
Visualization of melanoma tumor with lectin-conjugated rare-earth doped fluoride nanocrystals

MOLECULAR REPRODUCTION AND DEVELOPMENT 44:370-381 (1996)
Modifications of the Lectin Binding Pattern in the Rat Zona Pellucida After In Vivo Fertilization MANUEL AVILES,1.2 LOUAY JABER,2 MARIA TERESA CASTELLS,’ FREDERICK K.W. KAN,2 AND JOSE BALLESTA’ ‘Section of Histology and General Embryology, Department of Cell Biology, School of Medicine, University of Murcia, Murcia, Spain and ‘Department of Anatomy and Cell Biology, Faculty of Medicine, Queen’s University, Kingston, Ontario, Canada
ABSTRACT The zona pellucida (ZP) is an extra- cellular matrix surrounding the mammalian oocyte. It is involved in the sperm-egg adhesion phenomenon, induces the acrosome reaction, and participates in the late blockage to polyspermy. Thus, during the process of fertil- ization the cortical reaction is induced and the biochemical and biological properties of the ZP are modified. Some of these changes have been suggested to prevent the polyspermy. However, the mechanisms behind most of these changes are not well understood. Carbohydrate resi- dues of the ZP glycoproteins have been shown to play a key role in the early step of fertilization. In the present study, the changes produced in the terminal oligosaccha- ride sequences of the rat ZP glycoproteins after in vivo fertilization were investigated by means of lectin-gold cyto- chemistry. A comparative quantitative analysis of the den- sity of labeling in the ZP before and after fertilization was carried out by automatic counting of gold particles. The ZP of fertilized and unfertilized eggs were labeled by a battery of lectins including PNA, LFA, MAA, AAA, DSA, RCA I, and WGA. For all lectin studied in both fertilized and unfertilized eggs the labeling was preferentially located in the inner region of the ZP. After fertilization, binding of PNA, LFA, MAA, AAA, and DSA decreased in both inner and outer regions of the ZP. Labeling of RCA I-binding sites only decreased in the inner ZP, whereas reactivity to WGA was increased in the inner area of the ZP. Digestion of the thin-sections with neuraminidase prior to labeling with WGA resulted in a decrease of labeling for WGA binding sites. However, the labeling density of WGA binding sites was similar in both unfertilized and fertilized eggs upon treat- ment with neuraminidase.
The present results demonstrate that the oligosaccha- ride chains contained in the rat ZP are modified after fertil- ization of the oocyte. Cortical granules of the oocytes might be involved in these modifications by two mecha- nisms: 1) by hydrolysis of terminal carbohydrate residues of ZP glycoproteins by specific glycosidases contained in the granules; and 2) by addition of new glycoproteins to the ZP after the exocytosis of the cortical granules (cortical reaction). 0 1996 Wiley-Liss, Inc.
INTRODUCTION The zona pellucida (ZP) is an extracellular matrix
which surrounds mammalian oocytes and plays im- portant roles during the early steps of fertilization. Thus, ZP glycoproteins provide primary and secondary sperm receptors, induce acrosome reaction, and participate in the blockage to polyspermy (Wassarman and Bleil, 1982; Dunbar and Wolgemuth, 1984; Wassarman, 1987a; b).
The ZP is primarily composed of three to four glycopro- teins which are highly glycosylated (Wassarman, 1988). The carbohydrate residues of ZP glycoproteins play an important role in the sperm-egg recognition (Ahuja, 1982; Ahuja, 1985; Shalgi et al., 1986). Thus, ZP treated with specific enzymes (galactose oxidase, a-galactosi- dase, fucosidase, and N-acetylglucosaminidase), lectins, glycoconjugates and monosaccharides prevent the in vitro binding of sperm to eggs (Ahuja, 1982; Ahuja, 1985; Shalgi et al., 1986; Bleil and Wassarman, 1988; Miller et al., 1992). Following fusion of sperm and egg or activa- tion of egg, some changes in the properties of the egg cell surface (Stewart-Savage and Bavister, 1988; Cherr et al., 1988) and the ZP have been reported (Bleil and Wassarman, 1980; Bleil et al., 1981; Wassarman, 1988; Miller et al., 1992). One of the processes involved in bringing these changes is called the “zona reaction” and these changes are thought to be responsible for establish- ing a secondary block to polyspermy. This late blockage is due to the release of the content of cortical granules into the perivitelline space, a process known as “cortical reaction.” Several studies have established that cortical granules contain proteases and glycosidases (Moller and Wassarman, 1989; Miller et al., 1992,1994). Biochemical studies have reported that ZP glycoproteins of several mammalian species, including humans, are modified after fertilization (Bleil et al., 1981; Shabanowitz and O’Rand, 1989; Brown et al., 1990; Hatanaka et al., 1992). Thus, ZP2 and ZP3, the two major ZP-associated glyco- proteins in mouse, are transformed into ZP2f and ZP3f. In mouse, ZP3f has lost the ability to bind the sperm
Key Words: Lectins cytochemistry, Rat zona pellucida, Oocytes, Glycoconjugates, Fertilization
Received August 28, 1995; accepted November 28, 1995. Address reprint requests to Dr. Jose Ballesta, Section of Histology and General Embryology, Department of Cell Biology, University of Murcia, Murcia 30071, Spain.
0 1996 WILEY-LISS, INC.

