
HIGH HUNTING PRESSURE SELECTS FOR EARLIER BIRTH DATE: WILD BOAR AS A CASE STUDY

Animal Reproduction Science 82–83 (2004) 321–339
Post-testicular sperm environment and fertility
J.-L. Gatti∗, S. Castella, F. Dacheux, H. Ecroyd, S. Métayer,V. Thimon, J.-L. Dacheux
Equipe “Gamète Male et Fertilité”, UMR 6175 INRA-CNRS-Un. de Tours-Haras, Station de Physiologie de laReproduction et des Comportements, Institut National de la Recherche Agronomique, 37380 Nouzilly, France
Abstract
When mammalian spermatozoa exit the testis, they show a highly specialized morphology; how-ever, they are not yet able to carry out their task: to fertilize an oocyte. This property, that includesthe acquisition of motility and the ability to recognize and to fuse with the oocyte investments, isgained only after a transit through the epididymis during which the spermatozoa from the testistravel to the vas deferens. The exact molecular mechanisms that turn these cells into fertile gametesstill remain mysterious, but surface-modifying events occurring in response to the external mediaare key steps in this process. Our laboratory has established cartographies of secreted (secretomes)and present proteins (proteomes) in the epididymal fluid of different mammals and have shown theregionalized variations in these fluid proteins along the epididymis. We have found that the mainsecreted proteins are common in different species and that enzymatic activities, capable of control-ling the sperm surface changes, are present in the fluid. Our studies also indicate that the epididymalfluid is more complex than previously thought; it contains both soluble and particulate compart-ments such as exosome-like vesicles (epididymosomes) and certainly specific glycolipid-proteinmicelles. Understanding how these different compartments interplay to modify sperm componentsduring their transit will be a necessary step if one wants to control and to ameliorate sperm qualityand to obtain valuable fertility markers helpful to establish a male fertility based genetic selection.© 2004 Elsevier B.V. All rights reserved.
Keywords: Epididymis; Spermatozoa; Fertility; Male genital tract; Proteins
1. Introduction
During the past few decades, the artificial insemination (AI) technique has allowed rapidgenetic progress, a driving force for competitiveness in animal farming. Artificial insemi-nation is based on the use of a small number of males selected for different genetic criteria.
∗ Corresponding author. Tel.:+33 247 427 803; fax:+33 247 427 743.E-mail address: [email protected] (J.-L. Gatti).
0378-4320/$ – see front matter © 2004 Elsevier B.V. All rights reserved.doi:10.1016/j.anireprosci.2004.05.011
322 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
The dissemination of the superior genetic traits of these males is limited by the quantityand the quality of their gametes. Gamete production may be controlled by various factors,each contributing to the heritability of this parameter. In contrast, fertility, which involvesa complex physiological process and is controlled by multiple genetic loci and variousother factors (involving the central nervous system and nutrition among others) and gametedifferentiation and maturation, is not correlated with heritability (Nadarajah et al., 1988;Humblot and Ducrocq, 1996).
Fertile spermatozoa result from a successive series of complex morphological and sur-face modifications. The main morphological changes occur during spermatogenesis, wherea round germ cell turns into a highly organized cell with a specific shape, a greatly con-densed and “inactive” nucleus and a flagellar apparatus. During this process, the sur-
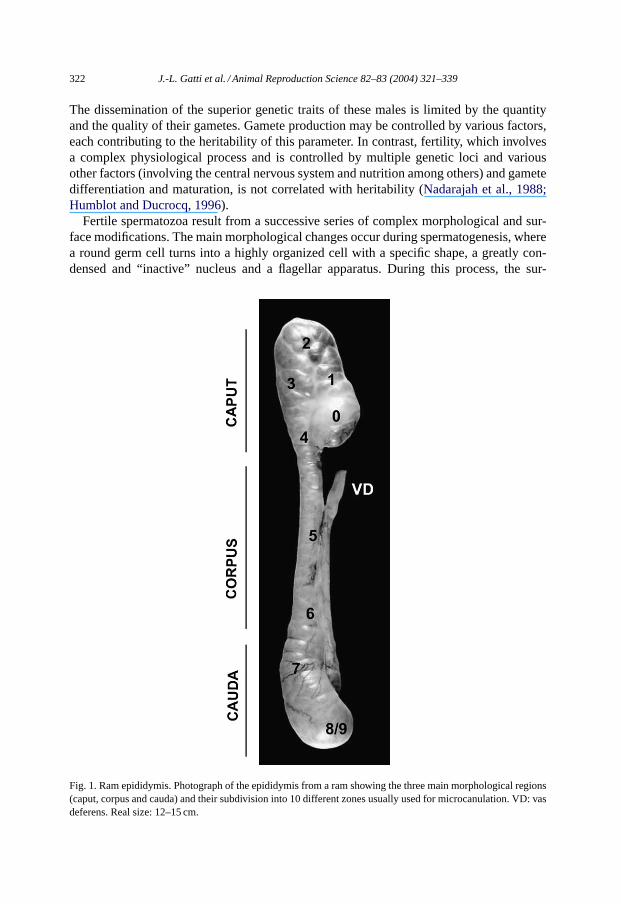
Fig. 1. Ram epididymis. Photograph of the epididymis from a ram showing the three main morphological regions(caput, corpus and cauda) and their subdivision into 10 different zones usually used for microcanulation. VD: vasdeferens. Real size: 12–15 cm.

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 323
face of this cell is organized in specific domains that are defined by distinct subsets ofnon-exchangeable lipids and proteins. Once this cell is liberated into the testicular fluid,it starts a long journey through the epididymis. Sperm quality, in terms of mobility andoocyte recognition, will be strongly dependent on the subtle post-testicular differentiationsthat occur in the epididymis. This accessory organ, anatomically connected to the testis,can be divided into three main regions: the caput, the corpus and the cauda (Fig. 1). Thisorgan shelters a several meters-long contorted duct (up to 60 m in large mammals), wherespermatozoa stay from several days to weeks depending upon the species (Yanagimachi,1994; Toshimori, 2003). In this tube, the media surrounding sperm will be completely re-newed. Its protein and other chemical compositions will be modified by the uptake andsecretion activities of the tubular epithelium (Yanagimachi, 1994). In the first region, theproteins from the rete testis fluid are replaced by newly secreted components. Thereafter,each region secretes specific components, the caput and the corpus being the most ef-fective. In parallel to the fluid changes, the sperm plasma membrane domains are pro-foundly remodeled by the degradation and/or liberation of testicular components, the ab-sorption and the integration of secreted proteins, and enzymatic surface-modifying eventslike glycosylation and deglycosylation of specific proteins (Dacheux et al., 2003; Tulsiani,2003).
In our laboratory, we have developed different methods from sperm movement analysisto two-dimensional poly-acrylamide gel electrophoresis (2D-PAGE) of the fluid and spermmembrane proteins, following micro-perfusion of the epididymal tubule. The 2D-PAGEmethod is a valuable tool which allows the identification and complete characterization ofproteins from complex biological samples when used in conjunction with mass spectrome-try. This paper will briefly review our recent findings and discuss the possibility they offerto describe the sperm maturation process.
2. Sperm maturation
Sperm maturation is evaluated by its final effect on the male gamete: acquisition ofmotility, the ability to recognize and bind to the zona pellucida, and to fuse with the oocyte.These different properties are acquired progressively and simultaneously as spermatozoatravel through the epididymis.
During the epididymal transit, the main morphological change that takes place in thespermatozoa is the migration of the cytoplasmic droplet, a remnant of the cytoplasm asso-ciated with testicular sperm. During transit from the caput to the corpus, this droplet slides,by an unknown mechanism, from the base of the head to the end of the intermediate pieceof the flagellum, and then shed from the spermatozoa either within the cauda fluid or dur-ing/after ejaculation depending upon the species. The necessity for the droplet migration isshown by the correlation between decreases in fertility and the proportion of spermatozoawith cytoplasmic droplets in the ejaculate. In different species, pathologies of spermatozoa(tail pin, decapitated spermatozoa) have been linked to a droplet migration problem (Bonetet al., 1992). It is worthy of note that the migration of the droplet occurs at the same timeas the motility of spermatozoa increases, but a direct relationship between these two eventshas not been demonstrated.

