine-phosphatases expressed inbrain, Proc. Natl. Acad. Sci. USA 87 (1990) 7000^7004.
[40] N.X. Krueger, M. Streuli, H. Saito, Structural diversity andevolution of human receptor-like protein tyrosine phospha-tases, EMBO J. 9 (1990) 3241^3252.
[41] M.F. Lin, R. Garcia-Arenas, M. Kawachi, F.F. Lin, Regu-lation of the expression of prostatic acid phosphatase inLNCaP human prostate carcinoma cells, Cell. Mol. Biol.Res. 39 (1993) 739^750.
[42] M.F. Lin, T.C. Meng, P.S. Rao, C. Chang, A.H. Schonthal,F.F. Lin, Expression of human prostatic acid phosphatase
correlates with androgen-stimulated cell proliferation inprostate cancer cell lines, J. Biol. Chem. 273 (1998) 5939^5947.
[43] P. Lin, P. Polverini, M. Dewhirst, S. Shan, P.S. Rao, K.Peters, Inhibition of tumor angiogenesis using a soluble re-ceptor establishes a role for Tie2 in pathologic vasculargrowth, J. Clin. Invest. 100 (1997) 2072^2078.
[44] J.A. McBain, A. Eastman, C.S. Nobel, G.C. Mueller, Apo-ptotic death in adenocarcinoma cell lines induced by buty-rate and other histone deacetylase inhibitors, Biochem. Phar-macol. 53 (1997) 1357^1368.
[45] T.C. Meng, M.S. Lee, M.F. Lin, Interaction between proteintyrosine phosphatase and protein tyrosine kinase is involvedin androgen-promoted growth of human prostate cancercells, Oncogene 19 (2000) 2664^2677.
[46] T.C. Meng, M.F. Lin, Tyrosine phosphorylation of c-ErbB-2is regulated by the cellular form of prostatic acid phospha-tase in human prostate cancer cells, J. Biol. Chem. 273(1998) 22096^22104.
[47] S. Mori, K. Murakami-Mori, B. Bonavida, Interleukin-6 in-duces G1 arrest through induction of p27(Kip1), a cyclin-dependent kinase inhibitor, and neuron-like morphology inLNCaP prostate tumor cells, Biochem. Biophys. Res. Com-mun. 257 (1999) 609^614.
[48] L.V. Nazareth, N.L. Weigel, Activation of the human an-drogen receptor through a protein kinase A signaling path-way, J. Biol. Chem. 271 (1996) 19900^19907.
[49] S.L. Parker, T. Tong, S. Bolden, P.A. Wingo, Cancer statis-tics, 1996, Ca Cancer J. Clin. 46 (1996) 5^27.
[50] Y. Qiu, L. Ravi, H.J. Kung, Requirement of ErbB2 forsignalling by interleukin-6 in prostate carcinoma cells, Na-ture 393 (1998) 83^85.
[51] Y. Qiu, D. Robinson, T.G. Pretlow, H.J. Kung, Etk/Bmx, atyrosine kinase with a pleckstrin-homology domain, is ane¡ector of phosphatidylinositol 3P-kinase and is involved ininterleukin 6-induced neuroendocrine di¡erentiation of pros-tate cancer cells, Proc. Natl. Acad. Sci. USA 95 (1998) 3644^3649.
[52] J.A. Rivero, S.E. Adunyah, Sodium butyrate stimulatesPKC activation and induces di¡erential expression of certainPKC isoforms during erythroid di¡erentiation, Biochem.Biophys. Res. Commun. 248 (1998) 664^668.
[53] D. Samid, A. Yeh, P. Prasanna, Induction of erythroid dif-ferentiation and fetal hemoglobin production in human leu-kemic cells treated with phenylacetate, Blood 80 (1992)1576^1581.
[54] J. Sap, P. D'Eustachio, D. Givol, J. Schlessinger, Cloningand expression of a widely expressed receptor tyrosine phos-phatase, Proc. Natl. Acad. Sci. USA 87 (1990) 6112^6116.
[55] R. Shen, T. Dorai, M. Szaboles, A.E. Katz, C.A. Olsson, R.Buttyan, Transdi¡erentiation of cultured human prostatecancer cells to a neuroendocrine cell phenotype in a hor-mone-depleted medium, Urol. Oncol. 3 (1997) 67^75.
[56] M.T. Spiotto, T.D. Chung, STAT3 mediates IL-6-inducedneuroendocrine di¡erentiation in prostate cancer cells, Pros-tate 42 (2000) 186^195.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^4342

[57] J. Su, L.T. Yang, J. Sap, Association between receptor pro-tein-tyrosine phosphatase RPTPalpha and the Grb2 adaptor.Dual Src homology (SH) 2/SH3 domain requirement andfunctional consequences, J. Biol. Chem. 271 (1996) 28086^28096.
[58] M. Tarle, N. Rados, Investigation on serum neurone-speci¢cenolase in prostate cancer diagnosis and monitoring: com-parative study of a multiple tumor marker assay, Prostate 19(1991) 23^33.
[59] D.A. Twillie, M.A. Eisenberger, M.A. Carducci, W.S. Hseih,W.Y. Kim, J.W. Simons, Interleukin-6: a candidate media-tor of human prostate cancer morbidity, Urology 45 (1995)542^549.
[60] W.G. van Inzen, M.P. Peppelenbosch, M.W. van den Brand,L.G. Tertoolen, S. de Laat, The role of receptor proteintyrosine phosphatase alpha in neuronal di¡erentiation ofembryonic stem cells, Brain Res. Dev. Brain Res. 91 (1996)304^307.
[61] M.H. Weinstein, A.W. Partin, R.W. Veltri, J.I. Epstein,Neuroendocrine di¡erentiation in prostate cancer: enhancedprediction of progression after radical prostatectomy, Hum.Pathol. 27 (1996) 683^687.
[62] X.H. Yang, K.T. Seow, S.M. Bahri, S.H. Oon, W. Chia,
Two Drosophila receptor-like tyrosine phosphatase genesare expressed in a subset of developing axons and pioneerneurons in the embryonic CNS, Cell 67 (1991) 661^673.
[63] M. Yoshida, M. Kijima, M. Akita, T. Beppu, Potent andspeci¢c inhibition of mammalian histone deacetylase both invivo and in vitro by trichostatin A, J. Biol. Chem. 265 (1990)17174^17179.
[64] S. Zelivianski, J. Dean, D. Madhavan, F.F. Lin, M.F. Lin,Expression of receptor protein tyrosine phosphatase alphamRNA in human prostate cancer cell lines, Mol. Cell. Bio-chem. 208 (2000) 11^18.
[65] L. Zeng, L. D'Alessandri, M.B. Kalousek, L. Vaughan, C.J.Pallen, Protein tyrosine phosphatase alpha (PTPalpha) andcontactin form a novel neuronal receptor complex linked tothe intracellular tyrosine kinase fyn, J. Cell Biol. 147 (1999)707^714.
[66] X.M. Zheng, C.J. Pallen, Expression of receptor-like proteintyrosine phosphatase alpha in rat embryo ¢broblasts acti-vates mitogen-activated protein kinase and c-Jun, J. Biol.Chem. 269 (1994) 23302^23309.
[67] X.M. Zheng, Y. Wang, C.J. Pallen, Cell transformation andactivation of pp60c-src by overexpression of a protein tyro-sine phosphatase, Nature 359 (1992) 336^339.
BBAMCR 14746 14-6-01 Cyaan Magenta Geel Zwart
S. Zelivianski et al. / Biochimica et Biophysica Acta 1539 (2001) 28^43 43




















