
Corticosteroids modulate the secretory processes of the rat intrahepatic biliary epithelium
12
GASTROENTEROLOGY 2002;122:1058-1069 Corticosteroids Modulate the Secretory Processes of the Rat Intrahepatic Biliary Epithelium DOMENICO ALVARO,* ALESSANDRO GIGLIOZZI,* LUCA MARUCCI,* GIANFRANCO ALPINI,§ BARBARA BARBARO,* RITA MONTERUBBIANESI,* LARA MINETOLA,* MARIA GRAZIA MANClNO,* JUAN F. MEDINA,II ADOLFO F. ATTILI,* and ANTONIO BENEDE]-I-I¢ *Division of Gastroenterology, Departmentof Clinical Medicine, UniversityLa Sapienza, Rome, Italy; tDepartment of Gastroenterology, University of Ancona, Ancona, Italy; §Departmentof Internal Medicine and Medical Physiology, Scott and White Hospital and The TexasA&M System Health Science Center, Collegeof Medicine, and Central Veterans Health Care, Temple, Texas; and "Laboratoryof Molecular Genetics, Division of Hepatologyand GeneTherapy, Universityof Navarra, Pamplona, Spain Background & Aims: We investigated the expression of glucocorticoid receptors (GcRs) in the intrahepatic bili- ary epithelium and the role of corticosteroids in the regulation of cholangiocyte secretion. Methods: GcR was studied by immunohistochemistry, reverse-transcription polymerase chain reaction, and Western blots. The ef- fects of dexamethasone and budesonide on biliary bi- carbonate excretion and H+/HCOa - transport processes were investigated in bile fistula rats, isolated intrahe- patic bile duct units (IBDUs), and purified cholangio- cytes. Results: GcRs were expressed by rat cholangio- cytes. Although acute administration of corticosteroids showed no effect, treatment for 2 days with dexameth- asone or budesonide increased (P < 0.05) biliary bicar- bonate concentration and secretion, which were blocked by the specific GcR antagonist, RU-486. IBDUs isolated from rats treated with dexamethasone or budesonide showed an increased (P < 0.05) activity of the Na+/H + exchanger (NHE1 isoform) and CI-/HC03- exchanger (AE2 member), which was blocked by RU-486. Protein expression of NHE1 and AE2 and messenger RNA for NH1 but not AE2 were increased (P < 0.05) in isolated cholangiocytes by dexamethasone treatment. Conclusions: The intrahepatic biliary epithelium ex- presses GcR and responds to corticosteroids by increas- ing bicarbonate excretion in bile. This is caused by corticosteroid-induced enhanced activities and protein expression of transport processes driving bicarbonate excretion in the biliary epithelium. A major function of the intrahepatic biliary epithe- lium is bicarbonate excretion in bile, which is mainly driven by the apical Na+-independent CI-/ HCO3- exchanger identified as isoform AE2 in rat and man and which is functionally coupled with the cystic fibrosis transmembrane regulator (CFTR). 1-6 In rat cholangiocytes, the activity of this anion exchanger is balanced by 2 acid extruders, the NHE1 isoform of Na+/H + exchanger and the Na+:HCO3 - cottansporter, which are both located in the basolateral pole of cholan- giocytes and, in concert with the anion exchanger, coop- erate in the regulation ofintracellular pH (pHi). >v In the intrahepatic biliary epithelium, bicarbonate biliary ex- cretion is regulated by a number of hormones/neuro- peptides. 1 Secretin, for example, induces bicarbonate choleresis by stimulating the CI-/HCO 3- exchanger ac- tivity in cholangiocytes. 2'4"~ The biliary epithelium is the specific target of damage in a group of chronic cholestatic liver diseases, the cholangiopathies, which represent a major indication for liver transplantation. 1 The most common acquired cholangiopathy is primary biliary cirrhosis (PBC), in which the expression and function of AE2 and secretin- induced choleresis are decreased. 6,*,9 In cystic fibrosis, the activity of the anion exchanger and bicarbonate excretion in bile is impaired because of a genetic CFTR defect3 ° Ursodeoxycholic acid (UDCA) is the only approved treatment for PBC, but clinical results are not highly satisfactory.~ 1 Recent reports indicate that budesonide, a corticosteroid with high receptor affinity and hepatic first pass clearance, ~2 improves the efficacy of UDCA in PBC Abbreviations used in this paper: BDL,bile duct ligation; CFTR, cystic fibrosis transmembrane regulator; DMSO, dimethylsulfoxide; EIPA, ethyl-n-isopropyl-amiloride; GAPDH, glyceraldehyde-3-phosphate dehy- drogenase; GcR, glucocorticoid receptor; IBDU, isolated rat bile duct unit; KRB, Krebs-Henseleit bicarbonate; PBC,primary biliary cirrhosis; pH=, intracellular pH; RT-PCR, reverse-transcription polymerase chain reaction; SR, secretin receptor. © 2002 by the American Gastroenterological Association 0016-5085/02/$35.00 doi:10.1053/gast.2002.32374
Transcript of Corticosteroids modulate the secretory processes of the rat intrahepatic biliary epithelium

GASTROENTEROLOGY 2002;122:1058-1069
Corticosteroids Modulate the Secretory Processes of the Rat Intrahepatic Biliary Epithelium
DOMENICO ALVARO,* ALESSANDRO GIGLIOZZI,* LUCA MARUCCI,*
GIANFRANCO ALPINI,§ BARBARA BARBARO,* RITA MONTERUBBIANESI,* LARA MINETOLA,* MARIA
GRAZIA MANClNO,* JUAN F. MEDINA,II ADOLFO F. ATTILI,* and ANTONIO BENEDE]-I-I¢ *Division of Gastroenterology, Department of Clinical Medicine, University La Sapienza, Rome, Italy; tDepartment of Gastroenterology, University of Ancona, Ancona, Italy; §Department of Internal Medicine and Medical Physiology, Scott and White Hospital and The Texas A&M System Health Science Center, College of Medicine, and Central Veterans Health Care, Temple, Texas; and "Laboratory of Molecular Genetics, Division of Hepatology and Gene Therapy, University of Navarra, Pamplona, Spain
Background & Aims: We investigated the expression of glucocorticoid receptors (GcRs) in the intrahepatic bili- ary epithelium and the role of corticosteroids in the regulation of cholangiocyte secretion. Methods: GcR was studied by immunohistochemistry, reverse-transcription polymerase chain reaction, and Western blots. The ef- fects of dexamethasone and budesonide on biliary bi- carbonate excretion and H+/HCOa - transport processes were investigated in bile fistula rats, isolated intrahe- patic bile duct units (IBDUs), and purified cholangio- cytes. Results: GcRs were expressed by rat cholangio- cytes. Although acute administration of corticosteroids showed no effect, treatment for 2 days with dexameth- asone or budesonide increased (P < 0.05) biliary bicar- bonate concentration and secretion, which were blocked by the specific GcR antagonist, RU-486. IBDUs isolated from rats treated with dexamethasone or budesonide showed an increased (P < 0.05) activity of the Na+/H + exchanger (NHE1 isoform) and CI-/HC03- exchanger (AE2 member), which was blocked by RU-486. Protein expression of NHE1 and AE2 and messenger RNA for NH1 but not AE2 were increased (P < 0.05) in isolated cholangiocytes by dexamethasone treatment. Conclusions: The intrahepatic biliary epithelium ex- presses GcR and responds to corticosteroids by increas- ing bicarbonate excretion in bile. This is caused by corticosteroid-induced enhanced activities and protein expression of transport processes driving bicarbonate excretion in the biliary epithelium.
A major function of the intrahepatic biliary epithe- lium is bicarbonate excretion in bile, which is
mainly driven by the apical Na+-independent CI-/ HCO3- exchanger identified as isoform AE2 in rat and man and which is functionally coupled with the cystic fibrosis transmembrane regulator (CFTR). 1-6 In rat
cholangiocytes, the activity of this anion exchanger is balanced by 2 acid extruders, the NHE1 isoform of Na+/H + exchanger and the Na+:HCO3 - cottansporter, which are both located in the basolateral pole of cholan- giocytes and, in concert with the anion exchanger, coop- erate in the regulation ofintracellular pH (pHi). >v In the intrahepatic biliary epithelium, bicarbonate biliary ex- cretion is regulated by a number of hormones/neuro- peptides. 1 Secretin, for example, induces bicarbonate choleresis by stimulating the CI-/HCO 3- exchanger ac- tivity in cholangiocytes. 2'4"~
The biliary epithelium is the specific target of damage in a group of chronic cholestatic liver diseases, the cholangiopathies, which represent a major indication for liver transplantation. 1 The most common acquired cholangiopathy is primary biliary cirrhosis (PBC), in which the expression and function of AE2 and secretin- induced choleresis are decreased. 6,*,9 In cystic fibrosis, the activity of the anion exchanger and bicarbonate excretion in bile is impaired because of a genetic CFTR defect3 °
Ursodeoxycholic acid (UDCA) is the only approved treatment for PBC, but clinical results are not highly satisfactory.~ 1 Recent reports indicate that budesonide, a corticosteroid with high receptor affinity and hepatic first pass clearance, ~2 improves the efficacy of UDCA in PBC
Abbreviations used in this paper: BDL, bile duct ligation; CFTR, cystic fibrosis transmembrane regulator; DMSO, dimethylsulfoxide; EIPA, ethyl-n-isopropyl-amiloride; GAPDH, glyceraldehyde-3-phosphate dehy- drogenase; GcR, glucocorticoid receptor; IBDU, isolated rat bile duct unit; KRB, Krebs-Henseleit bicarbonate; PBC, primary biliary cirrhosis; pH=, intracellular pH; RT-PCR, reverse-transcription polymerase chain reaction; SR, secretin receptor.
