
Synthetic peptides as putative therapeutic agents in transplantation medicine: application of...
13
Journal of Immunological Methods 241 (2000) 83–95 www.elsevier.nl / locate / jim Synthetic peptides as putative therapeutic agents in transplantation medicine: application of PEPSCAN to the identification of functional sequences in the extracellular domain of the interleukin-2 receptor beta chain (IL-2Rb) a b c c Helen Dionyssopoulou , Athanasia Mouzaki , Jerry Slootstra , Wouter Puijk , c a a, * Rob Meloen , Paul Cordopatis , Georgia Sotiropoulou a Laboratory of Pharmacognosy and Chemistry of Natural Products, Department of Pharmacy, School of Health Sciences, University of Patras, Patras GR-26500, Greece b Laboratory of Experimental Hematology and Transfusion Medicine, Department of Medicine, School of Health Sciences, University of Patras, Patras GR-26500, Greece c Department of Molecular Recognition, Institute for Animal Science and Health ( ID-DLO), P .O. Box 65, 8200 AB Lelystad, The Netherlands Received 6 January 2000; received in revised form 5 April 2000; accepted 26 April 2000 Abstract A desired treatment strategy in transplantation medicine is the selective targeting of alloreactive T cells without impairing antileukemic and antiviral activities. One approach is the synthesis of peptides that interfere with the binding of interleukin-2 (IL-2) to its high affinity receptor (IL-2R). This blocks the activation and proliferation of the antigen-activated T cells and the secretion of IL-2. The latter binds to its receptor, via the extracellular domain of the IL-2Rb chain, while its cytoplasmic domain is required for intracellular signal transduction. In this study, the PEPSCAN method was applied in order to identify antigenic sequences (epitopes) in the extracellular domain of the IL-2Rb. Based on the primary amino acid (aa) sequence of the IL-2Rb, a total of 239 overlapping dodecapeptides, spanning the entire sequence of IL-2Rb, were synthesized by PEPSCAN and their immunoreactivity was tested by ELISA using monoclonal antibodies (mAbs) specific for IL-2Rb such 85 96 as TU11, Mikb1, HuMikb1 and TU27. TU11 recognized a linear epitope located in the region R–Q . None of the 239 synthetic peptides was recognized by TU27. Mikb1 (and HuMikb1) recognized a discontinuous epitope formed by aa 106 148 170 202 located in the IL-2Rb domains L to P and E to A . Subsequently, synthetic peptides corresponding to the identified putative epitopic sequences were prepared by solid phase synthesis and their immunogenicity in vivo was assessed by raising polyclonal antibodies. Given that Mikb1 and HuMikb1 inhibit binding of IL-2 on the IL-2Rb, we addressed the question of whether the identified antigenic sequences serve as putative IL-2 binding domains. Synthetic peptides corresponding to these sequences were tested for their ability to compete with IL-2 for binding and, thereby, inhibit Abbreviations: aa, amino acid(s); ELISA, enzyme-linked immunosorbent assay; IL-2, interleukin-2; IL-2Ra, interleukin-2 receptor alpha chain; IL-2Rb, interleukin-2 receptor beta chain; IL-2Rg, interleukin-2 receptor gamma chain; IL-15, interleukin-15; mAb, monoclonal antibody; PBL, peripheral blood lymphocytes; rIL-2Rb, recombinant interleukin-2 receptor beta chain *Corresponding author. Tel.: 130-61-997-721; fax: 130-61-997-714. E-mail address: [email protected] (G. Sotiropoulou) 0022-1759 / 00 / $ – see front matter 2000 Elsevier Science B.V. All rights reserved. PII: S0022-1759(00)00212-X
Transcript of Synthetic peptides as putative therapeutic agents in transplantation medicine: application of...

