
Increased myocardial collagen and ventricular diastolic dysfunction in relaxin deficient mice: a...
10
Cardiovascular Research 57 (2003) 395–404 www.elsevier.com / locate / cardiores Increased myocardial collagen and ventricular diastolic dysfunction in relaxin deficient mice: a gender-specific phenotype a, b a b ,1 b * Xiao-Jun Du , Chrishan S. Samuel , Xiao-Ming Gao , Ling Zhao , Laura J. Parry , b Geoffrey W. Tregear a Experimental Cardiology Laboratory, Baker Medical Research Institute, PO Box 6492, St Kilda Road Central, Melbourne, Vic. 8008, Australia b Howard Florey Institute of Experimental Physiology and Medicine, University of Melbourne, Melbourne, Vic. 3010, Australia Received 19 March 2002; accepted 10 September 2002 Abstract Objective: To investigate cardiac phenotypes in mice deficient in the peptide hormone relaxin by gene targeting. Methods: 2 / 2 Echocardiography and cardiac catheterization were performed on male and female relaxin deficient (Rlx ) mice as well as 1 / 2 1 / 1 heterozygous (Rlx ) and wildtype (Rlx ) littermates aged between 8 and 24 months. Collagen expression and content in the heart were analysed by real-time PCR, hydroxyproline assay and histology. Results: Heart rate, blood pressures, left ventricular (LV) dimensions, fractional shortening and maximal and minimal dP/dt did not differ significantly between the three genotypes of either 2 / 2 gender at any age. However, 8–10-month-old Rlx males exhibited a greater transmitral flow velocity (A-wave) at the late LV diastolic 2 / 2 phase. Male Rlx mice aged between 12 and 24 months had significantly higher LV end-diastolic pressures, a 30% increase in atrial weight and 10–30% increases in lung and liver weights. Male mice also showed an age-dependent increase ( P,0.01) in LV collagen 2 / 2 content that was more pronounced in Rlx than control littermates ( P,0.01). Procollagen type-1 expression was also significantly 2 / 2 1 / 2 1 / 1 higher in the LV of Rlx males compared with either Rlx or Rlx males at 6, 9 and 12 months of age. Age-matched female 2 / 2 2 / 2 2 / 2 Rlx mice did not display any of these cardiac phenotypes seen in Rlx males. Conclusions: Male Rlx mice had impeded LV diastolic filling and increased atrial weights, most likely due to an increase in ventricular collagen content and chamber stiffness. These 2 / 2 2 / 2 phenotypes in the Rlx males were not observed in Rlx females, indicating the importance of other gender-related factors in cardiovascular function. 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved. Keywords: Fibrosis; Gender; Gene expression; Hormones; Ventricular function 1. Introduction and uterine cervix to facilitate birth [1–3]. Relaxin also stimulates growth and differentiation of the mammary Relaxin is a peptide hormone structurally similar to gland and nipple, both important in lactating females [1]. insulin and insulin-like growth factors, with two chains These effects of relaxin are achieved through its inhibition linked by disulfide bridges [1]. Relaxin concentrations are of collagen synthesis and activation of matrix metallop- highest in the ovary, placenta or serum of females during roteinases (MMPs) that degrade collagen [4,5]. The mouse pregnancy [1]. The main functions of relaxin are associated has two relaxin genes, M1 and M3, equivalent to the with female reproductive physiology and in particular human relaxin genes H2 and H3, respectively [6,7]. Late- 2 / 2 connective tissue remodeling in the interpubic symphysis pregnant mice rendered deficient in M1 relaxin (Rlx ) by gene targeting appear to deliver normally, although the pups are unable to suckle and do not survive [8]. This is *Corresponding author. Tel.: 161-3-8530-1294; fax: 161-3-8530- due to insufficient development of the mammary gland and 1100. nipple caused by an increased collagen content in these E-mail address: [email protected] (X.-J. Du). 1 Present address: Laboratory of Genetics and Physiology, NIDDK, NIH, Bethesda, MD 20892, USA. Time for primary review 29 days. 0008-6363 / 03 / $ – see front matter 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved. PII: S0008-6363(02)00663-6 at Beijing Normal University on May 29, 2013 http://cardiovascres.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Increased myocardial collagen and ventricular diastolic dysfunction in relaxin deficient mice: a...

Cardiovascular Research 57 (2003) 395–404www.elsevier.com/ locate/cardiores
I ncreased myocardial collagen and ventricular diastolic dysfunction inrelaxin deficient mice: a gender-specific phenotype
a , b a b ,1 b*Xiao-Jun Du , Chrishan S. Samuel , Xiao-Ming Gao , Ling Zhao , Laura J. Parry ,bGeoffrey W. Tregear
aExperimental Cardiology Laboratory, Baker Medical Research Institute, PO Box 6492, St Kilda Road Central, Melbourne, Vic. 8008, AustraliabHoward Florey Institute of Experimental Physiology and Medicine, University of Melbourne, Melbourne, Vic. 3010, Australia
Received 19 March 2002; accepted 10 September 2002
Abstract
Objective: To investigate cardiac phenotypes in mice deficient in the peptide hormone relaxin by gene targeting.Methods:2 /2Echocardiography and cardiac catheterization were performed on male and female relaxin deficient (Rlx ) mice as well as
1 /2 1 /1heterozygous (Rlx ) and wildtype (Rlx ) littermates aged between 8 and 24 months. Collagen expression and content in the heartwere analysed by real-time PCR, hydroxyproline assay and histology.Results: Heart rate, blood pressures, left ventricular (LV)dimensions, fractional shortening and maximal and minimal dP/dt did not differ significantly between the three genotypes of either
2 /2gender at any age. However, 8–10-month-old Rlx males exhibited a greater transmitral flow velocity (A-wave) at the late LV diastolic2 /2phase. Male Rlx mice aged between 12 and 24 months had significantly higher LV end-diastolic pressures, a 30% increase in atrial
weight and 10–30% increases in lung and liver weights. Male mice also showed an age-dependent increase (P,0.01) in LV collagen2 /2content that was more pronounced in Rlx than control littermates (P,0.01). Procollagen type-1 expression was also significantly
2 /2 1 /2 1 /1higher in the LV of Rlx males compared with either Rlx or Rlx males at 6, 9 and 12 months of age. Age-matched female2 /2 2 /2 2 /2Rlx mice did not display any of these cardiac phenotypes seen in Rlx males.Conclusions: Male Rlx mice had impeded LV
diastolic filling and increased atrial weights, most likely due to an increase in ventricular collagen content and chamber stiffness. These2 /2 2 /2phenotypes in the Rlx males were not observed in Rlx females, indicating the importance of other gender-related factors in
cardiovascular function. 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.
