
The Cloning and Characterization of a Murine Secretory Leukocyte Protease Inhibitor cDNA
11
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 232, 687–697 (1997) ARTICLE NO. RC976358 The Cloning and Characterization of a Murine Secretory Leukocyte Protease Inhibitor cDNA 1 Ralph J. Zitnik,* ,2 Jue Zhang,* Mohammed A. Kashem,² Tadahiko Kohno,² David E. Lyons,² Clifford D. Wright,² Eliot Rosen,‡ Itzhak Goldberg,‡ and Adrian C. Hayday§ *Department of Internal Medicine and §Department of Biology, Yale University, New Haven, Connecticut 06520; ²Amgen, Inc., Boulder, Colorado; and ‡Long Island Jewish Medical Center, Bronx, New York Received February 21, 1997 in order to maintain homeostasis and to prevent exces- Human secretory leukocyte protease inhibitor sive tissue destruction. Tight control of serine prote- (hSLPI) is produced by epithelial cells at mucosal sur- ases has been achieved both by the action of inhibitors faces, where it regulates both the neutrophil-mediated that are highly selective in their spectrum of inhibition, inflammation that characterizes inflammatory dis- and by secretion of less selective inhibitors at specific eases, and pathogens themselves via both antiprotease anatomical sites and/or in response to specific signals and ‘‘defensin-like’’ activities. Additionally, hSLPI may (reviewed in (2)). One inhibitor active against a variety regulate other processes such as cutaneous desquama- of proteases is the secretory leukocyte protease inhibi- tion and placental invasiveness. To better understand tor (SLPI), a 12 kDa protein secreted by epithelial cells the primary physiologic roles of SLPI, it will be im- at mucosal surfaces (3,4). SLPI is produced in response portant to establish a genetically tractable animal to cytokines and to the presence of serine proteases model, the most attractive candidate being the mouse. themselves (5,6). hSLPI has been extensively studied In this report, the cloning and characterization of mu- biochemically, and domains of the molecule which con- rine (m) SLPI is described. mSLPI is encoded by a sin- tain residues critical for the binding and inhibition of gle copy gene, and appears structurally highly similar a variety of proteases have been identified (7,8). The to hSLPI. At the same time, significant differences be- tween mSLPI and hSLPI are presented, notably a dif- expression pattern of hSLPI is consistent with a pro- ference in expression pattern, and a structural differ- posed role in vivo in the regulation of inflammatory ence in the protease binding site that correlates with a cell serine proteases. Indeed, the potent antiprotease difference in the spectrum of protease inhibiton. Such activity of hSLPI has prompted preliminary studies of species-specific evolution of this protease inhibitor is its therapeutic utility in emphysema and cystic fibrosis notable given that species-specific structure-function (reviewed in (9)). In addition, it has recently been sug- differences have previously been reported for the a-1 gested that hSLPI may be useful in modulating the antitrypsin family. q 1997 Academic Press airways inflammation which occurs in allergic rhinitis and asthma (10). At present however, many aspects of the biology of SLPI remain unelucidated. While its primary function Serine proteases play an important role in such di- may be the inhibition of inflammatory cell serine prote- verse biological processes as coagulation and fibrinoly- ases in the epithelium, several other potential func- sis, digestion, inflammation, angiogenesis, immunity, tions of SLPI have recently been described including tissue remodeling and wound healing (reviewed in (1)). the regulation of cutaneous desquamation, control of Precise regulation of proteolysis is critical, presumably musculoskeletal inflammation, and the regulation of placental invasiveness (11-14). While these studies have defined possible functions of SLPI, elucidation of 1 The sequence data in this manuscript was presented at The American Thoacic Society International Meeting, New Orleans, LA, the primary physiological role(s) will require animal in May 1996. studies in which the expression of SLPI can be deregu- 2 Address correspondence to Ralph J. Zitnik, M.D., Division of Pul- lated or ablated. In this regard, the capacity to produce monary and Critical Care Medicine, Department of Internal Medi- strains of mice with highly specific genetic modifica- cine, Yale University School of Medicine, LCI 105, 333 Cedar Street, tions, and to study the phenotypic consequences of New Haven, CT, 06520. Fax: (203) 785-3826. email: zitnik@ biomed.med.yale.edu. these alterations in murine models of human disease 0006-291X/97 $25.00 Copyright q 1997 by Academic Press All rights of reproduction in any form reserved. 687
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The Cloning and Characterization of a Murine Secretory Leukocyte Protease Inhibitor cDNA

BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 232, 687–697 (1997)ARTICLE NO. RC976358
The Cloning and Characterization of a Murine SecretoryLeukocyte Protease Inhibitor cDNA1
Ralph J. Zitnik,*,2 Jue Zhang,* Mohammed A. Kashem,† Tadahiko Kohno,† David E. Lyons,†Clifford D. Wright,† Eliot Rosen,‡ Itzhak Goldberg,‡ and Adrian C. Hayday§*Department of Internal Medicine and §Department of Biology, Yale University, New Haven, Connecticut 06520;†Amgen, Inc., Boulder, Colorado; and ‡Long Island Jewish Medical Center, Bronx, New York
Received February 21, 1997
in order to maintain homeostasis and to prevent exces-Human secretory leukocyte protease inhibitor sive tissue destruction. Tight control of serine prote-
(hSLPI) is produced by epithelial cells at mucosal sur- ases has been achieved both by the action of inhibitorsfaces, where it regulates both the neutrophil-mediated that are highly selective in their spectrum of inhibition,inflammation that characterizes inflammatory dis- and by secretion of less selective inhibitors at specificeases, and pathogens themselves via both antiprotease anatomical sites and/or in response to specific signalsand ‘‘defensin-like’’ activities. Additionally, hSLPI may
(reviewed in (2)). One inhibitor active against a varietyregulate other processes such as cutaneous desquama-of proteases is the secretory leukocyte protease inhibi-tion and placental invasiveness. To better understandtor (SLPI), a 12 kDa protein secreted by epithelial cellsthe primary physiologic roles of SLPI, it will be im-at mucosal surfaces (3,4). SLPI is produced in responseportant to establish a genetically tractable animalto cytokines and to the presence of serine proteasesmodel, the most attractive candidate being the mouse.themselves (5,6). hSLPI has been extensively studiedIn this report, the cloning and characterization of mu-biochemically, and domains of the molecule which con-rine (m) SLPI is described. mSLPI is encoded by a sin-tain residues critical for the binding and inhibition ofgle copy gene, and appears structurally highly similara variety of proteases have been identified (7,8). Theto hSLPI. At the same time, significant differences be-
tween mSLPI and hSLPI are presented, notably a dif- expression pattern of hSLPI is consistent with a pro-ference in expression pattern, and a structural differ- posed role in vivo in the regulation of inflammatoryence in the protease binding site that correlates with a cell serine proteases. Indeed, the potent antiproteasedifference in the spectrum of protease inhibiton. Such activity of hSLPI has prompted preliminary studies ofspecies-specific evolution of this protease inhibitor is its therapeutic utility in emphysema and cystic fibrosisnotable given that species-specific structure-function (reviewed in (9)). In addition, it has recently been sug-differences have previously been reported for the a-1 gested that hSLPI may be useful in modulating theantitrypsin family. q 1997 Academic Press airways inflammation which occurs in allergic rhinitis
and asthma (10).At present however, many aspects of the biology of