Journal of Immunological Methods 241 (2000) 83–95www.elsevier.nl / locate / jim
Synthetic peptides as putative therapeutic agents intransplantation medicine: application of PEPSCAN to theidentification of functional sequences in the extracellulardomain of the interleukin-2 receptor beta chain (IL-2Rb)
a b c cHelen Dionyssopoulou , Athanasia Mouzaki , Jerry Slootstra , Wouter Puijk ,c a a ,*Rob Meloen , Paul Cordopatis , Georgia Sotiropoulou
aLaboratory of Pharmacognosy and Chemistry of Natural Products, Department of Pharmacy, School of Health Sciences,University of Patras, Patras GR-26500, Greece
bLaboratory of Experimental Hematology and Transfusion Medicine, Department of Medicine, School of Health Sciences,University of Patras, Patras GR-26500, Greece
cDepartment of Molecular Recognition, Institute for Animal Science and Health (ID-DLO), P.O. Box 65, 8200 AB Lelystad,The Netherlands
Received 6 January 2000; received in revised form 5 April 2000; accepted 26 April 2000
Abstract
A desired treatment strategy in transplantation medicine is the selective targeting of alloreactive T cells without impairingantileukemic and antiviral activities. One approach is the synthesis of peptides that interfere with the binding of interleukin-2(IL-2) to its high affinity receptor (IL-2R). This blocks the activation and proliferation of the antigen-activated T cells andthe secretion of IL-2. The latter binds to its receptor, via the extracellular domain of the IL-2Rb chain, while its cytoplasmicdomain is required for intracellular signal transduction. In this study, the PEPSCAN method was applied in order to identifyantigenic sequences (epitopes) in the extracellular domain of the IL-2Rb. Based on the primary amino acid (aa) sequence ofthe IL-2Rb, a total of 239 overlapping dodecapeptides, spanning the entire sequence of IL-2Rb, were synthesized byPEPSCAN and their immunoreactivity was tested by ELISA using monoclonal antibodies (mAbs) specific for IL-2Rb such
85 96as TU11, Mikb1, HuMikb1 and TU27. TU11 recognized a linear epitope located in the region R–Q . None of the 239synthetic peptides was recognized by TU27. Mikb1 (and HuMikb1) recognized a discontinuous epitope formed by aa
106 148 170 202located in the IL-2Rb domains L to P and E to A . Subsequently, synthetic peptides corresponding to theidentified putative epitopic sequences were prepared by solid phase synthesis and their immunogenicity in vivo was assessedby raising polyclonal antibodies. Given that Mikb1 and HuMikb1 inhibit binding of IL-2 on the IL-2Rb, we addressed thequestion of whether the identified antigenic sequences serve as putative IL-2 binding domains. Synthetic peptidescorresponding to these sequences were tested for their ability to compete with IL-2 for binding and, thereby, inhibit
Abbreviations: aa, amino acid(s); ELISA, enzyme-linked immunosorbent assay; IL-2, interleukin-2; IL-2Ra, interleukin-2 receptor alphachain; IL-2Rb, interleukin-2 receptor beta chain; IL-2Rg, interleukin-2 receptor gamma chain; IL-15, interleukin-15; mAb, monoclonalantibody; PBL, peripheral blood lymphocytes; rIL-2Rb, recombinant interleukin-2 receptor beta chain*Corresponding author. Tel.: 130-61-997-721; fax: 130-61-997-714.E-mail address: [email protected] (G. Sotiropoulou)
0022-1759/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0022-1759( 00 )00212-X

84 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
107 118 178 199IL-2-induced proliferation of mitogen-stimulated human peripheral blood T cells. Sequences M–E and Y–Qprobably represent functional IL-2 binding domains on IL-2Rb, since these synthetic peptides significantly inhibited theproliferation of activated T cells and secretion of IL-2. 2000 Elsevier Science B.V. All rights reserved.
Keywords: Epitope mapping; PEPSCAN; Interleukin-2 receptor beta chain; Peptides
1. Introduction involved have not been identified. Although IL-2Rb
plays a critical role in IL-2-induced intracellularsignal transduction, the residues that are involved inIL-2 is a major regulatory cytokine of the humanIL-2 binding have not, as yet, been identified.immune system (Hatakeyama and Taniguchi, 1991;
Both IL-2Rb and IL-2Rg belong to the largeSmith, 1992) and, in healthy individuals, its expres-superfamily of class I cytokine receptors (Leonard,sion is restricted to the Th1 subset of T helper1996). Members of this family share characteristiclymphocytes (Mosmann, 1996). Binding of IL-2 tofunctional motifs on their extracellular domains.
its receptor IL-2R on the cell surface initiates signalThese include four conserved site-specific cysteine
transduction leading to lymphocyte proliferationresidues and a membrane proximal ‘WSXWS’ motif.