LECTIN BINDING PATTERN MODIFICATIONS 371
TABLE 1. Lectin Characteristics Taxonomic name of the source Abbreviation Carbohydrate binding specificity References Arach is hypogaea Ricinus communis Datura stramonium
Galanthus nivalis
Helix pomatia
Dolichos biflorus
L i m a f l a w s
Maackia amurensis
Sambucus nigra
Tetragonolobus purpureas
Ulex europaeus
Aleuria aurantia
Triticum vulgaris
PNA
DSA RCA-I
GNA
HPA
DBA
LFA
MAA
SNA
LTA
UEA-I
AAA
WGA
Terminal GalP1,3 GalNAc disaccharides Terminal GalP1,4 GlcNAc disaccharides Bi-, tri- and tetraantennary sugar chains in
which at least one N-acetyllactosamine is present
Terminal Manul,3 in high mannose N-linked oligosaccharides
Terminal a- and p-linked GalNAc residues
Terminal a-linked GalNAc residues
Neu5Acu2,3/6Gal ,Neu5Aca2, 3iGGalNAc
Neu5Acu2,3Gal~1,4G1cNAc
Neu5Aca2, 6GaliGalNAc
Type 2 chains containing Fuc residues on C-2 of the Gal of Galpl,4GlcNAc, with or without a second Fuc on the GlcNAc
Specificity as LTA. It prefers the monofucosyl form
Complex type N-linked oligosaccharides with a u1,6 fucosyl residue at the innermost GlcNAc. Terminal al,2-linked Fuc residues
terminal non-reducing GlcNAc or Neu5Ac Glycoconjugates with a high density of
Lotan et al., 1975 Green et al., 1987 Yamashita et al., 1987
Shibuya et al., 1988
Piller et al., 1990; Wu and Sugii, 1991
Piller et al., 1990; Wu and Sugii, 1991
Ravindranath et al., 1985; Mandal and Mandal, 1990
Wang and Cummings, 1988; Knibbs et al., 1991
Shibuya et al., 1987; Mandal and Mandal, 1990
Pereira and Kabat, 1974; Sugii and Kabat, 1982
Sugii and Kabat, 1982
Osawa and Tsuji, 1987; Yamashita et al., 1985
Debray et al., 1981
and does not induce the acrosome reaction (Bleil and Wassarman, 1980; Miller et al., 1994). However, the elec- trophoretic mobility of ZP3f appears unaltered following fertilization or egg activation (Bleil et al., 1981). It would indicate that the proteic portion of the glycoprotein is not modified. Previous studies had suggested that cortical granule proteases were responsible for the zona reaction (Gwatkin et al., 1973; 1974). Recently, an N-acetylgluco- saminidase has been detected in mouse cortical granules (Miller et al., 1994). Taken together, results from previ- ous studies suggest that 1) oligosacchaide side chains of ZP glycoproteins could be modified by specific cortical granule glycosidases and 2) these changes might be re- sponsible for the late blockage to polyspermy. Therefore, the aim of the present study was to characterize the terminal oligosaccharide side chains of ZP glycoproteins of both ova and fertilized ova by means of lectin cyto- chemistry. The comparative quantitative analysis of the lectin-gold binding in the ZP before and after fertiliza- tion provide the quantitative and qualitative data con- cerning the modifications of ZP glycoproteins during the early steps in the fertilization process.
MATERIALS AND METHODS Reagents
Polyethyleneglycol (PEG; MW 20,000), sodium citrate, potassium carbonate, sodium ascorbate, and tetrachlo- roauric acid (HAuC1, . 2H20) were obtained from Merck (Darmstadt, Germany). Trypsin inhibitor (ovomucoid)- gold labeled (10 nm), fetuin, bovine serum albumin (BSA), galactose oxidase (from Dactylium dendroides), neuraminidase (type V from Clostridium perfringens),
D-galactose, a-methyl mannopyranoside, L-fucose, D-N-acetylglucosamine, D-N-acetyl-galactosamine, N- acetylneuraminic acid, N-acetylneuraminyllactose were purchased from Sigma (Poole, Dorset, UK). Digoxigenin (DIG)-labeled lectins (MAA, GNA, SNA, PNA, and AAA) and sheep polyclonal IgG anti-DIG-antibodies were purchased from Boehringer Mannheim Biochemica (Barcelona, Spain). Donkey anti-sheep immunoglobulin IgG-gold conjugated antibodies were purchased from Biocell Research Ltd (Cardiff, UK). Mouse monoclonal antibodies against trisaccharides a-D-Gal( 1-3)(a-L-Fuc [1-21) P-D-Gal (anti B blood group) were obtained from Dako (Santa Barbara, CA, USA). Goat anti-mouse IgG + IgM-gold complex (15 nm) was from Biocell (Cardiff, UK). Unlabeled lectins were obtained from Sigma (Poole, Dorset, UK) (RCA I, DBA, DSA, WGA, UEA I, and LTA), Pharmacia (Sweden) (HPA), and Cal- biochem Corporation (La Jolla, CA) (LFA). The taxo- nomic names and specificity of lectins used in the present study are summarized in Table 1.
Collection and Preparation of Oocytes and Embryos for Lectin-Gold Cytochemistry
Sexually mature female Wistar rats (n = 5 ) were ob- tained from Charles River Laboratories (St-Constant, Quebec, Canada). They were stimulated to superovulate by an intraperitoneal injection (i.p.) of 15 IU pregnant mare’s serum gonadotropin (PMSG, Equinex; Ayerst, Montrkal, Qukbec, Canada) followed 48 hr later, by an i.p. injection of 15 IU human chorionic gonadotropin (hCG, APL; Ayerst). For collection of embryos, after injection with PMSG and hCG, females were placed with

372 M. AVILES ET AL.
fertile males in the evening of injection with hCG. The female rats showing vaginal plug and non-mated rats were killed by ether anaesthesia 17 hr after injection with hCG. The ventral abdominal wall was immediately cut open, and the oviducts were removed and placed in the well of a porcelain dish containing 0.1 M cacodylate buffer (pH 7.4). To collect oocyte-cumulus complexes, the ampullary portion of the oviduct was identified and torn open with fine steel tweezers under a dissecting microscope. The oocyte-cumulus complexes collected were fixed for 2 hr at 4°C by immersion in 2% glutaralde- hyde in 0.1 M cacodylate buffer (pH 7.4). At the end of the fixation, the specimens were washed three times in 0.1 M cacodylate buffer and kept overnight at 4°C in the same buffer.
For preparation of thin sections, the oocyte-cumulus were dehydrated in a series of graded methanols, infil- trated, and embedded in Lowicryl K4M according to rou- tine procedure. Superovulated oocytes were first located in 1 pm-thick sections of Lowicryl-embedded specimens by light microscopy. Thin sections were then cut with glass knives on a LKB ultramicrotome and mounted on formvar-carbon-coated nickel grids.
Preparation of Lectin-DIG Complexes HPA-, DBA-, and LTA-DIG complexes were prepared
using the DIG antibody labeling kit from Boehringer Mannheim Biochemica (Barcelona, Spain) as previously reported (Martinez-Menhrguez et al., 1993). The DIG- lectin complexes were purified using a Sephadex G-25 column. Eight samples of 500 ~1 were pooled from each column. The sample with a higher absorbance at 280 nm was selected for cytochemical procedures: 1.38 (HPA), 1.72 (DBA), and 0.48 (LTA).
Preparation of Lectin-Gold and Glycoprotein-Gold Complexes
Monodispersed colloidal gold solutions with a mean particle diameter of 20 nm were prepared according to Frens (1973).
Lectin- and protein-gold complexes were prepared by mixing 10 ml of colloidal gold with 1 ml of bidistilled water containing either 130 pg of RCA I (pH 8.0), 35 kg of fetuin (pH 6.01, or 375 pg of UEA I (pH 6.3). The minimal account of lectins or protein needed for stabilization of colloidal gold and the optimal pH were estimated by the method of Geoghegan and Ackerman (1977).
Cytochemical Labeling For lectin cytochemistry at the ultrastructural level,
colloidal gold was used as a marker. One-step, two-step, and three-step methods were applied.
The one-step method was used for RCA I and UEA I. The lectins were diluted ten-fold in PBS with 0.5% BSA, 0.05% Tween 20, and 0.05% Triton X-100. Thin sections of Lowicryl-embedded oocyte-cumulus complexes were incubated with the lectin for 1 hr at room temperature.