324 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
Spermatozoa obtained from the rete testis are almost immotile and are not able to showa straight-line displacement even after dilution in protective media. Progressive motilityacquisition is gradual as the spermatozoa moves along the epididymis. Thus, the number ofmotile cells seen after dilution increases in the corpus and reaches a maximum when theyenter the cauda, while their type of movement changes from wiggling to straight-line dis-placement (Dacheux and Paquignon, 1980; Yanagimachi, 1994). In the majority of species,spermatozoa motility in the epididymis is balanced by two important components: one con-cerning the maturation of the flagellar machinery (axoneme, dense fibers, etc.) and the othercontrolling this machinery to maintain the spermatozoa in an immotile state within the epi-didymis. The relationship between these two components may be complex since they mayuse similar cAMP-dependent signaling pathways. The flagellum of testicular spermatozoacan beat after removal of the plasma membrane in media that contain both ATP and cAMP,or when intact spermatozoa are mixed with phophodiesterase inhibitors, bicarbonate and/oregg yolk (Hoskins et al., 1975; Dacheux et al., 1979; Okamura et al., 1991; Patil et al.,2002). Moreover, the flagellar wave-form and the beat frequency observed under these con-ditions are close to those of mature spermatozoa (Ishijima and Witman, 1987). Thus, theimmotile state in the testis and the caput is due to an inhibition of the flagellar machineryby the low intracellular cAMP concentration. This inhibition is reversed when the sperma-tozoa cross the corpus epididymis, where cAMP increases in response to external signal(s)translated by the spermatozoa plasma membrane (Dacheux and Paquignon, 1980; Parisetet al., 1985). The nature of the signal and its exact pathway(s) are still unknown; how-ever, increased phosphorylation of proteins on the dense fibers and the axoneme complexhas been reported during the transit (Tash and Bracho, 1998; Eddy et al., 2003). In mostspecies, spermatozoa are immotile in the cauda epididymis; motility activation relies onan increase in intracellular cAMP and calcium, and changes in the phosphorylation statusof specific proteins (Huang and Vijayaraghavan, 2004; Inaba, 2003; Wade et al., 2003).The quiescent state of the spermatozoa in the cauda may be in part related to mechanicalconstraints induced by the presence of specific proteins produced by the epididymis, suchas immobilin in the rat that strongly increases the media viscosity (Usselman and Cone,1983). This protein appears to be species-specific and increased viscosity impairs only thepropagation of waves but not the ATP consumption or the oxidative metabolism in sperma-tozoa (Cardullo and Cone, 1986). Other factors of epididymal epithelial origin involved inthe control of spermatozoa motility include the ionic environment (calcium, pH, HCO3
−)and the presence or the absence of specific energy substrates to maintain a low spermatozoaATP concentration (Gatti et al., 1993; Wade et al., 2003; Wang et al., 2003).
Fertility of maturing spermatozoa can be measured by artificial insemination or in vitrofertilization. These studies also show an increasing fertility gradient between the caput andthe corpus with a maximum in the cauda, where fertility is equal or even better than thatobtained with ejaculated spermatozoa (Dacheux and Paquignon, 1980). The number ofspermatozoa able to recognize and bind to the oocyte investment increases in parallel to thenumber of motile spermatozoa, but the increase in binding is observed even at 4◦C when thespermatozoa motility is strongly reduced suggesting that these physiological properties areunrelated. Moreover, the number of spermatozoa able to bind to solubilized zona pellucida(indicating also indirectly the number of spermatozoa receptors for the zona) increasesstrongly in the corpus and cauda epididymis (Burkin and Miller, 2000). Recognition and

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 325
fusion involve specific components and domains on the spermatozoa surface. It is clear thatthe remodeling of the membrane in response to the epididymal secretions plays a major rolein controlling these properties.
3. Epididymal proteome and secretome
Proteomic analysis of the epididymal fluid obtained by one and two-dimensional gelelectrophoresis demonstrates that each of the proteins present in the fluid has a differ-ent distribution pattern along this organ (Fig. 2). The proteomic analysis reflects both
Fig. 2. Comparison between the proteome and secretome in ram and stallion. The fluid from the different epi-didymal zones of ram (A and B) and stallion (C and D) were obtained by microperfusion after incubation with35S-methionine and35S-cysteine. These fluids were separated by SDS–PAGE and silver-stained (B and D) anddried before fluorography to show the radio-labeled secreted proteins (A and C). The biosynthesis shows that theprotein secretion is highly regionalized, the caput and corpus being the most active regions, except for lactoferrinin the stallion. The silver stained gels show that the protein pattern changes in the fluid from different regionsof the tract. Some proteins are only transiently present and others accumulate in the caudal fluid. Comparisonbetween the proteome and secretome allows identification of proteins that are present in the fluid and not secretedsuggesting their spermatozoa origin (like (AB), gACE) (C0–C6: clusterin alpha and beta chains isoforms; Hex:hexosaminidase; Lf: lactoferrine; Gal: galactosidase; GPX: glutathione peroxidase; Alb: albumin).