(Waldmann, 1989; Taniguchi and Minami, 1993). This motif is predicted to be located on the floor ofIL-2R exists in low-, intermediate- and high-affinity the ligand binding site (Bazan, 1990). Single aaforms (Sugamura et al., 1995; Hanisch and Quiron, substitutions inside the ‘WSXWS’ motif block rec-1996; Leonard, 1996). IL-2Ra binds IL-2 with low ognition of the anti-IL-2Rb mAb Mikb1 which27affinity (K 510 M) and no signal can be trans-d inhibits IL-2 binding (Miyasaki et al., 1991). Crystalduced by this chain alone. IL-2Rb binds IL-2 with structures of complexes of class I receptors such asintermediate affinity and activates intracellular signal the human growth hormone–human growth hormone
210transduction. The high-affinity IL-2R (K 510 M)d receptor (DeVos et al., 1992), the human growthis a non-covalent complex of at least three trans- hormone–prolactin receptor (Somers et al., 1994),membrane proteins: IL-2Ra (p55, CD25, Tac), IL- and the erythropoietin agonist peptide–erythropoietin2Rb (p75, CD122) and IL-2Rg (p64, CD132, g ).c receptor (Livnah et al., 1996), revealed that theOn some cells such as natural killer (NK) cells, the WSXWS motif does not appear to play any role inIL-2Rb and IL-2Rg chains combine in the absence ligand binding or receptor–receptor interactions. Inof IL-2Ra to form fully functional receptor complex- contrast, it was demonstrated that analogs of IL-2Rb
29 132 133 134es with intermediate affinity (K 510 M). The with substitutions in S , H and Y do not bindd
IL-2Rg is also utilized by other cytokines, such as IL-2 (Imler et al., 1992). Therefore, the functional194 198IL-15, IL-4, IL-7, and IL-9 (Kondo et al., 1993; significance of the ‘ WSXWS ’ motif is still
Noguchi et al., 1993; Kimura et al., 1994). IL-15 is controversial. In addition, epitopes have been definedthe only cytokine other than IL-2 that shares the on the extracellular domain of IL-2Rb (Francois etIL-2Rb chain (Giri et al., 1994). al., 1995) by means of competition experiments
The identification of sequences on IL-2R subunits employing mAbs against the IL-2Rb, which competewhich are involved in binding IL-2 accompanied by for binding with IL-2, so they probably delineateintracellular signal transduction is essential to the domains on the IL-2Rb which participate in IL-2therapeutic manipulation of the IL-2 / IL-2R system. binding. However, the aa sequences of these an-Several putative functional segments (residues 1–6, tigenic determinants have not been defined.35–43 and 158–160) have been identified on the In this study, the PEPSCAN method (Geysen etIL-2Ra by site-directed mutagenesis where binding al., 1984; Meloen et al., 1991) was used in anof IL-2 was particularly sensitive to aa substitutions attempt to identify putative antigenic sequences in(Cullen et al., 1988; Robb et al., 1988). There is the extracellular domain of IL-2Rb. PEPSCAN isevidence of direct interaction between IL-2Rg and suitable for epitope mapping studies because itIL-2 (Voss et al., 1993) but the actual sequences allows the simultaneous synthesis on solid supports

H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 85
of hundreds of overlapping peptides (up to 12 aa in overnight at 48C. After washing three times withlength). The immunoreactivity of PEPSCAN syn- 0.05% Tween-80 in PBS for 10 min per wash,thetic peptides can, subsequently, be tested against peptides were incubated with rabbit anti-mousemultiple polyclonal or monoclonal Abs raised against peroxidase (rampo) (1 /1000) (Dako, Glostrup, Den-the entire protein. Recently, PEPSCAN was used for mark) for 1 h at 258C, excess secondary Ab wasthe identification of a functionally important se- removed and peroxidase substrate was added (2,29-quence on the b1-adrenoreceptor (Schneider et al., azino-di-3-ethyl-benzthiazoline sulfonate (ABTS),1998). Based on the sequence of the PEPSCAN 3% H O ) and allowed to react for 1 h. Absorbance2 2
peptide, novel peptidomimetics were designed, was measured at 405 nm with a multiscan ELISAwhich fully prevented the positive chronotropic plate reader (Titertek, Flow Laboratories, McLean,effect of anti-b1-adrenoreceptor autoantibodies from VA). After each assay, bound Abs were thoroughlythe serum of patients with dilated cardiomyopathy. removed after 1 h sonication in a heated (708C) bathIn addition, a modified version of PEPSCAN has solution of 1% sodium dodecylsulfate and 0.1%been used in a study of the interleukin-6 receptor 2-mercaptoethanol in PBS, according to an estab-(Halimi et al., 1995). lished protocol (Geysen et al., 1984). Pin-peptides