© 2002 by the American Gastroenterological Association 0016-5085/02/$35.00
doi:10.1053/gast.2002.32374

April 2002 CORTICOSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1059
patients, ~3 even if it is of no beneficial effects in patients
that do not respond to U D C A alone. 14 In pr imary scle-
rosing cholangitis, prednisone exerts shor t - term benefi-
cial effects when administered together with U D C A ,
whereas budesonide has no effects) 5 However, short
t reatment wi th corticosteroids may transiently improve
intrahepatic cholestasis of different origins (corticoste- roid whitewash), a6
Corticosteroids regulate secretory processes and the
activity of electrolyte transporters in different tissues
(kidney, colon, and i l eum)) 7-~9 However, the effects of
corticosteroids on the pathophysiology of the intrahe-
patic biliary epi thel ium are unknown. W e investigated
the effects of corticosteroids on the activities and protein
expression of electrolyte transporters involved in pHi
regulation and bicarbonate biliary excretion in the rat
intrahepatic biliary epithelium.
Materials and Methods
Male Wistar rats (225-260 g) were purchased from Charles River (Calco, Lecco, Italy). Reagents were purchased from Sigma Chemical Co. (St. Louis, MO) unless otherwise indicated. RU-486 was kindly provided by Roussel-Uclaf, Exelgyn, Paris, France. Bile duct ligation (BDL; 2 weeks) and preparation of intrahepatic rat bile duct units (IBDUs), iso- lated cholangiocytes, and hepatocytes were performed as pre- viously described. 2,2°-22 The study protocols were in compli- ance with our institution's guidelines.
Animal Treatment and Functional Studies
Dexamethasone or budesonide were prepared as a stock solution in dimethylsulfoxide (DMSO) and then diluted (1: 10,000) in Krebs-Henseleit bicarbonate (KRB) buffer + 0.2% bovine serum albumin to obtain the desired final concentra- tion. For acute experiments, dexamethasone (5 Dmol/L for 4 0 - 8 0 minutes) or budesonide (1 txmol/L for 4 0 - 8 0 minutes) was infused via jugular vein, and the effect on bile flow or bicarbonate biliary excretion was evaluated as described 2° and compared with basal values. Administration of KRB + bovine serum albumin + DMSO in control rats did not change bile flow and biliary bicarbonate excretion. To evaluate the effect of prolonged administration of corticosteroids, normal rats were treated for 2 days with dexamethasone, budesonide, or RU- 486 administered subcutaneously 3 times daily at the dose of 0.9, 0.15, and 1 mg, respectively. Normal rats, 2-week BDL rats, and rats treated for 2 days with dexamethasone or budes- onide were surgically prepared for bile collection as previously described. 2° After treatment with corticosteroids, normal rats were studied for bile secretion as described above or IBDU, cholangiocytes, or hepatocytes were isolated and cultured in media added with budesonide (1 btmol/L) or dexamethasone (5 btmol/L) for the study of electrolyte transporters.
Immunohistochemical Studies and Molecular Analysis of Glucocorticoid Receptors
For immunohistochemistry of glucocorticoid receptors (GcRs), liver sections were obtained from normal, dexametha- sone-treated, dexamethasone-control (cartier only), and 1- week BDL rats, incubated with the polyclonal primary anti- body against GcR (Santa Cruz Biotechnology Inc., Santa Cruz, CA; 1:20 in phosphate-buffered saline) for 2 hours at room temperature, and the reactive sites were detected using a DAKO LSAB kit (Copenhagen, Denmark) according to the manufacturer's instructions, followed by incubations for 5 minutes with 0.06% diaminobenzidine and 0.01% H202. In control sections, antibody binding to antigen was neutralized by preadsorption with the blocking peptide (Santa Cruz Bio- technology Inc.) according to the vendor's instructions. Im- munohistochemistry was also performed for cytokeratin 7 as described, 2t and bile duct mass was then measured by quan- titative morphometry. 2.
GcRs were also analyzed by reverse-transcription polymer- ase chain reaction (RT-PCR) in pure preparations of cholan- giocytes isolated from normal and BDL rats by immunomag- netic separation as described. 2~ Specific primers (sense 5'- GCTTCAGGATGTCATTACGGGGT-3 ' and antisense 5'- GTTTCTC-AAGCCTAGTATCGCCT-3' , expected fragment length 475 bp) designed from a sequence encoding for rat GcR 23 were used. Poly(A)+ messenger RNA (mRNA) from rat brain and yeast transfer RNA were, respectively, the pos- itive and negative controls. The fo l lowing RT-PCR conditions were used with 5 ng poly(A) + mRNA: 35 step cycles (30 seconds at 94°C, 30 seconds at 57°C, and 45 seconds at 72°C) by using a Taq DNA polymerase (PE Applied Biosystems, Foster City, CA). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as a housekeeping gene. 24
Measurement of the Activities of H+/HC03 - Transport Processes in IBDU
pHi of IBDU was measured using a microfluorimetric single-cell method (SPEX-AR-CM; Spex Industries, Edison, NJ) and 2,7,bis(carboxyethyl)-5(6)-carboxy-fluorescein-ace- toxymethyl-ester (Molecular Probes Inc., Eugene, OR) as a fluorescent pHi indicator. 2,3,2e The activities of C1 /HCO~- and Na+/H + exchangers and Na+-HCO3 - cotransporter were measured as described 2"3,22 and detailed above.
Western Blot Analysis of Electrolyte Transporters
Western blot analysis of GcR, NHE1, AE2, N+/K +- adenosine triphosphatase (ATPase), CFTR, and Na+/K+/2CI -
in isolated cholangiocytes or hepatocytes was performed as described 21 by using the following antibodies: (1) polyclonal primary antibody against GcR (Santa Cruz Biotechnology Inc.; dilution 1:200 in phosphate-buffered saline); (2) antibody anti-NHE1 isoform of the Na+/H + (dilution 1:300) (Chemi- con, Hofheim, Germany) and the immunocomplexes detected

1060 ALVARO ET AL. GASTROENTEROLOGY Vol. 122, No. 4
i
Figure 1. Immunohistochemistry for GcR in liver sections from normal, dexamethasone-treated, and 1-week BDL rats. (A) In normal liver, a specific reaction appears in the cytoplasm mostly in the apical area and in the perinuclear region of cholangiocytes (curved arrow). (B) After BDL, a pronounced nuclear positivity (arrow) as well as a diffuse cytoplasmic staining are evident. (C) A stronger nuclear and cytoplasmic positivity is evident in sections of dexamethasone-treated rats. in all situations, a positive reaction is also present in cells of the surrounding connective tissue (arrowhead) and hepatocytes (star). Control sections taken from (A1) normal, (B1) 1-week BDL, and (C1) dexamethasone-treated rats and incubated with the primary antibody neutralized by preabsorption with the blocking peptide show absence of specific staining. L, lumen. Original magnification 2 5 0 x .
using alkaline phosphatase-conjugated goat anti-mouse im- munoglobulin (Ig) G (dilution 1:4000) and NBT/BiCip as substrates (Sigma); (3) monoclonal antibody anti-AE2 aa 655- 668 (1:2500) (E. Martinez-Anso, J. Prieto, and Juan F. Me- dina, unpublished observation); (4) antibody anti-Na+/K + ATPase (Chemicon; 1:700); (5) polyclonal CFTR antibodies (Santa Cruz; dilution 1:100); and (6) T4 monoclonal antibody anti-Na+/K+/2C1- (1:3000), developed by C. Lytle. This an- tibody was obtained from the Developmental Studies Hybrid- oma Bank, developed under the auspices of the NICHD and maintained by the University of Iowa, Department of Biolog- ical Sciences, Iowa City, IA. Immunocomplexes were detected by horseradish-conjugated goat anti-mouse IgM (dilution 1:4000) (Chemicon), anti-chicken IgG, or anti-rabbit IgG, using an enhanced chemiluminescence Western blotting kit
(Amersham, Little Chalfont, England) and evaluated by auto- radiography.