For LFA, DSA, and WGA a two-step method was used. Ovomucoid-gold for DSA and WGA and fetuin-gold for
LFA were applied (Geoghegan and Ackerman, 1977; Roth, 1983; Egeaet al., 1989; Madrid et al., 1990; Castells et al., 1992).
For lectin-DIG labeling (MAA, AAA, DBA, PNA, HPA, AAA, and GNA) a three step-method was used (Martinez-Menarguez et al., 1992, 1993).
For monoclonal antibody labeling, grids were incu- bated in 1% BSA in PBS for 10 min and then with mouse monoclonal anti B blood group antibody (1:50) for 1 hr. After washing in PBS, the grids were incubated with gold labeled goat anti-mouse IgG + IgM diluted in PBS, 0.5% BSA, 0.05% Triton X-100, and 0.05% Tween 20.
After washing in the corresponding buffer and bidis- tilled water, the grids were counterstained with uranyl acetate and lead citrate and examined in an transmission electron microscope.
Controls The following controls were used: 1) Substitution of
conjugated and unconjugated molecules by the corres- ponding buffer; 2) Pre-incubation of the lectins with the corresponding hapten-sugar inhibitor used at a range of concentration from 0.1 to 1M: D-Gal (for PNA), a-methyl-a-Man (for GNA), D-GalNAc (for HPA and DBA), D-GlcNAc (for WGA), N-acetyl-lactosamine (for DSA and RCA I), L-Fuc (For UEA I, AAA, and LTA), Neu5Ac (for LFA), and N-acetylneuraminyllactose (for MAA and SNA); 3) Pre-incubation of the grids with neur- aminidase for LFA, SNA, and MAA and galactose oxidase for PNA.
Enzymatic Treatment Neuraminidase treatment. WGA shows an affin-
ity for GlcNAc and Neu5Ac residues. Digestion with neuraminidase followed by WGA staining was performed to establish the presence of sialic acid. Grids were treated with neuraminidase type V from Clostridiurn per- fringens (1 Uiml) in acetate buffer pH 5.0 at 37°C for 3 hr to remove Neu5Ac residues.
Controls Substitution of neuraminidase by acetate buffer pH
5.0. Galactose oxidase treatment. Ultrathin sections
were incubated with 50 U/ml of galactose oxidase in PBS at pH 7.4 for 24 hr at 37°C in a moist chamber.
Control Substitution of galactose oxidase by PBS (pH 7.4).
Quantitative Analysis At least five randomly selected areas of ZP of each un-
fertilized and fertilized eggs (five to ten) were photo- graphed on the electron microscope. Fifteen to twenty fields of both outer and inner ZP from each egg were used for the quantitative evaluation (Table 2). Quantitative evaluation was carried out by an Imco 10 image computer (Kontron Bildanalyse, Germany), using the Microm Im- age Processing software (Microm, Barcelona, Spain).
The density of labeling was determined by automatic

LECTIN BINDING PATTERN MODIFICATIONS 373
TABLE 2. Labeling Densities (Gold Particles/pm2) in the Rat ZP of Unfertilized and Fertilized Eggs*
Zona pellucida Unfertilized eggs Fertilized eggs
PNA" inner 25.70 ? 2.22 A 17.07 t 0.79 a 0
outer 7.30 ? 0.58 3.15 ? 0.39 0
MAA' inner 30.68 5 1.17 A 8.28 +- 0.40 outer 18.41 t 0.96 4.89 t 0.32 0
AAA' inner 18.99 t 1.73 A 10.15 t 0.62 0
outer 13.99 t 0.99 8.79 ? 0.83 WGAb inner 248.83 5 10.56 A 366.03 5 17.21 a
outer 205.21 t 10.43 212.03 t 12.68 NEU-WGAb inner 167.20 ? 8.81 A 166.00 t 10.73 8
outer 121.82 t 7.34 98.62 t 6.75 RCA I" inner 53.42 t 2.43 A 40.31 t 1.65 8
outer 40.43 5 2.06 26.70 t 1.31 0
LFAb inner 46.45 t 2.68 A 15.12 5 1.38 a 0
outer 12.69 5 0.87 5.03 t 0.60 DSAb inner 164.77 -t 3.58 A 95.89 t 2.25 8
outer 101.80 -t 3.60 38.50 t 1.92 0
*Labeling densities were expressed as mean 2- standard error (SE). "One step method. b T ~ o step method. "Three step method. Neu; neuraminidase. A Statistically significant differences between inner and outer areas of ZP in unfertilized eggs (P < 0.05). a Statistically significant differences between inner and outer areas of ZP in fertilized eggs (P < 0.05).
Statistically significant differences between fertilized and unfertilized eggs (P < 0.05).
counting of the gold particles enclosed in selected areas by a graphic digitizing tablet. Labeling densities were compared by Student's t-test (P < 0.05). A triple statisti- cal comparison was performed for: a) lectin labeling be- fore and after neuraminidase digestion; b) lectin labeling in the outer and inner ZP; and c) lectin labeling before and after fertilization.
RESULTS The labeling densities of various lectins are summa-
rized in Table 2. In the present study, the ZP has been arbitrarily divided into two zones of approximately equal thickness: inner and outer regions. The inner region is located close to the oocyte while the outer zone is facing the cumulus cells. These two regions show different morphological and cytochemical charac- teristics. For quantitative analysis, separation of the ZP into inner and outer zones was done on photomicro- graphs by drawing a line separating the ZP into two equal halves concentrically.
Unfertilized Tubal Eggs Zona pellucida. The ZP was labeled by WGA, PNA,
MAA, DSA, RCA I, AAA, and LFA (Figs. la , 3a, 4a, 5a, and 6a) but not reactive to HPA, LTA, UEA I, DBA, SNA, GNA, and anti-B antibody. The labeling density was always higher in the inner area of the ZP when compared with the outer zone (see Table 2). After neur- aminidase digestion a decrease in the WGA labeling was found in the ZP (Fig. lb).
Oocyte. In the ooplasm, several structures were la- beled. Cortical granules were detected beneath the plasma membrane of unfertilized eggs. They were mod-
erately labeled with WGA, PNA, DSA, RCA I, and LFA (Figs. la,c, 3a, 5a, and 6a). A weak labeling was detected with AAA. However, some cortical granules were unreac- tive to the above lectins. No reactivity was observed with the other lectins and antibody used in this study (Fig. 4a). After neuraminidase digestion, WGA labeling over cortical granules was abolished (Fig. lb,d). Multivesicu- lar bodies were labeled by DSA (Fig. 5a inset), LFA, RCA I, AAA, and WGA. The oocyte plasma membrane was intensely labeled by DSA (Fig. 5a).
Fertilized Tubal Eggs Zona pellucida. The lectins that were used to label
the ZP of unfertilized eggs were also employed for the detection of their corresponding binding sites in the ZP of fertilized eggs. A similar labeling pattern was obtained with these lectins in eggs before and after fertilization (Figs. 2,3b, 4b, 5b, and 6b). In fertilized eggs, the labeling densities of various lectins in the inner area of the ZP were higher than those of the outer area except for AAA where no significant difference was observed (see Ta- ble 2).
