
Autonomic, Neuroendocrine, and Immune Responses to Psychological Stress: The Reactivity Hypothesisa
Upload
independentCategory
view
0download
0
Cellular and Molecular Neurobiologv Vol. 16, No. 3, 1996
Gonadal Steroid Modulation of Neuroendocrine Transduction: A Transynaptic View
Rafaei Alonso-Solfs , t''- Pedro Abreu, 1 Ignacio L6pez- Coviella, 1 Guadalberto Herntindez, 1 Natalia Fajardo, 1 Francisco Hermindez-Dfaz, t Aniria Dfaz-Cruz, t and Alex Hermindez 1
Received March 16, 1995; accepted April 19, 1995
KEY WORDS: pineal gland; GT~_7 cells; norepinephrine; tyrosine hydroxylase: melatonin: cAMP; estradiol; progesterone; testosterone; adrenergic receptors.
SUMMARY
1. Steroid hormones act on neuronal communication through different mechanisms, ranging from transynaptic modulation of neurotransmitter synthesis and release to development and remodeling of synaptic circuitry. Due the wide distribution of putative brain targets for steroid hormones, acute Or sustained elevations of their circulating levels may affect, simultaneously, a variety of neuronal elements. In an elementary mode of interaction, steroids are able to modulate both the synthesis and release of a neurotransmitter at a particular synapse, and the response of its target postsynaptic cells. Using two neuroendoc- rine transducing systems--the rat pineal gland and the GT;_7 cell line--we have examined these interactions and the following findings are discussed in this article.
2. In the rat, pineal melatonin production is partially controlled by gonadal hormones. In females, melatonin synthesis and secretion is reduced during the night of proestrus, apparently as a consequence of elevated estradiol and progesterone levels. In males, circulating testosterone seems to be necessary to maintain the amplitude of the nocturnal melatonin peak.
Laboratory of Neuroendocrinology, Department of Physiology, University of La Laguna School of Medicine, 38320 Tenerife, Spain.
"~To whom correspondence should be addressed at Departamento de Fisiologla, Facultad de Medicina, La Cuesta, 38320 Tenerife, Spain.
357 0272.434019610600.0357509.5010 ~) 1996 Plenum Publishing Corporation

358 Alonso-Solfs et al.
3. Some gonadal effects on pineal activity are exerted on its noradrenergic input, since changes in circulating steroid hormone levels are able to induce acute modifications of tyrosine hydroxylase activity in pineal sympathetic nerve terminals.
4. Gonadal steroids are also able to regulate the response of pineal cells to adrenergic stimulation, since in vivo treatment of both male and female rats with steroid hormone blockers induces profound modifications in adrenergically- induced accumulation of cyclic AMP (cAMP) in dispersed pinealocytes.
5. Direct exposure of pineal cells from gonadectomized female and male rats to estradiol (E~) or testosterone (T), respectively, potentiates pinealocyte response to adrenergic activation. In addition, short-term (15 min) exposure to either progesterone (Pg) or progesterone coupled to bovine serum albumin (P-3-BSA) suppresses the E2-dependent potentiation of adrenergic response in female rat pinealocytes.
6. Exposure of GT1-7 cells to E2 completely blocked the norepinephrine (NE)-induced elevation of cAMP content. In E2-treated GT1_7 cells, additional exposure (15 min) to either Pg or P-3-BSA abolished E2-dependent inhibition of NE responsiveness. In addition, P-3-BSA alone increased basal cAMP levels in GT~_7 cells, regardless previous exposure to E2.
7. In conclusion, there are evidences, both from the current literature and from the present results, supporting the view that in some neuroendocrine systems gonadal hormones modulate neurotransmission by acting, simultaneously, at pre- and postsynaptic sites. The models presented here constitute appropriate examples of this transynaptic mode of steroid action and, therefore, may offer a useful approach to investigate steroid hormone actions on the brain.
INTRODUCTION
Steroid hormone actions on neuronal communication are diverse, highly adapt- able, and integrative. This assumption is based on the multiple mechanism of steroid-cell interactions (McEwen, 1991; Wehling, 1994), the stage and time dependency of neuronal responses (Fink, 1988; Fink et al., 1991), and the wide range of functional categories in the expression of hormonal effects (Arnold and Breedlove, 1985; Jones and Pfaff, 1991; McEwen, this issue). In an attempt to develop a conceptual hierarchic approach, the following points should be considered: 1) Steroid hormones can regulate cell activity by interacting with, at least, two types of receptors, the classical and well known intracellular receptor (Evans, 1988) and novel membrane receptors, that either directly gate ion channels or are suspectedly coupled to second messenger systems (Wehling, 1994); 2) As a result, steroid-receptor interaction will produce rapid, nongenomic changes in neuronal excitability (Schumacher, 1990; Joels and De Kloet, 1991), or delayed, long-lasting effects on neuronal gene expression (Beato, 1989); in addition, since evidence exists for second messenger-related modulation of transcriptional processes (Hadcock and Malbon, 1991; Moyer et al., 1993), the possibility of cross-talk between genomic and nongenomic steroid effects is open

Steroids and Neuroendoerine Transduction 359
to exploration; 3). In terms of synaptic transmission, steroids are able to affect a variety of neuronal phenomena, ranging from transynaptic modulation of neurotransmitter synthesis and release (Etgen et al., 1992) to development and remodeling of synaptic circuitry (McEwen and Woolley, 1994); finally, 4) By acting at several levels of the nervous system, and through different molecular mechanisms, changes in circulating steroid hormone levels participate in the coordination of related behaviors and neuroendocrine events (McEwen et al., 1987; Pfaff, 1989).
Considering the wide distribution of putative neuronal targets for steroid action, it can be expected that acute or sustained elevations of adrenal or gonadal hormone levels would be able to act, simultaneously, on a variety of neuronal elements. This may constitute an effective and economic mechanism by which a circulating steroid molecule is utilized in the coordination of different synaptic pathways involved in the expression of a particular behavior, or in the regulation of several related neuroendocrine adaptations. Steroids are known to modulate neurotransmission at particular synapses by two different ways: 1) regulating neurotransmitter synthesis and release (Dluzen and Ramfrez, 1989; Drouva et al., 1983; Schumacher et al., 1989), or 2) controlling the expression of postsynaptic receptors (Hruska, 1986; Kukstas et al., 1991; Schumacher et al., 1990) and receptor sensitivity (Harrelson and McEwen, 1987; Lambert et al., this issue; Marius et al., 1989; Pettiti and Etgen, 1990). Thus, it would be interesting to investigate how the same steroid molecule modulates, at the same type of synapse, both pre- and postsynaptic events, as well as whether there is a functional relationship between the two responses. Therefore, it would be useful to have a simple system where to isolate pre- and postsynaptic steroid effects. In this article, we present studies on two such systems in which transynaptic gonadal steroid actions can be studied at the cellular level: the rat pineal gland and the GTI_7 cell line.
Neuroendocrine Transduction at the Rat Pineal Gland
Neuroendocrine transduction at the mammalian pineal gland constitutes an excellent model to study steroid effects of neurons, since it is constituted, basically, by a single presynaptic noradrenergic input acting on pinealocyte adrenoceptors. In most species, pineal noradrenergic transmission is accelerated during the night, causing a nocturnal rise in the production of melatonin (Reiter, 1991). In the rat, norepinephrine (NE), released from postganglionic sympathetic nerve terminals arising from the superior cervical ganglia (SCG), and acting on B- and al-adrenergic receptors in the pinealocyte membrane, initiates a series of events leading to melatonin synthesis (Fig. 1) (Sugden, 1989). B-adrenoceptor stimulation activates adenyl cyclase through a stimulatory guanine nucleotide- binding regulatory protein (Gs), resulting in a rapid increase in intraceUular cyclic AMP (cAMP) levels. Activation of rat pinealoeyte otradrenoceptors markedly potentiates B-receptor-induced accumulation of cAMP via a mechanism which involves elevation of intracellular Ca2÷ (Sugden et al., 1987), generation of diacylglycerol (DG) from phosphatidylinositol (PI) (Ho and Klein, 1987), and

ros,
oo
O
..
..
.
Targ
et ce
lls
( Fi
g. 1
. Sc
hem
atic
mod
el o
f po
tent
ial
site
s fo
r st
eroi
d ac
ticm
s on
pin
eal-
mel
aton
in s
yste
m.
Firs
t, st
eroi
d ho
rmon
es
coul
d re
gula
te
pres
ynap
tie
nora
dren
ergi
c in
0ut
by
chan
ges
in
TH
ge
ne
tran
scri
ptio
n (1
),
or
by
mod
ifyi
ng
pre-
exis
ting
TH
mol
ecul
es (
2).
Seco
nd,
ster
oids
mig
ht i
nter
act
with
pin
ealo
cyte
s at
pos
tjun
ctio
nal
leve
l, ei
ther
by
acti
ng o
n m
embr
ane
rece
ptor
s (3
), o
r by
act
ivat
ing
intr
acel
lula
r re
cept
ors
and
regu
lati
ng s
peci
fic
tran
scri
ptio
n pr
oces
ses
(4).
Fina
lly,
ste
roid
hor
mon
es m
ay m
odul
ate
mel
aton
in t
arge
t ac
tion
by
inte
ract
ing
with
its
rec
epto
rs (
5).
r~

