
PHYSIOLOGICAL AND MOLECULAR CHARACTERIZATION OF SOME EGYPTIAN SHEEP BREEDS
Upload
independentCategory
view
9download
0
Genetic diversity of wild boar populations and domesticpig breeds (Sus scrofa) in South-western Europe
PAULO CÉLIO ALVES1,2*†, IRIS PINHEIRO1,2†, RAQUEL GODINHO1,JOAQUIN VICENTE3, CHRISTIAN GORTÁZAR3 and MASSIMO SCANDURA4
1CIBIO, Centro de Investigação em Biodiversidade e Recursos Genéticos, Universidade do Porto,Campus Agrário de Vairão, 4485-661 Vairão, Portugal2Departamento de Biologia, Faculdade de Ciências da Universidade do Porto 4099-002 Porto,Portugal3IREC, Instituto de Investigación en Recursos Cinegéticos (CSIC/UCLM/JCCLM), 13005 CiudadReal, Spain4Dipartimento di Zoologia e Genetica Evoluzionistica, Università di Sassari, via Muroni 25 07100Sassari, Italy
Received 5 January 2010; accepted for publication 16 June 2010bij_1530 797..822
The Eurasian wild boar, Sus scrofa, is an important game species and is widely distributed in Eurasia. The recentdemographic expansions allied to man mediated translocations and reintroductions, reshaped the genetic vari-ability of wild boar populations, and gave rise to management problems. Genetic variability and the populationstructure of the European populations are not well-known, in particular in the Iberian Peninsula. In the presentstudy, we assessed the phylogenetic relationship among 17 Iberian wild boar populations and several Eurasian wildand domestic pigs by sequencing 660 bp of the mitochondrial (mt)DNA control region. This analysis was extendedto the two autochthonous Portuguese pig breeds, Bísaro and Alentejano, and their relationship with Eurasian pigswas also established. High levels of gene diversity were found in Iberian wild boar and Portuguese domestic pigs,as well as a significant genetic difference among and within populations. Iberian wild boar and Portuguese pigbreeds belong to the main European clade but diverge with animals from Central Europe, showing a highproportion of private (Iberian) haplotypes. These results suggest the existence of glacial refugia for Sus scrofa inthe Iberian Peninsula during the Last Glacial Maximum. No signs of Asian mtDNA introgression from Asianpopulations were found in the present survey. © 2010 The Linnean Society of London, Biological Journal of theLinnean Society, 2010, 101, 797–822.
ADDITIONAL KEYWORDS: ancestry – domestication – Iberian Peninsula – mitochondrial DNA.
INTRODUCTION
The wild boar, Sus scrofa, is common in Eurasia andcan also be found in the Northwestern region ofAfrica. Studies based on morphological data suggestthe existence of four geographically distinct subspe-cies, with Sus scrofa scrofa occurring in Europe, WestAsia and Northwest Africa (Genov, 1999). However,the taxonomy of this species is far from being resolved
and several studies based on genetic markers areconsistent with the existence of more subspecies inboth Europe and Asia (Randi, 1995; Watanobe et al.,1999).
During the 18th and beginning of the 19th Centu-ries, mainly as a result of hunting pressure, wild boarunderwent an extreme population decline in severalEuropean countries, and even became extinct in someregions such as Britain (Apollonio, Randi & Toso,1988; Yalden, 1999). However, after World War II, adecrease in the number of predators and changes inagricultural practices lead to a substantial range
*E-mail: [email protected]†These authors contributed equally to this work.
Biological Journal of the Linnean Society, 2010, 101, 797–822. With 5 figures
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822 797

expansion of this species in some European countries(Sáez-Royuela & Tellería, 1986).
In Portugal, during the 19th Century, populationsof wild boar also declined, and in the beginning of the20th Century, this species was restricted to a fewNorthern locations and to small nuclei in the Centreand South of the country, occupying no more than4.75% of the continental territory (Bugalho, Carvalho& Borges, 1984). Nevertheless, in the last 30 years,large fields once exploited for agriculture becamegradually abandoned, and wild boar hunting wasforbidden, thereby allowing the Portuguese wild boarpopulations to recover. Currently, this species occu-pies all the continental territory, except for the largeurban centres (Fonseca & Correia, 2008; Ferreiraet al., 2009).
Similarly, in Spain, the fragmented population sur-viving after World War II increased in size and thespecies spread over most of the country (Tellería &Saez-Royuela, 1985). The species’ expansion mighthave been also shaped by translocations and reintro-ductions of animals, often crossbred in captivity withdomestic pigs. Thus, hybridization between animalswith distinct geographical origins and between wildboar and domestic pigs might be common and there-fore represent a threat to the genetic integrity ofnative wild populations (Apollonio et al., 1988;Vernesi et al., 2003). Although the consequences ofsuch practices can strongly affect the wild boargenetic diversity locally, Scandura et al. (2008)showed that the pattern of variation over a widegeographic scale appears to depend mostly on ancientdemographic processes (e.g. glacial retreats and post-glacial expansions).
Southern regions of Europe, especially the Mediter-ranean, were very important refugial areas forseveral animal and plant species during the Quater-nary glaciations (Hewitt, 1999; Sommer & Nada-chowski, 2006). The Iberian Peninsula providedfundamental refugia for numerous mammals, such aswild rabbits (Branco et al., 2002), hares (Alves et al.,2008) or brown bears (Tarbelet & Bouvet, 1994).Accordingly, Iberia may have harboured one or morerelict wild boar populations during the Last GlacialMaximum, which could have contributed to therecolonization of Central and Northern Europe afterthe climate had become milder. Previous geneticstudies sustain that Spanish and Central Europeanwild boar are closely related, clustering in one mainEuropean group (Alves et al., 2003; Scandura et al.,2008). However, an extensive survey covering most ofthe Iberian Peninsula, including Portugal, has notbeen carried out so far.
Although previous archaeological and geneticevidences suggest that domestication of the wild boartook place in the Near East, at approximately 9000
years BP (Epstein & Bichard, 1984), in the light ofcomprehensive genetic investigations, Larson et al.(2005; 2007) proposed a different scenario, suggestingat least six independent centres of domestication.In their reconstruction, even if domesticated NearEastern pigs entered Central Europe with earlyNeolithic human migrations, they left no descendantsamong European pig breeds (Larson et al., 2007). Oneor more European centres of domestication were pro-posed by these authors at the origin of modern Euro-pean pig breeds. One of these centres might well belocated in the Iberian peninsula.
In recent years, studies concerning the geneticdiversity of livestock have been improved. The estab-lishment of conservation programmes became a pri-ority in many countries because local domestic breedsrepresent an important genetic and ecological com-ponent of biodiversity (Hall & Bradley, 1995). InPortugal, there are two autochthonous pig breeds,Alentejano and Bísaro. Because of certain morpho-logical adaptations to Mediterranean environments,the former is considered a typical Mediterraneanbreed, as is the Iberian pig in Spain (Toro et al., 2000;Reis, 2003). The Bísaro breed, a descendant from theCeltic lineage, is well adapted to northern Iberianclimate (Reis, 2003). Historical records indicate thatBísaro was extensively crossed with several importedbreeds in an attempt to improve its performance(Miranda do Vale, 1949). As a result, this breed wasalmost extinct and, currently, it is under a strictconservation programme. Mitochondrial (mt)DNAand microsatellite markers have been used to assessthe genetic diversity of Iberian pigs and to establishtheir relationships with other European breeds (Toroet al., 2002; Alves et al., 2003; Fabuel et al., 2004).However, these studies did not include the Portu-guese breeds.
This present study aims to provide new insight intothe genetic diversity of the Iberian wild boar popula-tions and their phylogenetic relationships with otherEuropean and Asian populations, and to analyse, forthe first time, the genetic relationships between Por-tuguese and several other European domestic breeds.To achieve these goals, we sequenced 741 bp of themtDNA control region, and assessed the geneticdiversity of 17 Iberian wild boar populations andAlentejano and Bísaro domestic breeds.
MATERIAL AND METHODSSAMPLE COLLECTION AND DNA EXTRACTION
Tissue and blood samples were collected from 138hunted wild boar, comprising 129 animals capturedfrom 17 populations in the Iberian Peninsula, six inFrance and three in Austria. Sampling sites and
798 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

number of samples collected at each site are shownin Figure 1. Twenty-two tissue samples were alsoobtained from the two domestic Portuguese breeds,Bísaro (Bis) and Alentejano (Ale). Total genomic DNAwas extracted using standard protocols.
DNA AMPLIFICATION AND SEQUENCING
A 1244-bp fragment of the mtDNA control region,between sites 15 390 and 16 634 (according to thereference sequence AJ002189; Ursing & Arnason,1998), was amplified by polymerase chain reaction(PCR) with the primers: 5′-CGCCATCAGCACCCAAAGCT-3′ and 5′-ATTTTGGGAGGTTATTGTGTTGTA-3′, forward and reverse, respectively (Alveset al., 2003). PCR was performed in a 13-mL volumecontaining 100 ng of DNA, 1¥ reaction buffer (Ecogen),1.5 mM MgCl2, 300 mM of each dNTP, 0.15 mM of eachprimer and 0.5 U of Taq DNA polymerase (Ecogen).Thermocycling conditions were: 94 °C for 4 min, fol-lowed by 35 cycles at 94 °C for 30 s, 62 °C for 30 s, and72 °C for 30 s, with a final extension of 72 °C for 5 min.
PCR products were then purified using MicroSpinS-400 HR Columns (Amersham Biosciences) and usedas sequencing templates. A second primer set (forward:5′-CCGTGGGGGTTTCTATTGA-3′; reverse: 5′-TGGGCGATTTTAGGTGAGATGGT-3′) (Alves et al., 2003)was used to determine the nucleotide sequence ofa 741-bp fragment of the control region on an ABIPRISM 3100 DNA sequencer, using the BigDye Termi-nator, version 3.0, Cycle Sequencing kit (AppliedBiosystems) in accordance with the manufacturer’sinstructions. GenBank accession numbers for the hap-lotype sequences described in the present study areHM747196 to HM747222.
STATISTICAL ANALYSIS
Sequences were aligned by eye using BIOEDIT, version5.0.9 (Hall, 1999) and compared with the mtDNApig reference sequence (GenBank accession numberAJ002189 – Ursing & Arnason, 1998). The tandemrepeat motif of 10 bp inserted in the D-loop region waseliminated before the sequence analysis. Levels of
Figure 1. Sampling sites of wild boar populations from Central Europe and Iberian Peninsula. In brackets are shownthe number of samples collected at each site. Aus, Austria (Vienne); Mad, Madine; Pre, Prémian; Pyr, Pyrenees; Ast,Asturias; Veb, Valle del Ebro; Bur, Burgos; Gua, Guadalajara; Mun, Montes Universales; Tol, Toledo; Alb, Albacete; Cre,Ciudad Real; Jae, Jaén; Sev, Sevilla; Bra, Bragança; Ses, Serra da Estrela; Rmo, Reguengos de Monsaraz; Cer, Cercal;Vgu, Vale do Guadiana; Alg, Algarve.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 799
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

molecular diversity such as haplotype (h) and nucle-otide (pn) diversities, mean number of pairwise nucle-otide differences (p) and number of segregating sites(S) were independently calculated for wild boarand domestic breeds with ARLEQUIN, version 2.0(Schneider, Roessli & Excoffier, 2000). Parameters pn
and p were computed according to Kimura two-parameters distance method (Kimura, 1980). Popula-tion genetic structure was estimated among andwithin populations by an analysis of molecular vari-ance (AMOVA) (Excoffier, Smouse & Quattro, 1992).The variance components and its proportion ofvariation expressed as percentage were calculated.Moreover, the significance of the variance amongpopulations and FST were tested by random permu-tation, under the null hypothesis that all samples aredrawn from a global population. These tests were alsoperformed in ARLEQUIN, version 2.0 (Schneider et al.,2000).
Intraspecific mtDNA phylogenies, particularly inthe case of control region, are frequently difficult toobtain as a result of high rates of homoplasy, andmedian networks are often a better approach thanphylogenetic trees (Bandelt et al., 1995). Further-more, for large sample sizes or for a large number ofhaplotypes, the complexity of a network can be solvedwith a median-joining (MJ) network algorithm(Bandelt, Forster & Röhl, 1999). Based on a 660-bpfragment of the mtDNA control region, a MJ networkwas constructed with a combination of all domesticand wild haplotypes obtained in the present study,plus 156 published sequences, in accordance with themethodology outlined by Bandelt et al. (1999) usingNETWORK, version 4.1.0 (Röhl, 2000) (further detailsabout haplotypes, their accession numbers, and ref-erences are provided in the Appendix, Table A1). Amaximum parsimony calculation was used to elimi-nate nonparsimonious links (Bandelt et al., 1999). Asecond MJ network was created with the same pro-cedure, using the shorter region (411 bp) reported byScandura et al. (2008). To reveal phylogeographicrelationships between the Iberian and other Euro-pean wild populations, our 138 new wild boarsequences were added to an alignment of 230 previ-ously published mtDNA control regions from severalEuropean countries.