Oocyte. Occurrence of cortical granules in fertilized eggs was very scarce (Fig. 2). Lectin binding pattern associated with the oocyte plasma membrane and multi- vesicular bodies was similar to that of unfertilized eggs.
The lectin binding specificity was established by the results obtained after the preincubation of the lectin with the corresponding inhibitory sugar (Fig. 6c), substi- tution of conjugated and unconjugated molecules by the corresponding buffer and by incubation of the thin-sec- tions with neuraminidase or galactose oxidase.

374 M. AVILES ET AL.
Fig. 1. WGA followed by ovomucoid-gold. a: Unfertilized egg. An intense labeling is observed throughout the ZP. Note reactive cortical granules in the ooplasm (arrowheads). b: Unfertilized egg pretreated with neuraminidase. No reaction is observed over cortical granules (arrowheads). The reactivity in the ZP is decreased. c: Unfertilized egg.
Cortical granules (arrowheads) and the ZP are labeled. d Unfertilized egg pretreated with neuraminidase. Labeling over cortical granules (arrowheads) is abolished. CC, cumulus cell; CM, cumulus matrix; 0, oocyte; PVS, perivitelline space; ZP, zona pellucida. a: x 21,000; b: X 24,000; c: X 40,000; d: X 40,000.
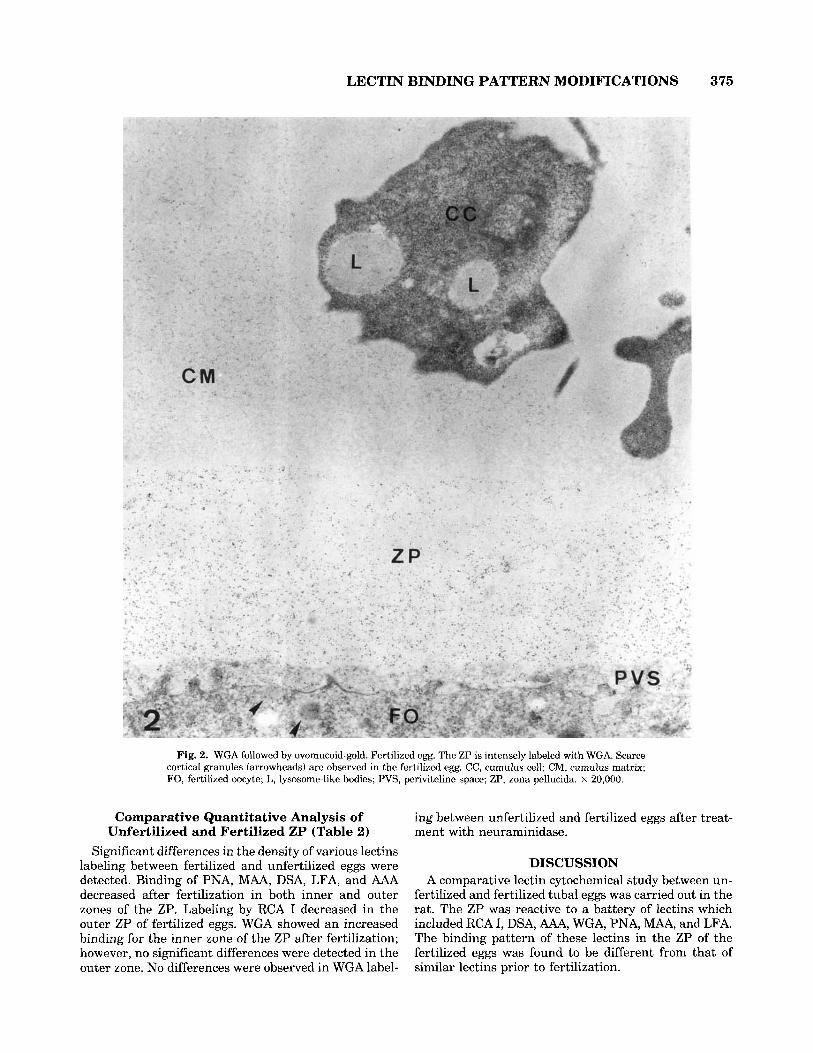
LECTIN BINDING PATTERN MODIFICATIONS 375
Fig. 2. WGA followed by ovomucoid-gold. Fertilized egg. The ZP is intensely labeled with WGA. Scarce cortical granules (arrowheads) are observed in the fertilized egg. CC, cumulus cell; CM, cumulus matrix; FO, fertilized oocyte; L, lysosome-like bodies; PVS, periviteline space; ZP, zona pellucida. x 20,000.
Comparative Quantitative Analysis of Unfertilized and Fertilized ZP (Table 2)
ing between unfertilized and fertilized eggs after treat- ment with neuraminidase.
Significant differences in the density of various lectins labeling between fertilized and unfertilized eggs were detected. Binding of PNA, MAA, DSA, LFA, and AAA decreased after fertilization in both inner and outer zones of the ZP. Labeling by RCA I decreased in the outer ZP of fertilized eggs. WGA showed an increased binding for the inner zone of the ZP after fertilization; however, no significant differences were detected in the outer zone. No differences were observed in WGA label-
DISCUSSION A comparative lectin cytochemical study between un-
fertilized and fertilized tuba1 eggs was carried out in the rat. The ZP was reactive to a battery of lectins which included RCA I, DSA, AAA, WGA, PNA, MAA, and LFA. The binding pattern of these lectins in the ZP of the fertilized eggs was found to be different from that of similar lectins prior to fertilization.

376 M. AVILES ET AL.
Fig. 3. PNA-DIGiSheep anti DIG/anti sheep IgG-gold. a: Unfertil- ized egg. The labeling intensity is higher in the inner region of the ZP. Cortical granules (arrowheads) are also labeled. b: Fertilized egg. Binding of PNA is decreased after fertilization. CC, cumulus cell; CM, cumulus matrix; FO, fertilized oocyte; m, mitochondria; 0, oocyte; IPVS, perivitelline space; ZP, zona pellucida. a: X 24,000; b: X 24,000.
Fig. 4. MAA-DIGiSheep anti DIG/ anti sheep IgG-gold. a: Unfertil- ized egg. The ZP is moderately labeled. Cortical granules (arrowheads) are not reactive. b Fertilized egg. The labeling over the ZP is decreased. CC, cumulus cell; CM, cumulus matrix; FO, fertilized oocyte; PVS, perivitelline space; ZP, zona pellucida. a: X 18,000; b: x 18,000.