Steroids and Neuroendocfine Transduction 361
translocation and activation of protein kinase C (PKC) (Sugden et al., 1985). lntracellular cAMP, as a second messenger, activates a cAMP-dependent protein kinase, increases mRNA transcription, and induces a transient rise in serotonin N-acetyltransferase (NAT), the rate-limiting enzyme in melatonin production (Klein et al., 1983). Recent data have shown that the pineal gland contains an isoform of CREM (cAMP-responsive element modulator) gen, named ICER (inducible cAMP early repressor), which acts as a potent repressor of gene transcription induced by cAMP (Stehele et al., 1993). Since the expression of this gene, which displays a circadian rhythmicity, is under adrenergic control, it could be responsible for the reduction of NAT activity following several hours of adrenergic stimulation.
In addition to the above mentioned adrenergic control, it has been shown in several species that physiological oscillations of gonadal steroids, or those induced by experimental manipulations, also affect pineal melatonin synthesis and secretion (Ozaki et al., 1978; Hern~indez et al., 1990a; Moujir et aL, 1990; Alonzo et al., 1993). Even though the real real nature of steroid-induced actions on pineal activity remains to be completely established, theoretically gonadal or adrenal hormones may act at different sites in the pineal-melatonin system (Fig. 1), including the interactions of melatonin with their target ceils (Vanecek et al., 1990; Zisapel and Anis, 1988; Zisapel et al., 1987). The studies presented here show that, in male and female rats, gonadal steroid hormones modulate both norepinephrine synthesis in pineal sympathetic terminals, and the response of pineal cells to adrenoceptor activation.
Neuroendocrine Transduction at the GnRH-Producing Neuronal Cell Line
The synthesis and secretion of GnRH is controlled by several neurotransmit- ter systems acting on neurons in the hypothalamus, with their own intrinsic oscillatory properties (Kordon et aL, 1994), which respond to the strong feedback exerted by gonadal steroids (Fink, 1988; Fink, 1994). Until very recently, it was considered that GnRH neurons do not contain estrogen receptors (Shivers et al., 1983), which lead to the assumption that hormone actions were mediated by interneurons. However, the recent report of estrogen receptor expression in an immortalized cell line (GT1) producing GnRH (Polletti et al., 1994), as well as its functional activation by estradiol (Negro-Vilar et al., this issue), together with the potential membrane effects of steroid hormones (Rarnirez, this issue), have completely changed the picture. Even though results obtained from tumoral cell lines must be interpreted cautiously, the possibility that GnRH neurons can respond to gonadal steroids through active receptors make these cells appropriate candidates for the study of steroid transynaptic effects. On the other hand, GT1 cells have provided an optimal model for the study of different aspects of GnRH regulation (Mellon et al., 1990; Weiner et al., 1992 ). These cells, cultured on coverslips, show oscillatory basal GnRH release which depends on voltage- sensitive Ca 2÷ influx (Krsmanovic et al., 1992), and whose synchronous activity appears to me mediated by cell-to-cell-contracts and the paracrine action of diffusible mediators (Martinez de la Escalera, 1992a). In addition, these cells

362 Alonso-Solfs et aL
express functional receptors for several neurotransmitters, which include norepi- nephrine (Martinez de la Escalera et al., 1992b), dopamine (Martfnez de la Escalera et al., 1992c), GABA (Martfnez de la Escalera et aL, 1994), glutamate (Spergel et aL 1994), or even GnRH (Cesnjaj et aL 1993). Therefore, we have decided to use these neurons as another model to investigate the effects of gonadal steroids on transynaptic responses. In a preliminary experiment, we report here the effects of oestradiol and progesterone on NE-induced cAMP accumulation in GT1 cells.
METHODS
Animals and Experimental Manipulations
Adult female and male Sprague-Dawley rats (175-200 g), housed in groups of 3-5 per cage, were maintained in a temperature-controlled room (23 + 2°C) under a light-dark cycle of 12:12 (60-80 ~Wcm -2) for at least 1 week before experimental manipulations. In experiments with female rats, vaginal smears were examined daily, and animals showing at least two consecutive 4-day estrus cycles were used. Gonadectomy was performed under ether anaesthesia on dates indicated for each particular experiment. Acute steroid injections were given subcutaneously (sc)in 0.2 ml of olive oil at times indicated. In male rats, chronic testosterone treatment was performed by means of testosterone propionate (TP)-filled silastic capsules (2 × 30 mm), implanted sc at the time of orchidec- tomy. In experiments with hormone blockers, normal cyclic female rats were injected at proestrus, 7 and 2 hours before the onset of darkness, with 4 mg RU486 (1 l13-[4-dimethylaminophenyl]-176-hydroxy-17(3-[prop-l-ynyl]-estra-9,9- dien-3-one), or with i mg Tamoxifen ([2]l-[p-dimethylaminoethoxyphenyl]l,2- diphenyl-l-butene) in 0.2 ml of olive oil as the vehicle. In the case of male rats, they were treated with daily injections of 10mg/kg Flutamide (4'-nitro-3'- trifluoromethyl-isobutyranilidine) during 10 days, or with 70mg/kg ethylene dimethane sulfonate (EDS, ethylene 1,2-dimethane) 1 week before sacrifice. In all experiments, animals were sacrificed by decapitation at the times indicated, and their pineals and SCG quickly dissected out and either stored frozen at -17°C until assays, or processed for pinealocytes incubation as described below.
Cell Cultures
Pineal cells were dispersed by trypsinization and DNA-ase treatment according to previously described procedures (Buda and Klein, 1978). Pineal- ocytes (200,000 cells m1-1) were plated onto 3 5 x 1 0 m m culture dishes and incubated under humidified atmosphere (95% CO2/5% O2) at 37°C in Dulbecco's modified Eagle's medium (DMEM) containing fetal bovine serum (FBS, 10%), L-Glutamine (1%), Penicillin G (60/zg/ml, and Streptomycin (10/zg/ml). Eighteen hours later, after complete degeneration of sympathetic nerve terminals, pinealocytes were transferred to polypropylene tubes (100,000 cells tube -1) and treated with 0.i mM 3-isobutyl-l-methylxantine (IBMX, a phosphodiesterase

Steroids and Neuroendocrine Transduction 363
inhibitor) for 60m in. Thereafter, they were treated with either the vehicle (water), 1 jzM isoprenaline (ISO, a B-agonist), 1/zM phenylephrine (PHE, an oq-agonist), or both (1/~M each) for 15rain. Ceils were then collected by centrifugation (10,000 x g, 30 s), homogenized by sonication in 100 ~I of 50 nM acetate buffer (pH 4.0), and incubated in a water bath at 90°C for 3 min. After centrifugation, cells were frozen at -70°C until assayed for cAMP. In experi- ments designed to evaluate direct steroid effects on pinealocyte response to adrenergic stimulation, pineal cells from gonadectomized rats were cultured for 5 days in DMEM as described above and then changed to DMEM containing steroid-free (Dextran-Charcoal-Stipped) FBS. Pinealocytes from ovariectomized female rats were exposed to 17B-estradiol (E2, 100nM) or the vehicle for 48 hours, and then to either progesterone (Pg) or progesterone conjugated to bovine serum albumin at carbon 3 (P-3-BSA)(20 nM each) for 15 min. Pineal- ocytes from orchidectomized male rats were exposed to testosterone (T) or E2 (100nM each) or vehicle for 48 hours. After steroid treatment, cells were stimulated with adrenergic agonists and processed as described above.
GTI_7 cells, generously provided by Dr. P. Mellon (San Francisco, CA) through Dr. A. Negro-Vilar (Philadelphia, PE), were grown in 35 × 10 mm tissue culture dishes. The basic medium was DMEM supplemented with 10% FBS, Glucose (4.5 mg/ml), Penicillin G (100U/ml), and Streptomycin (100/~g/ml). When they were 70% confluent, the medium was changed to phenol red-free DMEM containing 10% DCC-FBS and ceils cultured for 5 more days. On the third day, half of the dishes were supplemented with E2 (100 nM) or vehicle for 48 h. On the fifth day, some cells were additionally exposed to Pg, P-3-BSA (20 nM each), or vehicle for 15 min. Ten rain before harvesting cells were treated with 10 ~M NE or vehicle. After aspiration of the medium cells, were collected in 100/~1 of 50nM acetate buffer (pH4.0), homogenized by sonication, and incubated in a water bath at 90°C for 3 min. After centrifugation, cells were frozen at -70°C until assayed for cAMP content.
Biochemical Measurements
For determination of pineal melatonin content and TH activity, individual glands were homogenized by sonication (80 W, 15 s) in 200/zl of 0.25 M sucrose. Analysis were performed by high-performance liquid chromatography and fluorescence or electrochemical detection, respectively, as previously described (Abreu et al., 1987; Hem~indez et al., 1990b). Pineal TH content was determined by western blot analysis using a monoclonal anti-TH antibody (Boheringer, Germany) as described elsewhere (HernLndez et aL, 1994; Laemmli, 1970). Total RNA was isolated following standard procedures (Chomcynski and Sacchi, 1986). The eDNA was synthesized from approximately 2/zg of total RNA and amplified by PCR. The products of PCR, yielding a 272 bp fragment as that published previously for TH eDNA (Grima et al., 1985), were electrophoresed on polyacrylamide gels and measured by densitometry using an image analysis system (GS-670, BioRad, CA). Pinealocytes and GT1 cells cAmP content following adrenergic stimulation was determined in duplicate after acetylation, using an enzyme immunoassay kit (EIA RPN225, Amersham, UK).

