
Cell motility and mechanics in three-dimensional collagen matrices
30
Cell Motility and Mechanics in Three-Dimensional Collagen Matrices Frederick Grinnell 1 and W. Matthew Petroll 2 Departments of 1 Cell Biology and 2 Ophthalmology, University of Texas Southwestern Medical Center, Dallas, Texas 75390; email: [email protected], [email protected] Annu. Rev. Cell Dev. Biol. 2010. 26:335–61 First published online as a Review in Advance on May 3, 2010 The Annual Review of Cell and Developmental Biology is online at cellbio.annualreviews.org This article’s doi: 10.1146/annurev.cellbio.042308.113318 Copyright c 2010 by Annual Reviews. All rights reserved 1081-0706/10/1110-0335$20.00 Key Words biomechanics, extracellular matrix remodeling, Rho GTPases, tensional homeostasis, tissue engineering, wound healing Abstract Fibrous connective tissues provide mechanical support and frameworks for other tissues of the body and play an integral role in normal tis- sue physiology and pathology. Three-dimensional collagen matrices exhibit mechanical and structural features that resemble fibrous con- nective tissue and have become an important model system to study cell behavior in a tissue-like environment. This review focuses on motile and mechanical interactions between cells—especially fibroblasts—and collagen matrices. We describe several matrix contraction models, the interactions between fibroblasts and collagen fibrils at global and sub- cellular levels, unique features of mechanical feedback between cells and the matrix, and the impact of the cell-matrix tension state on cell mor- phology and mechanical behavior. We develop a conceptual framework to explain the balance between cell migration and collagen translocation including the concept of promigratory and procontractile growth factor environments. Finally, we review the significance of these concepts for the physiology of wound repair. 335 Annu. Rev. Cell Dev. Biol. 2010.26:335-361. Downloaded from www.annualreviews.org by University of Texas Southwestern Medical Center on 07/11/14. For personal use only.
Transcript of Cell motility and mechanics in three-dimensional collagen matrices

CB26CH14-Grinnell ARI 3 September 2010 19:34
Cell Motility and Mechanicsin Three-DimensionalCollagen MatricesFrederick Grinnell1 and W. Matthew Petroll2
Departments of 1Cell Biology and 2Ophthalmology, University of Texas SouthwesternMedical Center, Dallas, Texas 75390; email: [email protected],[email protected]
Annu. Rev. Cell Dev. Biol. 2010. 26:335–61
First published online as a Review in Advance onMay 3, 2010
The Annual Review of Cell and DevelopmentalBiology is online at cellbio.annualreviews.org
This article’s doi:10.1146/annurev.cellbio.042308.113318
Copyright c© 2010 by Annual Reviews.All rights reserved
1081-0706/10/1110-0335$20.00
Key Words
biomechanics, extracellular matrix remodeling, Rho GTPases,tensional homeostasis, tissue engineering, wound healing
Abstract
Fibrous connective tissues provide mechanical support and frameworksfor other tissues of the body and play an integral role in normal tis-sue physiology and pathology. Three-dimensional collagen matricesexhibit mechanical and structural features that resemble fibrous con-nective tissue and have become an important model system to study cellbehavior in a tissue-like environment. This review focuses on motileand mechanical interactions between cells—especially fibroblasts—andcollagen matrices. We describe several matrix contraction models, theinteractions between fibroblasts and collagen fibrils at global and sub-cellular levels, unique features of mechanical feedback between cells andthe matrix, and the impact of the cell-matrix tension state on cell mor-phology and mechanical behavior. We develop a conceptual frameworkto explain the balance between cell migration and collagen translocationincluding the concept of promigratory and procontractile growth factorenvironments. Finally, we review the significance of these concepts forthe physiology of wound repair.
335
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Contents
BEYOND REDUCTIONISM . . . . . . . . 336REGULATION OF MATRIX
CONTRACTION . . . . . . . . . . . . . . . . 337MATRIX MECHANICS . . . . . . . . . . . . . 341CELL MIGRATION. . . . . . . . . . . . . . . . . 343HIGH-RESOLUTION IMAGING
OF LOCAL MATRIXREMODELING . . . . . . . . . . . . . . . . . . 347
REGULATION OF LOCALMATRIX REMODELING BYSMALL G PROTEINS ANDMECHANICAL STIMULI. . . . . . . . 348
WOUND REPAIR ANDMATRIX CONTRACTION . . . . . . 350
CONCLUSIONS AND FUTUREDIRECTIONS . . . . . . . . . . . . . . . . . . . . 352
BEYOND REDUCTIONISM
Snatched from a life of obscurity and installed incontemporary glass and plastic palaces, cells are indanger of becoming Pygmalion’s proteges. Housedin more traditional residences constructed of waterand collagen instead of plastic or glass, do cells leadprimitive, less cultured lives?
Tom Elsdale and Jonathan Bard: “Collagensubstrata for studies on cell behavior” (Elsdale& Bard 1972)
In their 1924 review “Behavior of cells intissue cultures,” Lewis & Lewis summarizedthe contemporary field of tissue culture andits goal to reveal “new facts concerning thebehavior, structure, and physiology of theliving body cells” (Lewis & Lewis 1924).Diverse normal and malignant cell typeshad been observed to migrate out of tissuefragments onto the surface of glass coverslips,thereby making the cells accessible to inves-tigation. Cell pseudopodia were believed toprovide the mechanism for migration. Thenew methodology for establishing and growingtissue cells on glass coverslips rapidly replaced
the techniques of cell culture in 3D matricesprepared with clotted lymph and plasma thathad been introduced 15 years earlier.
When cells are cultured on glass coverslips,the physical framework with which the cellsinteract becomes invisible (literally and oftenfiguratively as well). Recognizing the impor-tance of hierarchical organization for biologicalfunction and structure, developmental biologistPaul Weiss continued to emphasize the need tolearn about cells not only as individuals (a re-ductionist approach) but also in the context ofthe interdependent relationship between cellsand their physical framework (a systems-basedapproach) (Weiss 1959). Figure 1a shows oneof the experiments used by Weiss to illustratesuch interdependence. Stretch (arrows) appliedat the corners of a plasma clot resulted in ori-entation of the fibrin. Mesenchymal cells in thestretched region of the clot developed bipolarmorphology parallel to the lines of tension. Inunstrained regions, where the fibrin lacked ten-sion, cells retained a more symmetric, stellateshape. Figure 1b illustrates a more complex ex-ample. In this case, three tissue fragments wereembedded in a fibrin matrix. Cellular activ-ity resulted in the development of tension andthe orientation of the fibrin between the frag-ments. These tension lines became tracks forsubsequent cell migration—evidence for con-tact guidance.
Acid-solubilized collagen, when coated oncoverslips and polymerized by ammonia vapor,forms an amorphous gel that can be used asa substrate to study cell-collagen interactions(Bornstein 1958). However, adjusting the col-lagen solution to physiological ionic strengthand pH first, and then initiating polymerizationby raising the temperature, produces hydratedcollagen lattices in which cells can be cultured(Elsdale & Bard 1972). Using their new tech-nique, Elsdale and Bard showed that fibroblastsexhibited features of shape, motility andgrowth in 3D collagen matrices not observedin routine 2D culture. Moreover, they made adistinction between cell behavior in restrainedmatrices (i.e., attached to culture dishes) versusfloating matrices (i.e., in culture medium).
336 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Under floating conditions there occurred acell-dependent “gradual collapse of the latticeto a dense, opaque body less than one-tenththe original size” (Elsdale & Bard 1972).
Subsequently, Michalopoulos and Pitot(1975) cultured liver parenchymal cells on thesurfaces of restrained and floating collagen ma-trices. They reported that cells on floating ma-trices caused the matrix to decrease in diameter,which they described as contraction. Also, cellson floating but not restrained matrices exhib-ited morphologic and biochemical character-istics of differentiated liver parenchymal cells,including formation of structures resemblingbile canaliculi (Michalopoulos & Pitot 1975).Similarly, Emerman & Pitelka (1977) observedthat mammary epithelial cells cultured on thesurfaces of floating collagen matrices causedmatrix contraction and retained morphologi-cal and biochemical characteristics of differen-tiated mammary epithelium including the de-velopment of alveolar-like structures within thematrices.
Although others had noticed the phe-nomenon of collagen matrix contraction,Eugene Bell made the contraction process itselfa problem for investigation. He suggested thatunderstanding how fibroblasts contract a col-lagen matrix into a tissue-like structure mightprovide insights into wound contraction (Bellet al. 1979). Moreover, he used contracted col-lagen matrices containing fibroblasts as the der-mal equivalent component of in vitro skin grafts(Bell et al. 1981), which became a foundingtechnology of the tissue engineering companyOrganogenesis.
Type I collagen is the major protein com-ponent of fibrous connective tissues, and fi-broblasts are the cell type primarily responsi-ble for collagen biosynthesis and remodeling.Because fibrous connective tissues provide me-chanical support and frameworks for the othertissues of the body, collagen matrix contractioncan influence diverse aspects of normal tissuephysiology and pathology. This review will fo-cus on motile and mechanical interactions be-tween cells—especially fibroblasts—and colla-gen matrices, the relationship between matrix
a
b
Figure 1Effect of regional tension on cell shape and migration. (a) Stretch (arrows)applied at the corners of a plasma clot results in orientation of the fibrin.Mesenchymal cells in the stretched region developed bipolar morphologyparallel to the lines of tension. In unstrained regions, cells retained a moresymmetric, stellate shape. (b) Cell migration along tension lines connectingthree tissue fragments in a fibrin clot. Reprinted from Weiss (1959) withpermission of the American Physical Society.
contraction and cell migration, and the signifi-cance of matrix contraction for cell physiology.Several recent reviews provide more generaldiscussions regarding the use of collagen andother 3D matrices to model the morphogene-sis of normal and malignant cells (e.g., Davis &Senger 2008, Griffith & Swartz 2006, Xu et al.2009, Yamada & Cukierman 2007).
REGULATION OF MATRIXCONTRACTION
Many cell types including fibroblasts, smoothmuscle cells, epithelial cells, and endothelial
www.annualreviews.org • Cells in 3D Collagen Matrices 337
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Cell traction(specific): rearwardforce transmitted to asubstrate during cellextension, analogousto a tire on the road
Tractional force(generic): forcetransmitted to asubstrate regardless ofmechanism
Stiffness: a measureof the resistanceoffered by an elasticbody to deformation(bending, stretching,or compression)
cells contract collagen matrices (Supplemen-tal Table 1; follow the Supplemental Mate-rial link from the Annual Reviews home page athttp://www.annualreviews.org). Therefore,matrix contraction is a highly conserved fea-ture of cell motile behavior similar to cell adhe-sion, spreading, and migration. Given the po-tential of matrix contraction to influence tissuestructure, understanding the basic mechanismsand regulation of collagen matrix contractionhas become an important design considerationfor tissue engineering (Brown et al. 2005, Clarket al. 2007, Lutolf & Hubbell 2005).
Although we use the word fibroblast in ageneric sense, it should be recognized that fi-broblasts represent a diverse cell population(Rinn et al. 2008, Sorrell & Caplan 2009). Con-siderable variation in matrix contraction by dif-ferent types of fibroblasts has been reported.For instance, papillary fibroblasts contract col-lagen matrices better than reticular fibroblasts(Schafer et al. 1989). Wound fibroblasts con-tract matrices better than dermal fibroblasts(Germain et al. 1994). Early passage fibro-blasts contract matrices better than establishedcell lines, which contract matrices better thanSV40-transformed cells (Steinberg et al. 1980).
The term collagen matrix contraction canbe used to refer to the final organization of thematrix or to the cellular mechanisms responsi-ble for achieving that organization. Early find-ings suggested that matrix contraction couldresult from cell traction (rearward force ex-erted during cell extension) or cell contraction(inward force exerted by cell shortening) (Bellet al. 1979, Bellows et al. 1982, Stopak & Harris1982). The expression “tractional force” is of-ten used in a generic sense to refer to any forceexerted on the matrix by cells regardless of themechanism. We will follow that trend but indi-cate the mechanism where appropriate.
Matrix contraction was initially found to de-pend on serum stimulation (Steinberg et al.1980); however, a wide range of physiologi-cally relevant growth factors have been shownto substitute for serum, many of which activatedifferent signaling pathways (SupplementalTable 2; follow the Supplemental Material
link from the Annual Reviews home page athttp://www.annualreviews.org). In the ab-sence of added growth factors, matrix contrac-tion becomes dependent on autocrine or (in thecase of mixed cell cultures) paracrine cell secre-tion. Also, collagen matrix contraction requiresintegrin-dependent adhesive interactions, andmany different collagen-binding integrins canfunction in matrix contraction including α1β1(Racine-Samson et al. 1997), α2β1 (Klein et al.1991, Schiro et al. 1991), α3β1 (Morales et al.2007), α11β1 (Tiger et al. 2001), and αvβ3(Cooke et al. 2000). No role in matrix con-traction has yet been described for collagen-binding tyrosine kinase receptors (Vogel et al.2006).
Given the multiple cell types, mechanismsfor exerting force, receptors, and signalingpathways involved in collagen matrix contrac-tion, rather than a singular view of the pro-cess, an overall conceptual framework can bedescribed best. Cells interacting with collagenmatrices form a system. Global factors such aswhether the matrix is floating or restrained aswell as local factors such as matrix stiffness in-fluence the pattern and mechanism of contrac-tion. Some of the most important variables thatwe will mention are low versus high cell-matrixtension states, cell tractional force relative tocollagen matrix stiffness, and promigratory ver-sus procontractile growth factor environments.
Figure 2 shows three interrelated modelsthat have been used to study the signaling,regulatory, and motor mechanisms involved incollagen matrix contraction (Grinnell 2000).Floating matrix contraction occurs when cell-containing matrices are released from the cul-ture surfaces soon after matrix polymeriza-tion. During and after contraction, the cellsexhibit few actin stress fibers and focal adhe-sions (low cell-matrix tension state). Restrainedmatrix contraction occurs when cells are in-cubated with matrices that remain attached tothe culture surfaces. During contraction, ma-trix height decreases, cells and collagen be-come aligned parallel to the culture surface,and prominent actin stress fibers and focal ad-hesions form (high cell-matrix tension state).
338 Grinnell · Petroll
Supplemental Material
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
LPA:lysophosphatidic acid
PDGF: platelet-derived growth factor
Stress-relaxation occurs upon release of re-strained matrices that previously have devel-oped tension. During stress-relaxation, actinstress fibers disappear, fibronectin matrix andfibronexus junctions are disrupted, and cellsand collagen lose their aligned orientation. Asa practical matter, floating matrix contractionand stress-relaxation are easily quantified bymeasuring the change in matrix diameter.