The presence of widely distributed haplotypes canbe the result of natural (demographic and rangeexpansion) or anthropogenic processes (translocationsand reintroductions). Populations that experienced ademographic expansion usually present unimodalmismatch distributions, in opposite of populations atdemographic equilibrium, which generate multimodaldistributions (Rogers & Harpending, 1992). Thesmoothness of the distribution can be quantified bythe raggedness index, which takes larger values in
stationary populations (Harpending, 1994). To deter-mine whether Iberian wild boar experienced a demo-graphic expansion, a mismatch analysis was carriedout in the overall Iberian population, and the signifi-cance of the observed values was assessed after10 000 replicates, using a parametric bootstrapapproach, under the null hypothesis that the observeddata fit the sudden expansion model (Schneider &Excoffier, 1999). The mismatch analysis and the rag-gedness index were performed in ARLEQUIN, version2.0 (Schneider et al., 2000). The Fu’s FS and Tajima’sD-tests of neutrality were calculated, and their sig-nificance values were estimated after 5000 replicates,under the null hypothesis of selective neutrality andpopulation equilibrium using ARLEQUIN, version 2.0(Schneider et al., 2000). The Fu’s FS test of neutralityis very sensitive to demographic expansion, generallyleading to large negative values (Fu, 1997). Tajima’sD-test of neutrality (Tajima, 1989a), although lesspowerful than FS, can be useful when trying to dis-tinguish the effects of population expansion versusmutation rate heterogeneity on mismatch distribu-tions. They have opposite effects on D because suddenexpansion leads to negative D-values, whereasuneven mutation rates shift it towards more positivevalues (Tajima, 1989a, b; Aris-Brosou & Excoffier,1996).
RESULTSCONTROL REGION DIVERSITY AND
POPULATION STRUCTURE
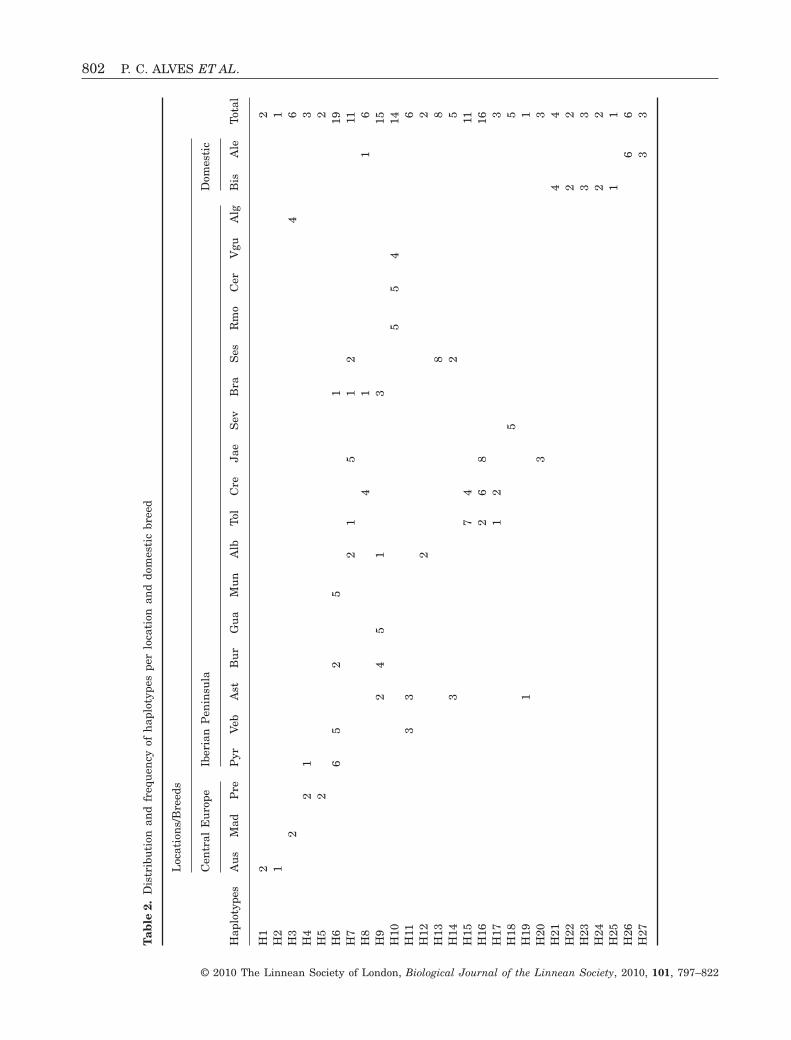
In a total of 160 sequences analyzed, 27 haplotypes(H1–H27) were defined by 24 variable sites. Theycomprise 22 transitions, one transversion, and oneinsertion/deletion of 1 bp at nucleotide 15 578(Table 1). The distribution of haplotypes per locationand breed is shown in Table 2. Five different wildboar mtDNA haplotypes were found in French andAustrian samples, two of which were also identified inthe Pyrenees (H4) and Algarve (H3). Some wild boarhaplotypes have high frequencies and are widely dis-tributed across the Iberian Peninsula (haplotypes H6,H7, H9, and H16). Nevertheless, some unique haplo-types are specific to a few locations, such as H12 inAlbacete, H18 in Sevilla, and H19 in Asturias. Thehighest haplotype diversity was observed amongnorth-western Iberia and central Spain populations,whereas the lowest levels were found in south-western Iberia. The domestic Portuguese breedsshare only one haplotype (H8) with the analyzedIberian wild boar. Bísaro shows the highest number ofhaplotypes and no common haplotypes were foundbetween this breed and Alentejano pigs (Table 2).Levels of gene diversity, number of segregating sites,
800 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

Tab
le1.
Var
iabl
epo
siti
ons
wit
hin
the
741-
bpfr
agm
ents
ofth
em
itoc
hon
dria
lD
NA
con
trol
regi
onfo
un
din
27(H
1–H
27)
hap
loty
pes
onbo
thw
ild
boar
and
dom
esti
cpi
gssa
mpl
edin
the
pres
ent
stu
dy
Hap
loty
pes
1545
015
558
1557
815
588
1560
415
616
1567
515
683
1569
515
714
1572
315
741
1575
815
822
1582
515
878
1593
616
010
1607
416
091
1609
216
127
1613
916
141
AJ0
0218
9T
A–
CG
TT
GC
TA
CT
AC
AA
AA
AC
AA
AH
1.
.–
..
..
..
..
..
..
..
..
..
G.
.H
2.
.–
T.
..
..
..
..
..
..
..
..
G.
.H
3.
T–
..
C.
..
C.
.C
..
..
..
..
G.
.H
4.
.–
..
..
..
..
.C
.T
..
..
G.
..
.H
5.
.–
..
..
..
..
.C
.T
G.
..
G.
..
.H
6C
.–
..
..
..
C.
..
..
G.
..
..
G.
.H
7.
.–
..
..
..
C.
.C
..
..
..
..
G.
.H
8.
.–
..
..
..
C.
..
..
..
..
..
..
.H
9.
.–
..
..
..
C.
.C
G.
..
..
..
..
.H
10.
.–
..
..
..
C.
.C
..
.G
..
..
..
.H
11.
.–
..
..
..
C.
.C
..
..
G.
..
G.
.H
12C
.–
..
..
..
C.
..
..
G.
..
.T
G.
.H
13.
.–
..
..
..
..
.C
G.
..
..
..
G.
.H
14.
.–
..
..
..
C.
..
..
..
..
..
G.
.H
15.
.–
.A
..
..
CG
.C
G.
..
..
..
G.
.H
16.
.–
..
..
..
CG
.C
..
.G
..
..
..
.H
17.
.–
..
..
..
C.
.C
G.
..
..
..
G.
.H
18.
.A
..
..
..
C.
TC
..
..
..
..
G.
.H
19.
.–
..
.C
..
C.
..
..
..
..
..
G.
.H
20C
.–
..
..
..
C.
..
..
..
..
..
G.
.H
21.
T–
..
..
..
C.
.C
..
..
..
..
G.
.H
22.
.–
..
..
..
C.
.C
..
..
..
..
.G
GH
23.
.–
..
..
A.
C.
..
G.
..
..
..
G.
.H
24.
T–
..
C.
..
C.
..
..
..
..
..
G.
.H
25.
T–
..
..
..
C.
TC
..
..
.G
..
..
.H
26.
.–
..
..
.T
C.
.C
..
..
..
..
GG
GH
27.
.–
..
..
..
C.
.C
..
..
..
..
GG
G
Seq
uen
ceid
enti
ties
and
inde
lsar
ere
pres
ente
dby
dots
and
dash
es,r
espe
ctiv
ely.
Nu
cleo
tide
posi
tion
sar
en
um
bere
dac
cord
ing
toth
ere
fere
nce
sequ
ence
Gen
Ban
kac
cess
ion
nu
mbe
rA
J002
189
(Urs
ing
&A
rnas
on,
1998
).
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 801
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Tab
le2.
Dis
trib
uti
onan
dfr
equ
ency
ofh
aplo
type
spe
rlo
cati
onan
ddo
mes
tic
bree
d
Hap
loty
pes
Loc
atio
ns/
Bre
eds
Tota
l
Cen
tral
Eu
rope
Iber
ian
Pen
insu
laD
omes
tic
Au
sM
adP
reP
yrV
ebA
stB
ur
Gu
aM
un
Alb
Tol
Cre
Jae
Sev
Bra
Ses
Rm
oC
erV
guA
lgB
isA
le
H1
22
H2
11
H3
24
6H
42
13
H5
22
H6
65
25
119
H7
21
51
211
H8
41
16
H9
24
51
315
H10
55
414
H11
33
6H
122
2H
138
8H
143
25
H15
74
11H
162
68
16H
171
23
H18
55
H19
11
H20
33
H21
44
H22
22
H23
33
H24
22
H25
11
H26
66
H27
33
802 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
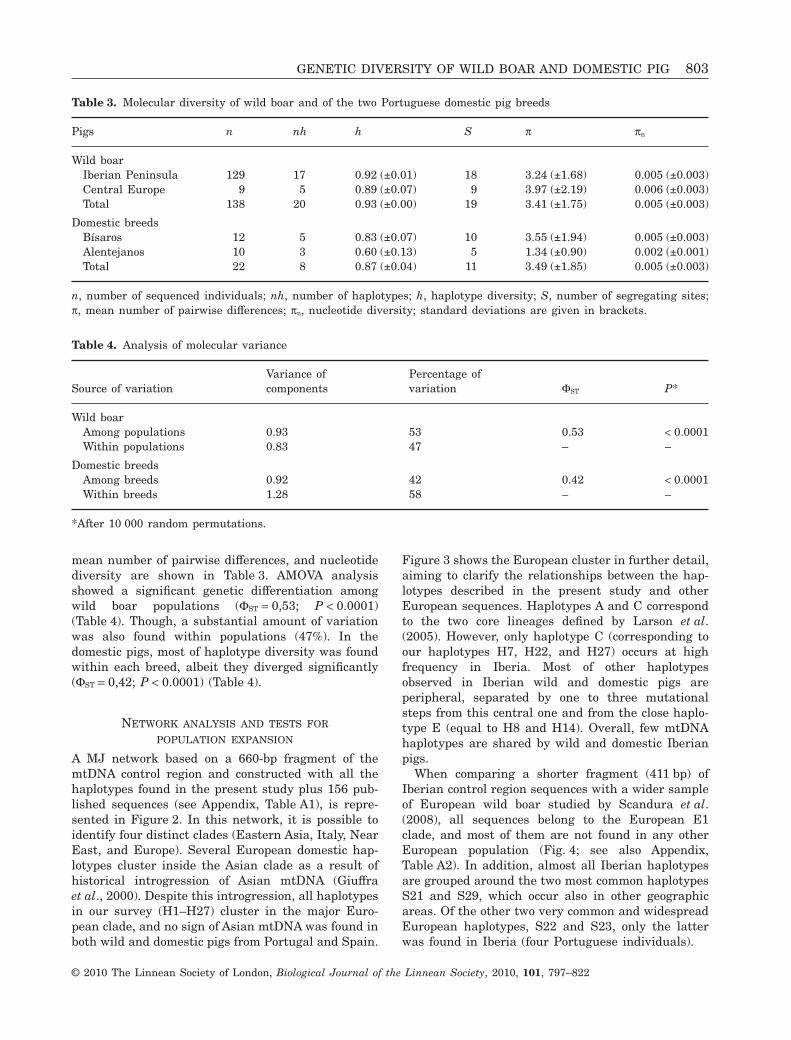
mean number of pairwise differences, and nucleotidediversity are shown in Table 3. AMOVA analysisshowed a significant genetic differentiation amongwild boar populations (FST = 0,53; P < 0.0001)(Table 4). Though, a substantial amount of variationwas also found within populations (47%). In thedomestic pigs, most of haplotype diversity was foundwithin each breed, albeit they diverged significantly(FST = 0,42; P < 0.0001) (Table 4).
