
Comparative analysis of amplified and nonamplified RNA for hybridization in cDNA microarray
Upload
independentCategory
view
1download
0
RESEARCH PAPER
A phloem-enriched cDNA library from Ricinus: insightsinto phloem function
C. Doering-Saad, H. J. Newbury, C. E. Couldridge, J. S. Bale and J. Pritchard*
School of Biosciences, University of Birmingham, Edgbaston, Birmingham B15 2TT, UK
Received 20 January 2006; Accepted 10 June 2006
Abstract
The aim of this study was to identify genes that are
expressed in the phloem. Increased knowledge of
phloem regulation will contribute to our understand-
ing of its many roles, from transport of solutes to
information about interactions with pathogens. A cDNA
library constructed from phloem-enriched sap exuding
from cut Ricinus communis (L.) hypocotyls was se-
quenced. To assess contamination from other tissues,
two libraries were constructed: one using the first
15 min of exudation and the other from sap collected
after 120 min of exudation had elapsed. Of 1012 clones
sequenced, 158 unique transcripts were identified. The
presence of marker molecules such as profilin, the low
occurrence of chloroplast-related mRNAs, and the
sieve element localization of constituent mRNA using
in situ hybridization were consistent with a phloem
origin of the sap. Functional analysis of the cDNAs
revealed classifications including ribosomal function,
interaction with the environment, transport, DNA/RNA
binding, and protein turnover. An analysis of the
closest Arabidopsis thaliana (L.) homologue for each
clone indicated that genes involved in cell localization,
protein synthesis, tissue localization, organ localiza-
tion, organ differentiation, and cell fate were repre-
sented at twice the level occurring in the whole
Arabidopsis genome. The transcripts found in this
phloem-enriched library are discussed in the context
of phloem function and the relationship between the
companion cell and sieve element.
Key words: Arabidopsis, cDNA, companion cell, functional
genomics, phloem, plasmodesmata, Ricinus, sieve element,
translocation, transport.
Introduction
Phloem is essential for plant function, having many roles,including the transport of solutes from sources such asphotosynthesizing leaves to sinks such as growing root tips.It can be argued that plant selection and breeding toincrease yield over the last 10 000 years has actually beendeveloping varieties with more efficient solute and waterpartitioning through the phloem. While its transport role iswell known, it also acts to transmit developmental signalsaround the plant (Lucas and Lee, 2004; Oparka, 2004) andis the site of immediate defence responses against patho-gens, ranging from viruses to aphids, and also for systemicsignalling (van Bel and Gaupels, 2004). However, despiteits importance, relatively little is known about the way inwhich phloem function is regulated. Indeed, debates aboutthe biophysical mechanism of transport have only recentlybeen fully resolved (van Bel, 2003b; Gould et al., 2004).
Crucial to an understanding of phloem function is thefact that early in development the sieve element loses itsnucleus. However, the associated companion cell retainsa nucleus and maintains sieve element integrity and func-tions. These include a requirement for membrane trans-porter proteins, systems to ameliorate oxidative damage,regulation of nitrogen metabolism and a range of pathogendefence systems.
Companion cells and sieve elements are connected byplasmodesmata. These are plasma membrane-bound poresin the cell wall through which smooth endoplasmic re-ticulum is insinuated: microfilaments may be associatedwith the central membrane core (Oparka, 2004).
The presence of mRNAs in the anucleate sieve element(Xoconostle-Cazares et al., 1999) raises the question oftheir role in this cell type. There are two reasons whymRNA might be present in the sieve element. First, it maybe dragged by bulk flow of solution from companion cell tosieve element and therefore reflect the gene expression in
* To whom correspondence should be addressed. E-mail: [email protected]: DIG, digoxigenenin; RT–PCR, reverse transcription–polymerase chain reaction.
Journal of Experimental Botany, Vol. 57, No. 12, pp. 3183–3193, 2006
doi:10.1093/jxb/erl082 Advance Access publication 25 August, 2006This paper is available online free of all access charges (see http://jxb.oxfordjournals.org/open_access.html for further details)
ª 2006 The Author(s).This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) whichpermits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

the companion cell; for example, the thioredoxin and actinmRNA detected in the sieve element of rice (Sasaki et al.,1998). Secondly, some mRNA may have a role in long-distance transfer of information, termed macromolecu-lar trafficking. In situ hybridization has demonstrated thelong-distance movements of mRNA through grafted tissue(Ruiz-Medrano et al., 1999). mRNA moving in the sieveelement can have a phenotypic effect distant from its source(Haywood et al., 2005). For example, in Arabidopsis,mRNA of the FLOWERING LOCUS (T) gene was traf-ficked to the shoot apex and activated other genes involvedin the initiation of flowering (Huang et al., 2005).
Since RNA can be detected in the sieve element (Sasakiet al., 1998; Doering Saad et al., 2002) and annotations canindicate cDNA function (Asano et al., 2002; Nakamonoet al., 2003; Vilaine et al., 2003), analysis of sieve elementcDNA can provide information about gene expression inthe companion cell. The presence of mRNA for a sucrosetransporter, an aquaporin, and a proton ATPase has beendemonstrated using aphid stylectomy to sample the sieveelement followed by reverse transcription–polymerasechain reaction (RT–PCR) (Doering-Saad et al., 2002).Immunolocalization and in situ hybridization can alsoprovide useful information; however, these approachespreclude the identification of genes or proteins not pre-dicted by existing models. An alternative approach is toidentify all genes expressed in phloem tissue. Such studiesare limited by the relatively small proportion of phloemtissue in any plant organ and the difficulty in obtaining pureor enriched samples. Gene expression has been studied inphloem-enriched samples from Apium graveolens L.(Vilaine et al., 2003) or using laser capture microdissection(Asano et al., 2002; Nakazono et al., 2003). Pure phloemsap can be obtained from cut aphid stylets (Pritchard,1996); however, insufficient sap could be obtained to makecDNA libraries, in part because sieve element sap hasa very low mRNA concentration (Doering-Saad et al.,2002). Based on a large body of work by Milburn, it hasbeen demonstrated that large quantities of ‘pure’ phloemsap can be collected from Ricinus communis plants(Milburn, 1970). This plant has been heavily used instudies of phloem function (Smith and Milburn, 1980;Schobert et al., 1998; Komor, 2000). Additionally it hasbeen demonstrated previously that the protein compositionof phloem sap obtained from Ricinus by exudation isqualitatively the same as that of pure sap obtained from thesame plant by aphid stylectomy, but different from wholeleaf or stem (Barnes et al., 2004). We are thereforeconfident that the sap obtained by exudation is representa-tive of Ricinus phloem sap.
In the present study, genes relevant to phloem functionwere identified by creating cDNA libraries from Ricinusphloem sap. The data obtained complement emerginginformation on the phloem transcriptome (Asano et al.,2002; Nakazono et al., 2003; Vilaine et al., 2003) and
Ricinus phloem protein libraries (Barnes et al., 2004) andthose from other plant species (Hayashi et al., 2000;Hoffmann-Benning et al., 2002; Walz et al., 2004). ThecDNAs cloned have been sequenced and, by identifyinghomologues in sequence databases, suggestions are madeconcerning the functions of the transcripts detected.
Materials and methods
Plant material and growth conditions
Seeds of R. communis (Sheffield’s Seed Company, New York) weresurface-sterilized and germinated on sterilized vermiculite in the darkat 25 8C for 3 d. Germinated seedlings were transferred to singlepots containing vermiculite saturated with Long Ashton NutrientSolution and grown at 25 8C in a 16:8 h long day photocycle anda PAR of ;200 mmol m�2 s�1.