Serum can stimulate fibroblast contractionof collagen matrices in all three of the mod-els shown in Figure 2. Research using re-strained matrices attached to a force moni-tor showed that serum and the serum lipidgrowth factor lysophosphatidic acid (LPA)were far more effective than platelet-derivedgrowth factor (PDGF) in stimulating contrac-tile force generation by fibroblasts in restrainedcollagen matrices (Kolodney & Elson 1993).Subsequent work demonstrated that PDGFand LPA stimulate collagen matrix contrac-tion by different mechanisms (Grinnell 2000).Figure 3 summarizes key steps in the sig-naling pathways involved (Rhee & Grinnell2006). Three points are worth special emphasis.First, cell-matrix tension state influences whichgrowth factors stimulate collagen matrix con-traction. Both PDGF and LPA stimulate float-ing matrix contraction; only LPA stimulatesstress-relaxation. Second, although LPA stim-ulates collagen matrix contraction at both lowand high cell-matrix tension states, differentsignaling pathways and motor mechanisms areinvolved. LPA-stimulated floating contraction(low cell-matrix tension state) involves p21-activating kinase 1 (PAK1) in cooperation withthe Rho effector mDia1; myosin II is not re-quired. LPA-stimulated stress-relaxation (highcell-matrix tension state) depends on Rho ki-nase and myosin II. Third, although stimu-lation of floating matrix contraction by bothPDGF and LPA involves PAK1 signaling, dif-ferent Rho effectors cooperate with PAK1: Rhokinase in the case of PDGF and mDia1 in thecase of LPA. In general, stimulation of matrixcontraction by serum and by LPA exhibit sim-ilar features.
Fibroblasts polymerizedin collagen matrix
Floating matrix contraction
(dermal equivalent)
ReleaseRestrained matrix
contraction(tension develops)
Culture
12 mm
4–6 mmStress-relaxation
(tension dissipated)
Release
Figure 2Models of collagen matrix contraction. Floating matrix contraction occurswhen cell-containing matrices are released from culture surfaces soon aftermatrix polymerization. Restrained matrix contraction occurs when cells areincubated with matrices that remain attached to the culture surfaces.Stress-relaxation occurs upon release of restrained matrices that previouslyhave developed tension. Adapted from Grinnell (2000).
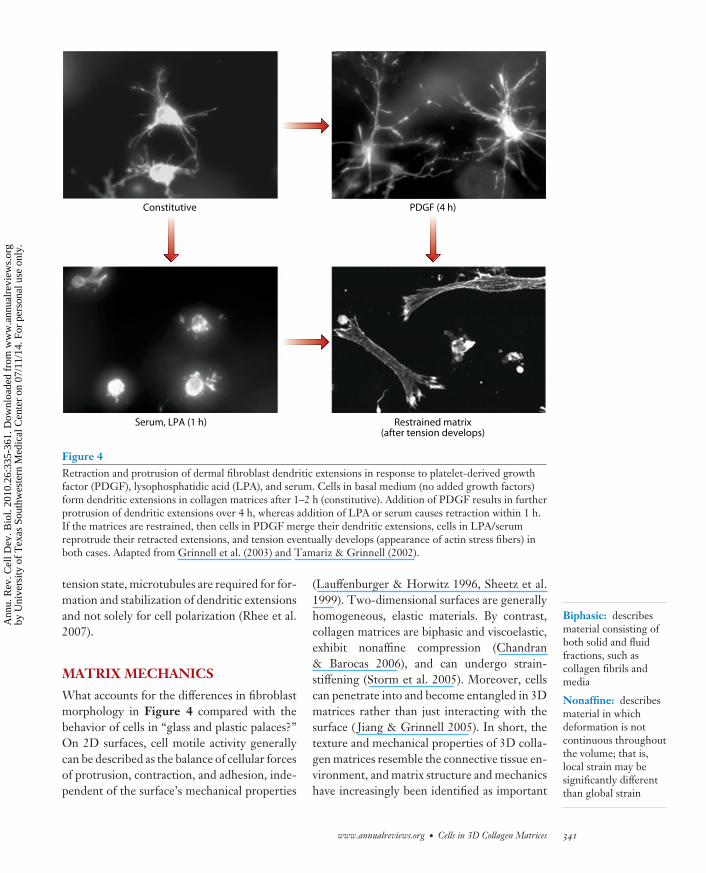
The images of cell morphology in Figure 4help explain the basis for the alternative reg-ulatory and motor mechanisms described inFigure 3. In newly polymerized collagen ma-trices, early passage human fibroblasts constitu-tively form thin, highly branched pseudopodialextensions, giving the cells a dendritic appear-ance. PDGF stimulates further protrusion ofthe dendritic extensions by a Rac- and PAK1-dependent mechanism. LPA or serum stim-ulates their retraction by a Rho/Rho kinase/myosin II–dependent mechanism (Rhee &Grinnell 2007). These cellular responses rep-resent exaggerated versions of the cell ruf-fling (PDGF) and contraction (serum or LPA)that fibroblasts exhibit on conventional 2D cul-ture surfaces (Hall 1998). In restrained collagenmatrices, cell morphology matures as tensiondevelops. In PDGF-containing medium, den-dritic extensions merge; in serum- and LPA-containing media, retracted extensions repro-trude. In both cases, stress fibers and focal
www.annualreviews.org • Cells in 3D Collagen Matrices 339
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
PDGF
PI3 kinase Gαi
RhoRho (basal) Rho
Rho kinaseMyosin IImDia1PAK1
Cell rufflingtraction
Floating matrixcontraction
Cell shorteningcontraction
Stress-relaxation
Rho kinaseMyosin II
LPA
a Low cell-matrix tension state b High cell-matrix tension state
LPA
Figure 3Growth factor regulation of collagen matrix contraction by dermal fibroblasts. Cell-matrix tension stateinfluences which growth factors stimulate collagen matrix contraction and the signaling pathways and motormechanisms involved. Gαi, G protein subunit αi; LPA, lysophosphatidic acid; PGDF, platelet-derivedgrowth factor; PI3 kinase, phosphatidylinositol 3-kinase. Adapted from Rhee & Grinnell (2006).
adhesions eventually develop (Tamariz &Grinnell 2002). Therefore, the cells themselvesare phenotypically different under the differentexperimental conditions shown in Figure 3.
In retrospect, it has become clear thatfloating matrix contraction describes outcome,not mechanism, and depends on the tractiongenerated by cell ruffling—hand-over-handcycles of local pseudopodial protrusion andretraction (Glaser et al. 1987). More thanone motor mechanism appears capable ofgenerating the mechanical force necessaryfor collagen translocation during ruffling. Forinstance, following Rac-dependent protrusion,collagen movement toward cells can occurduring Rho/Rho kinase/myosin II–dependentcell retraction (Meshel et al. 2005). Alterna-tively, corneal fibroblasts in 3D matrices can
pull collagen inward during cell protrusion(Petroll et al. 2008a, Roy et al. 1997). Incontrast to floating matrices, cell contraction(shortening) rather than cell ruffling providesthe mechanism for stress-relaxation (Grinnell2003, Tomasek et al. 2002).
Disrupting the cells’ actin cytoskeletonblocks all mechanisms of collagen matrix con-traction, but the effect of disrupting micro-tubules depends on the cell-matrix tensionstate. On one hand, if fibroblasts already areunder tension, then disrupting microtubules in-creases force generation (Brown et al. 1996,Kolodney & Wysolmerski 1992) and promotesstress-relaxation (Lee et al. 2003). On the otherhand, disrupting microtubules of cells in float-ing collagen matrices inhibits contraction (Bellet al. 1979, Lee et al. 2003). At a low cell-matrix
340 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
Constitutive
Serum, LPA (1 h) Restrained matrix(after tension develops)
PDGF (4 h)
Figure 4Retraction and protrusion of dermal fibroblast dendritic extensions in response to platelet-derived growthfactor (PDGF), lysophosphatidic acid (LPA), and serum. Cells in basal medium (no added growth factors)form dendritic extensions in collagen matrices after 1–2 h (constitutive). Addition of PDGF results in furtherprotrusion of dendritic extensions over 4 h, whereas addition of LPA or serum causes retraction within 1 h.If the matrices are restrained, then cells in PDGF merge their dendritic extensions, cells in LPA/serumreprotrude their retracted extensions, and tension eventually develops (appearance of actin stress fibers) inboth cases. Adapted from Grinnell et al. (2003) and Tamariz & Grinnell (2002).
Biphasic: describesmaterial consisting ofboth solid and fluidfractions, such ascollagen fibrils andmedia
Nonaffine: describesmaterial in whichdeformation is notcontinuous throughoutthe volume; that is,local strain may besignificantly differentthan global strain
tension state, microtubules are required for for-mation and stabilization of dendritic extensionsand not solely for cell polarization (Rhee et al.2007).
MATRIX MECHANICS
What accounts for the differences in fibroblastmorphology in Figure 4 compared with thebehavior of cells in “glass and plastic palaces?”On 2D surfaces, cell motile activity generallycan be described as the balance of cellular forcesof protrusion, contraction, and adhesion, inde-pendent of the surface’s mechanical properties
(Lauffenburger & Horwitz 1996, Sheetz et al.1999). Two-dimensional surfaces are generallyhomogeneous, elastic materials. By contrast,collagen matrices are biphasic and viscoelastic,exhibit nonaffine compression (Chandran& Barocas 2006), and can undergo strain-stiffening (Storm et al. 2005). Moreover, cellscan penetrate into and become entangled in 3Dmatrices rather than just interacting with thesurface ( Jiang & Grinnell 2005). In short, thetexture and mechanical properties of 3D colla-gen matrices resemble the connective tissue en-vironment, and matrix structure and mechanicshave increasingly been identified as important
www.annualreviews.org • Cells in 3D Collagen Matrices 341
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Tensionalhomeostasis: themaintenance ofconstant mechanicaltension within cells,matrices, or tissues
factors in determining cell behavior (Baro-cas & Tranquillo 1997, Georges & Janmey2005, Pedersen & Swartz 2005, Plant et al.2009, Silver et al. 2002, Wakatsuki et al. 2000).Cells and matrix interact to develop tensionalhomeostasis (Brown et al. 1998, Delvoye et al.1991, Paszek et al. 2005, Petroll et al. 2004b).Biomechanical features of contracted collagenmatrices resemble dermis (Ahlfors & Billiar2007). Application of external forces such ascyclic strain during contraction can influencecell orientation and final mechanical strength(Seliktar et al. 2000).
Based on rheometric measurements, theelastic modulus of newly polymerized 1–2 mg ml−1 collagen matrices, the concentra-tion range frequently utilized for collagen ma-trix contraction experiments, is less than 50 Pa(Barocas et al. 1995, Leung et al. 2007). By con-trast, the stiffness of the plastic polymers andglass used as tissue culture surfaces is >1 GPa.How best to determine the mechanical proper-ties of collagen matrices remains an open ques-tion. Rheometry describes bulk phase materialcharacteristics but provides limited informationabout local mechanical properties. Other meth-ods such as nanoindentation or atomic forcemicroscopy can be used to acquire the latterinformation.
Soft 2D surfaces increasingly are being usedto study how cells exert force on a substrateand how mechanical features of the substrateregulate cell motile function (Discher et al.2005, Munevar et al. 2001, Rehfeldt et al.2007). Increasing substrate stiffness increasesthe formation of actin stress fibers and focaladhesions (Engler et al. 2004, Pelham & Wang1997, Yeung et al. 2005). Overlaying fibroblastswith soft 2D surfaces can cause a switch from2D to more 3D morphology (Beningo et al.2004). However, only fibroblasts interactingwith collagen and other 3D matrices formdendritic extensions. As soon as cells beginto experience tension, dendritic extensionsundergo myosin II–dependent reorganizationinto more lamellar structures (Rhee et al.2007). For fibroblasts on the surfaces ofcollagen matrices, this switch occurs when the
collagen concentration of the matrix is in-creased from 1 mg ml−1 to 4 mg ml−1 (Rheeet al. 2010).
Understanding the dendritic features of fi-broblasts is particularly important because rest-ing connective tissues contain fibroblasts ina dendritic network (Goldsmith et al. 2004,Langevin et al. 2004, Salomon et al. 1988). In-teraction between this network and the sur-rounding collagen extracellular matrix pro-vides a mechanism for homeostatic tissuemechanosensing in response to diverse inputsranging from interstitial fluid flow (Heuchelet al. 1999, Pedersen et al. 2007) to stretch(Langevin et al. 2005) to tissue loss (Grinnellet al. 2003, Tomasek et al. 2002). Moreover,accumulated defects in the matrix as a resultof aging disrupt integration between the fi-broblast network and surrounding collagen ma-trix, thereby interfering with normal mechani-cal homeostasis (Fisher et al. 2008).
Collagen matrix contraction requires thefibril network to be interconnected, which al-lows local force to be propagated over long dis-tances (Barocas & Tranquillo 1997, Sawhney& Howard 2002, Vanni et al. 2003). Duringcontraction, cell-independent noncovalent in-teractions rapidly stabilize reorganized colla-gen fibrils. These interactions occur in the ab-sence of collagen telopeptides; however, thepresence of telopeptides increases contrac-tion and alters collagen fibril packing (Freyet al. 1995, Wolf et al. 2009, Woodley et al.1991). During restrained matrix contraction,the matrix itself develops tension. After re-lease, cell-independent matrix contraction oc-curs (Grinnell & Ho 2002, Marenzana et al.2006).
In addition to biologically polymerizedcollagen matrices and collagen matrices formedby electrospinning (Powell & Boyce 2009),fibroblasts can contract diverse other types ofmatrices ranging from fibrin (Tuan & Grinnell1989) to large-pore (140 μm) collagen–chondroitin sulfate composites (Freyman et al.2002). Increasing collagen matrix stiffness bychemical cross-linking with nonenzymaticglycation inhibits matrix contraction (Howard
342 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
TGFβ: transforminggrowth factor β
MMP: matrixmetalloproteinase
Porosity: the ratio ofthe volume of amaterial’s pores to itstotal volume
et al. 1996). Embedding noncompressibleelements into the matrix such as insolublecollagen fragments (Gentleman et al. 2004),single-walled carbon nanotubes (MacDonaldet al. 2005) or cross-linked hyaluronan (Mehraet al. 2006) slows down the rate of contraction.However, hyaluronan by itself has little effecton matrix contraction even under conditionsshown to alter matrix mechanics (Kreger& Voytik-Harbin 2009). Also, fibronectinprobably is not required for collagen matrixcontraction (Tomasek & Akiyama 1992). How-ever, fibronectin was shown to increase matrixcontraction by fibronectin-null fibroblasts(Hocking et al. 2000).
The ability of matrix composition and me-chanics to influence growth factor binding andactivation provides additional mechanisms forindirect regulation of collagen matrix contrac-tion (Macri et al. 2007, Wipff et al. 2007).Stimulation of contraction caused by throm-bospondin and inhibition of contraction causedby decorin have been linked to the ability ofthese matrix components to modify the avail-ability of transforming growth factor β (TGFβ)(Sakai et al. 2003, Zhang et al. 2009). Ma-trix components also can exert indirect ef-fects on contraction by modulating cell sig-naling. Tenascin C inhibits fibronectin-fibrinmatrix contraction by blocking cell signalingdownstream of Rho activation (Midwood &Schwarzbauer 2002).