326 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
the uptake/degradation of proteins and the secretion and accumulation of new compo-nents. It is a dynamic view of the proteins the epididymal tubule contains. The secre-tome, obtained by analyzing35S-methionine- or cysteine-labeled proteins secreted by eachzone of the epididymis gives a more static view, but allows to identify proteins that arerapidly degraded or reabsorbed and hence do not accumulate in the fluid (Syntin et al.,1996; Castella et al., 2003). Both methods are necessary to understand the complete ac-tivity of the epididymal epithelium, or to discover the presence of the proteins derivedfrom spermatozoa (Fig. 2). A comparison of the results obtained in different mammalshas been reviewed recently (Dacheux et al., 2003) and will be only briefly highlightedhere.
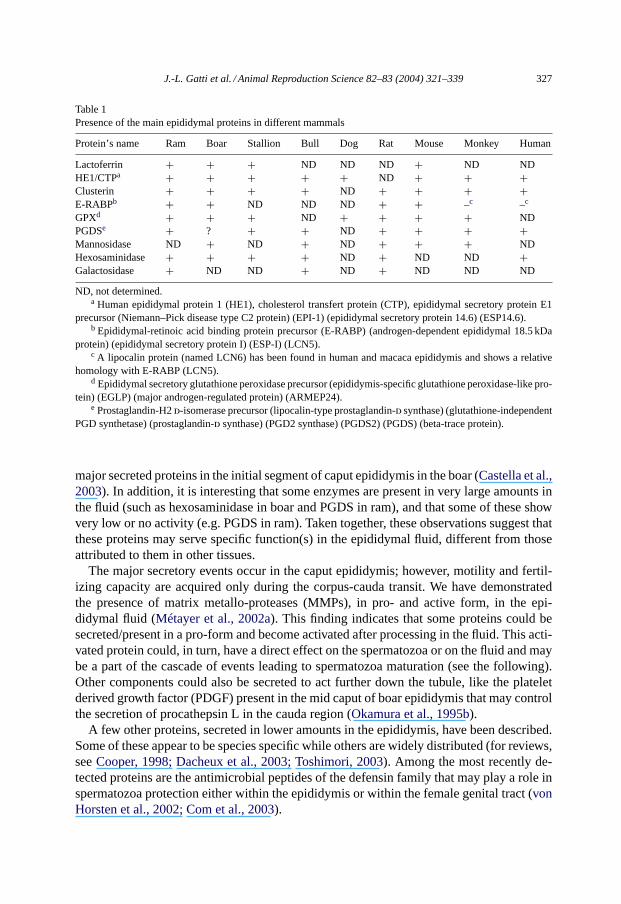
The majority of the proteins entering the epididymis from the rete testis disappears inthe proximal caput epididymis (Fig. 2). For example, clusterin and transferin, the mainproteins secreted by the Sertoli cells, are found in the rete testis fluid but not in the fluidfrom the proximal caput where these proteins are reabsorbed. The epithelial up-take ofspecific components is not restricted to the caput region. Almost all of the main componentsfound in epididymal fluid are the secretory products of the duct. This represents more thanone hundred different detectable proteins, with a high polymorphism (both in molecularweight and isoelectric point) due to glycosylations and other post-translational modifications(Hermo et al., 1991; Syntin et al., 1996; Fouchécourt et al., 2000). The most active regions arethe caput and corpus that synthesize about 80% of the compounds in the fluid, demonstratingthe high degree of regionalization of activities in this organ. In some cases, one protein canrepresent more than 50% of the secretion of one epididymal zone. In the horse and boar,newly synthesized clusterin is 60% of the secreted proteins of the distal caput (seeFig. 2D).Overall, in domestic mammals, about 10 different proteins account for 80–90% of thetotal epididymal secretion (Table 1). These proteins show restricted patterns of secretionand different and independent behavior, after secretion. These proteins are also found inthe epididymis of other mammalian species including human, dog and rodents, and havesimilar epididymal distributions (Table 1).
Almost no major secreted components seem specific for the epididymis and their role inepididymal spermatozoa maturation is still unknown. For example, clusterin is detected in alarge number of tissues and has different functions ranging from membrane lipid recyclingin apoptotic cell death, to a role as a chaperone to stress-induced secreted proteins (Jonesand Jomary, 2002). Knocking out this protein had no apparent effect on reproduction oron the normal physiology of the mouse (McLaughlin et al., 2000). Other proteins like theHE1/CTP/NP-C2, E-RABP, and PGDS (seeTable 1) are involved in the transport of hy-drophobic compounds such as cholesterol, retinoic acid and androgen, respectively (Rankinet al., 1992; Okamura et al., 1999; Fouchécourt et al., 2002). These proteins apparently donot bind to the spermatozoa; however, may act as signaling molecules for epididymal ac-tivity. Moreover, the major proteins from the rete testis fluid, clusterin and transferrin, arereplaced in the fluid by secreted clusterin and a member of the transferrin family, lactoferrin(Farnaud and Evans, 2003). It is hypothesized that these components may be involved inprotecting the spermatozoa cells against oxidation reaction and/or bacterial attacks, duringtheir transit and storage. Some of these components are secreted as specific isoforms in pre-cise regions of the epididymis and have only partial sequence homologies with other knownproteins. This is the case with the RNAse-like protein recently identified that is one of the
J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 327
Table 1Presence of the main epididymal proteins in different mammals
Protein’s name Ram Boar Stallion Bull Dog Rat Mouse Monkey Human
Lactoferrin + + + ND ND ND + ND NDHE1/CTPa + + + + + ND + + +Clusterin + + + + ND + + + +E-RABPb + + ND ND ND + + –c –c
GPXd + + + ND + + + + NDPGDSe + ? + + ND + + + +Mannosidase ND + ND + ND + + + NDHexosaminidase + + + + ND + ND ND +Galactosidase + ND ND + ND + ND ND ND
ND, not determined.a Human epididymal protein 1 (HE1), cholesterol transfert protein (CTP), epididymal secretory protein E1
precursor (Niemann–Pick disease type C2 protein) (EPI-1) (epididymal secretory protein 14.6) (ESP14.6).b Epididymal-retinoic acid binding protein precursor (E-RABP) (androgen-dependent epididymal 18.5 kDa
protein) (epididymal secretory protein I) (ESP-I) (LCN5).c A lipocalin protein (named LCN6) has been found in human and macaca epididymis and shows a relative
homology with E-RABP (LCN5).d Epididymal secretory glutathione peroxidase precursor (epididymis-specific glutathione peroxidase-like pro-
tein) (EGLP) (major androgen-regulated protein) (ARMEP24).e Prostaglandin-H2d-isomerase precursor (lipocalin-type prostaglandin-d synthase) (glutathione-independent
PGD synthetase) (prostaglandin-d synthase) (PGD2 synthase) (PGDS2) (PGDS) (beta-trace protein).
major secreted proteins in the initial segment of caput epididymis in the boar (Castella et al.,2003). In addition, it is interesting that some enzymes are present in very large amounts inthe fluid (such as hexosaminidase in boar and PGDS in ram), and that some of these showvery low or no activity (e.g. PGDS in ram). Taken together, these observations suggest thatthese proteins may serve specific function(s) in the epididymal fluid, different from thoseattributed to them in other tissues.
The major secretory events occur in the caput epididymis; however, motility and fertil-izing capacity are acquired only during the corpus-cauda transit. We have demonstratedthe presence of matrix metallo-proteases (MMPs), in pro- and active form, in the epi-didymal fluid (Métayer et al., 2002a). This finding indicates that some proteins could besecreted/present in a pro-form and become activated after processing in the fluid. This acti-vated protein could, in turn, have a direct effect on the spermatozoa or on the fluid and maybe a part of the cascade of events leading to spermatozoa maturation (see the following).Other components could also be secreted to act further down the tubule, like the plateletderived growth factor (PDGF) present in the mid caput of boar epididymis that may controlthe secretion of procathepsin L in the cauda region (Okamura et al., 1995b).
A few other proteins, secreted in lower amounts in the epididymis, have been described.Some of these appear to be species specific while others are widely distributed (for reviews,seeCooper, 1998; Dacheux et al., 2003; Toshimori, 2003). Among the most recently de-tected proteins are the antimicrobial peptides of the defensin family that may play a role inspermatozoa protection either within the epididymis or within the female genital tract (vonHorsten et al., 2002; Com et al., 2003).

328 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
4. Spermatozoa membrane modifications
Spermatozoa labeling with lectins, antibodies, radioactive precursors or other chemi-cal probes have shown that great changes occur in the lipids and protein composition ofthe spermatozoa membrane during transit (Dacheux et al., 1984; Voglmayr et al., 1985;Yanagimachi, 1994). These changes result from different mechanisms including redistri-bution or disappearance after proteolytic processing, action of glycolytic enzymes andintegration of newly synthesized components.
4.1. Redistribution or removal after proteolytic processing
This mechanism has been described mainly for proteins that are positioned in specificspermatozoa membrane domains during spermiogenesis. We have demonstrated that thegerminal form of the angiotensin-I converting enzyme (gACE) present in the epididy-mal fluid and the seminal plasma in mammals, is derived from spermatozoa (Gatti et al.,1999; Métayer et al., 2002b). Angiotensin-converting enzyme (ACE) plays a central rolein the renin–angiotensin system and thus in blood pressure control. ACE is responsiblefor the production of angiotensin II and is important in the elimination of bradykinin. Inthe male ACE is found in two different isoforms: the first one is somatic (sACE) and ispresent in a large number of tissues (brain, lung, kidney,. . . ), while the other, gACE, istestis-specific (Bernstein, 1998). The gACE is found exclusively in the germ cells; thisenzyme is derived from the same gene as sACE, but from an alternate promoter (Kumaret al., 1989; Langford et al., 1991). These two isoforms share only one of the two ac-tive domains of sACE: the C-terminal domain that anchors the protein in the cell mem-brane. We have demonstrated that in large mammals and rodents, the gACE present onthe surface of the mid-piece and the principal piece of the flagellum, is completely re-moved during spermatozoa transit through the caput (Fig. 3) (Gatti et al., 1999; Métayeret al., 2002b). The protein is then released into the fluid as an active form and can belater found in the seminal plasma (Métayer et al., 2001). We have been able to show thatthis release is the result of an active proteolytic process at the cell surface and that ittakes place in the epididymal zone where the cleavage occurs and short fragments con-taining the transmembrane and the intracellular sequences appear in the spermatozoa cell(Métayer, 2001). This is the only known example of a protein present on the spermato-zoa membrane, that is released into the fluid where it exerts an enzymatic function andmay directly have an influence on the activity of the epididymal epithelium (Leung et al.,1998).
Other testicular spermatozoa proteins are also proteolytically processed during transit.The best known examples are the proteins from the ADAM family including fertilin al-pha and beta (PH30), cyritestin, testase, etc. (for review, seeEvans, 1999). Fertilin beta,present on the entire testicular hamster spermatozoa head, is degraded by two successivecleavages during caput transit (Frayne et al., 1998). After processing, the distribution of thisprotein is restricted to the post-acrosomal domain of the spermatozoa. Other proteins trans-formed in the same way include hyaluronidase pH20 (2B1/Spam1), protein CE9 (EMM-PRIN/basigin/OX47/MC31/gp42/CD147) and alpha-d-mannosidase (Tulsiani et al., 1995;Hou et al., 1996; Deng et al., 1999; Seaton et al., 2000). None of the protease(s) responsible