SLPI remain unelucidated. While its primary functionSerine proteases play an important role in such di-may be the inhibition of inflammatory cell serine prote-verse biological processes as coagulation and fibrinoly-ases in the epithelium, several other potential func-sis, digestion, inflammation, angiogenesis, immunity, tions of SLPI have recently been described includingtissue remodeling and wound healing (reviewed in (1)). the regulation of cutaneous desquamation, control ofPrecise regulation of proteolysis is critical, presumably musculoskeletal inflammation, and the regulation ofplacental invasiveness (11-14). While these studieshave defined possible functions of SLPI, elucidation of1 The sequence data in this manuscript was presented at The
American Thoacic Society International Meeting, New Orleans, LA, the primary physiological role(s) will require animalin May 1996. studies in which the expression of SLPI can be deregu-
2 Address correspondence to Ralph J. Zitnik, M.D., Division of Pul- lated or ablated. In this regard, the capacity to producemonary and Critical Care Medicine, Department of Internal Medi- strains of mice with highly specific genetic modifica-cine, Yale University School of Medicine, LCI 105, 333 Cedar Street,
tions, and to study the phenotypic consequences ofNew Haven, CT, 06520. Fax: (203) 785-3826. email: [email protected]. these alterations in murine models of human disease
0006-291X/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
687
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
generacy, and I indicates inosine. PCR was performed in 60mM Triswill be a powerful means of understanding gene func-(pH 8.5), 15mM (NH4)SO4, 2.0mM MgCl2 and 2.5mM deoxyNTPs.tion. To this end, the cloning of a murine SLPI cDNA,The template was 1 ml of the reverse transcription reaction describedand the biochemical characterization of recombinant above. Amplifications were performed in a 50 ml reaction mix with
murine SLPI protein are presented in this report. 12.5 pmol of each primer, and 1 Unit Amplitaq polymerase. ThePCR reaction conditions were as follows: 5* denaturation (947C), 309annealing (427C), 1* extension (657C); followed by 35 cycles of 1*
MATERIALS AND METHODS denaturation (947C), 309 annealing (427C), and 1* extension (657C).Under these conditions, an amplified product of 53 bp was produced,which was subcloned into the pGEM-T vector (Promega) and se-Materials and reagents. Pyro-glutamyl-prolyl-valyl-(p)-nitroani-quenced using Sequenase (USB/Amersham, Arlington Heights, IL),lide (pyroGlu-Pro-Val-pNA) was obtained from Chromogenix (Frank-according to the manufacturer’s instructions. The DNA sequence re-lin, Ohio). Trasylol (aprotinin) was purchased from Pentexvealed that the portion of the PCR product amplified between the(Kankakee, IL). Human neutrophil elastase and human neutrophilprimers indeed encoded the appropriate segment of the protein se-cathepsin G were purchased from Calbiochem (La Jolla, CA).quence derived above.N-succinyl-alanyl-alanyl-prolyl-phenylalanyl-(p)-nitroanilide (N-suc-
A cDNA library, constructed by reverse transcription of r3T3Ala-Ala-Pro-Phe-pNA), and N-tosyl-L-phenyl-alanine-chloromethyl-mRNA using oligo d(T) priming, was directionally cloned into theketone (TPCK)-treated bovine pancreatic trypsin were from SigmaZAP Express l vector (Stratagene) at EcoRI and Xho I sites, and(St. Louis, MO). N-a-benzoyl-L-arginyl-(p)-nitroanilide (N-a-benzoyl-was maintained in the XL-1 Blue MRF* host strain according to theL-Arg-pNA), avian myeloblastosis virus (AMV) reverse tran-manufacturer’s instructions. cDNA clones were isolated by sequen-scriptase, and bovine pancreatic chymotrypsin were from Boeh-tial plaque purification according to standard techniques using as aringer-Mannheim Biochemicals (Indianapolis, IN). Recombinant hu-probe the cloned rt-PCR product described above (16). Library hy-man secretory leukocyte protease inhibitor was from R&D Systemsbridizations were performed at 607C in 61SSC, 51Denhardt’s solu-(Minneapolis, MN); Amplitaq polymerase was from Perkin-Elmer-tion, and 0.5% SDS, and washed in 21SSC, 0.1% SDS at 507C. ForCetus (Norwalk, CT); RNAsin was from Promega (Madison, WI), andeach positive clone, single-stranded filamentous phage were pro-restriction endonucleases and other DNA-modifying enzymes wereduced from the ZAP Express l vector by co-infection of the XL-1-from New England Biolabs (Beverly, MA). Geneticin (G418), andBlue MRF* host with ExAssist helper phage (Stratagene). Doubleother tissue culture antibiotics and additives were from GIBCO/BRLstranded pBK-CMV plasmid DNA was recovered after infection of(Gaithersberg, MD), and serum was either from Hyclone (Logan,the XLOLR strain (Stratagene) with single-stranded filamentousUT), or from Gemini Bio-Products (Calabasas, CA). Chinese hamsterphage, according to manufacturer’s instructions. Inserts were eitherovary (CHO) cells, line K1 were from American Type Culture Collec-sequenced directly within pBK-CMV, or were subcloned to pBlue-tion (Rockville, MD), and ras-transformed 3T3 cells, strain D4 (r3T3),script KS II (Stratagene) and then sequenced.were a gift from Dr Ermanno Gherardi (Imperial Cancer Research
Fund, London, UK). PCR and sequencing primers were synthesized Multiple species genomic southern analysis. A nylon membraneby the W.M. Keck Foundation Biotechnology Resource Core Facility blotted with 4 mg of EcoRI-digested genomic DNA from human, lowerat Yale University (New Haven, CT), while peptide sequencing and primate (Rhesus monkey), mouse (Balb/c), rat (Sprague-Dawley),synthesis was performed by the Laboratory for Macromolecular rabbit, cow, dog, chicken, and yeast (S. cerevisiae) was obtained fromAnalysis, Albert Einstein College of Medicine (Bronx, NY). Clontech (Palo Alto, CA), and was hybridized (at 657C in 51SSPE,
101Denhardt’s solution, 2% SDS, 10 mg/ml sonicated salmon spermCloning of the mSLPI cDNA. The mSLPI protein was isolatedDNA) with full length mSLPI cDNA (a32P-dATP -labeled using theduring an attempt to purify ‘‘scatter factor-inducing factor’’ (SFIF)Klenow fragment of E. coli DNA polymerase after random hexamerfrom the conditioned media (CM) of r3T3 cells. Purification includedpriming), and washed at 607C to a maximum stringency of 21SSC,sequential cation exchange chromatography, heparin sepharose af-0.05% SDS. Autoradiography was performed at 0807C for varyingfinity chromatography, and FPLC as described (15), producing a sin-periods of time using intensifying screens.gle major protein species that migrated at approximately 14 kDa and
12 kDa in SDS-PAGE under non-reducing and reducing conditions, Analysis of tissue expression of SLPI mRNA. A murine multiplerespectively; plus several minor bands (15). Edman degradation of tissue northern blot was obtained (Clontech) which contains 1-2 mgthe major band revealed: (A/G)KNDAIKIGA(C/W)PAKKPAQ(C/ per lane of poly A-enriched RNA isolated from various tissues ofW)L(I/K)LE(I/K)PQ, as an N-terminal sequence (residues in paren- Balb/C mice. The membrane was hybridized (at 427C in 51SSPE,theses indicate sequencing uncertainties). 101 Denhardt’s solution, 50% deionized formamide, 2% SDS and 10
To clone the cDNA encoding this peptide sequence, r3T3 cells were mg/ml sonicated salmon sperm DNA) with the full length mSLPIused as a source of RNA. r3T3 cells were maintained in Dulbecco’s cDNA (labeled as described above), and was washed at 507C to amodified Eagle’s medium supplemented with 100 U/ml penicillin G, maximum stringency of 0.21SSC, 0.1% SDS. To assess equivalency100 mg/ml streptomycin, 2mM L-glutamine, and 10% (v/v) heat-inac- of loading between lanes, the membrane was stripped and reprobedtivated fetal calf serum. Confluent cell monolayers were washed with with a cDNA encoding human b-actin (Clontech) that detects a 2.0PBS, and mRNA isolated by guanadinium isothiocyanate extraction kb message in all murine tissues, along with varying amounts of anand oligo-d(T) cellulose chromatography, according to the manufac- additional 1.8 kb message.turer’s instructions (Stratagene, La Jolla, CA). mRNA was reverse In other experiments tissue was harvested from normal Balb/c ortranscribed in 50mM Tris-HCl (pH 8.3), 10mM MgCl2, 10mM DTT, C57/BL.6 mice, and homogenized in Trizol reagent (GIBCO/BRL).and 4mM sodium pyrophosphate in a 20 ml reaction volume with 1- Total RNA was extracted, poly (A) fractions purified by oligo d(T)2 mg mRNA template, 10 U RNAsin, 3 mg random hexamers, mixed cellulose chromatography (Qiagen, Chatsworth, CA), and northerndeoxynucleotides each at a final concentration of 1.25mM, and 36 analysis performed as previously described (17).units of AMV reverse transcriptase at 427C for 30 minutes.
