
Distinguishing fibroblast promigratory and procontractile growth factor environments in 3-D collagen...
10
The FASEB Journal • Research Communication Distinguishing fibroblast promigratory and procontractile growth factor environments in 3-D collagen matrices Hongmei Jiang, Sangmyung Rhee, Chin-Han Ho, and Frederick Grinnell 1 Department of Cell Biology, University of Texas Southwestern Medical School, Dallas, Texas ABSTRACT Understanding growth factor function during wound repair is necessary for the development of therapeutic interventions to improve healing out- comes. In the current study, we compare the effects of serum and purified growth factors on human fibroblast function in three different collagen matrix models: cell migration in nested matrices, floating matrix contrac- tion, and stressed-released matrix contraction. The results of these studies indicate that platelet-derived growth factor (PDGF) is unique in its capacity to promote cell migration. Serum, lysophosphatidic acid, sphingosine-1-phophate (S1P), and endothelin-1 pro- mote stressed-released matrix contraction but not cell migration. In addition, we found that S1P inhibits fibroblast migration and treatment of serum to remove lipid growth factors or treatment of cells to interfere with S1P 2 receptor function increases serum promigra- tory activity. Our findings suggest that different sets of growth factors generate promigratory and procontrac- tile tissue environments for fibroblasts and that the balance between PDGF and S1P is a key determinant of fibroblast promigratory activity.—Jiang, H., Rhee, S., Ho, C.-H., Grinnell, F. Distinguishing fibroblast promi- gratory and procontractile growth factor environments in 3-D collagen matrices. FASEB J. 22, 2151–2160 (2008) Key Words: cell migration cell contraction platelet-derived growth factor lysophosphatidic acid sphingosine-1-phos- phate wound repair In 1979, Bell et al. (1) reported that fibroblast contraction of type I collagen matrices resulted in formation of tissue-like structures, which were called “dermal equivalents” in anticipation of their use as part of living skin grafts (2). Subsequent to Bell’s initial observations, three dimensional collagen and fibrin matrices containing fibroblasts and other cell types have been used to study cellular remodeling of connec- tive tissue-like matrices and to characterize iterative cellular adaptation to the changing biomechanical fea- tures of the three-dimensional (3-D) matrix environ- ment (3, 4). Mechanical remodeling of collagen and other extracellular matrix (ECM) molecules by fibro- blasts and myofibroblasts (5) has been implicated in diverse physiological processes including wound repair (6 – 8), fibrosis (9 –11), and malignant transformation (12–14). 3-D matrices differ from two-dimensional (2-D) pla- nar surfaces in a variety of ways, such as substratum stiffness and porosity and the distribution and density of matrix adhesion sites (3, 6). Of particular signifi- cance, collagen matrices exhibit viscoelastic properties that resemble connective tissues (15–18). These vis- coelastic characteristics, i.e., the ability of collagen and other molecules within the tissue matrix to stretch and slip past each other and undergo stable reorganization, permit connective tissue remodeling to occur (19). Surgical procedures take advantage of connective tissue viscoelastic properties when mechanical stretch is used for tissue expansion (20, 21). Unlike connective tissue matrices, ECM molecules adsorbed or covalently linked to conventional planar surfaces (e.g., plastic, glass, and polyacrylamide) tend to stay in register with each other and have little capacity to undergo stable, cell-mediated mechanical and molecular reorganization. Conse- quently, 3-D matrix models permit analysis of cell- matrix interactions that cannot be studied with ECM- coated materials (22). The matrix contraction model introduced by Bell et al. (1) often is referred to as floating matrix contrac- tion. The mechanism of floating matrix contraction involves cell motile activity but not cell contraction (23–25). If, however, fibroblasts develop mechanical stress before the matrices are released and are allowed to float in culture medium, then matrix contraction (stressed-released) depends on cell contraction (26 – 29). Floating and stressed-released matrix contraction show different patterns of growth factor stimulation. Serum, platelet-derived growth factor (PDGF), and lysophosphatidic acid (LPA) promote floating matrix contraction, whereas serum and LPA but not PDGF promote stressed-released matrix contraction (30). Re- cently, we observed yet another pattern of growth factor regulation in studies on fibroblast migration in nested collagen matrices (dermal equivalents surrounded by 1 Correspondence: Department of Cell Biology, University of Texas Southwestern Medical School, 5323 Harry Hines Boulevard, Dallas, TX 75390-9039, USA. E-mail: frederick. [email protected] doi: 10.1096/fj.07-097014 2151 0892-6638/08/0022-2151 © FASEB
Transcript of Distinguishing fibroblast promigratory and procontractile growth factor environments in 3-D collagen...

The FASEB Journal • Research Communication
Distinguishing fibroblast promigratory andprocontractile growth factor environments in 3-Dcollagen matrices
Hongmei Jiang, Sangmyung Rhee, Chin-Han Ho, and Frederick Grinnell1
Department of Cell Biology, University of Texas Southwestern Medical School, Dallas, Texas
ABSTRACT Understanding growth factor functionduring wound repair is necessary for the developmentof therapeutic interventions to improve healing out-comes. In the current study, we compare the effects ofserum and purified growth factors on human fibroblastfunction in three different collagen matrix models: cellmigration in nested matrices, floating matrix contrac-tion, and stressed-released matrix contraction. Theresults of these studies indicate that platelet-derivedgrowth factor (PDGF) is unique in its capacity topromote cell migration. Serum, lysophosphatidic acid,sphingosine-1-phophate (S1P), and endothelin-1 pro-mote stressed-released matrix contraction but not cellmigration. In addition, we found that S1P inhibitsfibroblast migration and treatment of serum to removelipid growth factors or treatment of cells to interferewith S1P2 receptor function increases serum promigra-tory activity. Our findings suggest that different sets ofgrowth factors generate promigratory and procontrac-tile tissue environments for fibroblasts and that thebalance between PDGF and S1P is a key determinant offibroblast promigratory activity.—Jiang, H., Rhee, S.,Ho, C.-H., Grinnell, F. Distinguishing fibroblast promi-gratory and procontractile growth factor environmentsin 3-D collagen matrices. FASEB J. 22, 2151–2160 (2008)
Key Words: cell migration � cell contraction � platelet-derivedgrowth factor � lysophosphatidic acid � sphingosine-1-phos-phate � wound repair
In 1979, Bell et al. (1) reported that fibroblastcontraction of type I collagen matrices resulted information of tissue-like structures, which were called“dermal equivalents” in anticipation of their use as partof living skin grafts (2). Subsequent to Bell’s initialobservations, three dimensional collagen and fibrinmatrices containing fibroblasts and other cell typeshave been used to study cellular remodeling of connec-tive tissue-like matrices and to characterize iterativecellular adaptation to the changing biomechanical fea-tures of the three-dimensional (3-D) matrix environ-ment (3, 4). Mechanical remodeling of collagen andother extracellular matrix (ECM) molecules by fibro-blasts and myofibroblasts (5) has been implicated indiverse physiological processes including wound repair
(6–8), fibrosis (9–11), and malignant transformation(12–14).
3-D matrices differ from two-dimensional (2-D) pla-nar surfaces in a variety of ways, such as substratumstiffness and porosity and the distribution and densityof matrix adhesion sites (3, 6). Of particular signifi-cance, collagen matrices exhibit viscoelastic propertiesthat resemble connective tissues (15–18). These vis-coelastic characteristics, i.e., the ability of collagen andother molecules within the tissue matrix to stretch andslip past each other and undergo stable reorganization,permit connective tissue remodeling to occur (19).Surgical procedures take advantage of connective tissueviscoelastic properties when mechanical stretch is usedfor tissue expansion (20, 21). Unlike connective tissuematrices, ECM molecules adsorbed or covalently linkedto conventional planar surfaces (e.g., plastic, glass, andpolyacrylamide) tend to stay in register with each otherand have little capacity to undergo stable, cell-mediatedmechanical and molecular reorganization. Conse-quently, 3-D matrix models permit analysis of cell-matrix interactions that cannot be studied with ECM-coated materials (22).
