
Identification of PARP homologues in human ejaculated sperm
9
Determination of Poly (ADP-ribose) polymerase (PARP) homologues in human ejaculated sperm and its correlation with sperm maturation Rajesh Jha, Ph.D., a Ashok Agarwal, Ph.D., HCLD., a Reda Mahfouz, M.D., a Uwe Paasch, M.D., b Sonja Grunewald, M.D., b Edmund Sabanegh, M.D., a Satya P. Yadav, Ph.D., c and Rakesh Sharma, Ph.D. a a Reproductive Research Center, Glickman Urological and Kidney Institute, and c Molecular Biotechnology Laboratory, Lerner Research Institute, Cleveland Clinic, Cleveland, Ohio; and b Department of Dermatology/Andrology Unit, University of Leipzig, Leipzig, Germany Objectives: To investigate the presence of Poly (ADP-ribose) polymerase (PARP) and evaluate its function in ejaculated spermatozoa. Design: Experimental study. Setting: Reproductive Research Center. Patient(s): Semen specimens from 18 healthy donors and 12 infertile males. Intervention(s): Ejaculated spermatozoa were subjected to sperm fractionation with a double-layer density gradi- ent, protein extraction, detection, immunoblotting, and mass spectrometry. Semen samples were exposed to PARP inducer staurosporine, or hydrogen peroxide with or without PARP inhibitor 3-aminobenzimide. Annexin V assay was examined for apoptosis by flow cytometry. Result(s): We detected 75 kDa, 63 kDa, and 60 kDa PARP homologues on the immunoblot of mature and immature sperm fraction isolated from human ejaculate, which were identified as PARP-1 (75 kDa), PARP-9 (63 kDa), and PARP-2 (60 kDa), respectively. Western blot analysis showed a positive correlation between the amount of PARP protein and sperm maturity. PARP proteins (75 kDa, 63 kDa, and 60 kDa) were evaluated after inducing apoptosis by hydrogen peroxide and staurosporine exposure. Conclusion(s): PARP-2 may have a role in prevention of oxygen species/oxidative stress and chemical (stauro- sporine)-induced sperm cell apoptosis. The presence of the PARP homologue, that is, PARP-1 (75 kDa), suggests its role in preventing damage of mature sperm. Additional studies are needed to delineate the role of PARP-9 in sperm physiology. The results from our study indicate an active role for PARPs in sperm cell physiology in pre- venting apoptosis. (Fertil Steril Ò 2009;91:782–90. Ó2009 by American Society for Reproductive Medicine.) Key Words: Sperm, DNA damage, PARP homologues, apoptosis, sperm maturity The maintenance of genomic integrity is very important to prevent cancer and other genetic diseases (1). Eukaryotic organisms protect DNA lesions by numerous mechanisms. During moderate DNA damage, cell cycle check point pro- teins such as p53 are activated, and assist in DNA repair mechanism through a roentgenogram crosscomplementing factor and other single strand breaks repair/base excision repair factors (DNA polymerase b and DNA ligase III) (2, 3). However, high doses of DNA damage result in a passive and unorganized mode of cell death known as necrosis (4, 5). Mono or Poly (ADP-ribosyl)ation is an immediate DNA damage-dependent posttranslational modification of histones and other nuclear proteins. This contributes to the survival of injured proliferating cells [reviewed in (6)]. This modifica- tion of Poly (ADP-ribosyl)ation is accomplished by Poly (ADP-ribose) polymerases (PARP) that help in identifying region of DNA breaks for recruitment of the DNA base exci- sion repair mechanism. It is also essential for the mainte- nance of genomic integrity and survival in response to genotoxic insults (7–10). During excessive DNA damage and in the presence of sufficient energy supply this enzyme is cleaved into 24-kDa and 84-kDa domains preventing DNA repair, thereby marking apoptosis (11–13). If a damaged cell lacks energy, PARP is cleaved into 60-kDa fragments along with ongoing necrosis. Poly (ADP-ribose) polymerases now constitute a large family of 18 proteins [reviewed in (6)]. Poly (ADP-ribose) metabolism is critical in a wide range of biologic structures and processes, including DNA repair and maintenance of genomic stability, Received October 26, 2007; revised and accepted December 31, 2007. R.J. has nothing to disclose. A.A. has nothing to disclose. R.M. has noth- ing to disclose. U.P. has nothing to disclose. S.G has nothing to dis- close. E.S. has nothing to disclose. S.P.Y. has nothing to disclose. R.S. has nothing to disclose. Presented at the 63rd Annual meeting of the American Society of Repro- ductive Medicine, Washington, DC, October 13–17, 2007. Supported by a Research Grant from the Glickman Urological and Kidney Institute and Research Programs Committee, Cleveland Clinic, Cleve- land, OH. Reprint requests: Ashok Agarwal, Ph.D., HCLD, Glickman Urological & Kidney Institute, and Departments of Obstetrics/Gynecology, Cleveland Clinic, 9500 Euclid Avenue, Desk A19.1, Cleveland, Ohio 44195 (FAX: 216-636-3118/216-445-6049; E-mail: [email protected]; Web site: http://www.clevelandclinic.org/reproductiveresearchcenter). Fertility and Sterility â Vol. 91, No. 3, March 2009 0015-0282/09/$36.00 Copyright ª2009 American Society for Reproductive Medicine, Published by Elsevier Inc. doi:10.1016/j.fertnstert.2007.12.079 782
-
Upload
anmalaiuniversity -
Category
Documents
-
view
2 -
download
0
Transcript of Identification of PARP homologues in human ejaculated sperm