An attempt to amplify the portion of the cDNA encoding residues Eukaryotic expression of mSLPI cDNA. The full length mSLPIopen reading frame was cloned into the pcDNA 3 eukaryotic expres-2-19 was directed by degenerate primer pools synthesized on the
basis of the N-terminal amino acid sequence above (Figure 1A). sion vector (Invitrogen, San Diego, CA), which also contains a neomy-cin resistance gene to allow for selection of transfectants.The upstream pool was: 5*AA(A/G)AA(C/T)GA(C/T)GCIATIAA(A/
G)AT3*; and the downstream pool was: 5*CA(C/T)TGIGCIGG(C/ CHO cells were maintained in F12 media supplemented with 100U/ml penicillin G, 100 mg/ml streptomycin, 2mM L-glutamine, andT)TT(C/T)TT3*, where residues in parentheses indicate primer de-
688
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
10% (v/v) heat-inactivated fetal calf serum. Cells were transfected Protease inhibition assays. Elastase activity was determined bymeasuring the amidolytic effect of human neutrophil elastase onwith plasmid DNA by calcium phosphate co-precipitation according
to standard methods (16). Stable transfectants were selected by the chromogenic substrate pyroGlu-Pro-Val-pNA (20). The assay wasperformed in duplicate in 96 well plates, in a total reaction volumegrowth in the neomycin analog G418 at 1000 mg/ml, and individual
clones isolated, expanded, and maintained in 400 mg/ml G418. Se- of 200 ml. Elastase (7.5 nM) was incubated either alone, or withvarying concentrations of human or murine SLPI for 15 minutes atrum-free conditioned media collected from stably transfected CHO
clones was centrifuged to remove cells and debris, and concentrated 377C, in 100mM Tris-HCl (pH 8.3), 0.96 M NaCl, 1% BSA. The sub-strate (0.3 mM) was added, and the residual activity in each wellusing a centrifugal concentrator with a 5kDa size exclusion mem-
brane (Millipore, Bedford, MA). Protein concentrations were mea- measured as change in absorbance at 405 nm after a 20 minuteincubation at 377C.sured using a modified Bradford method (BioRad, Hercules, CA).
Trypsin activity was determined by measuring the amidolytic ef-Prokaryotic expression of mSLPI cDNA, and purification of mSLPIfect of TPCK-treated bovine pancreatic trypsin on the chromogenicprotein. The mSLPI cDNA was modified for prokaryotic expressionsubstrate N-a-benzoyl-L-Arg-pNA (21). The assay was performed inusing a PCR approach. A plasmid containing the full-length mSLPIduplicate in 96 well plates in a total volume of 200 ml. Trypsin (45open reading frame used as a template was amplified with an up-nM) was incubated either alone, or with varying concentrations ofstream primer 5*- GTACGTCCATGGGCAAAAATGAT-3*, and ahuman or murine SLPI for 15 minutes at 377C in 50 mM Tris-HCldownstream primer 5*-TGGAATTCTTATCACATCGGGGG-3*, that(pH 7.8), 20 mM CaCl2. The substrate (0.5 mM) was then added,introduced several modifications to the mSLPI open reading frame:and the residual activity in each well was measured as change inthe signal peptide is removed, and a methionine initiator codon isabsorbance at 405 nm after a 20 minute incubation at 377C.introduced upstream from the codon encoding the glycine residue at
Chymotrypsin activity was determined by measuring the amidoly-the N-terminus of the mature secreted mammalian protein; an Ncotic effect of bovine pancreatic chymotrypsin on the chromogenic sub-I cloning site is introduced surrounding the initiator ATG, and astrate N-suc-Ala-Ala-Pro-Phe-pNA (22). The assay was performed inTAA terminator codon and EcoRI cloning site are introduced at theduplicate in 96- well plates in a total volume of 200 ml. Chymotrypsinend of the reading frame. The amplified product was subcloned and(12.5 nM) was incubated either alone, or with varying concentrationssequenced completely in both directions, and then subcloned to aof human or murine SLPI for 15 minutes at 377C in 100 mM Tris-prokaryotic expression vector driven by a synthetic promoter. E. coliHCl (pH 7.8), 10 mM CaCl2. The substrate (0.1 mM) was then added,strain FM15 was transformed with the mSLPI expression vector,and the residual activity in each well was measured as the changeand grown in a 40 liter fermentation system at 377C until an OD ofin absorbance at 405 nm after a 20 minute incubation at 377C.10 was reached. mSLPI expression was induced by addition of 0.5
Cathepsin G activity was determined by measuring the amidolyticmM IPTG for a period of 5 hours. Initial experiments revealed thateffect of human neutrophil cathepsin G on the chromogenic substrateunder these conditions mSLPI accounted for approximately 5% ofN-suc-Ala-Ala-Pro-Phe-pNA (23). The assay was performed in dupli-total cellular protein, and that 50% of the expressed protein wascate in 96-well plates in a total reaction volume of 200 ml. Cathepsinpresent in the soluble fraction (data not shown). Active recombinantG (16 nM) was incubated either alone, or with varying concentrationsmSLPI was then refolded and purified using previously publishedof human or murine SLPI for 15 minutes at 377C in 625 mM Tris-procedures (18,19). The purity of the resulting preparations was as-HCl (pH 7.5), 2.5 mM MgCl2, 0.125% Brij 35. The substrate (0.4 mM)sessed by SDS-PAGE.was then added, and residual activity in each well was measured as
Isolation of murine bronchoalveolar lavage fluid. C57/BL.6 mice the change in absorbance at 405 nm after a 15 minute at 377C.(maintained under specific pathogen-free conditions) were euthan-ized, and the trachea surgically isolated under sterile conditions, Determination of inhibition constants. The inhibitory effect of hu-cannulated with a 21G polypropylene catheter, and tied. A 2ml ali- man and murine SLPI against each proteolytic enzyme was analyzedquot of sterile PBS was instilled, and gently aspirated with a syringe. by calculating the inhibition constant (Ki) from the antiprotease de-Lavage fluid was centrifuged for 1 minute @ 14,000 g, 47C to remove terminations. The Ki was calculated assuming behavior as a tight-cells and debris, and stored at 0807C. Aliquots were thawed, concen- binding inhibitor by the equation:trated 25-50 fold using a centrifugal microconcentrator with a 5kDasize exclusion membrane (Millipore), and the protein concentration
vi/v0Å [((K*i / [I]/ [E]0) 20 4[I]0 [E]0)1/20 (K*i / [I]00 [E]0)]/2 [E]0measured before use.Immunoblotting of mSLPI protein. The peptide (A/G)KNDAIK-
IGACPAKKPAQC was synthesized and conjugated to keyhole limpet where K*i is the apparent inhibition constant, vi and v0 are the inhib-hemocyanin (KLH), injected subcutaneously into New Zealand White ited and uninhibited rates, and [I]0 and [E]0 are the total concentra-Rabbits along with complete Freund’s adjuvant, and booster immuni- tions of inhibitor and enzyme, respectively (24). The active site con-zations were performed in incomplete Freund’s adjuvant at regular centration of elastase, chymotrypsin, and cathepsin G were deter-intervals. All imunoblotting studies were performed with whole se- mined by titration with rhSLPI (3,25). The active site concentrationrum isolated from these animals. of trypsin was determined by titration with aprotinin (26). The KiProteins were separated by SDS-PAGE under reducing conditions
values were obtained by correcting the K*i values for the effect ofthrough 15% acrylamide gels. Prestained molecular weight stan-substrate concentration according to (27):dards (Biorad) were included. Proteins were electrophoretically
transferred to PVDF membranes (Immobilon-P, Millipore) in 25mMTris, 192mM glycine, 10% methanol, at 100V for 60 minutes at 47C. Ki Å IC50/(1 / [S]/Km).Membranes were blocked overnight at 47C in TTBS (100mM Tris-HCl (pH 7.5), 0.9% NaCl, 0.1% (w/v) Tween 20) in 3% (w/v) bovineserum albumin (BSA Fraction V, Sigma), washed in TTBS, and incu- Sequence and database analysis. The Swissprot, PIR, EMBL, and
GenBank molecular biology databases were searched using thebated in a 1:500 dilution of primary antibody in TTBS with 0.1% (w/v) BSA for 1 hour at room temperature before washing again in TTBS BLAST network service (National Center for Biotechnology Informa-
tion) and the FASTA program from the Genetics Computer Groupand incubation in a 1:250 dilution of biotinylated anti-rabbit IgG(Vector Laboratories, Burlingame, CA) for 30 minutes at room tem- (GCG) Wisconsin Sequence Analysis Software Package (University
of Wisconsin). Sequence analysis and multiple sequence alignmentsperature. After another wash in TTBS, membranes were incubatedin the Vectastain-ABC Elite reagent, washed, and developed using were performed using the GCG and MacDNASIS (Hitachi) sequence
analysis programs. The sequence information in this manuscript hasthe DAB substrate kit according to the manufacturer’s instructions.