Collection of phloem exudates
Eight-day-old seedlings were used for the collection of phloemexudate, with their hypocotyl hook emerging well above thevermiculite, while their cotyledons were still below the surface ofthe substrate. Plants were transferred into a high humidity chamberfor 2 h, after which the apex of the hypocotyl hook was severed andboth sides were blotted with sterilized filter paper. Both sides of thecut hypocotyl usually started to exude immediately, i.e. xylemexudate from the rootlet stump and phloem exudate from the coty-ledon stump. Phloem sap was collected over the period immediatelyfollowing cutting for 15 min. This sap was assumed to have thegreatest contamination and was used to construct the early library.Subsequently, phloem exudate was aspirated off the cotyledon stumpat 30, 45, 65, and 90 min to wash off contaminating material. Sap wasthen collected after 120 min and was used to construct the late library.Following collection, exudates (2–6 ll) were immediately buffered
with 2 vols of RNase inhibitor buffer [0.5 U ll�1 ANTI-RNase(Ambion), 1 U ll�1 RNAsin (Promega) in 10 mM TRIS (pH 8.0)]and frozen in liquid nitrogen.
Quality testing of exudates
In order to assess the quality of the exudate in terms of the presenceof phloem-enriched mRNAs but the absence of contaminatinggenomic DNA (leaching out of the cut parenchyma cells), RT–PCRand PCR were performed with primers for profilin on a small aliquotof collected phloem and xylem exudate.Briefly, reverse transcription was performed using 1.5 ll of
buffered exudate collected at the various time points using theSensiscript Reverse Transcription Kit (Qiagen) in the presence of anoligo(dT) primer (dT15) in 5 ll reactions. Multiple negative controls(containing water instead of exudate) and positive controls (contain-ing DNase I-treated total RNA from Ricinus cotyledons) wereincluded in the reverse transcription reactions. Entire reverse tran-scription reactions were amplified by gene-specific PCR withHotStarTaq (Qiagen) on a Hybaid OnmiGene cycler, using anintron-spanning primer pair for Ricinus profilin (RcRro_F 59-CGAACCAAACCCTTCCATCGAATTTC, RcPro_R 59-TATAG-CAGGTAGGCAAATGCAAGC).PCRs designed to detect minute traces of contaminating geno-
mic DNA were carried out with HotStarTaq on 1.5 ll of bufferedexudates for up to 45 cycles, alongside negative and positive controls(water and 10 pg of Ricinus genomic DNA, respectively).
Construction of cDNA libraries from phloem exudates
An early and a late library were constructed to test whether exudateswere contaminated by non-sieve element cells. Damaged cells
3184 Doering-Saad et al.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

neighbouring the sieve element would be expected to lose theircontents early during exudate collection so that sieve element sapcollected later would be expected to be more representative of sieveelement sap. The ‘early’ library was constructed from the phloemexudate collected 15 min after cutting the hypocotyl hook (i.e.t15). The ‘late’ library was constructed from the phloem exudatecollected after 120 min (i.e. t120). mRNA contamination fromsurrounding cut cells was reduced by aspirating off the exudate thataccumulated in the interim, thus washing the cut surface of thehypocotyl.As phloem exudate contains only minute traces of mRNA, a PCR-
amplified cDNA library was constructed (PCR cDNA libraryConstruction Kit, Stratagene) according to the manufacturer’s recom-mendations. As starting material, 5 ll of buffered phloem exudatewere used in the reverse transcription reaction and, after strandsynthesis and adaptor ligation, the double-stranded cDNA was PCRamplified for 29 cycles. The PCR-amplified product was cloned intopCMV-PCR (Stratagene) and transformed into Escherichia coliXL10 Gold. An analysis of the transformants revealed that ;95%of the clones contained an insert, averaging 750 nt in size. InsertcDNAs from isolated clones were amplified with conventional T3and T7 primers and sequenced. BLAST searches were performed onthe sequences obtained between March and May 2006. Those cloneswith significant matches (e <1�10), and clear Arabidopsis homo-logues were identified.
RNA extraction and northern analysis
Total RNA was extracted from source leaves, stem, and roots takenfrom 6-month-old Ricinus plants using the RNeasy Plant Kit(Qiagen). A 10 lg aliquot of total RNA was separated alongside3 lg of RNA size marker (Sigma) using conventional protocols.RNA was transferred onto a Bright Star Plus nylon membrane(Ambion) and fixed by UV cross-linking.Using primers directed against internal regions of the chosen
library clones (Table 1), gene-specific probes were generated. Anadditional probe directed against the 18S rRNA was generated usingthe QuantumRNA Plant 18S Internal Standard Kit (Ambion). PCR-amplified products were gel purified (QIAquick Gel purification Kit,Qiagen) and used to generate radioactive, randomly primed, andstripable probes (StripEZ DNA Kit, Ambion). Pre-hybridizationand hybridization (overnight at 42 8C) were carried out in Ambion’sformaldehyde-containing UltraHybe buffer. Blots were washed athigh stringency and radioactive signals detected by exposure of theblots to a phosphor image screen (Imaging Screen K, BioRad). Probe-specific signal intensities were quantified (Quantity One software,BioRad).Calculating the ratio between the values obtained with the gene-
specific probes and those obtained with the 18S rRNA internalstandard allowed signal intensities to be normalized. By employingthis strategy, small variations in the amounts of RNA loaded onto the
lanes of the original northern blot do not influence the accuracy of theresults.
In situ hybridization
Longitudinal and transverse tissue sections of 8-d-old Ricinushypocotyls and 6-month-old Ricinus stems were fixed in formalde-hyde and embedded in Paraplast prepared according to Jackson(1991). Ribbons of consecutive sections (15 lm thick) were mountedonto polylysine-coated slides (BDH). Slides were dewashed, rehy-drated, permeabilized by treatment with pronase, acetylated, andfinally dehydrated as previously described (Jackson, 1991). Onlyfreshly pretreated slides were used for in situ hybridization.Sequences were selected for northern analysis on the basis of the
confidence with which their function could be identified followingBLAST searching and an assessment of the relevance of these func-tions to the operation of sieve elements. These included sequencesencoding peroxiredoxin, ribosomal protein S25, an ABC transporter,glutathione transferase, a radical-induced cell death 1 protein (RCD1)involved in oxidative stress (Overmyer et al., 2005; Ahlfors et al.,2004), a sieve element-localized restricted TEV movement protein(RTM2) interacting with viruses (Chisholm et al., 2001), a potassiumtransporter, and an actin-depolymerizing factor. In situ sense andantisense probes were generated by PCR amplification of internalfragments using the primers listed in Table 1. Gel-purified PCRproducts (QIAquick Gel purification Kit, Qiagen) were subclonedinto EcoRV-linearized pZERO-2 (Invitrogen), where T7 and SP6promoter sites flank the cDNA insert. The sequence and orientation ofcDNA present in all subclones were verified by sequencing. Vectorswere linearized immediately downstream of the cDNA insert andpurified by phenol/chloroform extraction. Labelled RNA was gen-erated (DIG RNA labelling Kit, Roche) by run-off transcription usingeither T7 or SP6 RNA polymerase in the presence of digoxigenin(DIG)-UTP. The integrity and quantity of the resulting sense andantisense RNAs were estimated by running the RNA alongside aDIG-labelled standard of known concentration. Probes were hydro-lysed to ;200–150 nt in size and purified according to conventionalprotocols (‘Nonradioactive in situ Hybridisation Application Man-ual’, Roche, 2nd edn, p. 142).Freshly prepared slides were hybridized to hydrolysed and
denatured, DIG-labelled sense or antisense probes (1 lg ml�1 finalprobe concentration) for 16 h at 42 8C in a high humidity chamber,followed by removal of non-specifically hybridized probes. DIG-labelled hybrids were detected with alkaline phosphatase-conjugatedanti-DIG Fab fragments (3.75 U ml�1) as recommended (DIGNucleic Acid Detection Kit, Roche). The substrate solution forcolour development contained 10% polyvinyl alcohol (70–100 kDa)in addition to 5 mMMgCl2, 0.2 mM BCIP, and 0.2 mM NBT. Slideswere incubated vertically in colour development solution in thedark overnight at room temperature, after which washing in TEstopped the reaction. Slides were dehydrated by incubation inethanol, and dried.