364 Alonso-Soifs et al.
Chemicals
Most chemicals and reagents were obtained from Sigma Chemical Co. (Fancy Road, Poole, UK)~ RU486 was obtained from Dr. R. Deraedt (Roussel-Uclaf, Romainville, France) through Dr. J. E. S~inchez-Criado. EDS was synthesized according to standard procedures described elsewhere (Gaytan et aL, 1990).
Statistics
Quantitative data are expressed as Mean + SEM of 8-10 determinations. Significant differences between groups were evaluated by two-way analysis of variance, and post -hoe comparisons were made by Newmann-Keuls and Tukey tests. In some cases, due to large heterogeneity of variance size, log cAMP values were used for calculations.
RESULTS
Gonadal Effects on Pineal Melatonin Production
Studies in Female Rats
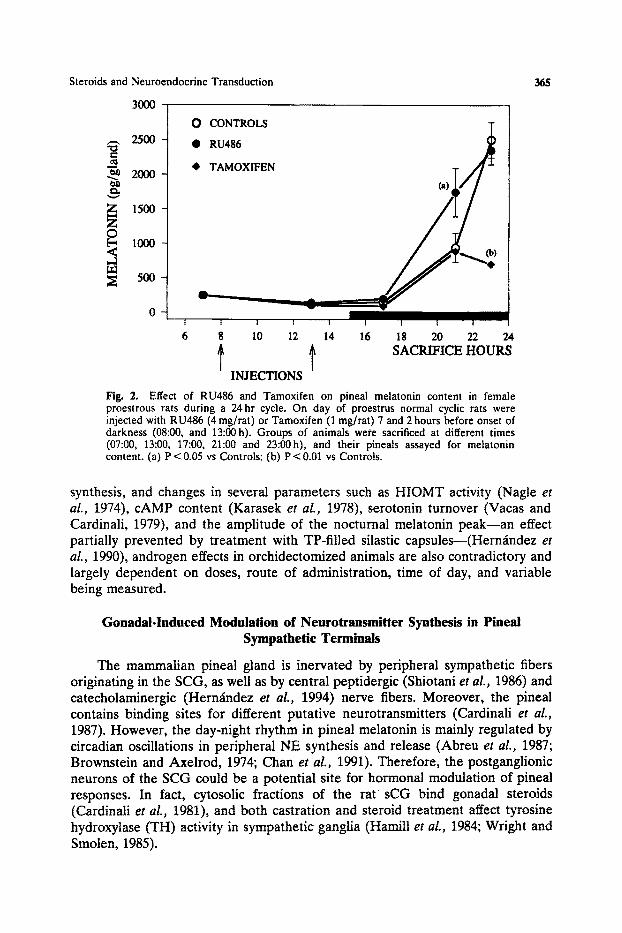
Even though analysis of hormonal-induced changes of pineal activity during female reproductive cycle is rather complicated, experimental evidence has been obtained in several species, including the human (Alonso et al., 1993; Brzezinski et al., 1988). In the rat, several biochemical parameters related to pineal melatonin synthesis have been shown to change during the estrous cycle. One of the most consistent findings is a reduction in pineal and urinary melatonin content during the night of proestrus, coinciding with high levels of E2 and Pg (Ozaki et al., 1978; Moujir et al., 1990). Since cyclic changes in urinary melatonin are not a reflection of hormone modifications in peripheral metabolism or renal clearance, it seems likely that ovarian hormones may exert an inhibitory action directly on pineal melatonin synthesis and secretion. It was shown tha t ovariectomy increases serum melatonin levels, and that administration of both E2 and Pg to ovariectomized rats reduced them to those of intact controls (Ozaki et al., 1978). In addition, treatment of ovariectomized animals with E2 and/or Pg completely blocks the isoproterenol-induced elevation of pineal melatonin content (Moujir et al., 1990). To further investigate the relationship between ovarian hormones and pineal melatonin, we treated normal cyclic rats with the antiestrogen Tamoxifen (1 mg/rat), the antiprogestagen RU486 (4 rag/rat), or vehicle, on day of proestrus and 7 and 2 hours before the onset of darkness. Animals were sacrificed at different times during a 25-hour cycle and their pineals analyzed for melatonin content. Figure 2 shows that RU486 accelerated the physiological nocturnal rise in pineal melatonin levels, while Tamoxifen seemed to restrain it.
Studies in Male Rats
In male rats, functional relationships between normal circulating levels of gonadal hormones and pineal activity are not clear and controversial results have been frequently reported (Alonso et al., 1993). Although most data indicate that castration of male rats causes a reduction in the rate of pineal melatonin
,-, 2500
~ 2000
v
Z 1500
O
O CONTROLS
- • RU486
• TAMOXIFEN
I
6
A
V
I I I
1 g 20 22 24 SACRIFICE HOURS
Fig. 2.
I I I I 1
8 10 12 14 16
t INJECTIONS
Steroids and Neuroendocrine Transduction
3OOO
Effect of RU486 and Tamoxifen on pineal melatonin content in female proestrous rats during a 24hr cycle. On day of proestrus normal cyclic rats were injected with RU486 (4 mg/rat) or Tamoxifen (1 mg/rat) 7 and 2 hours before onset of darkness (08:00, and 13:00h). Groups of animals were sacrificed at different times (07:00, 13:00, 17:00, 21:00 and 23:00h), and their pineats assayed for melatonin content. (a) P < 0.05 vs Controls; (b) P < 0.01 vs Controls.
365
synthesis, and changes in several parameters such as HIOMT activity (Nagle et al., 1974), cAMP content (Karasek et al., 1978), serotonin turnover (Vacas and Cardinali, 1979), and the amplitude of the nocturnal melatonin peak--an effect partially prevented by treatment with TP-filled silastic capsules--(Hern~ndez et al., 1990), androgen effects in orchidectomized animals are also contradictory and largely dependent on doses, route of administration, time of day, and variable being measured.
Gonadal-Induced Modulation of Neurotransmitter Synthesis in Pineal Sympathetic Terminals
The mammalian pineal gland is inervated by peripheral sympathetic fibers originating in the SCG, as well as by central peptidergic (Shiotani et al., 1986) and catecholaminergic (Hern~ndez et al., 1994) nerve fibers. Moreover, the pineal contains binding sites for different putative neurotransmitters (Cardinali et al., 1987). However, the day-night rhythm in pineal melatonin is mainly regulated by circadian oscillations in peripheral NE synthesis and release (Abreu et al., 1987; Brownstein and Axelrod, 1974; Chanet al., 1991). Therefore, the postganglionic neurons of the SCG could be a potential site for hormonal modulation of pineal responses. In fact, cytosolic fractions of the rat sCG bind gonadal steroids (Cardinali et aL, 1981), and both castration and steroid treatment affect tyrosine hydroxylase (TH) activity in sympathetic ganglia (Hamill et al., 1984; Wright and Smolen, 1985).
1.0 .8
o .6
,
m
c
O
...
< 1-
Z
k-
.4
0E
E
.2
0.0
SC
G T
H m
RN
A R
T-P
CR
ana
lysi
s
OV
X
OV
X
+ E
2
OV
X
OV
X
+ E
2
+P
Act
in
297
bp
TH
27
2 b
p
+P
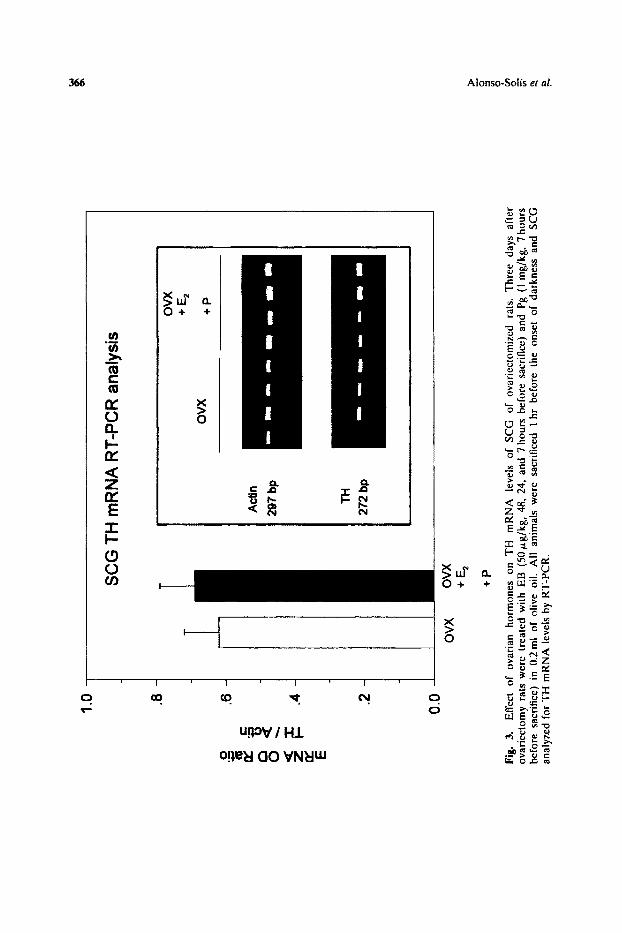
Fig.
3.
E
ffec
t of
ov
aria
n ho
rmon
es
on
TH
m
RN
A
leve
ls
of
SC
G
of
ovar
iect
omiz
ed
rats
. T
hree
da
ys
afte
r ov
arie
ctom
y ra
ts w
ere
trea
ted
wit
h E
B
(50
#g/k
g, 4
8, 2
4, a
nd 7
hou
rs b
efor
e sa
crif
ice)
and
Pg
(1
mg/
kg,
7 ho
urs
befo
re
sacr
ific
e) i
n 0.
2ml
of o
live
oil
. A
ll
anim
als
wer
e sa
crif
iced
1
hr
befo
re
the
onse
t of
dar
knes
s an
d S
CG
an
alyz
ed f
or T
H m
RN
A l
evel
s by
RT
-PC
R.
> =1

A 'ID
r-
~w
O
• r E
F-
e~
(a)
TH
CO
NT
EN
T
ov
x
ov
x
+P
Kd
56
-60
>
OV
X
OV
X
+E2
÷P
Fig,
4.
Eff
ect
of o
vari
an h
orm
ones
on
TH
act
ivit
y an
d qu
anti
ty i
n pi
neal
gla
nds
of o
vari
ecto
miz
ed r
ats.
Pi
neal
s w
ere
obta
ined
fro
m t
he e
xper
imen
tal
anim
als
desc
ribe
d in
Fig
. 3
and
assa
yed
for
TH
act
ivit
y an
d qu
anti
ty b
y H
PL
C a
nd W
este
rn-b
lot,
res
pect
ivel
y. (
a) P
< 0
.01
vs O
vx.
o.
13
.
2:
=1 o ,.q
t-
-i

368 Alonso-Solfs et al.
Studies in Female Rats
To investigate whether ovarian hormones affect NE synthesis in pineal sympathetic terminals, normal cyclic rats were ovariectomized and treated with estradiol benzoate (EB; 50 mg/kg; 2 and 1 days, and 7 hours before sacrifice) plus Pg (1 mg/kg; 7 hours before sacrifice). All animals were sacrificed 1 hour after the onset of darkness, and their pineals and SCG quickly removed and processed as described in methods section. Measurements of TH mRNA levels in SCG, by RT-PCR, and pineal TH contefit, by western blot, showed no effect of ovarian hormones (Figs 3 and 4b). However, pineal TH activity in ovariectomized animals was enhanced after treatment with EB and Pg (Fig. 4a). The magnitude of the variation in TH activity, although small, is quantitatively similar to that observed in the rat pineal between day and night (Abreu et aL, 1987), and hence sufficient to produce large and rapid changes in the rate of NE synthesis in pineal sympathetic terminals.
Studies in Male Rats
Male rats were subjected to orchidectomy and testosterone replacement by two different regimens: (A) TP-filled silastic capsules at the time of gonadectomy; and (B) three daily TP injections (40 tzg, sc, at 11:00, 15:00, and 19:00 h, starting with the gonadectomy. Rats were sacrificed 10 days after castration at different hour intervals, and their pineals assayed for TH activity. Figure 5 shows a
25
• ~ 20
15
10
o
Fig. 5.
0,)
CO Cx Cx + Tp
Effect of castration and testosterone treatment on day-night rhythms of pineal TH activity. Male rats were orchideetomized and implanted with testosterone propionate (TP)-filled silastic capsules (2 x 30 mm). Ten days later they were sacrificed at different times of the 24-hour cycle and their pineals assayed for TH activity. Since there were not differences depending of the light or dark period, values of several times are pooled in the figure. (a) P<0.05 vs Light; (b) P<0.01 vs CO-dark.