The matrix contraction model introduced by Bell etal. (1) often is referred to as floating matrix contrac-tion. The mechanism of floating matrix contractioninvolves cell motile activity but not cell contraction(23–25). If, however, fibroblasts develop mechanicalstress before the matrices are released and are allowedto float in culture medium, then matrix contraction(stressed-released) depends on cell contraction (26–29). Floating and stressed-released matrix contractionshow different patterns of growth factor stimulation.Serum, platelet-derived growth factor (PDGF), andlysophosphatidic acid (LPA) promote floating matrixcontraction, whereas serum and LPA but not PDGFpromote stressed-released matrix contraction (30). Re-cently, we observed yet another pattern of growth factorregulation in studies on fibroblast migration in nestedcollagen matrices (dermal equivalents surrounded by
1 Correspondence: Department of Cell Biology, Universityof Texas Southwestern Medical School, 5323 Harry HinesBoulevard, Dallas, TX 75390-9039, USA. E-mail: [email protected]
doi: 10.1096/fj.07-097014
21510892-6638/08/0022-2151 © FASEB

cell-free outer collagen matrices). In nested collagenmatrices, fibroblast migration is stimulated by PDGFbut not by serum or LPA (31).
Understanding the timing and functions of physio-logical growth factors in the wound environment isnecessary to develop therapeutic strategies to improvehealing outcomes (32–35). During wound repair, fibro-blasts are believed to migrate into the wound region,develop mechanical stress, and undergo contraction (7,9–11). The possibility that fibroblast migration andcontraction might be regulated by different growthfactors was already implicit in the original work thatidentified PDGF and serum lysophospholipids as acti-vators of the small G proteins Rac and Rho, respectively(36, 37). The current study was carried out to investi-gate the above possibility in more detail, using 3-Dmatrix assays to compare serum, PDGF, LPA, andseveral other growth factors that have been shownunder diverse conditions and with different cell types topromote matrix contraction, namely, transforminggrowth factor-� (TGF-�; ref. 38), sphingosine-1-phos-phate (S1P; ref. 39), epidermal growth factor (EGF; ref.40), endothelin-1 (endo-1; ref. 41), and basic fibroblastgrowth factor (bFGF; ref. 42).
The results indicate that PDGF is unique in itscapacity to promote human fibroblast migration. Se-rum, LPA, S1P, and endo-1 promote stressed-releasedmatrix contraction but not cell migration. In addition,we found that S1P inhibits fibroblast migration, andtreatment of serum to remove lipid growth factors ortreatment of cells to interfere with S1P2 receptor func-tion increases serum promigratory activity. Our find-ings are consistent with the idea that different sets ofgrowth factors generate promigratory and procontrac-tile tissue environments for fibroblasts and that thebalance between PDGF and S1P is a key determinant offibroblast promigratory activity.
MATERIALS AND METHODS
Materials
Type I collagen (3 mg/ml, Vitrogen) was purchased fromCohesion (Palo Alto, CA, USA). Dulbecco modified Eaglemedium (DMEM), TRIzol reagent, Superscript II, Opti-MEMI, oligofectamine, Platinum Pfx DNA polymerase, and 0.25%trypsin/EDTA solution were purchased from Invitrogen(Gaithersburg, MD, USA). Fetal bovine serum (FBS) andadult human serum (HS) were purchased from Gemini(Woodland, CA, USA). PDGF (BB isotype) was purchasedfrom Upstate Biotechnology (Lake Placid, NY, USA). Fattyacid-free bovine serum albumin (BSA), LPA, S1P, EGF,endo-1, bFGF, and activated charcoal were obtained fromSigma (St. Louis, MO, USA). TGF-�, Rho kinase inhibitorY27632, and S1P2 receptor inhibitor JTE-013 were obtainedfrom Calbiochem-Novabiochem (La Jolla, CA, USA). Fluoro-mount G was obtained from Southern Biotechnology Associ-ates (Birmingham, AL, USA).
S1P and other lipids were removed from serum by charcoalstripping (43). Ten milliliter serum samples were incubatedovernight at 4°C with 1 g of activated charcoal. Samples were
centrifuged at 2000 g for 10 min, and the supernatants werefiltered using a 0.22 mm cellular acetate filter.
Cell culture and collagen matrix measurements
Use of human foreskin fibroblasts was approved by theUniversity Institutional Review Board (Exemption 4). Earlypassage human foreskin fibroblasts (hTERT-immortalized;ref. 44) were cultured in DMEM supplemented with 10% FBSat 37°C in a 5% CO2 humidified incubator. All experimentswere carried out two or more times.
Methods for preparing collagen matrix cultures and mea-suring floating collagen matrix contraction, stressed-releasedmatrix contraction, and cell migration in nested collagenmatrices have been described previously (29, 31). Briefly, tomeasure floating matrix contraction, collagen matrices (200�l, 1.5 mg/ml collagen, 2�105 cells/matrix) were polymer-ized for 1 h, then released from the culture surface, andincubated for 4 h floating in basal medium (DMEM�5mg/ml BSA) and growth factors as shown. To measurestressed-released matrix contraction, collagen matrices asabove were cultured overnight in DMEM/10% FBS, rinsed,and then released from the culture surface and incubated for1 h floating in basal medium and growth factors as shown. Atthe end of the incubations, samples were fixed with 3%paraformaldehyde in phosphate buffered saline, and theextent of matrix contraction was measured as decrease inmatrix diameter.
To measure cell migration using nested collagen matrices,floating matrices were incubated 4 h in DMEM/10% FBS,after which the cell-containing contracted matrices werere-embedded in 200 �l of collagen matrices (1.5 mg/mlcollagen) and incubated for an additional 24 h in basalmedium and growth factors as shown. At the end of theincubations, samples were fixed as above and then stainedwith propidium iodine (Molecular Probes, Eugene, OR,USA) to detect cell nuclei. Cell migration index was calcu-lated by counting the average number of cells that hadmigrated out of dermal equivalents in five �10 microscopicfields selected arbitrarily. Each field included the border ofthe dermal equivalent (detected by dark field microscopy)and the furthest moving cells (detected by nuclear stainingwith propidium iodide).
To measure cell migration by scrape wounding, fibroblastswere incubated overnight on collagen-coated coverslips inDMEM/BSA and 0.1% FBS. The samples were scrapewounded with a pipette tip and then incubated an additional24 h in basal medium and growth factors as shown. At the endof the incubations, samples were fixed and then stained withpropidium iodine and with phalloidin to detect actin.
Actin staining, immunofluorescence microscopy, andimmunoblotting
Immunostaining of matrices was carried out as describedpreviously (31). For actin staining, we used Alexa Fluor 488-or 594-conjugated phalloidin (Molecular Probes). Imageswere collected at 22° with a Nikon Elipse 400 fluorescentmicroscope using �10/0.45, �20/0.75, and �40/0.75 NikonPlan Apo infinity corrected objectives (Nikon, Tokyo, Japan),Photometrics SenSys camera (Photometrics, Tucson, AZ,USA), and MetaVue acquisition software (Molecular Dynam-ics, Downingtown, PA, USA). Subsequent image processingwas carried out using Adobe Photoshop 5.5 or 7.0 (AdobeSystems, San Jose, CA, USA).
Dendritic cell index was determined using the MetaVueIntegrated Morphometry Analysis function. For each condi-tion, region measurements were made after outlining 20 cells.
2152 Vol. 22 July 2008 JIANG ET AL.The FASEB Journal

Dendritic index was calculated as perimeter2/4��area. Thedendritic index for a round cell is 1.0.