NETWORK ANALYSIS AND TESTS FOR
POPULATION EXPANSION
A MJ network based on a 660-bp fragment of themtDNA control region and constructed with all thehaplotypes found in the present study plus 156 pub-lished sequences (see Appendix, Table A1), is repre-sented in Figure 2. In this network, it is possible toidentify four distinct clades (Eastern Asia, Italy, NearEast, and Europe). Several European domestic hap-lotypes cluster inside the Asian clade as a result ofhistorical introgression of Asian mtDNA (Giuffraet al., 2000). Despite this introgression, all haplotypesin our survey (H1–H27) cluster in the major Euro-pean clade, and no sign of Asian mtDNA was found inboth wild and domestic pigs from Portugal and Spain.
Figure 3 shows the European cluster in further detail,aiming to clarify the relationships between the hap-lotypes described in the present study and otherEuropean sequences. Haplotypes A and C correspondto the two core lineages defined by Larson et al.(2005). However, only haplotype C (corresponding toour haplotypes H7, H22, and H27) occurs at highfrequency in Iberia. Most of other haplotypesobserved in Iberian wild and domestic pigs areperipheral, separated by one to three mutationalsteps from this central one and from the close haplo-type E (equal to H8 and H14). Overall, few mtDNAhaplotypes are shared by wild and domestic Iberianpigs.
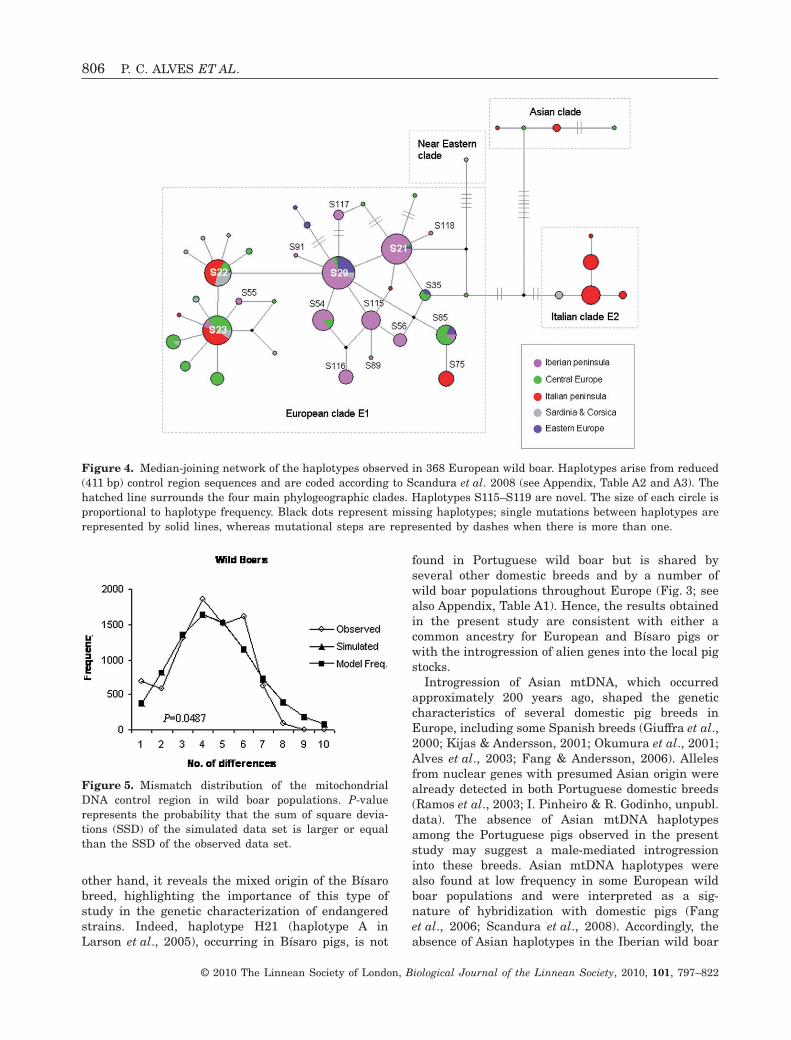
When comparing a shorter fragment (411 bp) ofIberian control region sequences with a wider sampleof European wild boar studied by Scandura et al.(2008), all sequences belong to the European E1clade, and most of them are not found in any otherEuropean population (Fig. 4; see also Appendix,Table A2). In addition, almost all Iberian haplotypesare grouped around the two most common haplotypesS21 and S29, which occur also in other geographicareas. Of the other two very common and widespreadEuropean haplotypes, S22 and S23, only the latterwas found in Iberia (four Portuguese individuals).
Table 3. Molecular diversity of wild boar and of the two Portuguese domestic pig breeds
Pigs n nh h S p pn
Wild boarIberian Peninsula 129 17 0.92 (±0.01) 18 3.24 (±1.68) 0.005 (±0.003)Central Europe 9 5 0.89 (±0.07) 9 3.97 (±2.19) 0.006 (±0.003)Total 138 20 0.93 (±0.00) 19 3.41 (±1.75) 0.005 (±0.003)
Domestic breedsBísaros 12 5 0.83 (±0.07) 10 3.55 (±1.94) 0.005 (±0.003)Alentejanos 10 3 0.60 (±0.13) 5 1.34 (±0.90) 0.002 (±0.001)Total 22 8 0.87 (±0.04) 11 3.49 (±1.85) 0.005 (±0.003)
n, number of sequenced individuals; nh, number of haplotypes; h, haplotype diversity; S, number of segregating sites;p, mean number of pairwise differences; pn, nucleotide diversity; standard deviations are given in brackets.
Table 4. Analysis of molecular variance
Source of variationVariance ofcomponents
Percentage ofvariation FST P*
Wild boarAmong populations 0.93 53 0.53 < 0.0001Within populations 0.83 47 – –
Domestic breedsAmong breeds 0.92 42 0.42 < 0.0001Within breeds 1.28 58 – –
*After 10 000 random permutations.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 803
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

The bell-shaped mismatch distribution (Fig. 5) andthe low and nonsignificant raggedness index (resultsnot shown) obtained for the Iberian wild boar popu-lation are consistent with demographic expansion.However, when the validity of the sudden expansionmodel was tested, the null hypothesis that theobserved data are compatible with the model wasrejected (P = 0.0487). Moreover, the Fu’s FS (-2.308)was not significant at the 5% level (P = 0.250) and theTajima’s D-test of neutrality (0.077) was not signifi-cantly different from zero (P > 0.1).
DISCUSSIONGENETIC VARIABILITY OF WILD BOAR POPULATIONS
Sequencing analysis of mitochondrial DNA controlregion revealed high levels of gene diversity in wildboar populations of the Iberian Peninsula. The resultsobtained by the AMOVA analysis show that most ofthe variation is observed among populations, a strongindication of their genetic structure. It is also possibleto recognize that a few haplotypes are exclusive tosome geographic locations (Table 2), providing evi-dence for this structure and for the uniqueness ofsome populations. Taken together with the high levelsof haplotype diversity obtained, this may reflect theexistence of glacial refugia for S. scrofa in the IberianPeninsula during the Last Glacial Maximum. On theother hand, haplotypes such as H6, H7, H9, and H16
are widely distributed, which suggests that they areancestral haplotypes or, alternatively, that they resultfrom expanding populations or from translocatedanimals. Considering that these haplotypes are at thetips of the network, they most likely have a recentorigin. Higher levels of haplotype variation have beenobserved in northern Portuguese and Spanish popu-lations, which is reasonable given that S. scrofa sur-vived in the wild in these areas when most of wildpopulations shrunk in both countries during the lastcentury (Bugalho et al., 1984). On the basis of arecent microsatellite investigation revealing a stronglatitudinal structuring in the Portuguese population,Ferreira et al. (2009) suggested that the wild popula-tions recovered almost simultaneously, albeit inde-pendently from diverse refuge areas. Their resultsindicate similar levels of diversity but a stronggenetic differentiation between Northern, Central,and Southern wild stocks. This contrasts with thelack of mitochondrial diversity we observed in theSouth, where only two haplotypes were found in fourgeographically distant locations. A different roleplayed by the two sexes in the recolonization processcan supposedly explain the observed differencebetween the molecular markers. The occurrence ofone haplotype (H3) shared between populations fromAlgarve and Northern France is likely to be a resultof a translocation. The closest genetic relationshipbetween H3 and other Iberian haplotypes, rather
Figure 2. Median-joining network based on a 660-bp fragment of mitochondrial DNA control region depicting therelationships between the main four clades described for wild and domestic pigs (Asia, Italy, Near East, and Europe). Thehaplotypes shown in grey correspond to animals of unknown status from either Corsica or Sardinia because the statusof so-called wild boar in both islands is controversial (Larson et al., 2005). Additional details about clades, haplotypes, andaccession numbers are provided in the Appendix (Table A1). Black dots represent missing haplotypes; mutations arerepresented by the nucleotide position in which they occurred (numeration according to the reference sequence GenBankaccession number AJ002189). Areas of circles are proportional to the number of sampled individuals.
804 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

than with H4 and H5 from Southern France (Fig. 3and Table 2), could be related to translocations fromIberia to France.
Demographic expansions, associated with post-glacial recolonization processes, lead to an almostPoisson distribution of pairwise nucleotide differences(Slatkin & Hudson, 1991), even though homoplasy atsome nucleotide sites can affect this pattern (Ber-torelle & Slatkin, 1995). This does not appear to bethe case for Iberian wild boar, where the mismatchanalysis fails to detect any signature of a past expan-sion, resulting from a rejection of the sudden expan-sion model and the nonsignificant tests of neutrality.Molecular diversity indices such as mean number ofpairwise nucleotide differences (p) and number ofsegregating sites (S) are also consistent with thisresult because they take lower values with anincreasing mutation rate heterogeneity (Aris-Brosou& Excoffier, 1996).
DOMESTIC PIG BREEDS
High levels of gene diversity were also found in thetwo Portuguese breeds. However, diversity valueswere slightly lower than wild boar, perhaps as aresult of a smaller sample size or to mtDNA incom-plete lineage sorting, reflecting the recent origin ofthese breeds (Hall & Bradley, 1995). These highvalues, especially in Bísaro, and the large geneticvariation within breeds can be interpreted as evi-dence of extensive crossbreeding with importedanimals. When comparing Portuguese pig sequenceswith other domestic breeds, it is possible to recognizethat almost all the haplotypes are shared withIberian pigs from Spain (e.g. haplotype C in Fig. 3).On the one hand, this supports the ancestry ofIberian pigs because the two Portuguese ancestralstrains of Alentejano contributed to their origin(Rodrigáñez et al., 2000; Toro et al., 2000); on the
Figure 3. Detail of the European clade shown in Fig. 2 depicting the relationships between wild and domestic pigs fromEurope. The A and C haplotypes correspond to the two core lineages described by Larson et al. (2005). Additional detailsabout clades, haplotypes, and accession numbers are provided in the Appendix (Table A1). Black dots represent missinghaplotypes, mutations are represented by the nucleotide position in which they occurred and areas of circles areproportional to the number of sampled individuals.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 805
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
other hand, it reveals the mixed origin of the Bísarobreed, highlighting the importance of this type ofstudy in the genetic characterization of endangeredstrains. Indeed, haplotype H21 (haplotype A inLarson et al., 2005), occurring in Bísaro pigs, is not
found in Portuguese wild boar but is shared byseveral other domestic breeds and by a number ofwild boar populations throughout Europe (Fig. 3; seealso Appendix, Table A1). Hence, the results obtainedin the present study are consistent with either acommon ancestry for European and Bísaro pigs orwith the introgression of alien genes into the local pigstocks.