Keywords: Fibrosis; Gender; Gene expression; Hormones; Ventricular function
1 . Introduction and uterine cervix to facilitate birth [1–3]. Relaxin alsostimulates growth and differentiation of the mammary
Relaxin is a peptide hormone structurally similar to gland and nipple, both important in lactating females [1].insulin and insulin-like growth factors, with two chains These effects of relaxin are achieved through its inhibitionlinked by disulfide bridges [1]. Relaxin concentrations are of collagen synthesis and activation of matrix metallop-highest in the ovary, placenta or serum of females during roteinases (MMPs) that degrade collagen [4,5]. The mousepregnancy [1]. The main functions of relaxin are associated has two relaxin genes, M1 and M3, equivalent to thewith female reproductive physiology and in particular human relaxin genes H2 and H3, respectively [6,7]. Late-
2 /2connective tissue remodeling in the interpubic symphysis pregnant mice rendered deficient in M1 relaxin (Rlx )by gene targeting appear to deliver normally, although thepups are unable to suckle and do not survive [8]. This is
*Corresponding author. Tel.:161-3-8530-1294; fax:161-3-8530- due to insufficient development of the mammary gland and1100.
nipple caused by an increased collagen content in theseE-mail address: [email protected](X.-J. Du).1Present address: Laboratory of Genetics and Physiology, NIDDK,
NIH, Bethesda, MD 20892, USA. Time for primary review 29 days.
0008-6363/03/$ – see front matter 2003 European Society of Cardiology. Published by Elsevier Science B.V. All rights reserved.PI I : S0008-6363( 02 )00663-6
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from

396 X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404
2 /2tissues [8,9]. Male Rlx mice also show phenotypes of Sonos 5500 ultrasound machine with a 15-MHz linear-higher collagen content in the prostate gland, poor tubular array transducer (Hewlett-Packard), as described previous-development and a reduction in prostate growth [10]. ly [20]. The first experiment was conducted on a total of
1 /1 1 /2 2 /2In recent years, evidence has accumulated to show that 60 male and female Rlx , Rlx , Rlx mice agedrelaxin acts on the cardiovascular system in both females between 12 and 24 months. Mice were anesthetizedand males. High affinity relaxin binding sites are localized (mixture of ketamine–xylazine–atropine at 60–12–0.6to the atria of male and female rats [11,12] and studies in mg/kg, respectively) and 2-dimensional guided M-modevitro and in vivo have demonstrated positive chronotropic traces crossing the LV were acquired at a sweep speed ofand probably inotropic responses to exogenous relaxin in 100 mm/s. The following parameters were measured,both genders [13–15]. Relaxin stimulates atrial natriuretic using the leading-edge technique: LV internal dimensionspeptide secretion from isolated perfused rat hearts [16] and of diastole and systole (LVIDd, LVIDs), LV externalalso causes a dose-dependent increase in coronary blood dimension of diastole (LVEDd) and wall thickness atflow via a nitric oxide-mediated mechanism [17]. Cultured diastole and systole. Measurements were taken, in blindatrial cardiocyctes derived from adult male rats produce fashion, from two cardiac cycles and the data averaged.and secrete immunoreactive relaxin [18], and a recent Fractional shortening (FS%) was calculated as (LVIDd2
study has demonstrated relaxin mRNA in the human heart LVIDs) /LVIDd. Wall thickening index (WTI) was the net[19]. Data also show that chronic heart failure patients increase in LV wall thickness in systole versus diastole. LV
3 3have increased myocardial relaxin gene expression and mass was calculated as (LVEDd –LVIDd )31.05.elevated plasma relaxin concentrations [19], suggesting A second echocardiographic experiment used 27 male
1 /1 1 /2 2 /2that cardiac relaxin may regulate heart function by an Rlx , Rlx and Rlx mice aged at 9 monthsautocrine/paracrine mechanism. (between 8 and 10 months). After completion of M-mode
To investigate the physiological significance of endog- image recording, the ventricular diastolic function wasenous relaxin in the heart, we determined cardiac function analyzed by long-axis 2-D image guided color Doppler
2 /2in both male and female Rlx mice. The ventricular recordings of blood flow velocities across the mitral valvesmyocardium was examined for chemical and morphologi- from the apical position while a standard lead-II ECG wascal abnormalities, with emphasis on interstitial collagen continuously recorded. Peak flow velocities, isovolumic-content and myocyte hypertrophy. To address the age- relaxation time (IVRT) and E-wave deceleration timedependent development of cardiac phenotypes in these (DT) were measured digitally in standard fashion [21,22].mice, animals were studied between 8 and 24 months of All measures were taken from seven to ten cardiac cyclesage for male mice and 12 and 24 months of age for female and averaged.mice. The results from this study point to a cardiacphenotype in male mice deficient in relaxin. Specifically, 2 .3. Cardiac catheterizationleft ventricular diastolic filling is impeded which is verylikely related to an increase in collagen content in the left Arterial blood pressures and LV pressures were mea-ventricle (LV). sured in groups of seven to 12 male and female mice, as
described previously [23]. Mice were anesthetized (pen-tobarbitone 80 mg/kg and atropine 0.6 mg/kg) and a 1.4 F
2 . Methods catheter (Millar Instruments, Houston) was inserted intothe LV via the right carotid artery. Aortic blood pressures,
2 .1. Animals heart rate, LV pressures and the maximal rate of increase ordecay of LV pressures, dP/dt or dP/dt , were ac-max min
All animal experiments were approved by the Baker quired digitally. After recording baseline parameters, aInstitute and Howard Florey Institute Animal Experimental b-adrenergic agonist isoproterenol was infused intraven-
2 /2Ethics Committees. The relaxin gene knockout (Rlx ) ously as a bolus at increasing doses of 50–2000 pg perstrain was generated at the Howard Florey Institute by mouse and functional responses were monitored.targeted deletion of the relaxin gene as described previous-
1 /2ly [8]. Heterozygous (Rlx ) mice were mated to obtain 2 .4. Morphological analysis1 /1 2 /2 1 /2wildtype (Rlx ), Rlx and Rlx littermates. All
mice used in these studies had the same mixed background Mice were killed at the end of the experiments by(129SV/C57Blk6J) and were aged between 9 and 24 pentobarbitone overdose. The hearts were isolated and themonths for males and 12 and 24 months for females. Tail LV, right ventricle and atria were separated and weighed.biopsies and PCR analysis of genomic DNA [8] confirmed The lungs, liver and kidney were also weighed and tibiathe genotype of each animal at weaning. length was measured. Ventricles were either frozen for
biochemical assay or fixed in 10% formalin in PBS.2 .2. Echocardiography Tissues were embedded in paraffin and transverse sections
(5 mm) were cut and stained with hematoxylin and eosin orTransthoracic echocardiography was performed using a 0.1% picrosirius red to stain collagen. Images of LV
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from

X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404 397
sections were gathered with a CCD video camera (Op- was demonstrated in parallel RT-PCR reactions that as-timas, BioScan, Edmonds, WA, USA), digitized and quan- sessed expression of glyceraldehyde phosphate dehydro-tified using Optimas 6.5 program. Interstitial collagen genase (GAPDH).content in the LV was determined following the methoddescribed previously [23]. The LV sections were sampled 2 .6. Statisticsin a systematic fashion and 15–20 fields in each LV wereanalyzed. The picrosirius red stained area was calculated as Results are expressed as mean6S.E.M. Between-groupa percentage of the total area within a field and the comparisons were made using one- or two-way analysis ofaveraged percentage was used. One section at the LVvariance (ANOVA) and unpaired Student’st-test as post-equator was used for measuring myocyte transverse cross-hoc analysis. Data from the real time PCR did not showsectional area. The average of 60–70 cells from randomly homogeneity of variance and were log transformed beforeselected fields was used. analysis by one-way ANOVA. Linear correlation was
performed for echo-derived LV mass and actual weights.2 .5. Collagen expression P,0.05 was considered statistically significant.