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 329
Fig. 3. gACE is shed from spermatozoa during the epididymal transit. Fluids from the different epididymal zoneswere tested with an antibody against ACE. This shows that the protein appears in the caput fluid. Immunolocaliza-tion with the same antibody indicates that the protein is mainly on the intermediate piece on testicular spermatozoa,while after transit, only a faint signal was observed on the acrosome of the spermatozoa from the cauda and in theejaculate.
for these transformations have been identified, but, serine-type proteases are thought to beinvolved (Frayne et al., 1998; Blobel, 2000).
A number of these proteins, present on spermatozoa and modified during the transit, havebeen described as essential for fertility (i.e., fertilin, cyritestin, gACE, etc.). These asser-tions are based on experimental results showing that the sperm–egg binding property isinhibited with antibodies or recombinant proteins and peptides, and that infertility occurs inmales following gene knockout (Esther et al., 1996; Evans, 2001). Surprisingly, almost allthe knockout mice for these surface proteins present similar phenotypes: although the sper-matozoa show normal morphology and motility, they have problems in traversing throughthe female genital tract and show a decreased ability to binding to the zona pellucida eventhough the involvement of these proteins have been assumed to be specifically at the levelof egg plasma membrane binding. The spermatozoa from some of these mice proved to befertile in vitro when mixed with eggs without zona pellucida. Interestingly, it was noted thatin some of these knock-out mice, the removal of one surface protein results in the decreaseor the absence of other surface components (Evans, 2001; Nishimura et al., 2001) suggest-ing that the spermatozoa surface architecture depends on the correct positioning of all itscomponents. Thus, the loss of one of these components may compromise the formation orthe proper maturation of other membrane proteins during epididymal transit which, in turn,may be reflected as lowered fertility rate.
4.2. Insertion of newly synthesized proteins
A large number of epididymal proteins have been reported to be integrated into thespermatozoa membrane during transit, however, it is not exactly known whether theseproteins are simply adsorbed to the spermatozoa surface, establish ionic interactions, orare integrated into the membrane (Cooper, 1998). Moreover, only a few of these pro-

330 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
teins have been consistently detected and described at the molecular level, in mammalianspecies.
4.2.1. Low strength interactionsClusterin is one of the secreted proteins found to be associated with the spermatozoa
membrane (Howes et al., 1998). However, only certain isoforms of this protein bind tospermatozoa membrane and this association may be related to the integrity of the sperma-tozoa (O’Bryan et al., 1994). The specific type of association is not known but clusterincould interact with cell membrane due to its chaperoning activity, or simply by adsorptionto the cell surface because of its large extracellular concentration.
4.2.2. Insertion after proteolytic processingIn boar, the alpha-mannosidase is secreted as a 135 kDa form, in the caput-corpus regions,
and a 27 kDa fragment of this protein is found to be inserted into the equatorial segment ofspermatozoa head (Okamura et al., 1992, 1995a). In mature spermatozoa, this fragment isconcentrated at the apical cap of the head.
The protein DE/AEG (acidic epididymal protein) has been described in several species.This is part of a family of cysteine-rich secretory proteins (CRISP) characterized by 16conserved cysteine residues that form a characteristic pattern of disulfide bonds (Brooks,1982; Kratzschmar et al., 1996). The protein AEG1/CRISP1 and AEG2/CRISP3 are presentin the epididymis but only AEG1/CRISP1 is epididymis-specific. TheAEG1 gene generatesdifferent mRNA transcripts and apparently produces two proteins: D and E. These proteinsmay be inserted into the spermatozoa membrane after the proteolytic removal of theircarboxy-terminal region (Roberts et al., 2002). In human spermatozoa, no CRISP1 proteinis detected but a new form of CRISP, linked to a GPI anchor, has been recently reported(Kratzschmar et al., 1996; Yudin et al., 2002).
4.2.3. Proteins with hydrophobic properties
(a) GPI anchored proteinsSpermatozoa cells are known to gain GPI anchored proteins during the epididymal
transit. A well-described example is the human epidiymal protein 5 (HE5-CDw52antigen) which is absent in testicular spermatozoa but present on mature male gametes(Kirchhoff et al., 1993, 1998). This protein is composed of a short peptidic sequence,is heavily glycosylated and is detected on mature lymphocyte. Its role in spermatozoaphysiology is still unknown.
(b) Hydrophobic proteinsWe detected two compounds, of 17 and 23 kDa molecular weight range, in the cauda
epididymis of the ram. Of these the 17 kDa protein has been immunolocalised to thespermatozoa flagella (Gatti et al., 2000). These proteins are strongly inserted sincerepeated washing could not remove them. These proteins present on the spermatozoasurface were also not removed by the phospholipase C treatment suggesting that they arenot inserted via a GPI anchor. The proteins released into the fluid could be extracted by amethanol–chloroform treatment or a TX114 partition, suggesting that they are stronglyhydrophobic and/or associated with membraneous or lipidic vesicles (see the following).