m R N A M e a s u r e m e n t s
Ribonuclease protection analysis was performed in iso- lated cholangiocytes using the Direct Lysis RPA kit (Ambion Inc., Austin, TX) as described21 The [(x-32p]-uridine triphos- phate (800 Ci/mmol; Amersham, Arlington Heights, IL) la- beled antisense riboprobes were synthesized from: (1) an 889- base pair (bp) complementary DNA (cDNA) subcloned in pGEM-3Zf vector, using T7 RNA-polymerase, for NHE1 isoform of the Na+/H + exchanger; (2) a 349-bp cDNA sub- cloned in PCRtmlI vector, using SP6 RNA-polymerase, for the AE2 member of the CI / HCO 3- exchanger; (3) a 318-bp cDNA for the secretin receptors (SRs) was transcribed from

April 2002 CORTICOSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1061
A
o3
13_
g
0
Z Z
G l u c o c o r t ~ c o i d R e c e D t o r m F I N A
4 7 5 b p
B
GcR
Cholangiocytes 106 kd . . . .
N+Dexa. N BDL
% increase vs normals
0 N+Dexa. BDL
Figure 2. (A) Cholangiocytes from both normal and BDL rats were examined for the presence of the message for GcR and GAPDH (the housekeeping gene) by RT-PCR. The message for GcR was expressed by normal cholangiocytes and up-regulated in cholangiocytes from BDL rats. GAPDH gene expression, which assures the comparability of the total mRNA used, was similar among normal and hyperplastic cholangiocytes from BDL rats. Poly (A) + mRNA from rat brain and yeast transfer RNA were the positive and negative controls, respectively. Representative experiment of n = 2. (B) Western blot analysis of GcR in cholangiocytes isolated from normal rats (N), rats treated for 2 days with dexamethasone (N+Dexa.), and BDL rats. Bar graph shows the percent increase of protein mass in comparison with normal rats. Data are mean _+ SE of n = 3. *P < 0.05 vs. normals.
pGEM4Z-SR (a gift of Dr. N. LaRusso). GAPDH was used as a housekeeping gene, 21
Statistical Analyses
Data are shown as arithmetic means - SE. Statistical analysis was conducted using the Student t test or the analysis of the variance when multiple comparisons were carried out.
Results
Immunohistochemistry for GcR (Figure 1) was performed in liver sections from normal, dexamethasone- treated, dexamethasone-control (carrier only), and 1-week BDL rats. In normal (Figure 1A) and in dexamethasone- control liver (not shown), a specific reaction appears in the cytoplasm, mostly in the apical area and in the perinuclear region of cholangiocytes. After 1 week BDL (Figure 1B), a pronounced nuclear positivity and a dif- fuse cytoplasmic staining is evident. A stronger nuclear and cytoplasmic positivity is evident in sections of dex- amethasone-treated rats (Figure 1C). In all situations, a positive reaction is also present in cells of the surround- ing connective tissue and hepatocytes. Control sections
taken from normal, 1-week BDL, dexamethasone-
treated, and dexamethasone-control rats and incubated
with the primary antibody neutralized by preabsorption
with the blocking peptide show the absence of specific
staining (Figure 1).
By RT-PCR, the message for the GcR (475 bp) was
expressed by cholangiocytes from normal rats and was
increased in cholangiocytes isolated from BDL rats,
whereas the expression of G A P D H mRNA (the house-
keeping gene) was unchanged (Figure 2A). By Western
blot, the GcR was significantly (P < 0.05) increased in
pure preparations of cholangiocytes isolated from BDL
rats (n = 3) or from normal rats treated with dexameth-
asone for 2 days (n = 3), in comparison with normal rats
(n = 3; Figure 2B). Acute intravenous administration of dexamethasone (5
gtmol/L, n = 10) or budesonide (1 ~mol/L, n = 6) for
4 0 - 8 0 minutes in normal or BDL rats induced no
significant changes in basal bile flow or bicarbonate
biliary excretion, which also remained stable when the 2
corticosteroids were interrupted (Figure 3A).

1062 ALVARO ET AL. GASTROENTEROLOGY Vol. 122, No. 4
A ~ .2
NORMAL RATS
• DEXA. 5 M • BUDE. 1 M V
0 2 0 4 0 6 0 8 0 1 0 0 T i m e ( ra in)
• DEXA. 5 M s-50 T • BUDE. i M
~._~
~ . ~
"" 30~-
B
0 2 0 4 0 6 0 8 0 1 O0 T ime (min)
• "= 1
~'2.5
1.5
70
g~
~ =
4~ o
2 BileFIow j [
&
' ' 7 -
O S
- 7 -
BDL RATS
• DEXA. 5 M • BUDE. 1 M
2 0 4 0 6 0 8 0 1 0 0 t 2 0 1 4 0 1 ( ; 0 1 8 0 T i m e (m in )
• DEXA. 5 M • BUDE. 1 M
2 0 4 0 6 0 8 0 1 0 0 1 2 0 1 4 0 1( ;0 1 8 0 T l r n e (mln)
H C O 3" output
T ] ,
i
0 i o C DEXA. BUDE. DEXA.+ C
RU-486 DEXA. BUDE. DEXA.+
RU-486
Figure 3. Effect of (A) acute or (B) chronic administrat ion of cort icosteroids on bile flow and bicarbonate biliary excretion in normal and BDL rats. (A) Dexa- methasone (5 ~LmoI/L, n = 10) or budesonide (1 i~mol/L, n = 6) administered for 4 0 - 8 0 minutes through the jugular vein in normal (left) or 2-week BDL (right) bile f istula rats showed no signif icant ef fect on bile flow and bicarbonate bil iary excretion. In addit ion, no signif- icant changes were observed when the 2 corticosteroids were interrupted. (B) A 2-day treat- ment of normal rats with dexa- methasone (DEXA. 0.9 mg /3 t imes daily, subcutaneously; n = 12) or budesonide (BUDE. 0 .15 mg/3 t imes daily, subcu- taneously; n = 12) induced a signif icant increase of bile flow (left) and bicarbonate biliary ex- cretion (right) compared with controls (C; n = 10) adminis- tered with the carrier alone (i.e,, DMSO+KRB). Rats treated with dexamethasone + RU-486 (n = 10), a specific GcR antagonist, showed a similar bile flow and bicarbonate biliary excretion to controls and significantly (P < 0.05) lower levels compared with rats treated with dexameth- asone alone. *P < 0.05 vs. con- trols; &P < 0.05 vs. dexametha- sone. Data are mean _+ SE.
Dexamethasone (5 Drool/L) administered to I B D U
perfused with KRB showed no effect on the C 1 - / H C O 3-
exchanger activity when compared with paired control
IBDU. In fact, the maximal rate of pHi alkalinization
after C I - removal as well as the maximal rate of pHi
recovery after C1- readmission (C1- removal = 0 .140 ---
0.011 vs. 0.133 + 0.010 [controls] p H units/min; C1-
readmission = 0.164 + 0.012 vs. 0 .170 + 0.011 p H
units/min; n = 10) were unaffected by dexamethasone. In contrast with acute administrat ion, the 2-day treat-
ment with dexamethasone (0.9 mg/3 times daily, sub-
cutaneously; n = 12) or budesonide (0.15 mg/3 times
daily, subcutaneously; n = 12) promoted an increase in
bile flow and bicarbonate biliary excretion, which were
significantly higher (P < 0.05) than paired controls (n =
10) administered with the carrier alone (i.e., D M S O +
KRB) (Figure 3B). Bile salt biliary excretion was similar
between rats treated for 2 days with dexamethasone
(36.6 - 2.9 nmol • min -1 • g wet liver -1) or budesonide
(37.9 --- 3.1 nmol • min -1 • g wet liver - i ) and controls
(35.3 --- 2.3 nmol • rain -1 • g wet l iver-i) . The effects of
dexamethasone on bile secretion were blocked by the GcR antagonist, RU486 . In fact, after dexamethasone
(0.9 mg, 3 times daily) + R U - 4 8 6 (1 mg, 3 times daily;
n = 10) for 2 days, bile flow and bicarbonate biliary
excretion were similar to controls (n = 10) but were
April 2002 CORTICOSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1063
lower (P < 0.05) compared with rats treated with dexa-
methasone alone (n = 12; Figure 3B). RU-486 alone (1 mg, 3 times daily; n = 5) showed no effect on basal bile flow and bicarbonate biliary excretion (controls; n = 5).