Steroids and Neuroendocrine Transduction 369
20
< " ~ 10
[-' ,-1 5
& 0
A
- O - . . . . <D - ' 0 " . . . . O - . . . . < > , - O - . . . . < 3 - . . . . O - . . . . <3,- . . . . 0
I I I } I I I I |
08 12 16 20 24 04 08 12 16
2O 0
10 ~'~
~2 ~q 0
0 ~q
3O
> , ~ 25
2. - r..) 15
~ 10 b., , ,.d
5 • o
0
, , (a) t i I e ,
: I ' I ira(a) . . I i a i t
• s s , I I
• w I • • I 1 i e I I
c---d" ~ 6 ' c- C" . . . . . . . . O- . . . . . . 0
I I I I I I I
08 12 16 20 24 04 08
TP INJECTIONS
25 O
>
10 --
,--t 5 0
,.-1
0 0 Z
Fig. 6. Effect of TP injections on TH activity and plasma testosterone levels of castrated male rats. Orchidectomized rats received three daily injections or oil vehicle (A) or 40/~g TP (B), at 11:00, 15:00, and 19:00 h, during ten consecutive days. Groups of animals were sacrificed at the times indicated, and their pineals and trunk blood assayed for TH activity and testosterone concentration, respectively. (a) P < 0.01 vs 14:00, 18:00, 22:00 h.
decrease in nocturnal TH activity, which abolishes the day-night rhythm. Treatment with TP-silastic capsules was unable to restore normal day-night rhythm, even though it elevated, both day and night values of enzyme activity. Moreover, when testosterone was given as daily injections for 10 days, pineal TH activity displayed acute oscillations apparently related to those of serum testosterone concentration (Fig. 6a, b). Since maximal levels of pineal TH activity appeared in animals killed at 16:00, 20:00, and 24:00 h, and peak values of serum T were shown at 12:00, 16:00, and 20:00 h, we can not conclude that TP injections were stimulatory nor inhibitory, and little could be said about the temporal relationships with pineal TH activity.
These data show that changes in circulating levels of gonadal hormones,

370 Alonso-Sol~etaL
similar to those occurring under physiological circumstances, may induce acute variations in pineal TH activity. In other neuronal systems, these changes are presumably produced via two different mechanisms: a) By changes in TH gene transcription and modulation of de novo synthesis of enzyme molecules (Blum et al., 1987); or b) By activation/deactivation of pre-existing TH molecules (Pasqualini et al., 1991), an effect that may involve changes in TH phosphoryla- tion by a direct nongenomic estrogenic action (Pasqualini et al., 1993; this issue). Therefore, even though modifications of enzyme synthesis can not entirely be ruled out from our experiments, it seems that gonadal steroids induced changes in the activity of existing TH molecules in pineal sympathetic nerve terminals, which in turn could influence the noradrenergic input received by pineal cells.
Gonadal.Induced Modulation of Pinealocyte Response to Adrenoceptor Action
In some brain areas, the interaction of neurotransmitters with their mem- brane receptors can be modified by previous exposure to gonadal hormones, both, in vivo (Pettiti and Etgen, 1990), and in vitro (Mauss et al., 1989). Moreover, earlier on, it was found that treatment of ovariectomized rats with estradiol inhibited the activation of adenyl cyclase by NE (Weiss and Crayton, 1970). Since NE-dependent increase in pineal melatonin synthesis is initiated with elevations of intracellular cAMP, we decided to investigate whether adrenergically-induced cAMP levels in pinealocytes, in vitro were influenced by previous treatment with gonadal hormone blockers, in vivo.
Adrenergic Stimulation o f Female Rat Pinealocytes
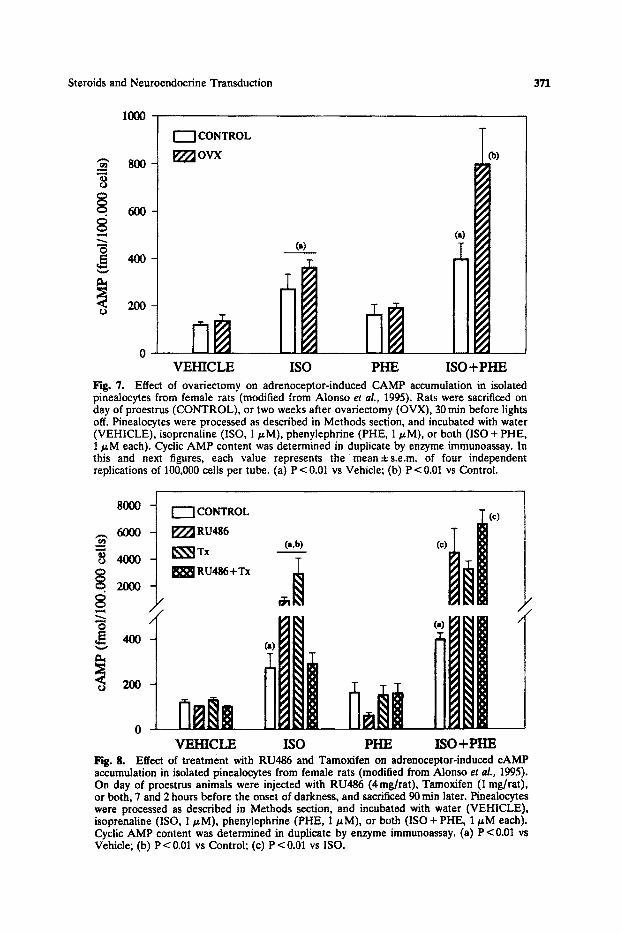
Sexually mature female rats were monitored for estrous cyclicity or ovariectomized two weeks before the experiment. Coinciding with the proestrus of control animals, and 7 and 2 hours before the onset of darkness, all animals were treated with RU486 (4 mg/rat), Tamoxifen (1 mg/rat), RU486 plus Tamoxi- fen, or vehicle (0.2 ml oil). Ninety min later rats were sacrificed and their pineals quickly dissected out and dispersed by trypsinization. Pinealocytes were incubated as described in Methods section, and cAMP levels determined after treatment for 15 min with 1 tzM ISO, PHE, or both. ISO, but not PHE, significantly increased cAMP levels regardless of gonadal status. The combination of ISO and PHE was unable to further modify the effect of ISO alone in control proestrous rats, but enhanced it by 2-fold in ovariectomized animals (Fig. 7). Treatment with RU486 or Tamoxifen, but not together, caused a significant potentiation of the ISO-induced accumulation of cAMP (Fig. 8). These results show that ctl- potentiation of B-stimulation of pineaolocyte adenyl cyclase is absent during the rat proestrous night (coinciding with high levels of E2 and Pg), which is in good agreement with the reduction of pineal melatonin synthesis observed during that period. Since treatment of normal cyclic rats with RU486 dramatically potentiated the oq-augmentation of B-induced cAMP accumulation, it may be suggested that circulating Pg exerts an inhibitory effect on a~- and B-receptor synergism. It
Steroids and Neuroendocrine Transduction
IOOO
371
r ~
8
[""]CONTROL
177~OVX 8OO
6 0 0 -
4 0 0 -
2OO
b)
0 V E H I C L E ISO P H E ISO + P H E
Fig. 7. Effect of ovariectomy on adrenoceptor-induced CAMP accumulation in isolated pinealocytes from female rats (modified from Alonso el al., 1995). Rats were sacrificed on day of proestrus (CONTROL), or two weeks after ovariectomy (OVX), 30 rain before lights off. Pinealocytes were processed as described in Methods section, and incubated with water (VEHICLE), isoprenaline (ISO, 1 p.M), phenylephrine (PHE, 1 p.M), or both (ISO + PHE, 1 ~M each). Cyclic AMP content was determined in duplicate by enzyme immunoassay. In this and next figures, each value represents the mean±s.e.m, of four independent replications of 100,000 cells per tube. (a) P < 0.01 vs Vehicle; (b) P < 0.01 vs Control.
80O0 CONTROL 0 ~ . 6000 [77~ RU486
"" ~ Tx (a,b) (¢)
"~ 4000 ~ RU486+Tx
ilia ilii o V E H I C L E ISO P H E I S O + P H E
Hg. 8. Effect of treatment with RU486 and Tamoxifen on adrenoceptor-induced cAMP accumulation in isolated pinealocytes from female rats (modified from Alonso et al., 1995). On day of proestrus animals were injected with RU486 (4rag/rat), Tamoxifen (1 rag/rat), or both, 7 and 2 hours before the onset of darkness, and sacrificed 90 rain later. Pinealocytes were processed as described in Methods section, and incubated with water (VEHICLE), isoprenaline (ISO, 1/.tM), phenylophrine (PHE, 1/~M), or both (ISO + PHE, 1/zM each). Cyclic AMP content was determined in duplicate by enzyme immunoassay. (a) P < 0.01 vs Vehicle; (b) P < 0.01 vs Control; (c) P < 0.01 vs ISO.
372 Alonso-Solfs et al.
should be pointed out that a similar situation occurs in slices from preoptic area and medial hypothalamus of estrogen-primed rats, where Pg abolishes the same response to NE (Pettiti and Etgen, 1990). Unfortunately, the effects of Tamoxifen can not be completely explained from these results. Treatment with Tamoxifen alone enhanced ISO-induced cAMP levels, which could be interpreted as a suppression of the inhibitory action of E2 on pineal B-receptors. However, when RU486 and Tamoxifen were given together, no effect at all on the B-response was observed, while al-potentiation of the B-response was still present. Whether or not these effects of Tamoxifen reflect some interference between its anti-estrogenic and estrogenic actions--that Tamoxifen may behave as a pure estrogen antagonist in some systems (Sutherland et al., 1977), and as a full agonist in others (Harper and Walpole, 1967)--, or they simply reflect an interaction between E2 and Pg on pineal gland regulation, awaits further investigation.
Adrenergic Stimulation o f Male Rat Pinealocytes
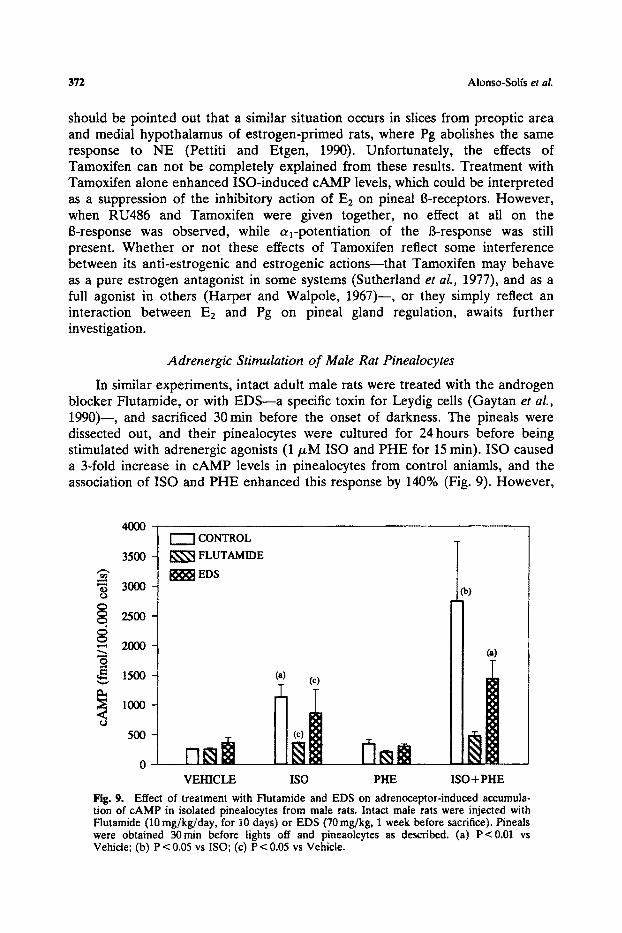
In similar experiments, intact adult male rats were treated with the androgen blocker Flutamide, or with EDS--a specific toxin for Leydig cells (Gaytan et al., 1990)--, and sacrificed 30 min before the onset of darkness. The pineals were dissected out, and their pinealocytes were cultured for 24 hours before being stimulated with adrenergic agonists (1/zM ISO and PHE for 15 rain). ISO caused a 3-fold increase in cAMP levels in pinealocytes from control aniamls, and the association of ISO and PHE enhanced this response by 140% (Fig. 9). However,
8 "6
4000
3500
3000
2500
2000
1500
1000
500
0
CONTROL
FLUTAMIDE
EDS
(a) (c)
VEHICLE ISO PHE
(i)
0i ISO + PHE
Fig. 9. Effect of treatment with Flutamide and EDS on adrenoceptor-induced accumula- tion of cAMP in isolated pinealocytes from male rats. Intact male rats were injected with Flutamide (10 mg/kg/day, for 10 days) or EDS (70 mg/kg, 1 week before sacrifice). Pineals were obtained 30m in before lights off and pineaolcytes as described. (a) P<0.01 vs Vehicle; (b) P < 0.05 vs ISO; (c) P < 0.05 vs Vehicle.

