
Zonadhesin Is Essential for Species Specificity of Sperm Adhesion to the Egg Zona Pellucida
Upload
nevada-renoCategory
view
1download
0
Developmental Biology 222, 99-109 (2000) doi:10.1006/dbio.2000.9707, available ol~lillc at http://www.idealibrary.com on | ~ f ~ ' ~ . @
Zona Pellucida Protein Binding Ability Sperm during Epididymal Maturation and the Acrosome Reaction
of Porcine
®
Heather R. Burkin and David J. Miller ~ Departmel~t of Animal Sciences, U~iw~rsity of Illinois at Urba~2a-Champaign, Urban& Illi~ois 61801
In many mammals, the first interaction between gametes during fertilization occurs when sperm contact the zona pellucida surrounding the egg. Although porcine sperm first contact the zona pellucida via their plasma membrane, the regions of the sperm surface that display zona receptors have not been determined. We have used the Alexa 488 fluorophore conjugated to solnbilized porcine zona pellucida proteins to observe zona receptors on live boar sperm. Zona proteins bound live, acrosome-intact sperm on the anterior portion of the sperm head, concentrated in a thin band over the acrosomal ridge. When sperm membranes were permeabilized by fixation or acrosome reactions induced by the ionophore A23187, zona binding was extended to a broad area covering the entire acrosomal region. Zona binding proteins were present in the acrosomes of sperm from all regions of the epididymis. In contrast, zona binding sites were found on the plasma membrane of most sperm from the corpus and cauda epididymis, but on only 6% of caput epididymal sperm. In conclusion, acrosome-intact boar sperm exhibit concentrated zona protein binding over the acrosomal ridge and acquire this binding in the corpus region of the epididymis, correlating with the developmental stage at which sperm gain the ability to fertilize oocytes. © 2000 Academic Press
Key Words: fertilization; zona pellucida; sperm; acrosome; epididymis.
INTRODUCTION
The zona pellucida is an extracellular glycoprotein coat that surrounds the oocyte and, in many n~ammals, provides the first barrier sperm must penetrate for fertilization to occur. The molecular interactions between the sperm and zona pellucida appear to be relatively species-specific (Peterson et al., 1980), but have been studied in most detail in the mouse. The mouse zona pellucida is composed of three glycoproteins termed ZP1, ZP2, and ZP3 in order of decreasing molecular weight (reviewed in Wassarman, 1988). In the mouse, a surface form of /3-1,4- galactosyltransferase serves as a primary sperm receptor for O-linked oligosaccharides of ZP3 (Miller et ai, 1992). Aggregation of/3-1,4-galactosyltransferase by ZP3 induces sperm to undergo the acrosome reaction (Macek et al., 1991 ), resulting in the release of the acrosomal vesicle. The sperm acrosome reaction must occur before sperm can
i To whom correspondence should be addressed. Fax: (217) 333- 8286. E-mail: [email protected].
0012-1606/00 $35.00 Copyright © 2000 by Academic Press All rights of reproduction in any form reserved.
penetrate the zona and reach the egg plasma membrane (Saling et aI., 1979). Unexpectedly, sperm from /3-1,4- galactosyltransferase knockout mice bind to the zona pel- lucida at higher levels than wild-type sperm although they are unable to undergo the acrosome reaction in response to ZP3 (Lu and Shur, 1997). Subsequent to the acrosome reaction, murine ZP2 may serve as a secondary sperm receptor by binding molecules on the sperm inner acroso- real membrane (Mortillo and Wassarman, 1991). Since knockout mice for either the ZP3 receptor /3-1,4- galactosyltransferase (Lu and Shur, 1997) or the putative ZP2 receptor proacrosin are fertile, there appears to be some level of redundancy in zona pellucida binding molecules on mouse sperm (Baba et al., 1994; Adham et al., 1997).
Because of the relative ease of obtaining gametes required for zona pellucida isolation and fertilization studies, the pig is a useful organism for comparing various steps in the fertilization process to those in the mouse. Like the mouse zona, the porcine zona pellucida is also composed of three : glycoproteins which we will refer to as ssZPA (MW 70 kDa), ssZPB (MW 59 kDa), and ssZPC (MW 46 k D a )
99

100 Burk in and Mil ler
according to recently proposed nomencla tu re (Harris et al., 1994; Hedrick, 1996). Based on sequence data, these glyco- proteins appear to be homologues of mouse ZP2, ZP1, and ZP3, respectively (Harris et al., 1994). Due to extensive glycosylation, porcine zona proteins migrate as broad bands on SDS-PAGE gels and are difficuk to separate biochemi- cally; however, recent data suggest that ssZPB and ssZPC are pr imari ly responsible for sperm binding (Yurewicz et aL, 1998}. Al though /3-1,4-galactosyltransferase can bind to porcine zona proteins roughly the size of ssZPB and ssZPC, this protein is not necessary or sufficient for at taching boar sperm to the intact zona pellucida in vi tro (Rebeiz and Miller, 1999). Direct knowledge of the location of zona pellucida receptors on boar sperm wil l be useful in deter- m i n i n g the potent ial roles of various zona b ind ing proteins that have been identified in the boar (Hardy and Gathers, 1994; Mori et al., 1995; Geng et al., 1997; Enssl in et al., 1998).
Al though binding and penetra t ion through the zona pel- lucida are essential for successful fertilization, the exact regions of sperm that are capable of b inding the zona pellucida have remained controversial (Jones et al., 1991; Yurewicz et al., 1993; Yonezawa et aI., 1995; Fazeli et al., 1995, 1997). We have used fluorescence microscopy to localize solubilized zona pellucida proteins on live acrosomeAntact and acrosome-reacted boar sperm. In addi- tion, we have determined the developmental stage at which boar sperm begin to display receptors for the zona pellucida.
MATERIALS AND METHODS
Isolation and Solubilization of Zona Pellucida Proteins
Zona petlucida proteins were prepared as described previously (Dunbar et aL, 1980; Rebeiz and Miller, 1999). Porcine ovaries were disrupted with razor blades in dmKRBT medimn {120 mM NaC1, 2 mM KC1, 2 mM CaC12, 10 mM NaHCO3, 1.2 mM MgSO4, 0.36 mM NaH~PO4, 5.6 mM glucose, 1.1 mM pyruvic acid, 25 mM TAPSO, 18.5 mM sucrose, 0,6% BSA, 10 units/nrl penicillin, 10 /,g/lnl streptomycin, pH 7.3) to free oocytes from follicles. Oocytes were centrifuged at 800g for 10 rain, followed by one wash -with dmKRBT. Pellets were homogenized with a Polytron in dmKRBT containing 1 >g/ml leupeptin, 1 >g/ml pepstatin, 10/zg/ml phenyl- methylsulfonyl fluoride, 0.3 mg/ml DNase 1, and 0.3 mg/ml hyaluronidase. Additional homogenizations were performed after the addition of NP-40 to 1% and repeated after the addition of deoxycholic acid to 1%. Homogenates were separated on two successive 50% Pereoll gradients at 44,000g for 70 min at 3°C. Zona pellucida fragments were recovered, washed several times with dmKRBT, and checked for purity under a microscope. Zona proteins were washed at least four times in 10 mM Tris, pH 9.5, 100 mM NaC1 and solubilized for 20 rain at 73°C. Protein concentra- tions were determined by BCA assay (Pierce) using BSA as a standard.