Introgression of Asian mtDNA, which occurredapproximately 200 years ago, shaped the geneticcharacteristics of several domestic pig breeds inEurope, including some Spanish breeds (Giuffra et al.,2000; Kijas & Andersson, 2001; Okumura et al., 2001;Alves et al., 2003; Fang & Andersson, 2006). Allelesfrom nuclear genes with presumed Asian origin werealready detected in both Portuguese domestic breeds(Ramos et al., 2003; I. Pinheiro & R. Godinho, unpubl.data). The absence of Asian mtDNA haplotypesamong the Portuguese pigs observed in the presentstudy may suggest a male-mediated introgressioninto these breeds. Asian mtDNA haplotypes werealso found at low frequency in some European wildboar populations and were interpreted as a sig-nature of hybridization with domestic pigs (Fanget al., 2006; Scandura et al., 2008). Accordingly, theabsence of Asian haplotypes in the Iberian wild boar
Figure 4. Median-joining network of the haplotypes observed in 368 European wild boar. Haplotypes arise from reduced(411 bp) control region sequences and are coded according to Scandura et al. 2008 (see Appendix, Table A2 and A3). Thehatched line surrounds the four main phylogeographic clades. Haplotypes S115–S119 are novel. The size of each circle isproportional to haplotype frequency. Black dots represent missing haplotypes; single mutations between haplotypes arerepresented by solid lines, whereas mutational steps are represented by dashes when there is more than one.
Figure 5. Mismatch distribution of the mitochondrialDNA control region in wild boar populations. P-valuerepresents the probability that the sum of square devia-tions (SSD) of the simulated data set is larger or equalthan the SSD of the observed data set.
806 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

populations can be indicative of a limited historicalgene flow from domestic stocks to wild boar, at leastin Spain.
PHYLOGENETIC RELATIONSHIPS BETWEEN IBERIAN
WILD BOAR AND OTHER EUROPEAN POPULATIONS
The MJ network clearly shows that Iberian wild anddomestic pigs belong to the main European clade,which is sharply differentiated from a second Euro-pean (Italian) clade, from the Near Eastern clade andfrom the East-Asian clade (Larson et al., 2005). Thetwo core lineages (A and C) described by Larson et al.(2005) are indeed the most internal ones, each ofwhich is surrounded by a star-like pattern, consistentwith a recent population expansion, as previouslydescribed both in wild and domestic populations(Fang & Andersson, 2006; Scandura et al., 2008).Although C-like haplotypes are found both in Iberianpigs and wild boar, A-like haplotypes have not beendetected so far in wild populations of the IberianPeninsula. These findings corroborate the hypothesisof a possible post-glacial expansion from differentglacial refugia proposed by Scandura et al. (2008),where the Iberian relict populations would have con-tributed to the recolonization with the spread ofC-like haplotypes (Fig. 4, S29, S21 and surroundinghaplotypes). Some of these haplotypes would havebeen retained by locally domesticated pig breeds. Thishypothesis would be also consistent with the ideaproposed by Larson and coworkers’ that numerouswild boar lineages have been domesticated (Larsonet al., 2005). However, we cannot rule out the alter-native hypothesis that only a restricted area ofdomestication in Central Europe does exist, fromwhich subsequent migration and/or translocation ofanimals to the rest of Europe occurred. The intermin-gling pattern seen in the network (Fig. 3) is consis-tent with the former hypothesis. Moreover, the factthat wild and domestic forms can naturally hybridize,and the additional problems with feral pigs, may alsohave affected the results (Randi, 1995; Goulding,2001; Wilson, 2004). A wider sampling in Europe andin the Near East would be needed to confirm orexclude these hypotheses.
The present study provides new insight into thegenetic diversity of Iberian wild boar populations anddomestic Portuguese pig breeds, and their relation-ships with other Eurasian pigs. It also provides astarting point for exploring the origins and structureof Iberian wild boar populations. Nevertheless,because there are large differences between sexeswith respect to dispersal and hybridization in domes-tic pigs, the present study should also be extended tonuclear markers and to the Y chromosome. Singlenucleotide polymorphisms are becoming a marker of
choice in population ecology, evolution, and conserva-tion (Morin, Luikart & Wayne, 2004), and thereforecan provide new insight into the real amount ofhybridization between wild and domestic pigs in theIberian Peninsula, as well as help to clarify theirgenetic relationships and identify their geneticsources.
ACKNOWLEDGEMENTS
We thank to Pedro Santos from University ofÉvora (Portugal), Klaus Hackländer from Universityof Veterinary Medicine (Vienna) and the OfficeNational de la Chasse (France) for providing samplesfrom wild boar. Samples from Bísaro and Alentejanopigs were kindly provided by João Santos e Silva andEstação Zootécnica Nacional (Portugal), respectively.R. Godinho worked under a postdoctoral grant fromFCT (SFRH/BPD/36021/2007).
REFERENCES
Alves PC, Melo-Ferreira J, Branco M, Suchentrunk F,Ferrand N, Harris J. 2008. Evidence for genetic similarityof two allopatric European hares (Lepus corsicanus and L.castroviejoi) based on nuclear genes sequences. MolecularPhylogenetics and Evolution 46: 1191–1197.
Alves E, Óvilo C, Rodríguez MC, Silió L. 2003. Mitochon-drial DNA sequence variation and phylogenetic relation-ships among Iberian pigs and other domestic and wild pigpopulations. Animal Genetics 34: 319–324.
Apollonio M, Randi E, Toso S. 1988. The systematics of thewild boar (Sus scrofa L.) in Italy. Bollettino Di Zoologia 3:213–221.
Aris-Brosou S, Excoffier L. 1996. The impact of populationexpansion and mutation rate heterogeneity on DNAsequence polymorphism. Molecular Biology and Evolution13: 494–504.
Bandelt HJ, Forster P, Sykes BC, Richards MB. 1995.Mitochondrial portraits of human populations using mediannetworks. Genetics 141: 743–753.
Bandelt HJ, Forster P, Röhl A. 1999. Median-joining net-works for inferring intraspecific phylogenies. MolecularBiology and Evolution 16: 37–48.
Bertorelle G, Slatkin M. 1995. The number of segregatingsites in expanding human populations, with implications forestimates of demographic parameters. Molecular Biologyand Evolution 12: 887–892.
Branco M, Monnerot M, Ferrand N, Templeton R. 2002.Postglacial dispersal of the European rabbit (Oryctolaguscuniculus) on the Iberian Peninsula reconstructed fromnested clade and mismatch analyses of mitochondrial DNAgenetic variation. Evolution 56: 792–803.
Bugalho JF, Carvalho JS, Borges JF. 1984. Situation dusanglier au Portugal. Symposium International sur le San-glier. Toulouse, France. Les Colloques de l’INRA 22: 112–121.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 807
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

Epstein H, Bichard M. 1984. Pig. In: Mason IL, ed. Evolu-tion of domesticated animals. London: Longman, 145–162.
Excoffier L, Smouse PE, Quattro JM. 1992. Analysis ofmolecular variance inferred from metric distances amongDNA haplotypes: application to human mitochondrial DNArestriction data. Genetics 131: 479–491.
Fabuel E, Barragán C, Silió L, Rodríguez MC, Toro MA.2004. Analysis of genetic diversity and conservation priori-ties in Iberian pigs based on microsatellite markers. Hered-ity 93: 104–113.
Fang M, Andersson L. 2006. Mitochondrial diversity inEuropean and Chinese pigs is consistent with populationexpansions that occurred prior to domestication. Proceed-ings of the Royal Society of London Series B, BiologicalSciences 273: 1803–1810.
Fang M, Berg F, Ducos A, Andersson L. 2006. Mitochon-drial haplotypes of European wild boar with 2n = 36 areclosely related to those of European domestic pigs with2n = 38. Animal Genetics 37: 459–464.
Ferreira E, Souto L, Soares AMVM, Fonseca C. 2009.Genetic structure of the wild boar population in Portugal:evidence of a recent bottleneck. Mammalian Biology 74:274–285.
Fonseca C, Correia F. 2008. O javali, Azevedo J, ed. Miran-dela: Colecção Património Natural Transmontano.
Fu YX. 1997. Statistical tests of neutrality of mutationsagainst population growth, hitchhiking and backgroundselection. Genetics 147: 915–925.
Genov P. 1999. A review of the cranial characteristics of thewild boar (Sus scrofa Linnaeus 1758), with systematic con-clusions. Mammal Review 29: 205–238.
Giuffra E, Kijas JMH, Amarger V, Carlborg Ö, Jeon J-T,Andersson L. 2000. The origin of the domestic pig: inde-pendent domestication and subsequent introgression. Genet-ics 154: 1785–1791.
Gongora J, Fleming P, Spencer PBS, Mason R,Garkavenko O, Meyer J-N, Droegemueller C, Lee JH,Moran C. 2004. Phylogenetic relationships of Australianand New Zealand feral pigs assessed by mitochondrialcontrol region sequence and nuclear GPIP genotype.Molecular Phylogenetics and Evolution 33: 339–348.
Goulding MJ. 2001. Possible genetic sources of free-livingwild boar (Sus scrofa) in southern. England. MammalReview 31: 245–248.
Hall TA. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series 41: 95–98.
Hall SJG, Bradley DG. 1995. Conserving livestock breedbiodiversity. Trends in Ecology and Evolution 10: 267–270.
Harpending HC. 1994. Signature of ancient populationgrowth in a low-resolution mitochondrial DNA mismatchdistribution. Human Biology 66: 591–600.
Hewitt GM. 1999. Post-glacial re-colonization of Europeanbiota. Biological Journal of the Linnean Society 68: 87–112.
Kijas JMH, Andersson L. 2001. A phylogenetic study of theorigin of the domestic pig estimated from the near-completemtDNA genome. Journal of Molecular Evolution 52: 302–308.
Kimura M. 1980. A simple method for estimating evolution-ary rate of base substitution through comparative studies ofnucleotide sequences. Journal of Molecular Evolution 16:111–120.
Larson G, Dobney K, Albarella U, Fang M, Matisoo-Smith E, Robins J, Lowden S, Finlayson H, Brand T,Willerslev E, Rowney-Conwy P, Andersson L, CooperA. 2005. Worldwide phylogeography of wild boar revealsmultiple centers of pig domestication. Science 307: 1618–1621.
Larson G, Albarella U, Dobney K, Rowley-Conwy P,Schibler J, Tresset A, Vigne JD, Edwards CJ,Schlumbaum A, Dinu A, Balacsescu A, Dolman G,Tagliacozzo A, Manaseryan N, Miracle P, VanWijngaarden-Bakker L, Masseti M, Bradley DG,Cooper A. 2007. Ancient DNA, pig domestication, and thespread of the Neolithic into Europe. Proceedings of theNational Academy of Sciences of the United States ofAmerica 104: 15276–15281.
Miranda do Vale J. 1949. Gado Bissulco. Suínos, Bovinos,Arietinos e Caprinos. Lisboa: Livraria Sá da Costa.
Morin PA, Luikart G, Wayne RK, for the SNP workshopgroup. 2004. SNPs in ecology, evolution and conservation.Trends in Ecology and Evolution 19: 208–216.
Okumura N, Kurosawa Y, Kobayashi E, Watanobe T,Ishiguro N, Yasue H, Mitsuhashi T. 2001. Genetic rela-tionship amongst the major non-coding regions of mitochon-drial DNAs in wild boars and several breeds of domesticatedpigs. Animal Genetics 32: 139–147.
Ramos AM, Mestre R, Gouveia S, Evans G, Zhang Y,Cardoso MF, Rothschild MF, Plastow G, Rangel-Figueiredo T. 2003. Use of type I DNA markers for initialgenetic characterization of two Portuguese swine breeds.Archivos de Zootecnia 52: 255–264.
Randi E. 1995. Conservation genetics of the genus Sus. IBEXJournal of Mountain Ecology 3: 6–12.
Randi E, D’Huart JP, Lucchini V, Aman R. 2002.Evidence of two genetically deeply divergent species ofwaterhog, Phacochoerus africanus and P. aethiopicus(Artiodactyla: Suiformes) in East Africa. MammalianBiology 67: 91–96.
Reis J. 2003. Suínos. Lisboa: Direcção Geral de Veterinária.Rodrigáñez J, Toro MA, Rodríguez C, Silió L. 2000.
Alleles survival from Portuguese and Spanish strains in apopulation of Iberian pig. In: Afonso JA, Tirapicos JL, eds.Tradition and innovation in Mediterranean pig production.Zaragoza: CIHEAM-EU, 57–61.
Rogers AR, Harpending H. 1992. Population growth makeswaves in the distribution of pairwise genetic differences.Molecular Biology and Evolution 9: 552–569.