Total collagen content and concentrations in the LVmyocardium were determined by hydroxyproline assay as
3 . Resultsdescribed previously [24]. Male mice (n544) aged be-tween 9 and 24 months and female mice aged between 12
3 .1. Echocardiographic changesand 24 months (n518) were studied. The results wereexpressed asmg hydroxyproline/mg dry weight. In a
There were no gross abnormalities in the cardiovascularseparate experiment, heart tissues were collected from 2 /21 /1 2 /2 system and no mortality in male or female Rlx mice of12-month-old male Rlx and Rlx mice. The atria
any age. Using echocardiography and catheterization tech-were separated from the ventricles and all tissues frozen inniques, we found no significant difference in heart rate,liquid nitrogen. Total RNA was extracted using RNA WizdP/dt , dP/dt , LV dimensions, wall thickness, FS ormax min(Ambion; Geneworks, Adelaide, South Australia). RealWTI between the three genotypes of males at 8–24 monthstime PCR was used to quantify gene expression ofof age or females at 12–24 months of age (Table 1).procollagen type 1 (COL-1) and proMMPs (proMMP-2,
2 /2Aortic blood pressures and LV pressures in the RlxproMMP-9 and proMMP-13). First strand cDNA synthesis1 /2 1 /1mice were also similar to those of the Rlx and Rlxused 1 or 1.5mg total RNA and 1.25 U/ml MultiScribe
mice. LV end-diastolic pressure (LVEDP) was unchangedreverse transcriptase (Applied Biosystems, Foster City,2 /2in male Rlx mice at 8–10 months of age but elevatedCA) in a 30-ml reaction as described previously [25]. A
at 12–24 months of age (P,0.05, Table 1). Although theresecond reverse transcription reaction used 30 ng RNA andwas a slight trend for an increase in LVEDP in the femalea series of ventricle RNA dilutions (100–0.001 ng) for the
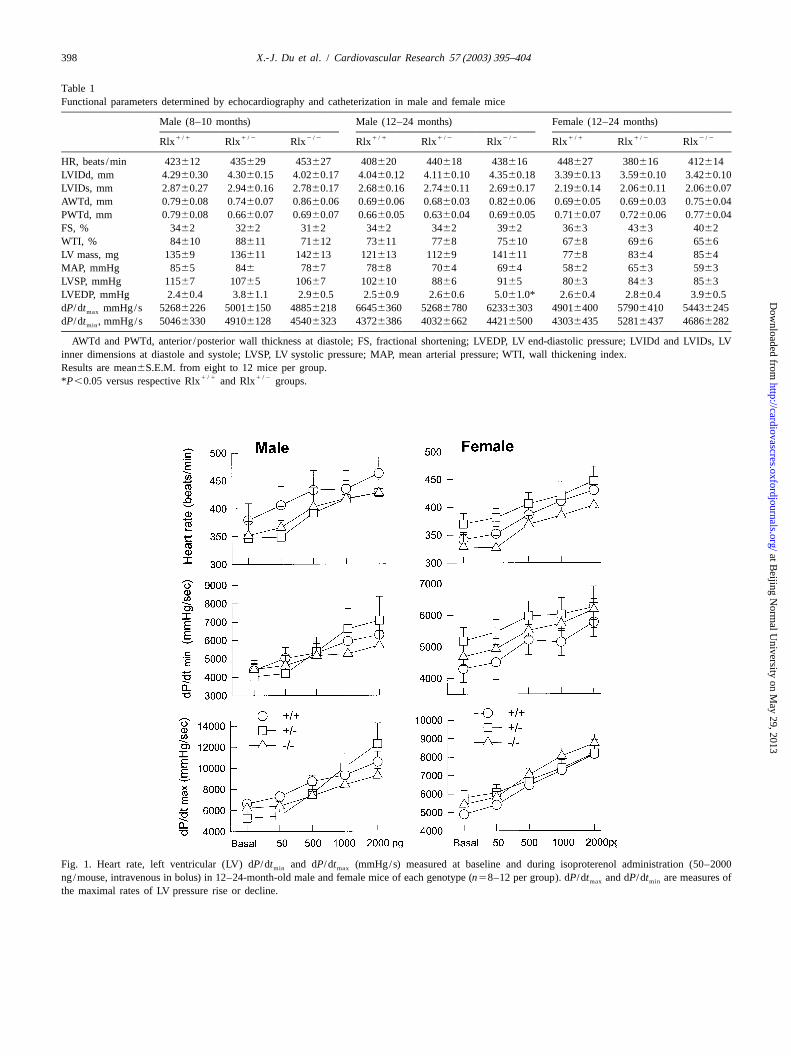
2 /2Rlx mice, it was not significant (Table 1). While theendogenous reference (18S ribosomal RNA) PCR re-catheter was positioned in the LV, the heart was challengedactions. Mouse-specific COL-1 and proMMP primers andby intravenous injection of isoproterenol. The responses ofprobes were designed using Primer Express (Appliedheart rate, dP/dt and dP/dt were similar in all threemax minBiosystems). The probes were labelled at the 59 end withgenotype groups of both genders (Fig. 1). These findingsFAM (6-carboxy fluorescein; Keystone Division,indicate normal chronotropic, inotropic and lusitropicBiosource International, Foster City, CA). The PCR re-
2 /2function in Rlx mice.actions were carried out in triplicate 25-ml volumes in anDiastolic function of the LV was further examined inABI PRISM 7700 Sequence Detector using the relative
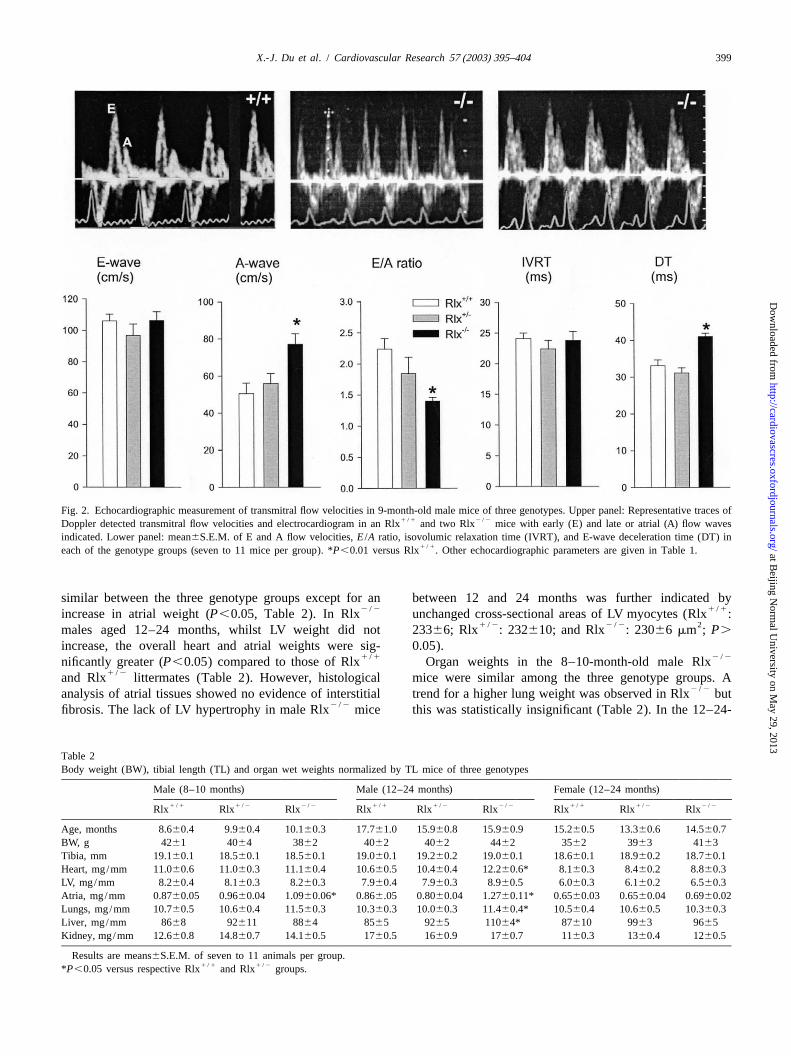
male mice aged 8–10 months by measuring Doppler flowstandard curve method, 96-well optical reaction plates and velocities across the mitral valves during the diastolicTaqMan PCR components (all Applied Biosystems) as
phase. All three genotype groups of mice had similar earlyoutlined previously [25]. In a further experiment, COL-1filling flow velocity (E-wave). However, the late fillingexpression was analysed in 6- and 9-month-old male
2 /21 /1 2 /2 flow velocity (A-wave) was higher in the Rlx males,Rlx and Rlx mice.