Table 1. Primers for the generation of probes used in northern analysis derived from sequences present in the early library (i.e. madefrom sap collected over the 15 min following hypocotyl excision)
Target gene Forward primer Reverse primer
Peroxiredoxin N6_F1 59-GGTTGCCAATCTTGAAGAAGG N6_R1 59-TTTTTGCATCCGAAACAAGCRibosomal protein S25 S5_F1 59-CAGATCCAATGGCACCAAAG S5_R1 59-GGCAGACAATACTCGAATGACABC transporter U5_F1 59-CACCACAAACGGGAAGATG U5_R1 59-TTTGCAGCAAATGCTCACCGlutathione transferase S8_F1 59-TGACCAGCAGAAGGTCTARG S8_R1 59-GCAGCAAGGAGAAAATGAAAGRadical-induced cell death RCD1 W10_F1 59-GTCAAGGTTCAAGCAATACAA W10_R1 59-CAAAGCCCTTCAAAGTTGTTCTEV movement 1 RTM2 M10_F1 59-ATGGCAACAAGAACAGGAAC M10_R1 59-CCTTAGGATTAAGAGCTGCTTCPotassium transporter M5_F1 59-CTTGAAATTTGTCGCGTCCC M5_R1 59-TCTTTGTTGGCTGAAGTAGTTCActin-depolymerizing factor O9_F1 59-AACTATGGCTAATGCTGCATC O9_R1 59-ACACACACCTTGCATAAGTTC
Phloem cDNAs 3185
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Results and discussion
Quality testing of exudates
Comparison of xylem exudate from the rooted stump ofsevered hypocotyls with phloem sap from the apical portionof the hypocotyl provided a method for assessing the purityof the sieve element sap used for cDNA production. Phloemsap had the expected chemical characteristics comparedwith xylem sap of high pH and osmotic pressure (phloempH of 7.360.06 compared with 5.860.06 xylem, phloemosmotic pressure of 1.560.07 MPa compared with0.760.06 MPa for xylem). Additionally, the proteincomposition of phloem sap obtained in this way fromRicinus was qualitatively the same as that of sap obtainedby aphid stylectomy (Barnes et al., 2004).
The phloem origin of the sap was further examined bytesting for the presence of proteins previously located in thephloem. Profilin was present in the phloem of Ricinushypocotyls, with lower expression in epidermis, pith, andxylem (Schobert et al., 2000). In this study, after reversetranscription and 25 cycles of sequence-specific PCR,profilin mRNA was always detected in phloem exudate,but never in xylem exudate, providing further evidencesupporting a sieve element origin for the exuding sap.
Preliminary analysis of the early and late libraries: Thecontents of cells damaged during cutting of the hypocotylwould be expected to be washed away during the exudatecollection procedure so that sieve element sap for the latelibrary would be expected to be more representative ofsieve element sap.Accordingly, the annotations of cDNAse-quences found in the early and late libraries were compared.
The inserts of 34 random clones from the early librarywere sequenced. BLAST searches were performed usingthese sequences between March and May 2006; 21 cloneshad both significant matches (e <1�10) in the databases andclear Arabidopsis homologues.
The inserts of 1012 clones of the late library weresequenced. A total of 267 of these had a significant match(e <1�10) to a plant sequence in the databases. Of these, 40%were represented only once, 14% twice, and the remaindermore than twice in the library. Thus 158 non-redundant,unique clones were identified. This compares with an analy-sis of 793 clones from a celery phloem-enriched cDNAlibrary in which 87% of the clones were singletons, 9.5%duplicates, and 3% were represented more than three times(Vilaine et al., 2003). The closest matching to an Arabi-dopsis gene was also recorded for each cDNA.
Northern and in situ analysis: The phloem specificity of thelibraries was further examined by determining the celllocalization of selected messages found in the library usingin situ hybridization. First, northern analysis was carriedout to determine which clones would be subsequentlyanalysed in more detail. The last round of probing wascarried out with the probe for 18S rRNA, and its signal
intensity quantified by phosphorimage analysis to correctthe signal intensities for differences in loading. Signalintensities were normalized by defining the signal strengthfrom the leaf as 1 (Fig. 1).
Based on the northern analysis, peroxiredoxin wasselected for use in in situ analysis since it was highlyexpressed in stem tissue and has been previously localizedto phloem (Rouhier et al., 2001). Hydrolysed sense andantisense probes corresponding to internal fragments of thecDNA present in the phloem library were used for in situhybridizations on transverse sections of Ricinus stem andhypocotyls. Peroxiredoxin antisense, but not the corre-sponding sense, probes showed specific staining of isolatedcells belonging to the phloem both in Ricinus hypocotyls,the tissue from which the library was constructed, and inadult stem transverse sections (Fig. 2a–d). In the youngertissue, there was some signal around the xylem not appar-ent in the older tissue. Thus, sap solute characteristics, thepresence of known phloem message, the northern analysisresults, and the sieve element localization of the phloemcDNA for peroxiredoxin were consistent with a sieveelement origin for the cDNA clones.
Assessing the purity of phloem sap: genes involved inphotosynthesis in early and late libraries
One strategy for assessing the degree of contamination ofthe cDNA libraries is to determine the frequency of clonesthat would be expected to be found in other cell types butnot in the sieve element–companion cell complex (Vilaineet al., 2003). On this basis, chloroplast-associated sequen-ces were looked for in both the early and late libraries. This
01234567
perox
iredo
xin
ribos
omal
protei
n S25
ABC trans
porte
r
glutat
hione
trans
feras
e
ATP8 (vir
al mov
emen
t)
K+-tran
sport
er
actin
depo
lym. fa
ctor
Relative in
ten
sity leaf
stem
root
Fig. 1. Relative expression of selected library clones in different tissuesof R. communis. Relative expression of selected mRNAs in leaf, stem,and root tissue of 6-month-old Ricinus plants. Northern blots containing10 lg of RNA of leaf, stem, and root tissue per lane were probed withradioactively labelled PCR-generated probes specific to internal sequen-ces of the genes indicated. Northern blots were also probed with a probedirected against the 18S rRNA. After hybridization, the signal intensitieswere quantified using a phosphorimager. The signal strengths of the gene-specific probes were expressed as a ratio (value obtained for gene x/valueobtained for 18S rRNA) to account for slight variations in the amountof total RNA loaded onto each lane of the original northern. Finally, theratio obtained for ‘leaf’ in each hybridization experiment was arbitrarilydefined as 1.
3186 Doering-Saad et al.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

is not straightforward, since sieve elements are known tocontain plastids (‘P-plastids’). Although these do not performphotosynthesis, their function(s) is unknown (Knoblauchand van Bel, 1998). They form part of the wide spectrumof plastid types and, for the purposes of this analysis, it wasassumed that they may be capable of all of the metabolicactivities found in the chloroplast except photosynthesis.