LECTIN BINDING PATTERN MODIFICATIONS 377
Fig. 5. DSA followed by ovomucoid-gold. a: Unfertilized egg. An intense labeling is observed in the ZP. Cortical granules (arrows) are specifically labeled. Inset: DSA shows a moderate affinity for the multi- vesicular bodies and associated vesicles (arrowheads) contained in the ooplasm. b Fertilized egg. The lectin binding in the ZP is decreased
Modifications of the Lectin Binding Pattern in the ZP After Fertilization
A previous study, using lectin histochemistry at the light microscopical level, reported that no differences occurred in the hamster ZP before and after fertilization (Yanagimachi and Nicolson, 1976). However, the failure of detection of modifications of lectin binding patterns in the above study may be due to the limited resolution of light microscopy. In the present study, lectin-gold cytochemistry, which is known to be a sensitive and reliable approach for specific labeling of sugar moieties with high resolution and specificity a t the ultrastruc- tural level (Roth, 1983; 1987), was used in combination with quantitative analysis to examine the distribution of binding sites of a battery of lectins in the rat ZP before and after fertilization.
In a previous study, the occurrence of L-Fucose resi- dues a1,6 linked to the inner core region of N-linked glycoproteins of rat ovarian ZP was demonstrated
after fertilization. Note the reactivity in the oocyte plasma membrane. CC, cumulus cell; CM, cumulus matrix; FO, fertilized oocyte; 0, oocyte; PVS, perivitelinespace; ZP, zona pellucida. a: X 24,000; inset: X 22,000, b: X 22,000.
(Avilks et al., 1994a). Fucose residues play a key role in the fertilization process. Thus, treatment with a-L- fucosidase destroys the sperm receptor activity of mouse ZP (Bleil and Wassarman, 1988). Several studies have also demonstrated that fucose and glycans containing fucose are potent inhibitors of in vitro fertilization in rat and other mammals (Ahuja, 1982; Huanget al., 1982; Shalgi et al., 1986; Mahony et al., 1991). In the present study, reactivity of the lectin from Aleuria aurantia (AAA), which specifically binds fucose residues linked to the core region of N-linked oligosaccharides (Yama- shita et al., 1985) decreased after fertilization. Moreover, the presence of fucose-binding proteins in the sperm plasma membrane of several mammals have been pre- viously reported (Topfer-Petersen et al., 1985; Jones et al., 1988). Recently, the existence of a-L-fucosidase in the plasma membrane of rat sperm head has also been reported (AvilBs et al., 1994b). This enzyme might be involved in the sperm-egg recognition and adhesion dur-
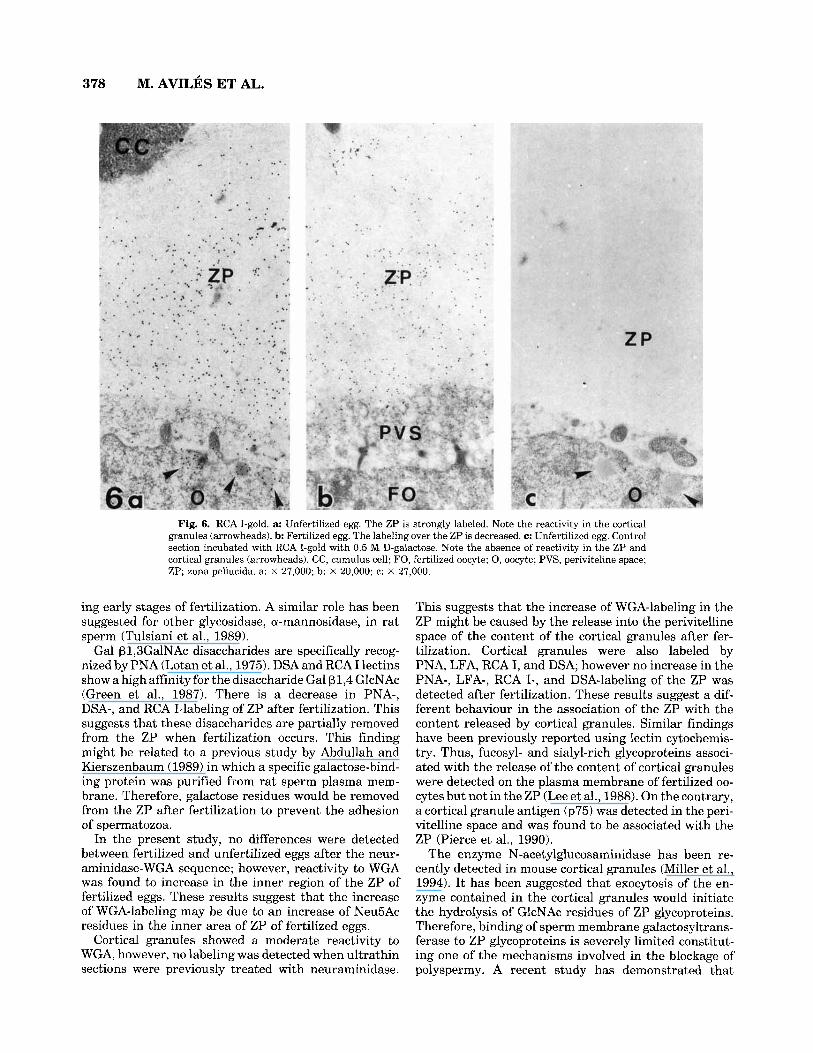
378 M. AVILES ET AL.
Fig. 6. RCA I-gold. a: Unfertilized egg. The ZP is strongly labeled. Note the reactivity in the cortical granules (arrowheads). b: Fertilized egg. The labeling over the Z P is decreased. c: Unfertilized egg. Control section incubated with RCA I-gold with 0.5 M D-galactose. Note the absence of reactivity in the ZP and cortical granules (arrowheads). CC, cumulus cell; FO, fertilized oocyte; 0, oocyte; PVS, periviteline space; ZP; zona pellucida. a: X 27,000; b: X 20,000; c: X 27,000.
ing early stages of fertilization. A similar role has been suggested for other glycosidase, a-mannosidase, in rat sperm (Tulsiani et al., 1989).
Gal f31,3GalNAc disaccharides are specifically recog- nized by PNA (Lotan et al., 1975). DSA and RCA I lectins show a high affinity for the disaccharide Gal @1,4 GlcNAc (Green et al., 1987). There is a decrease in PNA-, DSA-, and RCA I-labeling of ZP after fertilization. This suggests that these disaccharides are partially removed from the ZP when fertilization occurs. This finding might be related to a previous study by Abdullah and Kierszenbaum (1989) in which a specific galactose-bind- ing protein was purified from rat sperm plasma mem- brane. Therefore, galactose residues would be removed from the ZP after fertilization to prevent the adhesion of spermatozoa.
In the present study, no differences were detected between fertilized and unfertilized eggs after the neur- aminidase-WGA sequence; however, reactivity to WGA was found to increase in the inner region of the ZP of fertilized eggs. These results suggest that the increase of WGA-labeling may be due to an increase of Neu5Ac residues in the inner area of ZP of fertilized eggs.
Cortical granules showed a moderate reactivity to WGA, however, no labeling was detected when ultrathin sections were previously treated with neuraminidase.