Röhl A. 2000. Network. Clare: Fluxus Technology Ltd.Sáez-Royuela C, Tellería JL. 1986. The increased popula-
tion of the wild boar (Sus scrofa L.) in Europe. MammalReview 16: 97–101.
Scandura M, Iacolina L, Crestanello B, Pecchioli E, DiBenedetto MF, Russo V, Davoli R, Apollonio M, Ber-torelle G. 2008. Ancient vs. recent processes as factorsshaping the genetic variation of the European wild boar: are
808 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

the effects of the last glaciation still detectable? MolecularEcology 17: 1745–1762.
Schneider S, Excoffier L. 1999. Estimation of past demo-graphic parameters from the distribution of pairwise differ-ences when the mutation rates vary among sites:application to human mitochondrial DNA. Genetics 152:1079–1089.
Schneider S, Roessli D, Excoffier L. 2000. Arlequin 2.0. Asoftware for population genetic data analysis. Geneva:Genetics and Biometry Laboratory, Department of Anthro-pology, University of Geneva.
Slatkin M, Hudson RR. 1991. Pairwise comparisons of mito-chondrial DNA sequences in stable and exponential growingpopulations. Genetics 129: 555–562.
Sommer RS, Nadachowski A. 2006. Glacial refugia ofmammals in Europe: evidence from fossil records. MammalReview 36: 251–265.
Tajima F. 1989a. Statistical method for testing the neutralmutation hypothesis by DNA polymorphism. Genetics 123:585–595.
Tajima F. 1989b. The effect of change in population size onDNA polymorphism. Genetics 123: 597–601.
Tarbelet P, Bouvet J. 1994. Mitochondrial DNA poly-morphism, phylogeography, and conservation genetics ofthe brown bear (Ursus arctos) in Europe. Proceedings of theRoyal Society of London Series B, Biological Sciences 255:195–200.
Tellería JC, Saez-Royuela YC. 1985. L’évolutiondémographique du sanglier (Sus scrofa) en Espagne.Mammalia 49: 195–202.
Toro MA, Rodrigañez J, Silió L, Rodríguez C. 2000.Genealogical analysis of a close herd of Black hairlessIberian pigs. Conservation Biology 14: 1843–1851.
Toro M, Barragan C, Ovilo C, Rodrigañez J, RodríguezC, Silió L. 2002. Estimation of coancestry in Iberian pigsusing molecular markers. Conservation Genetics 3: 309–320.
Ursing BM, Arnason U. 1998. The complete mitochondrialDNA sequence of the pig (Sus scrofa). Journal of MolecularEvolution 47: 302–306.
Vernesi C, Crestanello B, Pecchioli E, Tartari D, Cara-melli D, Hauffe H, Bertorelle G. 2003. The geneticimpact of demographic decline and reintroduction in thewild boar (Sus scrofa): a microsatellite analysis. MolecularEcology 12: 585–595.
Watanobe T, Okumura N, Ishiguro N, Nakano M, MatsuiA, Sahara M, Komatsu M. 1999. Genetic relationship anddistribution of the Japanese wild boar (Sus scrofa leucomys-tax) and Ryukyu wild boar (Sus scrofa riukiuanus) analysedby mitochondrial DNA. Molecular Ecology 8: 1509–1512.
Wilson CJ. 2004. Rooting damage to farmland in Dorset,southern England, caused by feral wild boar Sus scrofa.Mammal Review 4: 331–335.
Yalden D. 1999. The history of British mammals. London:Poyser.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 809
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
APPENDIX
Table A1. Locations, status, accession numbers and references of the haplotypes used to construct the median-joiningnetwork depicted in Figs 2, 3, plus the haplotypes found in the present study
Code Status Breed LocationAccessionnumber
Haplotype(Larsonet al., 2005) Reference
Asian cladeGL77 Wild boar – Iran AY884638 – Larson et al. (2005)GL191 Wild boar – Russia AY884691 CB Larson et al. (2005)GL192 Wild boar – Russia AY884692 CB Larson et al. (2005)GL237 Wild boar – Russia AY884711 – Larson et al. (2005)SLEU1 Wild boar – Japan AB015084 – Watanobe et al. (1999)SLEU2 Wild boar – Japan AB015085 – Watanobe et al. (1999)SLEU3 Wild boar – Japan AB015086 – Watanobe et al. (1999)SRIU1 Wild boar – Ryukyu AB015089 – Watanobe et al. (1999)SRIU2 Wild boar – Ryukyu AB015090 – Watanobe et al. (1999)MJ01 Domestic pig Machado Jabugo Spain AY884782 D Larson et al. (2005)FRCR01 Domestic pig Creole France France AY884779 D Larson et al. (2005)DLAN11 Domestic pig Landrace UK AY884750 L Larson et al. (2005)DLAN13 Domestic pig Landrace France AY884760 D Larson et al. (2005)DLW02 Domestic pig Large white Germany AY884786 FH Larson et al. (2005)DLW06 Domestic pig Large white UK AY884762 FH Larson et al. (2005)DLW07 Domestic pig Large white UK AY884785 D Larson et al. (2005)DLW10 Domestic pig Large white Germany AY884772 GX Larson et al. (2005)BRIHS1 Domestic pig British old spot UK AY884757 FH Larson et al. (2005)LB01 Domestic pig Large black UK AY884784 GX Larson et al. (2005)DUKTAM Domestic pig Tamworth UK AY884767 D Larson et al. (2005)YORK Domestic pig Yorkshire ? AY243481 – Cho, I.C., Jung, Y.H.,
Jeon, J.T. and Kim, B.W.(unpubl. data)
BERK1 Domestic pig Berkshire ? AY429459 – Cho, I.C., Jung, Y.H.,Jeon, J.T. and Kim, B.W.(unpubl. data)
BERK2 Domestic pig Berkshire ? AF276936 – Cho, I.C., Jung, Y.H.,Jeon, J.T. and Kim, B.W.(unpubl. data)
LWH2 Domestic pig Large white ? AY232882 – Alves et al. (2003)LWH3 Domestic pig Large white ? AY232883 – Alves et al. (2003)PIET3 Domestic pig Pietrain ? AY232887 – Alves et al. (2003)DUR5 Domestic pig Duroc ? AY232879 – Alves et al. (2003)SBJA Domestic pig Spotted black
JabugoSpain AY232890 – Alves et al. (2003)
MEIS1 Domestic pig Meishan ? AY232888 – Alves et al. (2003)WAN Domestic pig Wan’na China AF276924 – Kim et al. (2002)
Near Eastern cladeGL55 Wild boar – Turkey AY884619 – Larson et al. (2005)GL59 Wild boar – Iran AY884622 BQ Larson et al. (2005)GL142 Wild boar – Armenia AY884680 BQ Larson et al. (2005)GL193 Wild boar – Armenia AY884693 – Larson et al. (2005)GL194 Wild boar – Armenia AY884694 – Larson et al. (2005)GL236 Wild boar – Armenia AY884710 BQ Larson et al. (2005)GL254 Wild boar – Iran AY884725 – Larson et al. (2005)GL270 Wild boar – Armenia AY884726 – Larson et al. (2005)GL271 Wild boar – Armenia AY884727 BQ Larson et al. (2005)
Italian cladeGL65 Unknown – Sardinia AY884628 – Larson et al. (2005)GL190 Unknown – Sardinia AY884690 – Larson et al. (2005)GL244 Wild boar – Italy AY884716 – Larson et al. (2005)GL245 Wild boar – Italy AY884717 BH Larson et al. (2005)
810 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A1. Continued
Code Status Breed LocationAccessionnumber
Haplotype(Larsonet al., 2005) Reference
GL246 Wild boar – Italy AY884718 BH Larson et al. (2005)GL247 Wild boar – Italy AY884719 BG Larson et al. (2005)GL248 Wild boar – Italy AY884720 BG Larson et al. (2005)GL249 Wild boar – Italy AY884721 – Larson et al. (2005)GL250 Wild boar – Italy AY884722 BH Larson et al. (2005)GL251 Wild boar – Italy AY884723 BH Larson et al. (2005)ITWB1 Wild boar – Italy AB015094 BH Watanobe et al. (1999)ITWB2 Wild boar – Italy AB015095 BG Watanobe et al. (1999)
European cladeGL52 Wild boar – Spain AY884616 – Larson et al. (2005)GL63 Wild boar – Germany AY884626 A Larson et al. (2005)GL71 Wild boar – France AY884633 AJ Larson et al. (2005)GL73 Wild boar – Morocco AY884635 C Larson et al. (2005)GL107 Wild boar – Germany AY884664 – Larson et al. (2005)GL108 Wild boar – Germany AY884665 A Larson et al. (2005)GL109 Wild boar – Germany AY884666 C Larson et al. (2005)GL110 Unknown – Corsica AY884667 A Larson et al. (2005)GL111 Unknown – Sardinia AY884668 A Larson et al. (2005)GL112 Wild boar – Holland AY884669 A Larson et al. (2005)GL113 Wild boar – Macedonia AY884670 – Larson et al. (2005)GL133 Feral – Norway AY884672 E Larson et al. (2005)GL141 Wild boar – Armenia AY884679 A Larson et al. (2005)GL143 Unknown – Corsica AY884681 – Larson et al. (2005)GL144 Wild boar – Sardinia AY884682 AJ Larson et al. (2005)GL221 Wild boar – Spain AY884697 – Larson et al. (2005)GL222 Wild boar – Portugal AY884698 – Larson et al. (2005)GL242 Wild boar – Spain AY884714 – Larson et al. (2005)GL252 Wild boar – Italy AY884724 A Larson et al. (2005)GL284 Unknown – Corsica AY884728 A Larson et al. (2005)GL285 Unknown – Corsica AY884729 A Larson et al. (2005)GL286 Unknown – Corsica AY884730 – Larson et al. (2005)GL287 Unknown – Corsica AY884731 BK Larson et al. (2005)GL288 Unknown – Sardinia AY884732 A Larson et al. (2005)GL289 Unknown – Sardinia AY884733 BK Larson et al. (2005)SWB1 Wild boar – Spain AY232868 BY Alves et al. (2003)SWB2 Wild boar – Spain AY232869 BY Alves et al. (2003)SWB3 Wild boar – Spain AY232870 BY Alves et al. (2003)SWB4 Wild boar – Spain AY232871 CA Alves et al. (2003)SWB5 Wild boar – Spain AY232872 CA Alves et al. (2003)SWB6 Wild boar – Spain AY232873 – Alves et al. (2003)SWB7 Wild boar – Spain AY232874 – Alves et al. (2003)FIN1 Wild boar – Finland AF535163 BA Gongora et al. (2004)FIN2 Wild boar – Finland AF535164 – Gongora et al. (2004)LCOR82 Unknown – Corsica AY884796 BK Larson et al. (2005)LSAR88 Unknown – Sardinia AY884795 BK Larson et al. (2005)DIBBL1 Domestic pig Iberian Black Spain AY884765 C Larson et al. (2005)DMIDW1 Domestic pig Middle White UK AY884770 C Larson et al. (2005)BRIL1 Domestic pig British lop
earedUK AY884756 C Larson et al. (2005)
BK01 Domestic pig Berkshire UK AY884783 A Larson et al. (2005)GERBUN Domestic pig German Bunte Germany AY884754 A Larson et al. (2005)DLEIC1 Domestic pig Leicoma UK AY884759 A Larson et al. (2005)DHAMP1 Domestic pig Hampshire UK AY884777 A Larson et al. (2005)DHAMP2 Domestic pig Hampshire Germany AY884758 – Larson et al. (2005)DLAN04 Domestic pig Landrace Denmark AY884747 BK Larson et al. (2005)DLAN05 Domestic pig Landrace Denmark AY884776 BK Larson et al. (2005)
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 811
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

Table A1. Continued
Code Status Breed LocationAccessionnumber
Haplotype(Larsonet al., 2005) Reference