S1P receptor gene silencing and reverse transcriptase-polymerase chain reaction (RT-PCR)
Small interfering RNA (siRNA) silencing of gene expressionin human fibroblasts was accomplished as described previ-ously (25). Primer pairs were designed by and obtained fromDharmacon (Chicago, IL, USA): S1P1 siRNA: 5�-GCUCAA-GACCGUAAUUAUCUU-3� and 5�-GAUAAUUACGGUCUU-GAGCUU-3�; and S1P2 siRNA: 5�-UUGCCAAGGUCAAGCU-GUAUU-3� and 5�-UACAGCUUGACCUUGGCAAUU-3�. Toobtain high-efficiency transfection, fibroblast cultures (60–70% confluent) were rinsed with antibiotic-free DMEM andtreated with trypsin-EDTA for 1 min to elicit cell rounding butnot detachment. Subsequently, antibiotic-free DMEM/10%FBS was added to quench the trypsin. After cells were rinsedwith antibiotic-free DMEM, they were incubated with 1 mlOpti-MEM I and oligofectamine containing 300 nM siRNA.After 24 h, 1 ml of DMEM/20% FBS was added to thetransfection medium and incubations were continued for anadditional 36 h, at which time cells were subcultured. Mock-transfected cells were treated with the transfection reagentbut no siRNA.
Total RNA was isolated from either mock or siRNA-treatedcells using the TRIzol reagent according to the manufactur-er’s instructions. First-strand cDNA was synthesized from 1 �gof the RNA in a final volume of 20 �l using 1 �g of randomprimers and Superscript II reverse transcriptase in the pres-ence of RNase inhibitor (RNase Out, Invitrogen, Gaithers-burg, MD, USA) according to the manufacturer’s instruc-tions. To assess mRNA silencing, cDNAs were subjected toPCR analysis with specific sets of primers. For either S1P1 orS1P2 specific amplification, primers were designed as follows:forward S1P1 (nucleotides 501–520): 5�-AACTGACCTCGGT-GGTGTTC-3�, reverse S1P1 (nucleotides 817–836): 5�-AGT-TATTGCTCCCGTTGTGG-3�; and forward S1P2 (nucleotides90–109): 5�-ACCATGGGCAGCTTGTACTC-3�, reverse S1P2(nucleotide 445–464): 5�-CAGGAGGCTGAAGACAGAGG-3�.PCR was performed with Platinum Pfx DNA Polymerase usingGene Amp PCR system 9700 (Applied Biosystems, Foster City,CA, USA). PCR with GAPDH primers was carried out as aninternal control. To ensure that the assay was in the linearrange, the cycle number and amounts of RNA were varied.
RESULTS
Growth factor specificity of cell migration andcollagen matrix contraction
Figure 1A compares the ability of diverse growth factorsto stimulate floating collagen matrix contraction. Theconcentrations selected were those shown previously bydifferent laboratories to have agonist activity. PDGF,LPA, and serum (FBS and HS) were the most active instimulating floating matrix contraction, although all ofthe growth factors tested appeared to have some ago-nist effect above basal medium (BSA).
Figure 1B compares fibroblast migration in nestedcollagen matrices using the same growth factors andconcentrations as in Fig. 1A. PDGF at 50 ng/ml ap-peared to be much more promigratory than any of theother growth factors tested including HS and FBS. In
HS, the PDGF concentration is �50 ng/ml (45). Levelsof PDGF in fetal serum are lower (46). No additionalstimulation of contraction occurred for any of thegrowth factors tested at lower concentrations (up tothree orders of magnitude).
Finally, Fig. 1C shows stressed-released matrix con-traction in the presence of the different growth factors.In this assay, PDGF had little procontractile activity.Serum, LPA, S1P, and endo-1 were potent agonists eventhough these growth factors did not stimulate cellmigration.
Taken together, the results in Fig. 1 demonstratedthat different sets of growth factors act as agonists forfloating matrix contraction, stressed-released matrixcontraction, and fibroblast migration in nested colla-
Figure 1. Growth factor and serum stimulation of collagenmatrix contraction and fibroblast migration in nested colla-gen matrices. A) Floating collagen matrix (FMC) was carriedout for 4 h in medium containing growth factors and serumat the concentrations indicated. Matrix contraction valuesshown are the averages � sd for duplicate samples from 3separate experiments. Starting matrix diameter 12 mm.B) Nested collagen matrices were incubated 24 h in mediumcontaining growth factors and serum at the concentrationsindicated. Cell migration index values shown are the aver-ages � sd of duplicate samples from 3 separate experiments.C) Stressed-released collagen matrix contraction (SRMC) wascarried out for 1 h in medium containing growth factors andserum as indicated. Matrix contraction values shown are theaverages � sd of duplicate samples. Starting matrix diame-ter 12 mm.
2153PROMIGRATORY VS. PROCONTRACTILE ENVIRONMENTS
gen matrices. PDGF is unique in its ability to promotefibroblast migration in nested matrices but a weakagonist for stressed-released matrix contraction. Serum,LPA, S1P, and endo-1 are potent agonists for stressed-released matrix contraction but not for cell migration.In contrast to stressed-released matrix contraction andcell migration, contraction of floating collagen matri-ces shows little growth factor specificity.
The results observed in Fig. 1B also suggested thatS1P might be an inhibitor of human fibroblast migra-tion in nested collagen matrices because the migrationindex fell below basal levels. S1P previously has beenshown to inhibit fibroblast chemotaxis in transwellmigration assays (47, 48). Additional experiments werecarried out to test the effects of S1P on PDGF-stimu-lated migration in nested collagen matrices. Figure 2shows that S1P at 1–10 �M blocked cell migrationstimulated by 50 ng/ml of PDGF, whereas adding LPAor FBS over a wide range of concentrations in combi-nation with PDGF had no effect.
Growth factor specificity of fibroblast migration in2-D after scrape wounding
To learn if the overall pattern of growth factor stimu-lation of migration observed in Fig. 1B was unique tocells in collagen matrices, experiments also were car-ried out using 2-D human fibroblast cultures. Figure 3shows images of cells 24 h after scrape wounding,double-stained with phalloidin for actin and propidiumiodide to identify cell nuclei. It can be seen from thephotomicrographs that PDGF was the most promigra-tory growth factor and S1P appeared to be inhibitorysimilar to our observations for cell migration in nestedcollagen matrices.
Differential effects of S1P and LPA on fibroblastdendritic extensions
Fibroblasts in collagen matrix form dynamic dendriticcell extensions. PDGF stimulates their protrusion,which depends on Rac and PAK1 activation. LPA causestheir retraction, which depends on Rho and Rhokinase/myosin II activation (25, 29, 49). Figure 4Ashows the appearance of fibroblasts in collagen matri-ces after 6 h. Consistent with previous studies, cells inbasal medium form short dendritic extensions thatincrease in size and complexity in the presence ofPDGF but retract in the presence of LPA. Similar toLPA, addition of serum or S1P resulted in retraction ofdendritic extensions. Retraction was Rho kinase depen-dent since addition of the Rho kinase inhibitor Y27632during the last 4 h of the incubations blocked retrac-tion. Endo-1 affected fibroblast dendritic extensionssimilarly as LPA (not shown).
Inhibiting Rho kinase not only blocked retractionbut also permitted dendritic extensions to reprotrude.Reprotrusion in the presence of S1P appeared to beless robust than with LPA or serum in the medium. Toquantify this effect, morphometric measurements weremade to determine dendritic cell index (perimeter2/4��area). Figure 4B shows that dendritic index variedmarkedly, so much so that it was useful to plot theresults on a log scale to help appreciate the differences.Fibroblasts incubated in S1P-containing medium had adendritic index �1–2 consistent with the round appear-ance of the cells (dendritic index1.0 for a round cell),whereas cells in basal medium had a dendritic index�20 and PDGF-treated cells had a dendritic index of100. Cells in LPA and serum-containing medium haddendritic indices �2–4. In the presence of Rho kinaseinhibitor, the dendritic index increased to �40 for LPAand �80 for FBS (�20-fold increases). With Rho kinaseinhibitor and S1P, the dendritic index increased only to�5 (�5-fold increase), confirming that S1P not onlycaused retraction of dendritic extensions but also inter-fered with reprotrusion of dendritic extensions underconditions in which Rho kinase was blocked.
Figure 4C shows that the effects of S1P were com-pletely reversible. When fibroblasts with dendritic ex-tensions that had retracted for 2 h were washed andswitched to PDGF-containing medium, then no differ-ences were observed between cells that had been inLPA or serum compared with S1P. Dendritic extensionsappeared to reprotrude completely in each case.