7
Determination of Poly (ADP-ribose) polymerase(PARP) homologues in human ejaculated spermand its correlation with sperm maturationRajesh Jha, Ph.D.,a Ashok Agarwal, Ph.D., HCLD.,a Reda Mahfouz, M.D.,a Uwe Paasch, M.D.,b
Sonja Grunewald, M.D.,b Edmund Sabanegh, M.D.,a Satya P. Yadav, Ph.D.,c and RakeshSharma, Ph.D.a
a Reproductive Research Center, Glickman Urological and Kidney Institute, and c Molecular Biotechnology Laboratory, Lerner
Research Institute, Cleveland Clinic, Cleveland, Ohio; and b Department of Dermatology/Andrology Unit, University of Leipzig,
Leipzig, Germany
Objectives: To investigate the presence of Poly (ADP-ribose) polymerase (PARP) and evaluate its function inejaculated spermatozoa.Design: Experimental study.Setting: Reproductive Research Center.Patient(s): Semen specimens from 18 healthy donors and 12 infertile males.Intervention(s): Ejaculated spermatozoa were subjected to sperm fractionation with a double-layer density gradi-ent, protein extraction, detection, immunoblotting, and mass spectrometry. Semen samples were exposed to PARPinducer staurosporine, or hydrogen peroxide with or without PARP inhibitor 3-aminobenzimide. Annexin V assaywas examined for apoptosis by flow cytometry.Result(s): We detected �75 kDa, �63 kDa, and �60 kDa PARP homologues on the immunoblot of mature andimmature sperm fraction isolated from human ejaculate, which were identified as PARP-1 (�75 kDa), PARP-9(�63 kDa), and PARP-2 (�60 kDa), respectively. Western blot analysis showed a positive correlation betweenthe amount of PARP protein and sperm maturity. PARP proteins (75 kDa, 63 kDa, and 60 kDa) were evaluated afterinducing apoptosis by hydrogen peroxide and staurosporine exposure.Conclusion(s): PARP-2 may have a role in prevention of oxygen species/oxidative stress and chemical (stauro-sporine)-induced sperm cell apoptosis. The presence of the PARP homologue, that is, PARP-1 (�75 kDa), suggestsits role in preventing damage of mature sperm. Additional studies are needed to delineate the role of PARP-9 insperm physiology. The results from our study indicate an active role for PARPs in sperm cell physiology in pre-venting apoptosis. (Fertil Steril� 2009;91:782–90. �2009 by American Society for Reproductive Medicine.)
Key Words: Sperm, DNA damage, PARP homologues, apoptosis, sperm maturity
Fertility and Sterility� Vol. 91, No. 3, March 2009Copyright ª2009 American Society for Reproductive Medicine,
82
a passive and unorganized mode of cell death known asnecrosis (4, 5).
Mono or Poly (ADP-ribosyl)ation is an immediate DNAdamage-dependent posttranslational modification of histonesand other nuclear proteins. This contributes to the survival ofinjured proliferating cells [reviewed in (6)]. This modifica-tion of Poly (ADP-ribosyl)ation is accomplished by Poly(ADP-ribose) polymerases (PARP) that help in identifyingregion of DNA breaks for recruitment of the DNA base exci-sion repair mechanism. It is also essential for the mainte-nance of genomic integrity and survival in response togenotoxic insults (7–10). During excessive DNA damageand in the presence of sufficient energy supply this enzymeis cleaved into 24-kDa and 84-kDa domains preventingDNA repair, thereby marking apoptosis (11–13). If a damagedcell lacks energy, PARP is cleaved into �60-kDa fragmentsalong with ongoing necrosis.
Poly (ADP-ribose) polymerases now constitute a large familyof 18 proteins [reviewed in (6)]. Poly (ADP-ribose) metabolismis critical in a wide range of biologic structures and processes,including DNA repair and maintenance of genomic stability,
The maintenance of genomic integrity is very important toprevent cancer and other genetic diseases (1). Eukaryoticorganisms protect DNA lesions by numerous mechanisms.During moderate DNA damage, cell cycle check point pro-teins such as p53 are activated, and assist in DNA repairmechanism through a roentgenogram crosscomplementingfactor and other single strand breaks repair/base excisionrepair factors (DNA polymerase b and DNA ligase III)(2, 3). However, high doses of DNA damage result in
Received October 26, 2007; revised and accepted December 31, 2007.
R.J. has nothing to disclose. A.A. has nothing to disclose. R.M. has noth-
ing to disclose. U.P. has nothing to disclose. S.G has nothing to dis-
close. E.S. has nothing to disclose. S.P.Y. has nothing to disclose.
R.S. has nothing to disclose.
Presented at the 63rd Annual meeting of the American Society of Repro-
ductive Medicine, Washington, DC, October 13–17, 2007.
Supported by a Research Grant from the Glickman Urological and Kidney
Institute and Research Programs Committee, Cleveland Clinic, Cleve-
land, OH.
Reprint requests: Ashok Agarwal, Ph.D., HCLD, Glickman Urological &
Kidney Institute, and Departments of Obstetrics/Gynecology, Cleveland
Clinic, 9500 Euclid Avenue, Desk A19.1, Cleveland, Ohio 44195 (FAX:
216-636-3118/216-445-6049; E-mail: [email protected]; Web site:
http://www.clevelandclinic.org/reproductiveresearchcenter).
0015-0282/09/$36.00Published by Elsevier Inc. doi:10.1016/j.fertnstert.2007.12.079

transcriptional regulation, centromere function and mitotic spin-dle formation, centrosomal function, structure and function ofvault particles, telomere dynamics, trafficking of endosomal ves-icles, apoptosis, and necrosis [reviewed in (6, 14)].