Using the best Arabidopsis homologue hit for eachclone, all of the sequences in both early and late librariesreferring to plastids (or chloroplasts) in their annotationswere identified. Extensive searches were carried out usingsequence databases to obtain all of the available information
about the function of the proteins encoded by these clones.For some of the predicted proteins, it was not possible toput forward a reasonable case that they were located inplastids (e.g. At1g06690, At3g09830, and At3g25560). Forothers, it appeared quite possible that they were located inP-plastids and other plastid types as well as chloroplasts(e.g. At1g75330 encoding ornithine carbamoyltransferase;At4g22590 encoding trehalose phosphatase; At5g35630encoding glutamate-ammonia ligase; and At1g80560encoding an enzyme involved in leucine synthesis).However, for two clones of the early library (At1g03130and At2g06520) and three clones of the late library(At1g03130, At3g27700, and Atcg00680), the predictedproteins were clearly involved in photosynthesis.
This crude comparative measure of contamination there-fore reveals two out of 21=9.5% (or, omitting duplicates,11.8%) photosynthetic sequences in the early library butonly three out of 267=1.1% (or, omitting duplicates, 1.9%)in the late library. For comparison, a search of the GOannotations for chloroplast/photosynthesis-related genesrevealed >4500 genes, or;20% of the Arabidopsis genome.Thus, the late library appears much less contaminated withphotosynthetic sequences than the early library, and bothearly and late libraries contain considerably fewer photo-synthetic-related sequences than are represented in thewhole Arabidopsis genome.
Characterization of major gene classes in the early andlate libraries
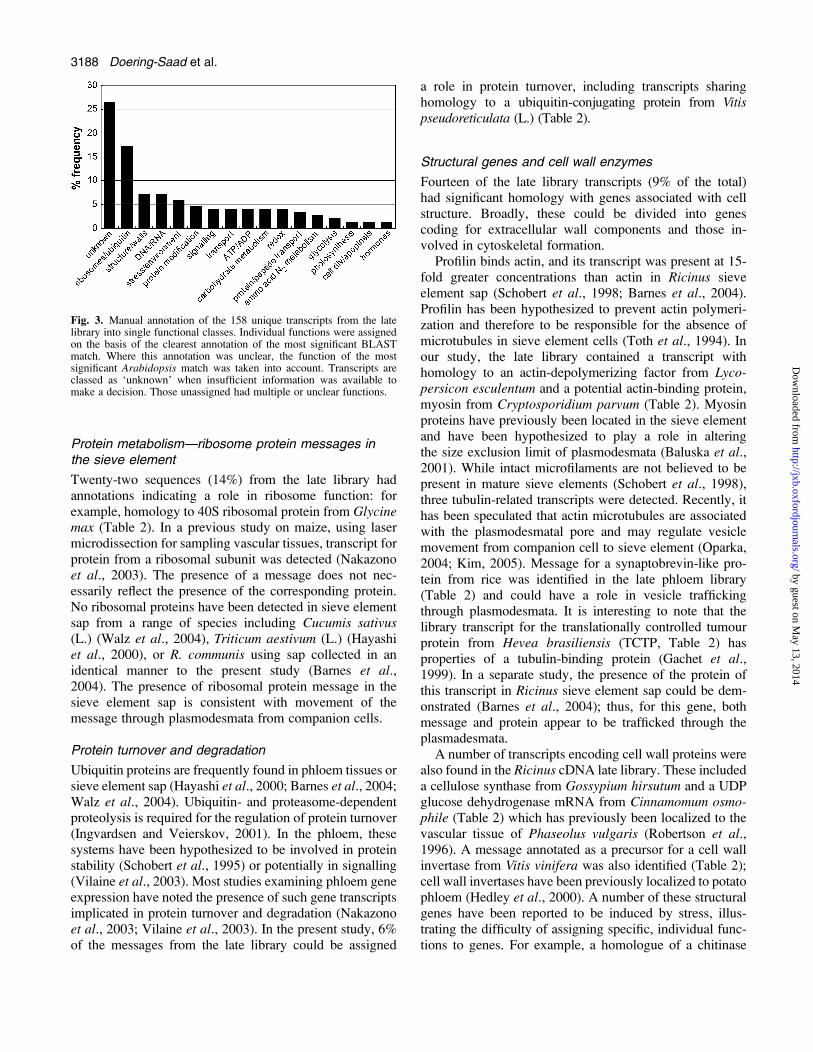
A total of 158 non-redundant sequences from the latelibrary were identified as having significant homology inthe databases. The full list of genes is shown in thesupplementary material available at JXB online. Thesesequences were derived from a wide range of plants (andother organisms), reflecting their use as experimentalspecies, and were divided into 19 individual functionalcategories based on their annotations. Twenty-five sequen-ces (16% of the total) were of unknown function (Fig. 3).It is expected that companion cells, and potentially sieveelements, will contain transcripts representing generallyexpressed genes in addition to a subset of transcripts relatedto the specific function of phloem tissues. Thus, thepresence of a transcript in the library does not necessarilyindicate an exclusively phloem-specific function. To dis-criminate between transcripts representing gene expressionthroughout the plant and those specific to the phloem, analternative strategy is to prepare differential libraries(Vilaine et al., 2003). However, a specific sieve element–companion cell complex function might be inferred fromthe annotation and current models of phloem function (vanBel, 2003a, b). In addition, a small but increasing numberof studies have catalogued a phloem location of transcriptsand proteins; some of these are common to those in thelibraries reported here.
Fig. 2. In situ hybridization of two cDNAs detected in the phloem sap.Sense and antisense probes were generated by PCR amplification ofinternal fragments of peroxiredoxin. DIG-labelled hybrids were detectedwith alkaline phosphatase-conjugated anti-DIG. (a, b) Peroxiredoxinsense and antisense labelling of sections of 8-d-old Ricinus hypocotyls.(c, d) Peroxiredoxin sense and antisense labelling of sections of 6-month-old Ricinus stem. X, xylem; P, phloem; C, cortex.
Phloem cDNAs 3187
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Protein metabolism—ribosome protein messages inthe sieve element
Twenty-two sequences (14%) from the late library hadannotations indicating a role in ribosome function: forexample, homology to 40S ribosomal protein from Glycinemax (Table 2). In a previous study on maize, using lasermicrodissection for sampling vascular tissues, transcript forprotein from a ribosomal subunit was detected (Nakazonoet al., 2003). The presence of a message does not nec-essarily reflect the presence of the corresponding protein.No ribosomal proteins have been detected in sieve elementsap from a range of species including Cucumis sativus(L.) (Walz et al., 2004), Triticum aestivum (L.) (Hayashiet al., 2000), or R. communis using sap collected in anidentical manner to the present study (Barnes et al.,2004). The presence of ribosomal protein message in thesieve element sap is consistent with movement of themessage through plasmodesmata from companion cells.
Protein turnover and degradation
Ubiquitin proteins are frequently found in phloem tissues orsieve element sap (Hayashi et al., 2000; Barnes et al., 2004;Walz et al., 2004). Ubiquitin- and proteasome-dependentproteolysis is required for the regulation of protein turnover(Ingvardsen and Veierskov, 2001). In the phloem, thesesystems have been hypothesized to be involved in proteinstability (Schobert et al., 1995) or potentially in signalling(Vilaine et al., 2003). Most studies examining phloem geneexpression have noted the presence of such gene transcriptsimplicated in protein turnover and degradation (Nakazonoet al., 2003; Vilaine et al., 2003). In the present study, 6%of the messages from the late library could be assigned
a role in protein turnover, including transcripts sharinghomology to a ubiquitin-conjugating protein from Vitispseudoreticulata (L.) (Table 2).
Structural genes and cell wall enzymes
Fourteen of the late library transcripts (9% of the total)had significant homology with genes associated with cellstructure. Broadly, these could be divided into genescoding for extracellular wall components and those in-volved in cytoskeletal formation.