In the present study, a total of 239 overlapping were ready to be re-used after washing three timesdodecapeptides were synthesized by PEPSCAN and with 0.05% Tween-80 in PBS and once with distilledtheir immunoreactivity was tested against several H O for 10 min per wash. Peptides were considered2
well-characterized monoclonal antibodies, which positively immunoreactive when ELISA absorbancespecifically recognize the IL-2Rb. In order to char- values were greater than three times the backgroundacterize functional antigenic sequences of the IL- (the mean of the lowest 25% of the absorbance of all2Rb, the classical PEPSCAN (Geysen et al., 1984) 239 dodecapeptides in the scan, i.e. the lowest 60was modified and used in combination with a absorbances) (Geysen et al., 1987). MAb TU27 wasradiolabeled ligand in vitro binding assay, the pro- purchased from Becton Dickinson, USA and MiKB1
duction of antibodies, and a proliferation assay of from CLB, The Netherlandsactivated human T cells. Two putative functionaldomains were identified on the IL-2Rb which com- 2.3. Production of polyclonal serapete with IL-2 for binding and significantly inhibitmitogen-induced proliferation of normal peripheral The following peptides were used for immuniza-
106 124 114blood T cells and secretion of IL-2. tions: LMAPISLQVVHVETHRCNI , VVHV-129 126 140ETHRCNISWEIS , WEISQASHYFERHLE ,
178 199YEFQVRVKPLQGEFTTWSPWSQ . Peptides2. Materials and methods were synthesized stepwise on phenyl–acetomethyl
(PAM) resin (Novabiochem, Switzerland) according2.1. Synthesis of rod-peptides to the Merrifield solid-phase method (Steward and
Young, 1984) and the classical t-butyloxycarbonyl /126 140Support-bound dodecapeptides were synthesized benzyl (tBoc/Bzl) chemistry. Peptides W–E
178 199as described previously (Geysen et al., 1984). Pep- and Y–Q were synthesized with a cysteine attides were synthesized on the polyethylene pins the N-terminus for coupling with keyhole limpetwhich form part of the holders that fit into 96-well haemocyanin (KLH). Then, peptides were cleavedmicrotitre plates (Greiner). from the polymer support with 10% HF and purified
by gel filtration on Sephadex G-25 with 2 M acetic2.2. ELISA on pins acid as eluent (Plaue and Briand, 1988) and HPLC
according to standard protocols (Walker, 1994).For ELISA procedures on pins, mAbs were dis- Female rabbits were immunized with peptides (1
solved in phosphate-buffer (pH 7.2) containing 5% mg/rabbit) conjugated to KLH and administeredovalbumin, 5% horse-serum and 1% Tween-80 to subcutaneously and intramuscularly (Langeveld etblock non-specific binding. mAbs were incubated al., 1994). Six weeks later the immunized animals

86 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
125were boosted. Freund’s complete and incomplete to remove unbound I-IL-2. Radioactivity boundadjuvants (Difco Laboratories, Michigan, USA) were on rods was measured in a g-counter (Riastar,used in the immunization protocol as described Hewlett Packard). All steps were carried out at roomelsewhere (Langeveld et al., 1994). Pre-immune sera temperature. Bound radioactivity was removed fromwere collected prior to the immunizations. Eight- rods by sonication for 1 h in a water-bath with 0.1 Mweek antisera were collected for testing 2 weeks NaH PO , 1% SDS and 0.1% 2-mercaptoethanol at2 4
after administration of the booster. 608C. Subsequently, rods were washed twice withH O, once with methanol and air-dried. Residual2
2.4. ELISA radioactivity on rods was measured (|180–500 cpm/rod) and subtracted from the total bound radioactivi-
Different dilutions of recombinant IL-2Rb ex- ty. Each peptide was synthesized in duplicate. Re-tracellular domain (rIL-2Rb ectodomain) (Sana et sults represent the average of two independental., 1994) in PBS (final amounts ranged from 0.06 to experiments. Reagents were purchased from Sigma.3.85 mg), were added to 96-well microtest plates(Nunc-Immuno plates maxisorp). Plates were left at 2.6. Cellular proliferation and IL-2 secretion48C overnight, washed three times with PBS con- assaystaining 0.1% BSA and 0.1% Tween-20 and un-occupied sites blocked for 2 h at 378C with PBS Human peripheral blood lymphocytes (PBL) werecontaining 1% BSA and 0.1% Tween-20. Then, rIL- isolated from the venous blood of adult donors2Rb ectodomain was incubated for 2 h at 378C with (Transfusion Centre, Patras University Hospital) bya 1/1000 dilution of the first antibody (antiserum Ficoll /Hypaque fractionation (Herzberg and Smith,against synthetic peptides, pre-immune serum or 1987). PBL were cultured in RPMI 1640 cultureTU11). The plates were washed at room temperature, medium (CM) containing 10% fetal bovine serumthree times with 0.05% Tween-80 in PBS, and once and antibiotics as described (Mouzaki et al., 1993).with H O (for 10 min per wash). Then, the plates2 Cell viability was assessed by Trypan-blue exclusionwere incubated for 2 h at 258C with 1/2000 goat (Mouzaki et al., 1993) and was always .90%.anti-rabbit peroxidase (or goat anti-mouse peroxi- Triplicate samples were measured in 5-ml RIA vials