Steroids and Neuroendocrine Transduction 373
in pinealocytes from aninals that had been treated with Flutamide or EDS, cAMP accumulation in response to ISO was reduced, and the at-potent iat ion of the B-response was suppressed.
Direct Effects of Gonadal Steroids on Pinealocyte Adrenoceptor Activation
Although the above data showed that, both, in male and female rats, gonadal hormone levels may modulate pinealocyte sensitivity to adrenergic stimulation, it remained unclear whether this was the result of an indirect action on norepinephrine synthesis and release from sympathetic nerve terminals which, in turn could mediate up- or down-regulation of pineal adrenoceptors. To investigate whether steroid hormones may affect pinealocyte responsiveness by a direct action, cAMP levels were measured in cultured pinealocytes after a l - and B-receptor activation, in the presence or absence of gonadal steroids in the medium. Pineal glands from male or female gonadectomized rats were obtained 1 hour before the onset of darkness, and their pinealocytes incubated for 6 days in steroid-free medium and then exposed to gonadal steroids and to adrenergic agonists as described in methods section. Treatment with ISO caused a 40-fold increase in cAMP levels in pinealocytes from rats of either sex, and treatment with ISO plus PHE enhanced it further by 50% (Figs. 10 and 11). In pinealocytes from ovariectomized female rats, exposure to E2 48 hours prior of adrenergic stimulation enhanced the response to ISO, alone or in combination with PH E. Moreover , additional exposure to either Pg of P-3-BSA, 15 rain before adrenergic stimulation, entirely suppressed E2-induced effect on cAMP levels (Fig. 10). In
1200
1000
o 200
I I VEHICLE (a)
\ ISO
ISO+PHE
CONTROL E x E2+Pg E2+Pb-BSA
Fig. 10. Effect of in vitro exposure to ovarian steroids on adrenoceptor-induced cAMP accumulation in isolated pinealocytes from female ovariectomized rats. Animals were sacrificed 30rain before lights off and pinealocytes processed as described in Methods section and incubated with E2 (100 riM), for 48 hours, Pg (20 nM), or P-3-BSA (20 nM), for 15 min each. Cyclic AMP levels were determined after 15 rain exposure to 1 p_M ISO, PHE, or both. (a) P < 0.01 vs Control.
374 Alonso-Solfs et aL
800 ~ | VEHICLE
[ N-~,so t~c*) i ~ ISO+ PHE (b) = 600
400
0
o CONTROL T E 2
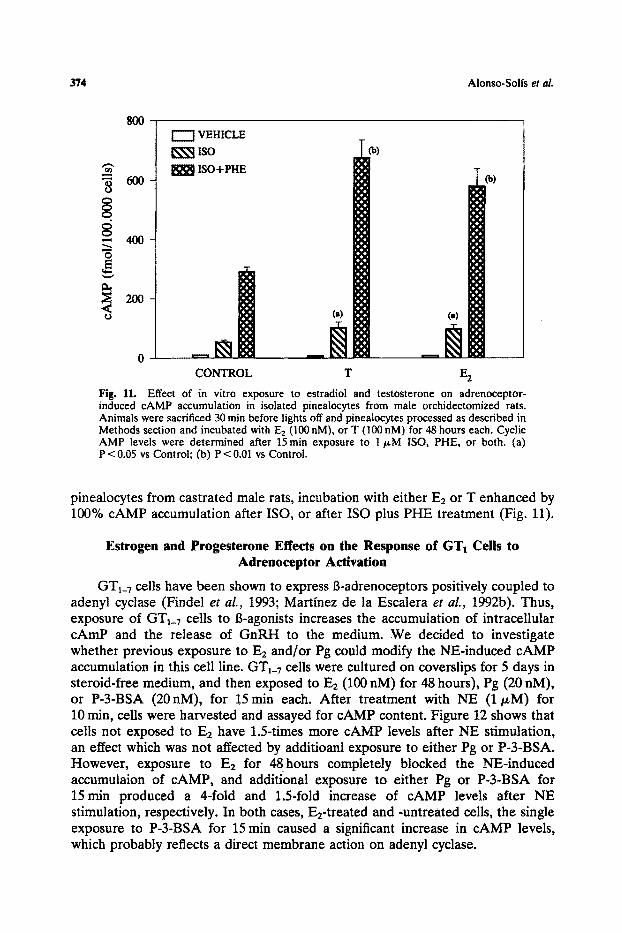
Fig. 11. Effect of in vitro exposure to estradiol and testosterone on adrenoceptor- induced cAMP accumulation in isolated pinealocytes from male orchidectomized rats. Animals were sacrificed 30 rain before lights off and pinealocytes processed as described in Methods section and incubated with E2 (100 nM), or T (100 nM) for 48 hours each, Cyclic AMP levels were determined after 15rain exposure to 1 t~M ISO, PHE, or both. (a) P < 0.05 vs Control; (b) P < 0.01 vs Control.
pinealocytes from castrated male rats, incubation with either E2 or T enhanced by 100% cAMP accumulation after ISO, or after ISO plus PHE treatment (Fig. 11).
Estrogen and Progesterone Effects on the Response of GT1 Cells to Adrenoceptor Activation
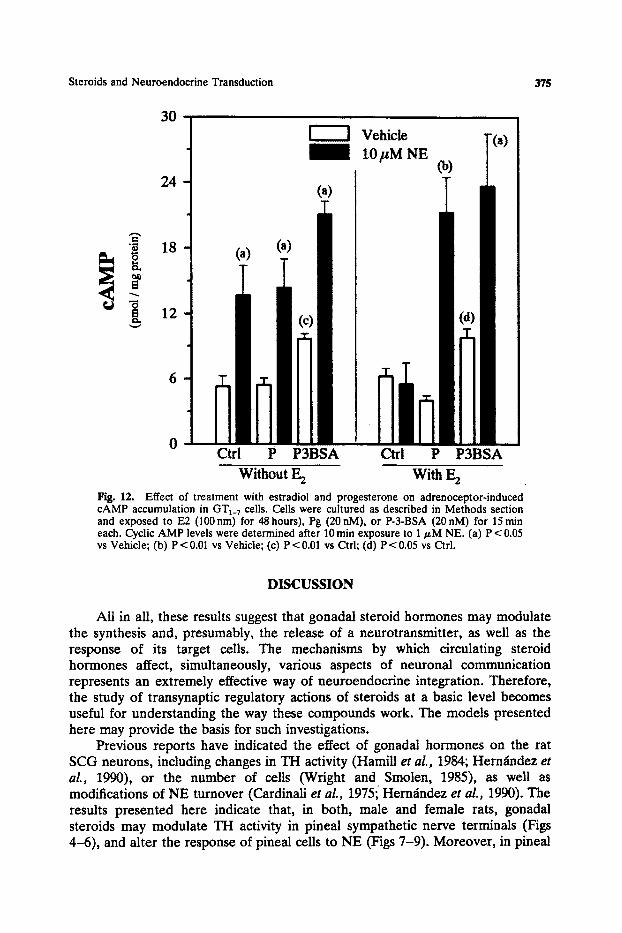
GTt_7 cells have been shown to express B-adrenoceptors positively coupled to adenyl cyctase (Findel et aL, 1993; Martinez de la Escalera et aL, 1992b). Thus, exposure of GTI_7 cells to B-agonists increases the accumulation of intracellular cAmP and the release of GnRH to the medium. We decided to investigate whether previous exposure to E2 and/or Pg could modify the NE-induced cAMP accumulation in this cell line. GT1_7 ceils were cultured on coverslips for 5 days in steroid-free medium, and then exposed to E2 (100 nM) for 48 hours), Pg (20 nM), or P-3-BSA (20nM), for 15min each. After treatment with NE (1/~M) for 10 min, cells were harvested and assayed for cAMP content. Figure 12 shows that cells not exposed to E2 have 1.5-times more cAMP levels after NE stimulation, an effect which was not affected by additioanl exposure to either Pg or P-3-BSA. However, exposure to E2 for 48hours completely blocked the N-E-induced accumulaion of cAMP, and additional exposure to either Pg or P-3-BSA for 15m in produced a 4-fold and 1.5-fold increase of cAMP levels after NE stimulation, respectively. In both cases, E2-treated and -untreated cells, the single exposure to P-3-BSA for 15 min caused a significant increase in cAMP levels, which probably reflects a direct membrane action on adenyl cyclase.
Steroids and Neuroendocrine Transduction 375
30
24
6
0
Vehicle T(a)
, t 0 p M N E (b)
(a)
~'a~ (a)
Ctrl P P3BSA nin Ctrl P P3BSA
W i t h o u t E 2 With E 2
Fig. 12. Effect of treatment with estradiol and progesterone on adrenoceptor-induced cAMP accumulation in GT1_7 cells. Cells were cultured as described in Methods section and exposed to E2 (100 nm) for 48 hours), Pg (20 nM), or P-3-BSA (20 nM) for 15 rain each. Cyclic AMP levels were determined after 10 min exposure to 1/zM NE. (a) P < 0,05 vs Vehicle; (b) P < 0.01 vs Vehicle; (c) P < 0.01 vs Ctd; (d) P < 0.05 vs Ctrl.
DISCUSSION
All in all, these results suggest that gonadal steroid hormones may modulate the synthesis and, presumably, the release of a neurotransmitter, as well as the response of its target cells. The mechanisms by which circulating steroid hormones affect, simultaneously, various aspects of neuronal communication represents an extremely effective way of neuroendocrine integration. Therefore, the study of transynaptic regulatory actions of steroids at a basic level becomes useful for understanding the way these compounds work. The models presented here may provide the basis for such investigations.
Previous reports have indicated the effect of gonadal hormones on the rat SCG neurons, including changes in TH activity (Hamill et al., 1984; Hermindez et al., 1990), or the number of cells (Wright and Smolen, 1985), as well as modifications of NE turnover (Cardinali et al., 1975i Hern~ndez et al., 1990). The results presented here indicate that, in both, male and female rats, gonadal steroids may modulate TH activity in pineal sympathetic nerve terminals (Figs 4-6), and alter the response of pineal cells to NE (Figs 7-9). Moreover, in pineal