Liver weight was not significantly affected by treat-
ment with dexamethasone, budesonide, or RU-486 (nor-
mal = 9.4 ± 0.3 g; dexamethasone = 10.1 + 0.3 g; budesonide = 9.7 + 0.4 g; RU-486 = 9.3 + 0.3 g;
dexamethasone + RU-486 = 9.5 +-- 0.4 g). This allows the exclusion that the increased bile flow and bicarbonate biliary excretion is caused by an effect of corticosteroids on liver cell mass.
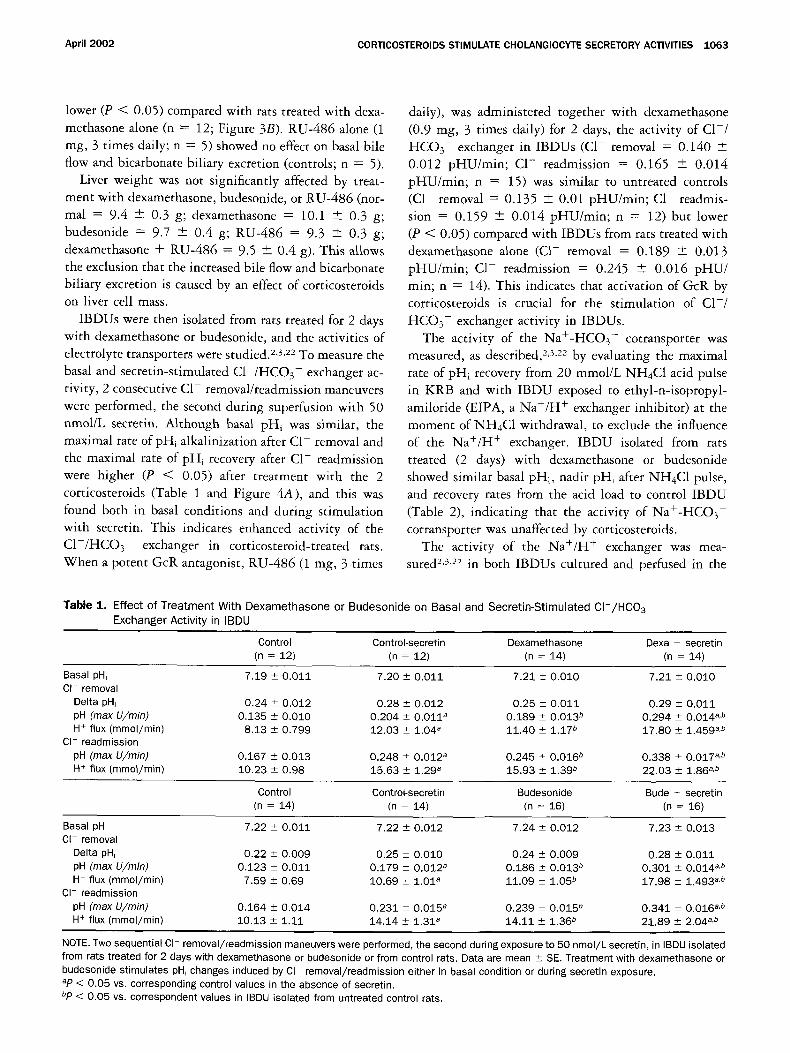
IBDUs were then isolated from rats treated for 2 days with dexamethasone or budesonide, and the activities of electrolyte transporters were studied. 2,3,22 To measure the
basal and secretin-stimulated C I - / H C O 3- exchanger ac-
tivity, 2 consecutive C1- removal/readmission maneuvers
were performed, the second during superfusion with 50 nmol/L secretin. Although basal p h i was similar, the maximal rate of pHi alkalinization after C1- removal and
the maximal rate of pHi recovery after C1- readmission were higher (P < 0.05) after treatment with the 2 corticosteroids (Table 1 and Figure 4A), and this was
found both in basal conditions and during stimulation with secretin. This indicates enhanced activity of the
C I - / H C O 3- exchanger in corticosteroid-treated rats. When a potent GcR antagonist, RU-486 (1 mg, 3 times
daily), was administered together with dexamethasone
(0.9 mg, 3 times daily) for 2 days, the activity of C1-/ H C O 3- exchanger in IBDUs (Cl- removal = 0.140 ---
0.012 pHU/min ; C1- readmission = 0.165 + 0.014 pHU/min ; n = 15) was similar to untreated controls
(C1- removal = 0.135 + 0.01 pHU/min ; C1- readmis- sion = 0.159 - 0.014 pHU/min ; n = 12) but lower (P < 0.05) compared with IBDUs from rats treated with
dexamethasone alone (C1- removal = 0.189 + 0.013 pHU/min ; C1- readmission = 0.245 + 0.016 p H U / min; n = 14). This indicates that activation of GcR by
corticosteroids is crucial for the stimulation of C1-/
H C O S exchanger activity in IBDUs. The activity of the Na+-HCO3 - cotransporter was
measured, as described, 2,3,22 by evaluating the maximal
rate of pHi recovery from 20 mmol/L NH4C1 acid pulse in KRB and with IBDU exposed to ethyl-n-isopropyl-
amiloride (EIPA, a N a + / H ÷ exchanger inhibitor) at the moment of NH4C1 withdrawal, to exclude the influence
of the N a + / H + exchanger. IBDU isolated from rats treated (2 days) with dexamethasone or budesonide
showed similar basal pHi, nadir pHi after NH4C1 pulse, and recovery rates from the acid load to control IBDU
(Table 2), indicating that the activity of Na+-HCO3 - corransporter was unaffected by corticosteroids.
The activity of the N a + / H + exchanger was mea- sured 2,3,22 in both IBDUs cultured and perfused in the
Table 1. Ef fec t o f T r e a t m e n t With D e x a m e t h a s o n e or B u d e s o n i d e on Basal and Sec re t i n -S t imu la ted C l - / H C 0 3 Exchanger Act iv i ty in IBDU
Control Control-secretin Dexamethasone Dexa + secretin (n = 12) (n = 12) (n = 14) (n = 14)
Basal pH, 7.19 + 0 .011 7.20 _+ 0 .011 7.21 + 0 ,010 7.21 + 0.010 Cl- removal
Delta pH~ 0.24 +_ 0.012 0.28 -+ 0 .012 0.25 _+ 0.011 0.29 _+ 0.011 pH (max U/min) 0,135 _+ 0.010 0 .204 + 0 .011 a 0 .189 _ 0.013 b 0 .294 _+ 0.014 a,b H + flux (mmol/min) 8.13 _+ 0.799 12.03 +_ 1.04 a 11.40 + 1.17 b 17.80 + 1.459 a,b
Cl- readmission
pH (max U/min) 0.167 + 0 .013 0 .248 + 0,012 a 0 .245 + 0 .016 ~ 0 .338 -+ 0.017 a,b H + flux (mmol/min) 10.23 + 0.98 15.63 + 1.29 a 15.93 _+ 1.39 b 22.03 -+ 1.86 a,b
Control Control-secretin Budesonide Bude + secretin (n = 14) (n = 14) (n = 16) (n = 16)
Basal pH, 7.22 + 0 .011 7.22 + 0 .012 7.24 _+ 0 .012 7,23 -+ 0,013 Cl- removal
Delta pH, 0.22 _+ 0 .009 0.25 _+ 0 .010 0.24 _+ 0.009 0,28 + 0.011 pH (max U/min) 0.123 - 0 ,011 0.179 _+ 0 .012 a 0 .186 _+ 0.013 b 0 .301 + 0 .014 a.b H + flux (mmol/min) 7.59 + 0.69 10.69 _+ 1.01 a 11.09 + 1.05 b 17.98 + 1.493 a.b
Cl- readmission
pH (max U/min) 0.164 + 0 .014 0.231 _+ 0.015 a 0 ,239 + 0 .015 b 0 .341 -+ 0 .016 a,b H + flux (mmol/min) 10.13 + 1.11 14.14 + 1.31 a 14.11 + 1.36 b 21.89 + 2.04 a,b
NOTE. Two sequential CI- removal/readmission maneuvers were performed, the second during exposure to 50 nmol /L secretin, in IBDU isolated from rats treated for 2 days with dexamethasone or budesonide or from control rats. Data are mean _+ SE. Treatment with dexamethasone or budesonide stimulates pH, changes induced by CI removal/readmission either in basal condition or during secretin exposure. ap < 0.05 vs. corresponding control values in the absence of secretin. bp < 0.05 VS. correspondent values in IBDU isolated from untreated control rats.