6dase) (Sigma). After washing, peroxidase substrate (1310 cells /ml) followed by addition of the indi-178 199 126 140 107 118ABTS and 2 ml 3% H O were added. After 45 min,2 2 cated peptide ( Y–Q , W–E , M–E ,
85 96the absorbance was measured at 405 nm with a R–Q and a control peptide, RREL-multiscan ELISA plate reader (Titertek, Flow Lab- FRQSLSQRETALV) over a concentration rangeoratories, McLean, VA). For competition experiments from 0.1 nM to 1000 nM with or without theantibodies were pre-incubated overnight at 48C with mitogen phytohaemagglutinin (PHA, 5 mg/ml).a 1000-fold excess of synthetic peptide. Final volumes were adjusted to 1 ml /vial with CM.
Cells were incubated for 2 days in a CO incubator125 22.5. In vitro binding of IL-2 and counted by an automated cell counter (SysmexSE-9500). The supernatants were collected and
The selected 52 immunoreactive rod-attached assayed for IL-2 concentration using the Quantikinedodecapeptides were tested for in vitro binding of IL-2 ELISA kit (R&D Systems). Samples were run125I-IL-2. Each rod-peptide was preincubated with in triplicate, in three independent experiments.200 ml of PBS containing 0.05% Tween 80, 0.5%ovalbumin and 0.5% horse serum overnight to blocknon-specific adsorption of protein. Then, rods were
125 3. Resultsincubated with 175 ml of 250 pM I-IL-2 (Amer-sham, specific activity 670 Ci /mmol) in the same
125solution overnight. When indicated, I-IL-2 was 3.1. Scanningmixed with a 200-fold excess of cold IL-2(Proleukin, Roche). Subsequently, rods were exten- Synthetic rod-attached peptides were used to mapsively washed with PBS containing 0.05% Tween-80 the antigenic sites on the extracellular domain of the

H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 87
IL-2Rb chain. By means of the PEPSCAN method, for their ability to recognize specifically the entireall possible 239 overlapping dodecapeptides (in steps IL-2Rb extracellular domain. Binding of antisera toof one aa residue) that can be derived from the the IL-2Rb was estimated by the absorbance mea-primary sequence, were synthesized. mAbs TU27 sured in a standard ELISA procedure in which(Takeshita et al., 1989), Mikb1 (Tsudo et al., 1989), various concentrations of the recombinant IL-2Rb
HuMikb1 (Hakimi et al., 1993), TU11 (Suzuki et al., extracellular domain (rIL-2Rb ectodomain) were1989) were tested against the synthetic rod-peptides. used to coat microtitre plates. Two of the polyclonal
126 140Peptides were considered to represent antigenic sites Abs, anti- W–E (rabbit E, Table 1) and anti-178 199if peaks occurred in a set of neighbouring peptides Y–Q (rabbit G, Table 1), exhibited considera-
and if at least one of the peaks in such a set bly higher binding affinity for rIL-2Rb than didamounted to more than three times the background. TU11, which is a specific and well-characterizedEach vertical bar in Fig. 1 represents the absorbance anti-IL-2Rb mAb (Fig. 2). Specificity was confirmedof the substrate solution. The TU11 mAb recognized in the same ELISA experiment by incubation ofa linear epitope, as shown by its binding to several rIL-2Rb ectodomain with pre-immune sera, whichneighbouring dodecapeptides. It was concluded that resulted in practically no signal (Fig. 2). Similarly,the epitope recognized by TU11 lies within residues the addition of excessive amounts of synthetic85 96 126 140REGVRWRVMAIQ (Fig. 1A). For Mikb1 mAb peptides used for immunization ( W–E and
178 199two major putative Ab-binding domains were iden- Y–Q , respectively) resulted in dramatically106 148 170 202tified: L–P and E–A (Fig. 1B). Inside reduced absorbance values (Fig. 2). Finally, the
126 140 178 199these sequences, Mikb1 binds to many non-neigh- specificity of anti- W–E and anti- Y–Qbouring dodecapeptides, although with low affinity. Abs was confirmed by the fact that they did notThe peaks depicted in Fig. 1B correspond to pep- recognize the rIL-2Rg ectodomain (data not shown).