This suggests that the increase of WGA-labeling in the ZP might be caused by the release into the perivitelline space of the content of the cortical granules after fer- tilization. Cortical granules were also labeled by PNA, LFA, RCA I, and DSA; however no increase in the PNA-, LFA-, RCA I-, and DSA-labeling of the ZP was detected after fertilization. These results suggest a dif- ferent behaviour in the association of the ZP with the content released by cortical granules. Similar findings have been previously reported using lectin cytochemis- try. Thus, fucosyl- and sialyl-rich glycoproteins associ- ated with the release of the content of cortical granules were detected on the plasma membrane of fertilized 00-
cytes but not in the ZP (Lee et al., 1988). On the contrary, a cortical granule antigen (p75) was detected in the peri- vitelline space and was found to be associated with the ZP (Pierce et al., 1990).
The enzyme N-acetylglucosaminidase has been re- cently detected in mouse cortical granules (Miller et al., 1994). It has been suggested that exocytosis of the en- zyme contained in the cortical granules would initiate the hydrolysis of GlcNAc residues of ZP glycoproteins. Therefore, binding of sperm membrane galactosyltrans- ferase to ZP glycoproteins is severely limited constitut- ing one of the mechanisms involved in the blockage of polyspermy. A recent study has demonstrated that

LECTIN BINDING PATTERN MODIFICATIONS 379
mouse ZP glycoproteins treated with N-acetylglucosami- nidase were not recognized by sperm membrane galacto- syltransferase (Miller et al., 1992). In the present study, no differences in the gold labeling densities in the ZP were detected between unfertilized and fertilized eggs after application of the sequence neuraminidase-WGA which specifically detects GlcNAc residues. This find- ing suggests that, in the rat, GlcNAc residues may not be involved in the blockage of polyspermy as has been suggested in the mouse. Different species-specific mechanisms for the blockage of polyspermy have been previously described. Thus, membrane galactosyltrans- ferase described in the mouse has not been detected in humans (Tulsiani et al., 1990).
In summary, the present study demonstrates that the oligosaccharide side chains of ZP glycoproteins are modi- fied by the fertilization of the oocyte. These findings provide further support to the hypothesis which suggests that the late blockage of polyspermy might be caused by the incorporation of cortical granule content into the ZP resulting in modifications of the oligosaccharide chains of ZP glycoproteins. Previous studies using im- munocytochemistry and lectin cytochemistry have re- ported that fertilization of the oocyte is accompanied by the release of the cortical granule content into the perivitelline space modifying the properties of the oocyte plasma membrane (Stewart-Savage and Bavister, 1988; Cherr et al., 1988) and the overlying acellular ZP (Barros and Yanagimachi, 1971; Wassarman, 1987a,b; 1990). These modifications caused the zona reaction and pre- vent the further entry of further spermatozoa into the oocyte (the so-called “block to polyspermy”). These changes are probably brought forth by specific glycosi- dases contained in the cortical granules; however, fur- ther studies are needed to confirm both the direct partici- pation of cortical granules in the modification of the ZP glycoproteins and the involvement of other enzymes in this process, particularly those released by the spermato- zoa during the acrosome reaction.
Uniformity of the ZP In a previous study, an asymmetric distribution of
some oligosaccharide chains in the rat ovarian ZP was reported (Aviles et al., 1994a). Thus, RCA I-labeling was higher in the outer ZP while binding sites of LFA, MAA, DSA, and AAA were homogeneously distributed and la- beling intensities of WGA and PNA were higher in the inner ZP. In the present study, ovulated eggs, both fertil- ized and unfertilized, also shows an asymmetric distribu- tion of lectin binding sites in the ZP. Thus, higher con- centrations of binding sites of PNA, MAA, WGA, RCA I, DSA, and LFA were detected in the inner area of the ZP; however AAA showed a uniform binding pattern only in the fertilized egg. A previous study has also re- ported a higher RCA I-labeling in the inner ZP of rat tuba1 eggs (Shalgi et al., 1991). In the hamster, changes in the lectin labeling of the ZP after ovulation (Roux and Kan, 1991) and changes in antibody labeling of the ZP and perivitelline space after fertilization (Kan et al., 1993; Kan and ROUX, 1995) have been reported. These
modifications might be partly caused by an internal re- distribution of the sugar residues after ovulation. Ovi- ductal secretions also contribute to the reported changes in the lectin bindingpattern of ZP (Roux and Kan, 1991). Previous studies have shown that some glycoproteins (called oviductins) secreted by the oviduct become associ- ated with the ZP of ovulated eggs (Araki et al., 1987; Boice et al., 1990; Buhi et al., 1993; Gandolfi et al., 1991; Kan et al., 1989,1993; Kapur and Johnson; 1985, 1986; Oikawa et al., 1988; Wegner and Killian, 1991). These results indicate that the ZP of postovulatory oocytes is further modified by the addition of oviduct-specific glycoproteins. However, the exact function(s) of these oviductins remain unclear and the physiological signifi- cance of the modifications of the ZP awaits to be defined. It is hoped that future studies will shed more light on the effects of ZP modifications on the process of fertilization.
ACKNOWLEDGMENTS We thank Mr. J . Moya, Mrs. M. D. L6pez-L6pez, and
Mrs. Gonzalez-Ulloa for technical assistance. We thank the Electron Microscopy Service and the Image Analysis Unit (Project CICYT IN89-0573) of the University of Murcia for excellent technical assistance. A portion of this work has been presented in abstract form at the 13th International Congress of Electron Microscopy. This work was supported by a grant from Comunidad Aut6noma de la Regi6n de Murcia PIB 93/04 (Spain) and a grant (MT-10904) from the Medical Research Council of Canada to F.W.K.K. (Canada). M.A. is supported by a fellowship (AP93 77506829) from the Ministerio de Educaci6n y Ciencia (Spain).
A substantial portion of this work was carried out during Manuel Aviles tenure as a Visiting Fellow in F.W.K.K. laboratory.
REFERENCES Abdullah M, Kierszenbaum AL (1989): Identification of rat testis galac-
tosyl receptor using antibodies to liver asialoglycoprotein receptor: purification and localization on surfaces of spermatogenic cells and sperm. J Cell Biol 108:367-375.
Ahuja KK (1982): Fertilization studies in the hamster: The role of cell surface carbohydrates. Exp Cell Res 140:353-362.
Ahuja KK (1985): Carbohydrate determinants involved in mammalian fertilization. Am J Anat 174:207-223.
Araki Y, Kurata S, Oikawa T, Yamashita T, Hiroi M, Naiki M, Sendo F (1987): A monoclonal antibody reacting with the zona pellucida of the oviductal but not with that of the ovarian egg of the golden hamster. J Reprod Immunol 11:193-208.
Avil6s M, Martinez-Menarguez JA, Castells MT, Madrid JF, Ballesta J (1994a): Cytochemical characterization of oligosaccharide side chains of the glycoproteins of rat zona pellucida: An ultrastructural study. Anat Rec 239:137-149.