689
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
been submitted to the GenBank database, and is listed under theaccession number (submitted).
RESULTS
The mSLPI cDNA Sequence and PredictedTranslation Product
The protein sequence for putative scatter factor-in-ducing factor described in Materials and Methods wasthe basis for a degenerate oligomer pool used to isolatea murine cDNA probe (Figure 1A). The sequence ofthe probe was in close agreement with the remainingsequence of the peptide, and was used to screen a cDNAlibrary created from r3T3 cDNA. On primary screeningof 0.5 1 106 plaques, 70 positive clones were identified.From this group the 35 most strongly hybridizingclones were selected and taken through tertiary screen-ing, after which 15 positive clones remained. Restric-tion digestion and end sequencing revealed that allclones contained the same open reading frame, andthree such clones were sequenced fully in both direc-tions (Figure 1B). The majority of the clones containedthe same 671 bp cDNA, which corresponded to the ma-jor mSLPI message observed in northern analysis ofr3T3 mRNA (data not shown). However one of thecDNA clones had an additional 350 bp of sequence lo-cated upstream of the beginning of the open readingframe. The additional 5* sequence is present in contigu-
FIG. 1. Cloning of the murine SLPI cDNA and comparison ofity within the mSLPI gene upstream of the major tran-the translation product to the human and porcine proteins. (A) Thescription start site, which indicates that this clone maystrategy used to produce a mSLPI cDNA probe is shown. The mSLPIrepresent the product of transcription from a minor protein sequence and corresponding cDNA sequence are shown. The
upstream alternative start site within the 5*-regula- upstream and downstream degenerate primer pools are shown intory region of the mSLPI gene (Zitnik RJ, et al., manu- lowercase. Residues at which degeneracy was introduced are indi-
cated by the presence of two nucleotides at that position, and thescript in preparation).position of inosine residues is indicated by ‘‘i’’. (B). Nucleotide se-The longest open reading frame is preceded by a con-quence of the mSLPI cDNA is shown, with the amino acid sequencesensus sequence (ACCATG) for mammalian transla- of the predicted translation product listed below the nucleotide se-
tion initiation as described by Kozak (28). The open quence. An asterisk indicates the position of the stop codon. Numbersin plainface refer to the nucleotide sequence, and boldface numbersreading frame encodes a predicted translation productrefer to the amino acid sequence. The underlined amino acid residues131 amino acids in length, and the stop codon is fol-constitute the signal peptide. (C). Multiple sequence alignment oflowed by a 265 bp 3*-untranslated region. The sequencethe human (h), porcine (p), and murine (m) SLPI proteins. For each
of the translation product is in good agreement with the protein, residues shown in lowercase designate the signal peptides,original protein sequence data. Since peptide sequence and residues shown in uppercase constitute the mature secreted pro-
teins. Underlined residues represent either putative (murine anddata used to create the probe was generated from mate-porcine), or demonstrated (human) serine protease binding sites.rial present in cell supernatants, the N-terminal resi-Closed circles indicate the positions of cysteine residues which aredue in the mature secreted protein can be assigned conserved between the three proteins.
to the glycine at position 26, and residues 1-25 likelyconstitute a hydrophobic signal peptide (Figure 1B).The predicted Mr of the full-length translation productis 14,308, while that of the secreted protein is 11,696. cDNA a strong candidate for the product of the murine
SLPI gene. At the amino acid level, mSLPI was 58%Comparison with published sequences revealed thestrongest match of the sequence to be with human and identical to hSLPI, and 61% identical to pSLPI. The
three proteins show a high degree of homology in theporcine (p) SLPI proteins (3,29,30). The sequences fromthe three species are compared in a multiple sequence signal peptide, with 80% identity between the human
and murine, and an 84% identity between the porcinealignment (Figure 1C). The striking organizationalsimilarity and sequence homology renders this murine and murine amino acid sequences. In addition, there
690
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
TABLE I gene (29). No bands were seen in DNA from chickenor yeast, even after lengthy exposures.Interspecies Differences in the Primary Structure
of SLPI Protease Binding Sites*Tissue Expression of mSLPI mRNA
Residueposition Human Porcine Murine In order to determine whether the expression pattern
of the mSLPI gene was similar to that observed in otherP5 Tyr Tyr Gln species, we analyzed the level of mSLPI mRNA in nor-P4 Gly Gly Ala
mal murine tissues. An mRNA of approximately 0.7P3 Gln Gln Argkbp was expressed strongly in lung and spleen, withP2 Cys Cys Cys
P1 Leu Met Met much lower levels observed in kidney, liver, and testis.P1* Met Met Met The expression in spleen contrasts strongly with theP2* Leu Leu Leu expression pattern in human (see Discussion). In car-P3* Asn Asn Asn
diac and skeletal muscle, only trace amounts of expres-P4* Phe Phe Phesion were seen. No expression was observed in brainP5* Phe Phe Phetissue, even after lengthy exposures (Figure 3). In other
* The amino acid sequence of the known human (h), and putative similar experiments comparing the mSLPI mRNA inporcine (p) and murine (m) SLPI protease binding sites in shown. murine tissues with that expressed in r3T3 cells, anThe assignment of protease inhibitor binding site positions (P5-P*5)
mRNA fragment migrating at an identical position wasis based on (8). The positions of residues where mSLPI and pSLPIobserved (data not shown).residues differ are underlined in boldface.