113 124 126tides: QVVHVETHRCNI , WEISQASHYFE-140 137 148 172RHLE , RHLEFEARTLSP , LTPDTQY- 1253.4. In vitro binding of I-IL-2183 191 202EFQVR , and FTTWSPWSQPLA . As ex-
pected, HuMikb1 showed similar specificity (dataSince Mikb1 (and HuMikb1) completely inhibit
not shown). TU27 probably binds to a nonlinearthe binding of IL-2 to the b chain, we tested whether
epitope which could not be detected, since nothe epitopic sequences identified for these mAbs bind
immunoreactivity above the background value wasIL-2 in vitro. A total of 52 peptides were resynthes-
observed with any of 239 rod peptides tested (Fig.ized on rods in duplicate and tested for their ability
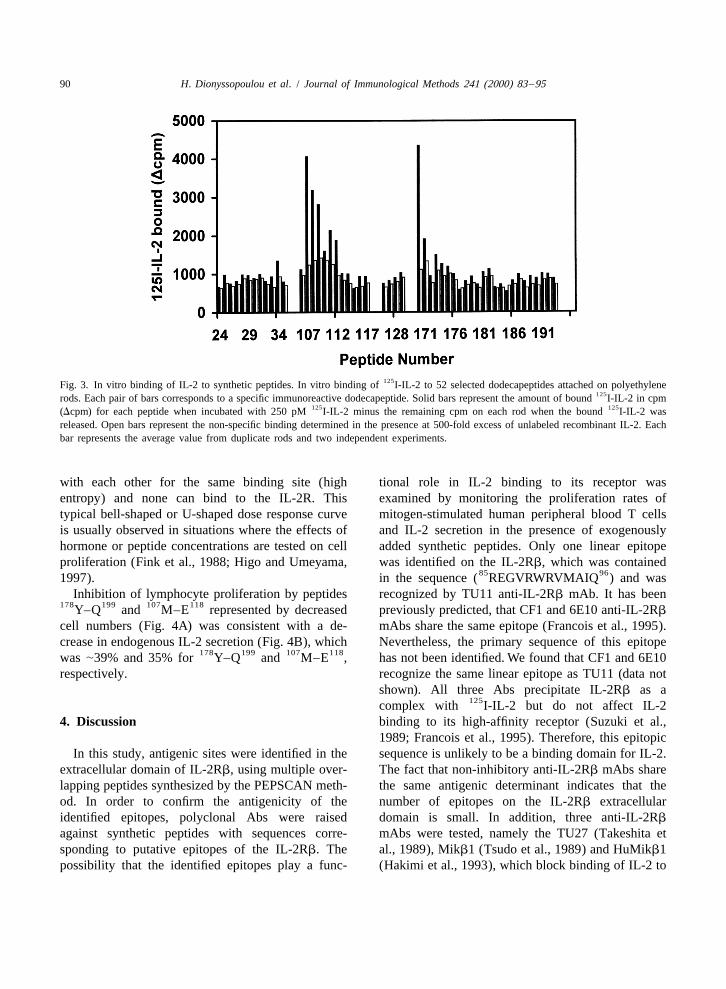
1C). 125to bind I-IL-2 in a one step in vitro assay (Tzartosand Remoundos, 1990). Peptides homologous to3.2. Antigenicity of the identified epitopesPEPSCAN immunoreactive sequences were selected.Peptides that gave no signal in PEPSCAN mapping
The antigenicity of the domains identified bywere included as negative controls. As shown in Fig.