Protein Biotinylation Solubilized porcine zona pellucida proteins were purified as
described above. Control proteins, fetuin and ovalbumin, were purchased from Sigma. Zona and control proteins were biotinytated with NHS-Sulfo Biotin (Pierce) according to the manufacturer's instructions. Proteins were dialyzed into 0. t M sodium phosphate buffer, pH 7.4. Twenty micrograms of NHS-Sulfo Biotin was added per 100 /*g protein and the reaction was incubated 2 h on ice. Unconjugated biotin was removed by dialysis into 0.1 M sodium phosphate buffer, pH 7.4. Biotinylated proteins were separated on SDS-PAGE gels, transferred to nitrocellulose membranes, and detected with avidin-atkaline phosphatase and substrates 5-bromo- 4-chloro-3-indotyl phosphate and nitroblue tetrazolium (Vector Laboratories) to verify that all proteins were equally biotinylated (not shown).
Preparation of Sperm Semen samples were collected from mature Yorkshire or Duroc
boars and either used immediately or diluted 1:4 in X-Cell Extender (IMV, Minneapolis, MN) for 24-48 h prior to use. Semen samples were washed through Percoll gradients containing 5.4 ml Percoll, 0.6 ml t0× HBS (1.3 M NaC1, 40 mM KC1, 10 mM CaC12, 5 mM MgCI~), and 4 ml dmTALP {2.1 mM CaC12, 3.1 mM KC1, 1.5 mM MgC12, 100 mM NaC1, 0.29 mM KHaPO4, 0.36% lactic acid, 26 mM NaHCO3, 0.6% BSA, 1 mM pyruvic acid, 20 mM Hepes, pH 7.3, 10 units/ml penicillin, 10 /*g/mt streptomycin) at 800g for t0 min. Sperm were washed with 15 ml dmTALP and pelleted for 5 min at 600g. Washed sperm were resuspended to a concentration of 2 × 10r/ml in dmTALP and capacitated 4 h at 39°C, 5% CO> In some experiments, the calcium ionophore A23187 was added to capaci- tated sperm to a final concentration of 10/~M for 1 h at 39°C, 5% COe to induce the acrosome reaction (Larson and Miller, 1999).
To obtain epididymal sperm, testes and epididymides were removed from anesthetized Duroc boars. Epididymides were dis- sected immediately or stored for 16 h in dmTALP at room tem- perature prior to dissection. Storage did not affect viability, motiL ity, or zona protein binding of recovered epididymal sperm (data not shown). Connective tissue was removed and each epididymis was divided into caput, corpus, and cauda sections as defined by Okamura et al. (1992). Individual sections were placed in petri dishes containing dmTALP and macerated with razor blades to release sperm. The sperm were pelleted 5 rain at 600g, followed by three washes with dmTALP. Washed epididymal sperm were resuspended to a concentration of 2 × 10r/ml and incubated for 4 h under capacitating conditions as described above.
Fluorescent Zona Protein Binding to Fixed Sperm Sperm were allowed to air dry on coverslips and fixed 10 min in
95% ethanol, 5% acetic acid at -20°C. Sperm were blocked for 2 h with 5% normal goat serum in solution A (127 mM NaCt, 5.3 mM KC1, 18.2 mM Hepes, pH 7.2, 0.02% azide). Biotinytated proteins were diluted to 250/*g/ml in solution A and incubated with sperm for 2 h in a moist chamber at room temperature. Coverslips were rinsed three times for 2 min with cold solution A. Proteins were detected with fluorescein-avidin DCS (Vector Laboratories) diluted to 13 >g/ml with solution A for 1 h in a moist chamber. Coverslips were rinsed as described above. In some experiments, sperm were counterstained for 5 rain with 10 ~g/ml rhodamine-labeled Pisum sativum agglntinin (Vector Laboratories) to verify acros0mal status (Melendrez et aL, 1994). Sperm were washed three tim.es with
Copyright © 2000 by Academic Press. All rights of reproduction in any form reserved.

Zona Protein Bindil?g to Sperm 101
solution A, mounted in SlowFade (Molecular Probes, Inc.), and sealed with nail polish. Fluorescence images were acquired under 630× magnification with a Sony DXC 9000 color video CCD camera.
Conjugation of Proteins to Alexa Fluor Dye
Porcine zona pellucida proteins were isolated al~d sotubilized as described above. Transferrin and BSA controls were obtained from. Sigma. After dialysis into PBS, pH 7.4, proteins were labeled with Alexa Fluor 488 (Molecular Probes) according to the manufac~ tnrer's instructions. One milligram of protein was resuspended in 0.5 ml PBS, then 50/~l of 1 M bicarbonate was added to adjust the pH. The protein solution was transferred to a vial of reactive Alexa dye and allowed to stir for 1 h at room tem.perature. The reaction was stopped with the addition of 17 >1 of 1 M hydroxylamine solution for 30 rain. Alexa-conjugated hydroxylamine was removed by extensive dialysis into PBS at 4°C.
Fluorescent Zona Protein Binding to Live Sperm
Boar sperm were washed and capacitated as described above. In some experiments, the ionophore A23t87 was added to induce the acrosome reaction as described above. For saturation experiments, Alexa-zona protein conjugate was added to 0.4 ml sperm to yield final concentrations of 10, 50, or 100 ~g/ml Alexa-zona and 2 × 107 sperm/ml. After approximately 30-min incubations, fluorescence images were acquired under 400?< magnification with a Sony DXC 9000 color video CCD canrcra. Analysis of fluorescence intensity was performed on a Macintosh computer using the NIH Image program (developed at the National Institutes of Health). Integrated densities were measured for three sperm per frame and averaged. Measurements were taken for three flames for each Alexa-zona protein concentration teste&
The 10 ~g/ml Alexa-zona protein concentration was used in subsequent experiments because it yielded an intense signal on the sperm head with minimal background fluorescence. One microliter of 0.5 lng/ml propidium iodide was added to each sample to detect dead or membrane~compromised sperm. Incubations were gener- ally between 15 and 60 rain; however, the fluorescent signal could be observed after as little as 5 min. Live sperm images were captured as described above.
Sperm-Egg Binding Inhibition Assays
For sperm-egg binding assays, porcine oocytes were collected and fixed as described (Rebeiz and Miller, 1999). Oocytes were aspirated from follicles with an 18-gauge needle and centrifuged at 600g for l0 inin. Follicular fluid was removed and individual oocytes were transferred to medium B (127 mM NaC1, 5.3 mM KC1, 18.2 mM Hepes, pH 7.2), Oocytes were vortexed 5 lnin to remove cumulus cells and washed three times in medium B. The washed oocytes were fixed in medium B containing 1.5 % formaldehyde for 10 min. After three additional washes in medium B, oocytes -were transferred to a drop of dmTALP, covered with mineral oil, and stored at 4°C until needed. Before use, oocytes were washed through 3 drops of dmTALP.
Sperm were washed as described above and the concentration w a s adjusted to 2 × 106 sperm/ml with dmTALP. Sperm were incubated under the capacitating conditions described above for 4 h. During the last 30 rain of capacitation, 90 /~1 of sperm was added to 10 /zl of 2 mg/ml protein (unlabeled zona protein,
Alexadabeled zona protein, or Alexa-labeled transferrin) or 10 ~1 of buffer control. Twenty microliters of sperm was added to triplicate 20~p~t dmTALP droplets containing 10 oocytes each. Sperm were allowed to bind oocytes at 39°C for 15 mh~. Oocytes were washed through one drop of dmTALP to remove loosely bound sperm and then transferred to 20-~1 droplets of 4.% paraformaldchyde. Bound sperm were counted using an inverted microscope. Statistical differences between treatments were analyzed with one-way ANOVA followed by the Tukey HSD test.