t 0 6 4 ALVARO ET AL. GASTROENTEROLOGY Vol. 122, No. 4
A Krb Glucon ate Krb Gluconate Krb
8 0 ~' t t t
7.6
7.4
~ 7.2
7.0
8.0
7.8
7.6
7.4
"~ 7.2 ¸
Dexamethasone 2 days + 36 hrs incub.
s e c r e t i n [ C o n t r o l s ]
400 800 1200 1600 TIME (sec.)
B H e p e ~ N H 4 C ~ Hepes ; H 4 C I EIPA
8.0 7.8 t [ Dexamethasone 7.6 [ 2days+36 hrs incub. 7.4 7.2
~ 7.8 -' 6.8
6.6 6.4
8.0
7.6
7.4 C o n t r o l s 7.2
-~ 7.0 - 6.8
6.6 6.4
0 400 800 1200 1600 2000 TIME (sec.)
Figure 4. Effect of a 2-day treatment with dexamethasone on the activity of the (A) CI-/HC03- and (/3) Na+/H + exchangers in IBDU. (A) Two consecutive acute Cl- removal/readmission maneuvers were performed, the second during superfusion with 50 nmol/L secretin. The maximal rate of pH, alkalinization after Cl removal and the maximal rate of pH, recovery after Cl- readmission were higher (P < 0.05) after treatment with dexamethasone (top) than controls (bot- tom), both in basal conditions and during secretin superfusion. (B) IBDU cultured and perfused with solutions nominally free of bicarbon- ate (L15 and HEPES, respectively). The recovery rates from NH4Ci- induced acidification were higher (P < 0.05) when measured in IBDUs from rats treated with dexamethasone (top) than from control rats (bottom). When EIPA was administered, only marginal pH, recovery was seen indicating that, in HEPES, the Na+/H + exchanger was the only transporter involved in pH~ recovery. Experiments are represen- tative of data reported in Tables 1 and 2.
absence (L15 and HEPES, respectively) or presence of bicarbonate (o~-MEM and KRB). In HEPES, the
N a + / H + exchanger was the only transport process in- volved in the recovery from acid load (i.e., NH4C1 pulse) and, in fact, when EIPA was administered at the moment of NH4C1 withdrawal (Figure 4B), only marginal pHi recovery was observed. Basal pHi was higher (P < 0.05;
Table 2) in IBDUs isolated from rats treated with dexa-
methasone or budesonide than in controls. Nadir p H i
after NH4C1 withdrawal was similar, but the recovery
rates from NH4C1 acidification were higher (P < 0.05)
when measured in IBDUs from rats treated with dexa-
methasone or budesonide (Table 2 and Figure 4B) than
controls. This indicates that the Na+ /H + exchanger ac-
tivity was enhanced by corticosteroids. In addition, dex-
amethasone-treated and control IBDUs showed a similar
pH1 when exposed to 50 ~mol/L EIPA in HEPES (dexa-
methasone + EIPA = 6.89 + 0.010, n = 5; controls +
EIPA = 6.90 + 0.011, n = 5) or 1 I~mol/L cariporide, a specific NHE1 inhibitor 7 (dexamethasone + caripo-
ride = 6.88 + 0.013, n = 6; controls + cariporide =
6.89 + 0.011, n = 5), suggesting that the higher basal
pHi of corticosteroid-treated IBDUs is caused by activa-
tion of the Na+ /H + exchanger, specifically the NHE1
isoform. To evaluate the activity of the N a + / H + ex-
changer in the presence of bicarbonate, IBDUs cultured
and perfused with oL-MEM and KRB, respectively, were
exposed to H2-DIDS (4,4'-diisothiocyanatodihydrostil- bene-2,2'-disulfonic acid disodium salt, 0.5 mmol/L;
Molecular Probes Inc., Eugene, OR) to exclude the in-
fluence of the Na+-HCO3 - cotransporter and then to
NH4C1 acid pulse. Under these experimental conditions,
basal pHi as well as nadir pHi after NH4C1 withdrawal were similar, but the recovery rates from NH4Cl-induced
acidification were higher (P < 0.05) after treatment with
dexamethasone or budesonide (Table 2). This indicates
that the Na+/H + exchanger activity was enhanced by cor-
ticosteroid treatment also in the presence of bicarbonate.
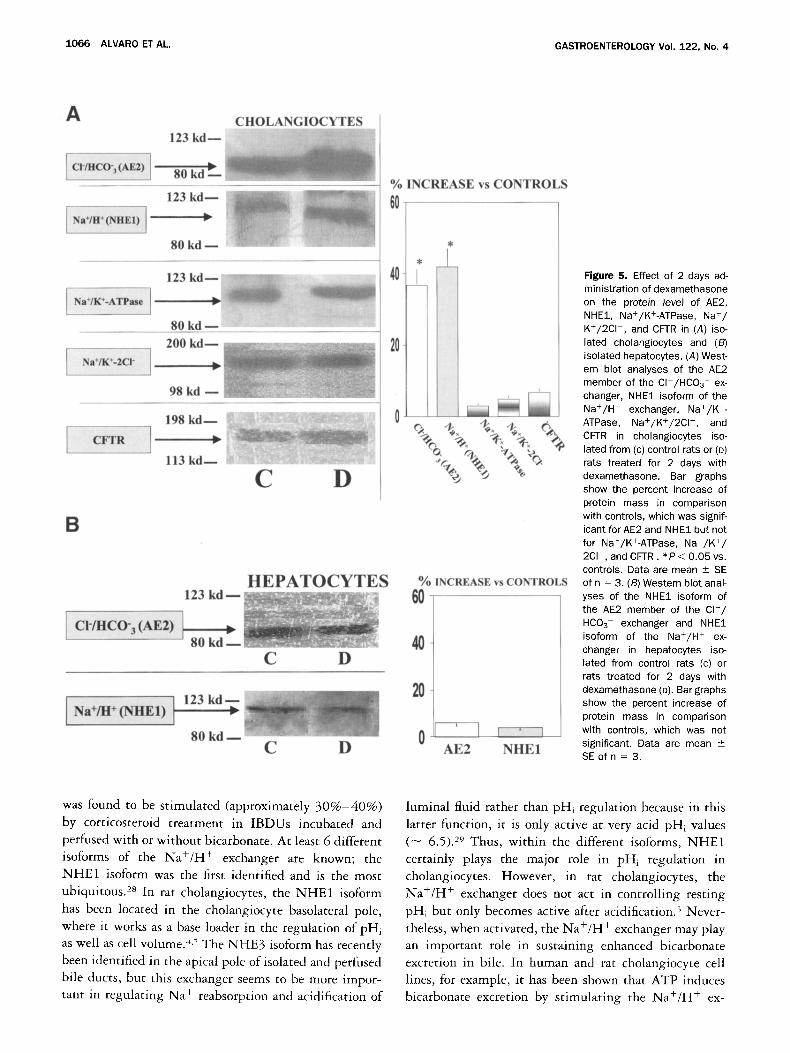
The protein mass of AE2 and N H E 1 were significantly
(P < 0.05) increased (AE2 = + 37% -+ 4%, n = 3;
NHE1 = + 42% + 3%, n = 3; Figure 5A) in cholan-
giocytes, but not in hepatocytes (Figure 5B) isolated
from rats treated for 2 days with dexamethasone, com-
pared with untreated controls.
We have also evaluated the effect of a 2-day treatment
with dexamethasone on the protein expression of the
Na+/K+-ATPase, Na+/K+-2C1 - , and CFTR in isolated
cholangiocytes (Figure 5A), but no changes were found.