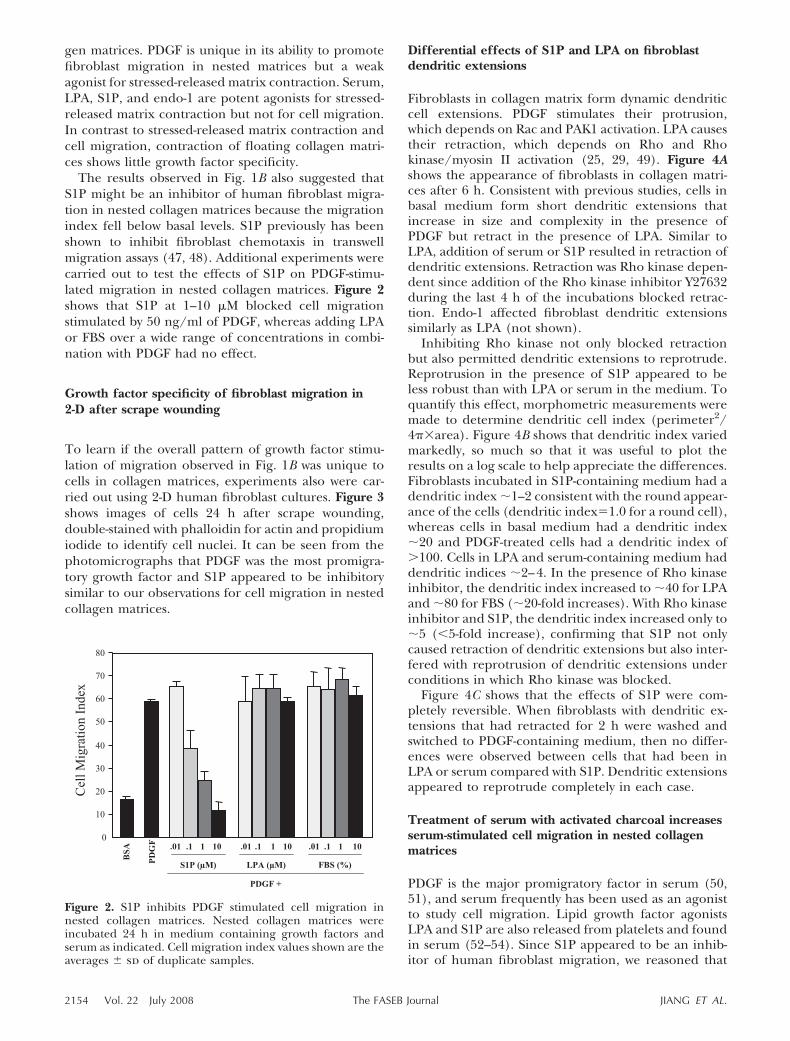
Treatment of serum with activated charcoal increasesserum-stimulated cell migration in nested collagenmatrices
PDGF is the major promigratory factor in serum (50,51), and serum frequently has been used as an agonistto study cell migration. Lipid growth factor agonistsLPA and S1P are also released from platelets and foundin serum (52–54). Since S1P appeared to be an inhib-itor of human fibroblast migration, we reasoned that
Figure 2. S1P inhibits PDGF stimulated cell migration innested collagen matrices. Nested collagen matrices wereincubated 24 h in medium containing growth factors andserum as indicated. Cell migration index values shown are theaverages � sd of duplicate samples.
2154 Vol. 22 July 2008 JIANG ET AL.The FASEB Journal

the presence of S1P in serum might be acting as aninhibitor to reduce serum promigratory activity. Incu-bation of serum with activated charcoal can remove S1Pand other lipid agonists (43), which was the methodused to identify serum lysophospholipids as activatorsof the small G protein Rho (37).
Human and FBS were treated with activated char-
coal. Figure 5A shows that no differences in the Coo-masie blue stained profile of polypeptides could bedetected after charcoal treatment. Figure 5B shows thatactivated charcoal treatment removed the activity fromserum responsible for retraction of dendritic exten-sions, and Fig. 5C shows that charcoal-treated serumhad much greater promigratory activity than control
Figure 3. Growth factor and serum stimulationof fibroblast migration on collagen-coated cov-erslips. Human fibroblast cultures were scrapewounded and incubated 24 h in medium con-taining growth factors and serum as indicatedusing concentrations shown in Fig. 1. At the endof the incubations, samples were fixed andstained for actin and propidium iodide (PI).Scale bar 300 �m.
Figure 4. S1P, LPA, and FBS stimulate retraction of fibroblastdendritic extensions in collagen matrices. A) Collagen matriceswere polymerized 1 h with 2 � 104 cells/matrix. After polymer-ization, samples were incubated 6 h in basal medium and 50ng/ml PDGF, 10 �M LPA, 1 �M S1P, or 10% FBS as indicatedwith 10 �M Y27632 added during the last 4 h where shown. Atthe end of the incubations, samples were fixed and stained foractin. B) Same as in A, except samples were subjected tomorphometric analysis. Dendritic index values shown are theaverages � sd for 20 cells. C) Collagen matrices were polymer-
ized 1 h with 2 � 104 cells/matrix. After polymerization, half the samples were incubated 6 h in basal medium and 1�M S1P, 10 �M LPA, and 10% FBS as indicated. After 2 h, half the samples were washed and switched toPDGF-containing medium. At the end of the incubations, samples were fixed and stained for actin.
2155PROMIGRATORY VS. PROCONTRACTILE ENVIRONMENTS
serum. These findings were consistent with the ideathat lipid agonists in serum (e.g., LPA and S1P) wereresponsible for retraction of dendritic extensions andinhibited human fibroblast migration. Since LPA doesnot inhibit cell migration as shown in Fig. 2, S1P was alikely candidate to explain the migration inhibitioneffect.
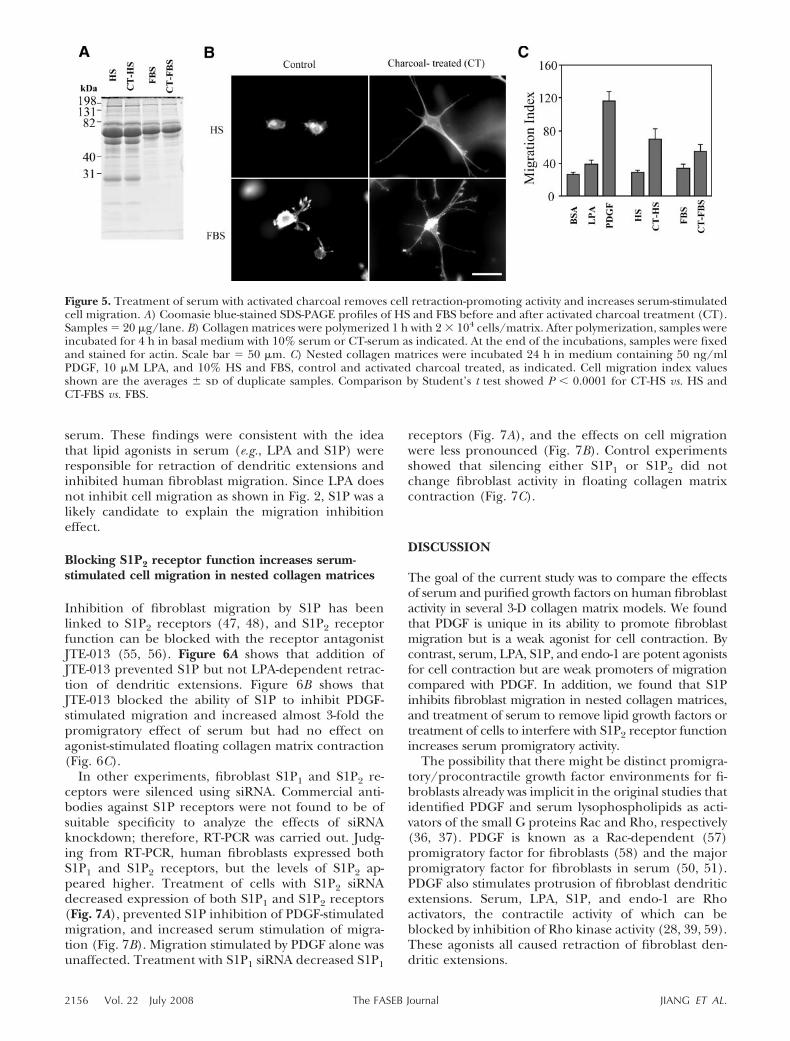
Blocking S1P2 receptor function increases serum-stimulated cell migration in nested collagen matrices
Inhibition of fibroblast migration by S1P has beenlinked to S1P2 receptors (47, 48), and S1P2 receptorfunction can be blocked with the receptor antagonistJTE-013 (55, 56). Figure 6A shows that addition ofJTE-013 prevented S1P but not LPA-dependent retrac-tion of dendritic extensions. Figure 6B shows thatJTE-013 blocked the ability of S1P to inhibit PDGF-stimulated migration and increased almost 3-fold thepromigratory effect of serum but had no effect onagonist-stimulated floating collagen matrix contraction(Fig. 6C).