PEPSCAN mapping as being recognized by Mikb1 107 1183 dodecapeptides MAPISLQVVHVE and(and HuMikb1) was tested in vivo. Four overlapping 170 181 125ETLTPDTQYEFQ exhibited the best I-IL-2106 124 114 129 126 140 178peptides ( L–I , V–S , W–E , Y–
binding, while significant binding reactivity was199Q ) homologous to the two immunoreactive re-observed for overlapping dodecapeptides adjacent to
gions on the extracellular domain of IL-2Rb were 107 118peptide MAPISLQVVHVE , suggesting thatprepared by conventional solid phase synthesis and
some residues on both ends of this sequence areused for immunization. Table 1 summarizes the 125essential for IL-2 binding. The specificity of I-IL-primary sequences and the Ab titers obtained for
2 binding was confirmed by successful competitioneach peptide.
with cold IL-2 as shown in Fig. 3 (open bars).
3.3. Cross-reactivity of anti-peptide polyclonalantibodies 3.5. Proliferation assays
Polyclonal Abs with the higher titers were tested We sought to test whether sequences identified by

88 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
Fig. 1. Identification of epitopes in the IL-2Rb chain using PEPSCAN. Scan of overlapping dodecapeptides homologous to the primary aasequence of the extracellular domain of human IL-2Rb chain precursor (including 26 residues of the signal peptide). Number 1 correspondsto the first residue of mature IL-2Rb. Peptides were reacted with (A) TU11 (35); (B) Mikb1 (33); (C) TU27 (32). Each peptide is identifiedby the number of its amino-terminal residue.

H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 89
Table 1 logical effect by competing with IL-2 for binding.Serological titers of anti-peptide antibodies raised in rabbits For this purpose, synthetic peptides homologous to
a b cPeptide Rabbit Titer the identified sequences were tested for their ability106 124 to inhibit the proliferation of mitogen-stimulatedIL-2Rb L–I A 3.5–4.0
126 140B 3.5 PBLs. Addition of the antigenic peptide W–E
114 129IL-2Rb V–S C 4 had no effect on lymphocyte proliferation (Fig. 4A).D 4.5–5.0 Therefore, it probably does not represent a binding126 140IL-2Rb W–E E 2.0–2.5 107 118domain for IL-2. However, peptides M–E andF 1.5–2.0 178 199
178 199 Y–Q elicited specific inhibitory effects on theIL-2Rb Y–Q G 3.5H 3.0–3.5 proliferation of PHA-stimulated PBL (Fig. 4A).
85 96a 106 124 Peptide R–Q and two randomly selected 17-aaPrimary aa sequence of the peptides IL-2Rb L–I :
106 124 114 129 114 peptides non-homologous to the IL-2Rb ectodomainLMAPISLQVVHVETHRCNI ; IL-2Rb V–S : VVHV-129 126 140 126ETHRCNISWEIS ; IL-2Rb W–E : WEISQASHYFER- (one not shown) were used as negative controls and
140 178 199 178HLE ; IL-2Rb Y–Q : YEFQVRVKPLQGEFTTWS- displayed no biological effect. PHA-stimulated PBLs199PWSQ . showed a bell-shaped dose response to peptideb Two rabbits /peptide were immunized.
c treatment, with maximum suppression of prolifer-All titers are expressed as 2Log (serum dilution at 3310 178 199ation of 67% and 60% for peptides Y–Q andbackground extinction). ELISA values are mean values.107 118M–E , respectively. The inhibition of cell prolif-
PEPSCAN mapping as putative epitopes, and shown eration reached peak values with 500 nM peptide andto elicit antibody production in vivo or binding of the effect was lost at higher peptide concentrationsIL-2 in vitro, represent IL-2 binding sequences on (1000 nM). This can be explained by the hypothesisthe IL-2Rb, and would, therefore, display a bio- that, at too high concentrations, peptides compete
Fig. 2. Binding of anti-peptide polyclonal sera to rIL-2Rb ectodomain. ELISA plates coated with increasing amounts of rIL-2Rb106 124 114 129 126 140 178 199ectodomain were incubated with anti- L–I , anti- V–S , anti- W–E , anti- Y–Q serums, TU11 mAb, pre-bleed E, pre-bleed
126 140 126 140G or a 1000-fold excess of the corresponding immunizing peptide ( WEISQASHYFERHLE for anti- W–E and178 199 178 199YEFQVRVKPLQGEFTTWSPWSQ for anti- Y–Q ).