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 331
5. Molecular mechanisms involved in the membrane modifications
We have shown that modifications of the spermatozoa surface involve specific mech-anisms: proteolysis of preexisting components, insertion of compounds after proteolysisor via their hydrophobic properties. Modifications of preexisting components suggest thatproteases are present either on the spermatozoa or secreted into the fluid and activated inspecific segments of the epididymis. This implies also that proteolysis must be limited andwell-controlled and that specific inhibitors of these proteases are present and/or secretedin specific areas. The mRNA for numerous proteases and protease inhibitors have been re-ported in the epididymis, but only a few of them have been detected in the epididymal fluid(Okamura et al., 1995b; Clauss et al., 2002; Jervis and Robaire, 2002; Métayer et al., 2002a).
The presence of several pro-enzymes and pro-proteins has been demonstrated in theepididymal fluids of different mammals. Recently, using the sensitive gel zymographymethod and immunochemistry, we have demonstrated the presence of MMPs in pro- andactive forms, in the epididymal fluids of different mammals. We have also detected Furin,a pro-hormone convertase 2 (PC-2), generally noted as an intracellular serine protease in-volved in the processing of numerous pro-proteins including the MMPs, in the fluid fromthe distal caput region of the ram. However, furin was not present in cauda fluid. Proteasescould be involved also in the modification of the fluid by cleaving secreted compoundswithout known activity, such as the prion protein (Gatti et al., 2002). Prion protein (Prp)is involved in the transmission of encephalopathies including the bovine spongiform en-cephalopathy, the Human Creutzfeld–Jackob disease and the sheep scrapie (Prusiner, 2001).Prp is a GPI membrane anchored glycoprotein normally present in numerous tissues. It isalso found in spermatozoa and genital tract fluids. We have observed that this protein issecreted in the epididymal fluid where it is present in different isoforms due to glycosylationand proteolytic processing. Our recent studies suggest also that these different isoforms arepartitioned into different compartments within the fluid (see the following).
Epididymal fluid is also a rich source of protease inhibitors; for example, a specifictissue inhibitor of MMPs, TIMP2, is found in the epididymal fluid of the boar, the ramand the horse (Métayer et al., 2002a). Other types of protease inhibitors detected includealpha-macroglobulin, cystatin-related epididymal protein (CRES) and a cysteine-rich pro-tein containing both Kunitz-type and WAP-type (four disulfide core) consensus sequencesnamed epididymal protease inhibitors (EPPIN) (Métayer et al., 2002a; Cornwall et al., 2003;Hsia and Cornwall, 2003; Sivashanmugam et al., 2003). Interestingly, it has been shownthat CRES is a potent inhibitor of furin convertase and therefore, may control the activityof this protein in the fluid.
In addition, proteins of the epididymal fluid and spermatozoa surface may also be targetsof different glycosydase enzymes that are secreted or activated in the fluid. These enzymeswould appear to be involved in the modification of the spermatozoa membrane properties,although, to date, only indirect proofs of their actions exist (Tulsiani, 2003).
5.1. How can secreted proteins be inserted in the spermatozoa plasma membrane?
This is an important point in elucidating the spermatozoa surface modifications occurringin the epididymis. As stated before, several components secreted in the epididymal fluid

332 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
need to undergo proteolysis before or during their insertion into spermatozoa membrane.This modification certainly increases their hydrophobic property or unmasks a specifichydrophobic sequence that allows this process to occur. Alternatively, this could allowrecognition by specific surface receptors.
More striking is the presence and the transfer of hydrophobic proteins with or withoutGPI anchor, in the epididymal fluid. The hydrophobic proteins in the fluid were thoughtto be linked to membrane vesicles, or included in lipid micelles (Kirchhoff et al., 1998).It was long known that small vesicles are present in the seminal plasma and that theyare derived from the prostate and seminal vesicular epithelium (Saez et al., 2003). Thesevesicles, named prostasomes or seminosomes, contain a specific subset of proteins and areable to transfer some of them to spermatozoa under certain conditions (Saez et al., 2003;Utleg et al., 2003). It has been suggested that such vesicles (named epididymosomes) maybe present in the epididymal fluid and that they may enable the transfer of specific proteinsto the spermatozoa, under specific conditions (Frenette et al., 2002; Saez et al., 2003). Wehave demonstrated (Gatti et al., 2003) that these vesicles indeed exist and they are foundonly in the cauda epididymal fluid where they represent a small fraction of the fluid proteins(Fig. 4). These vesicles contain specific proteins found also in prostasomes, but the majorcompounds present on these vesicles were not found on cauda or ejaculated spermatozoa(Gatti et al., in preparation). Thus, if these vesicles transfer proteins from the epithelium tothe spermatozoa in vivo, it must be only a subset of proteins with a very specific mechanismand not simply due to membrane fusion.
While a part of the hydrophobic fluid proteins are linked to epididymosomes, a largequantity remain in the bulk phase after the complete removal of these vesicles by high speed
Fig. 4. Epididymal fluid contains exosome-like vesicles with specific proteins. The cauda fluid (CEP) from theram (zone 8/9) was centrifuged twice at 15,000× g and the supernatant was centrifuged at high speed (50,000× g) for 2 h. The pellet obtained after this centrifugation was processed for electron microscopy (left). Thispellet was composed of small membrane-bound vesicles of 25–50 nm diameter. These vesicles, referred to asepididymosomes (Eps), show a different protein pattern from the CEP when separated by SDS–PAGE (right).

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 333
centrifugation (Gatti et al., 2003). This suggests that other types of transport exist, eitherin the form of very small vesicles or micelles. We recently observed by sucrose gradientseparation or gel filtration, that high molecular weight complexes (more than 1 MDa) existin the epididymal fluid. Some of the hydrophobic proteins are linked to these complexes,which also contain associated proteins that are currently under study in our laboratory.
6. Epididymal proteins and fertility
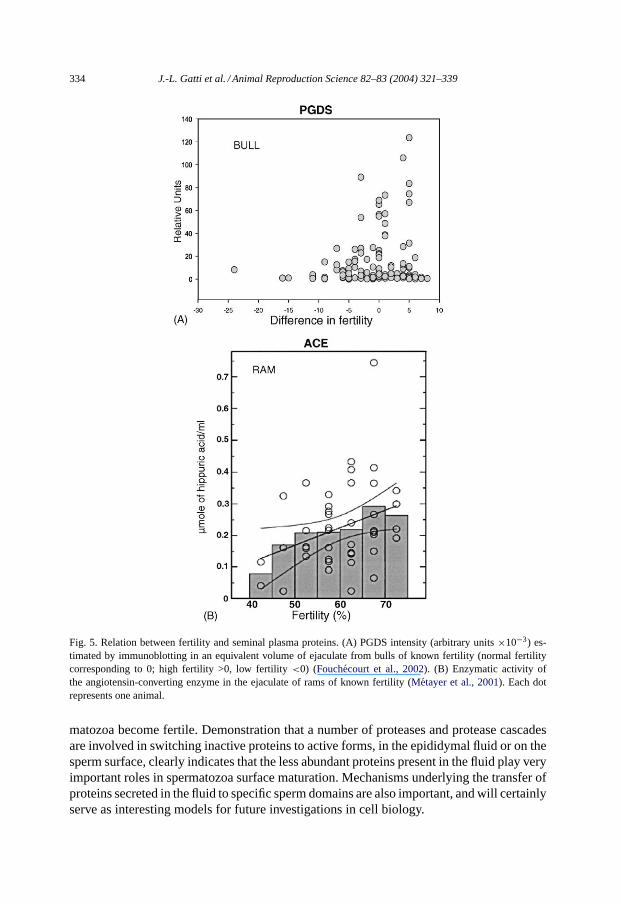
Numerous attempts have been made to correlate the fertility potential of the spermatozoawith the proteins present in the seminal plasma, including those originating from the epi-didymis. For example, it has been suggested that the quantity of prostaglandin D2 synthase(PGDS) in seminal plasma is correlated with the fertility of the bull (Gerena et al., 1998).While we found that a low PGDS content in bull seminal plasma is associated with lowerfertility, we also noted that bulls with high fertility displayed high or low concentrations ofthis protein (Fig. 5A), indicating that PGDS was not a good fertility marker and that it maynot even be a necessary factor for fertility (Fouchécourt et al., 2002).
Another approach is to use a spermatozoa protein added or modified during transit.Epididymis-secreted clusterin associated with spermatozoa has been used as a marker forfertility since it could be correlated to the number of abnormal spermatozoa in the ejaculate(Howes et al., 1998; Ibrahim et al., 2000). However, the correlation between clusterinpositive spermatozoa and fertility remains low. Several other proteins have been used toassess fertility in the bull, including one of the p26h-related proteins, p25b (a carbonylreductase-like protein), which is a glycosylphosphatidyl inositol-anchored protein foundon the spermatozoa and in epididymosomes (Frenette and Sullivan, 2001). Although levelsof this protein on the spermatozoa were found to be highly variable from male to male, ithas been suggested that lower levels could be associated with lower fertility.
Because we have demonstrated that all the ACE activity of the ram seminal plasma is dueto the gACE released from spermatozoa during transit, we made the assumption that thelevel of activity in this fluid could be a marker of maturation. We hypothesized that since thequantity of gACE in seminal plasma (released during epididymal spermatozoa transit) iscorrelated with the number of ejaculated spermatozoa, a deviation from the expected gACEcontent could be an indicator of interrupted spermatozoa maturation. We have measuredthe activity of gACE in the seminal plasma of young rams (Métayer et al., 2001) and havefound that the seminal plasma gACE activity was correlated to the number of spermatozoa(spermatozoa count) in the ejaculate. We noted that lower activity in seminal fluid wasassociated with lower fertility levels in animals (Fig. 5B); however, these animals did notdisplay the lowest spermatozoa count. This approach needs to be repeated with a largersample of animals and in different species in order to be validated.
7. Perspectives
Focusing on major epididymal proteins is the first step in the comprehension of thespermatozoa maturation environment; but, it may not be enough to understand how sper-
334 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
Fig. 5. Relation between fertility and seminal plasma proteins. (A) PGDS intensity (arbitrary units×10−3) es-timated by immunoblotting in an equivalent volume of ejaculate from bulls of known fertility (normal fertilitycorresponding to 0; high fertility >0, low fertility<0) (Fouchecourt et al., 2002). (B) Enzymatic activity ofthe angiotensin-converting enzyme in the ejaculate of rams of known fertility (Metayer et al., 2001). Each dotrepresents one animal.
matozoa become fertile. Demonstration that a number of proteases and protease cascadesare involved in switching inactive proteins to active forms, in the epididymal fluid or on thesperm surface, clearly indicates that the less abundant proteins present in the fluid play veryimportant roles in spermatozoa surface maturation. Mechanisms underlying the transfer ofproteins secreted in the fluid to specific sperm domains are also important, and will certainlyserve as interesting models for future investigations in cell biology.