Profilin binds actin, and its transcript was present at 15-fold greater concentrations than actin in Ricinus sieveelement sap (Schobert et al., 1998; Barnes et al., 2004).Profilin has been hypothesized to prevent actin polymeri-zation and therefore to be responsible for the absence ofmicrotubules in sieve element cells (Toth et al., 1994). Inour study, the late library contained a transcript withhomology to an actin-depolymerizing factor from Lyco-persicon esculentum and a potential actin-binding protein,myosin from Cryptosporidium parvum (Table 2). Myosinproteins have previously been located in the sieve elementand have been hypothesized to play a role in alteringthe size exclusion limit of plasmodesmata (Baluska et al.,2001). While intact microfilaments are not believed to bepresent in mature sieve elements (Schobert et al., 1998),three tubulin-related transcripts were detected. Recently, ithas been speculated that actin microtubules are associatedwith the plasmodesmatal pore and may regulate vesiclemovement from companion cell to sieve element (Oparka,2004; Kim, 2005). Message for a synaptobrevin-like pro-tein from rice was identified in the late phloem library(Table 2) and could have a role in vesicle traffickingthrough plasmodesmata. It is interesting to note that thelibrary transcript for the translationally controlled tumourprotein from Hevea brasiliensis (TCTP, Table 2) hasproperties of a tubulin-binding protein (Gachet et al.,1999). In a separate study, the presence of the protein ofthis transcript in Ricinus sieve element sap could be dem-onstrated (Barnes et al., 2004); thus, for this gene, bothmessage and protein appear to be trafficked through theplasmadesmata.
A number of transcripts encoding cell wall proteins werealso found in the Ricinus cDNA late library. These includeda cellulose synthase from Gossypium hirsutum and a UDPglucose dehydrogenase mRNA from Cinnamomum osmo-phile (Table 2) which has previously been localized to thevascular tissue of Phaseolus vulgaris (Robertson et al.,1996). A message annotated as a precursor for a cell wallinvertase from Vitis vinifera was also identified (Table 2);cell wall invertases have been previously localized to potatophloem (Hedley et al., 2000). A number of these structuralgenes have been reported to be induced by stress, illus-trating the difficulty of assigning specific, individual func-tions to genes. For example, a homologue of a chitinase
Fig. 3. Manual annotation of the 158 unique transcripts from the latelibrary into single functional classes. Individual functions were assignedon the basis of the clearest annotation of the most significant BLASTmatch. Where this annotation was unclear, the function of the mostsignificant Arabidopsis match was taken into account. Transcripts areclassed as ‘unknown’ when insufficient information was available tomake a decision. Those unassigned had multiple or unclear functions.
3188 Doering-Saad et al.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
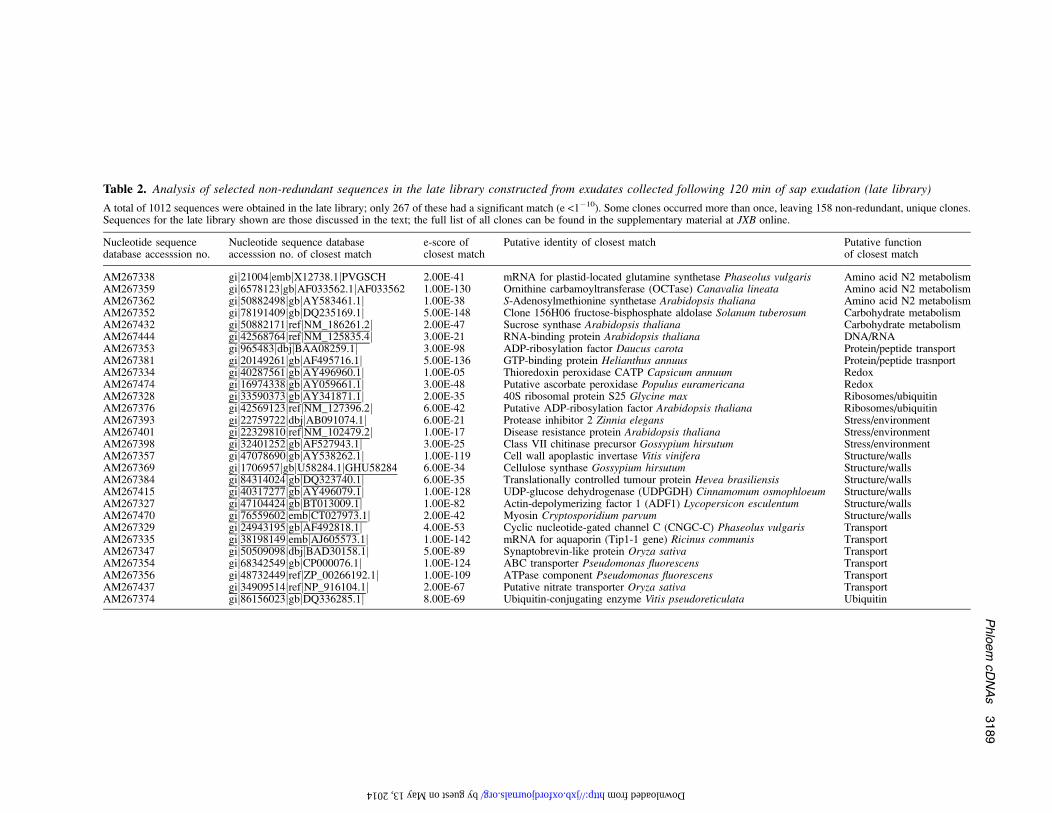
Table 2. Analysis of selected non-redundant sequences in the late library constructed from exudates collected following 120 min of sap exudation (late library)
A total of 1012 sequences were obtained in the late library; only 267 of these had a significant match (e <1�10). Some clones occurred more than once, leaving 158 non-redundant, unique clones.Sequences for the late library shown are those discussed in the text; the full list of all clones can be found in the supplementary material at JXB online.
Nucleotide sequencedatabase accesssion no.