Sperm DNA damage plays an important role in the patho-genesis of male infertility (15–17). Failure of fertilization inassisted reproduction technology, despite the use of morpho-logically normal and motile sperm, may be because of suchdamage (15–17). Apoptosis or programmed cell death in-volves a series of cellular, morphologic, and biochemicalalterations, leading to cell suicide without eliciting an inflam-matory response. The presence of a key apoptotic protein,Fas, on mature spermatozoa (18) has been previously de-tected, but it does not seem to be functional (19, 20). Otherinvestigators have shown the presence and externalizationof phosphatidylserine residues (21) and caspase activation(22). PARP cleavage has been reported as an apoptosis ornecrosis marker in other cell types depending on the typeof cleavage (23). This molecule has also been reported inthe testicular germ line maintenance and its development(24–26). However, its presence on ejaculated sperm is un-clear. Although two reports showed its presence (27, 28) an-other documents its absence (29). In male infertility andpathophysiology, apoptotic sperm represent poor sperm qual-ity, and this may be attributed to altered functions of normalapoptosis or oxidative stress (18, 26, 27, 29).
The objectives of our study were to investigate the pres-ence of PARP protein and its homologues in ejaculatedhuman sperm and to determine its relation to sperm maturity.We also assessed PARP protein levels through Western blot-ting after PARP modulation/sperm cell apoptosis induction.
MATERIALS AND METHODS
Sperm Sample Preparation
Following the approval of the institutional review board, se-men specimens from 18 healthy donors and 12 infertile maleswere collected for this study. Healthy donors were defined asa group of healthy male volunteers with sperm parameters inthe normal range (normozoospermic) according to the WorldHealth Organization guidelines (30). These included menwith both proven and unproven fertility. Infertile maleswere defined as men who were attending the male infertilityclinic with a history of infertility of at least 1 year.
Sperm count and motility was done before and after dou-ble-density centrifugation by a double-density gradientmethod (80% and 40% gradient; SAGE In vitro FertilizationInc., San Clemente, CA) (31, 32). The interphase between the40% to 80% gradient comprising the immature sperm wascarefully aspirated into a tube, washed, and resuspended insperm wash medium. This fraction consisted largely of im-mature, morphologically abnormal sperm with poor motility.Similarly, the pellet at the bottom of the 80% gradient wascarefully aspirated into another tube, washed, and resus-pended in sperm wash medium. This fraction consisted ofmorphologically normal, highly motile mature sperm.
Fertility and Sterility�
Both mature sperm fraction and immature sperm fractionwere washed with Quinn’s sperm wash media (SAGE In vitroFertilization Inc.), and final washing was done with phos-phate-bufferd saline (PBS; pH 7.4, 10 mM). The experimentaldesign is shown in Figure 1.
Sperm Protein Extraction
Sperm proteins were extracted according to the method ofShukla et al. (33). Briefly, postwashed sperm cells pelletswere solubilized in sperm solublization buffer comprisingof 187mM Tris HCl, pH 6.8, 2% SDS, 10% glycerol, 1% Tri-ton X-100, 1 mM PMSF, 1 mM EGTA, and 0.01% Aprotinin.Sperm suspension was homogenized using a mechanicalhomogenizer (Fisher Scientific, Hanover Park, IL) usingthree strokes at 10-second intervals. The homogenate wascentrifuged at 12, 000 � g for 20 minutes, and the resultantsupernatant was used as a sperm protein extract. All theseprocedures were performed at 4�C.
PARP Immunoblotting
Protein concentrations were estimated by BCA Protein AssayKit (Pierce Biotechnology, Inc., Rockford, IL). Proteins weredenatured with Laemmli buffer (Bio-Rad Laboratories, Her-culus CA) (34) and subsequently heated for 5 minutes at95�C. Equal concentration of protein (50 mg) was loaded
FIGURE 1
Schematic representation of the experimental studydesign.
Jha. PARP homologues in human spermatozoa. Fertil Steril 2009.
783

784 Jha et al. PARP homologues in human spermatozoa
done with 5% bovine serum albumin, followed by incubationwith 1:300 (vol./vol.) dilution of the anti-PARP-1 (goat anti-PARP-1, Santa Cruz; cat no. sc-1561). After washing, cover-slips with sperm cells adsorbed were incubated with 1:300(vol. by vol.) dilution of rabbit antigoat IgG antibody conju-gated with fluorescein isothiocyanate (Sigma; Cat no. F2016) for 1 hour at room temperature. Phosphate-buffered sa-line with Tween 20 (10 mM, pH 7.4) was used throughout theprocedure. Following each incubation, washing was repeatedthrice with PBS-Tat 5-minute intervals. The sperm cells werecounter stained in DAPI (4’, 6-diamidino-2-phenylindole)(Vector mounting media with DAPI, Vector Laboratories,Burlingame, CA). The cells were imaged on an upright fluo-rescence microscope with an exposure time of 1 second (Le-ica Microsystems Inc, Bannockburn, IL). Negative controlswere prepared by excluding the incubation of the cells inanti-PARP-1. These experiments were repeated five times,and representative images were obtained.
PARP Modulation Assay/PARP Induction by Apoptosis
Semen samples were collected from eight donors for thisstudy. Both mature and immature sperm fractions were sepa-rated using the double-density gradient centrifugationmethod as described above. PARP activation/apoptosis wasinduced by an oxidative stress inducer hydrogen peroxide(H2O2; 30 mM) and apoptosis by staurosporine (2.5 mM).PARP activity was inhibited by inhibitor 3-amino-benzamide(0.3 mM). Each mature and immature sperm fraction was re-suspended in HTF media and divided into five aliquots/group,that is, control/untreated; H2O2 treated; H2O2þ 3-aminoben-zamide; staurosporine alone; and staurosporine þ 3-amino-benzamide. All aliquots were incubated at 37�C for 1 hour.After incubation, mature and immature sperm aliquots wereprocessed for gel electrophoresis and Western blotting asmentioned above. PARP protein bands on immunoblot wereanalyzed by Tooltab TL100 (Nonlinear Dynamics).