The role of matrix metalloproteinases(MMPs) in collagen matrix contraction re-mains unclear. Fibroblasts from stomelysin-1null (Bullard et al. 1999) and MMP-9 nullbut not MMP-2 null mice ( Johnson & Galis2004) show reduced matrix contraction. How-ever, matrix contraction occurs in the presenceof serum, which contains high levels of solu-ble proteinase inhibitors, and little gross degra-dation of the matrix occurs during contrac-tion. Rather than soluble metalloproteinases,membrane-tethered MT1-MMP seems themost likely candidate to play a regulatory rolein collagen matrix contraction (Deryugina et al.1998) through its ability to modify the pericel-lular microenvironment (Itoh & Seiki 2006).
CELL MIGRATION
Thus far, our discussion has focused on globalcontraction of collagen matrices. What aboutthe experimental result described in Figure 1b?As already mentioned, Weiss recognized thattension between tissue fragments resulted in lo-cal fibrin alignment and that tension lines inthe fibrin matrix became the tracks along whichcells migrated. Stopak & Harris, working withtissue explants in collagen matrices, showed thatcellular mechanical activity was responsible forgenerating the tension. They emphasized therole of cell mechanics in connective tissue mor-phogenesis (Stopak & Harris 1982). Throughlocal matrix contraction, migrating cells can es-tablish tracks for their own migration (Friedlet al. 1997, Provenzano et al. 2006, Tranquillo1999) or for the migration of other cells(Gaggioli et al. 2007). Similarly, imposing ten-sion externally on polymeric 3D matrices alsocan establish aligned tracks for cell migra-tion (Guido & Tranquillo 1993, Raeber et al.2008).
If cells migrated only on the surfaces ofaligned fibrils in a 3D matrix, then individualfibrils might become singular migration paths(Doyle et al. 2009). But collagen matrices fre-quently contain fibrils that lack a singular ori-entation, and fibroblasts can interact with fib-rils oriented both parallel and perpendicular tothe direction of cell pseudopodia (Petroll et al.2008b). As a result, cell-induced alignment ofcollagen fibrils is not necessarily confined tothe axis of cell movement (Friedl et al. 1998,Petroll et al. 2004a). Moreover, the porosityof 3D matrices establishes the possibility ofdual modes of cell migration—mesenchymaland amoeboid (Friedl & Brocker 2000). In mes-enchymal migration, the cells attach to and pullon the matrix. In amoeboid migration, cells pro-trude through the spaces and push off the ma-trix. With tumor cells, Rho activation favors theamoeboid mode of migration; Rac activationfavors the mesenchymal mode (Sanz-Morenoet al. 2008).
Matrix porosity makes possible multiplemigratory mechanisms but also establishes
www.annualreviews.org • Cells in 3D Collagen Matrices 343
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
Dermalequivalent
Reembed incell-free matrix
Act
in
Bead
s
Restrained nestedmatrix
Floating nestedmatrix
a
b
c
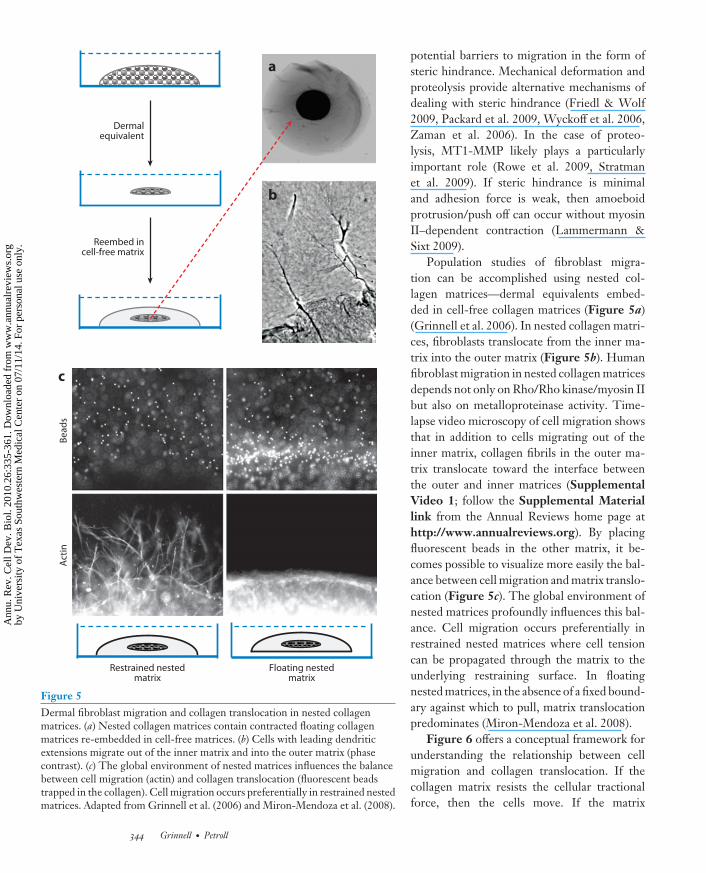
Figure 5Dermal fibroblast migration and collagen translocation in nested collagenmatrices. (a) Nested collagen matrices contain contracted floating collagenmatrices re-embedded in cell-free matrices. (b) Cells with leading dendriticextensions migrate out of the inner matrix and into the outer matrix (phasecontrast). (c) The global environment of nested matrices influences the balancebetween cell migration (actin) and collagen translocation (fluorescent beadstrapped in the collagen). Cell migration occurs preferentially in restrained nestedmatrices. Adapted from Grinnell et al. (2006) and Miron-Mendoza et al. (2008).
potential barriers to migration in the form ofsteric hindrance. Mechanical deformation andproteolysis provide alternative mechanisms ofdealing with steric hindrance (Friedl & Wolf2009, Packard et al. 2009, Wyckoff et al. 2006,Zaman et al. 2006). In the case of proteo-lysis, MT1-MMP likely plays a particularlyimportant role (Rowe et al. 2009, Stratmanet al. 2009). If steric hindrance is minimaland adhesion force is weak, then amoeboidprotrusion/push off can occur without myosinII–dependent contraction (Lammermann &Sixt 2009).
Population studies of fibroblast migra-tion can be accomplished using nested col-lagen matrices—dermal equivalents embed-ded in cell-free collagen matrices (Figure 5a)(Grinnell et al. 2006). In nested collagen matri-ces, fibroblasts translocate from the inner ma-trix into the outer matrix (Figure 5b). Humanfibroblast migration in nested collagen matricesdepends not only on Rho/Rho kinase/myosin IIbut also on metalloproteinase activity. Time-lapse video microscopy of cell migration showsthat in addition to cells migrating out of theinner matrix, collagen fibrils in the outer ma-trix translocate toward the interface betweenthe outer and inner matrices (SupplementalVideo 1; follow the Supplemental Materiallink from the Annual Reviews home page athttp://www.annualreviews.org). By placingfluorescent beads in the other matrix, it be-comes possible to visualize more easily the bal-ance between cell migration and matrix translo-cation (Figure 5c). The global environment ofnested matrices profoundly influences this bal-ance. Cell migration occurs preferentially inrestrained nested matrices where cell tensioncan be propagated through the matrix to theunderlying restraining surface. In floatingnested matrices, in the absence of a fixed bound-ary against which to pull, matrix translocationpredominates (Miron-Mendoza et al. 2008).
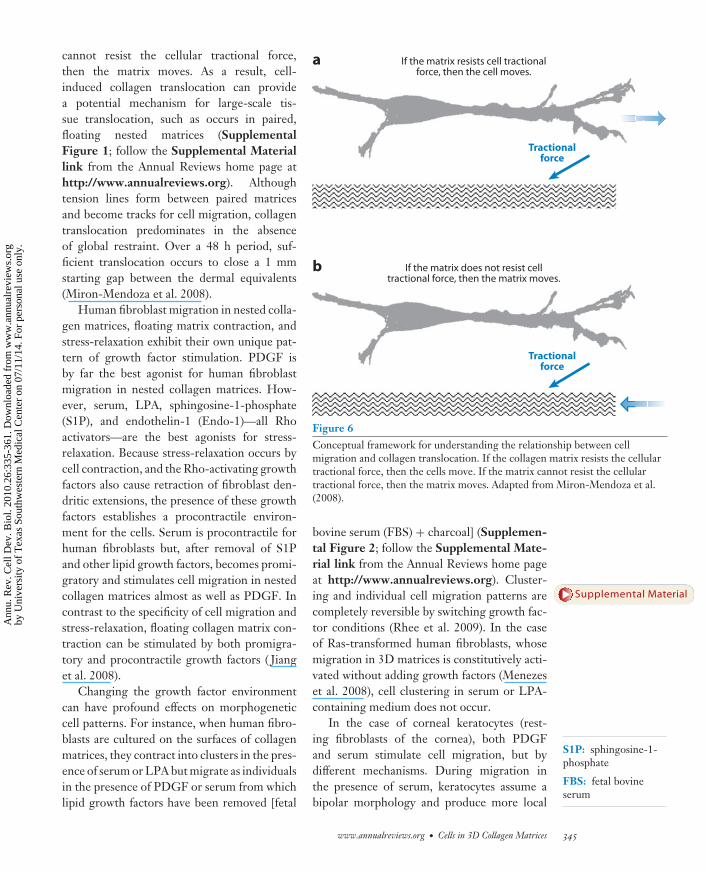
Figure 6 offers a conceptual framework forunderstanding the relationship between cellmigration and collagen translocation. If thecollagen matrix resists the cellular tractionalforce, then the cells move. If the matrix
344 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
S1P: sphingosine-1-phosphate
FBS: fetal bovineserum
cannot resist the cellular tractional force,then the matrix moves. As a result, cell-induced collagen translocation can providea potential mechanism for large-scale tis-sue translocation, such as occurs in paired,floating nested matrices (SupplementalFigure 1; follow the Supplemental Materiallink from the Annual Reviews home page athttp://www.annualreviews.org). Althoughtension lines form between paired matricesand become tracks for cell migration, collagentranslocation predominates in the absenceof global restraint. Over a 48 h period, suf-ficient translocation occurs to close a 1 mmstarting gap between the dermal equivalents(Miron-Mendoza et al. 2008).
Human fibroblast migration in nested colla-gen matrices, floating matrix contraction, andstress-relaxation exhibit their own unique pat-tern of growth factor stimulation. PDGF isby far the best agonist for human fibroblastmigration in nested collagen matrices. How-ever, serum, LPA, sphingosine-1-phosphate(S1P), and endothelin-1 (Endo-1)—all Rhoactivators—are the best agonists for stress-relaxation. Because stress-relaxation occurs bycell contraction, and the Rho-activating growthfactors also cause retraction of fibroblast den-dritic extensions, the presence of these growthfactors establishes a procontractile environ-ment for the cells. Serum is procontractile forhuman fibroblasts but, after removal of S1Pand other lipid growth factors, becomes promi-gratory and stimulates cell migration in nestedcollagen matrices almost as well as PDGF. Incontrast to the specificity of cell migration andstress-relaxation, floating collagen matrix con-traction can be stimulated by both promigra-tory and procontractile growth factors ( Jianget al. 2008).
Changing the growth factor environmentcan have profound effects on morphogeneticcell patterns. For instance, when human fibro-blasts are cultured on the surfaces of collagenmatrices, they contract into clusters in the pres-ence of serum or LPA but migrate as individualsin the presence of PDGF or serum from whichlipid growth factors have been removed [fetal
a If the matrix resists cell tractionalforce, then the cell moves.
b If the matrix does not resist celltractional force, then the matrix moves.
Tractionalforce
Tractionalforce
Figure 6Conceptual framework for understanding the relationship between cellmigration and collagen translocation. If the collagen matrix resists the cellulartractional force, then the cells move. If the matrix cannot resist the cellulartractional force, then the matrix moves. Adapted from Miron-Mendoza et al.(2008).
bovine serum (FBS) + charcoal] (Supplemen-tal Figure 2; follow the Supplemental Mate-rial link from the Annual Reviews home pageat http://www.annualreviews.org). Cluster-ing and individual cell migration patterns arecompletely reversible by switching growth fac-tor conditions (Rhee et al. 2009). In the caseof Ras-transformed human fibroblasts, whosemigration in 3D matrices is constitutively acti-vated without adding growth factors (Menezeset al. 2008), cell clustering in serum or LPA-containing medium does not occur.
In the case of corneal keratocytes (rest-ing fibroblasts of the cornea), both PDGFand serum stimulate cell migration, but bydifferent mechanisms. During migration inthe presence of serum, keratocytes assume abipolar morphology and produce more local
www.annualreviews.org • Cells in 3D Collagen Matrices 345
Supplemental Material
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
a 10% FBS
b PDGF 50 ng ml–1
40 μm
40 μm
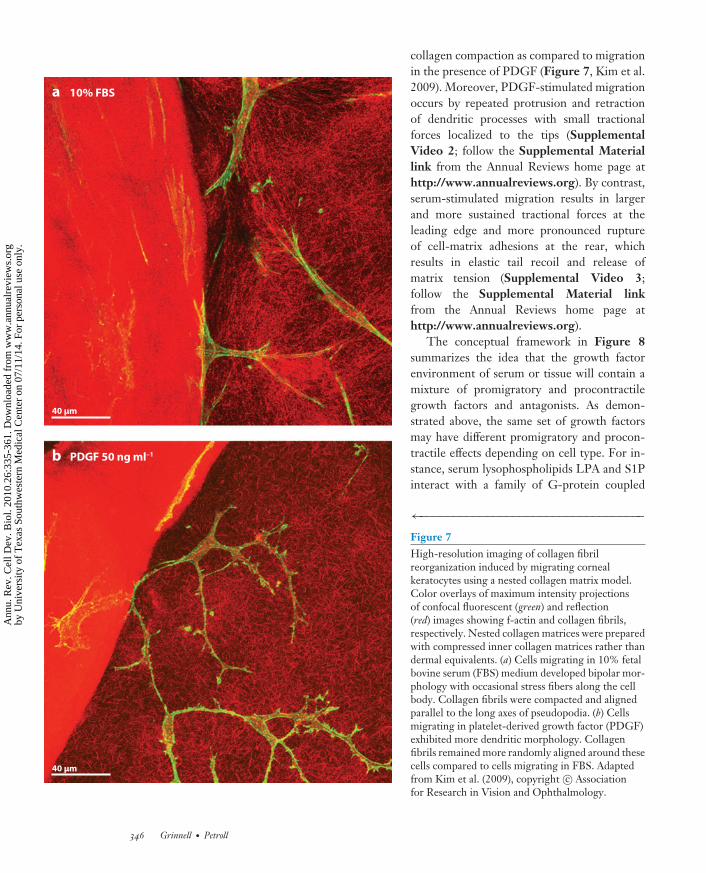
collagen compaction as compared to migrationin the presence of PDGF (Figure 7, Kim et al.2009). Moreover, PDGF-stimulated migrationoccurs by repeated protrusion and retractionof dendritic processes with small tractionalforces localized to the tips (SupplementalVideo 2; follow the Supplemental Materiallink from the Annual Reviews home page athttp://www.annualreviews.org). By contrast,serum-stimulated migration results in largerand more sustained tractional forces at theleading edge and more pronounced ruptureof cell-matrix adhesions at the rear, whichresults in elastic tail recoil and release ofmatrix tension (Supplemental Video 3;follow the Supplemental Material linkfrom the Annual Reviews home page athttp://www.annualreviews.org).