Bleil JD, Wassarman PM (1980): Mammalian sperm-egg interaction: Identification of a glycoprotein in mouse egg zonae pellucidae pos- sessing receptor activity for sperm. Cell 20:873-882.
Bleil JD, Beall CF, Wassarman PM (1981): Mammalian sperm-egg in- teraction: Fertilization of mouse eggs triggers modification of the major zona pellucida glycoprotein, ZP-2. Dev Biol 86:189-197.
Bleil JD, Wassarman PM (1988): Galactose at the nonreducing termi- nus of 0-linked oligosaccharides of mouse egg zona pellucida glyco- protein ZP3 is essential for the glycoproteins sperm receptor activity. Proc Natl Acad Sci U S A 85:6778-6782.
Boice ML, McCarthy TJ, Mavrogianis PA, Fazleabas AT, Verhage HG

380 M. AVILES ET AL.
(1990): Localization of oviductal glycoproteins within the zona pellu- cida and perivitelline space of ovulated ova and early embryos in baboons (Papio anubis). Biol Reprod 43:340-346.
Brown CR, Clarke N, Aiken M, Bavister BD (1990): Changes in the composition of the hamster zona pellucida after fertilization in vivo but not in vitro. J Reprod Fert 90:447-454.
Buhi WC, O’Brien B, Alvarez IM, Erdos G, Dubois D (1993): Immuno- gold localization of porcine oviductal secretory proteins within the zona pellucida, perivitelline space, and plasma membrane of oviduc- tal and uterine oocytes and early embryos. Biol Reprod 48:1274 1283.
Castells MT, Ballesta J, Madrid JF, Martinez-Menarguez JA, Aviles M (1992): Ultrastructural localization of glycoconjugates in human bronchial glands: the subcellular organization of N- and 0-linked oligosaccharide chains. J Histochem Cytochem 40:265-274.
Cberr GN, Drobnis EZ, Katz DF (1988): Localization of cortical granule constituents before and after exocytosis in the hamster egg. J Exp Zoo1 246%-93.
Debray H, Decout D, Stecker G, Spik G, Montreuil J (1981): Specificity of twelve lectins towards oligosaccharides and glycopeptides related to N-glycosylproteins. Eur J Biochem 117:41-55.
Dunbar BS, Wolgemuth DJ (1984): Structure and function of the mammalian zona pellucida, a unique extracellular matrix. In BH
Satir (ed): “Modern Cell Biology”, Vol3. New York: Alan R. Liss, pp
Egea G, Goldstein IJ, Roth J (1989): Light and electron microscopic detection of (3 Gal h 1,4 GlcNAc bl) sequences in asparagine-linked oligosaccharides with the Datura stramonium lectin. Histochemis- try 92515-522.
Frens G (1973): Controlled nucleation for the regulation of the particle size in monodisperse gold suspensions. Nature Phys Sci 241:20-22.
Gandolfi F, MoldinaS, Brevini TAL, Galli C, Moor RM, LauriaA (1991): Oviduct ampullary epithelium contributes a glycoprotein to the zona pellucida, perivitelline space and blastomeres membrane of sheep embryos. Eur J Bas Appl Histochem 35~383-392.
Geoghegan WD, Ackerman GA (1977): Adsorption of horseradish perox- idase, ovomucoid and anti-immunoglobulin to colloidal gold for the indirect detection of concanavalin A, wheat germ and goat anti-hu- man immunoglobulin G on cell surfaces at the electron microscopic level: A new method, theory and application. J Histochem Cyto- chem 25:1187-1200.
Green ED, Brodbeck RM, Baenziger JU (1987): Lectin affinity high performance liquid chromatography. Interactions of N-glycanase- released oligosaccharides with Ricinus communis agglutinin I and Ricinus communis agglutinin 11. J Biol Chem 262:12030-12037.
Gwatkin RBL, Williams DT, Hartmann JF, Kniazuk M (1973): The zona reaction of hamster and mouse eggs: Production in vitro by a trypsin-like protease from cortical granules. J Reprod Fert 32: 259-265.
Gwatkin RBL, Williams DT (1974): Heat sensitivity of the cortical granule protease from hamster eggs. J Reprod Fert 39:153-155.
Hatanaka Y, Nagai T, Tobita T, Nakano M (1992): Changes in the properties and composition of zona pellucida of pigs during fertiliza- tion in vitro. J Reprod Fert 95431440.
Huang TTF, Ohzu E, Yanagimachi R (1982): Evidence suggesting that L-fucose is part of a recognition signal for sperm-zona pellucida at- tachment in mammals. Gamete Res 5:355-361.
Jones R, Brown CR, Lancaster T (1988): Carbohydrate-binding proper- ties of boar sperm proacrosin and assessment of its role in sperm- egg recognition and adhesion during fertilization. Development 102:781-792.
Kan FWK, St-Jacques S, Bleau G (1989): Immunocytochemical evidence for the transfer of an oviductal antigen to the zona pellucida of hamster ova after ovulation. Biol Reprod 40:585-598.
Kan FWK, Roux E, Bleau G (1993) Immunolocalization of oviductin in endocytic compartments in the blastomeres of developing embryos in the golden hamster. Biol Reprod 48:77-88.
Kan FWK, Roux E (1995) Elaboration of an oviductin by the oviductal epithelium in relation to embryo development as visualized by immu- nocytochemistry. Microsc Res Tech 31:478-487.
Kapur RP, Johnson LV (1985): An oviductal fluid glycoprotein associ- ated with ovulated mouse ova and early embryos. Dev Biol112:89-93.
Kapur RP, Johnson LV (1986): Selective sequestration of an oviductal
77-111.
fluid glycoprotein in the perivitelline space of mouse oocytes and embryos. J Exp Zoo1 238:249-260.
Knibbs RN, Goldstein IJ, Ratcliffe RM, Shibuya N (1991): Characteriza- tion of the carbohydrate binding specificity of the leukoagglutinating lectin from Maackia amurensis. Comparison with other sialic-acid specific-lectins. J Biol Chem 266:83-88.
Lee SH, Ahuja KK, Gilburt DJ, Whittingham DG (1988): The appear- ance of glycoconjugates associated with cortical granule release dur- ing mouse fertilization. Development 102595-604.
Lotan R, Skutelsky E, Danon D, Sharon N (1975): The purification, composition, and specificity of the anti-T lectin from peanut (Arachis hypogaea). J Biol Chem 250:8518-8523.
Madrid JF, Ballesta J, Castells MT, Hernhdez F (1990): Glycoconju- gate distribution in the human fundic mucosa revealed by lectin- and glycoprotein-gold cytochemistry. Histochemistry 95: 179-187.
Mahony MC, Oehninger S, Clark GF, Acosta AA, Hodgen GD (1991): Fucoidin inhibits the zona pellucida-induced acrosome reaction in human spermatozoa. Contraception 44:657-665.
Mandal C, Mandal C (1990): Sialic acid binding lectins. Experientia 46:433-441.