Nucleotide sequence databaseaccesssion no. of closest match
e-score ofclosest match
Putative identity of closest match Putative functionof closest match
AM267338 gij21004jembjX12738.1jPVGSCH 2.00E-41 mRNA for plastid-located glutamine synthetase Phaseolus vulgaris Amino acid N2 metabolismAM267359 gij6578123jgbjAF033562.1jAF033562 1.00E-130 Ornithine carbamoyltransferase (OCTase) Canavalia lineata Amino acid N2 metabolismAM267362 gij50882498jgbjAY583461.1j 1.00E-38 S-Adenosylmethionine synthetase Arabidopsis thaliana Amino acid N2 metabolismAM267352 gij78191409jgbjDQ235169.1j 5.00E-148 Clone 156H06 fructose-bisphosphate aldolase Solanum tuberosum Carbohydrate metabolismAM267432 gij50882171jrefjNM_186261.2j 2.00E-47 Sucrose synthase Arabidopsis thaliana Carbohydrate metabolismAM267444 gij42568764jrefjNM_125835.4j 3.00E-21 RNA-binding protein Arabidopsis thaliana DNA/RNAAM267353 gij965483jdbjjBAA08259.1j 3.00E-98 ADP-ribosylation factor Daucus carota Protein/peptide transportAM267381 gij20149261jgbjAF495716.1j 5.00E-136 GTP-binding protein Helianthus annuus Protein/peptide trasnportAM267334 gij40287561jgbjAY496960.1j 1.00E-05 Thioredoxin peroxidase CATP Capsicum annuum RedoxAM267474 gij16974338jgbjAY059661.1j 3.00E-48 Putative ascorbate peroxidase Populus euramericana RedoxAM267328 gij33590373jgbjAY341871.1j 2.00E-35 40S ribosomal protein S25 Glycine max Ribosomes/ubiquitinAM267376 gij42569123jrefjNM_127396.2j 6.00E-42 Putative ADP-ribosylation factor Arabidopsis thaliana Ribosomes/ubiquitinAM267393 gij22759722jdbjjAB091074.1j 6.00E-21 Protease inhibitor 2 Zinnia elegans Stress/environmentAM267401 gij22329810jrefjNM_102479.2j 1.00E-17 Disease resistance protein Arabidopsis thaliana Stress/environmentAM267398 gij32401252jgbjAF527943.1j 3.00E-25 Class VII chitinase precursor Gossypium hirsutum Stress/environmentAM267357 gij47078690jgbjAY538262.1j 1.00E-119 Cell wall apoplastic invertase Vitis vinifera Structure/wallsAM267369 gij1706957jgbjU58284.1jGHU58284 6.00E-34 Cellulose synthase Gossypium hirsutum Structure/wallsAM267384 gij84314024jgbjDQ323740.1j 6.00E-35 Translationally controlled tumour protein Hevea brasiliensis Structure/wallsAM267415 gij40317277jgbjAY496079.1j 1.00E-128 UDP-glucose dehydrogenase (UDPGDH) Cinnamomum osmophloeum Structure/wallsAM267327 gij47104424jgbjBT013009.1j 1.00E-82 Actin-depolymerizing factor 1 (ADF1) Lycopersicon esculentum Structure/wallsAM267470 gij76559602jembjCT027973.1j 2.00E-42 Myosin Cryptosporidium parvum Structure/wallsAM267329 gij24943195jgbjAF492818.1j 4.00E-53 Cyclic nucleotide-gated channel C (CNGC-C) Phaseolus vulgaris TransportAM267335 gij38198149jembjAJ605573.1j 1.00E-142 mRNA for aquaporin (Tip1-1 gene) Ricinus communis TransportAM267347 gij50509098jdbjjBAD30158.1j 5.00E-89 Synaptobrevin-like protein Oryza sativa TransportAM267354 gij68342549jgbjCP000076.1j 1.00E-124 ABC transporter Pseudomonas fluorescens TransportAM267356 gij48732449jrefjZP_00266192.1j 1.00E-109 ATPase component Pseudomonas fluorescens TransportAM267437 gij34909514jrefjNP_916104.1j 2.00E-67 Putative nitrate transporter Oryza sativa TransportAM267374 gij86156023jgbjDQ336285.1j 8.00E-69 Ubiquitin-conjugating enzyme Vitis pseudoreticulata Ubiquitin
Phloem
cDNAs
3189
by guest on May 13, 2014 http://jxb.oxfordjournals.org/ Downloaded from

precursor from cotton was found in the late library;chitinase precursors are involved in cell wall remodellingand are widely implicated in interactions with variouspathogens (Kasprzewska, 2003). A homologue of thecellulose synthase from the late library was induced inphloem fibres by mechanical stress (Wu et al., 2000).
Interaction with DNA/RNA
Six per cent of transcripts from the late library had anno-tations related to DNA or RNA binding. Of these, twocoded for RNA-binding proteins from Arabidopsis, anexample being listed in Table 2. These may be chaperonesin long-distance transport of other macromolecules (Lucasand Lee, 2004). The remainder possessed annotationsimplying involvement in regulation of transcription, DNAbinding, and RNA processing which might be expectedto occur in companion cells and not in the anucleate sieveelements. However, if mRNA moves through the phloemsap as information, then RNA binding and chaperoningsystems will be important in sieve element function.
Carbohydrate metabolism
The phloem pathway is central to carbohydrate transport,and five cDNAs from the late library encoded proteinswith involvement in carbohydrate metabolism. These in-cluded a message for a sucrose synthase from Arabidopsis(Table 2); a homologue of this protein was previouslyfound to be located in companion cells in maize (Nolteand Koch, 1993) and may be involved in processing phloem-translocated sucrose (Komatsu et al., 2001; Konishi et al.,2004). An aldolase transcript from potato (Table 2) waspresent, and immunolocalization studies have demonstratedthe occurrence of an aldolase protein in cucumber sievetubes and companion cells (Chen et al., 2004).
Redox
In a long-lived, anucleate, yet highly active cell type, suchas the sieve element, oxidative stress is a potential problem(Walz et al., 2002). Four cDNAs encoded products thathave functions related to oxidative stress including anascorbate peroxidase from Populus euramericana and athioredoxin peroxidase from Capsicum annuum (Table 2).Thioredoxin is one of the dominant proteins found in thesieve element and may also be involved in chaperoningproteins through plasmodesmata (Ishiwatari et al., 1998).
Amino acid metabolism
Phloem is important in the translocation of amino acids todeveloping sinks. In addition, the amino acid compositionis considered important in determining the performance ofphloem-feeding herbivores such as aphids and whitefly. Anumber of transcripts from the late library encoded proteinswith roles in amino acid metabolism, including a methio-nine synthetase, a glutamine synthase, and an ornithine
carbamoyltransferase (Table 2). Proteins for the first twotranscripts have previously been detected in phloem sap(Hoffmann-Benning et al., 2002). Interestingly, transcriptsfor glutamine synthase were up-regulated by phloem-feeding aphid infestation in both tobacco leaves (Voelckelet al., 2004) and celery phloem (Divol et al., 2005).
Transport
Given the fundamental role of the phloem in long-distancetransport, it is perhaps surprising that only 6% of late librarytranscripts related to solute or water transport comparedwith 9.2% in the whole Arabidopsis genome. A messagefor an aquaporin from R. communis was found (Table 2), ashas been previously reported in barley sieve element sap(Doering-Saad et al., 2002), and both message and proteinwere localized to spinach phloem (Fraysse et al., 2005).However, aquaporins were not selectively expressed incelery phloem tissue (Vilaine et al., 2003). The late libraryalso contained an ABC transporter and a nitrate transportercDNA, along with a CNGC ion channel (Table 2), perhapstransporting K+. Interestingly, this transporter has beennoted to interact with the Ca2+ calmodulin-binding systempreviously observed in phloem (Hayashi et al., 2000;Barnes et al., 2004). Also in the transporter classification,transcripts for a synaptobrevin-like protein were observed.These proteins are believed to be involved in vesicletrafficking presumably via interaction with plasmodesmatalmembranes (Oparka, 2004). Two transcripts had homologywith components of a membrane ATPase which operatesin the sieve element–companion cell complex (Doering-Saad et al., 2002). A further seven transcripts were anno-tated as being involved in peptide/protein transport. Manyof these may be involved in protein trafficking, includinga homologue of Helianthus annuus GTP-binding protein(RAN) mRNA previously located to companion cells, andan ADP-ribosylation factor from Daucus carota believedto act in cytoskeletal remodelling and protein trafficking(Kiyosue and Shinozaki, 1995).
Interaction with the environment
The phloem may be the site of interaction with manypathogens and is also a potential site of signal transductionduring environmental perturbations. In our study, a tran-script with strong homology to a protease inhibitor tran-script from Zinnia elegans was found in the late library(Table 2). A cysteine-proteinase inhibitor has also beenfound in a protein library from Ricinus phloem (Barneset al., 2004), and a homologue was up-regulated follow-ing aphid infestation in Sorghum (Zhu-Salzman et al.,2004). A transcript for a homologue to an Arabidopsisthaliana disease resistance protein occurred in the latelibrary; a related transcript was found in the phloem ofrice (Asano et al., 2002), and messages for similarly anno-tated proteins were also up-regulated by aphids in Sorghum
3190 Doering-Saad et al.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

(Zhu-Salzman et al., 2004). Proteins annotated as havinga role in disease resistance have been found in phloem byother workers (Hoffmann-Benning et al., 2002) includingan ultraviolet-B-repressible protein from Pisum sativum.