The conceptual framework in Figure 8summarizes the idea that the growth factorenvironment of serum or tissue will contain amixture of promigratory and procontractilegrowth factors and antagonists. As demon-strated above, the same set of growth factorsmay have different promigratory and procon-tractile effects depending on cell type. For in-stance, serum lysophospholipids LPA and S1Pinteract with a family of G-protein coupled
←−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−−Figure 7High-resolution imaging of collagen fibrilreorganization induced by migrating cornealkeratocytes using a nested collagen matrix model.Color overlays of maximum intensity projectionsof confocal fluorescent (green) and reflection(red) images showing f-actin and collagen fibrils,respectively. Nested collagen matrices were preparedwith compressed inner collagen matrices rather thandermal equivalents. (a) Cells migrating in 10% fetalbovine serum (FBS) medium developed bipolar mor-phology with occasional stress fibers along the cellbody. Collagen fibrils were compacted and alignedparallel to the long axes of pseudopodia. (b) Cellsmigrating in platelet-derived growth factor (PDGF)exhibited more dendritic morphology. Collagenfibrils remained more randomly aligned around thesecells compared to cells migrating in FBS. Adaptedfrom Kim et al. (2009), copyright c© Associationfor Research in Vision and Ophthalmology.
346 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
receptors that various cell types express dif-ferentially. Depending on receptor expression,S1P potentially can activate cell contraction orcell migration signaling pathways. The S1P1 re-ceptor links to Rac activation and cell migra-tion, whereas the S1P2 receptor links to Rhoactivation, Rac inhibition, and inhibition of cellmigration (Sugimoto et al. 2003).
Ironically, most research on mechanismsof cell migration have been carried out inserum-containing medium. For many celltypes, serum will establish a procontractilerather than promigratory environment, i.e.,Rho rather than Rac activation (Ridley &Hall 1992). Because the presence of a pro-contractile or promigratory environment canalter the balance between the cellular forcesof protrusion, contraction, and adhesion, thegrowth factor environment has the potential toinfluence basic mechanisms and patterns of cellmotility, especially in the tissue-like 3D matrixenvironment, where cell tractional force andmatrix stiffness are closely balanced. IncreasingRho activation will result in an increase incell membrane tension and tensegrity forces(Ingber 1997, Sheetz & Dai 1996) tendingto impede cell spreading in 3D matrices(Grinnell 2003, Rhee et al. 2010). Conversely,increasing Rac activation stimulates formationof dendritic extensions (Grinnell 2003) andincreases random fibroblast migration in 3Dmatrices (Pankov et al. 2005). Cell durotaxis,which has been demonstrated for cells mi-grating in serum-containing media on 2Dsurfaces (Lo et al. 2000) and inside 3D matrices(Hadjipanayi et al. 2009), may turn out to beunique to the procontractile growth factorenvironment.
HIGH-RESOLUTION IMAGINGOF LOCAL MATRIXREMODELING
Imaging collagen matrices that contain micro-spheres has been useful for learning about thelocal and global movements of collagen in re-sponse to cell motile activity (Miron-Mendozaet al. 2008, Roy et al. 1997, Tamariz & Grinnell
S1P PDGF LPA
Extensionand migration
Retractionand contraction
Serum/tissue growth factor environment
Figure 8Conceptual framework for understanding promigratory versus procontractilegrowth factor environments. The growth factor environment of serum or tissuewill contain a mixture of promigratory and procontractile growth factors andantagonists. The same set of growth factors may have different promigratoryand procontractile effects depending on cell type; effects on dermal fibroblastsare depicted here. LPA, lysophosphatidic acid; PDGF, platelet-derived growthfactor; S1P, sphingosine-1-phosphate. Adapted from Jiang et al. (2008).
2002), but such experiments do not providedetailed information on the structural reorga-nization of the matrix. Advances in imagingtechnology combined with live cell labelingtechniques increasingly has made such studiespossible. The three most commonly usedmethods are differential interference contrast(DIC), confocal reflection microscopy, andconfocal microscopy combined with secondharmonic generation imaging.
Bard and Hay introduced DIC imagingto visualize corneal embryonic fibroblastmigration in situ and in collagen matrices(Bard & Hay 1975). This approach alloweddirect visualization of the local collagen fibrilorganization surrounding the cells. Improvedoptics and digital imaging now allow theseprocesses to be studied with high temporaland spatial resolution (Supplemental Video4; follow the Supplemental Material linkfrom the Annual Reviews home page athttp://www.annualreviews.org) (Petroll &Ma 2003). Tracking landmarks on collagenfibrils in the DIC images provides an accuratemeasure of matrix deformation without theneed for embedded microspheres (Vanni et al.2003).
www.annualreviews.org • Cells in 3D Collagen Matrices 347
Supplemental Material
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
DIC analysis of green fluorescent protein(GFP)-zyxin expressing fibroblasts spread-ing on or within collagen matrices showsthat new focal adhesions form at the tips ofpseudopodia while existing adhesions movebackward. Simultaneously, adhesions at thebase of pseudopodia move toward those at thetip, resulting in a region of contractile-likeshortening and matrix compression (Sup-plemental Figure 3, Supplemental Video5; follow the Supplemental Material linkfrom the Annual Reviews home page athttp://www.annualreviews.org) (Petroll &Ma 2003). These findings show that pseu-dopodia have distinct mechanical zones atthe leading edge and base (Petroll & Ma2003, Vanni et al. 2003). Similar observationsregarding different mechanical zones havebeen made using traction force microscopyon cells on 2D surfaces (Munevar et al. 2001).However, whereas groups of adjacent focaladhesions form within a cell lamellipodium on2D surfaces, in 3D matrices focal adhesionstend to form and treadmill in a linear patternalong individual collagen fibrils. The alignmentbetween pseudopodial focal adhesions andcollagen fibrils helps account for the contactguidance of cell migration along matrix fibrilsoriginally observed by Weiss (1959) and subse-quently confirmed by many other laboratories.
Taking advantage of refractive index differ-ences within the matrix, Friedl et al. introducedconfocal reflection imaging to generate high-contrast optical section images of matrixfibrils (Friedl et al. 1998). Time-lapse confocalreflection imaging has been particularly usefulin permitting direct observation of the sub-cellular mechanics underlying amoeboid andmesenchymal modes of migration through 3Dmatrices (Wolf et al. 2003). Using this method,the organization of collagen matrix remodelingaround single cells and between adjacent cellscan be visualized at high resolution (Supple-mental Figure 4; follow the SupplementalMaterial link from the Annual Reviews homepage at http://www.annualreviews.org) (Kimet al. 2006, Petroll et al. 2004a). An alternativeto confocal reflection microscopy that permits
both deeper penetration and higher-resolutionimaging of the matrix is second harmonic gen-eration imaging using two-photon microscopy(Friedl 2004, Yeh et al. 2004). With both ofthese techniques, signal intensity depends onthe orientation of the collagen fibrils beingimaged ( Jawerth et al. 2009).
Confocal imaging provides more detailedinformation about 3D collagen matrix reorga-nization than does DIC imaging. Nevertheless,longer acquisition times and potential photo-toxicity can limit the usefulness of this technol-ogy for time-lapse studies of very rapid eventsin living cells. Thus, wide-field fluorescent andDIC imaging are generally used to study rapidchanges in protein organization and matrix de-formation that occur within minutes, whereasconfocal and multiphoton imaging are used toassess more gradual changes in cell-inducedcompaction and alignment of collagen fibrils.
REGULATION OF LOCALMATRIX REMODELING BYSMALL G PROTEINS ANDMECHANICAL STIMULI
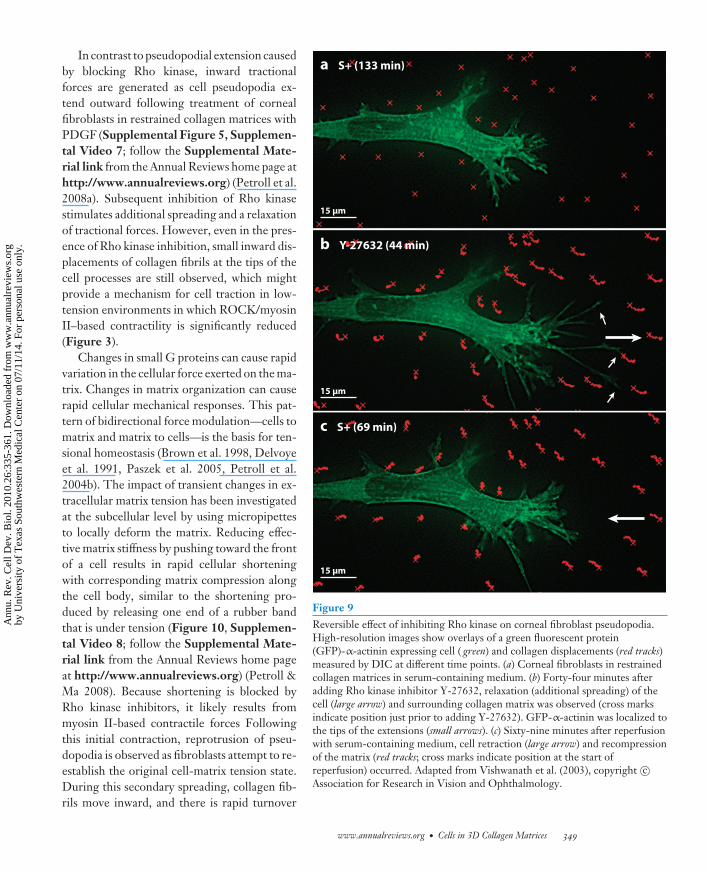
As described in previous sections, the Rho-family GTPases play a central role in the regula-tion of cell motile activity in collagen matrices.For instance, Rho activation causes dendriticcell extensions to retract; Rac activation causesthe extensions to protrude further (Figure 4).High-resolution imaging has been used tostudy the effects of Rho and Rac activation oncell motile activity in relationship to collagenremodeling. When corneal fibroblasts on topof or within restrained 3D collagen matrices arestimulated with LPA or serum, local collagencompression accompanies retraction of cellpseudopodia (Petroll et al. 2008b). Conversely,inhibiting Rho kinase results in extension of cellpseudopodia and reversible relaxation of cell-induced tension on the matrix (Figure 9, Sup-plemental Video 6; follow the SupplementalMaterial link from the Annual Reviews homepage at http://www.annualreviews.org)(Vishwanath et al. 2003).
348 Grinnell · Petroll
Supplemental Material
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
In contrast to pseudopodial extension causedby blocking Rho kinase, inward tractionalforces are generated as cell pseudopodia ex-tend outward following treatment of cornealfibroblasts in restrained collagen matrices withPDGF (Supplemental Figure 5, Supplemen-tal Video 7; follow the Supplemental Mate-rial link from the Annual Reviews home page athttp://www.annualreviews.org) (Petroll et al.2008a). Subsequent inhibition of Rho kinasestimulates additional spreading and a relaxationof tractional forces. However, even in the pres-ence of Rho kinase inhibition, small inward dis-placements of collagen fibrils at the tips of thecell processes are still observed, which mightprovide a mechanism for cell traction in low-tension environments in which ROCK/myosinII–based contractility is significantly reduced(Figure 3).
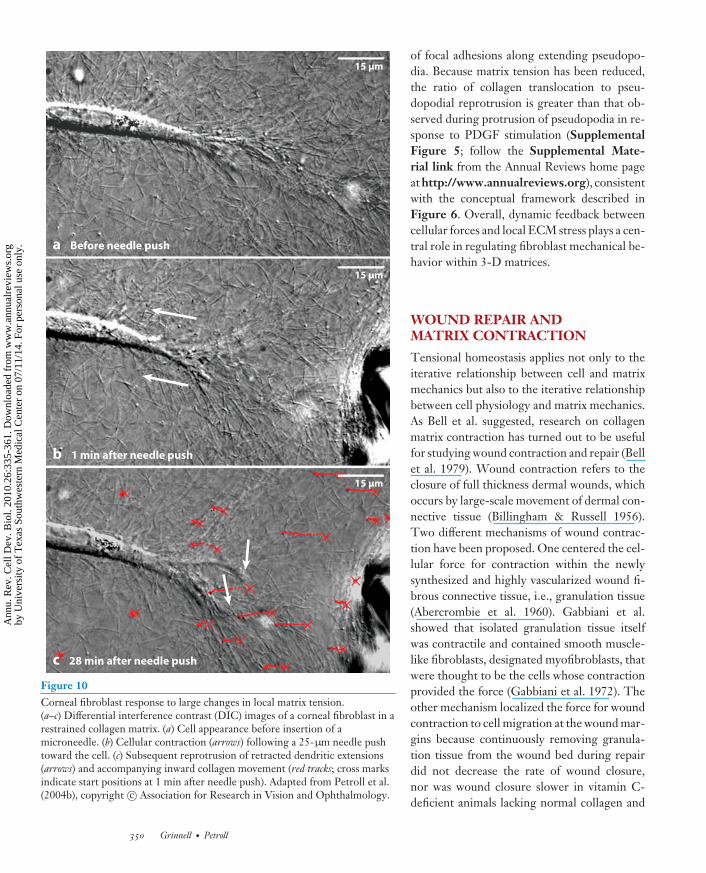
Changes in small G proteins can cause rapidvariation in the cellular force exerted on the ma-trix. Changes in matrix organization can causerapid cellular mechanical responses. This pat-tern of bidirectional force modulation—cells tomatrix and matrix to cells—is the basis for ten-sional homeostasis (Brown et al. 1998, Delvoyeet al. 1991, Paszek et al. 2005, Petroll et al.2004b). The impact of transient changes in ex-tracellular matrix tension has been investigatedat the subcellular level by using micropipettesto locally deform the matrix. Reducing effec-tive matrix stiffness by pushing toward the frontof a cell results in rapid cellular shorteningwith corresponding matrix compression alongthe cell body, similar to the shortening pro-duced by releasing one end of a rubber bandthat is under tension (Figure 10, Supplemen-tal Video 8; follow the Supplemental Mate-rial link from the Annual Reviews home pageat http://www.annualreviews.org) (Petroll &Ma 2008). Because shortening is blocked byRho kinase inhibitors, it likely results frommyosin II-based contractile forces Followingthis initial contraction, reprotrusion of pseu-dopodia is observed as fibroblasts attempt to re-establish the original cell-matrix tension state.During this secondary spreading, collagen fib-rils move inward, and there is rapid turnover
a S+ (133 min)
15 μm
b Y-27632 (44 min)
c S+ (69 min)
15 μm
15 μm
Figure 9Reversible effect of inhibiting Rho kinase on corneal fibroblast pseudopodia.High-resolution images show overlays of a green fluorescent protein(GFP)-α-actinin expressing cell ( green) and collagen displacements (red tracks)measured by DIC at different time points. (a) Corneal fibroblasts in restrainedcollagen matrices in serum-containing medium. (b) Forty-four minutes afteradding Rho kinase inhibitor Y-27632, relaxation (additional spreading) of thecell (large arrow) and surrounding collagen matrix was observed (cross marksindicate position just prior to adding Y-27632). GFP-α-actinin was localized tothe tips of the extensions (small arrows). (c) Sixty-nine minutes after reperfusionwith serum-containing medium, cell retraction (large arrow) and recompressionof the matrix (red tracks; cross marks indicate position at the start ofreperfusion) occurred. Adapted from Vishwanath et al. (2003), copyright c©Association for Research in Vision and Ophthalmology.