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 335
Studies on the transcriptome and the proteome of the testis and epididymis are underway.Transcriptome analysis will provide a basis to understand how the activity and regionaliza-tion of the epididymis is controlled while the availability of new techniques, such as massspectrometry, will enable the analysis of the less abundant proteins present in the epididy-mal fluid. In parallel, efforts should be focused on the identification and characterizationof spermatozoa surface proteins that are modified, in different mammalian species. Thisshould lead to a better understanding of the complex and interconnected mechanisms bywhich spermatozoa cell functions are developed during spermatogenesis and maturation.
Semen quality is currently judged by simple criteria (spermatozoa concentration, mobil-ity, abnormal cells) that may not always accurately predict fertility of a sample or explainthe differences between males. Even after these tests, important variations in fertility fol-lowing insemination persists. Several approaches have been suggested to ameliorate theerrors of screening, such as in vitro fertilization and hemi-zona binding assays; but they areexpensive and time-consuming and do not always serve as good predictors (Braundmeierand Miller, 2001; Rodriguez-Martinez, 2003). Increasing our knowledge on spermatozoaepididymal maturation should provide specific molecular markers in order to develop newcriteria that are more accurate and objective in predicting and improving male fertility.
Acknowledgements
This work is supported by grants from the “Région Centre”, GIS-prion and GENANIMAL-2003.
References
Bernstein, K.E., 1998. Examining the renin–angiotensin system one hundred years after its discovery. Nephrologie19, 391–395.
Blobel, C.P., 2000. Functional processing of fertilin: evidence for a critical role of proteolysis in sperm maturationand activation. Rev. Reprod. 5, 75–83.
Bonet, S., Briz, M., Fradera, A., Egozcue, J., 1992. Origin, development and ultrastructure of boar spermatozoawith folded tails and with two tails. Hum. Reprod. 7, 523–528.
Braundmeier, A.G., Miller, D.J., 2001. The search is on: finding accurate molecular markers of male fertility. J.Dairy Sci. 84, 1915–1925.
Brooks, D.E., 1982. Purification of rat epididymal proteins “D” and “E”, demonstration of shared immunologicaldeterminants, and identification of regional synthesis and secretion. Int. J. Androl. 5, 513–524.
Burkin, H., Miller, D.J., 2000. Zona pellucida protein binding ability of porcine sperm during epididymalmaturation and the acrosome reaction. Dev. Biol. 222, 99–109.
Cardullo, R.A., Cone, R.A., 1986. Mechanical immobilization of rat sperm does not change their oxygenconsumption rate. Biol. Reprod. 34, 820–830.
Castella, S., Fouchecourt, S., Teixeira Gomes, A.P., Vinh, J., Belghari, M., Dacheux, F., Dacheux, J.L., 2003.Identification of a member of a new RNase a family specifically secreted by epididymal caput epithelium. Biol.Reprod. 70, 319–328.
Clauss, A., Lilja, H., Lundwall, A., 2002. A locus on human chromosome 20 contains several genes expressingprotease inhibitor domains with homology to whey acidic protein. Biochem. J. 368, 233–242.
Com, E., Bourgeon, F., Evrard, B., Ganz, T., Colleu, D., Jegou, B., Pineau, C., 2003. Expression of antimicrobialdefensins in the male reproductive tract of rats, mice, and humans. Biol. Reprod. 68, 95–104.