leading to a significant reduction in the ratio of E/A wavesThe presence of relaxin gene transcripts in the mouse(E /A ratio; Fig. 2). IVRT was similar between the threeatria and ventricles was assessed by reverse transcription
2 /2groups in keeping with an unchanged dP/dt in Rlxmin(RT)-PCR with mouse-specific relaxin oligonucleotide2 /2mice measured by catheter. However, Rlx mice had aprimers (100 ng/ml: forward 59-
significant lengthening of DT compared to other twoGTGAATATGCCCGTGAATTGATC and reverse 59-groups (P,0.01; Fig. 2), indicating a lower passiveAGCGTCGTATCGAAAGG CTCT) and 5-ml cDNA tem-compliance of the LV [21].plate in a 50-ml PCR reaction. Primers were selected to
span the intron/exon junction, thus favoring PCR products(150-bp) arising only from cDNA rather than genomic 3 .1.1. Morphometry
2 /2contamination. The quality of first strand cDNA synthesis In 8–10-month-old Rlx males, heart weight was
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from
398 X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404
Table 1Functional parameters determined by echocardiography and catheterization in male and female mice
Male (8–10 months) Male (12–24 months) Female (12–24 months)1 /1 1 /2 2 /2 1 /1 1 /2 2 /2 1 /1 1 /2 2 /2Rlx Rlx Rlx Rlx Rlx Rlx Rlx Rlx Rlx
HR, beats /min 423612 435629 453627 408620 440618 438616 448627 380616 412614LVIDd, mm 4.2960.30 4.3060.15 4.0260.17 4.0460.12 4.1160.10 4.3560.18 3.3960.13 3.5960.10 3.4260.10LVIDs, mm 2.8760.27 2.9460.16 2.7860.17 2.6860.16 2.7460.11 2.6960.17 2.1960.14 2.0660.11 2.0660.07AWTd, mm 0.7960.08 0.7460.07 0.8660.06 0.6960.06 0.6860.03 0.8260.06 0.6960.05 0.6960.03 0.7560.04PWTd, mm 0.7960.08 0.6660.07 0.6960.07 0.6660.05 0.6360.04 0.6960.05 0.7160.07 0.7260.06 0.7760.04FS, % 3462 3262 3162 3462 3462 3962 3663 4363 4062WTI, % 84610 88611 71612 73611 7768 75610 6768 6966 6566LV mass, mg 13569 136611 142613 121613 11269 141611 7768 8364 8564MAP, mmHg 8565 846 7867 7868 7064 6964 5862 6563 5963LVSP, mmHg 11567 10765 10667 102610 8866 9165 8063 8463 8563LVEDP, mmHg 2.460.4 3.861.1 2.960.5 2.560.9 2.660.6 5.061.0* 2.660.4 2.860.4 3.960.5dP/dt mmHg/s 52686226 50016150 48856218 66456360 52686780 62336303 49016400 57906410 54436245max
dP/dt , mmHg/s 50466330 49106128 45406323 43726386 40326662 44216500 43036435 52816437 46866282min
AWTd and PWTd, anterior /posterior wall thickness at diastole; FS, fractional shortening; LVEDP, LV end-diastolic pressure; LVIDd and LVIDs, LVinner dimensions at diastole and systole; LVSP, LV systolic pressure; MAP, mean arterial pressure; WTI, wall thickening index.Results are mean6S.E.M. from eight to 12 mice per group.
1 /1 1 /2*P ,0.05 versus respective Rlx and Rlx groups.
Fig. 1. Heart rate, left ventricular (LV) dP/dt and dP/dt (mmHg/s) measured at baseline and during isoproterenol administration (50–2000min max
ng/mouse, intravenous in bolus) in 12–24-month-old male and female mice of each genotype (n58–12 per group). dP/dt and dP/dt are measures ofmax min
the maximal rates of LV pressure rise or decline.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from
X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404 399
Fig. 2. Echocardiographic measurement of transmitral flow velocities in 9-month-old male mice of three genotypes. Upper panel: Representative traces of1 /1 2 /2Doppler detected transmitral flow velocities and electrocardiogram in an Rlx and two Rlx mice with early (E) and late or atrial (A) flow waves
indicated. Lower panel: mean6S.E.M. of E and A flow velocities,E /A ratio, isovolumic relaxation time (IVRT), and E-wave deceleration time (DT) in1 /1each of the genotype groups (seven to 11 mice per group). *P,0.01 versus Rlx . Other echocardiographic parameters are given in Table 1.
similar between the three genotype groups except for an between 12 and 24 months was further indicated by2 /2 1 /1increase in atrial weight (P,0.05, Table 2). In Rlx unchanged cross-sectional areas of LV myocytes (Rlx :
1 /2 2 /2 2males aged 12–24 months, whilst LV weight did not 23366; Rlx : 232610; and Rlx : 23066mm ; P.