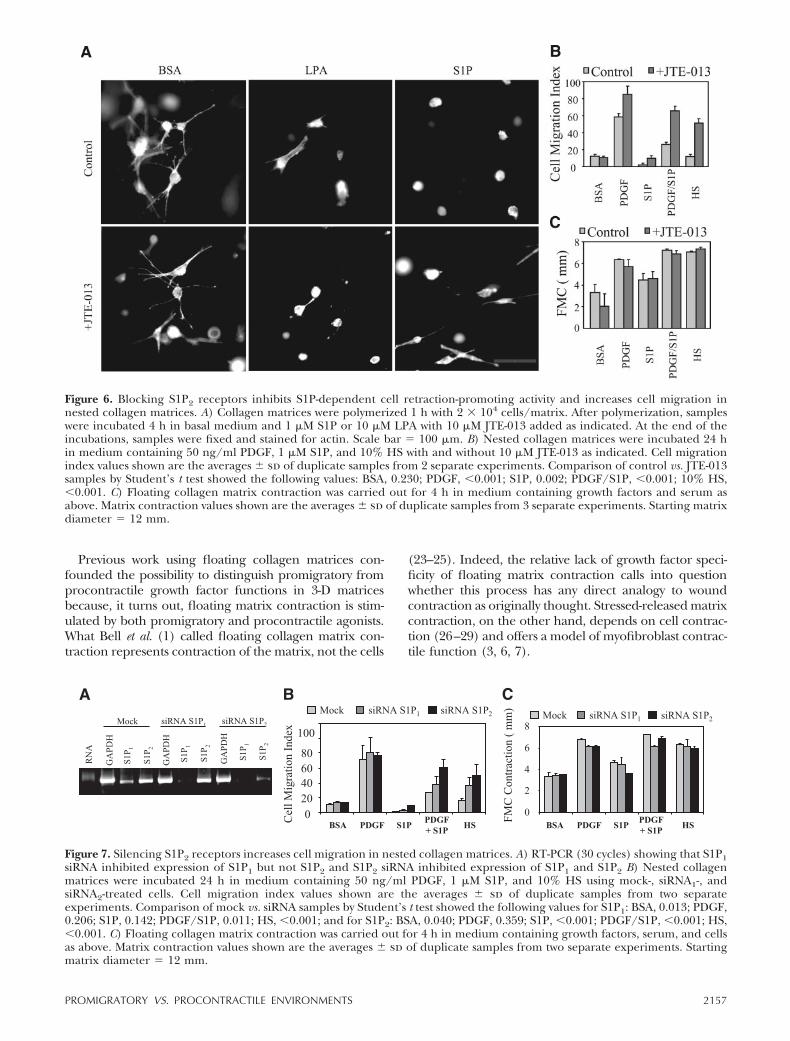
In other experiments, fibroblast S1P1 and S1P2 re-ceptors were silenced using siRNA. Commercial anti-bodies against S1P receptors were not found to be ofsuitable specificity to analyze the effects of siRNAknockdown; therefore, RT-PCR was carried out. Judg-ing from RT-PCR, human fibroblasts expressed bothS1P1 and S1P2 receptors, but the levels of S1P2 ap-peared higher. Treatment of cells with S1P2 siRNAdecreased expression of both S1P1 and S1P2 receptors(Fig. 7A), prevented S1P inhibition of PDGF-stimulatedmigration, and increased serum stimulation of migra-tion (Fig. 7B). Migration stimulated by PDGF alone wasunaffected. Treatment with S1P1 siRNA decreased S1P1
receptors (Fig. 7A), and the effects on cell migrationwere less pronounced (Fig. 7B). Control experimentsshowed that silencing either S1P1 or S1P2 did notchange fibroblast activity in floating collagen matrixcontraction (Fig. 7C).
DISCUSSION
The goal of the current study was to compare the effectsof serum and purified growth factors on human fibroblastactivity in several 3-D collagen matrix models. We foundthat PDGF is unique in its ability to promote fibroblastmigration but is a weak agonist for cell contraction. Bycontrast, serum, LPA, S1P, and endo-1 are potent agonistsfor cell contraction but are weak promoters of migrationcompared with PDGF. In addition, we found that S1Pinhibits fibroblast migration in nested collagen matrices,and treatment of serum to remove lipid growth factors ortreatment of cells to interfere with S1P2 receptor functionincreases serum promigratory activity.
The possibility that there might be distinct promigra-tory/procontractile growth factor environments for fi-broblasts already was implicit in the original studies thatidentified PDGF and serum lysophospholipids as acti-vators of the small G proteins Rac and Rho, respectively(36, 37). PDGF is known as a Rac-dependent (57)promigratory factor for fibroblasts (58) and the majorpromigratory factor for fibroblasts in serum (50, 51).PDGF also stimulates protrusion of fibroblast dendriticextensions. Serum, LPA, S1P, and endo-1 are Rhoactivators, the contractile activity of which can beblocked by inhibition of Rho kinase activity (28, 39, 59).These agonists all caused retraction of fibroblast den-dritic extensions.
Figure 5. Treatment of serum with activated charcoal removes cell retraction-promoting activity and increases serum-stimulatedcell migration. A) Coomasie blue-stained SDS-PAGE profiles of HS and FBS before and after activated charcoal treatment (CT).Samples 20 �g/lane. B) Collagen matrices were polymerized 1 h with 2 � 104 cells/matrix. After polymerization, samples wereincubated for 4 h in basal medium with 10% serum or CT-serum as indicated. At the end of the incubations, samples were fixedand stained for actin. Scale bar 50 �m. C) Nested collagen matrices were incubated 24 h in medium containing 50 ng/mlPDGF, 10 �M LPA, and 10% HS and FBS, control and activated charcoal treated, as indicated. Cell migration index valuesshown are the averages � sd of duplicate samples. Comparison by Student’s t test showed P � 0.0001 for CT-HS vs. HS andCT-FBS vs. FBS.
2156 Vol. 22 July 2008 JIANG ET AL.The FASEB Journal
Previous work using floating collagen matrices con-founded the possibility to distinguish promigratory fromprocontractile growth factor functions in 3-D matricesbecause, it turns out, floating matrix contraction is stim-ulated by both promigratory and procontractile agonists.What Bell et al. (1) called floating collagen matrix con-traction represents contraction of the matrix, not the cells
(23–25). Indeed, the relative lack of growth factor speci-ficity of floating matrix contraction calls into questionwhether this process has any direct analogy to woundcontraction as originally thought. Stressed-released matrixcontraction, on the other hand, depends on cell contrac-tion (26–29) and offers a model of myofibroblast contrac-tile function (3, 6, 7).