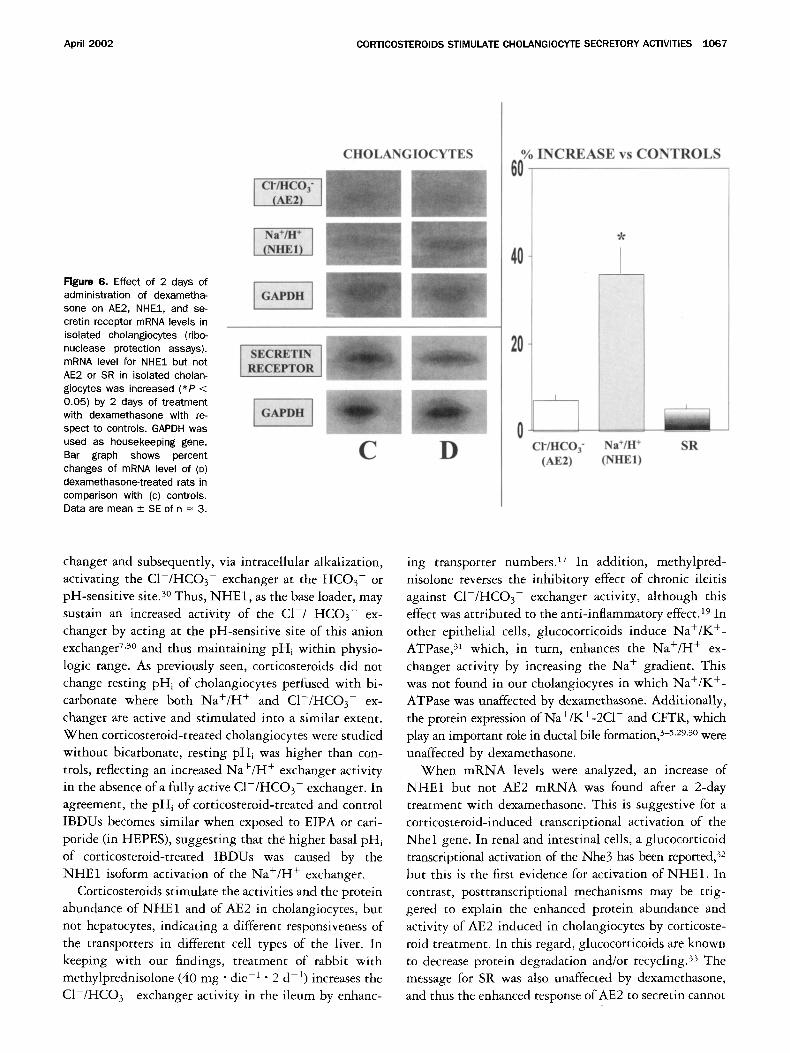
The m R N A levels for NHE1 in isolated cholangio-
cytes were increased (P < 0.05) by the 2-day treatment
with dexamethasone compared with controls ( + 36% +
7%; n = 3, Figure 6), whereas m R N A levels for AE2,
SR, and G A P D H , the housekeeping gene, were un-
changed.
We have excluded that changes in protein mass and m R N A levels were caused by an effect of corticosteroids
on cholangiocyte mass because bile duct volume after 2
days of dexamethasone (0.29% + 0.02%, n = 4) was
similar to controls (0.28% _+ 0.03%, n = 4).

April 2002 CORTICOSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1065
Table 2. Effect of Treatment With Dexamethasone or Budesonide on the Activities of the Na+:HC03 - Cotransporter and Na+/H + Exchanger
Nadir pH, after pH~ recovery rate H + flux Conditions Basal pHi NH4Cl pulse (U/min) (mmol/min)
Na+:HC03 - Cotransporter KRB + EIPA controls (n = 16) 7.20 _+ 0.012 KRB + EIPA DEXA (n = 16) 7.21 + 0.013 KRB + EIPA BUDE (n = 16) 7.22 +- 0.014
Na+/H + Exchanger Hepes controls (n = 18) 7.00 4- 0.010 Hepes DEXA (n = 18) 7.06 4- 0.011 a Hepes BUDE (n = 18) 7.08 _+ 0.013 a KRB + H2-DIDS controls (n = 16) 7.06 _+ 0.011 KRB + H2-DIDS DEXA (n = 16) 7.10 4- 0.013 a KRB + H2-DIDS BUDE (n = 16) 7.11 -+ 0.013 a
6.68 4- 0.014 0.159 _+ 0.018 10.09 4- 0.91 6.70 _+ 0.013 0.164 _+ 0.019 10.72 4- 0.96 6.71 +_ 0.015 0.169 _+_ 0.021 10.90 + 0.93
6.61 _+ 0.014 0.147 -+ 0.015 10.35 _+ 1.03 6.64 + 0.013 0.216 4- 0.016 a 13.93 -+ 1.36 a 6.65 4- 0.015 0.237 4- 0.017 a 14.41 4- 1.39 a 6.64 4- 0.012 0.048 _+ 0.002 3.49 4- 0.40 6.66 _+ 0.013 0.072 _+ 0.003 5.14 _+ 0.53 a 6.67 + 0.013 0.079 _+ 0.003 5.37 4- 0.57 a
NOTE. The activity of the Na+:HC03 - cotransporter was evaluated in IBDU, cultured in bicarbonate enriched media (e-MEM), and perfused with bicarbonate-rich solution (KRB) by measuring the maximal rate of pH~ recovery from 20 mmol/L NH4CI acid pulse and during exposure to amiloride analogue EIPA (50 i~moi/L), administered at the moment of NH4CI withdrawal. The activity of the Na+/H + exchanger was measured either in IBDU cultured and perfused with solutions nominally free of bicarbonate (L15 and HEPES, respectively) or enriched with bicarbonate (e-MEM and KRB, respectively). In HEPES, the maximal rate of PHi recovery from 20 mmol/L NH4CI acid pulse was measured. In KRB, IBDUs were superperfused with 0.5 mmol/L H2-DIDS to exclude the influence of the Na÷:HC03 - cotransporter. Treatment with dexamethasone or budesonide for 2 days increased the activity of the Na+/H ÷ exchanger both in HEPES and KRB, whereas that of the Na+:HC03 cotransporter was unaffected. Data are mean _+ SE. ap < 0.05 vs. the correspondent values measured in untreated control IBDU.
Discussion
The main findings emerging from this study demonstrate that: (1) dexamethasone and budesonide, at pharmacologic doses, stimulate bile flow and bicarbonate biliary excretion after a 2-day administration, but not after acute infusion; (2) the stimulatory effect of pro- longed treatment with corticosteroids on bicarbonate excretion is associated with increased activities of C1-/ HCO 3- and Na+H + exchangers in IBDUs as well as enhanced protein abundance of the AE2 member of CI - /HCO 3- exchanger and NHE1 isoform of the Na+/H + exchanger in pure preparations of isolated
cholangiocytes; (3) mRNA levels of NHE1 but not AE2 in isolated cholangiocytes were increased by prolonged treatment with corticosteroids; and (4) the stimulatory effects of corticosteroids on bicarbonate biliary excretion and on C1-/HCO 3- exchanger require activation of GcR, their effects being blocked by RU-486, a specific GcR antagonist. 25
We first demonstrated the expression of GcR in situ in the rat liver by immunohistochemistry and in isolated cholangiocytes by both RT-PCR and immunoblotting, and this confirms previous studies in the human liver, e6
The protein mass of GcR (Western blot) was increased in rats treated for 2 days with dexamethasone as well as in BDL rats where corticosteroid serum levels are increased as a consequence of cholestasis. 2v This indicates that GcR expression in cholangiocytes is induced by corticosteroids and suggests a possible role of these hormones in sus- taining the increased secretory activities of prolifer-
ating cholangiocytes typical of BDL rats. 2° Then, we
investigated the effect of pharmacologic doses of dexa- methasone, a synthetic corticosteroid, and budesonide, a corticosteroid of new generation, 12 on bile flow and bicarbonate biliary excretion. Although these 2 cortico-
steroids showed no effect after acute intravenous admin- istration, a 2-day treatment with dexamethasone or
budesonide induced a significant increase in bile flow and bicarbonate biliary excretion without significant effects
on liver cell mass or on bile duct mass. We then assessed whether a 2-day treatment with corticosteroids influ-
ences the activity, protein mass, and mRNA level of electrolyte transporters in IBDUs or isolated cholangio- cytes. IBDUs isolated from rats treated with dexameth-
asone or budesonide showed a C1-/HCO 3- exchanger activity that was increased by 40%-50% over controls
both in basal conditions and after stimulation with se-
cretin. Protein expression of AE2, the member CI- / HCO 3- exchanger identified in cholangiocytes, 4,6 was also increased by corticosteroid treatment in isolated
cholangiocytes but not in hepatocytes, suggesting that the activation of this anion exchanger in the intrahepatic
biliary epithelium plays a role in the enhanced bicarbon-
ate biliary excretion induced by prolonged treatment with corticosteroids. In the regulation of pHi in cholan- giocytes, the CI - /HCO3- exchanger works as a base extruder, and in this function it is counterbalanced by 2 base loaders, the Na+:HCO} - cotransporter and the
Na+/H + exchanger. Although the Na+:HCO3 - cotrans- porter was unaffected, the Na+/H + exchanger activity
1066 ALVARO ET AL. GASTROENTEROLOGY Vol. 122, No. 4
A
CI'/HCO'3 (AE2) ]
CHOLANGIOCYTES 123 k d m . < '.-°:""o =~-~:" :
80 kd ~ :II!
Na+/H+ (NHE1) ]
123 k d ~
80 kd
I Na+/K+-ATPase
123 k d - - ~ . .