increase, the overall heart and atrial weights were sig- 0.05).1 /1 2 /2nificantly greater (P,0.05) compared to those of Rlx Organ weights in the 8–10-month-old male Rlx
1 /2and Rlx littermates (Table 2). However, histological mice were similar among the three genotype groups. A2 /2analysis of atrial tissues showed no evidence of interstitial trend for a higher lung weight was observed in Rlx but
2 /2fibrosis. The lack of LV hypertrophy in male Rlx mice this was statistically insignificant (Table 2). In the 12–24-
Table 2Body weight (BW), tibial length (TL) and organ wet weights normalized by TL mice of three genotypes
Male (8–10 months) Male (12–24 months) Female (12–24 months)1 /1 1 /2 2 /2 1 /1 1 /2 2 /2 1 /1 1 /2 2 /2Rlx Rlx Rlx Rlx Rlx Rlx Rlx Rlx Rlx
Age, months 8.660.4 9.960.4 10.160.3 17.761.0 15.960.8 15.960.9 15.260.5 13.360.6 14.560.7BW, g 4261 4064 3862 4062 4062 4462 3562 3963 4163Tibia, mm 19.160.1 18.560.1 18.560.1 19.060.1 19.260.2 19.060.1 18.660.1 18.960.2 18.760.1Heart, mg/mm 11.060.6 11.060.3 11.160.4 10.660.5 10.460.4 12.260.6* 8.160.3 8.460.2 8.860.3LV, mg/mm 8.260.4 8.160.3 8.260.3 7.960.4 7.960.3 8.960.5 6.060.3 6.160.2 6.560.3Atria, mg/mm 0.8760.05 0.9660.04 1.0960.06* 0.866.05 0.8060.04 1.2760.11* 0.6560.03 0.6560.04 0.6960.02Lungs, mg/mm 10.760.5 10.660.4 11.560.3 10.360.3 10.060.3 11.460.4* 10.560.4 10.660.5 10.360.3Liver, mg/mm 8668 92611 8864 8565 9265 11064* 87610 9963 9665Kidney, mg/mm 12.660.8 14.860.7 14.160.5 1760.5 1660.9 1760.7 1160.3 1360.4 1260.5
Results are means6S.E.M. of seven to 11 animals per group.1 /1 1 /2*P ,0.05 versus respective Rlx and Rlx groups.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from
400 X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404
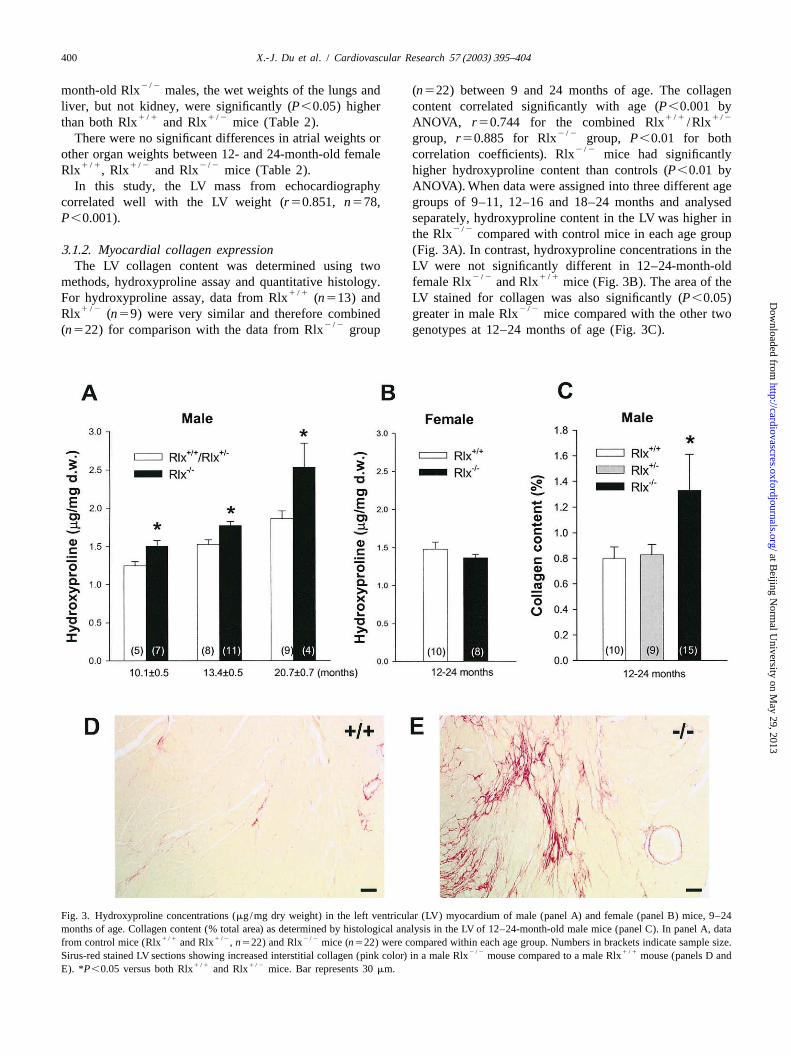
2 /2month-old Rlx males, the wet weights of the lungs and (n522) between 9 and 24 months of age. The collagenliver, but not kidney, were significantly (P,0.05) higher content correlated significantly with age (P,0.001 by
1 /1 1 /2 1 /1 1 /2than both Rlx and Rlx mice (Table 2). ANOVA, r50.744 for the combined Rlx /Rlx2 /2There were no significant differences in atrial weights or group,r50.885 for Rlx group, P,0.01 for both
2 /2other organ weights between 12- and 24-month-old female correlation coefficients). Rlx mice had significantly1 /1 1 /2 2 /2Rlx , Rlx and Rlx mice (Table 2). higher hydroxyproline content than controls (P,0.01 by
In this study, the LV mass from echocardiography ANOVA). When data were assigned into three different agecorrelated well with the LV weight (r50.851, n578, groups of 9–11, 12–16 and 18–24 months and analysedP,0.001). separately, hydroxyproline content in the LV was higher in
2 /2the Rlx compared with control mice in each age group3 .1.2. Myocardial collagen expression (Fig. 3A). In contrast, hydroxyproline concentrations in the
The LV collagen content was determined using two LV were not significantly different in 12–24-month-old2 /2 1 /1methods, hydroxyproline assay and quantitative histology. female Rlx and Rlx mice (Fig. 3B). The area of the
1 /1For hydroxyproline assay, data from Rlx (n513) and LV stained for collagen was also significantly (P,0.05)1 /2 2 /2Rlx (n59) were very similar and therefore combined greater in male Rlx mice compared with the other two
2 /2(n522) for comparison with the data from Rlx group genotypes at 12–24 months of age (Fig. 3C).