DLAN06 Domestic pig Landrace Finland AY884775 N Larson et al. (2005)DLAN08 Domestic pig Landrace Norway AY884748 – Larson et al. (2005)DLAN09 Domestic pig Landrace Iceland AY884749 E Larson et al. (2005)DLAN10 Domestic pig Landrace UK AY884774 BK Larson et al. (2005)DLAN12 Domestic pig Landrace UK AY884773 BK Larson et al. (2005)DLAN14 Domestic pig Landrace Germany AY884787 A Larson et al. (2005)DPIE03 Domestic pig Pietrain Germany AY884769 R Larson et al. (2005)DPIE05 Domestic pig Pietrain France AY884766 A Larson et al. (2005)DBIS01 Domestic pig Bísaro Portugal AY884755 A Larson et al. (2005)DDUR02 Domestic pig Duroc UK AY884746 A Larson et al. (2005)DDUR03 Domestic pig Duroc Germany AY884778 E Larson et al. (2005)DNEGCA Domestic pig Negro Canaria Spain AY884752 – Larson et al. (2005)BRISA1 Domestic pig Bristish
saddlebackUK AY884780 – Larson et al. (2005)
DLW05 Domestic pig Large white UK AY884761 – Larson et al. (2005)DLW08 Domestic pig Large white France AY884763 M Larson et al. (2005)DHM01 Domestic pig Hungary
MangalicaHungary AY884764 BK Larson et al. (2005)
DSWELI Domestic pig SwedishLinderod
Sweden AY884751 AF Larson et al. (2005)
WELSH Domestic pig Welsh China AF276937 U Kim et al. (2002)HAMP Domestic pig Hampshire ? AY429460 – Cho, I.C., Jung, Y.H.,
Jeon, J.T. and Kim, B.W.(unpubl. data)
DUR Domestic pig Duroc ? AY243482 – Cho, I.C., Jung, Y.H.,Jeon, J.T. and Kim, B.W.(unpubl. data)
DUR1 Domestic pig Duroc ? AY232875 M Alves et al. (2003)DUR2 Domestic pig Duroc ? AY232876 M Alves et al. (2003)DUR3 Domestic pig Duroc ? AY232877 A Alves et al. (2003)DUR4 Domestic pig Duroc ? AY232878 BK Alves et al. (2003)DUR6 Domestic pig Duroc ? AY232879 V Alves et al. (2003)DUR7 Domestic pig Duroc ? AY232880 – Alves et al. (2003)PIET1 Domestic pig Pietrain ? AY23O820 – Yue G.H. (unpubl. data)PIET2 Domestic pig Pietrain ? AY232886 A Alves et al. (2003)LWH1 Domestic pig Large white ? AY230822 A Yue G.H. (unpubl. data)LAND1 Domestic pig Landrace ? AY232884 A Alves et al. (2003)LAND2 Domestic pig Landrace ? AY232885 Q Alves et al. (2003)HMAN Domestic pig Hungary
MangalitzaHungary AY232892 A Alves et al. (2003)
BASQ Domestic pig Basque Spain AY232891 A Alves et al. (2003)IP1 Domestic pig Blond
AlentejanoSpain AY232842 BZ Alves et al. (2003)
IP2 Domestic pig BlondAlentejano
Spain AY232843 BZ Alves et al. (2003)
IP3 Domestic pig Black hairless Spain AY232844 C Alves et al. (2003)IP4 Domestic pig Portuguese red Spain AY232845 C Alves et al. (2003)IP5 Domestic pig Black hairless Spain AY232846 – Alves et al. (2003)IP6 Domestic pig Black hairless Spain AY232847 – Alves et al. (2003)IP7 Domestic pig Black hairless Spain AY232848 – Alves et al. (2003)IP8 Domestic pig Black hairless Spain AY232849 W Alves et al. (2003)IP9 Domestic pig Black hairless Spain AY232850 C Alves et al. (2003)IP10 Domestic pig Black hairless Spain AY232851 – Alves et al. (2003)IP11 Domestic pig Black hairless Spain AY232852 O Alves et al. (2003)IP12 Domestic pig Black hairless Spain AY232853 C Alves et al. (2003)IP13 Domestic pig Black hairless Spain AY232854 – Alves et al. (2003)
812 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A1. Continued
Code Status Breed LocationAccessionnumber
Haplotype(Larsonet al., 2005) Reference
IP14 Domestic pig Black hairless Spain AY232855 C Alves et al. (2003)IP15 Domestic pig Iberian red Spain AY232856 A Alves et al. (2003)IP16 Domestic pig Iberian red Spain AY232857 C Alves et al. (2003)IP17 Domestic pig Iberian red Spain AY232858 BK Alves et al. (2003)IP18 Domestic pig Iberian red Spain AY232859 C Alves et al. (2003)IP19 Domestic pig Iberian red Spain AY232860 – Alves et al. (2003)IP20 Domestic pig Iberian red Spain AY232861 BZ Alves et al. (2003)IP21 Domestic pig Black hairy Spain AY232862 O Alves et al. (2003)IP22 Domestic pig Black hairy Spain AY232863 – Alves et al. (2003)IP23 Domestic pig Black hairy Spain AY232864 E Alves et al. (2003)IP24 Domestic pig Black hairy Spain AY232865 – Alves et al. (2003)IP25 Domestic pig Black hairy Spain AY232866 C Alves et al. (2003)IP26 Domestic pig Iberian blond Spain AY232867 A Alves et al. (2003)H1 N Present studyH2 – Present studyH3 BK Present studyH4 – Present studyH5 – Present studyH6 – Present studyH7 C Present studyH8 E Present studyH9 – Present studyH10 – Present studyH11 – Present studyH12 – Present studyH13 – Present studyH14 E Present studyH15 – Present studyH16 BY Present studyH17 – Present studyH18 – Present studyH19 – Present studyH20 – Present studyH21 A Present studyH22 C Present studyH23 – Present studyH24 – Present studyH25 – Present studyH26 O Present studyH27 C Present study
When haplotypes were common to those described by Larson et al. (2005), letter codes are given.
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 813
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. List of the European wild boar sequences used to construct the median-joining network shown in Fig. 4
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
EWB1 Wild boar 1 Poland AF136555 S034 Giuffra et al. (2000)EWB2 Wild boar 1 Poland AF136556 S029 Giuffra et al. (2000)EWB3 Wild boar 1 Italy AF136563 S012 Giuffra et al. (2000)SwWB Wild boar 1 Sweden AF304203 S022 Kijas & Andersson (2001)
Wild boar 1 Germany D42180 S004 Okumura et al. (2001)Wild boar 2 Italy AB015094 S011 Okumura et al. (2001)Wild boar 1 Italy AB015095 S012 Okumura et al. (2001)
EWB4 Wild boar 1 Italy AB059651 S033 Okumura et al. (2001)Wild boar 1 Bulgaria AJ314542 S029 Randi et al. (2002)Unknown 1 Sardinia AJ314544 S022 Randi et al. (2002)
FIN1 Wild boar 1 Finland AF535163 S034 Gongora et al. (2004)FIN2 Wild boar 1 Finland AF535164 S049 Gongora et al. (2004)SWB1 Wild boar 1 Spain AY232868 S054 Alves et al. (2003)SWB2 Wild boar 1 Spain AY232869 S054 Alves et al. (2003)SWB3 Wild boar 1 Spain AY232870 S054 Alves et al. (2003)SWB4 Wild boar 1 Spain AY232871 S055 Alves et al. (2003)SWB5 Wild boar 1 Spain AY232872 S055 Alves et al. (2003)SWB6 Wild boar 1 Spain AY232873 S021 Alves et al. (2003)SWB7 Wild boar 1 Spain AY232874 S056 Alves et al. (2003)GL52 Wild boar 1 Spain AY884616 S089 Larson et al. (2005)GL63 Wild boar 1 Germany AY884626 S022 Larson et al. (2005)GL65 Unknown 1 Sardinia AY884628 S061 Larson et al. (2005)GL71 Wild boar 1 France AY884633 S065 Larson et al. (2005)GL107 Wild boar 1 Germany AY884664 S023 Larson et al. (2005)GL108 Wild boar 1 Germany AY884665 S022 Larson et al. (2005)GL109 Wild boar 1 Germany AY884666 S029 Larson et al. (2005)GL110 Unknown 1 Corsica AY884667 S022 Larson et al. (2005)GL111 Unknown 1 Sardinia AY884668 S022 Larson et al. (2005)GL112 Wild boar 1 Holland AY884669 S022 Larson et al. (2005)GL113 Wild boar 1 Macedonia AY884670 S021 Larson et al. (2005)GL143 Unknown 1 Corsica AY884681 S083 Larson et al. (2005)GL144 Unknown 1 Sardinia AY884682 S065 Larson et al. (2005)GL190 Unknown 1 Sardinia AY884690 S061 Larson et al. (2005)GL220 Unknown 1 Corsica AY884696 S090 Larson et al. (2005)GL221 Wild boar 1 Spain AY884697 S029 Larson et al. (2005)GL222 Wild boar 1 Portugal AY884698 S091 Larson et al. (2005)GL242 Wild boar 1 Spain AY884714 S091 Larson et al. (2005)GL244 Wild boar 1 Italy AY884716 S069 Larson et al. (2005)GL245 Wild boar 1 Italy AY884717 S011 Larson et al. (2005)GL246 Wild boar 1 Italy AY884718 S011 Larson et al. (2005)GL247 Wild boar 1 Italy AY884719 S012 Larson et al. (2005)GL248 Wild boar 1 Italy AY884720 S012 Larson et al. (2005)GL249 Wild boar 1 Italy AY884721 S012 Larson et al. (2005)GL250 Wild boar 1 Italy AY884722 S011 Larson et al. (2005)GL251 Wild boar 1 Italy AY884723 S011 Larson et al. (2005)GL252 Wild boar 1 Italy AY884724 S022 Larson et al. (2005)GL284 Unknown 1 Corsica AY884728 S022 Larson et al. (2005)GL285 Unknown 1 Corsica AY884729 S022 Larson et al. (2005)GL287 Unknown 1 Corsica AY884731 S023 Larson et al. (2005)GL288 Unknown 1 Sardinia AY884732 S022 Larson et al. (2005)GL289 Unknown 1 Sardinia AY884733 S023 Larson et al. (2005)LNSar88 Unknown 1 Sardinia AY884795 S023 Larson et al. (2005)LCorsica82 Unknown 1 Corsica AY884796 S023 Larson et al. (2005)
814 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
LeifFrance Wild boar 1 France AY884815 S092 Larson et al. (2005)S4 Wild boar 1 France DQ379232 S023 Fang et al. (2006)Sm10 Wild boar 1 France DQ379233 S023 Fang et al. (2006)78 Wild boar 1 Belgium DQ379234 S031 Fang et al. (2006)81 Wild boar 1 Belgium DQ379235 S054 Fang et al. (2006)83 Wild boar 1 Belgium DQ379236 S054 Fang et al. (2006)89 Wild boar 1 Belgium DQ379237 S054 Fang et al. (2006)96 Wild boar 1 Belgium DQ379238 S104 Fang et al. (2006)105 Wild boar 1 Belgium DQ379239 S104 Fang et al. (2006)113 Wild boar 1 Belgium DQ379240 S009 Fang et al. (2006)114 Wild boar 1 Belgium DQ379241 S021 Fang et al. (2006)mitS1 Wild boar 1 France DQ379242 S085 Fang et al. (2006)mitS2 Wild boar 1 France DQ379243 S085 Fang et al. (2006)mitS3 Wild boar 1 France DQ379244 S111 Fang et al. (2006)Sm1 Wild boar 1 France DQ379245 S085 Fang et al. (2006)Sm2 Wild boar 1 France DQ379246 S085 Fang et al. (2006)Sm3 Wild boar 1 France DQ379247 S085 Fang et al. (2006)Sm4 Wild boar 1 France DQ379248 S085 Fang et al. (2006)Sm5 Wild boar 1 France DQ379249 S085 Fang et al. (2006)Sm6 Wild boar 1 France DQ379250 S085 Fang et al. (2006)Sm7 Wild boar 1 France DQ379251 S085 Fang et al. (2006)Sm8 Wild boar 1 France DQ379252 S085 Fang et al. (2006)Sm9 Wild boar 1 France DQ379253 S022 Fang et al. (2006)S1 Wild boar 1 France DQ379254 S092 Fang et al. (2006)S2 Wild boar 1 France DQ379255 S092 Fang et al. (2006)S3 Wild boar 1 France DQ379256 S092 Fang et al. (2006)S6 Wild boar 1 France DQ379257 S092 Fang et al. (2006)S7 Wild boar 1 France DQ379258 S092 Fang et al. (2006)S8 Wild boar 1 France DQ379259 S092 Fang et al. (2006S9 Wild boar 1 France DQ379260 S092 Fang et al. (2006S10 Wild boar 1 France DQ379261 S092 Fang et al. (2006)AR16 Wild boar 1 Italy EU362409 S075 Scandura et al. (2008)AR17 Wild boar 1 Italy EU362410 S023 Scandura et al. (2008)AR18 Wild boar 1 Italy EU362411 S075 Scandura et al. (2008)AR20 Wild boar 1 Italy EU362412 S022 Scandura et al. (2008)AR21 Wild boar 1 Italy EU362413 S011 Scandura et al. (2008)AR22 Wild boar 1 Italy EU362414 S022 Scandura et al. (2008)AR23 Wild boar 1 Italy EU362415 S022 Scandura et al. (2008)AR24 Wild boar 1 Italy EU362416 S011 Scandura et al. (2008)FO1 Wild boar 1 Italy EU362417 S075 Scandura et al. (2008)FO2 Wild boar 1 Italy EU362418 S023 Scandura et al. (2008)FO3 Wild boar 1 Italy EU362419 S023 Scandura et al. (2008)FO4 Wild boar 1 Italy EU362420 S023 Scandura et al. (2008)FO5 Wild boar 1 Italy EU362421 S022 Scandura et al. (2008)FO6 Wild boar 1 Italy EU362422 S075 Scandura et al. (2008)FO7 Wild boar 1 Italy EU362423 S075 Scandura et al. (2008)FO8 Wild boar 1 Italy EU362424 S022 Scandura et al. (2008)FO9 Wild boar 1 Italy EU362425 S075 Scandura et al. (2008)FO10 Wild boar 1 Italy EU362426 S023 Scandura et al. (2008)SI1 Wild boar 1 Italy EU362427 S023 Scandura et al. (2008)SI2 Wild boar 1 Italy EU362428 S023 Scandura et al. (2008)SI3 Wild boar 1 Italy EU362429 S023 Scandura et al. (2008)SI4 Wild boar 1 Italy EU362430 S023 Scandura et al. (2008)