Figure 6. Blocking S1P2 receptors inhibits S1P-dependent cell retraction-promoting activity and increases cell migration innested collagen matrices. A) Collagen matrices were polymerized 1 h with 2 � 104 cells/matrix. After polymerization, sampleswere incubated 4 h in basal medium and 1 �M S1P or 10 �M LPA with 10 �M JTE-013 added as indicated. At the end of theincubations, samples were fixed and stained for actin. Scale bar 100 �m. B) Nested collagen matrices were incubated 24 hin medium containing 50 ng/ml PDGF, 1 �M S1P, and 10% HS with and without 10 �M JTE-013 as indicated. Cell migrationindex values shown are the averages � sd of duplicate samples from 2 separate experiments. Comparison of control vs. JTE-013samples by Student’s t test showed the following values: BSA, 0.230; PDGF, �0.001; S1P, 0.002; PDGF/S1P, �0.001; 10% HS,�0.001. C) Floating collagen matrix contraction was carried out for 4 h in medium containing growth factors and serum asabove. Matrix contraction values shown are the averages � sd of duplicate samples from 3 separate experiments. Starting matrixdiameter 12 mm.
Figure 7. Silencing S1P2 receptors increases cell migration in nested collagen matrices. A) RT-PCR (30 cycles) showing that S1P1siRNA inhibited expression of S1P1 but not S1P2 and S1P2 siRNA inhibited expression of S1P1 and S1P2 B) Nested collagenmatrices were incubated 24 h in medium containing 50 ng/ml PDGF, 1 �M S1P, and 10% HS using mock-, siRNA1-, andsiRNA2-treated cells. Cell migration index values shown are the averages � sd of duplicate samples from two separateexperiments. Comparison of mock vs. siRNA samples by Student’s t test showed the following values for S1P1: BSA, 0.013; PDGF,0.206; S1P, 0.142; PDGF/S1P, 0.011; HS, �0.001; and for S1P2: BSA, 0.040; PDGF, 0.359; S1P, �0.001; PDGF/S1P, �0.001; HS,�0.001. C) Floating collagen matrix contraction was carried out for 4 h in medium containing growth factors, serum, and cellsas above. Matrix contraction values shown are the averages � sd of duplicate samples from two separate experiments. Startingmatrix diameter 12 mm.
2157PROMIGRATORY VS. PROCONTRACTILE ENVIRONMENTS

Our observation that S1P inhibits fibroblast migra-tion in nested collagen matrices extends previous workdemonstrating that S1P blocks fibroblast chemotaxis intranswell migration assays (47, 48). S1P interacts withseveral heterotrimeric G-protein receptors (53) andaffects cells differently depending on their complementof S1P receptors (60–63). Inhibitory effects of S1P onfibroblasts have been associated with activation of S1P2receptors (47, 48) coupled to Rho stimulation and Racinhibition. Inhibition of cell migration by S1P cannotsimply be attributed to Rho activation and retraction ofdendritic extensions since neither LPA nor seruminhibited PDGF-stimulated cell migration even thoughboth agonists activate Rho and retraction of dendriticextensions. Direct measurement of Rho and Rac activityof fibroblasts in collagen matrices is technically difficultgiven the low cell number and high concentration ofcollagen with which one has to contend. Nevertheless,our observations on dendritic extensions provide indi-rect support for the Rho activation/Rac inhibition idea.S1P-treated cells were unable to fully protrude den-dritic extensions when Rho kinase activity was blocked,and protrusion is believed to be Rac dependent (25,49).
Inhibition of migration by S1P can explain whyserum has much less effect on human fibroblast migra-tion compared with PDGF even though PDGF is themajor promigratory factor in serum (50, 51). Consis-tent with this possibility, we found that treatment ofserum with activated charcoal, which removes lipidagonists (43), including those responsible for Rhoactivation (37), resulted in an increase in the promi-gratory activity of serum. Similarly, blocking S1P2 re-ceptor function with JTE-013 prevented S1P inhibitionof PDGF-stimulated migration and increased serumpromigratory activity.
Previous work showed that deletion of S1P2 receptorsfrom mouse embryo fibroblasts (MEFs) increased se-rum-stimulated MEF chemotaxis (48). We found thatsilencing S1P2 receptors by siRNA blocked S1P inhibi-tion of PDGF promigratory activity and increased se-rum promigratory activity for the treated cells. In theabsence of specific antibodies, receptor silencing wasdemonstrated by RT-PCR. Based on this method, siRNAsilencing of S1P1 receptors also appeared to increaseserum promigratory activity, an observation that re-quires future explanation. Taken together, the findingssuggest that S1P-stimulated retraction of dendritic ex-tensions and inhibition fibroblast migration depend onthe S1P2 receptor pathway and that the presence of S1Pin serum decreases serum promigratory activity forhuman fibroblasts.
That different sets of growth factors generate promi-gratory vs. procontractile tissue environments for fibro-blasts has important implications with respect to woundrepair. It is now well understood that the dynamicallychanging growth factor environment plays a key role inrepair (64–67). Until recently (68), most general re-views of the repair process have tended to ignore thelipid growth factors. These agonists likely are the key
activators for fibroblast and myofibroblast contraction.Regulation of fibroblast migration by the balance be-tween PDGF and S1P has additional implications. Con-ditions that can interfere with appropriate levels ortiming of growth factors in the wound environmentsuch as loss of proteinase regulation in chronic wounds(69, 70) are believed to result in abnormal repair (71,72). In general, it has been assumed that growth factorsupplementation should improve wound repair by com-pensating for insufficient levels of growth factors (32,33). Replenishment of PDGF will be of little conse-quence, however, if S1P levels in the wound environ-ment are elevated sufficiently so that the environmentis unfavorable for fibroblast migration.
In conclusion, our findings support previous workemphasizing the potential importance of the platelet-derived lipid agonists in wound repair (52, 54, 68, 73).As summarized in Fig. 8, the lipid agonists LPA and S1Pare key factors in determining connective tissue remod-eling processes that involve contractile activity of fibro-blasts (and myofibroblasts). In addition, the balancebetween PDGF and S1P is a key determinant of fibro-blast promigratory activity in the wound environment.Ironically, most studies on the mechanism of fibroblastmigration have been carried out in serum-containingmedium using cells in 2-D culture. Pharmacologicaland other interventions that have been reported toincrease cell migration could have resulted from en-hancement of the PDGF promigratory pathway orinterference with the S1P inhibitory pathway.
We thank W. Snell and M. Petroll for helpful advice andsuggestions. This research was supported by U.S. NationalInstitutes of Health grant GM-31321.
REFERENCES
1. Bell, E., Ivarsson, B., and Merrill, C. (1979) Production of atissue-like structure by contraction of collagen lattices by humanfibroblasts of different proliferative potential in vitro. Proc. Natl.Acad. Sci. U. S. A. 76, 1274–1278
Figure 8. Regulation of fibroblast migration and contractionby serum and the tissue environment. Different sets of growthfactors generate promigratory vs. procontractile tissue envi-ronments for fibroblasts. The balance between PDGF and S1Pis a key determinant of fibroblast promigratory activity. Seetext for other details.