Induction of the Acrosmne Reaction with Alexa- Zona Proteins
The biological activity of the Alexa~conjugated zona proteins was also tested by the ability to induce the acrosome reaction. Boar sperm were washed and capacitated as described above at a con- centration of 2 × l0 ~ sperm/ml. Ninety microliters of sperm was added to 10 ~1 of buffer control or 2 mg/mt zona protein (unlabeled or Atexa conjugated) and incubated i h at 39°C, 5% CO~. To assess acrosom.at status, spema were fixed in 4% paraformaldehyde and stained with Coomassie G-250 as described (Larson and Miller, 1999). Acrosomal status was determined for 200 sperm per slide. Triplicate slides were analyzed for each treatment, Statistical significance was assessed using one-way ANOVA followed by Tukey's HSD test.
RESULTS
Zona Proteins Localize to the Acrosomal and Postacrosomal Regions of Fixed Boar Sperm
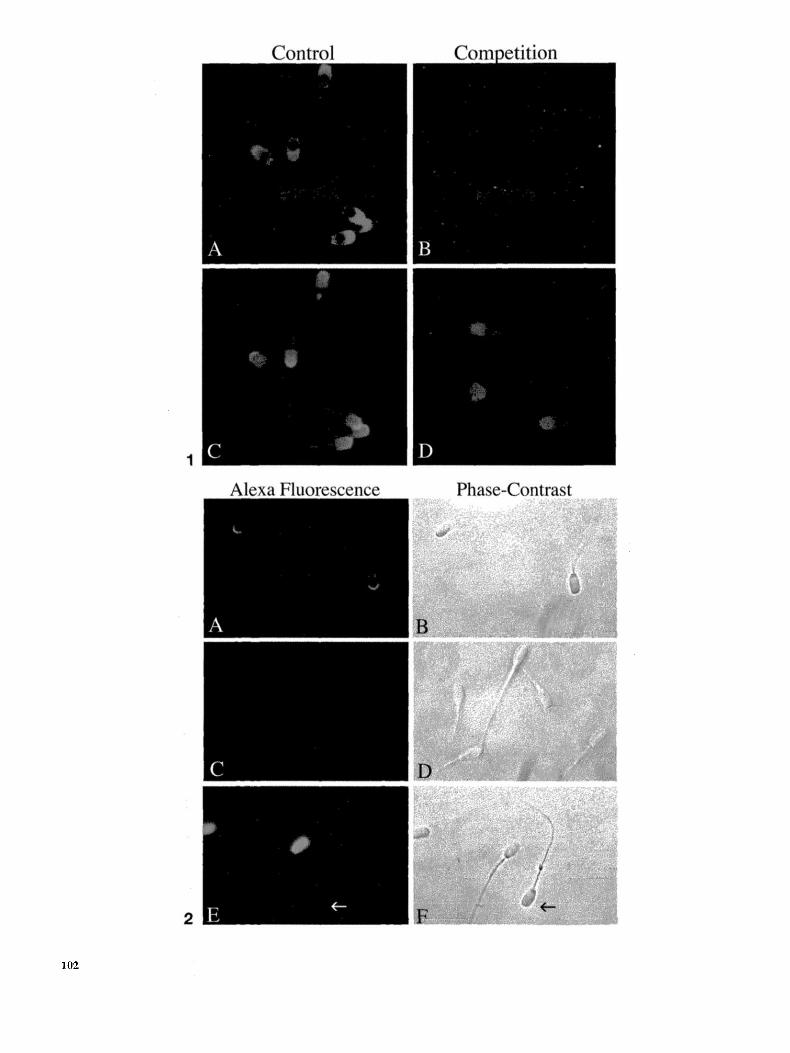
In ini t ia l experiments, fixed boar sperm were incubated with biot inylated zona pellucida or control proteins and fluoresceinated avidin. At concentrat ions of 1, 0.5, and 0.25 mg/ml, solubilized zona peflucida proteins bound to both the acrosomal region and the base of the fixed sperm head (Fig. 1A). To verify acrosomal status, sperm were counter- s tained wi th rhodamine-P, sativum agglutinin, which binds acrosome-intact sperm (Fig. 1C; Melendrez et al., 1994). Similar labeling patterns were observed when solu- bilized zona proteins were incubated with sperm fixed in ethanol or paraformaldehyde (data no t shown). The glyco- proteins fe tuin and ova lbumin were used as controls and did not label sperm. When fixed boar sperm were preincu- bated for 2 h with a 60-fold excess of solubilized zona proteins (15 mg/ml), the zona fluorescence signal was blocked (Fig. 1B). An excess of the control glycoprotein fetuin (15 mg/ml )d id not block zona binding to sperm (data not shown).
Zona Pellucida Binding Is Concentrated at the Acrosomal Ridge of Live, Acrosome-Intact Sperm
Because it was possible that sperm fixation might affect zona protein binding, we repeated the experiments de- scribed above on live sperm. When live sperm were incu- bated wi th biotinylated zona proteins and detected wi th fluoresceinated avidin, only about 10% of live sperm exhib- ited the acrosomal labeling pat tern ini t ia l ly observed in
Copyright © 2000 by Academic Press. All rights of reproduction in any form reserved.
Control Comoetition
Alexa Fluorescence
B
Phase-Contrast
C
102
2

Zone Protein Binding to Sperm 103
fixed sperm (data not shown). Because this method of detection required long incubation t imes and multiple centrifugation steps, many sperm became i lnmotile and the sperm membranes may have become permeable. Another potential disadvantage wi th live sperm and long incuba- tions is that the zona proteins themselves rnight induce changes in the sperm membrane.
To e l iminate these problems, we used zona proteins conjugated directly to the Alexa 488 fluorophore~ The 10 / ,g/ml concent ra t ion of zona pellucida proteins used for this detect ion sys tem was far lower than the 125 /xg/ml required to induce the acrosome react ion in boar sperm (Berger et aL, 1989). Furthermore, even at high zona protein concentra t ions , only approximate ly 15% of boar sperm typical ly undergo the acrosome react ion (Berger et aI., 1989), For these reasons, the Alexa-zona conjugate was unl ikely to induce physiological changes in live sperm under the condi t ions used. Alexa-zona proteins were added to live sperm, along wi th propid ium iodide to detect dead or membrane -compromised sperm. In initial experiments, three populat ions of sperm were observed. The first popula t ion of sperm stained s t rongly wi th propidium iodide and exhibi ted little Alexa-zona label- ing. This popula t ion probably consis ted of dead sperm that had lost their acrosomes. The second popula t ion stained wi th propid ium iodide and bound zona proteins over the acrosomal vesicle, s imilar to the labeling pa t te rn observed in fixed sperm. These sperm m a y have died wi thou t losing their acrosomal contents . Interest ingly, nearly all of the sperm that exhibited the broad acrosomal labeling pat tern originally seen in fixed sperm appeared dead, as they were stained wi th propid ium iodide. A third popula t ion of sperm did not stain wi th propid ium iodide and exhibited concent ra ted Alexa-zona f luorescence over the acrosomal ridge (Fig. 2A). This appears to be the region of live, acrosome-in tac t sperm that init iates zona pellucida binding as all mot i l e sperm belonged to this third populat ion. No differences in Alexa-zona binding were observed be tween capaci tated and uncapaci ta ted boar sperm samples or in samples diluted in extender (data no t shown).