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 815
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
SI5 Wild boar 1 Italy EU362431 S023 Scandura et al. (2008)SI6 Wild boar 1 Italy EU362432 S023 Scandura et al. (2008)SI7 Wild boar 1 Italy EU362433 S023 Scandura et al. (2008)SI9 Wild boar 1 Italy EU362434 S023 Scandura et al. (2008)SA1 Wild boar 1 Italy EU362435 S076 Scandura et al. (2008)SA2 Wild boar 1 Italy EU362436 S023 Scandura et al. (2008)SA3 Wild boar 1 Italy EU362437 S077 Scandura et al. (2008)SA4 Wild boar 1 Italy EU362438 S075 Scandura et al. (2008)SA5 Wild boar 1 Italy EU362439 S023 Scandura et al. (2008)SA6 Wild boar 1 Italy EU362440 S015 Scandura et al. (2008)SA7 Wild boar 1 Italy EU362441 S015 Scandura et al. (2008)SA8 Wild boar 1 Italy EU362442 S075 Scandura et al. (2008)SA9 Wild boar 1 Italy EU362443 S012 Scandura et al. (2008)SA10 Wild boar 1 Italy EU362444 S015 Scandura et al. (2008)FI27 Wild boar 1 Italy EU362445 S023 Scandura et al. (2008)FI29 Wild boar 1 Italy EU362446 S023 Scandura et al. (2008)FI30 Wild boar 1 Italy EU362447 S023 Scandura et al. (2008)FI31 Wild boar 1 Italy EU362448 S022 Scandura et al. (2008)FI33 Wild boar 1 Italy EU362449 S022 Scandura et al. (2008)FI34 Wild boar 1 Italy EU362450 S022 Scandura et al. (2008)FI36 Wild boar 1 Italy EU362451 S075 Scandura et al. (2008)FI37 Wild boar 1 Italy EU362452 S022 Scandura et al. (2008)FI40 Wild boar 1 Italy EU362453 S012 Scandura et al. (2008)FI47 Wild boar 1 Italy EU362454 S075 Scandura et al. (2008)MR1 Wild boar 1 Italy EU362455 S011 Scandura et al. (2008)MR2 Wild boar 1 Italy EU362456 S011 Scandura et al. (2008)MR3 Wild boar 1 Italy EU362457 S011 Scandura et al. (2008)MR4 Wild boar 1 Italy EU362458 S011 Scandura et al. (2008)MR5 Wild boar 1 Italy EU362459 S011 Scandura et al. (2008)MR6 Wild boar 1 Italy EU362460 S011 Scandura et al. (2008)MR8 Wild boar 1 Italy EU362461 S075 Scandura et al. (2008)MR9 Wild boar 1 Italy EU362462 S011 Scandura et al. (2008)MR11 Wild boar 1 Italy EU362463 S075 Scandura et al. (2008)MR12 Wild boar 1 Italy EU362464 S075 Scandura et al. (2008)MR14 Wild boar 1 Italy EU362465 S011 Scandura et al. (2008)SR2 Wild boar 1 Italy EU362466 S022 Scandura et al. (2008)SR6 Wild boar 1 Italy EU362467 S022 Scandura et al. (2008)SR10 Wild boar 1 Italy EU362468 S022 Scandura et al. (2008)SR11 Wild boar 1 Italy EU362469 S011 Scandura et al. (2008)SR12 Wild boar 1 Italy EU362470 S022 Scandura et al. (2008)SR15 Wild boar 1 Italy EU362471 S022 Scandura et al. (2008)SR16 Wild boar 1 Italy EU362472 S022 Scandura et al. (2008)SR17 Wild boar 1 Italy EU362473 S022 Scandura et al. (2008)SR18 Wild boar 1 Italy EU362474 S022 Scandura et al. (2008)SR20 Wild boar 1 Italy EU362475 S022 Scandura et al. (2008)CP1 Wild boar 1 Italy EU362476 S011 Scandura et al. (2008)CP6 Wild boar 1 Italy EU362477 S078 Scandura et al. (2008)CP8 Wild boar 1 Italy EU362478 S012 Scandura et al. (2008)CP10 Wild boar 1 Italy EU362479 S012 Scandura et al. (2008)CP13 Wild boar 1 Italy EU362480 S012 Scandura et al. (2008)CP14 Wild boar 1 Italy EU362481 S012 Scandura et al. (2008)CP15 Wild boar 1 Italy EU362482 S012 Scandura et al. (2008)CP16 Wild boar 1 Italy EU362483 S012 Scandura et al. (2008)
816 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
CP20 Wild boar 1 Italy EU362484 S078 Scandura et al. (2008)CP26 Wild boar 1 Italy EU362485 S078 Scandura et al. (2008)SS1 Unknown 1 Sardinia EU362486 S061 Scandura et al. (2008)SS4 Unknown 1 Sardinia EU362487 S022 Scandura et al. (2008)SS16 Unknown 1 Sardinia EU362488 S022 Scandura et al. (2008)SS33 Unknown 1 Sardinia EU362489 S022 Scandura et al. (2008)SS36 Unknown 1 Sardinia EU362490 S029 Scandura et al. (2008)SS37 Unknown 1 Sardinia EU362491 S029 Scandura et al. (2008)SS38 Unknown 1 Sardinia EU362492 S029 Scandura et al. (2008)SS42 Unknown 1 Sardinia EU362493 S079 Scandura et al. (2008)SS43 Unknown 1 Sardinia EU362494 S080 Scandura et al. (2008)SS45 Unknown 1 Sardinia EU362495 S081 Scandura et al. (2008)SS48 Unknown 1 Sardinia EU362496 S022 Scandura et al. (2008)SS50 Unknown 1 Sardinia EU362497 S082 Scandura et al. (2008)GO1 Wild boar 1 Gorice EU362498 S083 Scandura et al. (2008)GO4 Wild boar 1 Gorice EU362499 S083 Scandura et al. (2008)GO5 Wild boar 1 Gorice EU362500 S083 Scandura et al. (2008)GO6 Wild boar 1 Gorice EU362501 S083 Scandura et al. (2008)GO11 Wild boar 1 Gorice EU362502 S083 Scandura et al. (2008)GO13 Wild boar 1 Gorice EU362503 S083 Scandura et al. (2008)GO15 Wild boar 1 Gorice EU362504 S083 Scandura et al. (2008)GO18 Wild boar 1 Gorice EU362505 S083 Scandura et al. (2008)GO19 Wild boar 1 Gorice EU362506 S083 Scandura et al. (2008)SP2 Wild boar 1 Spain EU362507 S021 Scandura et al. (2008)SP3 Wild boar 1 Spain EU362508 S021 Scandura et al. (2008)SP6 Wild boar 1 Spain EU362509 S021 Scandura et al. (2008)SP8 Wild boar 1 Spain EU362510 S021 Scandura et al. (2008)SP9 Wild boar 1 Spain EU362511 S021 Scandura et al. (2008)SP10 Wild boar 1 Spain EU362512 S021 Scandura et al. (2008)SP12 Wild boar 1 Spain EU362513 S021 Scandura et al. (2008)SP13 Wild boar 1 Spain EU362514 S021 Scandura et al. (2008)SP15 Wild boar 1 Spain EU362515 S021 Scandura et al. (2008)FR2 Wild boar 1 France EU362516 S023 Scandura et al. (2008)FR3 Wild boar 1 France EU362517 S023 Scandura et al. (2008)FR6 Wild boar 1 France EU362518 S023 Scandura et al. (2008)FR8 Wild boar 1 France EU362519 S023 Scandura et al. (2008)FR10 Wild boar 1 France EU362520 S023 Scandura et al. (2008)FR11 Wild boar 1 France EU362521 S022 Scandura et al. (2008)FR13 Wild boar 1 France EU362522 S023 Scandura et al. (2008)FR15 Wild boar 1 France EU362523 S023 Scandura et al. (2008)FR16 Wild boar 1 France EU362524 S023 Scandura et al. (2008)FR20 Wild boar 1 France EU362525 S023 Scandura et al. (2008)AS1 Wild boar 1 Austria EU362526 S084 Scandura et al. (2008)AS3 Wild boar 1 Austria EU362527 S029 Scandura et al. (2008)AS4 Wild boar 1 Austria EU362528 S035 Scandura et al. (2008)AS5 Wild boar 1 Austria EU362529 S084 Scandura et al. (2008)AS6 Wild boar 1 Austria EU362530 S084 Scandura et al. (2008)AS10 Wild boar 1 Austria EU362531 S084 Scandura et al. (2008)AS11 Wild boar 1 Austria EU362532 S035 Scandura et al. (2008)AS12 Wild boar 1 Austria EU362533 S035 Scandura et al. (2008)AS13 Wild boar 1 Austria EU362534 S023 Scandura et al. (2008)AS14 Wild boar 1 Austria EU362535 S084 Scandura et al. (2008)PO8 Wild boar 1 Poland EU362536 S029 Scandura et al. (2008)
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 817
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
PO11 Wild boar 1 Poland EU362537 S085 Scandura et al. (2008)PO12 Wild boar 1 Poland EU362538 S029 Scandura et al. (2008)PO14 Wild boar 1 Poland EU362539 S085 Scandura et al. (2008)PO16 Wild boar 1 Poland EU362540 S085 Scandura et al. (2008)PO24 Wild boar 1 Poland EU362541 S085 Scandura et al. (2008)PO25 Wild boar 1 Poland EU362542 S029 Scandura et al. (2008)PO27 Wild boar 1 Poland EU362543 S029 Scandura et al. (2008)UN1 Wild boar 1 Hungary EU362544 S029 Scandura et al. (2008)UN2 Wild boar 1 Hungary EU362545 S029 Scandura et al. (2008)UN6 Wild boar 1 Hungary EU362546 S029 Scandura et al. (2008)UN9 Wild boar 1 Hungary EU362547 S029 Scandura et al. (2008)UN10 Wild boar 1 Hungary EU362548 S029 Scandura et al. (2008)UN15 Wild boar 1 Hungary EU362549 S029 Scandura et al. (2008)UN17 Wild boar 1 Hungary EU362550 S029 Scandura et al. (2008)UN18 Wild boar 1 Hungary EU362551 S029 Scandura et al. (2008)UN24 Wild boar 1 Hungary EU362552 S035 Scandura et al. (2008)UN28 Wild boar 1 Hungary EU362553 S035 Scandura et al. (2008)Jav01Ses Wild boar 1 Portugal S056 Present studyJav14Ses Wild boar 1 Portugal S056 Present studyJav15Ses Wild boar 1 Portugal S021 Present studyJav21Ses Wild boar 1 Portugal S056 Present studyJav27Ses Wild boar 1 Portugal S056 Present studyJav28Ses Wild boar 1 Portugal S056 Present studyJav29Ses Wild boar 1 Portugal S029 Present studyJav30Ses Wild boar 1 Portugal S056 Present studyJav31Ses Wild boar 1 Portugal S029 Present studyJav33Ses Wild boar 1 Portugal S056 Present studyJav34Ses Wild boar 1 Portugal S021 Present studyJav35Ses Wild boar 1 Portugal S056 Present studyJav36Cer Wild boar 1 Portugal S029 Present studyJav37Cer Wild boar 1 Portugal S029 Present studyJav38Cer Wild boar 1 Portugal S029 Present studyJav39Cer Wild boar 1 Portugal S029 Present studyJav40Cer Wild boar 1 Portugal S029 Present studyJav41Rmo Wild boar 1 Portugal S029 Present studyJav42Rmo Wild boar 1 Portugal S029 Present studyJav43Rmo Wild boar 1 Portugal S029 Present studyJav44Rmo Wild boar 1 Portugal S029 Present studyJav45Rmo Wild boar 1 Portugal S029 Present studyJav83Bra Wild boar 1 Portugal S021 Present studyJav84Bra Wild boar 1 Portugal S029 Present studyJav85Bra Wild boar 1 Portugal S021 Present studyJav86Bra Wild boar 1 Portugal S115 Present studyJav87Bra Wild boar 1 Portugal S115 Present studyJav89Bra Wild boar 1 Portugal S115 Present studyJav47Vgu Wild boar 1 Portugal S029 Present studyJav50Vgu Wild boar 1 Portugal S029 Present studyJav54Vgu Wild boar 1 Portugal S029 Present studyJav56Vgu Wild boar 1 Portugal S029 Present studyJav59Alg Wild boar 1 Portugal S023 Present studyJav61Alg Wild boar 1 Portugal S023 Present studyJav65Alg Wild boar 1 Portugal S023 Present studyJav67Alg Wild boar 1 Portugal S023 Present study