80 kd ~ ' ~ ' ~
Na+/K+2CI - 2 0 0 k d - -
9 8 k d - -
CT'I'R
C D
B
[ Cl-/HC ,(AE2)
H E P A T O C Y T E S 123 kd ~ - ~
1 .= I 8 0 k d m - - - - _ . . . . . , -~.~
C D
[ Na+/W(r m D I 123 kd. 7 1
80 kd m C D
% INCREASE vs CONTROLS
60
40
20
)
%°°.% +'. +,.
°/o INCREASE vs CONTROLS 60
40
20
0 I I A E 2 N H E 1
Figure 5. Effect of 2 days ad- ministration of dexamethasone on the protein level of AE2, NHE1, Na+/K+-ATPase, Na+/ K+/2Cl - , and CFTR in (A) iso- lated cholangiocytes and (B) isolated hepatocytes. (A) West- ern blot analyses of the AE2 member of the Cl /HCO3 ex- changer, NHE1 isoform of the Na+/H + exchanger, Na+/K +- ATPase, Na+/K+/2CI- , and CFTR in cholangiocytes iso- lated from (c) control rats or (D) rats treated for 2 days with dexamethasone. Bar graphs show the percent increase of protein mass in comparison with controls, which was signif- icant for AE2 and NHE1 but not for Na+/K+-ATPase, Na+/K+/ 2Cl , and CFTR. * P < 0.05 vs. controls. Data are mean + SE of n = 3. (B) Western blot anal- yses of the NHE1 isoform of the AE2 member of the C l - / HCO 3- exchanger and NHE1 isoform of the Na+/H + ex- changer in hepatocytes iso- lated from control rats (c) or rats treated for 2 days with dexamethasone (D). Bar graphs show the percent increase of protein mass in comparison with controls, which was not significant. Data are mean _+ SE of n - 3.
was found to be stimulated (approximately 3 0 % - 4 0 % ) by corticosteroid treatment in IBDUs incubated and perfused with or without bicarbonate. At least 6 different isoforms of the Na+/H + exchanger are known; the NHE1 isoform was the first identified and is the most ubiquitous. 2s In rat cholangiocytes, the NHE1 isoform has been located in the cholangiocyte basolateral pole, where it works as a base loader in the regulation of pHi as well as cell volume. <~ The NHE3 isoform has recently been identified in the apical pole of isolated and perfused bile ducts, but this exchanger seems to be more impor- tant in regulating Na + reabsorption and acidification of
luminal fluid rather than pHi regulation because in this latter function, it is only active at very acid pHi values (-- 6.5). 29 Thus, within the different isoforms, NHE1 certainly plays the major role in pHi regulation in cholangiocytes. However, in rat cholangiocytes, the Na+/H + exchanger does not act in controlling resting pHi but only becomes active after acidification) Never- theless, when activated, the Na+/H + exchanger may play an important role in sustaining enhanced bicarbonate excretion in bile. In human and rat cholangiocyte cell lines, for example, it has been shown that ATP induces bicarbonate excretion by stimulating the Na+/H + ex-
April 2002 CORTICOSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1067
Figure 6. Effect of 2 days of administration of dexametha- sone on AE2, NHE1, and se- cretin receptor mRNA levels in isolated cholangiocytes (ribo- nuclease protection assays). mRNA level for NHE1 but not AE2 or SR in isolated cholan- giocytes was increased (*P < 0.05) by 2 days of treatment with dexamethasone with re- spect to controls. GAPDH was used as housekeeping gene. Bar graph shows percent changes of mRNA level of (D) dexamethasone-treated rats in comparison with (c) controls. Data are mean +_ SE of n = 3.
CHOLANGIOCYTES
[ ctf=qcof
RI~CEPTOR
t C D
% I N C R E A S E vs C O N T R O L S
60
40
20
0 C I - [ I - I C O 3 -
(AE2)
qt
Na+/H + SR (NH~I)
changer and subsequently, via intracellular alkalization, activating the CI- /HCO 3- exchanger at the HCO3- or pH-sensitive site. 3° Thus, NHE 1, as the base loader, may sustain an increased activity of the C1-/ HCO 3- ex- changer by acting at the pH-sensitive site of this anion exchanger 7,3° and thus maintaining pHi within physio- logic range. As previously seen, corticosteroids did not change resting pHi of cholangiocytes perfused with bi- carbonate where both Na+/H + and C1-/HCO 3- ex- changer are active and stimulated into a similar extent. When corticosteroid-treated cholangiocytes were studied without bicarbonate, resting ph i was higher than con- trols, reflecting an increased Na+/H + exchanger activity in the absence of a fully active CI- /HCO 3- exchanger. In agreement, the pH i of corticosteroid-treated and control IBDUs becomes similar when exposed to EIPA or cari- poride (in HEPES), suggesting that the higher basal pHi of corticosteroid-treated IBDUs was caused by the NHE1 isoform activation of the Na+/H ÷ exchanger.
Corticosteroids stimulate the activities and the protein abundance of NHE1 and of AE2 in cholangiocytes, but not hepatocytes, indicating a different responsiveness of the transporters in different cell types of the liver. In keeping with our findings, treatment of rabbit with methylprednisolone (40 mg• die -1 • 2 d -I) increases the CI- /HCO 3- exchanger activity in the ileum by enhanc-
ing transporter numbers. 17 In addition, methylpred- nisolone reverses the inhibitory effect of chronic ileitis against C1-/HCO 3- exchanger activity, although this effect was attributed to the anti-inflammatory effect, i9 In other epithelial ceils, glucocorticoids induce Na+/K +- ATPase, 31 which, in turn, enhances the Na+/H + ex- changer activity by increasing the Na + gradient. This was not found in our cholangiocytes in which Na+/K +- ATPase was unaffected by dexamethasone. Additionally, the protein expression of Na+/K+-2C1 - and CFTR, which play an important role in ductal bile formation, 3-5,29,3° were unaffected by dexamethasone.
When mRNA levels were analyzed, an increase of NHE1 but not AE2 mRNA was found after a 2-day treatment with dexamethasone. This is suggestive for a corticosteroid-induced transcriptional activation of the Nhel gene. In renal and intestinal cells, a glucocorticoid transcriptional activation of the Nhe3 has been reported) 2 but this is the first evidence for activation of NHE1. In contrast, posttranscriptional mechanisms may be trig- gered to explain the enhanced protein abundance and activity of AE2 induced in cholangiocytes by corticoste- roid treatment. In this regard, glucocorticoids are known to decrease protein degradation and/or recycling) 3 The message for SR was also unaffected by dexamethasone, and thus the enhanced response of AE2 to secretin cannot

1068 ALVARO ET AL. GASTROENTEROLOGY Vol. 122, No. 4
be due to enhanced transcription of this receptor. We cannot exclude, however, that secretin signal transduc- tion pathway is influenced by corticosteroids.
The stimulation of both bicarbonate excretion and the C1-/HCO 3- exchanger by corticosteroids requires acti- vation of GcRs being inhibited by their specific antag- onist, RU-486. Many biological effects ofglucocorticoids are mediated by GcR, 34 and the results of the present study indicate that the modulatory effect on cholangio- cyte secretory processes is an additional example.
Certainly, the increased cholangiocyte secretory activ- ities are not the only mechanism of the stimulatory effect of corticosteroids on bile flow. It has been shown that prolonged 35 treatment with corticosteroids in the rat or even single bolus of hydrocortisone in the dog 36 induce an increased erythritol clearance (a marker of canalicular flow) without influence, as in our study, on bile salt biliary secretion. Thus, when considering the present and previous studies, the choleretic effect of corticosteroids seems to occur because of stimulation of bile salt-inde- pendent canalicular bile flow and ductal flow.
In conclusion, we have shown that the secretory pro- cesses underlying bicarbonate excretion in the intrahe- patic biliary epithelium are modulated by corticoste- roids, of which the effects are mediated through the glucocorticoid receptor. Wi th regard to the latter, it is noteworthy that UDCA may activate glucocorticoid re- ceptors, 37 and this could represent a mechanism of the anticholestatic effect of this drug. Our findings could have clinical implications for the treatment of cholan- giopathies where bicarbonate excretion is impaired and where corticosteroids could act not only as anti-inflam- matory agents but also by improving the secretory pro- cesses. Finally, our findings could introduce a mechanism for the beneficial effect of short treatment with cortico- steroids on the intrahepatic cholestasis of a different etiology, the so-called corticosteroid whitewash. ~6
References 1. Roberts SK, Ludwig J, LaRusso NF. The pathobiology of biliary
epithelia. Gastroenterology 1997;112:269-279. 2. Mennone A, Alvaro D, Cho WK, Boyer JL. Isolation of small
polarized bile duct units. Proc Natl Acad Sci U S A 1995;92: 6527-6531.
3. Alvaro D, Cho WK, Mennone A, Boyer JL. Effect of secretin on intracellular pH regulation in isolated rat bile duct epithelial cells. J Clin Invest 1993;92:1314-1325.