Eukaryotic Expression of mSLPI cDNA
is complete conservation of the position of cysteine resi- To investigate the protein encoded by the cloned cDNA,dues between the three mature proteins. Furthermore, CHO cells were stably transfected either with an expres-analysis of the mSLPI protein sequence reveals that sion vector carrying the mSLPI open reading frame, orthe N-terminal half of the protein is approximately 30% with an empty expression vector as a negative control.homologous to the C-terminal half, similar to hSLPI Transfectants were screened for mSLPI mRNA expres-(3). Mutagenesis and crystallographic studies have es- sion by northern analysis using the full length cDNA astablished that the residues surrounding the leucine in a probe. No SLPI message was detected in untransfectedposition 72 of the mature hSLPI protein (position 96 ofthe unprocessed protein sequence) comprise the serineprotease binding site (7,8). Leu72 itself constitutes theprimary peptide (P1) residue which interacts with theserine protease primary substrate binding site (S1);and the amide downstream of Leu72 is the scissile bondwhich would be hydrolyzed in a cleavable substrate(24). The corresponding region of the predicted mSLPIprotein shows a high degree of conservation withhSLPI, the important exception being the presence of amethionine residue in the murine protein at a positionanalogous to the Leu72 in hSLPI (Figure 1C, Table I).
The mSLPI Gene Is Present in Single Copy
To determine whether mSLPI mRNA is the productof a single gene or a family of closely related genes,mSLPI cDNA was hybridized to EcoRI-digested geno-mic DNA from a variety of species (Figure 2). ThecDNA probe hybridized strongly with a murine geno-mic fragment of approximately 3.3 kbp. There wasstrong cross-hybridization with a number of frag-ments from rat genomic DNA, and weak cross-hy-
FIG. 2. Multiple Species Southern Analysis. A membrane con-bridization with fragments from human, monkey,taining EcoRI-digested genomic DNA from various species as indi-dog, cow, and rabbit. The single band observed in cated was probed with mSLPI cDNA. The position of DNA molecular
human genomic DNA was about 3.8 kb in size, consis- weight standards is shown at left. The position of the 3.8 kbp EcoRI human (h) fragment is indicated by an arrow at left.tent with the reported structure of the human SLPI
691
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC
Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
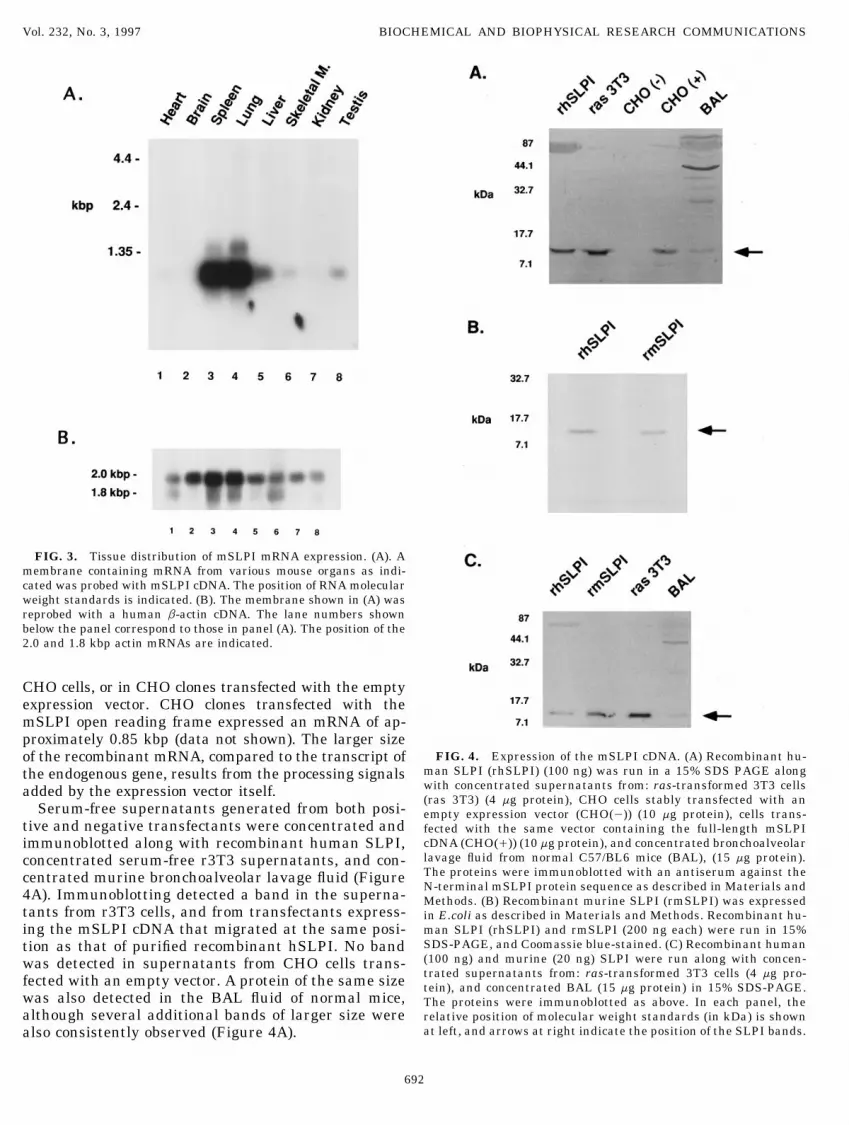
FIG. 3. Tissue distribution of mSLPI mRNA expression. (A). Amembrane containing mRNA from various mouse organs as indi-cated was probed with mSLPI cDNA. The position of RNA molecularweight standards is indicated. (B). The membrane shown in (A) wasreprobed with a human b-actin cDNA. The lane numbers shownbelow the panel correspond to those in panel (A). The position of the2.0 and 1.8 kbp actin mRNAs are indicated.
CHO cells, or in CHO clones transfected with the emptyexpression vector. CHO clones transfected with themSLPI open reading frame expressed an mRNA of ap-proximately 0.85 kbp (data not shown). The larger size
FIG. 4. Expression of the mSLPI cDNA. (A) Recombinant hu-of the recombinant mRNA, compared to the transcript ofman SLPI (rhSLPI) (100 ng) was run in a 15% SDS PAGE alongthe endogenous gene, results from the processing signalswith concentrated supernatants from: ras-transformed 3T3 cellsadded by the expression vector itself.(ras 3T3) (4 mg protein), CHO cells stably transfected with an
Serum-free supernatants generated from both posi- empty expression vector (CHO(0)) (10 mg protein), cells trans-tive and negative transfectants were concentrated and fected with the same vector containing the full-length mSLPI
cDNA (CHO(/)) (10 mg protein), and concentrated bronchoalveolarimmunoblotted along with recombinant human SLPI,lavage fluid from normal C57/BL6 mice (BAL), (15 mg protein).concentrated serum-free r3T3 supernatants, and con-The proteins were immunoblotted with an antiserum against thecentrated murine bronchoalveolar lavage fluid (FigureN-terminal mSLPI protein sequence as described in Materials and4A). Immunoblotting detected a band in the superna- Methods. (B) Recombinant murine SLPI (rmSLPI) was expressed
tants from r3T3 cells, and from transfectants express- in E.coli as described in Materials and Methods. Recombinant hu-man SLPI (rhSLPI) and rmSLPI (200 ng each) were run in 15%ing the mSLPI cDNA that migrated at the same posi-SDS-PAGE, and Coomassie blue-stained. (C) Recombinant humantion as that of purified recombinant hSLPI. No band(100 ng) and murine (20 ng) SLPI were run along with concen-was detected in supernatants from CHO cells trans-trated supernatants from: ras-transformed 3T3 cells (4 mg pro-fected with an empty vector. A protein of the same size tein), and concentrated BAL (15 mg protein) in 15% SDS-PAGE.
was also detected in the BAL fluid of normal mice, The proteins were immunoblotted as above. In each panel, thealthough several additional bands of larger size were relative position of molecular weight standards (in kDa) is shown
at left, and arrows at right indicate the position of the SLPI bands.also consistently observed (Figure 4A).