376 Alonso-Solfs et al.
cells from gonadectomized male and female rats, gonadal steroids may directly modulate cAMP accumulation after adrenoceptor activation. Our results from male rat pinealocytes, indicating an enhancement of NE-induced cAMP accumulation after treatment with T and E2 (Fig. 11), agree with early demonstrations of steroid uptake and metabolism by male rat pineals in organ cultures (Cardinali et aL, 1974). The blockage by Pg of the response to adrenergic stimulation induced by E2 in female rat pinealocytes (Fig. 10), also agrees with the effects of, in vivo, treatment with ovarian hormone blockers on pineal melatonin levels (Fig. 2). Since exposure of pineal cells to Pg produced the same effect when this hormone was coupled to BSA, it seems likely that these actions may be exerted at the level of the plasma membrane, as it has been previously reported in preparations of rat hypothalamic slices (Pettiti and Etgen, 1992). Whether these results are only the consequence of a direct action of steroids on the pinealocyte, or also indirectly mediated through other cell types to which pinealocytes respond, remains to be clarified. Nevertheless, since adrenergically- induced modulation of rat pineal androgen and estrogen metabolism has been shown (Cardinali et al., 1982; 1983), these findings may represent an example of a self-modulatory mechanism by which a hormone controls its own metabolism through reciprocal relationships with certain neurotransmitter systems (Slonso et aL, 1993). The simplicity of the rat pineal organ may constitute an extremely useful model for dissecting out the different components of such mechanisms.
Ovarian steroids control the synthesis (Rosie et al., 1990) and release (Drouva et al., 1983; Levine and Ramfrez, 1980; Sharkar and Fink, 1978) of LHRH from hypothalamic neurons. Since some reports suggested that LHRH neurons do not contain estrogen or progesterone receptors (Shivers et al., 1983; Fox et al., 1990), it was generally assumed that steroids effects must be mediated by interneurons. According to this view, ovarian steroids would modulate the synthesis and release of neurotransmitters from different interneurons which, in turn, could stimulate or inhibit LHRH output (Barraclough and Wise, 1982; Fink, 1994; Kalra and Kalra, 1983; Kordon et al., 1994). However, other reports indicated that gonadal steroids might also exert direct actions on GnRH neurons (Drouva et aL, 1983; Hoffman et al., 1990; Lee et al., 1990; Kim and Ramfrez, 1985). Using the GnRH producing neuronal cell line it has been recently shown that these cells possess specific binding sites for gonadal steroids, as well as the enzymatic machinery through which sex hormones regulate LHRH delivery (Poletti et aL, 1994). Moreover, by using RT-PCR it has been possible to demonstrate that GTa_7 cells express estrogen receptors in culture, and that they are functionally activated by exposure to E2 (Negro-Vilar et al., this issue). Our preliminary results confirm these findings, indicating that both E2 and Pg may exert direct actions on GT1_7 cells and modulate their response to NE stimulation (Fig. 12).
Among other receptors, GT~_v cells express the Bl-adrenoceptor, which is functionally coupled to adenyl cyclase (Findel et al., 1993; Martfnez de la Escalera et al., 1992b). Treatment of these cells with B~-agonists induces a rapid accumulation of cAMP, and stimulates the secretion of GnRH (Martinez de la Escalera et al. 1992b). Our results strongly suggest that E2 may exert a direct

Steroids and Neuroendocrine Transduction 377
inhibitory action on NE-induced cAMP accumulation, and that Pg, probably by acting at the level of the cell membrane, abolishes such effects in these cells. To our knowledge, this may constitutes the first indication of a direct effect of ovarian steroids on the response of GnRH producing cells to a neurotransmitter, and it agrees well with previous reports from in vivo experiments. Thus, Pg enhances the activation of LHRH neurons (Lee et al., 1990), and increases LHRH mRNA levels in the hypothalamus of ovariectomized E2-primed rats (Kim et al., 1994), an effect which is completely dependent on the integrity of hypothalamic NE transmission. On the other hand, E2 treatment of ovariec- tomized rats suppresses isoproterenol-induced stimulation of adenyl cyclase in membranes from medial hypothalamus-preoptic area, apparently by uncoupling the B-receptor from the guanine nucleotide-binding protein (Ungar et al., 1993). Finally, some studies on hypothalamic slices have found the opposite, namely that in preoptic area and hypothalamic slices from ovariectomized estradiol-primed rats, Pg or P-3-BSA suppressed NE-induced cAMP formation (Pettiti and Etgen, 1992). However, these later results do not contradict our findings; since brain slices are heterogeneous in nature and NE-induced changes in cAMP levels may represent the effects on different types of neurons.
(6)
( 2 )
\ \
• / t J ) l //,, / j / /
Fig. 13. Scheme summarizing potential sites for transynaptic steroid actions. Steroid hor- mones may affect neurotransmitter synthesis and/or release by actin on presynaptic neurons (1, 2). Steroids may also modulate postsynaptic responses by interacting on membrane (3) or intraceUular receptors (4). In addition, steroids may be metabolized by glial cells into other neuroactive steroid molecules (5). Finally, as a working hypothesis, steroid effects on postsynaptic neurons might induce the release of unknown mediators which, in turn, would potentially affect presynaptic events (6).

378 Alonso-Solfs et at.
In summary, we present here enough evidences in support of gonadal steroid regulation of neuronal function, both at pre- and postsynaptic levels. In the rat pineal gland, sex hormones paritally control melatonin production by affecting NE synthesis in sympathetic nerve terminals, and by modulating pinealocyte response to adrenergic stimulation. In GnRH secreting cells, gonadalhormones control noradrenergic responses and cAMP formation. These examples constitute appropriate paradigms of transynaptic steroid regulation (Fig. 13), as well as excellent models for the study of complexity of steroid hormone actions on the brain.
ACKNOWLEDGMENTS
This work was supported by DGICYT PM92-0160 and GAC 93/002 to R. A., University of La Laguna 227-068/93 to P.A., and GAC 92/069 to I.L.-C. The authors would like to thank Dr. J. E. Sfinchez-Criado and Dr. F. L6pez for helpful ideas and assistance with hormone blockers experiments and GTI_7 cells culture, respectively.
REFERENCES
Abreu, P., Santana, C., Hcm~ndez, G., Calzadilla, C. H., and Alonso, R. (1987). Day-night rhythm of rat pineal tyrosine hydroxylass activity as determined by high-performance liquid chromatog- raphy with amperometric detection. J. Neurochem. 48:665-668.
Alonso, R., Abreu, P., and Fajardo, N. (1993). Steroid influences on pineal melatonin production. In Yu, H.-S., and Reiter, R. J. (eds.), Melatonin. Biosynthesis, Physiological Effects, and Clinical Applications, CRC Press, Boca Raton, FL, pp. 73-105.
Arnold, A. P., and Breediove, S. M. (1985). Organizational and activational effects of sex steroids on brain and behavior: A reanalysis. Hormones Behav. 19:.469-498.
Barraclough, C. A., and Wise, P. M. (1982). The role of catecholamines in the regulation of pituitary luteinizing hormone and follicle-stimulating hormone secretion. Endocrine Rev. 3:91-119.
Beato, M. (1989). Gene regulation by steroid hormones. Cell 56-.335-344. Blum, M., McEwen, B. S., and Roberts, J. L. (1987). Transcriptional analysis of tyrosine hydroxylase
gene expression in the tuberoinfundibular dopaminergic neurons of the rat arcuate nucleus after estrogen treatment. J. Biol. Chem. 262".817-821.
Brownstein, M., and Axelrod, J. (1974). Pineal gland: 24 hour rhythm in norepinephrine turnover. Science 148:163-165.
Brzezinski, A., Lynch, H. J., Seibel, M. M., Deng, M. H., and Nader, T. M. (19.88). The circadian rhythm of plasma melatonin during the normal menstrual cycle and in amen9rrheic women. J. Clin. Endocrinol. Metab. 66".891-895.
Buda, M., and Klein, D. C. (1978). Suspension culture of pinealocytes: Regulation of N- acetyltransferase activity: Endocrinology 103:1483-1493.
Cardinali, D. P. (1979). Nuclear receptor estrogen complex in the pineal gland. Modulation by sympathetic nerves, Neuroendocrinology 24:333-346.
Cardinali, D. P., Nagle, C. A', and Rosner, J. M. (1974). Metabolic fate of androgens in the pineal organ: Uptake, binding to cytoplasmic proteins and conversion of testosterone into 50-reduced metabolites. Endocrinology 95-179-187.
Cardinali, D. P., Vacas, M. I., and Gejman, P. V. (1981). The sympathetic superior cervical ganglia as peripheral neuroendocrine centers. J. Neural Trans. 52:1-21.
Cardinali, D. P., Ritta, M. N., and Gejman, P. V. (1982). Norepinephrine stimulates testosterone