Fig. 3. Hydroxyproline concentrations (mg/mg dry weight) in the left ventricular (LV) myocardium of male (panel A) and female (panel B) mice, 9–24months of age. Collagen content (% total area) as determined by histological analysis in the LV of 12–24-month-old male mice (panel C). In panel A, data
1 /1 1 /2 2 /2from control mice (Rlx and Rlx ,n522) and Rlx mice (n522) were compared within each age group. Numbers in brackets indicate sample size.2 /2 1 /1Sirus-red stained LV sections showing increased interstitial collagen (pink color) in a male Rlx mouse compared to a male Rlx mouse (panels D and
1 /1 1 /2E). *P,0.05 versus both Rlx and Rlx mice. Bar represents 30mm.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from
X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404 401
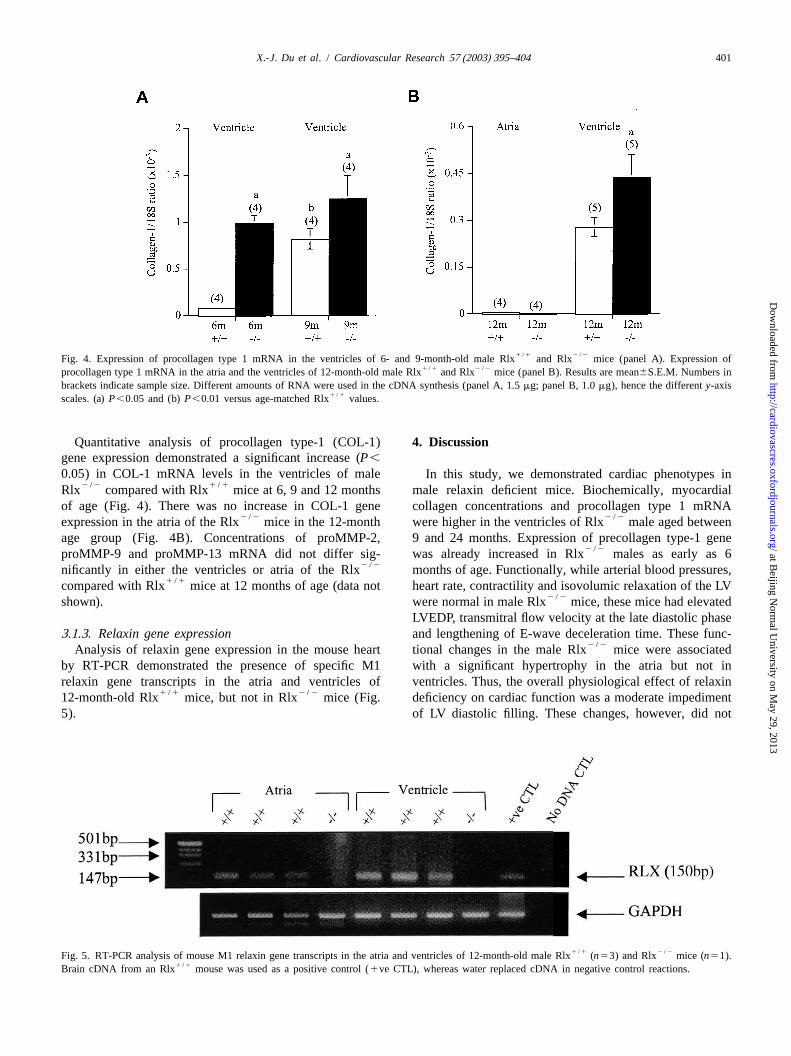
1 /1 2 /2Fig. 4. Expression of procollagen type 1 mRNA in the ventricles of 6- and 9-month-old male Rlx and Rlx mice (panel A). Expression of1 /1 2 /2procollagen type 1 mRNA in the atria and the ventricles of 12-month-old male Rlx and Rlx mice (panel B). Results are mean6S.E.M. Numbers in
brackets indicate sample size. Different amounts of RNA were used in the cDNA synthesis (panel A, 1.5mg; panel B, 1.0mg), hence the differenty-axis1 /1scales. (a)P,0.05 and (b)P,0.01 versus age-matched Rlx values.
Quantitative analysis of procollagen type-1 (COL-1) 4 . Discussiongene expression demonstrated a significant increase (P,
0.05) in COL-1 mRNA levels in the ventricles of male In this study, we demonstrated cardiac phenotypes in2 /2 1 /1Rlx compared with Rlx mice at 6, 9 and 12 months male relaxin deficient mice. Biochemically, myocardial
of age (Fig. 4). There was no increase in COL-1 gene collagen concentrations and procollagen type 1 mRNA2 /2 2 /2expression in the atria of the Rlx mice in the 12-month were higher in the ventricles of Rlx male aged between
age group (Fig. 4B). Concentrations of proMMP-2, 9 and 24 months. Expression of precollagen type-1 gene2 /2proMMP-9 and proMMP-13 mRNA did not differ sig- was already increased in Rlx males as early as 6
2 /2nificantly in either the ventricles or atria of the Rlx months of age. Functionally, while arterial blood pressures,1 /1compared with Rlx mice at 12 months of age (data not heart rate, contractility and isovolumic relaxation of the LV
2 /2shown). were normal in male Rlx mice, these mice had elevatedLVEDP, transmitral flow velocity at the late diastolic phase
3 .1.3. Relaxin gene expression and lengthening of E-wave deceleration time. These func-2 /2Analysis of relaxin gene expression in the mouse heart tional changes in the male Rlx mice were associated
by RT-PCR demonstrated the presence of specific M1 with a significant hypertrophy in the atria but not inrelaxin gene transcripts in the atria and ventricles of ventricles. Thus, the overall physiological effect of relaxin
1 /1 2 /212-month-old Rlx mice, but not in Rlx mice (Fig. deficiency on cardiac function was a moderate impediment5). of LV diastolic filling. These changes, however, did not
1 /1 2 /2Fig. 5. RT-PCR analysis of mouse M1 relaxin gene transcripts in the atria and ventricles of 12-month-old male Rlx (n53) and Rlx mice (n51).1 /1Brain cDNA from an Rlx mouse was used as a positive control (1ve CTL), whereas water replaced cDNA in negative control reactions.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from

402 X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404
lead to more pronounced cardiac dysfunction or mortality. may have led to a moderate increase in LV chamberstiffness and ventricular diastolic dysfunction.The cardiac phenotypes were not observed in aged-
2 /2 The reason for the increased lung and liver weights inmatched female Rlx littermates.2 /2male Rlx mice is not clear. This might be secondary toIn this study, we show that both atria and ventricles are
organ congestion due to impeded ventricular filling. How-putative sources of relaxin in the mouse heart, similar toever, this possibility is unlikely considering a well pre-the human [19]. High affinity relaxin binding sites are alsoserved dP/dt and dP/dt and only a modest increaselocalised to the rodent atria [11,12], implying that relaxin max min
2 /2in LVEDP in the aged male Rlx mice. A recent studyhas a direct action on the heart to influence cardiovascular2 /2reported that the Rlx mice had an age-related increaseactivity. These include chronotropic and inotropic effects
in collagen content and histological abnormalities in thestimulated by administration of relaxin both in vivo and inlung [30], providing an explanation for the increased lungvitro [13–15]. In this study, we did not observe any
2 /21 /1 2 /2 weights in the Rlx males in the current study. Itsignificant difference between Rlx and Rlx mice inremains to be established whether a collagen build-up alsoblood pressures, heart rate and ventricular inotropy andoccurs in the liver of these mice. Thus, the increasedlusitropy (dP/dt and dP/dt ) under basal and iso-max min
weights of lungs and liver are very likely the consequenceproterenol-stimulated conditions. Other cardiovascular ef-of elevated organ collagen content due directly to relaxinfects of exogenous relaxin are coronary, pulmonary anddeficiency.renal vasodilatation [17,26,27] and stimulation of release
There is now good evidence to show that relaxin actsof vasoactive peptides such as atrial natriuretic peptidedirectly on human fibroblasts of different organs [5,31,32][16] and vasopressin [28]. The vascular action of relaxin isto decrease the synthesis and secretion of interstitialmediated by nitric oxide and endothelin (ET-1) throughcollagens. Relaxin also increases the expression ofendothelin ET receptors [19,27,29]. We found that endo-B
proMMP-1, proMMP-2 and proMMP-3 and decreasesthelium-dependent vasodilatation and norepinephrine-in-production of tissue inhibitors of metalloproteinasesduced vasoconstriction in perfused aortic rings were
1 /1 2 /2 (TIMPs) in the human and porcine uterus [5,33]. Thesecomparable in Rlx and Rlx mice of both sexes (ourdata suggest a mechanism by which relaxin regulatesunpublished data), suggesting that relaxin deficiency doescollagen concentrations in tissues. In our study, an increasenot have a profound effect on vascular tone. Thus,in procollagen gene expression and collagen content in thealthough the cardiovascular system responds to exogenous
2 /2ventricles of Rlx mice was not accompanied by arelaxin, the long-term absence of endogenous relaxin insignificant alteration in MMP expression. However, MMPsmale and non-pregnant female mice does not result inare secreted in a proMMP form and are activated bysignificant alterations in hemodynamics.cleavage of the propeptide sequence. It has been demon-The functional measurement revealed a greater late-strated that relaxin treatment increases MMP activityphase flow velocity of LV diastolic filling due to atrialwithout affecting MMP protein expression in the porcinecontraction and lengthening of DT in 8–10-month-old
2 /2 uterus [33]. We did not examine MMP activity in themale Rlx mice, and a modestly elevated LVEDP in2 /22 /2 ventricles of Rlx mice, so it is possible that changes in12–24-month-old male Rlx mice. The atrial weight
MMP activity did occur without changes in MMP genewas increased in both age groups. Histological analysis ofexpression. A second explanation is that the level of MMPatrial tissues showed no evidence of interstitial fibrosis.