4. Spirli C, Granato A, Zsembery A, Anglani F, Okolics~nyi L, La- Russo NF, Crepaldi G, Strazzabosco M. Functional polarity of Na+/H + and CI-/HC03- exchangers in rat cholangiocyte cell line. Am J Physiol 1998;275:G1236-G1245.
5. Fitz JG, Basavappa S, McGill J, Melhus O, Cohn JA. Regulation of membrane chloride currents in rat bile duct epithelial cells. J Clin Invest 1993;91:319-328.
6. Medina JF, Martinez-Anso' E, V&zquez J J, Prieto J. Decreased
anion exchanger 2 immunoreactivity in the liver of patients with primary biliary cirrhosis. Hepatology 1997;25:12-17.
7. HLibner C, Stremmel W, Elsing C. Sodium, hydrogen exchange type 1 and bile ductular secretory activity in the guinea pig. Hepatology 2000;31:562-571.
8. Prieto J, Garcia N, Marti-Climent JM, Penuelas I, Richter JA, Medina JF. Assessment of biliary bicarbonate secretion in hu- mans by positron emission tomography. Gastroenterology 1999; 117:167-172.
9. Melero S, Spirli C, Medina JF, Durier E, Neuberger J, Prieto J, Strazzabosco M. Defective regulation of Cl-/HC03- and Na+/H + exchangers in cholangiocytes from PBC patients (abstr). Hepatol- ogy 2000;32:299a.
10. Colombo C, Batte77ati PM, Strazzabosco M, Podda M. Liver and biliary problems in cystic fibrosis. Semin Liver Dis 1998;18:227- 235.
11. Beuers U, Boyer JL, Paumgartner G. Ursodeoxycholic acid in cholestasis: potential mechanisms of action and therapeutic applications. Hepatology 1998;28:1449-1453.
12. Ryrfeldt A, Andersson P, Edsbacker S, Tonnesson M, Davies D, Pauwels R. Pharmacokinetics and metabolism of budesonide, a selective glucocorticoid. Eur J Respir Dis 1982;122(suppl):86- 95.
13. Leuschner M, Maier KP, Schlichting J, Strahl S, Herrmann G, Dahm HH, Ackermann H, Happ J, Leuschner U. Oral budesonide and ursodeoxycholic acid for treatment of primary biliary cirrho- sis: results of a prospective double-blind trial. Gastroenterology 1999;117:918-925.
14. Angulo P, Jorgensen RA, Keach JC, Dickson ER, Smith C, Lindor KD. Oral budesonide in the treatment of patients with primary biliary cirrhosis with a suboptimal response to ursodeoxycholic acid. Hepatology 2000;31:318-323.
15. van Hoogstraten H J, Vleggaar FP, Boland G J, van Steenbergen W, Griffioen P, Hop WC, van Hattum J, van Berge Henegouwen GP, Schalm SW, van Buuren HR. Budesonide or prednisone in com- bination with ursodeoxycholic acid in primary sclerosing cholan- gitis: a randomized double-blind pilot study. Am J Gastroenterol 2000;95:2015-2022.
16. Sherlock S, Dooley J. Diseases of the liver and biliary system. lOth ed. Oxford: Biackwell Scientific, 1997:1-714.
17. Foster ES, Zimmerman TW, Hayslett JP, Binder HJ. Corticosteroid alteration of active electrolyte transport in rat distal colon. Am J Physiol 1983;245:G688-G695.
18. Unwin R, Capasso G, Giebisch G. Bicarbonate transport along the loop of Henle effects of adrenal steroids. Am J Physiol 1995; 268:F234-F239.
19. Coon S, Sundaram U. Mechanism of glucocorticoid-mediated reversal of inhibition of CI-/HC03- during chronic ileitis. Am J Physiol 2000;278:G570-G577.
20. Alpini G, Lenzi R, Sarkozi L, Tavoloni N. Biliary physiology in rats with bile ductular cell hyperplasia. J Clin Invest 1988;81:569- 578.
21. Alvaro D, Alpini G, Onori P, Franchitto A, Glaser SS, Le Sage G, Folli F, Gaudio E. Estrogens stimulate proliferation of rat intrahe- patic biliary epithelium. Gastroenterology 2000;119:1681-1691.
22. Alvaro D, Della Guardia P, Bini A, Gigliozzi A, Piat C, Capocaccia L. Effect of glucagon on intracellular pH regulation in isolated rat hepatocyte couplets. J Clin Invest 1995;96:665-675.
23. Miesfeld R, Rusconi S, Godowski P J, Maler BA, Okret S, WikstrSm AC, Gustafsson JA, Yamamoto KR. Genetic complementation to a glucocorticoid receptor deficiency by expression of cloned re- ceptor cDNA. Cell 1986;46:389-399.
24. Fort P, Marty L, Piechaczyk M, el Sabrouty S, Dani C, Jeanteur P, Blanchard JM. Various rat adult tissues express only one major mRNA species from the glyceraldehyde-3-phosphate-dehydroge- nase multigenic family. Nucleic Acids Res 1985;13:1431-1442.
25. Bardon S, Vignon F, Chalbos D, Rochefort H. RU486, a progestin

April 2002 CORT|COSTEROIDS STIMULATE CHOLANGIOCYTE SECRETORY ACTIVITIES 1069
and glucocorticoid antagonist, inhibits the growth of breast can- cer cells via the progesterone receptor. J Clin Endocrinol Metab 1985;60:692-697.
26. Oakley RH, Webster JC, Sar M, Parker CR, Cidlowski JA. Expres- sion and subcellular distribution of the beta-isoform of the human glucocorticoid receptor. Endocrinology 1997;138:5028-5038.
27. Tjandra K, Kubes P, Rioux K, Swain MG. Endogenous glucocorti- coids inhibit neutrophil recruitment to inflammatory sites in cho- lestatic rats. Am J Physiol 1996;270:G821-G825.
28. Fliegel L, Murtazina R, Dibrov P, Harris C, Moor A, Fernandez- Rachubinski FA. Regulation and characterization of the Na+/H + exchanger. Biochem Cell Biol 1998;76:735-741.
29. Singh SKG, Cavestro M, Mennone A, Boron WF, Boyer JL. Distinct apical and basolateral Na+/H + exchangers in isolated intrahe- patic bile duct segments (abstr). Gastroenterology 1997;112: A1384.
30. Zsembery A, Spirli C, Granato A, LaRusso NF, Okolicsanyi L, Strazzabosco M. Purinergic regulation of acid/base transport in human and rat biliary epithelial cell lines. Hepatolegy 1998;28: 914-920.
31. Rayson BM, Edelman IS. Glucocorticoid stimulation of Na-K-AT- Pase in superfused distal segments of kidney tubules in vitro. Am J Physiol 1982;243:F463-F470.
32. Kandasamy RA, Orlowski J. Genomic organization and glucocor- ticoid transcriptional activation of the rat Na+/H + exchanger Nhe3 gene. J Biol Chem 1996;271:10551-10559.
33. Ambuhl PM, Yang X, Peng Y, Preisig PA, Moe OW, Alpern RJ. Glucocorticoids enhance acid activation of the Na+/H + ex- changer 3 (NHE3). J Clin Invest 1999;103:429-435.
34. Bamberger CM, Schulte HM, Chrousos GP. Molecular determi- nants of glucocorticoid receptor function and tissue sensitivity to glucocorticoids. Endocr Rev 1996;17:245-261.
35. Zsigmond G, Solymoss B. Increased canalicular bile production induced by pregnenolone-16~x-carbonitrile, spironolattone and cortisol in rats. Proc Soc Exp Biol Med 1974;145:631-635.
36. Macarol V, Morris TQ, Baker K J, Bradley SE. Hydrocortisone choleresis in the dog. J Clin Invest 1970;49:1714-1723.
37. Tanaka H, Makino I. Ursodeoxycholic acid-dependent activation of the glucocorticoid receptor. Biochem Biophys Res Commun 1992;188:942-948.
Received March 26, 2001. Accepted December 6, 2001. Address requests for reprints to: Domenico Alvaro, M.D., Diparti-
mento di Medicina Clinica, Divisione di Gastroenterologia, Viale Uni- versit~ 37, 00185 Rome, Italy. e-mail: [email protected]; fax: (39) 06-444-0806.
Supported by MURST (40% funds) grants MM9806210866, MM06215421 /2 , MM06215421, and 9906022315-003 (to D.A. and A.B.), by a grant award from Scott and White Hospital and the Texas A&M University System, by NIH grant DK54811, and by a VA Merit Award (to G.A.).




