Homology with Arabidopsis
Assigning cDNAs to functions can be criticized as beingsubjective because most sequences fall into more than onefunctional category. To overcome these difficulties and toexploit the unrivalled genetic information available, theclosest homologue to each of the cDNAs from the latelibrary was determined in the Arabidopsis genome. Func-tional categorization of the Arabidopsis genes obtained inthis way could then be determined using the GO annota-tions from the TAIR web site. To facilitate this, theAnnotation Superviewer facility provided by NicholasProvart on the Arabidopsis Functional Genomics Toolson the BBC web site (http://bbc.botany.utoronto.ca/ntools/cgi/bin/ntools_classification_superviewer.cgi) was used(Provart and Zhu, 2003). An advantage of this approachis that the frequency of occurrence of each functionalcategory can be compared with its frequency of occurrencein the whole Arabidopsis genome although caution mustbe exercised in interpretation since the cDNA library will
be biased towards the more abundant messages (Fig. 4).Generally, as might be expected for a library constructedfrom a single cell type, most functional categories for thelate library differed in frequency from those of the wholeArabidopsis genome. Specifically, genes involved in celllocalization, protein synthesis, tissue localization, organlocalization, organ differentiation, and cell fate were over-represented >2-fold compared with the whole Arabidopsisgenome. Those categories for transposable elements andunclassified proteins were under-represented by more thana half.
Conclusions
The transcripts identified in the late phloem library encodefunctions that are consistent with current models of phloemfunction, and consolidate and extend the results of otherstudies examining mRNA (Nakazono et al., 2003; Divolet al., 2005) and protein (Barnes et al., 2004; Walz et al.,2004) occurrence in sieve elements. This information,which is complemented by transcriptional profiling studies,is valuable since it informs models of phloem function.Although little of use can be stated about the transcripts forwhich no annotation is currently available, these could
Fig. 4. Functional categorization of the closest homologues in the Arabidopsis genome of transcripts occurring in the Ricinus phloem sap cDNA library(‘normed’ to frequency of class overall ID numbers in the Arabidopsis genome). Error bars are 99% confidence intervals calculated from standarddeviations derived from bootstrapping 100 times. Annotation was performed using Annotation Superviewer (Provart and Zhu, 2003). The sum of thefunctional frequencies is greater than the number of sequences entered as genes can be categorized in more than one functional class.
Phloem cDNAs 3191
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

eventually be particularly useful in extending our un-derstanding of phloem function. For the cDNAs identifiedhere, reverse genetic approaches can be used in the modelspecies A. thaliana to gain experimental evidence about thespecific functions of their homologues. At its simplest, thisinvolves the use of individuals homozygous for loss-of-function mutations (resulting from insertion of T-DNA ora transposon), followed by detailed physiological analysesto detect possible changes in phloem-related function.
Supplementary data
Supplementary data can be found at JXB online.
Acknowledgements
We wish to thank BBSRC for studentships to CC and Syngentafor support to CD-S. The report was improved by the comments oftwo anonymous referees.
References
Ahlfors R, Lang S, Overmyer K, et al. 2004. Arabidopsis radical-induced cell death1 belongs to the WWE protein–protein in-teraction domain protein family and modulates abscisic acid,ethylene, and methyl jasmonate responses. The Plant Cell 16,1925–1937.
Asano T, Masumura T, Kusano H, Kikuchi S, Kurita A,Shimada H, Kadowaki K. 2002. Construction of aspecialized cDNA library from plant cells isolated by lasercapture microdissection: toward comprehensive analysis of thegenes expressed in the rice phloem. The Plant Journal 32,401–408.
Baluska F, Cvrckova F, Kendrick-Jones J, Volkmann D. 2001.Sink plasmodesmata as gateways for phloem unloading. MyosinVIII and calreticulin as molecular determinants of sink strength?Plant Physiology 126, 39–46.
Barnes A, Bale J, Constantinidou C, Ashton P, Jones A,Pritchard J. 2004. Determining protein identity from sieveelement sap in Ricinus communis L. by quadrupole time of flight(Q-TOF) mass spectrometry. Journal of Experimental Botany 55,1473–1481.
Chen ZH, Walker RP, Tecsi LI, Lea PJ, Leegood RC. 2004.Phosphoenolpyruvate carboxykinase in cucumber plants is in-creased both by ammonium and by acidification, and is present inthe phloem. Planta 219, 48–58.
Chisholm ST, Parra MA, Anderberg RJ, Carrington JC. 2001.Arabidopsis RTM1 and RTM2 genes function in phloem to restrictlong-distance movement of tobacco etch virus. Plant Physiology127, 1667–1675.
Divol F, Vilaine F, Thibivilliers S, Amselem J, Palauqui JC,Kusiak C, Dinant S. 2005. Systemic response to aphid infestationby Myzus persicae in the phloem of Apium graveolens. PlantMolecular Biology 57, 517–540.
Doering-Saad C, Newbury HJ, Bale JS, Pritchard J. 2002. Useof aphid stylectomy and RT–PCR for the detection of trans-porter mRNAs in sieve elements. Journal of ExperimentalBotany 53, 631–637.
Fraysse LC, Wells B, McCann MC, Kjellbom P. 2005. Specificplasma membrane aquaporins of the PIP1 subfamily are expressedin sieve elements and guard cells. Biology of the Cell 97, 519–534.
Gachet Y, Tournier S, Lee M, Lazaris-Karatzas A, Poulton T,Bommer UA. 1999. The growth-related, translationally controlledprotein P23 has properties of a tubulin binding protein andassociates transiently with microtubules during the cell cycle.Journal of Cell Science 112, 1257–1271.
Gould N, Thorpe MR, Minchin PEH, Pritchard J, White PJ.2004. Solute is imported to elongating root cells of barley asa pressure driven flow of solution. Functional Plant Biology 31,391–397.
Hayashi H, Fukuda A, Suzui N, Fujimaki S. 2000. Proteins in thesieve element–companion cell complexes: their detection, local-ization and possible functions. Australian Journal of PlantPhysiology 27, 489–496.
Haywood V, Yu TS, Huang NC, Lucas WJ. 2005. Phloem long-distance trafficking of gibberellic acid-insensitive RNA regulatesleaf development. The Plant Journal 42, 49–68.
Hedley PE, Maddison AL, Davidson D, Machray GC. 2000.Differential expression of invertase genes in internal and externalphloem tissues of potato (Solanum tuberosum L.). Journal ofExperimental Botany 51, 817–821.
Hoffmann-Benning S, Gage DA, McIntosh L, Kende H,Zeevaart JAD. 2002. Comparison of peptides in the phloem sapof flowering and non-flowering Perilla and lupine plants usingmicro-bore HPLC followed by matrix-assisted laser desorption/ionizationtime-of-flight mass spectrometry. Planta 216, 140–147.
Huang T, Bohlenius H, Eriksson S, Parcy F, Nilsson O. 2005. ThemRNA of the Arabidopsis gene FT moves from leaf to shoot apexand induces flowering. Science 309, 1694–1696.
Ingvardsen C, Veierskov B. 2001. Ubiquitin- and proteasome-dependent proteolysis in plants. Physiologia Plantarum 112,451–459.
Ishiwatari Y, Fujiwara T, McFarland KC, Nemoto K,Hayashi H, Chino M, Lucas WJ. 1998. Rice phloem thiore-doxin h has the capacity to mediate its own cell-to-cell transportthrough plasmodesmata. Planta 205, 12–22.
Jackson D. 1991. In situ hybridisation in plants. In: Gurr SJ,McPherson MJ, Bowles DJ, eds. Molecular plant pathology,a practical approach, Vol. 1. Oxford: Oxford University Press,163–174.
Kasprzewska A. 2003. Plant chitinases—regulation and function.Cell Molecular Biology Letters 8, 809–824.