Steroids and Neuroendocrine Transduction 379
aromatization and inhibits 50 reduction via 6-adrenoceptors in rat pineal gland. Mol. Cell. Endocrinol. 28:199-209.
Cardinali, D., Vacas, M. I., Ritta, M. N., and Gejman, P. V. (1983). Neurotransmitter-controlled steroid hormone receptors in the central nervous system. Neurochem. Int. 5:185-192.
Cardinali, D. P., Vacas, M. I , and Rosenstein, R. E. (1987). Muttifactorial control of pineal melatonin synthesis: An analysis through binding sites. In Reiter, R. J., and Fraschini, F. (Eds.), Advances in Pineal Research, Vol. 2, John Libbey, London, pp. 51-59.
Cesnjaj, M., Krsmanovic, L. Z., Catt, K. J., and Stojilkovic, S. S.(1993). Autocrine induction of c-fos expression in GT1 neuronal cells by gonadotropin-releasing hormone. Endocrinology 133:3042- 3045.
Chan, Y. S., Cheung, Y. M., and Pang, S. F. (1991). Rhythmic release pattern of pineal melatonin in rodents. Neuroendocrinology 53 (Suppl. 1):69-67.
Chomcynski, P,, and Sacchi, N. (1986). Single-step method of RNA isolation by acid guanidium thiocyanate-phenolchloroform extraction. Anal, Biochem. 162:156-159.
Dluzen, D. E., and Ramirez, V. D. (1989). Progesterone effects upon dopamine release from the corpus striatum in female rats. I. Evidence for interneuronal control. Brain Res. 476:332-337.
Drouva, S. V., Laplante, E., and Kordon, C. (1983). Effects of ovarian steroids on in vitro release of LHRH from mediobasal hypothalamus. Neuroendocrinology 37:336-341.
Etgen, A. M., Ungar, S., and Pettiti, N. (1992). Estradiol and progesterone modulation of norepinephrine neurotransmission: Implications for the regulation of female reproductive behavior. J. Neuroendocrinot. 4.255-271.
Evans, R. (1988). The steroid and thyroid hormone receptor superfamily. Science 240:.889-895. Findell, P. R., Wong, K. H., Jackman, J. K., and Daniels, D. V. (1993). Bl-Adrenergic and dopamine
(D1)-receptors coupled to adenylyl cyclase activation in GT1 gonadotropin-releasing hormone meuroscretory cells. Endocrinology 132:682-688.
Fink, G. (1988). The G. W. Harris Lecture. Steroid control of brain and pituitary function. Q. J. Exp. Physiol, 73.257-293.
Fink, G. (1994). Molecular principles from neuroendocrine models: Steroid control of central neurotransmission. In Bloom, F. (Ed.), Neuroscience: From the Molecular to the Cognitive, Progress in Brain Research, Vol 110, Elsevier, Amsterdam, pp. 139-147.
Fink, G., Rosei, R., and Thomson, E. (1991). Steroid actions on hypothalamic neurons with special reference to estrogen control of luteinizing hormone-releasing biosynthesis and release. In Fuxe, K., and Agnati, L. F. (Eds.), Volume Transmission in the Brain: Novel Mechanisms for Neural Transmission, Raven Press, New York, pp. 195-211.
Fox, S. R., Harlan, R. E., Shivers, B. D., and Pfaff, D. W. (1990). Chemical characterization of neuroendocrine targets for progesterone in the female rat brain and pituitary. Neuroendo- crinology 51:276-283.
Gaytan, F., Bellido, C., Aceitero, J., Aguilar, E., and S~tnchez-Criado, J. E. (1990). Leyding cell involvement in the paractine regulation of mast cells in the testicular interstititum of the rat. Biol. Reprod. 43"665-671.
Grima, B., Lamoroux, A., Blanot, F., Faucon Biguet, N., and Mallet, J. (1985). Proc. Natl. Acad. Sci. USA 82:617-621.
Hadcock, J. R., and Malbon, C. C. (1991). Regulation of receptor expression by agonists: Transcriptional and post-transcriptional controls. TINS 14:242-247.
Hamill, R. W., Earley, C. J., and Guernsey, L. A. (1984). Hormonal regulation of adult sympathetic neurons: The effects of castration on tyrosine hydroxylase activity. Brain Res. 299:.331-337.
Harper, M. J. K., and Walpole, A. L. (1967). A new derivative of triphenylethylene: Effects on implantation and mode of action in rats..I. Repro& Fertil. 13.'101-119.
Harrelson, A., and McEwen, B. S. (1987). Gonadal steroid modulation of neurotransmitter-stimulated cAMP accumulation in the hippocamous of the rat. Brain Res. 404:89-94.
Hern~dez, G., Abreu, P., Alonso, R., Santa, C., Moujir, F., and Calzadilla, C. H. (1990a). Castration reduces the nocturnal rise of pineal melatonin levels in the male rat by impairing its noradrenergic input. J. Neuroendocrinol. 2:777-782.
Hern~ndez, G., Abreu, P., Alonso, R., and Calzadilla, C. H. (1990b). Determination of pineal melatonin by high-performance liquid chromatography with electrochemical detection: Applica- tion for rhythm studies and tissue explants. J. Pineal Res. 8:11.-19.
Hern~dez, G., Bello, A. R., Gonz~lez-Coviella, I. L., Abreu, P., Fajardo, N., Reiter, R. J., Hermindez, A., and Alonso, R. (1994). Tyrosine hydroxylase activity in peripherally denervated rat pineal gland. Neurosci. Len. 17"7:131-134.
Ho, A. K., and Klein, D. C. (1987). Phosphatidylinositol phosphodiesterase (phospholipase C) activity in the pineal gland: characterization and photoneural regulation. J. Neurochem. 48:1933-1038.

380 Alonso-Solis et al.
Hoffman, G. E., Lee, W.-S., Attardi, B., Yann, V., and Fitzsimons, M. D. (1990). Luteinizing hormone-releasing hormone neurons express c-los antigen after steroid activation. Endocrinology 126:1736-1741.
Hruska, R, E. (1986). Elevation of striatal dopamine receptors by estrogen: dose and time studies. J. Neurochera. 47:1908-1915.
Joell, M., and De Kloet, R. (1991). Control of neuronal excitability by corticosteroid hormones. TINS 15:25-30.
Jones, K. J., and Pfaff, D. W. (1991). Emerging tenets in the mechanism of gonadal steroid action on hypothalamic neurons. In Motta, M. (ed.), Brain Endocrinology, Raven Press, New York, pp. 153-175.
Kim, K., and Ramirez, V. D. (1985). In vitro luteinizing hormone releasing hormone release from superfused rat hypothalami: Site of action of progesterone and effect of estrogen priming. Endocrinology 116:252-258.
Kim, K., Lee, B. J., Cho, B. N., Kang, S. S., Choi, W. S., Park, S. D., Lee, C. D., Cho, W. K., and Wuttke, W. (1994). Blockade of noradrenergic neurotransmission with diethyldithiocarbamic acid decreases the mRNA levels of gonadotropin-releasing hormone in the hypothalamus of ovariectomized, steroid-treated prepubertal rats. Neuroendocrinology 59:539-544.
Klein, D. C., Sugden, D., and Weller, J. L. (1983). Postsynaptic ct-adrenergic receptor potentiates 8-adrenergic stimulation of pineal serotonin N-acetyltransferase, Proc. Natl. Acad. Sci. USA 80:.599-603.
Kordon, C., Drouva, S. V., Martfnez de la Rscalera, G., and Wiener, R. (1994). Role of classic and peptide neuromediators in the neuroendocrine regulation of Iuteinizing hormone and prolactin. In Knobil, E., Neill, E., et al. (eds.), The Physiology of Reproduction, Raven Press, New York, Vol. I, pp. 1621-1681.
Krsmanovic, L. Z., Stojilkovic, S. S., Merelli, F., Dufour, S. M., Virmani, M. A., and Catt, K. J. (1992). Calcium signaling and episodic secretion of gonadotropin-releasing hormone in hypothal- amic neurons. Proc. Natl. Acad. Sci. USA 89:8462-8466.
Kukstas, L. A., Domec, C., Bascles, L , Bonnet, J., Verrier, D., Israel, J.-M., and Vincent, j.-D. (1991). Different expression of the two dopaminergic D2 receptors, D2415 and D2444, in two types of lactotroph, each characterized by their response to dopamine, and modification of expression by sex steroids. Endocrinology 129:1101-1102.
Laemmli, U. K. (1970). Cleavage of structural proteins during assembly of the head of bacteriophate T 4. Nature 227:680-685.
Lee, W.-S., Smith, S., and Hoffman, G. E. (1990). Progesterone enhances the surge of luteinizing hormone by increasing the activation of luteinizing hormone-releasing hormone neurons. Endocrinology 127:2604-2606.
Levine, J. E., and Ramirez, V. D. (1980). In vivo release of luteinizing hormone-releasing hormone estimated with push-pull cannulae from the mediobasal hypothalami of ovariectomized, steroid primed rats. Endocrinology 107:1782-1790.
Maus, M., Bertrand, P., Drouva, S., Rasolonjanahary, Kordon, C., Glowinski, J., Premont, J., and Enjalbert, A. (1989). Differential modulation of D1 and D2 dopamine-sensitive adenylate cyclases by 17g-estradiol in cultured striatal neurons and anterior pituitary cells. J. Neurochem. 52:410-418.
Martfnez de la Escalera, G., Choi, A. L. H., and Weiner, R. I. (1992a). Generation and synchronization of gonadotropin-releasing hormone (GnRH) pulses: Intrinsic properties of the GTI-1 GnRH neuronal cell line. Proc. Natl. Acad. Sci. USA 89"1852-1855.
Martfnez de ia Escalera, G., Gallo, F., Choi, A. L. H., and Weiner, R. I. (1992b), Betal-adrenergic regulation of the GT1 GnRH neuronal cell lines: Stimulation of GnRH release via receptors positively coupled to adenylate cyclase. Endocrinology 131:1397-1402.
Martfnez de la Escalera, G., Gallo, F., Choi, A. L. H., and Weiner, R. I. (1992c). Dopaminergic regulation of the GT 3 gonadotropin-releasing hormone (GnRH) neuronal cell lines: stimulation of GnRH release via Dl-receptors positively coupled to adenylate cyclase. Endocrinology 131:2965-2971.
Martfnez de la Escalera, G., Choi, A. L. H., and Weiner, R. I. (1994). Biphastic regulation of GnRH secretion in GT1 cell lines. Neuroendocrinotogy $9:.420-425.
McEwen, B. S. (t991). Non-genomic and genomic effects of steroids on neural activity. TIPS 12:141-147.
McEwan, B. S., and Woolley, C. S. (1994). Estradoil and progesterone regulate neuronal structure and synaptic connectivity in adult as well as developing brain. Exp. Geront. 29:.431-436.
McEwen, B. S., Jones, R., and Pfaff, D. (1987). Hormonal control of sexual behaviour in the female rat: Molecular, cellular and neurochemical studies. Biol. Reprod. 36:37-45.