Sperm Apoptosis Analysis by Annexin V Assay
Annexin V- FITC (BD Biosciences, San Jose, CA; Cat. No.51-65874X) was used for assessment of sperm apoptosis.Sperm were washed with cold PBS and then resuspended in1� annexin V binding buffer (10 mM HEPES/NaOH(pH7.4), 140 mM NaCl, 2.5 mM CaCl2). To get the concen-tration 1.0 � 106 sperm/mL, 2.5 mL Annexin þ 2.5 mL Pro-pidium iodide were added. All test and control tubes weregently mixed and incubated for 15 minutes at 25�C in thedark then 400 mL of PBS was added into each tube. After in-cubation samples were analyzed by flow cytometry (FACS-can, BD, San Jose, CA) at a low flow rate using Cellquestsoftware for about 10,000, and FACs data files were analyzedusing FlowJo software version 6.4.2 at the flow core facilityof the Lerner Research Institute, Cleveland Clinic.
Statistical Analysis
Results are presented as the mean � standard error as well asmedian and interquartile ranges for sperm count and motility.
onto each well and separated on a 12% SDS-PAGE for the pro-tein profile verification. SDS-PAGE resolved proteins werevisualized with 0.1% Coomassie (Brilliant Blue R-250;Sigma St. Louis, MO) stained in 10% acetic acid and 40%methanol. Excess Coomassie stain was removed by additionalwash with 10% acetic acid and 40% methanol. Proteins wereresolved on SDS-PAGE then electroblotted onto nitrocellu-lose membrane (Bio-Rad) in the presence of 25 mM Trisbase, 192 mM glycine, and 20%v/v methanol (34, 35). Theblotted membranes were incubated with 5% nonfat milk fol-lowed by anti-PARP-1 (Santa Cruz, Santa Cruz, CA; cat no.sc-1561) and rabbit antigoat horseradish peroxidase (HRP)conjugated (sc-2768) (1:1,000) dilutions.
Incubation with primary antibody and blocker (5% nonfatmilk) was done for 2 hours, whereas incubation with secondaryantibody was for 1 hour. Phosphate-buffered saline (10 mM,pH 7.4 containing 0.1% Tween-20; PBS-T) was used through-out the procedure. Protein bands were visualized by diamino-benzidene tetrahydrochloride reaction (Vector Laboratories,Burlingame, CA). Secondary antibody specificity was verifiedby incubating with secondary antibody only. PARP immuno-positive bands were quantified by Totallan 100 software (Non-linear Dynamics Ltd, Newcastle upon Tyne, UK).
PARP Protein Identification by Peptide MassFingerprinting
Protein extract from mature sperm from rwo donors and oneinfertile patient was used for this experiment. Proteins wereresolved on 12% SDS-PAGE; three lanes were used for im-munoblotting and other three lanes were stained with coomas-sie (Brillant blue R-250). The three Western blotted laneswere probed with PARP-1 antibody to detect the PARP-1-im-munopositive band. The corresponding PARP-1-immunopos-itive bands in the SDS-PAGE were excised and digested byTrypsin using Trypsin In-Gel Digest Kit (Cat. No. PP 0100,Sigma). The reaction was performed for 12 hours, followingwhich the tryptic peptides were detected by MALDI-Tof-Tof(Applied Biosystem, Foster City, CA) at the Cleveland ClinicLerner Research Institute core facility. Bioinformatics analy-ses were performed through ExPASy Proteomics Server (Pro-tein Prospector; MS-Fit) to identify the corresponding proteinbased on the peptide mass fingerprinting.
Immunolocalization of PARP on the Ejaculated HumanSperm
Immunolocalization of PARP was done on donors and infer-tile mature and immature sperm fractions according to themethod of Shukla et al. (33). Briefly, spermatozoa weresmeared onto clean glass coverslips precoated with 0.05%poly-L-lysine. These smears were allowed to dry at roomtemperature. The coverslips were treated with 10% formalinfor 10 minutes to fix the sperm, followed by neutralization in0.5 M ammonium chloride for another 10 minutes. Spermwere permeabilized by 0.25% Triton X-100 for 20 minutes,followed by chilled ethanol for 10 seconds. Blocking was
Vol. 91, No. 3, March 2009

FIGURE 2
Detection of PARP proteins 75 kDa, 63 kDa, and 60 kDa, employing Western blot and their densitometric analysisfor evaluating the protein level in donors and infertile men between (A) immunoblot of mature and immaturesperm fraction; (B) 75 kDa band intensity in mature and immature sperm fraction; (C) 60 and 63 kDa bandintensity in mature and immature sperm fraction; and (D) validation of primary antibody specificity byimmunoblotting with secondary antibody alone.
Jha. PARP homologues in human spermatozoa. Fertil Steril 2009.
Fertility and Sterility� 785

TABLE 1Analysis of the bands by peptide mass fingerprinting identified these as PARP-1, PARP-9, and PARP-2cleaved fragment, respectively.
Observedbands on theimmunoblot(kDa)
Querymatch/percentage Species
Name of the protein(PARP homolog)
GeneAccession # MW (Da)
75 31/40 (77%) human Poly [ADP-ribose]polymerase 1 (PARP-1)(ADPRT) (NAD(þ) ADP-ribosyl transferase 1)
P09874 112,954
63 24/43 (55%) human Poly [ADP-ribose]polymerase 9 (PARP-9)
Q8IXQ6 96,344
60 13/23 (56%) human Tankyrase-2 (TANK2)(TRF1-interacting ankyrin-related ADP-ribosepolymerase 2) (PARP-2)
Q9H2K2 126,919
Jha. PARP homologues in human spermatozoa. Fertil Steril 2009.
A Student’s t test was used for comparison of two data sets.Significance was defined as P<.05. This statistical analysiswas performed using Microsoft Excel software.
RESULTS
The median (25th and 75th percentile) for sperm parametersin unprocessed liquefied seminal ejaculates used in these ex-periments were: donors, concentration (�106/mL) 49 (37.2,97.7); percent motility, 59.3 (47.8, 70.6); and range of roundcell count was 0.01–2 � 106/mL. For patients, concentration(�106/mL) 28.1 (17.9, 52.9); percent motility: 57.2 (51.2,62.8); and range of round cell count was 1.0–5.0 � 106/mL.