2158 Vol. 22 July 2008 JIANG ET AL.The FASEB Journal

2. Bell, E., Sher, S., Hull, B., Merrill, C., Rosen, S., Chamson, A.,Asselineau, D., Dubertret, L., Coulomb, B., Lapiere, C., Nus-gens, B., and Neveux, Y. (1983) The reconstitution of living skin.J. Invest. Dermatol. 81, 2s–10s
3. Cukierman, E., Pankov, R., and Yamada, K. M. (2002) Cellinteractions with three-dimensional matrices. Curr. Opin. CellBiol. 14, 633–639
4. Grinnell, F. (1994) Fibroblasts, myofibroblasts, and woundcontraction. J. Cell Biol. 124, 401–404
5. Desmouliere, A., Chaponnier, C., and Gabbiani, G. (2005)Tissue repair, contraction, and the myofibroblast. Wound RepairRegen. 13, 7–12
6. Grinnell, F. (2003) Fibroblast biology in three-dimensionalcollagen matrices. Trends Cell Biol. 13, 264–269
7. Tomasek, J. J., Gabbiani, G., Hinz, B., Chaponnier, C., andBrown, R. A. (2002) Myofibroblasts and mechano-regulation ofconnective tissue remodelling. Nat. Rev. Mol. Cell Biol. 3, 349–363
8. Clark, R. A., Ghosh, K., and Tonnesen, M. G. (2007) Tissueengineering for cutaneous wounds. J. Invest. Dermatol. 127,1018–1029
9. Darby, I. A., and Hewitson, T. D. (2007) Fibroblast differentia-tion in wound healing and fibrosis. Int. Rev. Cytol. 257, 143–179
10. Hinz, B. (2007) Formation and function of the myofibroblastduring tissue repair. J. Invest. Dermatol. 127, 526–537
11. Abraham, D. J., Eckes, B., Rajkumar, V., and Krieg, T. (2007)New developments in fibroblast and myofibroblast biology:implications for fibrosis and scleroderma. Curr. Rheumatol. Rep.9, 136–143
12. Paszek, M. J., Zahir, N., Johnson, K. R., Lakins, J. N., Rozenberg,G. I., Gefen, A., Reinhart-King, C. A., Margulies, S. S., Dembo,M., Boettiger, D., Hammer, D. A., and Weaver, V. M. (2005)Tensional homeostasis and the malignant phenotype. CancerCell 8, 241–254
13. Comoglio, P. M., and Trusolino, L. (2005) Cancer: the matrix isnow in control. Nat. Med. 11, 1156–1159
14. Beacham, D. A., and Cukierman, E. (2005) Stromagenesis: thechanging face of fibroblastic microenvironments during tumorprogression. Semin. Cancer Biol. 15, 329–341
15. Barocas, V. H., Moon, A. G., and Tranquillo, R. T. (1995) Thefibroblast-populated collagen microsphere assay of cell tractionforce–Part 2: Measurement of the cell traction parameter.J. Biomech. Eng. 117, 161–170
16. Wakatsuki, T., Kolodney, M. S., Zahalak, G. I., and Elson, E. L.(2000) Cell mechanics studied by a reconstituted model tissue.Biophys. J. 79, 2353–2368
17. Roeder, B. A., Kokini, K., Sturgis, J. E., Robinson, J. P., andVoytik-Harbin, S. L. (2002) Tensile mechanical properties ofthree-dimensional type I collagen extracellular matrices withvaried microstructure. J. Biomech. Eng. 124, 214–222
18. Ahlfors, J. E., and Billiar, K. L. (2007) Biomechanical andbiochemical characteristics of a human fibroblast-produced andremodeled matrix. Biomaterials 28, 2183–2191
19. Silver, F. H., Siperko, L. M., and Seehra, G. P. (2002) Mechano-biology of force transduction in dermal tissue. Skin Res. Tech. 8,1–21
20. Johnson, T. M., Lowe, L., Brown, M. D., Sullivan, M. J., andNelson, B. R. (1993) Histology and physiology of tissue expan-sion. J. Dermatol. Surg. Oncol. 19, 1074–1078
21. Wilhelmi, B. J., Blackwell, S. J., Mancoll, J. S., and Phillips, L. G.(1998) Creep vs. stretch: a review of the viscoelastic propertiesof skin. Ann. Plast. Surg. 41, 215–219
22. Yamada, K. M., and Cukierman, E. (2007) Modeling tissuemorphogenesis and cancer in 3-D. Cell 130, 601–610
23. Ehrlich, H. P., and Rajaratnam, J. B. (1990) Cell locomotionforces versus cell contraction forces for collagen lattice contrac-tion: an in vitro model of wound contraction. Tissue Cell 22,407–417
24. Skuta, G., Ho, C. H., and Grinnell, F. (1999) Increased myosinlight chain phosphorylation is not required for growth factorstimulation of collagen matrix contraction. J. Biol. Chem. 274,30163–30168
25. Rhee, S., and Grinnell, F. (2006) P21-activated kinase 1: conver-gence point in PDGF- and LPA-stimulated collagen matrixcontraction by human fibroblasts. J. Cell Biol. 172, 423–432
26. Mochitate, K., Pawelek, P., and Grinnell, F. (1991) Stressrelaxation of contracted collagen gels: disruption of actin
filament bundles, release of cell surface fibronectin, and down-regulation of DNA and protein synthesis. Exp. Cell Res. 193,198–207
27. Yanase, M., Ikeda, H., Matsui, A., Maekawa, H., Noiri, E.,Tomiya, T., Arai, M., Yano, T., Shibata, M., Ikebe, M., Fujiwara,K., Rojkind, M., and Ogata, I. (2000) Lysophosphatidic acidenhances collagen gel contraction by hepatic stellate cells:association with rho-kinase. Biochem. Biophys. Res. Commun. 277,72–78
28. Parizi, M., Howard, E. W., and Tomasek, J. J. (2000) Regulationof LPA-promoted myofibroblast contraction: Role of Rho, myo-sin light chain kinase, and myosin light chain phosphatase. Exp.Cell Res. 254, 210–220
29. Abe, M., Ho, C. H., Kamm, K. E., and Grinnell, F. (2003)Different molecular motors mediate platelet-derived growthfactor and lysophosphatidic acid-stimulated floating collagenmatrix contraction. J. Biol. Chem. 278, 47707–47712
30. Grinnell, F., Ho, C. H., Lin, Y. C., and Skuta, G. (1999)Differences in the regulation of fibroblast contraction of float-ing versus stressed collagen matrices. J. Biol. Chem. 274, 918–923
31. Grinnell, F., Rocha, L. B., Iucu, C., Rhee, S., and Jiang, H.(2006) Nested collagen matrices: a new model to study migra-tion of human fibroblast populations in three dimensions. Exp.Cell Res. 312, 86–94
32. Davidson, J. M. (2007) Growth factors: the promise and theproblems. Int. J. Low. Extrem. Wounds 6, 8–10
33. Fu, X., Li, X., Cheng, B., Chen, W., and Sheng, Z. (2005)Engineered growth factors and cutaneous wound healing: suc-cess and possible questions in the past 10 years. Wound RepairRegen. 13, 122–130
34. Werner, S., and Grose, R. (2003) Regulation of wound healingby growth factors and cytokines. Physiol. Rev. 83, 835–870
35. Cross, K. J., and Mustoe, T. A. (2003) Growth factors in woundhealing. Surg. Clin. North Am. 83, 531–545
36. Ridley, A. J., Paterson, H. F., Johnston, C. L., Diekmann, D., andHall, A. (1992) The small GTP-binding protein rac regulatesgrowth factor-induced membrane ruffling. Cell 70, 401–410
37. Ridley, A. J., and Hall, A. (1992) The small GTP-binding proteinrho regulates the assembly of focal adhesions and actin stressfibers in response to growth factors. Cell 70, 389–399
38. Tingstrom, A., Heldin, C. H., and Rubin, K. (1992) Regulationof fibroblast-mediated collagen gel contraction by platelet-derived growth factor, interleukin-1 alpha and transforminggrowth factor-beta 1. J. Cell Sci. 102, 315–322
39. Cooke, M. E., Sakai, T., and Mosher, D. F. (2000) Contraction ofcollagen matrices mediated by alpha2beta1A and alpha(v)beta3integrins. J. Cell Sci. 113, 2375–2383
40. Smith, K. D., Wells, A., and Lauffenburger, D. A. (2006)Multiple signaling pathways mediate compaction of collagenmatrices by EGF-stimulated fibroblasts. Exp. Cell Res. 312, 1970–1982
41. Guidry, C., and Hook, M. (1991) Endothelins produced byendothelial cells promote collagen gel contraction by fibro-blasts. J. Cell Biol. 115, 873–880
42. Abe, M., Sogabe, Y., Syuto, T., Yokoyama, Y., and Ishikawa, O.(2007) Evidence that PI3K, Rac, Rho, and Rho kinase areinvolved in basic fibroblast growth factor-stimulated fibroblast-collagen matrix contraction. J. Cell Biochem.