818 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
Jve01Jae Wild boar 1 Spain S054 Present studyJve02Jae Wild boar 1 Spain S054 Present studyJve46Jae Wild boar 1 Spain S054 Present studyJve47Jae Wild boar 1 Spain S054 Present studyJve48Jae Wild boar 1 Spain S029 Present studyJve49Jae Wild boar 1 Spain S021 Present studyJve50Jae Wild boar 1 Spain S021 Present studyJve66Jae Wild boar 1 Spain S021 Present studyJve67Jae Wild boar 1 Spain S054 Present studyJve68Jae Wild boar 1 Spain S054 Present studyJve69Jae Wild boar 1 Spain S054 Present studyJve70Jae Wild boar 1 Spain S054 Present studyJve71Jae Wild boar 1 Spain S029 Present studyJve72Jae Wild boar 1 Spain S029 Present studyJve73Jae Wild boar 1 Spain S029 Present studyJve74Jae Wild boar 1 Spain S029 Present studyJve11Cre Wild boar 1 Spain S054 Present studyJve12Cre Wild boar 1 Spain S054 Present studyJve13Cre Wild boar 1 Spain S054 Present studyJve14Cre Wild boar 1 Spain S054 Present studyJve15Cre Wild boar 1 Spain S054 Present studyJve34Cre Wild boar 1 Spain S054 Present studyJve35Cre Wild boar 1 Spain S116 Present studyJve36Cre Wild boar 1 Spain S116 Present studyJve37Cre Wild boar 1 Spain S116 Present studyJve38Cre Wild boar 1 Spain S115 Present studyJve39Cre Wild boar 1 Spain S116 Present studyJve40Cre Wild boar 1 Spain S115 Present studyJve82Cre Wild boar 1 Spain S021 Present studyJve83Cre Wild boar 1 Spain S021 Present studyJve84Cre Wild boar 1 Spain S021 Present studyJve85Cre Wild boar 1 Spain S021 Present studyJve16Tol Wild boar 1 Spain S116 Present studyJve17Tol Wild boar 1 Spain S116 Present studyJve18Tol Wild boar 1 Spain S116 Present studyJve19Tol Wild boar 1 Spain S116 Present studyJve20Tol Wild boar 1 Spain S116 Present studyJve21Tol Wild boar 1 Spain S054 Present studyJve22Tol Wild boar 1 Spain S116 Present studyJve23Tol Wild boar 1 Spain S054 Present studyJve24Tol Wild boar 1 Spain S116 Present studyJve25Tol Wild boar 1 Spain S029 Present studyJve26Tol Wild boar 1 Spain S115 Present studyJve76Sev Wild boar 1 Spain S117 Present studyJve77Sev Wild boar 1 Spain S117 Present studyJve78Sev Wild boar 1 Spain S117 Present studyJve79Sev Wild boar 1 Spain S117 Present studyJve80Sev Wild boar 1 Spain S117 Present studyJve86Ast Wild boar 1 Spain S115 Present studyJve87Ast Wild boar 1 Spain S115 Present studyJve88Ast Wild boar 1 Spain S021 Present studyJve89Ast Wild boar 1 Spain S021 Present studyJve90Ast Wild boar 1 Spain S021 Present study
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 819
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Table A2. Continued
Code StatusNumber ofindividuals Location
Accessionnumber Haplotype Reference
Jve91Ast Wild boar 1 Spain S029 Present studyJve92Ast Wild boar 1 Spain S117 Present studyJve94Ast Wild boar 1 Spain S029 Present studyJve95Ast Wild boar 1 Spain S029 Present studyJve102Pyr Wild boar 1 Spain S021 Present studyJve104Pyr Wild boar 1 Spain S085 Present studyJve105Pyr Wild boar 1 Spain S021 Present studyJve106Pyr Wild boar 1 Spain S021 Present studyJve110Pyr Wild boar 1 Spain S021 Present studyJve111Pyr Wild boar 1 Spain S021 Present studyJve112Pyr Wild boar 1 Spain S021 Present studyJve115Veb Wild boar 1 Spain S021 Present studyJve116Veb Wild boar 1 Spain S021 Present studyJve117Veb Wild boar 1 Spain S021 Present studyJve119Veb Wild boar 1 Spain S021 Present studyJve120Veb Wild boar 1 Spain S029 Present studyJve124Veb Wild boar 1 Spain S029 Present studyJve125Veb Wild boar 1 Spain S029 Present studyJve127Veb Wild boar 1 Spain S021 Present studyJve143Gua Wild boar 1 Spain S115 Present studyJve145Gua Wild boar 1 Spain S115 Present studyJve144Gua Wild boar 1 Spain S115 Present studyJve146Gua Wild boar 1 Spain S115 Present studyJve147Gua Wild boar 1 Spain S115 Present studyJve128Mun Wild boar 1 Spain S021 Present studyJve129Mun Wild boar 1 Spain S021 Present studyJve131Mun Wild boar 1 Spain S021 Present studyJve132Mun Wild boar 1 Spain S021 Present studyJve133Mun Wild boar 1 Spain S021 Present studyJve134Bur Wild boar 1 Spain S115 Present studyJve135Bur Wild boar 1 Spain S115 Present studyJve136Bur Wild boar 1 Spain S021 Present studyJve138Bur Wild boar 1 Spain S115 Present studyJve140Bur Wild boar 1 Spain S115 Present studyJve141Bur Wild boar 1 Spain S021 Present studyJve148Alb Wild boar 1 Spain S029 Present studyJve151Alb Wild boar 1 Spain S021 Present studyJve155Alb Wild boar 1 Spain S115 Present studyJve157Alb Wild boar 1 Spain S021 Present studyJve159Alb Wild boar 1 Spain S029 Present studyJvFr26Mad Wild boar 1 France S023 Present studyJvFr27Mad Wild boar 1 France S023 Present studyJvFr28Pre Wild boar 1 France S085 Present studyJvFr32Pre Wild boar 1 France S085 Present studyJvFr28bPre Wild boar 1 France S085 Present studyJvFr32bPre Wild boar 1 France S085 Present studyJvAust01 Wild boar 1 Austria S035 Present studyJvAust07 Wild boar 1 Austria S036 Present studyJvAust07b Wild boar 1 Austria S119 Present study
Codes are adapted to match those used by Scandura et al. (2008) (i.e. haplotype S021 corresponds to their haplotypeH021). Novel haplotypes (S115–S119) are shown in bold.
820 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

Table A3. Frequency of the 42 haplotypes found in European wild boar, whose relationships are shown in themedian-joining network in Fig. 4
Geographicarea
Region:reference
Asian clade (A)
Neareasternclade European clade (E1)
S04 S09 S15 S33 S90 S21 S22 S23 S29 S31 S34 S35 S49 S54 S55 S56 S65 S75 S76 S77 S79 S80
Iberianpeninsula
Spain: Alveset al. (2003)
1 3 2 1
Spain: Larsonet al. (2005)
1
Spain: Scanduraet al. (2008)
9
Spain: presentstudy
30 14 16
Portugal: Larsonet al. (2005)
Portugal:present study
4 4 17 8
CentralEurope
France: Larsonet al. (2005)
1
France: Fanget al. (2006
1 2
France:Scanduraet al. (2008)
1 9
France: presentstudy
2
Belgium: Fanget al. (2006
1 1 1 3
Holland: Larsonet al. (2005)
1
Germany:Okumuraet al. (2001
1
Germany:Larson et al.(2005)
2 1 1
Austria:Scanduraet al. (2008)
1 1 3
Austria: presentstudy
2
North-EastItaly:Scanduraet al. (2008)
Italianpeninsula
Italy: Giuffraet al. (2000
Italy: Okumuraet al. (2001
1
Centre-SouthItaly: Larsonet al. (2005)
1
Centre-SouthItaly:Scanduraet al. (2008)
3 18 18 13 1 1
Sardinia &Corsica
Sardinia: Randiet al. (2002
1
Sardinia: Larsonet al. (2005)
2 2 1
Sardinia:Scanduraet al. (2008)
4 3 1 1 1
Corsica: Larsonet al. (2005)
1 3 2
EasternEurope
Poland: Giuffraet al. (2000
1 1
Poland:Scanduraet al. (2008)
4
Hungary:Scanduraet al. (2008)
8 2
Macedonia:Larson et al.(2005)
1
Bulgaria: Randiet al. (2002
1
Sweden: Kijas &Andersson(2001)
1
Finland:Gongora et al.(2004)
1 1
Total 1 1 3 1 1 46 35 41 51 1 2 6 1 22 2 9 2 13 1 1 1 1
GENETIC DIVERSITY OF WILD BOAR AND DOMESTIC PIG 821
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822

Table A3. Continued
Geographicarea
Region:reference
European clade (E1) Italian clade (E2)
TotalNumber ofHaplotypesS81 S82 S83 S84 S85 S89 S91 S92 S104 S111 S115 S116 S117 S118 S119 S11 S12 S61 S69 S78
Iberianpeninsula
Spain: Alveset al. (2003)
7 4
Spain: Larsonet al. (2005)
1 1 3 3
Spain: Scanduraet al. (2008)
9 1
Spain: presentstudy
1 15 11 5 1 93 8
Portugal: Larsonet al. (2005)
1 1 1
Portugal:present study
3 36 5
CentralEurope
France: Larsonet al. (2005)
1 2 2
France: Fanget al. (2006)
10 8 1 22 5
France:Scanduraet al. (2008)
10 2
France: presentstudy
4 6 2
Belgium: Fanget al. (2006
2 8 5
Holland: Larsonet al. (2005)
1 1
Germany:Okumuraet al. (2001)
1 1
Germany:Larson et al.(2005)
4 3
Austria:Scanduraet al. (2008)
5 10 4
Austria: presentstudy
1 3 3
North-EastItaly:Scanduraet al. (2008)
9 9 1
Italianpeninsula
Italy: Giuffraet al. (2000)
1 1 1
Italy: Okumuraet al. (2001)
2 1 4 3
Centre-SouthItaly: Larsonet al. (2005)
4 3 1 9 4
Centre-SouthItaly:Scanduraet al. (2008)
12 8 3 77 9
Sardinia &Corsica
Sardinia: Randiet al. (2002)
1 1
Sardinia: Larsonet al. (2005)
2 7 4
Sardinia:Scanduraet al. (2008)
1 1 12 7
Corsica: Larsonet al. (2005)
1 7 4
EasternEurope
Poland: Giuffraet al. (2000)
2 2
Poland:Scanduraet al. (2008)
4 8 2
Hungary:Scanduraet al. (2008)
10 2
Macedonia:Larson et al.(2005)
1 1
Bulgaria: Randiet al. (2002)
1 1
Sweden: Kijas &Andersson(2001)
1 1
Finland:Gongora et al.(2004)
2 2
Total 1 1 10 5 20 1 1 9 2 1 18 11 5 1 1 18 13 3 1 3 368 43
Coded are adapted to match those used by Scandura et al. (2008) (i.e. haplotype S021 corresponds to their haplotype H021). Haplotypes are partitioned into the four cladescorresponding to Eastern Asia (A), Near East (NE), Europe (E1), and Italy (E2).
822 P. C. ALVES ET AL.
© 2010 The Linnean Society of London, Biological Journal of the Linnean Society, 2010, 101, 797–822
Copyright © 2022 FDOKUMEN




