Steroids and Neuroendocrine Transduction 381
Mellon, P. L., Wind]e, J. J., Goldsmith, P. C., Padula, Roberts, J, L., and Weiner, R. I. (1990). Immortalization of hypothalamic GnRH neurons by genetically targeted tumorogenesis. Neuron 5:1-10.
Moujir, F., Bord6n, R., Santana, C., Hern~ndez, G,~ Abreu, P., and Alonso, R. (1990). Ovarian steroids block the isoproterenol-induced elevation of pineal melatonin production in female rats. Neurosci. Lett. 119.'12-14.
Moyer, M. L., Borror, K. C., Bona, B. J., De Franco, D. B., and Nordeen, S. K. (1993). Modulation of cell signaling pathways can enhance or impair glucoconicoid.induced gene expression without altering the state of receptor phosphorylation. Z Biol. Chem. 268:22,933-22,940.
Nagle, C. A., Cardinali, D. P., and Rosner, J. M. (1974). Effects of castration and testosterone administration on pineal and retinal hydroxyindot-O-methyltransferase of male rats. Neuroendocrinology 14:14-23.
Ozaki, Y., Wurtman, R. J., Alonso, R., and Lynch, H. J. (1978). Melatonin secretion decreases during the proestrous stage of the rat estrous cycle. Proc. Natl. Acad. Sci. USA 75:531-534.
Pasqualini, C., Leviel, V., Guiben, B., Faucon-Biguet, N., and Kerdelhu6, B. (1991). Inhibitory actions of acute estradiol treatment on the activity and quantity of tyrosine hydroxylase in the median eminence of ovariectomized rats. J. Neuroendocrinol. 3:.575-580.
Pasqualini, C., Guiben, B., and Leviel, V. (1993). Short-term inhibitory effect of estradiol on tyrosine hydroxylase activity in tuberoinfundibular dopaminergic neurons in vitro. J. Neurochem. 60:.1707-1713.
Pettiti, N., and Etgen, A. M. (1990). Alpha~-adrenoceptor augmentation of B-stimulated cAMP formation is enhanced by estrogen and reduced by progesterone in rat hypothalamic slices. Z Neurosci. 10:.2842-2849.
Pettiti, N., and Etgen, A. M. (1992). Progesterone promotes rapid desentization of alpha~-adrenergic receptor augmentation of cAMP formation in rat hypothalamic slices. Neuroendocrinology 55:1-8.
Pfaff, D. W. (1989). Patterns of steroid hormone effects on electrical and molecular events in hypothalamic neurons. Mol. NeurobioL 3:.135-154.
Poletti, A., Melcangi, R. C., Negri-Cesi, P., Maggi, R., and Martini, L. (1994). Steroid binding and metabolism in the luteinizing hormone-releasing hormone-producing neuronal cell line GTI-1. Endocrinology 135:2623-2628.
Reiter, R. J. (1991). Pineal gland. Interface between the photoperiodic environment and the endocrine system. TEM 2:13-29.
Rosie, R., Thomson, E., and Fink, G. (1990). Estrogen postiive feedback stimulates the synthesis of LHRH mRNA in neurons of the rostal diencepbalon. 3". Endocrinol. 124:285-289.
Schumacher, M. (1990). Rapid membrane effects of steroid hormones: An emerging concept in neuroendocrinology. TINS 13:359-361.
Schumacher, M., Coirini, H., Frankfurt, M., and McEwen, B. S. (1989). Localized actions of progesterone in hypothalamus involve oxytocin. Pro¢. Natl. Acad. Sci. USA 86:6798-6801.
Schumacher, M., Coirini, H., Pfaff, D. W., and McEwen, B. S. (1990). Behavioral effects of progesterone associated with rapid modulation of oxytocin receptors. Science 250:.691-694.
Sharkar, D. K., and Fink, G. (1979). Effects of gonadal steroids on output of luteinizing hormone releasing factor into pituitary stalk blood in the female rat. J. Endocrinol. 80:.303-313.
Shiotani, Y., Shiosaka, S., Emson, P. C., Hilyard, C. J., Girgis, S., and Maclntyre, I. (1986). Distribution of substance P (SP)-, calcitonin gene-related peptide (CGRP)-, vasoaetive intestinal polypeptide (VIP)-, and neuropeptide NPY)-containing nerve fibers in the pineal gland of gerbils. Neurosci. Len. 70:.187-192.
Shivers, B. D., Harlan, R. E., Morrel, J. E., and Pfaff, D. W, (1983). Absence of oestradiol concentration in cell nuclei of LHRH-immunoreactive neurons. Nature 304"345-347.
Spergel, D. J., Krsmanovic, L. Z., Stojilkobic, S. S., and Catt, K. J. (1994). Glutamate modulates [Ca2÷]i gonadotropin-releasing hormone secretion in immortalized hypothalamic GT1-7 neurons. N euroendocrinology 59:.309-317.
Stehle, J. H., Foulkes, N., Molina, C., Simonneaux, U., Pevet, P., and Sassone-Corsi, P. (1993). Adrenergic signals direct rhythmic expression of transcriptional repressor CREM in the pineal gland. Nature 365:314-320.
Sugden, D. (1989). Melatonin biosynthesis in the mammalian pineal gland. Experientia 45:922-932. Sugden, D., Vanecek, J., Klein, D. C., Thomas, T. P., and Anderson, W. B. (1985). Activation of
protein kinase C potentiates isoprenaline-induced cyclic AMP accumulation in rat pinealocytes. Nature 314:359-361.
Sugden, A. L., Sugden, D., and Klein, D. C. (1987). arAdrenoceptor activation elevates cytosolic calcium in rat pinealocytes by increasing net influx. Z Biol. Chem. 262:741-745.

382 Alonso-Solfs et al.
Sutherland, R., Mester, J., and Baulieu, E. E. (1977). Tamoxifen as a "pure" anti-oestrogen in chick oviduct, Nature 267:434-435.
Ungar, S., Makman, M. H., Morris, S. A., and Etgen, A. M. (1993). Estrogen uncouples 8-adrenergic receptor from the stimulatory guanine nucletodie-binding protein in female rat hypothalamus. Endocrinology 133:2818-2826.
Vacas, M. I., and Cardinali, D. P. (1979). Effect of castration and reproductive hormones on pineal serotonin metabolism in rats. Neuroendocrinology 28:187-195.
Vacas, M. I., Lowenstein, P. R., and Cardinali, D. P. (1979). Characterization of a cytosol progesterone receptor in bovine pineal gland. Neuroendocrinology 24:84-89.
Vanecek, J., Kosak, E., and Vorlicek, J. (1990). Daily changes in melatonin binding sites and the effect of castration. Mol, Cell. Endocrinol. 73:165-170.
Weiner, R. I., Wetsel, W., Goldsmith, P., Martfnez de la Escalera, G., Windle, J., Padula, C., Choi, A., Megro-Vilar, A., and Mellon, P. (1992). Gonadotropin-releasing hormone cell lines. In Ganong, W. F., and Martini, L (eds.), Frontiers in Neuroendocrinology, Vol. 13, Raven Press, New York, pp. 95-119.
Weiss, B., and Crayton, J. (1970). Gonadal hormones as regulators of rats pineal adenylate cyclase activity. Endocrinology 87:527-533.
Wehling, M. (1994). Nongenomic actions of steroid hormones. TEM 5:347-353. Wright, L. L., and Smolen, A. J. (1985). Effects of neonatal castration or treatment with
dihydrotestosterone on number of neurons in the rat superior cervical sympathetic ganglion. Deo. Brain Res. 20:.314-316.
Zisapel, N., and Anis, Y. (1988). Impact of circulating testosterone on iodomelatonin binding sites in the male rat brain. Mol. Cell. Endocrinol. 60:119-126.
Zisapel, N., Shaharabani, M., and Laudon, M. (1987). Regulation of melatonin's activity in the female rat brain by estradiol: Effects on neurotransmitter release and on iodomelatonin binding site. Neuroendocrinology 46:207-216.
Copyright © 2022 FDOKUMEN




