2 /2gene expression was very low in the hearts of Rlx miceTherefore, the weight increase in the atria is likely due toso we were unable to detect reductions in mRNA levels.hypertrophy of atrial myocytes. These changes are bestAdditionally, the low levels of MMPs present may notexplained by an increased ventricular impedance at the latehave been sufficient to inhibit collagen deposition observeddiastolic phase, resulting in atrial hypertrophy. An age-
2 /2in the Rlx mice. This possibility is supported by ourdependent increase in collagen content in the LV was2 /2 data showing progressive increases in LV collagen withobserved in Rlx males from 9 to 24 months of age. The
aging despite the presence of proMMP mRNA.LV diastolic function was studied in the 8–10-month-old2 /2 Recent studies using experimental animal models ofbut not in the 12–24-month-old male Rlx mice.
fibrosis in various organs have shown that administrationHowever, considering an age-dependent build-up of col-2 /2 of relaxin reduces the extent of fibrosis [30–35]. Thelagen in the LV, it is likely that the aged Rlx mice
2 /2would exhibit a more severe diastolic dysfunction than that cardiac phenotypes observed in male Rlx mice are inseen in their younger counterparts. Further experiments line with these studies and further support the view thatshowed increased procollagen type-1 gene expression in relaxin also regulates collagen synthesis and breakdown in
2 /2the ventricles of 6-, 9- and 12-month-old male Rlx non-reproductive tissues [30–35]. Dschietzig et al. re-mice. However, there was no indication of LV hypertrophy ported that myocardial relaxin expression and plasmain these mice as neither LV weight nor myocyte cross- relaxin concentrations increase profoundly in patients withsectional areas were significantly different from wildtype moderate to severe congestive heart failure [19]. Althoughmice. Thus, long-term disruption of endogenous relaxin in the implications of this finding remain unclear, relaxin maymale mice resulted in elevated LV interstitial collagen that function as a compensatory mediator and a marker for
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from

X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404 403
heart failure. Relaxin dilates coronary, renal and pulmon- Elodie Percy for technical assistance, Dr Ross Bathgate forary arteries by stimulating NO generation [29,36] and discussion and Dr Elaine Unemori for providing some ofantagonising vasoconstrictive peptides [26], and protects the real-time PCR primers and probes. This study wasthe myocardium from ischemia/ reperfusion injury [36]. supported by Australian National Health and MedicalWith all documented actions of exogenous relaxin, the Research Council Block Grants to the Baker Medicalcardiac phenotypes reported in the present study, and Research Institute and Howard Florey Institute. Dr Laurareported changes in relaxin levels in the failing heart [19], Parry holds an Australian Research Council QEII Fellow-it is feasible to speculate that relaxin plays an important ship.compensatory role in cardiovascular diseases. Furthermore,relaxin or relaxin-like compounds might form a class ofanti-fibrotic agents in pathological conditions, includingheart disease. R eferences
Although the mouse has two relaxin genes (mRlx-1 andmRlx-3) [6,7], it is unlikely that there is an upregulation in [1] Sherwood OD. In: 2nd ed, Relaxin. The physiology of reproduction,
vol. 1, New York: Raven Press, 1994, pp. 861–1009.mRlx-3 expression in the hearts of mRlx-1 knockout mice.1 /1 [2] Samuel CS, Butkus A, Coghlan JP, Bateman JF. The effect ofmRlx-3 transcripts are detected in the hearts of Rlx
relaxin on collagen metabolism in the non-pregnant rat pubicmice but only by RT-PCR [6], suggesting that mRlx-3symphysis: the influence of estrogen and progesterone in regulating
expression is extremely low in this organ. We have found relaxin activity. Endocrinology 1996;137:3884–3890.that mRlx-3 expression is not upregulated in the lungs of [3] Bryant-Greenwood GD, Schwabe C. Human relaxins: chemistry and
2 /2 biology. Endocrinol Rev 1994;15:5–26.12-month-old Rlx mice [30]. Furthermore, the re-[4] Samuel CS, Coghlan JP, Bateman JF. Effects of relaxin, pregnancyproductive and cardiac phenotypes associated with in-
and parturition on collagen metabolism in the rat pubic symphysis. Jcreased collagen and tissue fibrosis in mice lacking mRlx-1Endocrinol 1998;159:117–125.
are still clearly evident [8–10], despite the probable [5] Goldsmith LT, Weiss G, Palejwala S. Mechanism of relaxin actionpresence of the mRlx-3. This implies that mRlx-3 does not in human cervix. In: Tregear GW, Ivell R, Bathgate RAD, Wade JD,
editors, Relaxin 2000, Dordrecht: Kluwer Academic Publishers,compensate for the lack of mRlx-1 and does not influence2001, pp. 285–289.the phenotypes associated with mRlx-1 deficiency in mice.
[6] Bathgate RA, Samuel CS, Burazin TC et al. Human relaxin gene 3Unexpectedly, the cardiac phenotype was only observed(H3) and the equivalent mouse relaxin (M3) gene: novel members2 /2in male but not female Rlx mice. It has been reported of the relaxin peptide family. J Biol Chem 2002;277:1148–1157.
that matrix-remodeling activity of relaxin depends on or is [7] Evans BA, John M, Fowler KJ et al. The mouse relaxin gene:nucleotide sequence and expression. J Mol Endocrinol 1993;10:15–potentiated by female hormones, particularly estrogen
2 /2 23.[1,2]. The lack of cardiac phenotypes in female Rlx[8] Zhao L, Roche PJ, Gunnersen JM et al. Mice without a functionalmice suggests that other female hormones or factors
relaxin gene are unable to deliver milk to their pups. Endocrinology2 /2compensate for the lack of relaxin in female Rlx mice. 1999;140:445–453.Interestingly, recent studies in other gene-targeted mouse [9] Zhao L, Samuel CS, Tregear GW, Beck F, Wintour EM. Collagenstrains have shown that gender influences the development studies in late pregnant relaxin null mice. Biol Reprod 2000;63:697–
703.of cardiac phenotypes [37–40]. In these mouse strains,[10] Samuel CS, Amento EP. Relaxin knockout mice demonstratemales usually display more pronounced phenotypes than
delayed growth and have underdeveloped male reproductive tracts.females. Thus, gender is an important factor in theIn: Tregear GW, Ivell R, Bathgate RAD, Wade JD, editors, Relaxin
development of cardiac specific phenotypes in mice. 2000, Dordrecht: Kluwer Academic Publishers, 2001, pp. 221–223.In summary, our study provides evidence that deficiency [11] Tan YY, Wade JD, Tregear GW, Summers RJ. Quantitative au-
toradiographic studies of relaxin binding in rat atria, uterus andof endogenous relaxin results in an age-dependent increasecerebral cortex: characterization and effects of oestrogen treatment.in myocardial collagen content, atrial hypertrophy andBr J Pharmacol 1999;127:91–98.impeded ventricular diastolic filling but preserved myocar-
[12] Osheroff PL, Ho WH. Expression of relaxin mRNA and relaxindial function. These data indicate that under physiological receptors in postnatal and adult rat brains and hearts. Localizationconditions, relaxin is required for regulation of myocardial and developmental patterns. J Biol Chem 1993;268:15193–15199.