Martinez-Menarguez JA, Ballesta J , Aviles M, Castells MT, Madrid JF (1992): Cytochemical characterization of glycoproteins in the devel- oping acrosome of rats. An ultrastructural study using lectin histo- chemistry, enzymes and chemical deglycosylation. Histochemistry 97: 439-449.
Martinez-Menbrguez JA, Aviles M, Castells MT, Madrid JF, Ballesta J (1993): Glycosylation in Golgi apparatus of early spermatids of rat. A high resolution lectin cytochemical study. Eur J Cell Biol 61: 21-33.
Miller DJ, Macek MB, Shur BD (1992): Complementarity between sperm surface P-1,4-galactosyl-transferase and egg-coat ZP3 medi- ates sperm-egg binding. Nature 357589-593.
Miller DJ, Gong X, Decker G, Shur BD (1994): Egg cortical granule N-Acetylglucosaminidase is required for the mouse zona block to polyspermy. J Cell Biol 123:1431-1440.
Moller CC, Wassarman PM (1989): Characterization of a proteinase that cleaves zona pellucida glycoprotein ZP2 following activation of mouse eggs. Dev Biol 132:103-112.
Oikawa T, Sendai Y, Kurata S, Yanagimachi R (1988): A glycoprotein of oviductal origin alters biochemical properties of the zona pellucida of hamster egg. Gamete Res 19:113-122.
Osawa T, Tsuji T (1987): Fractionation and structural assessment of oligosaccharides and glycopeptides by use of immobilized lectins. Annu Rev Biochem 56:21-42.
Pereira MEA, Kabat EA (1974): Specificity of purified hemagglutinin (lectin) from Lotus tetragonolobus. Biochemistry 13:3184-3192.
Pierce KE, Siebert MC, Kopf G, Schultz RM, Calarco PG (1990): Charac- terization and localization of a mouse egg cortical granule antigen prior to and following fertilization or egg activation. Dev Biol 141:381-392.
Piller V, Piller F, Cartron J P (1990): Comparison of the carbohydrate- binding specificities of seven N-acetyl-D-galactosamine-recognizing lectins. Eur J Biochem 191:461-466.
Ravindranath MH, Higa HH, Cooper EL, Paulson JC (1985): Purifica- tion and characterization of an 0-acetylsialic acid-specific lectin from a marine crab Cancer antennarius. J Biol Chem 260:8850-8856.
Roth J (1983): Application of lectin-gold complexes for electron micro- scopic localization of glycoconjugates on thin sections. J Histochem Cytochem 31:987-999.
Roth J (1987): Light and electron microscopic localization of glycoconju- gates with gold labeled reagents. Scan Electro Microsc 1:695-704.
Roux E, Kan FWK (1991): Changes of glycoconjugate contents of the zona pellucida during oocyte growth and development in the golden hamster: a quantitative study. Anat Rec 230:347-360.
Shabanowitz RB, O’Rand M (1989): Molecular changes in the human zona pellucida associated with fertilization and human sperm-zona interactions. Ann N Y Acad Sci 541:621-632.
Shalgi RM, Matityahu A, Nebel L (1986): The role of carbohydrates in sperm-egg interaction in rats. Biol Reprod 34:446-452.
Shalgi R, Maymon R, Bar-Shira B, Amihai D, Skutelsky E (1991): Distribution of lectin receptors sites in the zona pellucida of follicular and ovulated rat oocytes. Mol Reprod Dev 29:365-372.
Shibuya N, Goldstein IJ, Broekaert WF, Nsimba-Lubaki M, Peeters B,

LECTIN BINDING PATTERN MODIFICATIONS 381
Pneumans WJ (1987): The elderberry (Sambucus nzgra L.) bark lectin recognizes the Neu5Ac(a2-6)Gal/GalNAc sequence. J Biol Chem 262:1596-1601.
Shibuya N, Goldstein IJ, Van Damme E m , Pneumans WJ (1988): Binding properties of mannose-specific lectin from snowdrop (Galan- thus nzualis) bulb. J Biol Chem 263:728-734.
Stewart-Savage J , Bavister BD (1988): A cell surface block to poly- spermy occurs in golden hamster eggs. Dev Biol 128:150-157.
Sugii S, Kabat EA (1982): Further immunochemical studies on the combining sites of Lotus tetragonolobus and Ulex europaeus I and I1 lectins. Carbohydr Res 99:99-101.
Topfer-Petersen E, Friess AE, Schill WB (1985): Evidence for a fucose- binding protein in boar spermatozoa. Histocbemistry 83:139-145.
Tulsiani DRP, Skudlarek MD, Orgebin-Crist MC (1989): Novel a-D- mannosidase of rat sperm plasma membranes: Characterization and potential role in sperm-egg interactions. J Cell Biol 109:1257-1267.
Tulsiani DRP, Skudlarek MD, Orgebin-Crist MC (1990): Human sperm plasma membranes possess a-D-mannosidase activity but no galacto- syltransferase activity. Biol Reprod 42343-858.
Wang WC, Cummings RD (1988): The immobilized leukoagglutinin from the seeds of Muackia umurensis binds with high affinity to complex-type Asn-linked 01-2,3 to penultimate galactose residues. J Biol Chem 263:4576-4585.
Wassarman PM, Bleil JD (1982): The role of zona pellucida glycopro- teins as regulators of sperm-egg interactions in the mouse In “Cellu- lar Recognition.” New York: Alan R. Liss, pp 845-863.
Wassarman PM (1987a): Early events in mammalian fertilization. Annu Rev Cell Biol 3:109-142.
Wassarman PM (1987b): The biology and chemistry of fertilization. Science 235:553-560.
Wassarman PM (1988): Zona pellucida glycoproteins. Annu Rev Bio- chem 57:415442.
Wassarman PM (1990): Profile of a mammalian sperm receptor. Devel- opment 108:l-17.
Wegner CC, Killian GJ (1991): In vitro and in vivo association of an oviduct estrus associated protein with bovine zona pellucida. Mol Reprod Dev 29:77-84.
Wu AM, Sugii S (1991): Coding and classification of D-galactose, N-acetyl-D-galactosamine, and -D-Galp-[1-3(4)1p-D-GlcpNAc, speci- ficities of applied lectins. Carbohydr Res 213:127-143.
Yamashita K, Kochibe N, Ohkura T, Ueda I, Kobata A (1985): Fraction- ation of L-fucose-containing oligosaccharides on immobilized Aleuria aurantia lectin. J Biol Chem 260:4688-4693.
Yamashita K, Totani K, Ohkura T, Takasaki S, Goldstein IJ, Kobata A (1987): Carbohydrate binding properties of complex-type oligosac- charides on immobilized Datura stramonium lectin. J Biol Chem
Yanagimachi R, Nicolson GL (1976): Lectin-binding properties of ham- ster egg zona pellucida and plasma membrane during maturation and preimplantation. Exp Cell Res 100:249-257.
262:1602-1607.
Copyright © 2022 FDOKUMEN




