Experiments were performed to show that the Alexa-
conjugated zone proteins displayed the same biological activity as unlabeled zona proteins. First, capacitated sperm were preincubated with unlabeled zona protein, Alexa-zona protein, Alexa-transferrin, or a buffer control. At 0.2 rag/ ml, Alexa-zona protein inhibited sperm binding to the intact zona pellucida to the same extent as unlabeled zona proteins (Fig. 3A). An average of 23.8 + 2.23 control sperm bound to each oocyte. Preincubation of sperm with either unlabeled zona protein or Alexa---zona protein inhibited sperm-egg binding (9.67 + 3.0 bound sperm/oocyte and 2.73 + 1.15 sperm/oocyte bound, respectively), Preincnba- tion with Alexa-transferrin did not inhibit sperm-egg bind- ing (21.7 + 3.42 bound sperm/oocyte}, In the second series of experiments, we determined if Alexaqabeled zona pro- teins could induce the sperm acrosome reaction to the same extent as unlabeled zona protein (Fig. 3B). At a concentra- tion of 0.2 mg/ml, Alexa-labeled zona protein induced the acrosome reaction in the same percentage of sperm as unlabeled zone proteins (8.8 + 0.21 and 9,7 + 1.2%, respectively). Only 5.3 -+ 0.82% of control sperm appeared acrosome reacted. These results are typical of boar sperm, which acrosome react relatively poorly in response to soluble zona protein (Berger et el., 1989). Because the Alexa-zona proteins bound to sperm, inhibited sperm-egg binding, and induced the sperm acrosome reaction, we concluded that zona proteins retain biological activity when conjugated to Alexa.
Several experi ments were performed to show that binding of Alexa-zona to sperm is saturable and specific. First, the signal was blocked with the addition of a 100-fold excess of unlabeled zona proteins (Fig. 2B} and no Alexa signal was detected when two labeled control proteins (transferrin and BSA) were incubated with live sperm (Fig. 2C). Second, we incubated sperm with different concentrat ions of Alexa- zona proteins and quantitated the average intensity of the fluorescent signal per sperm. The fluorescence pattern observed was the same at all concentrat ions tested. Al- though the fluorescence intensi ty between individual sperm was variable, Fig. 4 shows that the average fluores- cence intensi ty reached a plateau at approximately 50 /*g/ml. Although this concentrat ion is consistent with previously determined saturating concentrations of both
FIG. 1. Localization of solubilized zone pellucida proteins on fixed boar sperm. (A) Zona proteins bind to a broad area over the acrosomal vesicle and the base of the fixed sperm head. This image is representative of an experiment performed more than six times with at least three separate zona protein preparations. (B) The signal can be displaced with the addition of a 60-fold excess of unlabeled zona proteins. Sperm acrosomal status was verified by counterstaining with rhodamine-P, sativum agglutinin, which labels fixed acrosome-intact sperm (C and D). FIG. 2. Localization of solubilized zona pellucida proteins on live sperm. Alexa-labeled zone proteins and propidiurn iodide were added to live boar sperm. Alexa-zona proteins bound live, acrosome~intact sperm over the anterior head region concentrated over the acrosomal ridge (A). This labeling pattern was observed in more than six experiments performed with two different preparations of zona proteins. Addition of a 100-fold excess of unlabeled zone proteins displaced the signal (C), The Alexaqabeted control glycoprotein, transferrin, did not bind to live sperm, indicated by the arrow in (E). Dead sperm stained with propidium iodide are visible since fluorescence images were captured using a filter set that allowed detection of both red and green fluorochromes simultaneously (A, C, E). Corresponding phase-contrast images are shown (B, D, F).
Copyright © 2000 by Academic Press, All rights of reproduction in any form reserved.

104 Burk in and Mi]ler
A Sperm-Egg Binding Inhibition
G' 30 © ©
E K
<
2 0 -
/ 0 -
0 i
C o n t r o l Unlabe led A l e x a Zo tm Zona
A l e x a Transferr in
B Sperm Acrosome Reaction
& ~cJ
¢)
o <
g
C o n t r o l Unlabe led Zona
N
Alexa- Zona
FIG. 3. Alexa-coniugated zona proteins retain biological activity. {A) Capacitated sperm were preincubated with 0.2 mg/ml protein or buffer control and then allowed to bind porcine oocytes. The average number of sperm bound per oocyte was determined for triplicate droplets containing 10 oocytes each. Atexa-zona proteins inhibited sperm-egg binding to the same degree as unlabeled zona proteins. Alexa-transferrin did not inhibit sperm binding to the intact zona pellueida. (B) Triplicate capacitated sperm samples were incubated with 0.2 mg/ml protein or buffer control for 1 h and acrosomal status was detemained by Coomassie staining (Larson and Miller, 1999). Atexa .zona protein induced the sperm acrosome reaction to the same extent as unlabeled zona protein. Graphs are representative of experiments performed at least three times. Asterisks indicate samples that are significantly different from the control (P < 0.05).
routine [Thaler and Cardullo, 1996) and porcine (Yurewics et al., 1998) soluble zona proteins, it is slightly lower than the concentrat ion of soluble zona proteins required to induce the boar sperm acrosome reaction (125 mg/ml; Berger et al., 1989). This difference may reflect variations in potency between zona protein preparations or l imitat ions in quanti tat ing fluorescence. Therefore, live boar sperm with uncompromised membranes bound labeled zona pro- teins wi th a saturable and specific pattern that was differ- ent from the pattern observed on fixed or membrane- compromised sperm.