43. Lee, M. J., Van Brocklyn, J. R., Thangada, S., Liu, C. H., Hand,A. R., Menzeleev, R., Spiegel, S., and Hla, T. (1998) Sphin-gosine-1-phosphate as a ligand for the G protein-coupled recep-tor EDG-1. Science 279, 1552–1555
44. Rhee, S., Jiang, H., Ho, C. H., and Grinnell, F. (2007) Microtu-bule function in fibroblast spreading is modulated according tothe tension state of cell-matrix interactions. Proc. Natl. Acad. Sci.U. S. A. 104, 5425–5430
45. Huang, J. S., Huang, S. S., and Deuel, T. F. (1983) Humanplatelet-derived growth factor: radioimmunoassay and discoveryof a specific plasma-binding protein. J. Cell Biol. 97, 383–388
46. Olutoye, O. O., Yager, D. R., Cohen, I. K., and Diegelmann, R. F.(1996) Lower cytokine release by fetal porcine platelets: apossible explanation for reduced inflammation after fetalwounding. J. Pediatr. Surg. 31, 91–95
47. Sugimoto, N., Takuwa, N., Okamoto, H., Sakurada, S., andTakuwa, Y. (2003) Inhibitory and stimulatory regulation of Racand cell motility by the G12/13-Rho and Gi pathways integrated
2159PROMIGRATORY VS. PROCONTRACTILE ENVIRONMENTS

downstream of a single G protein-coupled sphingosine-1-phos-phate receptor isoform. Mol. Cell. Biol. 23, 1534–1545
48. Goparaju, S. K., Jolly, P. S., Watterson, K. R., Bektas, M., Alvarez,S., Sarkar, S., Mel, L., Ishii, I., Chun, J., Milstien, S., and Spiegel,S. (2005) The S1P2 receptor negatively regulates platelet-derived growth factor-induced motility and proliferation. Mol.Cell. Biol. 25, 4237–4249
49. Grinnell, F., Ho, C. H., Tamariz, E., Lee, D. J., and Skuta, G.(2003) Dendritic fibroblasts in three-dimensional collagen ma-trices. Mol. Biol. Cell 14, 384–395
50. Li, W., Fan, J., Chen, M., Guan, S., Sawcer, D., Bokoch, G. M.,and Woodley, D. T. (2004) Mechanism of human dermalfibroblast migration driven by type I collagen and platelet-derived growth factor-BB. Mol. Biol. Cell 15, 294–309
51. Gao, Z., Sasaoka, T., Fujimori, T., Oya, T., Ishii, Y., Sabit, H.,Kawaguchi, M., Kurotaki, Y., Naito, M., Wada, T., Ishizawa, S.,Kobayashi, M., Nabeshima, Y., and Sasahara, M. (2005) Deletionof the PDGFR-beta gene affects key fibroblast functions impor-tant for wound healing. J. Biol. Chem. 280, 9375–9389
52. Yatomi, Y., Igarashi, Y., Yang, L., Hisano, N., Qi, R., Asazuma, N.,Satoh, K., Ozaki, Y., and Kume, S. (1997) Sphingosine 1-phos-phate, a bioactive sphingolipid abundantly stored in platelets, isa normal constituent of human plasma and serum. J. Biochem.121, 969–973
53. Goetzl, E. J., and An, S. (1998) Diversity of cellular receptors andfunctions for the lysophospholipid growth factors lysophosphatidicacid and sphingosine 1-phosphate. FASEB J. 12, 1589–1598
54. Eichholtz, T., Jalink, K., Fahrenfort, I., and Moolenaar, W. H.(1993) The bioactive phospholipid lysophosphatidic acid isreleased from activated platelets. Biochem. J. 291, 677–680
55. Osada, M., Yatomi, Y., Ohmori, T., Ikeda, H., and Ozaki, Y.(2002) Enhancement of sphingosine 1-phosphate-induced mi-gration of vascular endothelial cells and smooth muscle cells byan EDG-5 antagonist. Biochem. Biophys. Res. Commun. 299, 483–487
56. Arikawa, K., Takuwa, N., Yamaguchi, H., Sugimoto, N.,Kitayama, J., Nagawa, H., Takehara, K., and Takuwa, Y. (2003)Ligand-dependent inhibition of B16 melanoma cell migrationand invasion via endogenous S1P2 G protein-coupled receptor.Requirement of inhibition of cellular RAC activity. J. Biol. Chem.278, 32841–32851
57. Anand-Apte, B., Zetter, B. R., Viswanathan, A., Qiu, R. G., Chen,J., Ruggieri, R., and Symons, M. (1997) Platelet-derived growthfactor and fibronectin-stimulated migration are differentiallyregulated by the Rac and extracellular signal-regulated kinasepathways. J. Biol. Chem. 272, 30688–30692
58. Seppa, H., Grotendorst, G., Seppa, S., Schiffmann, E., andMartin, G. R. (1982) Platelet-derived growth factor is chemotac-tic for fibroblasts. J. Cell Biol. 92, 584–588
59. Yee, H. F., Jr., Melton, A. C., and Tran, B. N. (2001) RhoA/rho-associated kinase mediates fibroblast contractile force genera-tion. Biochem. Biophys. Res. Commun. 280, 1340–1345
60. Peters, S. L., and Alewijnse, A. E. (2007) Sphingosine-1-phos-phate signaling in the cardiovascular system. Curr. Opin. Phar-macol. 7, 186–192
61. Takuwa, Y. (2002) Subtype-specific differential regulation ofRho family G proteins and cell migration by the Edg familysphingosine-1-phosphate receptors. Biochim. Biophys. Acta 1582,112–120
62. Watterson, K., Sankala, H., Milstien, S., and Spiegel, S. (2003)Pleiotropic actions of sphingosine-1-phosphate. Prog. Lipid Res.42, 344–357
63. Spiegel, S., English, D., and Milstien, S. (2002) Sphingosine1-phosphate signaling: providing cells with a sense of direction.Trends Cell Biol. 12, 236–242
64. Diegelmann, R. F., and Evans, M. C. (2004) Wound healing: anoverview of acute, fibrotic and delayed healing. Front. Biosci. 9,283–289
65. Martin, P. (1997) Wound healing–aiming for perfect skinregeneration. Science 276, 75–81
66. Aiba-Kojima, E., Tsuno, N. H., Inoue, K., Matsumoto, D.,Shigeura, T., Sato, T., Suga, H., Kato, H., Nagase, T., Gonda, K.,Koshima, I., Takahashi, K., and Yoshimura, K. (2007) Charac-terization of wound drainage fluids as a source of soluble factorsassociated with wound healing: comparison with platelet-richplasma and potential use in cell culture. Wound Repair. Regen. 15,511–520
67. Li, J., Chen, J., and Kirsner, R. (2007) Pathophysiology of acutewound healing. Clin. Dermatol. 25, 9–18
68. Watterson, K. R., Lanning, D. A., Diegelmann, R. F., andSpiegel, S. (2007) Regulation of fibroblast functions by lyso-phospholipid mediators: potential roles in wound healing.Wound Repair. Regen. 15, 607–616
69. Wysocki, A. B., Staiano-Coico, L., and Grinnell, F. (1993)Wound fluid from chronic leg ulcers contains elevated levels ofmetalloproteinases MMP-2 and MMP-9. J. Invest. Dermatol. 101,64–68
70. Grinnell, F., and Zhu, M. (1996) Fibronectin degradation inchronic wounds depends on the relative levels of elastase,alpha1-proteinase inhibitor, and alpha2-macroglobulin. J. In-vest. Dermatol. 106, 335–341
71. Menke, N. B., Ward, K. R., Witten, T. M., Bonchev, D. G., andDiegelmann, R. F. (2007) Impaired wound healing. Clin. Derma-tol. 25, 19–25
72. Chen, W. Y., and Rogers, A. A. (2007) Recent insights into thecauses of chronic leg ulceration in venous diseases and impli-cations on other types of chronic wounds. Wound Repair. Regen.15, 434–449
73. Moolenaar, W. H., van Meeteren, L. A., and Giepmans, B. N.(2004) The ins and outs of lysophosphatidic acid signaling.Bioessays 26, 870–881
Received for publication September 21, 2007.Accepted for publication January 17, 2008.
2160 Vol. 22 July 2008 JIANG ET AL.The FASEB Journal




