692
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
a potent inhibitor of a variety of proteases includingProduction of Recombinant mSLPI andelastase, cathepsin G, trypsin, chymotrypsin and mastDemonstration of Its Antiprotease Activitycell chymase (3,7,25,31,32). Based on comparison of the
In order to produce mSLPI in sufficient quantity and relative association rate constants and inhibition con-purity to undertake studies of its antiprotease function, stants that characterize the interaction between hu-we expressed recombinant mSLPI protein in E. coli. man SLPI and these proteases, it has been assumedThe mSLPI open reading frame was modified to yield that the major physiologic substrate for SLPI is neutro-a translation product in E. coli corresponding to the phil elastase. SLPI is present in large amounts in vari-mature secreted mammalian protein. However, for the ous mucosal secretions including tears, saliva, sputum,SLPI protein to function, correct folding and disulfide tracheal secretions, cervical fluid, and seminal fluid,bond formation must occur. Based on the high degree of where it is conventionally thought to play an importantconservation of both primary and predicted secondary role in the defense of epithelial surfaces against proteo-structure between the murine and human proteins, we lytic damage ((3,33-37), and references therein).employed a strategy of expression, in vitro refolding However, it has recently been appreciated that theand purification used previously to produce functional biology of SLPI is considerably more complex. SLPIhSLPI in E. coli. (18,19). The purified product was 95% is expressed in human articular and costal cartilage,pure when analyzed by SDS-PAGE and stained with articular meniscus, and intervertebral disc tissue; andCoomassie blue (Figure 4B). Recombinant murine and studies in vitro have demonstrated constitutive expres-human SLPI were immunoblotted along with serum- sion of SLPI by normal human articular chondrocytesfree concentrated r3T3 supernatants, and BAL fluid and annulus fibrosus fibrochondrocytes in primary cul-from normal mice (Figure 4C). An identical band of ture (11,38,39). Thus SLPI has been hypothesized toappropriate size was observed in each lane. play a role in the attenuating the inflammation and
The capacity of purified recombinant (r) mSLPI to tissue destruction that occur during various arthriti-function in vitro as a serine protease inhibitor was then des. In addition, SLPI is expressed in skin, where itassessed. Human neutrophil elastase (HNE) activity may regulate the process of physiologic desquamationwas efficiently inhibited by pre-incubation with by inhibiting a stratum corneum chymotryptic proteasermSLPI in the low nanomolar concentration range (12). Moreover, the isolation of SLPI as the major ser-(Figure 5A). An inhibition ofú90% of the HNE activity ine antiprotease in psoriatic plaques implicates it as aoccurred at [rmSLPI] Å 50 nM. The inhibiton constant potential modulator of cutaneous inflammation (14). In(Ki) based on a tight-binding interaction model was 1.4 mammalian species with noninvasive epitheliochorialnM. When tested in parallel, rhSLPI inhibited HNE placentation, such as pig, horse and cow, SLPI iswith a Ki of 0.2 nM (Figure 5B). rmSLPI was also a strongly expressed by the uterine endometrium duringpotent chymotrypsin inhibitor. At [rmSLPI] Å 240nM, mid- and late phase pregnancy. In contrast, those spe-ú90% of bovine chymotrypsin activity was inhibited, cies with invasive hemochorial placentation such asand a Ki of 5 nM was observed (Figure 5C), similar to rodents and humans do not show such endometrialthe 2.4 nM Ki observed for the interaction between SLPI production, implying that trophoblastic invasive-rhSLPI and bovine chymotrypsin (Figure 5D). rmSLPI ness may ordinarily be regulated, at least in part, bywas a less potent inhibitor of human neutrophil cathep- SLPI (13). Finally, in addition to its antiprotease activ-sin G (HNCG). At [rmSLPI] Å0.5 mM, only 80% inhib- ity, SLPI also functions in vitro as an antibiotic, in aiton of HNCG was observed, and the Ki values for fashion similar to that of the defensin family of pep-rmSLPI and rhSLPI were 93 nM, and 13nM, respec- tides (40). When taken together, these observationstively (Figure 5E, 5F). In sum, mSLPI is a potent inhib- suggest that in addition to protecting epithelial sur-itor of several proteases with a decreasing gradation of faces, SLPI may function more broadly to regulateactivity from HNE through HNCG, that follows the many physiologic and pathologic processes that requiresame gradation shown by hSLPI. However, in marked tightly controlled proteolysis.contrast to hSLPI, mSLPI was a poor inhibitor of tryp- To distinguish which of the many hypothesized rolessin. Even at micromolar concentrations, rmSLPI was of SLPI are of physiologic importance requires thenot able to completely inhibit bovine trypsin (data not study of the biology of SLPI in an animal species whichshown). The Ki values for the inhibition of bovine tryp- can be genetically manipulated. To this end, we reportsin by rmSLPI and rhSLPI were 646 and 44 nM, respec- the cloning and analysis of a cDNA encoding the mu-tively. rine homologue of SLPI. The cDNA encodes a predicted
translation product of 14 kDa. Because the N-terminusDISCUSSION of the secreted protein begins at residue 26, residues
1-25 likely constitute a signal peptide. The predictedsize of the mature secreted protein is approximately 12SLPI is a serine protease inhibitor which is produced
by epithelial cells at mucosal surfaces. Human SLPI is kDa. mSLPI appears to be encoded by a single gene,
693
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
AID BBRC 6358 / 6924$$6358 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
and cross-hybridization experiments suggest that a mains. The protease binding region of human SLPIis contained within the C-terminal domain, while thesimilar gene exists in many mammalian species, in-
cluding the reported 3.8 kbp EcoRI fragment of human N-terminal domain is responsible both for heparinbinding and for the ‘‘defensin-like’’ properties of theDNA (29). The mSLPI mRNA is expressed in a variety
of tissues, generally in a similar pattern as that ob- molecule (7,33,40,45,46). Interaction of the N-termi-nus with heparin increases both the association rateserved in the human. The exception to this is the very
high expression observed in murine spleen. The physio- constant, and the binding affinity for proteases(33,45). Formal proof that the homologous regions oflogic target for mSLPI within the spleen of mice is not
obvious, although the novel ‘‘MTSP’’ proteases recently mSLPI share similar properties will be a further fo-cus of future investigations. Third, it is notable thatcloned from mouse spleen (41) represent an intriguing
possibility, and an important focus of future study. the interspecies sequence conservation in the signalpeptide of these proteins is even greater than thatWhen analyzed by SDS-PAGE, mSLPI expressed
and secreted by CHO cells migrates at a slightly higher seen the mature peptide, and all of these signal pep-tides are also highly homologous to the elafin signalapparent molecular weight than that which would be
expected based on its predicted size. This is most likely peptide (43). Since these peptides are not normallyconserved over and above the maintenance of hydro-due to the substantial net positive charge on the mole-
cule at physiologic pH (the predicted pI of m- and phobicity, the striking sequence similarity suggeststhat in these protease inhibitors, the leader regionshSLPI are 8.8 and 8.9, respectively). rhSLPI also exhib-
ited this altered electrophoretic pattern both in the may additionally contain important and specific sig-nals for regulated trafficking and secretion.present studies as well as those of others (3). Our im-
munoblotting studies also detected a similar protein in The differences in structure between the hSLPI andmSLPI may provide some explanation for differencesthe BAL fluid of normal mice, though a number of
higher molecular weight bands were also seen. These in the spectrum of antiprotease activity exhibited bymSLPI. The mSLPI protein inhibited human neutro-bands may be due to nonspecific antibody cross reactiv-
ity with other proteins in BAL, to aggregation of the phil elastase, bovine chymotrypsin, and human neutro-phil cathepsin G; but compared to hSLPI, it was a nota-mSLPI protein, or to complexes of SLPI with proteases
(or with other proteins) which are not dissociated under bly poor inhibitor of bovine trypsin. The protease bind-ing sites of the three SLPI proteins from the predictedthe conditions used for SDS-PAGE.