Live, Acrosome-Reacted Sperm Show Increased Zona Binding
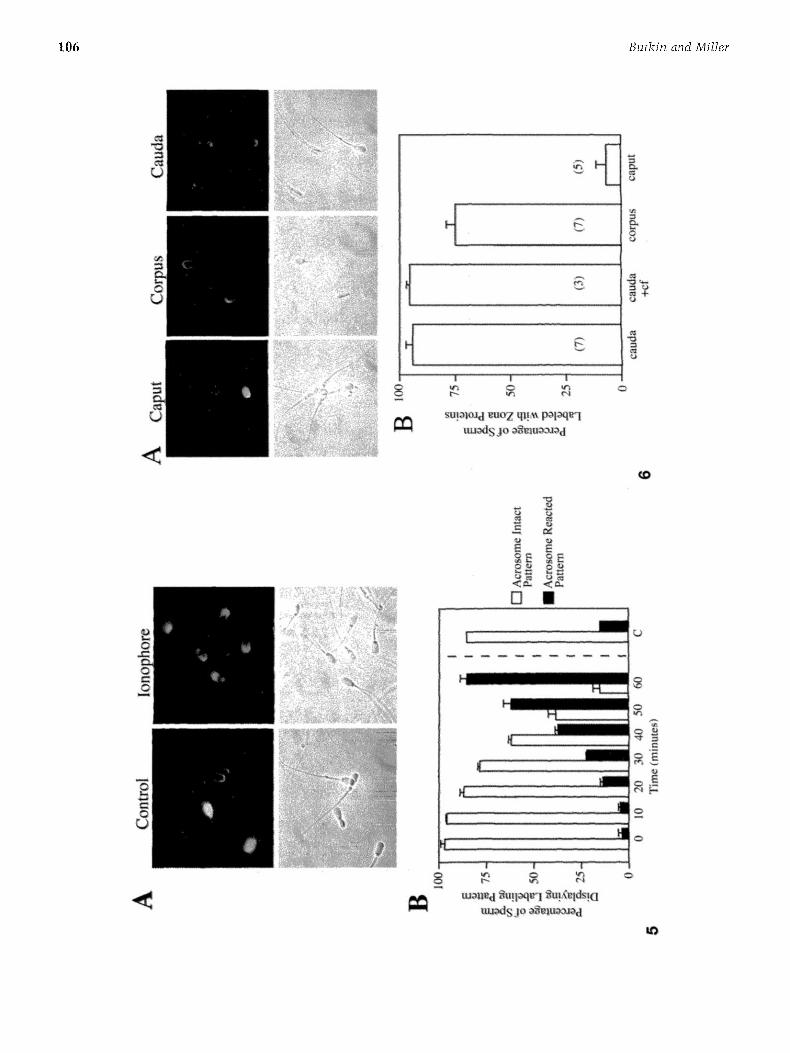
If the sperm acrosome reaction exposes acrosomal pro- teins with roles in secondary zona binding (Yanagimachi, 19941, one might predict tha t soluble zona proteins would label the entire acrosome following calcium ionophore treatment. When sperm were treated with the calcium ionophore A2318 7 to induce the acrosome reaction, most of the live (propidium iodide negative) sperm exhibited strong zona protein labeling extending from the acrosomal ridge to the equatorial region (Fig. 5AI. This labeling pattern appears identical to the signal originally seen over the acrosomal region of ethanol-fixed sperm populations (Fig. 1A). Sperm were removed and examined at various t ime points after the addition of ionophore. The proportion of live sperm with strong acrosomal labeling increased over t ime (Fig. 5B), suggesting that this labeling pattern is characteristic of acrosome-reacted sperm. Rhodamine-P. s a t i v u m agglutinin labeling was used to confirm the percentage of sperm undergoing the acrosome reaction in response to ionophore at each t ime point {data not shown).
Boar Sperm Acquire Zona Binding Ability during Epididymal Maturation
In order to determine the developmental stage at which boar sperm begin to display functional zona pellucida receptors, live sperm from different regions of the epididy- mis were incubated with Alexa-conjugated zona proteins. Acrosome-intact sperm from different regions of the epi- didymis varied in their ability to bind Alexa-zona proteins
25009'
20000 -
15000 -
"~ 10000 -
5000 -
T
I r i f 25 50 75 100
A l e x a - Z o n a Protein Concentrat ion
t25
FIG. 4. Saturability of the Alexa-zona fluorescence signal. Ca- pacitated boar sperm were incubated with 10, 50, or 100 b~g/ml Alexa-zona protein for 30 rain. Sperm fluorescence images were captured and analyzed with NIH hnage software. Measurements of integrated fluorescence density were taken for three sperm per image. The average sperm intensity for three images at each concentration was plotted. The Alexa--zona fluorescence signal reached saturation at approximately 50/xg/ml.
Copyright © 2000 by Academic P re s s Al l rights of reproduct ion in any form rese rved

Zona Prote in Bh?ding to Spem? 105
(Fig. 6). Labeling was detected on 93.8 + 2.9% of live sperm from the cauda epididymis and 74.9 + 3.6% of live sperrn from the corpus. In contrast, only 6.4 + 4~3% of propidium iodide-negative caput epidi@mal spen.n bound zona pro- teins, and the signal was less intense than in the other sperm populations. Cauda epididymal sperm were still able to bind zona proteins after incubation in fluid recovered from. the first caput sperm wash, suggesting that boar caput epididymal fluid does not contain an inhibitor of zona binding. Incubation of sperm from each region of the epididymis under capacitating conditions did not affect zona protein binding (unpublished observations). Interest- ingly, our preliminary observations indicate that ionophore~treated sperm from all regions of the epididylnis exhibit intense Alexa-zona protein binding, suggesting that zona-binding proteins inside the acrosome are functional at an earlier stage in sperm maturation than zona-binding proteins on the sperm surface.
DISCUSSION
By using the sensitive Alexa fluorophore directly conju- gated to solubilized porcine zona proteins, we have ob- served zona receptors on live boar sperm. In acrosome- intact sperm, these receptors are concentrated at the acrosomal ridge. Ionophore~treated sperm bind zona pro- teins with a stronger signal over a considerably larger area of the acrosomal region. Because this intense signal is present on all naembrane-permeabilized boar sperm, zona receptors on the plasma membrane may be overwhelmed by acrosomal zona binding proteins and therefore indistin- guishable on fixed sperm.. Our observations are consistent with electron microscopy images of dn vdtro fertilization captured by Peterson et aL (1981), which show acrosome- intact sperm initiating contact with the egg zona pellucida at the acrosomal ridge. Subsequent to induction of the acrosome reaction, boar sperm appear to contact the zona pellucida over a broad region extending from the acrosomal ridge to the equatorial region (Peterson et al., 1981).
Although it is theoretically possible that the addition of labeled zona proteins to live sperm could cause the acro- some reaction or other changes in the boar sperm mem- brane, the experimental conditions we used make this possibility unlikely. First, the concentration of zona pro- teins used is an order of magnitude lower than the concen- tration required to induce the acrosome reaction in boar sperm (Berger et ai., 1989). Second, the labeling times used were as short as 5 min, which does not approach the time required for the boar sperm, acrosome reaction (Berger et aL, 1989; Fazeli et aI., 1995). Finally, boar sperm generally undergo low rates of soluble zona-induced acrosome reac- tions (<15%) compared to sperm from other species (Berger et aL, 1989).
Early mouse in v i tro fertilization experiments suggested that sperm first bound to the zona at the apical ridge over the acrosolne (Saling et al., 1979). In contrast, direct label-
ing experiments have localized mZP3 binding to the entire mouse sperm, head (Bleil and Wassarman, 1986; Mortillo and Wassarman, 1991). Due to the morphological differ- ences between porcine and murine sperm, caution should be exercised when extrapolating these results to spade- shaped boar sperm. Our results show that zona protein binding is concentrated over the boar sperm acrosomal ridge.
In dom.estic species such as the sheep (Dziuk and Dick- man, 1965) and pig (P. J. Dziuk, personal communication), the concave side of the sperm head preferentially attaches to the zona pellucida during fertilization in viw). In our experiments, we could not detect increased zona binding on one side of the sperm, suggesting that the reason for this preferential at tachment is not due to the location of zona receptors on the sperm plasma membrane.
In the boar, radiolabeled ssZPB and ssZPC (also called ZP3c~ and ]3) together have been generally localized to the anterior head region of capacitated, acrosome-intact boar sperm after fixation (Yurewicz et aI., 1993). Other research- ers observed no zona protein binding to acrosome-intact boar sperm, but did observe binding over the acrosomal region of cold-shock-permeabilized sperm (Jones et aL, 1991; Yonezawa et al., 1995). These apparently conflicting data may be explained by our observations that the Alexa- zona signal observed in acrosome-reaeted or membrane- compromised sperm is far more intense than"the signal detected on the surface of live spern~. Fazeli et al. (1997) found that acrosome~intact boar sperm initiate binding to the intact zona pellucida in vitro, suggesting that the signal we observe on live, acrosome-intact sperm is likely to detect a receptor(s) involved in prim.ary sperm binding. The strong signal which becomes apparent only when sperm undergo the acrosome reaction or other forms of membrane permeabilization may detect secondary zona receptors in- side the acrosome. Because the signal on membrane- damaged sperm is much stronger, it is likely that some previous detection methods were not sensitive enough to reveal the primary sperm receptor labeling pattern, tt is unclear why acrosome-reacted sperm cannot initiate bind- ing to the intact zona pellucida (Fazeli et al., 1997), since they appear capable of binding soluble zona proteins.