Postwash sperm parameter of mature sperm fractions were:donors, concentration (�106/mL) 56.0 (39.7, 99.2); percent mo-tility: 75.0 (66.0, 84.0); patients, concentration (�106/mL): 34.2(17.0, 58.7); and percent motility: 61.0 (45.7, 72.1). Immaturesperm fraction parameters were: donors, sperm concentration(�106/mL) 27.0 (19.9, 39.7); and percent motility, 33.3 (24.2,43.9); patients, sperm concentration (�106/mL) 14.3 (7.4,29.7) and percent motility, 43.1 (28.7, 49.7).
Detection of PARP Protein on the Human Ejaculated Sperm
Anti-PARP-1 detected �75-kDa,�63-kDa, and �60-kDa pro-tein bands on the immunoblot in mature and immature sperm(Fig. 2A). In mature sperm of all investigated samples, the�75-kDa (P<.017) as well as the �63-kDa and �60-kDa(P<.04) PARP homologues exhibited elevated protein levelsin donors when compared with infertile males (Fig. 2B–C). Incontrast, investigation of the immature sperm fraction did notshow any significant differences in the 75-kDa, 63-kDa, and60-kDa protein levels in donors and infertile males (Fig. 2B–C).Primary antibody specificity was validated by immunoblottingof the secondary antibody alone (negative control) (Fig. 2D).
786 Jha et al. PARP homologues in human spermatozoa
Sperm PARP-1 Homologue Identification
The above-mentioned PARP-1 immunopositive bands (75kDa, 63 kDa, and 60 kDa) were subjected to peptide mass fin-ger printing and mass spectroscopy for their identification.Peptide mass finger printing analysis identified these to bePARP-1, PARP-9, and PARP-2, respectively (see Table 1).
PARP Immunolocalization on the Sperm
Immunolocalization of PARP on mature sperm from donorsshowed intense localization on the head region, particularlynear the acrosomal region (Fig. 3A). In the immature spermfraction it was abnormally distributed and present as a distinctband at the equatorial region of the sperm head (Fig. 3B). Thepresence of PARP on the mature and immature sperm frac-tion of infertile patients exhibited a similar distribution asseen in mature and immature sperm fraction from donors,but the intensity of fluorescence was relatively weak (Fig.3C and D). Specificity of PARP-1 was confirmed by labelingwith secondary antibody alone (Fig. 3E).
PARP Modulation Assay
PARP-1 antibody showed �75-kDa, 63-kDa, and 63-kDaPARP-immunopositive bands on the blot of donors and infer-tile mature as well as in immature sperm fractions (Fig. 4Aand B). In mature sperm, incubation with H2O2 alone resultedin a diminished 75-kDa PARP homologue, which showed fur-ther depletion when 3-aminobenzamide was added (P<.045)(Fig. 4A and C). The 75-kDa PARP of the immature spermfraction was not affected by H2O2 at all (Fig. 4B), but the60-kDa PARP homologue (PARP-2) was not detectable afterincubation with H2O2 in both the absence and the presence of3-aminobenzamide in mature and immature sperm (Fig. 4A,B, and E).
Vol. 91, No. 3, March 2009

FIGURE 3
Immunolocalization study of PARP-1 on (A) mature and (B) immature donor sperm, and (C) mature and (D)immature sperm from infertile patients. Crossreactivity was tested by incubating with secondary antibody alone(E).
Jha. PARP homologues in human spermatozoa. Fertil Steril 2009.
In contrast, incubation with staurosporine induced nochanges in the 75-kDa PARP homologue in mature spermbut led to a reduction of this homologue in immature sperm(Fig. 4C), particularly when 3-aminobenzamide/PARP inhib-itor was added (P<.001). In mature sperm, the 60-kDa PARPhomologue band intensity was slightly reduced by treatmentwith staurosporine, whereas 3-aminobenzamide þ stauro-sporine depleted further the levels of the 60-kDa protein(P<.004) (Fig. 4E). The immature sperm fraction showeda complete loss of the 60-kDa PARP homologue after incuba-tion with staurosporine and hydrogen peroxide (both withand without 3-aminobenzamide; Fig. 4E). Interestingly, the�63-kDa PARP homologue exhibited no significant changesin all the groups studied (Fig. 4D).
Fertility and Sterility�
Sperm Cell Apoptosis Analysis by Annexin V Assay
Treatment with H2O2 significantly elevated the portion ofannexin V positive sperm (P<.003) regardless of the combi-nation with 3-aminobenzamide in mature and immaturesperm. Treatment with staurosporine did not significantlyalter the amount of annexin V positive sperm, but additionof 3-aminobenzamide significantly increased annexin Vstaining (P<.0005; Fig. 4F). This was true for both matureand immature sperm.