www.annualreviews.org • Cells in 3D Collagen Matrices 349
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
c 28 min after needle push
b 1 min after needle push
a Before needle push
15 μm
15 μm
15 μm
Figure 10Corneal fibroblast response to large changes in local matrix tension.(a–c) Differential interference contrast (DIC) images of a corneal fibroblast in arestrained collagen matrix. (a) Cell appearance before insertion of amicroneedle. (b) Cellular contraction (arrows) following a 25-μm needle pushtoward the cell. (c) Subsequent reprotrusion of retracted dendritic extensions(arrows) and accompanying inward collagen movement (red tracks; cross marksindicate start positions at 1 min after needle push). Adapted from Petroll et al.(2004b), copyright c© Association for Research in Vision and Ophthalmology.
of focal adhesions along extending pseudopo-dia. Because matrix tension has been reduced,the ratio of collagen translocation to pseu-dopodial reprotrusion is greater than that ob-served during protrusion of pseudopodia in re-sponse to PDGF stimulation (SupplementalFigure 5; follow the Supplemental Mate-rial link from the Annual Reviews home pageat http://www.annualreviews.org), consistentwith the conceptual framework described inFigure 6. Overall, dynamic feedback betweencellular forces and local ECM stress plays a cen-tral role in regulating fibroblast mechanical be-havior within 3-D matrices.
WOUND REPAIR ANDMATRIX CONTRACTION
Tensional homeostasis applies not only to theiterative relationship between cell and matrixmechanics but also to the iterative relationshipbetween cell physiology and matrix mechanics.As Bell et al. suggested, research on collagenmatrix contraction has turned out to be usefulfor studying wound contraction and repair (Bellet al. 1979). Wound contraction refers to theclosure of full thickness dermal wounds, whichoccurs by large-scale movement of dermal con-nective tissue (Billingham & Russell 1956).Two different mechanisms of wound contrac-tion have been proposed. One centered the cel-lular force for contraction within the newlysynthesized and highly vascularized wound fi-brous connective tissue, i.e., granulation tissue(Abercrombie et al. 1960). Gabbiani et al.showed that isolated granulation tissue itselfwas contractile and contained smooth muscle-like fibroblasts, designated myofibroblasts, thatwere thought to be the cells whose contractionprovided the force (Gabbiani et al. 1972). Theother mechanism localized the force for woundcontraction to cell migration at the wound mar-gins because continuously removing granula-tion tissue from the wound bed during repairdid not decrease the rate of wound closure,nor was wound closure slower in vitamin C-deficient animals lacking normal collagen and
350 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.
CB26CH14-Grinnell ARI 3 September 2010 19:34
granulation tissue synthesis (reviewed in Gross1996).
The discovery of α-smooth muscle actinexpression as a biochemical marker for my-ofibroblast differentiation made it possible tocharacterize more precisely the appearanceand function of these cells (Skalli et al. 1986).Myofibroblasts sometimes appear late ingranulation tissue relative to wound closure(Berry et al. 1998, Doillon et al. 1987, Nedelecet al. 2000), and they are not required forfetal wounds to close (Martin 2007). Also, inthe tenascin-C knockout mouse repair occursnormally even though appearance of myofi-broblasts is delayed (Tamaoki et al. 2005).Therefore, at least under some circumstances,wound fibroblasts are sufficient for woundcontraction and closure. However, in mechan-ically restrained wounds that exhibit delayedclosure, the myofibroblast population increasesmarkedly (Hinz et al. 2001, Squier 1981).
Figure 11 shows the interrelationshipsbetween the above findings and collagenmatrix contraction models. At low mechanicalresistance (floating collagen matrices), cellmigration can provide the force for large-scaletissue movement, as observed with pairednested collagen matrices (SupplementalFigure 1; follow the Supplemental Materiallink from the Annual Reviews home page athttp://www.annualreviews.org). As mechan-ical resistance increases (restrained collagenmatrices), a high cell-matrix tension statedevelops and cell contraction becomes themechanism for force generation (Figure 3).In addition to mechanical resistance, thechange from migration to contraction requiresswitching from promigratory to procontractilegrowth factor environments (Figures 3 and8). If differentiation signals such as TGFβ
are present in the growth factor environment,then fibroblasts in restrained collagen matriceswill differentiate into myofibroblasts and exertgreater contractile force (Hinz 2009, Tomaseket al. 2002). The contractile force exerted byfibroblasts and myofibroblasts in combinationwith matrix biosynthesis ultimately resultsin wound closure and dissipation of tissue
Fibroblast
Floating
Restra
ined matri
x
Cell type causing wound contraction
Mec
hani
sm o
f wou
nd c
ontr
acti
onM
igra
tion
Cont
ract
ion
Myofibroblast
Figure 11Conceptual framework relating collagen matrixcontraction and wound contraction. At lowmechanical resistance (floating collagen matrices),cell migration can provide the force for large-scaletissue movement. As mechanical resistance increases(restrained collagen matrices), cell contractionbecomes the mechanism for force generation. Ifdifferentiation signals are present in the growthfactor environment, then fibroblasts experiencinghigh mechanical resistance differentiate intomyofibroblasts. Adapted from Grinnell (2003).
tension (stress-relaxation). Continued activityof myofibroblasts after wound closure occursunder pathological wound conditions such ascontracture and hypertrophic scar (Abrahamet al. 2007, Tomasek et al. 2002, Wynn 2008).
Myofibroblasts appear in tissue in contextsother than wound repair. For example, thesecells play a central role in fibrotic disease(Abraham et al. 2007, Desmouliere et al. 2003,Wynn 2008) and tumor progression (Butcheret al. 2009, Desmouliere et al. 2004, Weigelt& Bissell 2008). Most evidence suggeststhat fibroblasts differentiate into myofibro-blasts only when they are stimulated withappropriate growth factors when in a highcell-matrix tension state (e.g., in restrainedbut not floating collagen matrices) (Hinz2009, Tomasek et al. 2002). Interstitial fluidflow through collagen matrices containingfibroblasts also provides a myofibroblast-activating signal by increasing matrix tension
www.annualreviews.org • Cells in 3D Collagen Matrices 351
Supplemental Material
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
FAK: focal adhesionkinase
PI3 kinase:phosphatidylinositol3-kinase
and autocrine production of TGFβ (Ng et al.2005). However, the ability of endothelin-1 topromote differentiation of fibroblasts into my-ofibroblasts in floating collagen matrices showsthat with some growth factors, high cell-matrixtension may not be required for myofibroblastdifferentiation (Shi-Wen et al. 2004).
Studies on fibroblast proliferation andbiosynthesis in collagen matrices have con-tributed to the mechanoregulated model ofwound repair. Figure 12 summarizes thesefindings. In floating collagen matrices, fibro-blasts are turned off and exhibit little ma-trix biosynthesis or proliferation. In restrainedmatrices, cells are turned on. They prolifer-ate and synthesize and accumulate new ex-tracellular matrix (Eckes et al. 2006, Grinnell2003). After stress-relaxation, cells rapidly be-come quiescent as a result of disrupted signal-ing to cell cycle regulatory proteins (Fringer &Grinnell 2001). The 3D matrix environmentuniquely regulates focal adhesion kinase activa-tion (phospho FAK397) (Cukierman et al. 2001,Wei et al. 2008), and decreased FAK activa-tion probably accounts for cell quiescence infloating collagen matrices (Klein et al. 2009).Also, a subpopulation of fibroblasts in floating
Wounding
Resting fibroblastCell proliferation OFF
Matrix biosynthesis OFF
(Floating collagenmatrix)
Activated fibroblastCell proliferation ON
Matrix biosynthesis ON
(Restrained collagenmatrix)
Low cell-matrix tension High cell-matrix tension
Cell proliferation and biosynthesisCell migration and contraction
(Stress-relaxation)
Figure 12Mechanoregulated model of wound contraction. Inresting tissue, fibroblasts are turned off and exhibitlittle matrix biosynthesis or proliferation. Ingranulation tissue, fibroblasts are activated. Overtime, the combination of cell proliferation,biosynthesis, migration, and contraction returns thetissue from an activated to resting state. Adaptedfrom Grinnell (2003).
collagen matrices becomes apoptotic (Grinnellet al. 1999). Apoptosis results from decreasedphosphatidylinositol 3-kinase (PI3 kinase) sig-naling at least in part as a result of phosphataseand tensin homolog (PTEN) activation (Nhoet al. 2006). Increased expression of cell sur-face tissue transglutaminase may protect hyper-trophic scar cells from undergoing apoptosis infloating collagen matrices (Linge et al. 2005).
In relationship to wound repair, floatingcollagen matrices resemble normal tissue, andrestrained collagen matrices resemble granula-tion tissue. Fibroblasts in normal tissue are in alow cell-matrix tension state—the fibrocytes ofclassical histology. When these cells develophigh cell-matrix tension as part of tissue homeo-stasis or in response to mechanical wounding,they are activated to proliferate and synthesizenew connective tissue matrix. After dissipationof the high cell-matrix tension state throughthe combination of matrix biosynthesis andcontraction, fibroblasts turn off and granula-tion tissue regresses. External pressure therapy(Costa et al. 1999) or closing experimentalwounds by skin flapping releases internaltension and causes granulation tissue to regressrapidly through cell quiescence and apoptosis(Carlson et al. 2002, Darby et al. 2002). Increas-ing tissue tension by external skin splinting(Hinz et al. 2001) or injection of dermal fillers(Wang et al. 2007) increases matrix biosynthe-sis. In addition, increasing tissue tension resultsin decreased myofibroblast apoptosis and for-mation of hypertropic scars (Aarabi et al. 2007).Finally, mice with PTEN-null fibroblasts intheir mammary glands show increased matrixremodeling and formation of epithelial tumors(Trimboli et al. 2009), perhaps because of thefailure of myofibroblasts to undergo apoptosis.
CONCLUSIONS ANDFUTURE DIRECTIONS
Almost one hundred years have passed sincepublication of the Lewis and Lewis article dis-cussed at the beginning of this review. Sincethen, research on cells in 2D culture has pro-vided remarkable insights into cell structure
352 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
and function. Weiss’s hierarchical approach ofstudying cells in 3D matrices lagged behind butnow is beginning to catch up. Advances in imag-ing technology combined with live and fixed celllabeling techniques increasingly makes possiblehigh-resolution analysis of cell-matrix interac-tions at the global and subcellular levels.
In this review we focus on fibroblasts inter-acting with 3D collagen matrices, which exhibitmechanical and structural features that resem-ble fibrous connective tissue. Fibroblasts are thecell type primarily responsible for fibrous con-nective tissue biosynthesis and organization. Fi-broblast tractional force and matrix stiffness areclosely balanced in collagen matrices. Conse-quently, cells show a high degree of plasticityincluding the ability to develop dendritic mor-phology. Fibroblasts in 3D collagen matricesare capable of rapidly protruding and retract-ing cell extensions in response to growth fac-tor stimulation or physical manipulation of thematrix. The small G proteins Rac and Rho areparticularly important regulators of protrusionand retraction. Movement of cell extensions cancause inward (toward the cell body) movementof collagen fibrils not only as the extensions re-tract but also as they protrude.
Cell tractional force can remodel andcompact matrix fibrils into stably reorganizedstructures. Similar to cell adhesion, spreading,and migration, collagen matrix contraction hasturned out to be a highly conserved featureof cell motile behavior exhibited not only byfibroblasts but also by many other cell types.Global factors such as whether the matrix isfloating or restrained as well as local factors suchas matrix stiffness affect the cell-matrix tensionstate and influence cell signaling pathways,cytoskeletal function, and motor mechanismsrequired for contraction. Cells interactingwith collagen matrices form an interactivemechanical system. Tensional homeostasisdescribes this mechanical reciprocity.
The porosity of the matrix makes it pos-sible for cells to penetrate and move throughthe matrix and not just over its surface. Thegeometry of the matrix establishes limited do-mains of cell adhesion along matrix fibrils.
Pseudopodial adhesions in 3D matrices tend toform and treadmill in a linear pattern along in-dividual collagen fibrils, which helps accountfor contact guidance. However, in a matrix lack-ing oriented fibrils, fibroblasts can interact withfibrils oriented both parallel and perpendicularto the direction of cell pseudopodia. At a globallevel, cell migration and collagen translocationcan couple together to produce large-scale tis-sue movement.
We also emphasize the importance of dis-tinguishing promigratory and procontractilegrowth factor environments, which can alterthe balance between the cellular forces of pro-trusion, contraction, and adhesion. These dif-ferences will influence basic mechanisms andpatterns of cell motility, especially in the tissue-like 3D matrix environment, where cell trac-tional force and matrix stiffness are closelybalanced.
Finally, we discuss the relationship betweencollagen matrix contraction models and woundrepair. We suggest that the relative importanceof fibroblasts and myofibroblasts for woundcontraction depends on tissue resistance. In themechanoregulated model of wound repair, cellphysiology links with cell-matrix tension state.
What about the future? For tissue cells on2D surfaces, cell motile activity has been de-scribed in terms of a balance between cellularforces of protrusion, contraction, and adhesion.To successfully model cell mechanical behav-ior in 3D matrices, a new generation of engi-neered biocompatible matrices will be requiredin which matrix porosity, stiffness, and adhesionligand domains are defined and can be varied in-dependently so that their individual effects oncell behavior can be determined.
New types of engineered matrices also areneeded to permit more detailed investigationsof the influence of matrix fibril organizationon cell motility and mechanics. Most currentmethods for preparing 3D fibrillar matricesresult in matrices in which fibrils lack anyparticular organization or become aligned uni-axially. However, tissues exhibit more complexarchitectures such as collagen fibril bundlesand orthogonally arranged collagen sheets.
www.annualreviews.org • Cells in 3D Collagen Matrices 353
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Incorporation of these features into 3D matrixmodels could provide unique insights intocell-matrix interactions and might help clarifythe role of proteoglycans and other matrixcomponents in modulating cell behavior in thetissue environment.