We observed binding of solubilized zona proteins over the postacrosomal region of most fixed sperm examined regar& less of acrosomal status, fixation method, or detection method. This labeling pattern is consistent with observa- tions by Tsubamoto et al. (1996), in which antibodies to a recombinant form of ssZPA were used to detect binding to fixed, acrosome-reacted sperm from a number of species, including the boar. The ability of ssZPA to bind acrosome- intact boar sperm was not determined by these investiga- tors. Since we did not observe this pattern on any live sperm, our data suggest that zona binding to the postacro- somal region of the sperm head is induced by fixation and may not be physiologically relevant in the boar.
The developmental stage at which sperm begin to display zona pellucida receptors has also been the subject of debate.
Copyright © 2000 by Academic Press. All rights of reproduction in any form reserved,
106 Burkin and Miller
I
T Q
D |
¢D
tt~

Z o n a Protein Birlding to Sperm 107
Since the ability of sperm to bind to the zona pellucida is an essential step in successful fertilization and boar sperm acquire the ability to fertilize oocytes during epididymal maturat ion (Holtz and Smidt, 1976; Hunter et cd., 1976), we were interested in determining whether boar epididymat sperm could bind zona proteins. Solubilized porcine zona proteins bound to most sperm from the canda and corpus regions of the epididymis. In contrast, few caput sperm bound solubilized zona proteins, and these exhibited a far less intense signal compared to sperm from other epididy- real regions. This observation is consistent with in vitro binding assay data collected by Peterson et aL (1984), in which boar caput sperm did not bind to the zona pellucida. Interestingly, mouse caput epididymal sperm are able to bind to the intact zona pellucida after extensive washing to remove epididymal fluids (McLaughlin and Shut, 1987) and can undergo the acrosome reaction in response to solubi- lized zona proteins (Biegler et al., 1994). In these previous zona binding experiments only sperm that bound to eggs were assessed (Peterson et al,, 1984; McLaughlin and Shut, 1987), but our procedure allowed sampling of all sperm in the population. In addition, there may be variation among species in the changes sperm undergo during epididymal maturation. Since we observed little binding of solubilized zona proteins to caput boar sperm even after extensive washing to remove epididymal fluid, our results support the hypothesis of Peterson et al. (1984) that changes related to zona binding ability occur during epididymal transit in the boar. One possible mechanism for these changes might be the "unmasking" of zona receptors already present on the spemr surface during passage through the epididymis. This unmasking might occur through proteolysis or other e n z y matic activity. A second possibility is that receptors se- creted into the epididymal fluid are added to sperm during epididymal maturat ion (Lakoski et al., 1988).
After ejaculation, mature sperm undergo capacitation in the female reproductive tract. The molecular aspects of capacitation are not clear, but capacitated sperm, exhibit
increased moti l i ty and membrane changes (reviewed in Suarez, 1996; Visconti m~d Kopf, 1998}. Al though capacita- tion nray be required for zona pellucida binding in some species (Gwatkin and Williams, t978), uncapacitated boar sperm bind to intact zona _iz? vitro (Peterson et aL, 1984). Harkema et al. (1998} used flow cytonretry to measure fluorescein-conjugated zona proteins bound to boar sperm. They observed a large increase in the number of sperm with strong zona fluorescence after ionophore t reatment and a smaller increase after a 3-h incubation in capacitation medium~ Although we used 4oh incubation times to allow capacitation, we did not detect differences in Alexa-zona binding between capacitated and uncapacitated boar sperm. Our results support the hypothesis of Peterson et al. (1984) that boar sperm have no capacitation requirements for primary zona binding.
Many studies have indicated that sperm-zona pellucida interactions occur preferentially between gametes from the same species. Russel et at. (1979) showed that bull sperm would bind zona-intact pig oocytes, but at a far lower frequency than boar sperm. Under our experinrental condi- tions, solubilized Atexa-labeled pig zona proteins did not bind either capacitated or uncapacitated live bovine sperm (unpublished observations). In contrast, solubilized zona proteins did bind to fixed or ionophore-treated bovine sperm in a pattern similar to that seen in porcine sperm (unpublished observations). These data suggest that the species specificity observed at the level of the zona pellu- cida is due to divergence of primary and not secondary sperm receptors and are consistent wi th the lack of species- specific binding of zona proteins by proacrosin (Hardy and Garbers, 1994).
Candidate zona receptors have been localized to fixed boar sperm and most display a strong acrosomal labeling pattern (Mori et al., 1995; Ensslin et al., 1998). This pattern is similar to the pattern observed when labeled zona pellu- cida proteins were incubated with fixed, ionophoreotreated, or cold-shock-permeabilized sperm. /3-1,4-Gatactosyl-
FIG. 5. Binding of zona pellucida proteins is increased in acrosome~reacting sperm. Live, capacitated boar sperm were incubated with the calcium ionophore A23187 to induce the acrosome reaction, followed by the addition of Alexa-zona proteins and propidium iodide. The top row in (A) shows fluorescence images of Alexa-zona and propidium iodide, the bottom corresponding phase-contrast images. Acrosome-reacted sperm showed an increased area of zona protein binding, extending from the acrosomal ridge to the equatorial region of live spcrm, whereas controls without ionophore did not. For time-course experiments (B), samples were removed and at least 200 sperm counted at 10-rain intervals after the addition of ionophore. The percentage of live sperm displaying zona binding in a thin baud over the acrosomal ridge decreased with time (white bars), while the percentage displaying stroug acrosomal fluorescence increased [black bars). Controls to which ionophore was not added ("C') were counted after 60 min. The graph shows the average percentages of sperm ± standard error for duplicate samples and is representative of three experiments. Some error bars are too small to be seen. FIG. 6. Boar sperm acquire the ability to bind zoua pellucida proteins during epididymal maturation. Sperm from the caput, corpus, and cauda regions of the boar epididymis were capacitated and incubated with Alexa-zoua proteins and propidium iodide. IA) Most caput sperm do not bind zona proteins, and those that do display a very faint signal. In contrast, most corpus and cauda sperm exhibit intense zona protein binding. (B) Quantitation of live, acrosome-intact sperm displaying zona fluorescence. Approximately 93% of cauda sperm and 75% of corpus sperm bound zona proteins. In contrast, only about 6.4% of caput sperm bound zona proteins after extensive washing. Ninety-five percent of cauda sperm incubated for 4 h in caput fluid (+cf 1 retained the ability to bind zona proteins. Bars represent averages ± standard error. Sample nmnbers are indicated in parentheses and all except cauda sperm incubated in caput fluid represent sperm from three epididymides.
Copyright © 2000 by Academic Press. All rights of reproduction in any form reserved.