Comparison of similarities and differences in struc- P5 to the P*5 residues are illustrated in Table I. Theprimary structure of the murine protein differs fromture between human and mouse SLPI may give im-
portant insights into the biochemical function of the hSLPI at the P5, P4, and P3 residues, but probably themost critical difference is the presence of methionineprotein. There are several conserved features: First,
the position of cysteine residues within each domain rather than leucine at the P1 position. By extrapolationof findings on oxidation of both the non-P1 methionineis highly conserved, forming a pattern termed a ‘‘4-
disulfide core’’. A number of antiprotease molecules residues in hSLPI, and the P1 methionine in humanalpha-1 antitrypsin (a1AT), it is likely that the pres-share this motif including human, porcine, and mu-
rine SLPI, the skin specific proteinase inhibitor ence of a P1 methionine residue renders mSLPI andpSLPI significantly more vulnerable to oxidative inac-elafin, the red sea turtle proteinase inhibitor, and
eNAP-2, a neutrophil product which inhibits the bac- tivation ((47,48), and references therein). Moreover,mutagenesis studies performed on hSLPI have shownterial serine proteinases subtilisin and proteinase K,
and which like SLPI, has ‘‘defensin’’-like antibiotic that substitutions at the P1 position have profoundeffects on the spectrum of SLPI antiprotease activityactivity (3,4,42-44). While this pattern of disulfide
bonds is assumed to stabilize the folded protein, the (7). A change of the 72Leu in hSLPI to positivelycharged residues decreases the Ki for trypsin 1000-fold,exact function is not known. Second, analysis of the
mSLPI sequence reveals that there is substantial in- while substitution with a hydrophobic phenylalanineresidue increases the Ki against trypsin 10-fold, whileternal homology between the N- and C-terminal por-
tions of the protein. In hSLPI, this pattern of internal simultaneously decreasing the Ki against chymotryp-sin to the same extent. Therefore, it is plausible thathomology dividies the molcule into two functional do-
FIG. 5. Comparison of the antiprotease activities of purified recombinant murine SLPI (rmSLPI) and recombinant human SLPI (rhSLPI).Inhibition of human neutrophil elastase by rmSLPI (A), and rhSLPI (B); inhibition of bovine chymotrypsin by rmSLPI (C), and rhSLPI(D);and inhibition of human neutrophil cathepsin G by rmSLPI (E), and rhSLPI (F) are shown. In each panel, the concentration of SLPI isshown on the abscissa, and the fractional protease activity at each inhibitor concentration compared with uninhibited activity is on theordinate. For each experiment, an inhibiton constant (Ki) is shown, which was calculated as described in Materials and Methods assuminga tight-binding model of inhibiton. The curves shown are representative of 2-3 replicate inhibiton experiments performed with each protease.
695
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
the presence of this P1 methionine alone is sufficient radation similar to SFIF, but has not yet been success-fully cloned (53,54). The exact identity of these HGF/to render mSLPI a poor inhibitor of trypsin.
Differences in the structure of antiproteases that af- SF-inducing bioactivities remains unclear at present,but will be the subject of ongoing experiments.fect their spectrum of inhibition have previously been
illustrated by studies of a-1 antitrypsin (a1AT). a1AT In conclusion, we have cloned the murine SLPI cDNAand characterized the protein it encodes. This proteinhas diversified in mice and humans via different ge-
netic mechanisms. Murine a1AT differs from the hu- is produced in the normal murine lung, and has potentantielastase activity. The availability of reagents forman gene in that extensive gene duplication has oc-
curred, in contrast to human a1AT, where polymor- the murine SLPI homologue will allow us to better ad-dress many of the remaining issues regarding the inphism is generated through myriad mutations at a
single locus (reviewed in (49)). The P1 residues vary in vivo biological function of SLPI. Structure-functionstudies using mutagenized recombinant mSLPI andthe five murine a1AT gene products, and the substitu-
tions have functional consequences. Two of the a1AT murine proteases should increase our understanding ofthe biochemistry of protease-antiprotease interactions,gene products have a P1 methionine residue, similar
to that of human a1AT, and mSLPI. The two gene prod- and the evolutionary divergence of this process be-tween species. Finally, the production of geneticallyucts with a tyrosine in the P1 position have diminished
anti-elastase activity when compared to those having a modified murine strains in which the SLPI gene is over-expressed or disrupted, and examination of the behav-P1 methionine. Another gene product with a P1 leucine
residue has essentially no measurable antiprotease ac- ior of these animals in various murine models of humandisease should be rewarding. Such experiments maytivity. Interestingly, similar to mSLPI, none of the mu-
rine ‘‘a1-antitrypsin’’ gene products is active against lead to a better understanding of the role that SLPIplays in host defense against pathogens, in the normalmurine trypsin (50-52). Thus, the differences that we
note between human and murine SLPI are reminiscent reproductive process, and in cutaneous, musculoskele-tal, and pulmonary inflammatory diseases.of those in a1AT, and may have been in part driven by
simultaneous dissimilarities that have evolved in thecognate murine proteases. ACKNOWLEDGMENTS
The protein sequence used to clone mSLPI was iden-The technical assistance of Susane Pyle and Ann Hartman is grate-tified during an attempt to isolate a ‘‘scatter factor in-
fully appreciated. This work was supported in part by NIH grantducing factor’’ (SFIF) bioactivity in the supernatantsK08-HL03087.of ras-transformed 3T3 cells (15). This SFIF activity
induced fibroblasts to produce hepatocyte growth fac-REFERENCEStor-scatter factor (HGF/SF), and had a number of
physical properties in common with SLPI including1. Cheronis, J. C., and Repine, J. E. (1993) in Proteases, proteaseacid-stability, a relative resistance to proteolytic degra- inhibitors, and protease-derived peptides: Importance in human
dation, and similar size. However, while mSLPI is se- pathophysiology and therapeutics. Agents and Actions Supple-ments (Brune, K., and Parnham, M. J., Eds.), Birkhauser Verlag,creted by r3T3 cells (Figure 4), recombinant mSLPIBasel.protein expressed in both mammalian and prokaryotic
2. Potempa, J., Korzus, E., and Travis, J. (1994) J. Biol. Chem.systems had no discernible SFIF activity (data not269, 15957–15960.shown). There are a number of possibilities which
3. Thompson, R. C., and Ohlsson, K. (1986) Proc. Natl. Acad. Sci.might explain this discrepancy. First, the isolation ofUSA 83, 6692–6696.