DISCUSSION
PARP has been involved in various sperm cell functions suchas gamete differentiation (during spermiogenesis) (36),
787
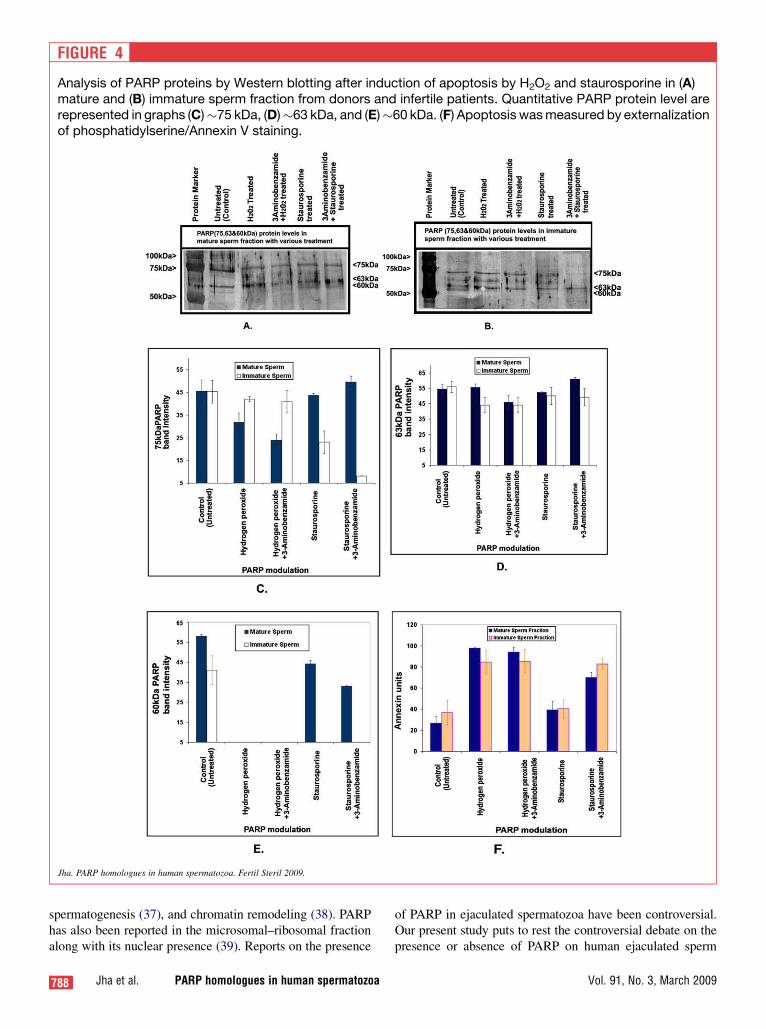
FIGURE 4
Analysis of PARP proteins by Western blotting after induction of apoptosis by H2O2 and staurosporine in (A)mature and (B) immature sperm fraction from donors and infertile patients. Quantitative PARP protein level arerepresented in graphs (C)�75 kDa, (D)�63 kDa, and (E)�60 kDa. (F) Apoptosis was measured by externalizationof phosphatidylserine/Annexin V staining.
Jha. PARP homologues in human spermatozoa. Fertil Steril 2009.
spermatogenesis (37), and chromatin remodeling (38). PARPhas also been reported in the microsomal–ribosomal fractionalong with its nuclear presence (39). Reports on the presence
788 Jha et al. PARP homologues in human spermatozoa
of PARP in ejaculated spermatozoa have been controversial.Our present study puts to rest the controversial debate on thepresence or absence of PARP on human ejaculated sperm
Vol. 91, No. 3, March 2009

(27–29). In this study we have shown the presence of PARP-1(�75 kDa) and its homologues PARP-9 (�63 kDa) and PARP-2 (�60 kDa) in both mature and immature sperm. PARP-1 wasnot detected as expected at 112 kDa; instead, the presence of 75kDa, 63 kDa, and 60 kDa raised the question of its presence. Wetherefore investigated and confirmed PARP-1 immunopositiveproteins by peptide mass fingerprinting analysis. These wereconfirmed as PARP-1, PARP-2, and PARP-9.
The presence of low PARP levels in mature sperm of infer-tile patients compared with the mature sperm of donors sug-gests a role of PARP and its homologues in male infertility.This also corresponded to the weaker fluorescence intensityseen in sperm from subfertile males. Furthermore, low levelsof PARP-1, -2, and -9 seen in the immature sperm fractions ofboth donors and infertile males compared with the maturefractions indicates a potential role of these homologues duringsperm maturation.
PARP homologues have a variety of cellular functions;among them, DNA damage detection and repair are mostimportant (6, 36, 40). Particularly, PARP-1 and PARP-2 areinvolved in DNA repair mechanisms. DNA fragmentationis higher in infertility patients as well as in immature sperm(6, 36, 40). It can be speculated that in such sperm the re-duced levels of PARP homologues might be responsiblefor the inefficiency to protect/repair posttesticular spermlesions.
PARP was found to be present on the anterior portion of thesperm head covering the acrosomal region in the mature frac-tion of donors and comparatively lower concentration of themature fraction of infertile patients. Their localization de-scribed abnormal distribution on the immature sperm fractionof both the fertile and infertile patient. PARP-1 proteins ex-hibited restricted to the central head region in normal spermphysiology, but expresses elsewhere than this position. Thepresence of high PARP-2 protein levels in the donors showsits involvement in genomic DNA integrity. Its involvementin the DNA repair mechanism against genotoxic agents hasalso been demonstrated (10, 41), and abnormalities in thePARP-2 content may affect sperm maturity.
We next examined the effect of oxidative stress and apo-ptosis by exposing sperm fractions to an external oxidativestress inducer, H2O2, and an apoptosis inducer, staurosporine.We did not observe any change in the 63-kDa band, indicat-ing that PARP-9 may not be significantly affected by oxida-tive stress and sperm apoptosis. In contrast, we observeda moderate reduction in 75-kDa PARP-1 in mature spermin the presence of oxidative stress and after induction of ap-optosis in the immature sperm, and this was exaggerated byaddition of the PARP-inhibitor. The 60-kDa PARP-2 wasnot detectable under oxidative stress in both the mature andimmature sperm. In the immature fraction, PARP-2 was notdetectable following induction of apoptosis by staurosporine.The strong modulation of PARP-2 suggests its involvement inresponse to oxidative stress and especially to pro-apoptoticstimuli in immature sperm.