Much remains to be learned about thelow cell-matrix tension environment. Relativelylittle is known about the dendritic fibroblast.Fibroblasts and other cell types appear capableof generating myosin II–independent tractionalforces in the low cell-matrix tension environ-
ment. Are other myosins responsible, or does adifferent type of molecular motor produce theseforces?
Lastly, development of new probes andimaging techniques to allow direct assessmentof mechanical signaling events in living cellsshould continue to improve our understandingof how cell motility in 3D matrices is regulated.Application of multiphoton imaging techniquesand novel experimental animal models will in-creasingly allow many of these processes to bestudied in vivo.
DISCLOSURE STATEMENT
The authors are not aware of any affiliations, memberships, funding, or financial holdings thatmight be perceived as affecting the objectivity of this review.
ACKNOWLEDGMENTS
Research in our laboratories has been supported by grants from the National Institutes of Health,GM031321 to F.G. and EY013322 to W.M.P., and from Research to Prevent Blindness, Inc. toW.M.P.
LITERATURE CITED
Aarabi S, Bhatt KA, Shi Y, Paterno J, Chang EI, et al. 2007. Mechanical load initiates hypertrophic scarformation through decreased cellular apoptosis. FASEB J. 21:3250–61
Abercrombie M, James DW, Newcombe JF. 1960. Wound contraction in rabbit skin, studied by splinting thewound margins. J. Anat. Lond. 94:170–82
Abraham DJ, Eckes B, Rajkumar V, Krieg T. 2007. New developments in fibroblast and myofibroblast biology:implications for fibrosis and scleroderma. Curr. Rheumatol. Rep. 9:136–43
Ahlfors JE, Billiar KL. 2007. Biomechanical and biochemical characteristics of a human fibroblast-producedand remodeled matrix. Biomaterials 28:2183–91
Bard JBL, Hay ED. 1975. The behavior of fibroblasts from the developing avian cornea: morphology andmovement in situ and in vitro. J. Cell Biol. 67:400–18
Barocas VH, Moon AG, Tranquillo RT. 1995. The fibroblast-populated collagen microsphere assay of celltraction force—Part 2: Measurement of the cell traction parameter. J. Biomech. Eng. 117:161–70
Barocas VH, Tranquillo RT. 1997. An anisotropic biphasic theory of tissue-equivalent mechanics: the interplayamong cell traction, fibrillar network deformation, fibril alignment, and cell contact guidance. J. Biomech.Eng. 119:137–45
Bell E, Ehrlich HP, Buttle DJ, Nakatsuji T. 1981. Living tissue formed in vitro and accepted as skin-equivalenttissue of full thickness. Science 211:1052–54
Bell E, Ivarsson B, Merrill C. 1979. Production of a tissue-like structure by contraction of collagen lattices byhuman fibroblasts of different proliferative potential in vitro. Proc. Natl. Acad. Sci. USA 76:1274–78
Bellows CG, Melcher AH, Aubin JE. 1982. Association between tension and orientation of periodontal liga-ment fibroblasts and exogenous collagen fibers in collagen gels in vitro. J. Cell Sci. 58:125–38
Beningo KA, Dembo M, Wang YL. 2004. Responses of fibroblasts to anchorage of dorsal extracellular matrixreceptors. Proc. Natl. Acad. Sci. USA 101:18024–29
Berry DP, Harding KG, Stanton MR, Jasani B, Ehrlich HP. 1998. Human wound contraction: collagenorganization, fibroblasts, and myofibroblasts. Plast. Reconstr. Surg. 102:124–31; discussion 32–34
354 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Billingham RE, Russell PS. 1956. Studies on wound healing, with special reference to the phenomenon ofcontracture in experimental wounds in rabbits’ skin. Ann. Surg. 144:961–81
Bornstein MB. 1958. Reconstituted rat-tail collagen used as substrate for tissue cultures on coverslips inmaximow slides and roller tubes. Lab. Investig. 7:134–37
Brown RA, Prajapati R, McGrouther DA, Yannas IV, Eastwood M. 1998. Tensional homeostasis in dermalfibroblasts: mechanical responses to mechanical loading in three-dimensional substrates. J. Cell Physiol.175:323–32
Brown RA, Talas G, Porter RA, McGrouther DA, Eastwood M. 1996. Balanced mechanical forces and mi-crotubule contribution to fibroblast contraction. J. Cell Physiol. 169:439–47
Brown RA, Wiseman M, Chuo CB, Cheema U, Nazhat SN. 2005. Ultrarapid engineering of biomimeticmaterials and tissues: fabrication of nano- and microstructures by plastic compression. Adv. Funct. Mater.15:1762–70
Bullard KM, Mudgett J, Scheuenstuhl H, Hunt TK, Banda MJ. 1999. Stromelysin-1-deficient fibroblastsdisplay impaired contraction in vitro. J. Surg. Res. 84:31–34
Butcher DT, Alliston T, Weaver VM. 2009. A tense situation: forcing tumor progression. Nat. Rev. Cancer9:108–22
Carlson MA, Longaker MT, Thompson JS. 2002. Granulation tissue regression induced by musculocutaneousadvancement flap coverage. Surgery 131:332–37
Chandran PL, Barocas VH. 2006. Affine versus nonaffine fibril kinematics in collagen networks: theoreticalstudies of network behavior. J. Biomech. Eng. 128:259–70
Clark RA, Ghosh K, Tonnesen MG. 2007. Tissue engineering for cutaneous wounds. J. Investig. Dermatol.127:1018–29
Cooke ME, Sakai T, Mosher DF. 2000. Contraction of collagen matrices mediated by α2β1A and αvβ3integrins. J. Cell Sci. 113:2375–83
Costa AM, Peyrol S, Porto LC, Comparin JP, Foyatier JL, Desmouliere A. 1999. Mechanical forces inducescar remodeling. Study in nonpressure-treated versus pressure-treated hypertrophic scars. Am. J. Pathol.155:1671–79
Cukierman E, Pankov R, Stevens DR, Yamada KM. 2001. Taking cell-matrix adhesions to the third dimension.Science 294:1708–12
Darby IA, Bisucci T, Pittet B, Garbin S, Gabbiani G, Desmouliere A. 2002. Skin flap-induced regressionof granulation tissue correlates with reduced growth factor and increased metalloproteinase expression.J. Pathol. 197:117–27
Davis GE, Senger DR. 2008. Extracellular matrix mediates a molecular balance between vascular morpho-genesis and regression. Curr. Opin. Hematol. 15:197–203
Delvoye P, Wiliquet P, Leveque JL, Nusgens BV, Lapiere CM. 1991. Measurement of mechanical forcesgenerated by skin fibroblasts embedded in a three-dimensional collagen gel. J. Investig. Dermatol. 97:898–902
Deryugina EI, Bourdon MA, Reisfeld RA, Strongin A. 1998. Remodeling of collagen matrix by human tumorcells requires activation and cell surface association of matrix metalloproteinase-2. Cancer Res. 58:3743–50
Desmouliere A, Darby IA, Gabbiani G. 2003. Normal and pathologic soft tissue remodeling: role of themyofibroblast, with special emphasis on liver and kidney fibrosis. Lab. Investig. 83:1689–707
Desmouliere A, Guyot C, Gabbiani G. 2004. The stroma reaction myofibroblast: a key player in the controlof tumor cell behavior. Int. J. Dev. Biol. 48:509–17
Discher DE, Janmey P, Wang YL. 2005. Tissue cells feel and respond to the stiffness of their substrate. Science310:1139–43
Doillon CJ, Hembry RM, Ehrlich HP, Burke JF. 1987. Actin filaments in normal dermis and during woundhealing. Am. J. Pathol. 126:164–70
Doyle AD, Wang FW, Matsumoto K, Yamada KM. 2009. One-dimensional topography underlies three-dimensional fibrillar cell migration. J. Cell Biol. 184:481–90
Eckes B, Zweers MC, Zhang ZG, Hallinger R, Mauch C, et al. 2006. Mechanical tension and integrin α2β1regulate fibroblast functions. J. Investig. Dermatol. Symp. Proc. 11:66–72
Elsdale T, Bard J. 1972. Collagen substrata for studies on cell behavior. J. Cell Biol. 54:626–37
www.annualreviews.org • Cells in 3D Collagen Matrices 355
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Emerman JT, Pitelka DR. 1977. Maintenance and induction of morphological differentiation in dissociatedmammary epithelium on floating collagen membranes. In Vitro 13:316–28
Engler A, Bacakova L, Newman C, Hategan A, Griffin M, Discher D. 2004. Substrate compliance versusligand density in cell on gel responses. Biophys. J. 86:617–28
Fisher GJ, Varani J, Voorhees JJ. 2008. Looking older: fibroblast collapse and therapeutic implications. Arch.Dermatol. 144:666–72
Frey J, Chamson A, Raby N, Rattner A. 1995. Collagen bioassay by the contraction of fibroblast-populatedcollagen lattices. Biomaterials 16:139–43
Freyman TM, Yannas IV, Yokoo R, Gibson LJ. 2002. Fibroblast contractile force is independent of the stiffnesswhich resists the contraction. Exp. Cell Res. 272:153–62
Friedl P. 2004. Dynamic imaging of cellular interactions with extracellular matrix. Histochem. Cell Biol. 122:183–90
Friedl P, Brocker EB. 2000. The biology of cell locomotion within three-dimensional extracellular matrix.Cell. Mol. Life Sci. 57:41–64
Friedl P, Maaser K, Klein CE, Niggemann B, Krohne G, Zanker KS. 1997. Migration of highly aggressiveMV3 melanoma cells in 3-dimensional collagen lattices results in local matrix reorganization and sheddingof α2 and β1 integrins and CD44. Cancer Res. 57:2061–70
Friedl P, Wolf K. 2009. Proteolytic interstitial cell migration: a five-step process. Cancer Metastasis Rev.28:129–35
Friedl P, Zanker K, Brocker E-B. 1998. Cell migration strategies in 3-D extracellular matrix: differences inmorphology, cell matrix interactions, and integrin function. Microsc. Res. Tech. 43:369–78
Fringer J, Grinnell F. 2001. Fibroblast quiescence in floating or released collagen matrices: contribution ofthe ERK signaling pathway and actin cytoskeletal organization. J. Biol. Chem. 276:31047–52
Gabbiani G, Hirschel BJ, Ryan GB, Statkov PR, Majno G. 1972. Granulation tissue as a contractile organ. Astudy of structure and function. J. Exp. Med. 135:719–34
Gaggioli C, Hooper S, Hidalgo-Carcedo C, Grosse R, Marshall JF, et al. 2007. Fibroblast-led collectiveinvasion of carcinoma cells with differing roles for RhoGTPases in leading and following cells. Nat. CellBiol. 9:1392–400
Gentleman E, Nauman EA, Dee KC, Livesay GA. 2004. Short collagen fibers provide control of contractionand permeability in fibroblast-seeded collagen gels. Tissue Eng. 10:421–27
Georges PC, Janmey PA. 2005. Cell type-specific response to growth on soft materials. J. Appl. Physiol.98:1547–53
Germain L, Jean A, Auger FA, Garrel DR. 1994. Human wound healing fibroblasts have greater contractileproperties than dermal fibroblasts. J. Surg. Res. 57:268–73
Glaser BM, Cardin A, Biscoe B. 1987. Proliferative vitreoretinopathy. The mechanism of development ofvitreoretinal traction. Ophthalmology 94:327–32
Goldsmith EC, Hoffman A, Morales MO, Potts JD, Price RL, et al. 2004. Organization of fibroblasts in theheart. Dev. Dyn. 230:787–94
Griffith LG, Swartz MA. 2006. Capturing complex 3D tissue physiology in vitro. Nat. Rev. Mol. Cell Biol.7:211–24
Grinnell F. 1994. Fibroblasts, myofibroblasts, and wound contraction. J. Cell Biol. 124:401–4Grinnell F. 2000. Fibroblast-collagen-matrix contraction: growth-factor signaling and mechanical loading.
Trends Cell Biol. 10:362–65Grinnell F. 2003. Fibroblast biology in three-dimensional collagen matrices. Trends Cell Biol. 13:264–69Grinnell F, Ho CH. 2002. Transforming growth factor β stimulates fibroblast-collagen matrix contraction
by different mechanisms in mechanically loaded and unloaded matrices. Exp. Cell Res. 273:248–55Grinnell F, Ho CH, Tamariz E, Lee DJ, Skuta G. 2003. Dendritic fibroblasts in three dimensional collagen
matrices. Mol. Biol. Cell 14:384–95Grinnell F, Rocha LB, Iucu C, Rhee S, Jiang H. 2006. Nested collagen matrices: a new model to study
migration of human fibroblast populations in three dimensions. Exp. Cell Res. 312:86–94Grinnell F, Zhu M, Carlson MA, Abrams JM. 1999. Release of mechanical tension triggers apoptosis of human
fibroblasts in a model of regressing granulation tissue. Exp. Cell Res. 248:608–19
356 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Gross J. 1996. Getting to mammalian wound repair and amphibian limb regeneration: a mechanistic link inthe early events. Wound Repair Regen. 4:190–202
Guido S, Tranquillo RT. 1993. A methodology for the systematic and quantitative study of cell contactguidance in oriented collagen gels. Correlation of fibroblast orientation and gel birefringence. J. Cell Sci.105(Part 2):317–31
Hadjipanayi E, Mudera V, Brown RA. 2009. Guiding cell migration in 3D: a collagen matrix with gradeddirectional stiffness. Cell Motil. Cytoskelet. 66:121–29
Hall A. 1998. Rho GTPases and the actin cytoskeleton. Science 279:509–14Heuchel R, Berg A, Tallquist M, Ahlen K, Reed RK, et al. 1999. Platelet-derived growth factor β receptor
regulates interstitial fluid homeostasis through phosphatidylinositol-3′ kinase signaling. Proc. Natl. Acad.Sci. USA 96:11410–15
Hinz B. 2010. The myofibroblast: paradigm for a mechanically active cell. J. Biomech. 43:146–55Hinz B, Mastrangelo D, Iselin CE, Chaponnier C, Gabbiani G. 2001. Mechanical tension controls granulation
tissue contractile activity and myofibroblast differentiation. Am. J. Pathol. 159:1009–20Hocking DC, Sottile J, Langenbach KJ. 2000. Stimulation of integrin-mediated cell contractility by fibronectin
polymerization. J. Biol. Chem. 275:10673–82Howard EW, Benton R, Ahern-Moore J, Tomasek JJ. 1996. Cellular contraction of collagen lattices is inhibited
by nonenzymatic glycation. Exp. Cell Res. 228:132–37Ingber DE. 1997. Tensegrity: the architectural basis of cellular mechanotransduction. Annu. Rev. Physiol.