mSLPI protein from the supernatants with SFIF activ-4. Seemuller, U., Arnhold, M., Fritz, H., Wiedenmann, K., Mach-
ity may be artifactual. mSLPI is produced in large leidt, W., Heinzel, R., Appelhans, H., Gassen, H.-G., andamounts by r3T3 cells and the correct SFIF protein Lottspeich, F. (1986) FEBS Letters 199(1), 43–48.may have been a minor component which co-purified 5. Maruyama, M., Hay, J. G., Yoshimura, K., Chu, C.-S., and Crys-
tal, R. G. (1994) J. Clin. Invest. 94, 368–375.with mSLPI. Consistent with this possibility is the6. Sallenave, J.-M., Shulmann, J., Crossley, J., Jordana, M., andpresence of several smaller bands in the vicinity of the
Gauldie, J. (1994) Am. J. Resp. Cell Mol. Biol. 11, 733–741.major SFIF band seen on SDS-PAGE (15). Second,7. Eisenberg, S. P., Hale, K. K., Heimdal, P., and Thompson, R. C.there may be a physiologically relevant association be-
(1990) J. Biol. Chem. 14, 7976–7981.tween SLPI and SFIF, whereby SLPI may bind with8. Grutter, M. G., Fendrich, G., Huber, R., and Bode, W. (1988)(or protect from proteolytic degradation) another factor
EMBO J. 7(2), 345–351.or factors necessary in order to fully reconstitute the
9. Vogelmeier, C., Gillisen, A., and Buhl, R. (1996) Chest 110,SFIF bioactivity. It is intriguing that a scatter factor- 261S–266S.inducing bioactivity isolated from the serum of injured 10. Lee, C., Igarashi, Y., Hohman, R. J., Kaulbach, H., White, M. V.,rats, termed ‘‘injurin’’ was reported to have physio- and Kaliner, M. A. (1993) Am. Rev. Resp. Dis. 147, 710–716.chemical properties including low molecular weight, 11. Jacoby, A. S., Melrose, J., Robinson, B. G., Hyland, V. J., and
Ghosh, P. (1993) Eur. J. Biochem. 218, 951–957.acid and heat stability, and resistance to protease deg-
696
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC

Vol. 232, No. 3, 1997 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
12. Franzke, C. W., Baici, A., Bartels, J., Christophers, E., and Wie- forov, T., and Dijkman, J. H. (1990) Bioch. Biophys. Acta. 1038,178–185.dow, O. (1996) J. Biol. Chem. 271, 21896–21890.
33. Ying, Q.-L., Kemme, M., and Simon, S. R. (1994) Biochem. 33,13. Simmen, R. C. M., Michel, F. J., Fliss, A. E., Smith, L. C., and5445–5450.Fliss, M. F. V. (1992) Endocrinology 130(4), 1957–1965.
34. Casslen, B. (1986) Acta Obstet. Gynecol. Scand. 65, 121–124.14. Wiedow, O., Young, J. A., Davison, M. D., and Christophers, E.(1993) J. Invest. Dermatol. 101, 305–309. 35. Kramps, J. A., Franken, C., Meijer, C. J. L. M., and Dijkman,
J. H. (1981) J. Histochem. Cytochem. 29(6), 712–719.15. Rosen, E., Joseph, A., Jin, L., Rockwell, S., Elias, J., Knessel, J.,Wines, J., J, M., Kluger, M., Goldberg, I., and Zitnik, R. J. (1994) 36. Franken, C., Meijer, C. J. L. M., and Dijkman, J. H. (1989) J.J. Cell Biol. 127(1), 225–234. Biochem. Cytochem. 37(4), 493–498.
16. Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular 37. Ohlsson, K., Bjartell, A., and Lilja, H. (1995) J. Androl. 16(1),Cloning: A Laboratory Manual, Second ed., Cold Spring Harbor 64–74.Laboratory Press, Cold Spring Harbor, NY. 38. Bohm, B., Deutzmann, R., and Burkhardt, H. (1991) Biochem.
17. Zitnik, R. J., Kotloff, R. M., Latifpour, J., Zheng, T., Whiting, J. 274, 269–273.N. L., Scwalb, J., and Elias, J. A. (1994) J. Immunol. 152, 1419– 39. Andrews, J. L., Melrose, J., and Ghosh, P. (1992) Biol. Chem.1427. Hoppe Selyer 373, 111–118.
18. Kohno, T., Carmichael, D. F., Sommer, A., and Thompson, R. C. 40. Hiemstra, P. S., Maassen, R. J., Stolk, J., Heinzel-Wieland, G. J.,(1990) Methods Enzymol. 185, 187–195. Steffens, G. J., and Dijkman, J. H. (1996) Infect. Immun. 64(11),
19. Seely, R. J., and Young, M. D. (1991) in ACS Symposium Series 4520–4524.(Georgiou, G., and DeBernandez-Clark, E., Eds.), Vol. 470, pp. 41. Fukusen, N., and Aoki, Y. (1996) J. Biochem. 119, 633–638.206–216, American Chemical Society, Washington, DC. 42. Farmer, S. J., Fliss, A. E., and Simmen, R. C. M. (1990) Mol.
20. Kramps, J. A., van Twisk, C., and van der Linden, A. C. (1983) Endocrinol. 4, 1095–1104.Scand. J. Clin. Lab. Invest. 43, 427–432. 43. Molhuizen, H. O. F., Alkemade, H. A. C., Zeeuwen, P. L. J. M.,
21. Somorin, O., Tokura, S., Nishi, N., and Noguchi, J. (1979) J. de Jongh, G. J., Wieringa, B., and Scalkwijk, J. (1993) J. Biol.Biochem. 85, 157–162. Chem. 268(16), 12028–12032.
22. DelMar, E. G., Largman, C., Brodrick, J. W., and Geokas, M. C. 44. Couto, M. A., Harwig, S. S. L., and Lehrer, R. I. (1993) Infect.(1979) Anal. Biochem. 99, 316–320. Immun. 61(7), 2991–2994.
23. Groutas, W. C., Brubaker, M. J., Venkataraman, R., Epp, J. B., 45. Faller, B., Mely, Y., Gerard, D., and Bieth, J. (1992) Biochem.Stanga, M. A., and McClenahan, J. J. (1992) Arch. Biochem. Bio- 31, 8285–8290.phys. 294(1), 144–146. 46. Mellet, P., Ermolieff, J., and Bieth, J. G. (1995) Biochem. 34,
24. Barrett, A. J., and Salvesen, G. (1986) in Proteinase Inhibitors, 2645–2652.Vol. 12, Research Monographs in Cell and Tissue Physiology 47. Luisetti, M., and Travis, J. (1996) Chest 110, 278S–283S.(Dingle, J. T., and Gordon, J. L., Eds.), Elsevier, Amsterdam.
48. Schuster, A., Hansen, G., Zubrod-Eichert, C., and Wahn, V.25. Walter, M., Plotnick, M., and Schechter, N. M. (1996) Arch. Bio- (1996) Ped. Res. 40(5), 732–737.
chem. Biophys. 327(1), 81–88.49. Eriksson, S. (1996) Chest 110(6), 237S–242S.
26. Petersen, L. C., Sprecher, C. A., Foster, D. C., Blumberg, H., Ha-50. Borriello, F., and Krauter, K. S. (1991) Proc. Natl. Acad. Sci.mamoto, T., and Kisiel, W. (1996) Biochem. 35, 262–272.
USA 88, 9417–9421.27. Bieth, J. G. (1984) Biochem. Med. 32, 387–397. 51. Paterson, T., and Moore, S. (1996) Biochem. Biophys. Res. Comm.28. Kozak, M. (1986) Cell 44, 283–292. 219, 64–69.29. Stetler, G., Brewer, M. T., and Thompson, R. C. (1986) Nuc. 52. Takahara, H., and Sinohara, H. (1983) J. Biochem. 93, 1411–
Acids. Res. 14, 7883–7896. 1419.30. Simmen, R. C. M., Badinga, L., and Michel, F. J. (1993) Mol. 53. Okazaki, H., Matsumoto, K., and Nakamura, T. (1994) Bioch.
Cell. Endocrinol. 97, 101–108. Biophys. Acta. 1220, 291–298.31. Smith, C. E., and Johnson, D. A. (1985) Biochem. J. 225, 463– 54. Matsumoto, K., Tajima, H., Hamanque, M., Kohno, S., Kino-
472. shita, T., and Nakamura, T. (1992) Proc. Natl. Acad. Sci. USA89, 3800–3804.32. Kramps, J. A., van Twisk, C., Appelhans, H., Meckelein, B., Niki-
697
AID BBRC 6358 / 6924$$$501 03-10-97 09:56:45 bbrcgs AP: BBRC



