108 Burk in and Mi l ler
transferase, a receptor for mouse ZP3, has been local ized to the acrosomal ridge of live, ac rosome- in tac t boar sperm (Larson and Miller , 1997), but is not required for spe rm-egg binding in the pig (Miller and Rebeiz, 1999). Another candidate, zonadhesin, appears to bind porcine but not bovine zona {Hardy and Gathers, 1995). However , the local- i za t ion of zonadhesin on boar sperm has not been reported. Since the physiological roles of m a n y of these receptors are unclear , i t r emains to be de te rmined whe the r these candi- dates have roles in p r imary or secondary sperm binding. sp56 is a zona-binding prote in tha t was original ly isola ted from mouse sperm and thought to be located on the sperm surface (Bleil and Wassarman, 1990; Cheng et aI., 1994; Bookbinder et al., 1995). Subsequent e lect ron microscopy exper iments have shown that sp56 and its guinea pig homologue AM67 are localized w i t h i n the acrosome (Foster et al., 1997). Consider ing the differences in zona binding pa t te rns tha t we observed be tween fixed and live sperm, i t is impor t an t to de te rmine whe ther candidate zona receptors can be def ini t ively local ized to the p lasma membrane of l ive boar sperm.
Al though the basic steps of fer t i l iza t ion are wel l con- served (Yanagimachi, 1994), compar ison of the pig and mouse models reveals differences at the level of p r imary zona pe l luc ida binding, inc luding the deve lopmenta l stage at w h i c h funct ional p r imary zona receptors appear on the sperm surface. Since the abi l i ty of sperm to bind to the zona pel luc ida has been correlated wi th fer t i l i ty in some species (Yogev et aI., 1996; Zhang et at., 1998), a bet ter unders tand- ing of the molecular in teract ions be tween sperm receptors and the zona pel luc ida wi l l be impor t an t in the develop- m e n t of infer t i l i ty tests and design of contraceptives.
ACKNOWLEDGMENTS
We thank Russ Wischover at the University of Illinois Moorman Swine Farm for providing boar semen samples and assisting with surgeries, Dr. Charles Graves and Jeffery Escobar for help with surgeries, and Dr. Stephen Kaufman and Dr. Dean Burkin for assistance with image capture. This work was supported by the Illinois Council for Food and Agricultural Research, the University of Illinois Agricultural Experiment Station as part of Hatch Project ILLU-35-0335, the United States Department of Agriculture NRI (99~35203), and the University of Illinois Research Board. H.R.B. was supported by United States Public Health Service Fellowship 5 T32 HD07028.
REFERENCES
Adham, I. M., Nayernia, K., and Engel, W. (1997), Spermatozoa lacking acrosin protein show delayed fertilization. MoL Repro& Dev. 46, 370-376.
Baba, T., Azmna, S., Kashiwabara, S., and Toyoda, Y. (1994). Sperm from mice carrying a targeted mutation of the acrosin gent can penetrate the oocyte zona pcllucida and effect fertilization. L Biol. Chem. 269, 31845--,31849.
Berger, T., Turner, K. O., Meizel, S., and Hedrick, J. L. (1989). Zona pellucida-induced acrosome reaction in boar sperm. Biol. Re- prod. 40, 525-530.
Biegler, B. E., Aarons, D. J., George, B. C., and Poirier, G. R. (1994). Induction of physiological acrosome reactions in caput epididy~ real spermatozoa of mice. ]. Reprod. Fertil. 100, 219-224.
Bleil, J. D., and Wassarman, P. M. (1986). Autoradiographic visual- ization of the mouse egg's sperm receptor bound to sperm. J. Cell Biol. 102, 1363-1371.
Bleil, J. D., and Wassarlnan, P. M. (1990). Identification of a ZP3-binding protein on acrosome-intact mouse sperm by pho- toaffinity crosslinking. Proc. Nat. Acad. Sci. USA 87, 5563-5567.
Bookbinder, L. H., Cheng, A., and Bleil, J. D. (1995). Tissue- and species-specific expression of sp56, a mouse sperm fertilization protein. Science 269, 86-89.
Cheng, A, Le, T., Palacios, M., Bookbinder, L. H., Wassarman, P. M., Suzuki, F., and Bleil, J. D. (1994). Sperm-egg recognition in the mouse: Characterization of sp56, a sperm protein having specific affinity for ZP3. J. Cell Biol. 125, 867-878.
Dunbar, B. S., Wardrip, N. J., and Hedrick, J. L. (t980). Isolation, physiochemical properties, and macrom.olecular composition of zona pellucida from porcine oocytes. Biochemistry 19, 356-365.
Dziuk, P. J., and Dickman, Z. (1965). Sperm penetration through the zona pe]lucida of the sheep egg. J~ Exp. ZooL 158, 237-240.
Ensslin, M., Vogel, T., Calvete, J. J., Thole, H. H., Sehmidtke, J., Matsuda, T., and Topfer~Petersen, E. (1998). Molecular cloning and characterization of P47, a novel boar sperm-associated zona- pellucida-binding protein homologous to a family of mammalian secretory proteins. Biol. Repro& 58, 1057-1064.
Fazeli, A., Hage, W. J., Cheng, F., Voorhout, W. F., Marl;s, A., Berets, M. M., and Colenbrander, B. (1997). Acrosome-intact boar sperm initiate bindil~g to the homologous zona pellucida in vitro. Biol. Reprod. 56, 430-438,
Fazeli, A. R., Holt~ C., Steenweg, W., Bcvers, M. M., Holt, W. V., and Colenbrander, B. (1995). Development of a sperm hem.izona binding assay for boar semen. Theriogenology 44, 17-27.
Foster, J. A., Friday, B. B., Maulit, M. T., Blobel/C., Winffey, V. P., Olson, G. E., Kim, K. S., and Gerton, G. L. (1997}. AM67, a secretory component of the guinea pig sperm acrosomal matrix, is related to mouse sperm protein sp56 and the complement component 4-binding proteins. J. Biol Chem. 272, 12714-12722
Geng, J., Raub, T. J., Baker, C. A., Sawada, G. A, Ma, L., and Elhammer, A. P. (1997). Expression of a P-selectin tigand in zona pellucida of porcine oocytes and P-selectin on acrosomal mem- brane of porcine sperm cells. Potential implications for their involvement in sperm--egg interactions. [. Ceil Biol. 137, 743- 754.
Gwatkin, R. B. L., and Williams, D. T. (1978). Bovine and hamster zona solutions exhibit receptor activity for capacitated but not for uncapacitated sperm. Gamete Res. 1, 259-263.
Hardy, D. M., and Garbers, D. L. (1994). Species-specific binding of sperm proteins to the extracellular matrix (zona pellucida) of the egg. J. Biol. Chem. 269, 19000-19004.
Hardy, D. M., and Garbers, D. L. (1995}. A sperm membrane protein that binds in a species-specific manner to the egg extracellular matrix is homologous to yon Willebrand factor. J. Biol. Chem. 270, 26025-26028~
Harkema, W., Harrison, R. A. P, Miller, N. G. A., Topper, E. K, and Woelders, H. (1998). Enhanced binding of zona pellucida proteins to the acrosomal region of intact boar spermatozoa in response to fertilizing conditions: A flow cytometric study. Biol. Repro& 58, 421-430.
Copyright © 2000 by Academic Press. All rights of reprod,ction in any form reserved.

Zone Protein Binding to Sperm 109
Harris, J. D., Hibler, D. W., Fontenot, G. K., Hsu, K. T., and Yurewicz, E. C. (1994). Cloning and characterization of zona pellucida genes and cDNAs from a variety of mammalian spe- cies: The ZPA, ZPB, and ZPC gene families. DNA Seq. 4, 361--393.