Fertility and Sterility�
We modulated PARPs function by inducing sperm apopto-sis via H2O2 and staurosporine in the physiology of isolatedmature and immotile sperm fraction of the ejaculated sperm.Evaluation of these protein functions explains the role ofPARP-2 and PARP-1 in providing a protective mechanismagainst oxidative stress in normal fertile individuals. ThisPARP-dependent mechanism is probably inefficient in infer-tile men; hence, the sperm are more susceptible to damage.Earlier reports documented apoptosis via oxidative stress(42) and chemically induced (43) through caspase-dependentand -independent pathways. Our results showed preventionof apoptosis in the presence of the PARP homologue,whereas PARP inhibition by 3-aminobenzamide increasedapoptosis by twofold. Moreover, PARP-2 levels were alsoinfluenced by apoptotic-inducing agents like hydrogen per-oxide (36). Evaluation of the externalization of phosphatidyl-serine measured by annexin-V staining showed a significantinduction of apoptosis by H2O2. These oxidative stress-induced membrane changes were not preventable by PARP.However, staurosporine-induced cell apoptosis can be pro-tected by PARP proteins as the PARP inhibitor 3-aminoben-zamide escalates phosphatidylserine externalization.
In conclusion, we have demonstrated that human ejacu-lated sperm contain PARP homologues, PARP-1 (�75kDa), PARP-2 (�60 kDa), and PARP-9 (�63 kDa). LowerPARP levels negatively affect sperm maturity, and conse-quently, may affect male fertility. PARP-1 and PARP-2may have a role in conferring protection to the ejaculatedsperm, particularly following exposure to oxidative stressand apoptosis inductors.
Acknowledgments: We acknowledge the support of our Clinical Andrology
laboratory staff in enrolling the donors and patients for the study.
REFERENCES1. Hanahan D, Weinberg RA. The hallmarks of cancer. Cell 2000;100:
57–70.
2. Dasika GK, Lin SC, Zhao S, Sung P, Tomkinson A, Lee EY. DNA dam-
age-induced cell cycle checkpoints and DNA strand break repair in
development and tumorigenesis. Oncogene 1999;18:7883–99.
3. Griffiths SD, Clarke AR, Healy LE, Ross G, Ford AM, Hooper ML, et al.
Absence of p53 permits propagation of mutant cells following genotoxic
damage. Oncogene 1997;14:523–31.
4. Bonfoco E, Krainc D, Ankarcrona M, Nicotera P, Lipton SA. Apoptosis
and necrosis: two distinct events induced, respectively, by mild and in-
tense insults with N-methyl-D-aspartate or nitric oxide/superoxide in
cortical cell cultures. Proc Natl Acad Sci USA 1995;92:7162–6.
5. Majno G, Joris I. Apoptosis, oncosis, and necrosis. An overview of cell
death. Am J Pathol 1995;146:3–15.
6. Ame JC, Spenlehauer C, de Murcia G. The PARP superfamily. Bioessays
2004;26:882–93.
7. Trucco C, Oliver FJ, de Murcia G, Menissier-de Murcia J. DNA repair
defect in poly(ADP-ribose) polymerase-deficient cell lines. Nucleic
Acids Res 1998;26:2644–9.
8. d’Adda di Fagagna F, Hande MP, Tong WM, Lansdorp PM, Wang ZQ,
Jackson SP. Functions of poly(ADP-ribose) polymerase in controlling
telomere length and chromosomal stability. Nat Genet 1999;23:76–80.
9. Vodenicharov MD, Sallmann FR, Satoh MS, Poirier GG. Base excision
repair is efficient in cells lacking poly(ADP-ribose) polymerase 1.
Nucleic Acids Res 2000;28:3887–96.
789

790 Jha et al. PARP homologues in human spermatozoa
27. Blanc-Layrac G, Bringuier AF, Guillot R, Feldmann G. Morphological
and biochemical analysis of cell death in human ejaculated spermatozoa.
Cell Mol Biol (Noisy-le-grand) 2000;46:187–97.
28. Wundrich K, Paasch U, Leicht M, Glander HJ. Activation of caspases in
human spermatozoa during cryopreservation—an immunoblot study.
Cell Tissue Banking 2006;7:81–90.
29. Taylor SL, Weng SL, Fox P, Duran EH, Morshedi MS, Oehninger S, et al.
Somatic cell apoptosis markers and pathways in human ejaculated
sperm: potential utility as indicators of sperm quality. Mol Hum Reprod
2004;10:825–34.
30. World Health organization. WHO laboratory manual for the examination
of human semen and sperm-cervical mucus interaction. 4th ed. Cam-
bridge: Cambridge University Press, 1999.
31. Allamaneni SS, Agarwal A, Rama S, Ranganathan P, Sharma RK. Com-
parative study on density gradients and swim-up preparation techniques
utilizing neat and cryopreserved spermatozoa. Asian J Androl 2005;7:
86–92.
32. Moustafa MH, Sharma RK, Thornton J, Mascha E, Abdel-Hafez MA,
Thomas AJ Jr, et al. Relationship between ROS production, apoptosis
and DNA denaturation in spermatozoa from patients examined for infer-
tility. Hum Reprod 2004;19:129–38.
33. Shukla S, Jha RK, Laloraya M, Kumar PG. Identification of non-
mitochondrial NADPH oxidase and the spatio-temporal organization
of its components in mouse spermatozoa. Biochem Biophys Res
Commun 2005;331:476–83.
34. Laemmli UK. Cleavage of structural proteins during the assembly of the
head of bacteriophage T4. Nature 1970;227:680–5.
35. Towbin H, Staehelin T, Gordon J. Electrophoretic transfer of proteins
from polyacrylamide gels to nitrocellulose sheets: procedure and some
applications. 1979. Biotechnology 1992;24:145–9.
36. Dantzer F, Mark M, Quenet D, Scherthan H, Huber A, Liebe B, et al. Pol-
y(ADP-ribose) polymerase-2 contributes to the fidelity of male meiosis
I and spermiogenesis. Proc Natl Acad Sci USA 2006;103:14854–9.
37. Maymon BB, Cohen-Armon M, Yavetz H, Yogev L, Lifschitz-Mercer B,
Kleiman SE, et al. Role of poly(ADP-ribosyl)ation during human sper-
matogenesis. Fertil Steril 2006;86:1402–7.