336 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
Cooper, T.G., 1998. Interactions between epididymal secretions and spermatozoa. J. Reprod. Fertil. Suppl. 53,119–136.
Cornwall, G.A., Cameron, A., Lindberg, I., Hardy, D.M., Cormier, N., Hsia, N., 2003. The cystatin-relatedepididymal spermatogenic protein inhibits the serine protease prohormone convertase 2. Endocrinology 144,901–908.
Dacheux, J.L., O’Shea, T., Paquignon, M., 1979. Effects of osmolality, bicarbonate and buffer on the metabolismand motility of testicular, epididymal and ejaculated spermatozoa of boars. J. Reprod. Fertil. 55, 287–296.
Dacheux, J.L., Paquignon, M., 1980. Relations between the fertilizing ability, motility and metabolism ofepididymal spermatozoa. Reprod. Nutr. Dev. 20, 1085–1099.
Dacheux, J.L., Paquignon, M., Lanneau, M., 1984. Sequential analysis of the epididymal sperm maturation processin the boar. Ann. N.Y. Acad. Sci. 438, 526–529.
Dacheux, J.L., Gatti, J.L., Dacheux, F., 2003. Contribution of epididymal secretory proteins for spermatozoamaturation. Microsc. Res. Tech. 61, 7–17.
Deng, X., Czymmek, K., Martin-DeLeon, P.A., 1999. Biochemical maturation of spam1 (ph-20) during epididymaltransit of mouse sperm involves modifications of N-linked oligosaccharides. Mol. Reprod. Dev. 52, 196–206.
Eddy, E.M., Toshimori, K., O’Brien, D.A., 2003. Fibrous sheath of mammalian spermatozoa. Microsc. Res. Tech.61, 103–115.
Esther Jr., C.R., Howard, T.E., Marino, E.M., Goddard, J.M., Capecchi, M.R., Bernstein, K.E., 1996. Micelacking angiotensin-converting enzyme have low blood pressure, renal pathology, and reduced male fertility.Lab. Invest. 74, 953–965.
Evans, J.P., 1999. Sperm disintegrins, egg integrins, and other cell adhesion molecules of mammalian gameteplasma membrane interactions. Front. Biosci. 4, D114–131.
Evans, J.P., 2001. Fertilin beta and other adams as integrin ligands: insights into cell adhesion and fertilization.Bioessays 23, 628–639.
Farnaud, S., Evans, R.W., 2003. Lactoferrin—a multifunctional protein with antimicrobial properties. Mol.Immunol. 40, 395–405.
Fouchécourt, S., Métayer, S., Locatelli, A., Dacheux, F., Dacheux, J.L., 2000. Stallion epididymal fluid proteome:qualitative and quantitative characterization secretion and dynamic changes of major proteins. Biol. Reprod.62, 1790–1803.
Fouchécourt, S., Charpigny, G., Reinaud, P., Dumont, P., Dacheux, J.L., 2002. Mammalian lipocalin-typeprostaglandin D2 synthase in the fluids of the male genital tract: putative biochemical and physiologicalfunctions. Biol. Reprod. 66, 458–467.
Frayne, J., Jury, J.A., Barker, J.A., Hall, L., 1998. The MDC family of proteins and their processing duringepididymal transit. J. Reprod. Fertil. Suppl. 53, 149–155.
Frenette, G., Sullivan, R., 2001. Prostasome-like particles are involved in the transfer of p25b from the bovineepididymal fluid to the sperm surface. Mol. Reprod. Dev. 59, 115–121.
Frenette, G., Lessard, C., Sullivan, R., 2002. Selected proteins of “prostasome-like particles” from epididymalcauda fluid are transferred to epididymal caput spermatozoa in bull. Biol. Reprod. 67, 308–313.
Gatti, J.L., Chevrier, C., Paquignon, M., Dacheux, J.L., 1993. External ionic conditions, internal ph and motilityof ram and boar spermatozoa. J. Reprod. Fertil. 98, 439–449.
Gatti, J.L., Druart, X., Guerin, Y., Dacheux, F., Dacheux, J.L., 1999. A 105- to 94-kilodalton protein in theepididymal fluids of domestic mammals is angiotensin I-converting enzyme (ACE) evidence that sperm are thesource of this ACE. Biol. Reprod. 60, 937–945.
Gatti, J.L., Druart, X., Syntin, P., Guerin, Y., Dacheux, J.L., Dacheux, F., 2000. Biochemical characterization oftwo ram cauda epididymal maturation-dependent sperm glycoproteins. Biol. Reprod. 62, 950–958.
Gatti, J.L., Métayer, S., Moudjou, M., Andréoletti, O., Lantier, F., Dacheux, J.L., Sarradin, P., 2002. Prion proteinis secreted in soluble forms in the epididymal fluid and proteolytically processed and transported in seminalplasma. Biol. Reprod. 67, 393–400.
Gatti, J.L., Belghazi, M., Métayer, S., Dacheux, F., Dacheux, J.L., 2003. Proteomic analysis of epididymal fluidexosome-like vesicles (epididymosomes). In: 28th Annual Meeting of the American Society of Andrology,Phoenix, Arizona, p. 36.
Gerena, R.L., Irikura, D., Urade, Y., Eguchi, N., Chapman, D.A., Killian, G.J., 1998. Identification of a fertility-associated protein in bull seminal plasma as lipocalin-type prostaglandind synthase. Biol. Reprod. 58, 826–833.

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 337
Hermo, L., Wright, J., Oko, R., Morales, C.R., 1991. Role of epithelial cells of the male excurrent duct system ofthe rat in the endocytosis or secretion of sulfated glycoprotein-2 (clusterin). Biol. Reprod. 44, 1113–1131.
Hoskins, D.D., Hall, M.L., Munsterman, D., 1975. Induction of motility in immature bovine spermatozoa by cyclicamp phosphodiesterase inhibitors and seminal plasma. Biol. Reprod. 13, 168–176.
Hou, S.T., Ma, A., Jones, R., Hall, L., 1996. Molecular cloning and characterization of rat sperm surface antigen2B1. a glycoprotein implicated in sperm-zona binding. Mol. Reprod. Dev. 45, 193–203.
Howes, E.A., Hurst, S., Laslop, A., Jones, R., 1998. Cellular distribution and molecular heterogeneity of mac393antigen (clusterin, beta-chain) on the surface membrane of bull spermatozoa. Mol. Hum. Reprod. 4, 673–681.
Hsia, N., Cornwall, G.A., 2003. Cres2 and cres3: new members of the cystatin-related epididymal spermatogenicsubgroup of family 2 cystatins. Endocrinology 144, 909–915.
Huang, Z., Vijayaraghavan, S., 2004. Increased phosphorylation of a distinct subcellular pool of proteinphosphatase, pp1{gamma}2, during epididymal sperm maturation. Biol. Reprod. 70, 439–447.
Humblot, P., Ducrocq, V., 1996. Genetic determinism of sperm production in bulls. Contracept. Fertil. Sex. 24,617–623.
Ibrahim, N.M., Gilbert, G.R., Loseth, K.J., Crabo, B.G., 2000. Correlation between clusterin-positive spermatozoadetermined by flow cytometry in bull semen and fertility. J. Androl. 21, 887–894.
Inaba, K., 2003. Molecular architecture of the sperm flagella: molecules for motility and signaling. Zool. Sci.Tokyo 20, 1043–1056.
Ishijima, S., Witman, G.B., 1987. Flagellar movement of intact and demembranated, reactivated ram spermatozoa.Cell Motil. Cytoskeleton 8, 375–391.
Jervis, K.M., Robaire, B., 2002. Changes in gene expression during aging in the brown norway rat epididymis.Exp. Gerontol. 37, 897–906.
Jones, S.E., Jomary, C., 2002. Clusterin. Int. J. Biochem. Cell. Biol. 34, 427–431.Kirchhoff, C., Krull, N., Pera, I., Ivell, R., 1993. A major mRNA of the human epididymal principal cells, HE5,
encodes the leucocyte differentiation cdw52 antigen peptide backbone. Mol. Reprod. Dev. 34„ 8–15.Kirchhoff, C., Osterhoff, C., Pera, I., Schroter, S., 1998. Function of human epididymal proteins in sperm
maturation. Andrologia 30, 225–232.Kratzschmar, J., Haendler, B., Eberspaecher, U., Roosterman, D., Donner, P., Schleuning, W.D., 1996. The human
cysteine-rich secretory protein (crisp) family. Primary structure and tissue distribution of crisp-1, crisp-2 andcrisp-3. Eur. J. Biochem. 236, 827–836.
Kumar, R.S., Kusari, J., Roy, S.N., Soffer, R.L., Sem, G.C., 1989. Structure of testicular angiotensin-convertingenzyme. A segmental mosaic isozyme. J. Biol. Chem. 264, 16754–16758.
Langford, K.G., Shai, S.Y., Howard, T.E., Kovac, M.J., Overbeek, P.A., Bernstein, K.E., 1991. Transgenic micedemonstrate a testis-specific promoter for angiotensin-converting enzyme. J. Biol. Chem. 266, 15559–15562.
Leung, P.S., Chan, H.C., Chung, Y.W., Wong, T.P., Wong, P.Y., 1998. The role of local angiotensins andprostaglandins in the control of anion secretion by the rat epididymis. J. Reprod. Fertil. Suppl. 53, 15–22.
McLaughlin, L., Zhu, G., Mistry, M., Ley-Ebert, C., Stuart, W.D., Florio, C.J., Groen, P.A., Witt, S.A., Kimball,T.R., Witte, D.P., Harmony, J.A., Aronow, B.J., 2000. Apolipoprotein j/clusterin limits the severity of murineautoimmune myocarditis. J. Clin. Invest. 106, 1105–1113.
Métayer, S., 2001. Protéome épididymaire, mise en évidence de protéases et role dans la maturation post-gonadiquedes spermatozoıdes. Doctorat de l’Université de Tours. Université de Tours, Tours, France.
Métayer, S., Dacheux, F., Guerin, Y., Dacheux, J.L., Gatti, J.L., 2001. Physiological and enzymatic properties ofthe ram epididymal soluble form of germinal angiotensin I-converting enzyme. Biol. Reprod. 65, 1332–1339.
Métayer, S., Dacheux, F., Dacheux, J.L., Gatti, J.L., 2002a. Comparison, characterization, and identification ofproteases and protease inhibitors in epididymal fluids of domestic mammals. Matrix metalloproteinases aremajor fluid gelatinases. Biol. Reprod. 66, 1219–1229.
Métayer, S., Dacheux, F., Dacheux, J.L., Gatti, J.L., 2002b. Germinal angiotensin I-converting enzyme is totallyshed from the rodent sperm membrane during epididymal maturation. Biol. Reprod. 67, 1763–1767.
Nadarajah, K., Burnside, E.B., Schaeffer, L.R., 1988. Genetic parameters for fertility of dairy bulls. J. Dairy Sci.71, 2730–2734.
Nishimura, H., Cho, C., Branciforte, D.R., Myles, D.G., Primakoff, P., 2001. Analysis of loss of adhesive functionin sperm lacking cyritestin or fertilin beta. Dev. Biol. 233, 204–213.
O’Bryan, M.K., Murphy, B.F., Liu, D.Y., Clarke, G.N., Baker, H.W., 1994. The use of anticlusterin monoclonalantibodies for the combined assessment of human sperm morphology and acrosome integrity. Hum. Reprod.9, 1490–1496.