Hedrick, J. L. (1996). Comparative structural and antigenic proper- ties of zona peltucida glycoproteins./. Reprod. Fertil. (Suppl.) 50, 9-17.
Holtz, W., and Smidt, D. (1976). The fertilizing capacity of epididy- real spermatozoa in the pig. 1. Reprod. Fertil. 46, 227--..229.
Hunter, R. H. F., Holtz, W., and Henfrcy, P. J. (1976). Epididylnal function in relation to the fertilizing ability of sperlnatozoa. J. Repro& Fertil. 46, 463-466.
Jones, R. (t991). Interaction of zona pellucida glycoproteins, sul- phated carbohydrates and synthetic polymers with proacrosin, the putative egg~binding protein from mammalian spermatozoa. Development 111, 1155-1163.
Lakoski, K. A., Carton, C. P., Cabot, C. L., and Soling, P. M. (1988). Epididymal maturation and the acrosome reaction in mouse sperm: Response to zona pellucida develops coincident with modification of M42 antigen. Biol. Reprod. 38, 221~233
Larson, J. L., and Miller, D. J. (1997)~ Sperm from a variety of mammalian spedes express/31,4-galactosyltransferase on their surface. BioI. Repmd. 57~ 442-445.
Larson, J. L., and Miller, D. J. (1999). Simple histochelnical stain for acrosomes on sperm from several species. Mol. Reprod. Dev. 52, 445-449.
Luj Q., and Shut, B. D. (1997). Sperm from /3-1,4 galactosyl- transferase-null mice are refractory to ZP3-induced acrosome reactions and penetrate the zona pellucida poorly. Development 124, 4121-4131.
Macek, M. B., Lopez, L. C, and Shut, B. D. (1991). Aggregation of fl-l,4-gatactosyltransferase on mouse sperm induces the acro~ some reaction. Dev. Biol. 147, 440-444.
McLaughlin, J. D., and Shut, B. D. (1987). Binding of caput epididy~ real mouse sperm to the zone petlucida. Dev. Biol. 124, 557-56t.
Melendrez, C., Meizel, S., and Berger, T. (1994). Comparison of the ability of progesterone and heat solubilized porcine zona pellu~ cida to initiate the porcine sperm acrosome reaction in vitro. Mol. Repro& Dev. 39, 433-438.
Miller, D. J., Macek, M. B., and Shur, B. D. (1992). Complementa~ rity between sperm surface j3-1,4-galactosyltransferase and egg- coat ZP3 mediates sperm-egg binding. Nature 357, 589-593.
Mori, E., Kashiwabara, S., Baba, T., lnagaki, Y., and Mori, T. (1995). Amino acid sequences of porcine Sp38 and proacrosin required for binding to the zona pellucida. Dev. Biol. 168, 575-583.
Mortillo, S., and Wassarman, P. M. (19911. Differential binding of golddabeled zona pellucida giycoproteins mZP2 and mZP3 to mouse sperm membrane components. Development 113, I41- 149.
Okamura, N., Dacheaux, F., Venien, A., Onoe, S., Huet, J. C., and Dacheaux, J. L. (1992). Localization of a maturation-dependent epididymal sperm surface antigen recognized by a monoclonal antibody raised against a 135-kilodalton protein in porcine epi- didymal fluid. Biol. Reprod. 47, 1040-1052.
Peterson, R. N., Russel, L. D., Bundman, D., Conway, M, and Freund, M, [1981). The interaction of living boar sperm and sperm plasma membrane vesicles with the porcine zona pellu- cida. Dev. Biol. 84, 144-156.
Peterson, R. N., Russel, L. D., Bundman, D., and Freund, M. (1980). Sperm-egg interactions: Evidence for boar sperm plasma mem~ brane receptors for porcine zona peltucida. Science 207, 73~-74.
Peterson, R. N., Russell, L. D., and Hunt, W. P. (1984). Evidence for specific binding of uncapacitated boar spermatozoa to porcine zonae pettucidae in vitro~ ]. Eytz Zool. 231, 137-147.
Rebeiz, M., aM Miller, D. J~ (19991. Porcine sperm sm'face /31,4galactosyltransferase binds to the zona pellucida but is not necessary or sufficient to mediate spermozona pellucida binding. MoL Reprod. Dev. 54, 379-387.
Russel, L., BuMman, D., and Freund, M. (1979). Sperm--egg inter~ actions: Evidence for boar sperm plasma membrane receptors for porcine zona pel]ucida. Science 207, 73-74.
Sating, P. M., Sowinski, J., and Storey, B. T. (I979). An ultrastruc- tnral study of epididymat mouse spermatozoa binding to zonae pellucidae in vitro: Sequential relationship to the acrosome reaction. J. Exp. Zool. 21)9, 229-238.
Suarez, S.S. (1996). Hyperactivated motility in sperm./. Al~ctrol. 17, 33t---335.
Thaler, C. D., and Cardullo, R. A. (1996). The initial molecular interaction between mouse sperm and the zona peltucida is a complex binding event./. Biol. Chem. 271, 23289-23297.
Tsubamoto, H., Hasagawa, A., Inoue, N., Yamasaki, N., and Koyama, K. (1996). Binding of recombinant pig zona peltucida 1 [ZP1) to acrosome-reacted spermatozoa. 7. Rel)rod. Fe~til. Suppl. 50, 63-67.
Visconti, P. E., and Kopf, G. S. (1998). Regulation of protein phosphorytation during sperm capacitation. Biol. Reprod. 59, 1-6.
Wassarman, P. M. (t988). Zona pellucida glycoproteins. Annn. Rev. Biochem. 57, 4t5-442.
Yanagimachi, R. (1994). Malnmalian fertilization. In "The Physi- ology of Reproduction" (E. Knobil and J. D. Neill, Eds.J, 2nd ed. Raven Press, New York.
Yogev, L., Gamzu, R., Hauser, R., Botchan, A., Alnit, A., Lessing, J. B., Paz, G., and Yavetz, H. {1996). Correlation between sperm penetration into the human zona pellucida and in vitro fertiliza- tion rates. Andrologia 29, 71-75.
Yonezawa, N., Hatanaka, Y., Takeyam.a, H., and Nakano, M. [1995). Binding of pig sperm receptor in the zone pellucida to the boar sperm acrosome. J. Repro& Fertil. 103, 1-8.
Yurewicz, E. C., Pack, B. A., Arlnant, D. R., and Sacco, A. G. (1993). Porcine zona pellucida ZP3c~ glycoprotein mediates binding of the biotinqabeled Mr 55,000 family (ZP3) to boar sperm mem- brane vesicles. MoI. Repro& Dev. 36, 382-389.
Yurewicz, E. C., Sacco, A. G., Gupta, S. K., Xu, N., and Gage, D. A. (t998). Hetero-otigomerization-dependent binding of pig oocyte zone pellucida glycoproteins ZPB and ZPC to boar sperm mem- brane vesicles. J. Biol. Chem. 273, 7488-7494.
Zhang, B. R., Larsson, B., Lundeheim, N., and Rodriguez-Martinez, H. (1998). Sperm characteristics and zona pellucida binding in relation to field fertility of frozen-thawed semen from dairy AI bulls. Int. I. Amtrol. 21, 207-216.
Received for publication September 27, 1999 Revised March 14, 2000
Accepted March 17, 2000
Copyright © 2000 by Academic Press. All rights of reproduction m any form reserved.
Copyright © 2022 FDOKUMEN



