59:575–99Itoh Y, Seiki M. 2006. MT1-MMP: a potent modifier of pericellular microenvironment. J. Cell Physiol. 206:1–8Jawerth LM, Munster S, Vader DA, Fabry B, Weitz DA. 2009. A blind spot in confocal reflection microscopy:
the dependence of fiber brightness on fiber orientation in imaging biopolymer networks. Biophys. J.98:L1–3
Jiang H, Grinnell F. 2005. Cell-matrix entanglement and mechanical anchorage of fibroblasts in three-dimensional collagen matrices. Mol. Biol Cell. 16:5070–76
Jiang H, Rhee S, Ho C-H, Grinnell F. 2008. Distinguishing fibroblast promigratory and procontractile growthfactor environments in 3D collagen matrices. FASEB J. 22:2151–60
Johnson C, Galis ZS. 2004. Matrix metalloproteinase-2 and -9 differentially regulate smooth muscle cellmigration and cell-mediated collagen organization. Arterioscler. Thromb. Vasc. Biol. 24:54–60
Kim A, Lakshman N, Karamichos D, Petroll WM. 2010. Growth factor regulation of corneal keratocytedifferentiation and migration in compressed collagen matrices. Investig. Ophthalmol. Vis. Sci. 51:864–75
Kim A, Lakshman N, Petroll WM. 2006. Quantitative assessment of local collagen matrix remodeling in 3-Dculture: the role of Rho kinase. Exp. Cell Res. 312:3683–92
Klein CE, Dressel D, Steinmayer T, Mauch C, Eckes B, et al. 1991. Integrin α2β1 is upregulated in fi-broblasts and highly aggressive melanoma cells in three-dimensional collagen lattices and mediates thereorganization of collagen I fibrils. J. Cell Biol. 115:1427–36
Klein EA, Yin L, Kothapalli D, Castagnino P, Byfield FJ, et al. 2009. Cell-cycle control by physiological matrixelasticity and in vivo tissue stiffening. Curr. Biol. 19:1511–18
Kolodney MS, Elson EL. 1993. Correlation of myosin light chain phosphorylation with isometric contractionof fibroblasts. J. Biol. Chem. 268:23850–55
Kolodney MS, Wysolmerski RB. 1992. Isometric contraction by fibroblasts and endothelial cells in tissueculture. J. Cell Biol. 117:73–82
Kreger ST, Voytik-Harbin SL. 2009. Hyaluronan concentration within a 3D collagen matrix modulates matrixviscoelasticity, but not fibroblast response. Matrix Biol. 28:336–46
Lammermann T, Sixt M. 2009. Mechanical modes of ‘amoeboid’ cell migration. Curr. Opin. Cell Biol. 21:636–44
Langevin HM, Bouffard NA, Badger GJ, Iatridis JC, Howe AK. 2005. Dynamic fibroblast cytoskeletal responseto subcutaneous tissue stretch ex vivo and in vivo. Am. J. Physiol. Cell Physiol. 288:C747–56
Langevin HM, Cornbrooks CJ, Taatjes DJ. 2004. Fibroblasts form a body-wide cellular network. Histochem.Cell Biol. 122:7–15
Lauffenburger DA, Horwitz AF. 1996. Cell migration: a physically integrated molecular process. Cell 84:359–69
www.annualreviews.org • Cells in 3D Collagen Matrices 357
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Lee DJ, Ho CH, Grinnell F. 2003. LPA-stimulated fibroblast contraction of floating collagen matrices doesnot require Rho kinase activity or retraction of fibroblast extensions. Exp. Cell Res. 289:86–94
Leung LY, Tian D, Brangwynne CP, Weitz DA, Tschumperlin DJ. 2007. A new microrheometric approachreveals individual and cooperative roles for TGF-β1 and IL-1β in fibroblast-mediated stiffening ofcollagen gels. FASEB J. 21:2064–73
Lewis WH, Lewis MR. 1924. Behavior of cells in tissue cultures. In General Cytology, ed. EV Cowdry,pp. 384–447. Chicago: Univ. Chicago Press
Linge C, Richardson J, Vigor C, Clayton E, Hardas B, Rolfe K. 2005. Hypertrophic scar cells fail to undergo aform of apoptosis specific to contractile collagen—the role of tissue transglutaminase. J. Investig. Dermatol.125:72–82
Lo CM, Wang HB, Dembo M, Wang YL. 2000. Cell movement is guided by the rigidity of the substrate.Biophys. J. 79:144–52
Lutolf MP, Hubbell JA. 2005. Synthetic biomaterials as instructive extracellular microenvironments for mor-phogenesis in tissue engineering. Nat. Biotechnol. 23:47–55
MacDonald RA, Laurenzi BF, Viswanathan G, Ajayan PM, Stegemann JP. 2005. Collagen-carbon nanotubecomposite materials as scaffolds in tissue engineering. J. Biomed. Mater. Res. A 74:489–96
Macri L, Silverstein D, Clark RA. 2007. Growth factor binding to the pericellular matrix and its importancein tissue engineering. Adv. Drug Deliv. Rev. 59:1366–81
Marenzana M, Wilson-Jones N, Mudera V, Brown RA. 2006. The origins and regulation of tissue tension:identification of collagen tension-fixation process in vitro. Exp. Cell Res. 312:423–33
Martin P. 2007. Wound healing and inflammation studies in genetically tractable organisms. Int. Congr. Ser.1302:3–16
Mehra TD, Ghosh K, Shu XZ, Prestwich GD, Clark RA. 2006. Molecular stenting with a crosslinked hyaluro-nan derivative inhibits collagen gel contraction. J. Investig. Dermatol. 126:2202–9
Menezes GC, Miron-Mendoza M, Ho CH, Jiang H, Grinnell F. 2008. Oncogenic Ras-transformed humanfibroblasts exhibit differential changes in contraction and migration in 3D collagen matrices. Exp. CellRes. 314:3081–91
Meshel AS, Wei Q, Adelstein RS, Sheetz MP. 2005. Basic mechanism of three-dimensional collagen fibertransport by fibroblasts. Nat. Cell Biol. 7:157–64
Michalopoulos G, Pitot HC. 1975. Primary culture of parenchymal liver cells on collagen membranes. Mor-phological and biochemical observations. Exp. Cell Res. 94:70–78
Midwood KS, Schwarzbauer JE. 2002. Tenascin-C modulates matrix contraction via focal adhesion kinase-and Rho-mediated signaling pathways. Mol. Biol. Cell 13:3601–13
Miron-Mendoza M, Seemann J, Grinnell F. 2008. Collagen fibril flow and tissue translocation coupled tofibroblast migration in 3D collagen matrices. Mol. Biol. Cell 19:2051–58
Morales SA, Mareninov S, Prasad P, Wadehra M, Braun J, Gordon LK. 2007. Collagen gel contraction byARPE-19 cells is mediated by a FAK-Src dependent pathway. Exp. Eye. Res. 85:790–98
Munevar S, Wang Y, Dembo M. 2001. Traction force microscopy of migrating normal and H-ras transformed3T3 fibroblasts. Biophys. J. 80:1744–57
Nedelec B, Ghahary A, Scott PG, Tredget EE. 2000. Control of wound contraction. Basic and clinical features.Hand. Clin. 16:289–302
Ng CP, Hinz B, Swartz MA. 2005. Interstitial fluid flow induces myofibroblast differentiation and collagenalignment in vitro. J. Cell Sci. 118:4731–39
Nho RS, Xia H, Diebold D, Kahm J, Kleidon J, et al. 2006. PTEN regulates fibroblast elimination duringcollagen matrix contraction. J. Biol. Chem. 281:33291–301
Packard BZ, Artym VV, Komoriya A, Yamada KM. 2009. Direct visualization of protease activity on cellsmigrating in three-dimensions. Matrix. Biol. 28:3–10
Pankov R, Endo Y, Even-Ram S, Araki M, Clark K, et al. 2005. A Rac switch regulates random versusdirectionally persistent cell migration. J. Cell Biol. 170:793–802
Paszek MJ, Zahir N, Johnson KR, Lakins JN, Rozenberg GI, et al. 2005. Tensional homeostasis and themalignant phenotype. Cancer Cell 8:241–54
Pedersen JA, Boschetti F, Swartz MA. 2007. Effects of extracellular fiber architecture on cell membrane shearstress in a 3D fibrous matrix. J. Biomech. 40:1484–92
358 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Pedersen JA, Swartz MA. 2005. Mechanobiology in the third dimension. Ann. Biomed. Eng. 33:1469–90Pelham RJ Jr, Wang Y. 1997. Cell locomotion and focal adhesions are regulated by substrate flexibility. Proc.
Natl. Acad. Sci. USA 94:13661–65Petroll WM, Cavanagh HD, Jester JV. 2004a. Dynamic three-dimensional visualization of collagen matrix
remodeling and cytoskeletal organization in living corneal fibroblasts. Scanning 26:1–10Petroll WM, Ma L. 2003. Direct, dynamic assessment of cell-matrix interactions inside fibrillar collagen
lattices. Cell Motil. Cytoskelet. 55:254–64Petroll WM, Ma L. 2008. Localized application of mechanical and biochemical stimuli in 3-D culture. Dev.
Dyn. 237:2726–36Petroll WM, Ma L, Kim A, Ly L, Vishwanath M. 2008a. Dynamic assessment of fibroblast mechanical activity
during Rac-induced cell spreading in 3-D culture. J. Cell Physiol. 217:162–71Petroll WM, Ma L, Ly L, Vishwanath M. 2008b. Analysis of the pattern of subcellular force generation by
corneal fibroblasts following Rho activation. Eye Contact Lens 34:65–70Petroll WM, Vishwanath M, Ma L. 2004b. Corneal fibroblasts respond rapidly to changes in local mechanical
stress. Invest. Ophthalmol. Vis. Sci. 45:3466–74Plant AL, Bhadriraju K, Spurlin TA, Elliott JT. 2009. Cell response to matrix mechanics: focus on collagen.
Biochim. Biophys. Acta 1793:893–902Powell HM, Boyce ST. 2009. Engineered human skin fabricated using electrospun collagen-PCL blends:
morphogenesis and mechanical properties. Tissue Eng. A 15:2177–87Provenzano PP, Eliceiri KW, Campbell JM, Inman DR, White JG, Keely PJ. 2006. Collagen reorganization
at the tumor-stromal interface facilitates local invasion. BMC Med. 4:38Racine-Samson L, Rockey DC, Bissell DM. 1997. The role of α1β1 integrin in wound contraction: a quan-
titative analysis of liver myofibroblasts in vivo and in primary culture. J. Biol. Chem. 272:30911–17Raeber GP, Lutolf MP, Hubbell JA. 2008. Part II: Fibroblasts preferentially migrate in the direction of
principal strain. Biomech. Model. Mechanobiol. 7:215–25Rehfeldt F, Engler AJ, Eckhardt A, Ahmed F, Discher DE. 2007. Cell responses to the mechanochemical
microenvironment—implications for regenerative medicine and drug delivery. Adv. Drug Deliv. Rev.59:1329–39
Rhee S, Grinnell F. 2006. P21-activated kinase 1: convergence point in PDGF- and LPA-stimulated collagenmatrix contraction by human fibroblasts. J. Cell Biol. 172:423–32
Rhee S, Grinnell F. 2007. Fibroblast mechanics in 3D collagen matrices. Adv. Drug Deliv. Rev. 59:1299–305Rhee S, Ho CH, Grinnell F. 2010. Promigratory and procontractile growth factor environments differentially
regulate cell morphogenesis. Exp. Cell Res. 316:232–44Rhee S, Jiang H, Ho CH, Grinnell F. 2007. Microtubule function in fibroblast spreading is modulated ac-
cording to the tension state of cell-matrix interactions. Proc. Natl. Acad. Sci. USA 104:5425–30Ridley AJ, Hall A. 1992. The small GTP-binding protein rho regulates the assembly of focal adhesions and
actin stress fibers in response to growth factors. Cell 70:389–99Rinn JL, Wang JK, Liu H, Montgomery K, van de Rijn M, Chang HY. 2008. A systems biology approach to
anatomic diversity of skin. J. Investig. Dermatol. 128:776–82Rowe RG, Li XY, Hu Y, Saunders TL, Virtanen I, et al. 2009. Mesenchymal cells reactivate Snail1 expression
to drive three-dimensional invasion programs. J. Cell Biol. 184:399–408Roy P, Petroll WM, Cavanagh HD, Chuong CJ, Jester JV. 1997. An in vitro force measurement assay to
study the early mechanical interaction between corneal fibroblasts and collagen matrix. Exp. Cell Res.232:106–17
Sakai K, Sumi Y, Muramatsu H, Hata K, Muramatsu T, Ueda M. 2003. Thrombospondin-1 promotesfibroblast-mediated collagen gel contraction caused by activation of latent transforming growth factorβ-1. J. Dermatol. Sci. 31:99–109
Salomon D, Saurat JH, Meda P. 1988. Cell-to-cell communication within intact human skin. J. Clin. Invest.82:248–54
Sanz-Moreno V, Gadea G, Ahn J, Paterson H, Marra P, et al. 2008. Rac activation and inactivation controlplasticity of tumor cell movement. Cell 135:510–23
Sawhney RK, Howard J. 2002. Slow local movements of collagen fibers by fibroblasts drive the rapid globalself-organization of collagen gels. J. Cell Biol. 157:1083–91
www.annualreviews.org • Cells in 3D Collagen Matrices 359
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Schafer IA, Shapiro A, Kovach M, Lang C, Fratianne RB. 1989. The interaction of human papillary andreticular fibroblasts and human keratinocytes in the contraction of three-dimensional floating collagenlattices. Exp. Cell Res. 183:112–25
Schiro JA, Chan BM, Roswit WT, Kassner PD, Pentland AP, et al. 1991. Integrin α2β1 (VLA-2) mediatesreorganization and contraction of collagen matrices by human cells. Cell 67:403–10
Seliktar D, Black RA, Vito RP, Nerem RM. 2000. Dynamic mechanical conditioning of collagen-gel bloodvessel constructs induces remodeling in vitro. Ann. Biomed. Eng. 28:351–62
Sheetz MP, Dai J. 1996. Modulation of membrane dynamics and cell motility by membrane tension. TrendsCell Biol. 6:85–89
Sheetz MP, Felsenfeld D, Galbraith CG, Choquet D. 1999. Cell migration as a five-step cycle. Biochem. Soc.Symp. 65:233–43
Shi-Wen X, Chen Y, Denton CP, Eastwood M, Renzoni EA, et al. 2004. Endothelin-1 promotes myofibroblastinduction through the ETA receptor via a rac/phosphoinositide 3-kinase/Akt-dependent pathway and isessential for the enhanced contractile phenotype of fibrotic fibroblasts. Mol. Biol. Cell 15:2707–19
Silver FH, Siperko LM, Seehra GP. 2002. Mechanobiology of force transduction in dermal tissue. Skin Res.Technol. 8:1–21
Skalli O, Ropraz P, Trzeciak A, Benzonana G, Gillessen D, Gabbiani G. 1986. A monoclonal antibody againstα-smooth muscle actin: a new probe for smooth muscle differentiation. J. Cell Biol. 103:2787–96
Sorrell JM, Caplan AI. 2009. Fibroblasts—a diverse population at the center of it all. Int. Rev. Cell Mol. Biol.276:161–214
Squier CA. 1981. The effect of stretching on formation of myofibroblasts in mouse skin. Cell Tissue Res.220:325–35
Steinberg BM, Smith K, Colozzo M, Pollack R. 1980. Establishment and transformation diminish the abilityof fibroblasts to contract a native collagen gel. J. Cell Biol. 87:304–8
Stopak D, Harris AK. 1982. Connective tissue morphogenesis by fibroblast traction. I. Tissue culture obser-vations. Dev. Biol. 90:383–98
Storm C, Pastore JJ, MacKintosh FC, Lubensky TC, Janmey PA. 2005. Nonlinear elasticity in biological gels.Nature 435:191–94