38. Meyer-Ficca ML, Scherthan H, Burkle A, Meyer RG. Poly(ADP-ribosy-
l)ation during chromatin remodeling steps in rat spermiogenesis. Chro-
mosoma 2005;114:67–74.
39. Concha II, Figueroa J, Concha MI, Ueda K, Burzio LO. Intracellular dis-
tribution of poly(ADP-ribose) synthetase in rat spermatogenic cells. Exp
Cell Res 1989;180:353–66.
40. Schreiber V, Dantzer F, Ame JC, de Murcia G. Poly(ADP-ribose): novel
functions for an old molecule. Nat Rev 2006;7:517–28.
41. Dantzer F, Giraud-Panis MJ, Jaco I, Ame JC, Schultz I, Blasco M, et al.
Functional interaction between poly(ADP-Ribose) polymerase 2 (PARP-
2) and TRF2: PARP activity negatively regulates TRF2. Mol Cell Biol
2004;24:1595–607.
42. Liu Z, Lin H, Ye S, Liu QY, Meng Z, Zhang CM, et al. Remarkably high
activities of testicular cytochrome c in destroying reactive oxygen spe-
cies and in triggering apoptosis. Proc Natl Acad Sci USA 2006;103:
8965–70.
43. Zhang XD, Gillespie SK, Hersey P. Staurosporine induces apoptosis of
melanoma by both caspase-dependent and -independent apoptotic path-
ways. Mol Cancer Ther 2004;3:187–97.
10. de Murcia JM, Niedergang C, Trucco C, Ricoul M, Dutrillaux B,
Mark M, et al. Requirement of poly(ADP-ribose) polymerase in recovery
from DNA damage in mice and in cells. Proc Natl Acad Sci USA
1997;94:7303–7.
11. Kaufmann SH, Desnoyers S, Ottaviano Y, Davidson NE, Poirier GG.
Specific proteolytic cleavage of poly(ADP-ribose) polymerase: an early
marker of chemotherapy-induced apoptosis. Cancer Res 1993;53:
3976–85.
12. Yung TM, Satoh MS. Functional competition between poly(ADP-ribose)
polymerase and its 24-kDa apoptotic fragment in DNA repair and tran-
scription. J Biol Chem 2001;276:11279–86.
13. D’Amours D, Sallmann FR, Dixit VM, Poirier GG. Gain-of-function of
poly(ADP-ribose) polymerase-1 upon cleavage by apoptotic proteases:
implications for apoptosis. J Cell Sci 2001;114:3771–8.
14. Burkle Poly(ADP-ribose) A. The most elaborate metabolite of NADþ.
FEBS J 2005;272:4576–89.
15. Benchaib M, Braun V, Lornage J, Hadj S, Salle B, Lejeune H, et al.
Sperm DNA fragmentation decreases the pregnancy rate in an assisted
reproductive technique. Hum Reprod 2003;18:1023–8.
16. Benchaib M, Lornage J, Mazoyer C, Lejeune H, Salle B, Francois
Guerin J. Sperm deoxyribonucleic acid fragmentation as a prognostic
indicator of assisted reproductive technology outcome. Fertil Steril
2007;87:93–100.
17. Borini A, Tarozzi N, Bizzaro D, Bonu MA, Fava L, Flamigni C, et al.
Sperm DNA fragmentation: paternal effect on early post-implantation
embryo development in ART. Hum Reprod 2006;21:2876–81.
18. Sakkas D, Mariethoz E, St. John JC. Abnormal sperm parameters in hu-
mans are indicative of an abortive apoptotic mechanism linked to the
Fas-mediated pathway. Exp Cell Res 1999;251:350–5.
19. Paasch U, Grunewald S, Said TM, Agarwal A, Sharma RK, Glander HJ.
Apoptotis signal transduction of human spermatozoa in response to
physiological and pathological stimuli. Biotechnologietagung 2004;3.
20. Paasch U, Agarwal A, Gupta AK, Sharma RK, Grunewald S,
Thomas AJ Jr, et al. Apoptosis signal transduction and the maturity status
of human spermatozoa. Ann N Y Acad Sci 2003;1010:486–8.
21. Barroso G, Morshedi M, Oehninger S. Analysis of DNA fragmentation,
plasma membrane translocation of phosphatidylserine and oxidative
stress in human spermatozoa. Hum Reprod 2000;15:1338–44.
22. Paasch U, Grunewald S, Fitzl G, Glander HJ. Deterioration of plasma
membrane is associated with activated caspases in human spermatozoa.
J Androl 2003;24:246–52.
23. Oberhammer F, Wilson JW, Dive C, Morris ID, Hickman JA,
Wakeling AE, et al. Apoptotic death in epithelial cells: cleavage of
DNA to 300 and/or 50 kb fragments prior to or in the absence of internu-
cleosomal fragmentation. EMBO J 1993;12:3679–84.
24. Alcivar AA, Hake LE, Hecht NB. DNA polymerase-beta and poly(AD-
P)ribose polymerase mRNAs are differentially expressed during the
development of male germinal cells. Biol Reprod 1992;46:201–7.
25. Di Meglio S, Tramontano F, Cimmino G, Jones R, Quesada P. Dual role
for poly(ADP-ribose)polymerase-1 and -2 and poly(ADP-ribose)glyco-
hydrolase as DNA-repair and pro-apoptotic factors in rat germinal cells
exposed to nitric oxide donors. Biochim Biophys Acta 2004;1692:35–44.
26. Sinha Hikim AP, Lue Y, Diaz-Romero M, Yen PH, Wang C,
Swerdloff RS. Deciphering the pathways of germ cell apoptosis in the
testis. J Steroid Biochem Mol Biol 2003;85:175–82.
Vol. 91, No. 3, March 2009




