[13] Thomas GR, Vandlen R. The purely chronotropic effects of relaxincollagen content and hence maintenance of a normalin the rat isolated heart. J Pharm Pharmacol 1993;45:927–928.ventricular diastolic function. These cardiac phenotypes in
2 /2 2 /2 [14] Kakouris H, Eddie LW, Summers RJ. Cardiac effects of relaxin inRlx males were not observed in Rlx females,rats. Lancet 1992;339:1076–1078.
indicating the importance of other gender-related factors in [15] Parry LJ, Poterski RS, Summerlee AJS, Jones SA. Mechanism ofcardiovascular function. the haemotensive action of porcine relaxin in anaesthetized rats. J
Neuroendocrinol 1990;2:53–58.[16] Toth M, Taskinen P, Ruskoaho H. Relaxin stimulates atrial nat-
riuretic peptide secretion in perfused rat heart. J EndocrinolA cknowledgements1996;150:487–495.
[17] Bani-Sacchi T, Bigazzi M, Bani D, Mannaioni PF, Masini E.The authors thank Chongxin Zhao for mouse breeding Relaxin-induced increased coronary flow through stimulation of
and genotyping, Helen Gehring, Dr Martyn Lewis and nitric oxide production. Br J Pharmacol 1995;116:1589–1594.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from

404 X.-J. Du et al. / Cardiovascular Research 57 (2003) 395–404
[18] Taylor MJ, Clark CL. Evidence for a novel source of relaxin: atrial [30] Samuel CS, Zhao C, Bathgate RAD et al., Relaxin deficient mice arecardiocytes. J Endocrinol 1994;143:R5–R8. associated with an age-related progression of pulmonary fibrosis.
[19] Dschietzig T, Richter C, Bartsch C et al. The pregnancy hormone FASEB J (in press).relaxin is a player in human heart failure. FASEB J 2001;15:2187– [31] Unemori EN, Amento EP. Relaxin modulates synthesis and secretion2195. of procollagenase and collagen by human dermal fibroblasts. J Biol
[20] Gao XM, Dart AM, Dewar E, Jennings G, Du XJ. Serial ech- Chem 1990;265:10681–10685.ocardiographic assessment of left ventricular dimensions and func- [32] Unemori EN, Pickford LB, Salles AL et al. Relaxin induces antion after myocardial infarction in mice. Cardiovasc Res extracellular matrix-degrading phenotype in human lung fibroblasts2000;45:330–338. in vitro and inhibits lung fibrosis in a murine model in vivo. J Clin
[21] Nishmura RA, Abel MD, Hatle LK, Tajik AJ. Assessment of Invest 1996;98:2739–2745.diastolic function of the heart: background and current applications [33] Lenhart JA, Ryan PL, Ohleth KM, Palmer SS, Bagnell CA. Relaxinof Doppler echocardiography. Mayo Clin Proc 1989;64:181–204. increases secretion of matrix-metalloproteinase-2 and matrix
[22] Knollmann BC, Blatt SA, Horton K et al. Inotropic stimulation metalloproteinase-9 during uterine and cervical growth and remodel-induces cardiac dysfunction in transgenic mice expressing a troponin ling in the pig. Endocrinology 2001;142:3941–3949.T (I79N) mutation liked to familial hypertrophic cardiomyopathy. J [34] Williams EJ, Benyon RC, Trim N et al. Relaxin inhibits effectiveBiol Chem 2001;276:10039–10048. collagen deposition by cultured hepatic stellate cells and decreases
[23] Du XJ, Autelitano DJ, Dilley RJ, Wang B, Dart AM, Woodcock EA. rat liver fibrosis in vivo. Gut 2001;49:577–583.b -Adrenergic receptor overexpression exacerbates development of [35] Garber SL, Mirochnik Y, Brecklin CS et al. Relaxin decreases renal2
heart failure after aortic stenosis. Circulation 2000;101:71–77. interstitial fibrosis and slows progression of renal disease. Kidney[24] Young M, Fullerton M, Dilley R, Funder J. Mineralocorticoids, Int 2001;59:876–882.
hypertension, and cardiac fibrosis. J Clin Invest 1994;93:2578– [36] Bani D, Masini E, Bello MG, Bigazzi M, Sacchi TB. Relaxin2583. protects against myocardial injury caused by ischemia and reperfu-
[25] Siebel AL, Gehring HM, Nave CD, Bathgate RAD, Borchers CE, sion in rat heart. Am J Pathol 1998;152:1367–1376.Parry LJ. The upregulation of mesotocin receptors in the tammar [37] Kadokami T, McTiernan CF, Kubota T, Frye CS, Feldman AM.wallaby myometrium is pregnancy-specific and independent of Sex-related survival differences in murine cardiomyopathy areestrogen. Biol Reprod 2002;66:1237–1243. associated with differences in TNF-receptor expression. J Clin
[26] Fisher C, MacLean M, Morecroft I et al. Is the pregnancy hormone Invest 2000;106:589–597.relaxin also a vasodilator peptide secreted by the heart? Circulation [38] Gao XM, Dart AM, Percy E, Du XJ. Sex differences in car-2002;106:292–295. diomyopathy phenotype in mice overexpressingb -adrenoceptors in2
[27] Danielson LA, Kercher LJ, Conrad KP. Impact of gender and the heart. J Mol Cell Cardiol 2001;33:A37, (abstract).endothelin on renal vasodilation and hyperfiltration induced by [39] Cross HR, Lu L, Steenbergen C, Philipson KD, Murphy E.
1 21relaxin in conscious rats. Am J Physiol 2000;279:R1298–R1304. Overexpression of the cardiac Na /Ca exchanger increases[28] Geddes BJ, Parry LJ, Summerlee AJS. Brain angiotensin-II partially susceptibility to ischemia/ reperfusion injury in male, but not female,
mediates the effects of relaxin on vasopressin and oxytocin release transgenic mice. Circ Res 1998;83:1215–1223.in anesthetized rats. Endocrinology 1994;134:1188–1192. [40] Geisterfer-Lowrance AA, Christe M, Conner DA et al. A mouse
[29] Failli P, Nistri S, Quattrone S et al. Relaxin up-regulates inducible model of familial hypertrophic cardiomyopathy. Sciencenitric oxide synthase expression and nitric oxide generation in rat 1996;272:731–734.coronary endothelial cells. FASEB J 2002;16:252–254.
at Beijing N
ormal U
niversity on May 29, 2013
http://cardiovascres.oxfordjournals.org/D
ownloaded from




