Kim JY. 2005. Regulation of short-distance transport of RNA andprotein. Current Opinion in Plant Biology 8, 45–52.
Kiyosue T, Shinozaki K. 1995. Cloning of a carrot cDNA fora member of the family of ADP-ribosylation factors (ARFs) andcharacterization of the binding of nucleotides by its product afterexpression in Escherichia coli. Plant and Cell Physiology 36,849–856.
Knoblauch M, van Bel AJE. 1998. Sieve tubes in action. The PlantCell 10, 35–50.
Komatsu A, Moriguchi T, Koyama K, OmuraM, Akihama T.2001. Analysis of sucrose synthase genes in citrus suggestsdifferent roles and phylogenetic relationships. Journal of Exper-imental Botany 53, 61–71.
Konishi T, Ohmiya Y, Hayashi T. 2004. Evidence that sucroseloaded into the phloem of a poplar leaf is used directly by sucrosesynthase associated with various b-glucan synthases in the stem.Plant Physiology 134, 1146–1152.
Komor E. 2000. Source physiology and assimilate transport: theinteraction of sucrose metabolism, starch storage and phloemexport in source leaves and the effects on sugar status in phloem.Australian Journal of Plant Physiology 27, 497–505.
3192 Doering-Saad et al.
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from

Lucas WJ, Lee JY. 2004. Plant cell biology—plasmodesmata asa supracellular control network in plants. Nature Reviews Molec-ular Cell Bioliogy 5, 712–726.
Milburn JA. 1970. Phloem exudation from castor bean: induction by
massage. Planta 95, 272–276.Morsomme P, Boutry M. 2000. The plant plasma membrane
H+-ATPase: structure, function and regulation. Biochimica etBiophysica Acta 1465, 1–16.
Nakazono M, Qiu F, Borsuk LA, Schnable PS. 2003. Laser-
capture microdissection, a tool for the global analysis of geneexpression in specific plant cell types: identification of genes
expressed differentially in epidermal cells or vascular tissues ofmaize. The Plant Cell 15, 583–596.
Nolte KD, Koch KE. 1993. Companion-cell specific localizationof sucrose synthase in zones of phloem loading and unloading.
Plant Physiology 101, 899–905.Oparka KJ. 2004. Getting the message across: how do plant cellsexchange macromolecular complexes? Trends in Plant Science 9,33–41.
Overmyer K, Brosche M, Pellinen R, Kuittinen T, Tuominen H,Ahlfors R, Keinanen M, Saarma M, Scheel D, Kangasjarvi J.2005. Ozone-induced programmed cell death in the Arabidopsis
radical-induced cell death1 mutant. Plant Physiology 137,1092–1104.
Pritchard J. 1996. Aphid stylectomy reveals an osmotic stepbetween sieve element and cortical cells in barley roots. Journalof Experimental Botany 47, 1519–1524.
Provart N, Zhu T. 2003. A browser-based functional classificationSuperViewer for Arabidopsis genomics. Currents in Computa-tional Molecular Biology 2003, 271–272.
Robertson D, Smith C, Bolwell GP. 1996. Inducible UDP-glucosedehydrogenase from French Bean (Phaseolus vulgaris L.) locatesto vascular tissue and has alcohol dehydrogenase activity. Bio-chemistry 313, 311–317.
Rouhier N, Gelhaye E, Sautiere PE, Brun A, Laurent P,Tagu D, Gerard J, de Fay E, Meyer Y, Jacquot JP. 2001.Isolation and characterization of a new peroxiredoxin frompoplar sieve tubes that uses either glutaredoxin or thioredoxin
as a proton donor. Plant Physiology 127, 1299–1309.Ruiz-Medrano R, Xoconostle-Cazares B, Lucas WJ. 1999.
Phloem long-distance transport of CmNACP mRNA: implicationsfor supracellular regulation in plants.Development 126, 4405–4419.
Sasaki T, Chino M, Hayashi H, Fujiwara T. 1998. Detection ofseveral mRNA species in rice phloem sap. Plant and CellPhysiology 39, 895–897.
Schobert C, Baker L, Szederkenyi J, Grossmann P, Komor E,Hayashi H, Chino M, Lucas WJ. 1998. Identification of
immunologically related proteins in sieve-tube exudate collectedfrom monocotyledonous and dicotyledonous plants. Planta 206,245–252.
Schobert C, Gottschalk M, Kovar DR, Staiger CJ, Yoo BC,Lucas WJ. 2000. Characterization of Ricinus communis phloemprofilin, RcPRO1. Plant Molecular Biology 42, 719–730.
Schobert C, Grossmann P, Gottschalk M, Komor E,Pecsvaradi A, Zurnieden U. 1995. Sieve-tube exudate fromRicinus communis L. seedlings contains ubiquitin and chape-rones. Planta 196, 205–210.
Shirsat AH, Thomson HEC, Elliott KA. 2003. The Brassicanapus extA extensin gene negative regulatory region controlsexpression in response to mechanical stresses. Plant, Cell andEnvironment 26, 1647–1655.
Smith JAC, Milburn JA. 1980. Osmoregulation and the controlof phloem-sap composition in Ricinus communis l. Planta 148,28–34.
Toth KF, Wang Q, Sjolund RD. 1994. Monoclonal-antibodiesagainst phloem p-protein from plant-tissue cultures. 1. Microscopyand biochemical analysis. American Journal of Botany 81,1370–1377.
van Bel AJE. 2003a. The phloem, a miracle of ingenuity. Plant, Celland Environment 26, 125–149.
van Bel AJE. 2003b. Transport phloem: low profile, high impact.Plant Physiology 131, 1509–1510.
van Bel AJE, Gaupels F. 2004. Pathogen-induced resistance andalarm signals in the phloem. Molecular Plant Pathology 5,495–504.
Vilaine F, Palauqui JC, Amselem J, Kusiak C, Lemoine R,Dinant S. 2003. Towards deciphering phloem: a transcriptomeanalysis of the phloem of Apium graveolens. The Plant Journal36, 67–81.
Voelckel C, Weisser WW, Baldwin IT. 2004. An analysis ofplant–aphid interactions by different microarray hybridizationstrategies. Molecular Ecology 13, 3187–3195.
Walz C, Giavalisco P, Schad M, Juenger M, Klose J, Kehr J.2004. Proteomics of curcurbit phloem exudate reveals a network ofdefence proteins. Phytochemistry 65, 1795–1804.
Walz C, Juenger M, Schad M, Kehr J. 2002. Evidence for thepresence and activity of a complete antioxidant defence system inmature sieve tubes. The Plant Journal 31, 189–197.
Williams LE, Lemoine R, Sauer N. 2000. Sugar transporters inhigher plants—a diversity of roles and complex regulation.Trends in Plant Science 5, 283–290.
Wu LG, Joshi SP, Chiang VL. 2000. A xylem-specific cellulosesynthase gene from aspen (Populus tremuloides) is responsive tomechanical stress. The Plant Journal 22, 495–502.
Xoconostle-Cazares B, Yu X, Ruiz-Medrano R, Wang HL,Monzer J, Yoo BC, McFarland KC, Franceschi VR, LucasWJ. 1999. Plant paralog to viral movement protein that potentiatestransport of mRNA into the phloem. Science 283, 94–98.
Zhu-Salzman K, Salzman RA, Ahn JE, Koiwa H. 2004.Transcriptional regulation of sorghum defense determinantsagainst a phloem-feeding aphid. Plant Physiology 134,420–431.
Phloem cDNAs 3193
by guest on May 13, 2014
http://jxb.oxfordjournals.org/D
ownloaded from
Copyright © 2022 FDOKUMEN




