Stratman AN, Saunders WB, Sacharidou A, Koh W, Fisher KE, et al. 2009. Endothelial cell lumen and vas-cular guidance tunnel formation requires MT1-MMP-dependent proteolysis in 3-dimensional collagenmatrices. Blood 114:237–47
Sugimoto N, Takuwa N, Okamoto H, Sakurada S, Takuwa Y. 2003. Inhibitory and stimulatory regulationof Rac and cell motility by the G12/13-Rho and Gi pathways integrated downstream of a single Gprotein-coupled sphingosine-1-phosphate receptor isoform. Mol. Cell Biol. 23:1534–45
Tamaoki M, Imanaka-Yoshida K, Yokoyama K, Nishioka T, Inada H, et al. 2005. Tenascin-C regulatesrecruitment of myofibroblasts during tissue repair after myocardial injury. Am. J. Pathol. 167:71–80
Tamariz E, Grinnell F. 2002. Modulation of fibroblast morphology and adhesion during collagen matrixremodeling. Mol. Biol. Cell 13:3915–29
Tiger CF, Fougerousse F, Grundstrom G, Velling T, Gullberg D. 2001. α11β1 integrin is a receptor forinterstitial collagens involved in cell migration and collagen reorganization on mesenchymal nonmusclecells. Dev. Biol. 237:116–29
Tomasek JJ, Akiyama SK. 2005. Fibroblast-mediated collagen gel contraction does not require fibronectin-α5β1 integrin interaction. Anat. Rec. 234:153–60
Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. 2002. Myofibroblasts and mechano-regulationof connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 3:349–63
Tranquillo RT. 1999. Self-organization of tissue-equivalents: the nature and role of contact guidance. Biochem.Soc. Symp. 65:27–42
Trimboli AJ, Cantemir-Stone CZ, Li F, Wallace JA, Merchant A, et al. 2009. Pten in stromal fibroblastssuppresses mammary epithelial tumours. Nature 461:1084–91
Tuan TL, Grinnell F. 1989. Fibronectin and fibrinolysis are not required for fibrin gel contraction by humanskin fibroblasts. J. Cell Physiol. 140:577–83
Vanni S, Lagerholm BC, Otey C, Taylor DL, Lanni F. 2003. Internet-based image analysis quantifies con-tractile behavior of individual fibroblasts inside model tissue. Biophys. J. 84:2715–27
360 Grinnell · Petroll
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

CB26CH14-Grinnell ARI 3 September 2010 19:34
Vishwanath M, Ma L, Jester JV, Otey CA, Petroll WM. 2003. Modulation of corneal fibroblast contractilitywithin fibrillar collagen matrices. Investig. Ophthalmol. Vis. Sci. 44:4724–35
Vogel WF, Abdulhussein R, Ford CE. 2006. Sensing extracellular matrix: an update on discoidin domainreceptor function. Cell Signal. 18:1108–16
Wakatsuki T, Kolodney MS, Zahalak GI, Elson EL. 2000. Cell mechanics studied by a reconstituted modeltissue. Biophys. J. 79:2353–68
Wang F, Garza LA, Kang S, Varani J, Orringer JS, et al. 2007. In vivo stimulation of de novo collagenproduction caused by cross-linked hyaluronic acid dermal filler injections in photodamaged human skin.Arch. Dermatol. 143:155–63
Wei WC, Lin HH, Shen MR, Tang MJ. 2008. Mechanosensing machinery for cells under low substratumrigidity. Am. J. Physiol. Cell Physiol. 295:C1579–89
Weigelt B, Bissell MJ. 2008. Unraveling the microenvironmental influences on the normal mammary glandand breast cancer. Semin. Cancer Biol. 18:311–21
Weiss P. 1959. Cellular dynamics. Rev. Mod. Phys. 31:11–20Wipff PJ, Rifkin DB, Meister JJ, Hinz B. 2007. Myofibroblast contraction activates latent TGF-β1 from the
extracellular matrix. J. Cell Biol. 179:1311–23Wolf K, Alexander S, Schacht V, Coussens LM, van Andrian UH, et al. 2009. Collagen-based cell migration
models in vitro and in vivo. Semin. Cell. Dev. Biol. 20:931–41Wolf K, Mazo I, Leung H, Engelke K, van Andrian UH, et al. 2003. Compensation mechanism in tumor
cell migration: mesenchymal-amoeboid transition after blocking of pericellular proteolysis. J. Cell Biol.160:267–77
Woodley DT, Yamauchi M, Wynn KC, Mechanic G, Briggaman RA. 1991. Collagen telopeptides (cross-linking sites) play a role in collagen gel lattice contraction. J. Investig. Dermatol. 97:580–85
Wyckoff JB, Pinner SE, Gschmeissner S, Condeelis JS, Sahai E. 2006. ROCK- and myosin-dependent matrixdeformation enables protease-independent tumor-cell invasion in vivo. Curr. Biol. 16:1515–23
Wynn TA. 2008. Cellular and molecular mechanisms of fibrosis. J. Pathol. 214:199–210Xu R, Boudreau A, Bissell MJ. 2009. Tissue architecture and function: dynamic reciprocity via extra- and
intracellular matrices. Cancer Metastasis Rev. 28:167–76Yamada KM, Cukierman E. 2007. Modeling tissue morphogenesis and cancer in 3D. Cell 130:601–10Yeh AT, Kao B, Jung WG, Chen Z, Nelson JS, Tromberg BJ. 2004. Imaging wound healing using optical
coherence tomography and multiphoton microscopy in an in vitro skin-equivalent tissue model. J. Biomed.Opt. 9:248–53
Yeung T, Georges PC, Flanagan LA, Marg B, Ortiz M, et al. 2005. Effects of substrate stiffness on cellmorphology, cytoskeletal structure, and adhesion. Cell Motil. Cytoskelet. 60:24–34
Zaman MH, Trapani LM, Siemeski A, MacKellar D, Gong H, et al. 2006. Migration of tumor cells in 3Dmatrices is governed by matrix stiffness along with cell-matrix adhesion and proteolysis. Proc. Natl. Acad.Sci. USA 103:10889–94
Zhang Z, Garron TM, Li XJ, Liu Y, Zhang X, et al. 2009. Recombinant human decorin inhibits TGF-β1-induced contraction of collagen lattice by hypertrophic scar fibroblasts. Burns 35:527–37
www.annualreviews.org • Cells in 3D Collagen Matrices 361
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

AR425-FM ARI 14 September 2010 18:54
Annual Reviewof Cell andDevelopmentalBiology
Volume 26, 2010
ContentsEnzymes, Embryos, and Ancestors
John Gerhart � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 1
Control of Mitotic Spindle LengthGohta Goshima and Jonathan M. Scholey � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �21
Trafficking to the Ciliary Membrane: How to Get Acrossthe Periciliary Diffusion Barrier?Maxence V. Nachury, E. Scott Seeley, and Hua Jin � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �59
Transmembrane Signaling ProteoglycansJohn R. Couchman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � �89
Membrane Fusion: Five Lipids, Four SNAREs, Three Chaperones,Two Nucleotides, and a Rab, All Dancing in a Ring on Yeast VacuolesWilliam Wickner � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 115
Tethering Factors as Organizers of Intracellular Vesicular TrafficI-Mei Yu and Frederick M. Hughson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 137
The Diverse Functions of Oxysterol-Binding ProteinsSumana Raychaudhuri and William A. Prinz � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 157
Ubiquitination in Postsynaptic Function and PlasticityAngela M. Mabb and Michael D. Ehlers � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 179
α-Synuclein: Membrane Interactions and Toxicityin Parkinson’s DiseasePavan K. Auluck, Gabriela Caraveo, and Susan Lindquist � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 211
Novel Research Horizons for Presenilins and γ-Secretases in CellBiology and DiseaseBart De Strooper and Wim Annaert � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 235
Modulation of Host Cell Function by Legionella pneumophilaType IV EffectorsAndree Hubber and Craig R. Roy � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 261
A New Wave of Cellular ImagingDerek Toomre and Joerg Bewersdorf � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 285
Mechanical Integration of Actin and Adhesion Dynamicsin Cell MigrationMargaret L. Gardel, Ian C. Schneider, Yvonne Aratyn-Schaus,
and Clare M. Waterman � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 315
vii
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

AR425-FM ARI 14 September 2010 18:54
Cell Motility and Mechanics in Three-Dimensional Collagen MatricesFrederick Grinnell and W. Matthew Petroll � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 335
Rolling Cell AdhesionRodger P. McEver and Cheng Zhu � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 363
Assembly of Fibronectin Extracellular MatrixPurva Singh, Cara Carraher, and Jean E. Schwarzbauer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 397
Interactions Between Nuclei and the Cytoskeleton Are Mediatedby SUN-KASH Nuclear-Envelope BridgesDaniel A. Starr and Heidi N. Fridolfsson � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 421
Plant Nuclear Hormone Receptors: A Role for Small Moleculesin Protein-Protein InteractionsShelley Lumba, Sean R. Cutler, and Peter McCourt � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 445
Mammalian Su(var) Genes in Chromatin ControlBarna D. Fodor, Nicholas Shukeir, Gunter Reuter, and Thomas Jenuwein � � � � � � � � � � � � � � 471
Chromatin Regulatory Mechanisms in PluripotencyJulie A. Lessard and Gerald R. Crabtree � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 503
Presentation Counts: Microenvironmental Regulation of Stem Cells byBiophysical and Material CuesAlbert J. Keung, Sanjay Kumar, and David V. Schaffer � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 533
Paramutation and DevelopmentJay B. Hollick � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 557
Assembling Neural Crest Regulatory Circuits intoa Gene Regulatory NetworkPaola Betancur, Marianne Bronner-Fraser, and Tatjana Sauka-Spengler � � � � � � � � � � � � � � � 581
Regulatory Mechanisms for Specification and Patterning of PlantVascular TissuesAna Cano-Delgado, Ji-Young Lee, and Taku Demura � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 605
Common Factors Regulating Patterning of the Nervousand Vascular SystemsMariana Melani and Brant M. Weinstein � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 639
Stem Cell Models of Cardiac Development and DiseaseKiran Musunuru, Ibrahim J. Domian, and Kenneth R. Chien � � � � � � � � � � � � � � � � � � � � � � � � � � 667
Stochastic Mechanisms of Cell Fate Specification that Yield Randomor Robust OutcomesRobert J. Johnston, Jr. and Claude Desplan � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 689
A Decade of Systems BiologyHan-Yu Chuang, Matan Hofree, and Trey Ideker � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � � 721
viii Contents
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.

AnnuAl Reviewsit’s about time. Your time. it’s time well spent.
AnnuAl Reviews | Connect with Our expertsTel: 800.523.8635 (us/can) | Tel: 650.493.4400 | Fax: 650.424.0910 | Email: [email protected]
New From Annual Reviews:
Annual Review of Statistics and Its ApplicationVolume 1 • Online January 2014 • http://statistics.annualreviews.org
Editor: Stephen E. Fienberg, Carnegie Mellon UniversityAssociate Editors: Nancy Reid, University of Toronto
Stephen M. Stigler, University of ChicagoThe Annual Review of Statistics and Its Application aims to inform statisticians and quantitative methodologists, as well as all scientists and users of statistics about major methodological advances and the computational tools that allow for their implementation. It will include developments in the field of statistics, including theoretical statistical underpinnings of new methodology, as well as developments in specific application domains such as biostatistics and bioinformatics, economics, machine learning, psychology, sociology, and aspects of the physical sciences.
Complimentary online access to the first volume will be available until January 2015. table of contents:•What Is Statistics? Stephen E. Fienberg•A Systematic Statistical Approach to Evaluating Evidence
from Observational Studies, David Madigan, Paul E. Stang, Jesse A. Berlin, Martijn Schuemie, J. Marc Overhage, Marc A. Suchard, Bill Dumouchel, Abraham G. Hartzema, Patrick B. Ryan
•The Role of Statistics in the Discovery of a Higgs Boson, David A. van Dyk
•Brain Imaging Analysis, F. DuBois Bowman•Statistics and Climate, Peter Guttorp•Climate Simulators and Climate Projections,
Jonathan Rougier, Michael Goldstein•Probabilistic Forecasting, Tilmann Gneiting,
Matthias Katzfuss•Bayesian Computational Tools, Christian P. Robert•Bayesian Computation Via Markov Chain Monte Carlo,
Radu V. Craiu, Jeffrey S. Rosenthal•Build, Compute, Critique, Repeat: Data Analysis with Latent
Variable Models, David M. Blei•Structured Regularizers for High-Dimensional Problems:
Statistical and Computational Issues, Martin J. Wainwright
•High-Dimensional Statistics with a View Toward Applications in Biology, Peter Bühlmann, Markus Kalisch, Lukas Meier
•Next-Generation Statistical Genetics: Modeling, Penalization, and Optimization in High-Dimensional Data, Kenneth Lange, Jeanette C. Papp, Janet S. Sinsheimer, Eric M. Sobel
•Breaking Bad: Two Decades of Life-Course Data Analysis in Criminology, Developmental Psychology, and Beyond, Elena A. Erosheva, Ross L. Matsueda, Donatello Telesca
•Event History Analysis, Niels Keiding•StatisticalEvaluationofForensicDNAProfileEvidence,
Christopher D. Steele, David J. Balding•Using League Table Rankings in Public Policy Formation:
Statistical Issues, Harvey Goldstein•Statistical Ecology, Ruth King•Estimating the Number of Species in Microbial Diversity
Studies, John Bunge, Amy Willis, Fiona Walsh•Dynamic Treatment Regimes, Bibhas Chakraborty,
Susan A. Murphy•Statistics and Related Topics in Single-Molecule Biophysics,
Hong Qian, S.C. Kou•Statistics and Quantitative Risk Management for Banking
and Insurance, Paul Embrechts, Marius Hofert
Access this and all other Annual Reviews journals via your institution at www.annualreviews.org.
Ann
u. R
ev. C
ell D
ev. B
iol.
2010
.26:
335-
361.
Dow
nloa
ded
from
ww
w.a
nnua
lrev
iew
s.or
gby
Uni
vers
ity o
f T
exas
Sou
thw
este
rn M
edic
al C
ente
r on
07/
11/1
4. F
or p
erso
nal u
se o
nly.




