338 J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339
Okamura, N., Onoe, S., Sugita, Y., Paquignon, M., Dacheux, F., Dacheux, J.L., 1991. Water insoluble fraction ofegg yolk maintains porcine sperm motility by activating adenylate cyclase. Mol. Reprod. Dev. 28, 136–142.
Okamura, N., Dacheux, F., Venien, A., Onoe, S., Huet, J.C., Dacheaux, J.L., 1992. Localization of a maturation-dependent epididymal sperm surface antigen recognized by a monoclonal antibody raised against a 135-kilodalton protein in porcine epididymal fluid. Biol. Reprod. 47, 1040–1052.
Okamura, N., Tamba, M., Liao, H.J., Onoe, S., Sugita, Y., Dacheux, F., Dacheux, J.L., 1995a. Cloning ofcomplementary DNA encoding a 135-kilodalton protein secreted from porcine corpus epididymis and itsidentification as an epididymis-specific alpha-mannosidase. Mol. Reprod. Dev. 42, 141–148.
Okamura, N., Tamba, M., Uchiyama, Y., Sugita, Y., Dacheux, F., Syntin, P., Dacheux, J.L., 1995b. Direct evidencefor the elevated synthesis and secretion of procathepsin L in the distal caput epididymis of boar. Biochim.Biophys. Acta 1245, 221–226.
Okamura, N., Kiuchi, S., Tamba, M., Kashima, T., Hiramoto, S., Baba, T., Dacheux, F., Dacheux, J.L., Sugita,Y., Jin, Y.Z., 1999. A porcine homolog of the major secretory protein of human epididymis, HE1, specificallybinds cholesterol. Biochim. Biophys. Acta 1438, 377–387.
Pariset, C.C., Feinberg, J.M., Dacheux, J.L., Weinman, S.J., 1985. Changes in calmodulin level and cAMP-dependent protein kinase activity during epididymal maturation of ram spermatozoa. J. Reprod. Fertil. 74,105–112.
Patil, S.B., Kulanand, J., Padma, P., Shivaji, S., 2002. Reactivation of motility of demembranated hamsterspermatozoa: role of protein tyrosine kinase and protein phosphatases. Andrologia 34, 74–86.
Prusiner, S.B., 2001. Shattuck lecture—neurodegenerative diseases and prions. N. Engl. J. Med. 344, 1516–1526.Rankin, T.L., Ong, D.E., Orgebin-Crist, M.C., 1992. The 18-kda mouse epididymal protein (mep 10) binds retinoic
acid. Biol. Reprod. 46, 767–771.Roberts, K.P., Ensrud, K.M., Hamilton, D.W., 2002. A comparative analysis of expression and processing of the
rat epididymal fluid and sperm-bound forms of proteins D and E. Biol. Reprod. 67, 525–533.Rodriguez-Martinez, H., 2003. Laboratory semen assessment and prediction of fertility: still utopia? Reprod.
Domest. Anim. 38, 312–318.Saez, F., Frenette, G., Sullivan, R., 2003. Epididymosomes and prostasomes: their roles in post-testicular maturation
of the sperm cells. J. Androl. 24, 149–154.Seaton, G.J., Hall, L., Jones, R., 2000. Rat sperm 2b1 glycoprotein (pH20) contains a c-terminal sequence motif
for attachment of a glycosyl phosphatidylinositol anchor. Effects of endoproteolytic cleavage on hyaluronidaseactivity. Biol. Reprod. 62, 1667–1676.
Sivashanmugam, P., Hall, S.H., Hamil, K.G., French, F.S., O’Rand, M.G., Richardson, R.T., 2003. Characterizationof mouse Eppin and a gene cluster of similar protease inhibitors on mouse chromosome 2. Gene 312, 125–134.
Syntin, P., Dacheux, F., Druart, X., Gatti, J.L., Okamura, N., Dacheaux, J.L., 1996. Characterization andidentification of proteins secreted in the various regions of the adult boar epididymis. Biol. Reprod. 55, 956–974.
Tash, J.S., Bracho, G.E., 1998. Identification of phosphoproteins coupled to initiation of motility in live epididymalmouse sperm. Biochem. Biophys. Res. Commun. 251, 557–563.
Toshimori, K., 2003. Biology of spermatozoa maturation: an overview with an introduction to this issue. Microsc.Res. Tech. 61, 1–6.
Tulsiani, D.R., NagDas, S.K., Skudlarek, M.D., Orgebin-Crist, M.C., 1995. Rat sperm plasma membranemannosidase: localization and evidence for proteolytic processing during epididymal maturation. Dev. Biol.167, 584–595.
Tulsiani, D.R., 2003. Glycan modifying enzymes in luminal fluid of rat epididymis: are they involved in alteringsperm surface glycoproteins during maturation? Microsc. Res. Tech. 61, 18–27.
Usselman, M.C., Cone, R.A., 1983. Rat sperm are mechanically immobilized in the caudal epididymis by“immobilin,” a high molecular weight glycoprotein. Biol. Reprod. 29, 1241–1253.
Utleg, A.G., Yi, E.C., Xie, T., Shannon, P., White, J.T., Goodlett, D.R., Hood, L., Lin, B., 2003. Proteomic analysisof human prostasomes. Prostate 56, 150–161.
Voglmayr, J.K., Sawyer Jr., R.F., Dacheux, J.L., 1985. Glycoproteins: a variable factor in surface transformationof ram spermatozoa during epididymal transit. Biol. Reprod. 33, 165–176.
von Horsten, H.H., Derr, P., Kirchhoff, C., 2002. Novel antimicrobial peptide of human epididymal duct origin.Biol. Reprod. 67, 804–813.
Wade, M.A., Jones, R.C., Murdoch, R.N., Aitken, R.J., 2003. Motility activation and second messenger signallingin spermatozoa from rat cauda epididymidis. Reproduction 125, 175–183.

J.-L. Gatti et al. / Animal Reproduction Science 82–83 (2004) 321–339 339
Wang, D., King, S.M., Quill, T.A., Doolittle, L.K., Garbers, D.L., 2003. A new sperm-specific Na+/H+ exchangerrequired for sperm motility and fertility. Nat. Cell Biol. 5, 1117–1122.
Yanagimachi, R., 1994. Mammalians Fertilization, 2nd ed. Raven Press, New York, NY, USA.Yudin, A.I., Li, M.W., Robertson, K.R., Tollner, T., Cherr, G.N., Overstreet, J.W., 2002. Identification of a novel
GPI-anchored crisp glycoprotein, mak248, located on the posterior head and equatorial segment of cynomolgusmacaque sperm. Mol. Reprod. Dev. 63, 488–499.
Copyright © 2022 FDOKUMEN




