90 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
125Fig. 3. In vitro binding of IL-2 to synthetic peptides. In vitro binding of I-IL-2 to 52 selected dodecapeptides attached on polyethylene125rods. Each pair of bars corresponds to a specific immunoreactive dodecapeptide. Solid bars represent the amount of bound I-IL-2 in cpm
125 125(Dcpm) for each peptide when incubated with 250 pM I-IL-2 minus the remaining cpm on each rod when the bound I-IL-2 wasreleased. Open bars represent the non-specific binding determined in the presence at 500-fold excess of unlabeled recombinant IL-2. Eachbar represents the average value from duplicate rods and two independent experiments.
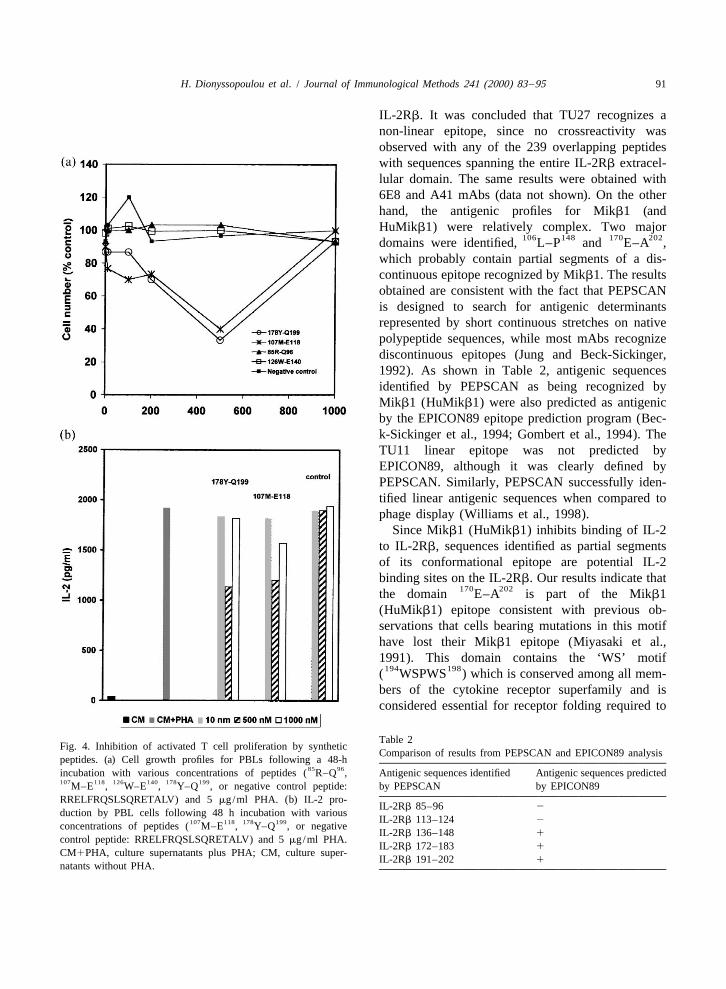
with each other for the same binding site (high tional role in IL-2 binding to its receptor wasentropy) and none can bind to the IL-2R. This examined by monitoring the proliferation rates oftypical bell-shaped or U-shaped dose response curve mitogen-stimulated human peripheral blood T cellsis usually observed in situations where the effects of and IL-2 secretion in the presence of exogenouslyhormone or peptide concentrations are tested on cell added synthetic peptides. Only one linear epitopeproliferation (Fink et al., 1988; Higo and Umeyama, was identified on the IL-2Rb, which was contained
85 961997). in the sequence ( REGVRWRVMAIQ ) and wasInhibition of lymphocyte proliferation by peptides recognized by TU11 anti-IL-2Rb mAb. It has been
178 199 107 118Y–Q and M–E represented by decreased previously predicted, that CF1 and 6E10 anti-IL-2Rb
cell numbers (Fig. 4A) was consistent with a de- mAbs share the same epitope (Francois et al., 1995).crease in endogenous IL-2 secretion (Fig. 4B), which Nevertheless, the primary sequence of this epitope
178 199 107 118was |39% and 35% for Y–Q and M–E , has not been identified. We found that CF1 and 6E10respectively. recognize the same linear epitope as TU11 (data not
shown). All three Abs precipitate IL-2Rb as a125complex with I-IL-2 but do not affect IL-2
4. Discussion binding to its high-affinity receptor (Suzuki et al.,1989; Francois et al., 1995). Therefore, this epitopic
In this study, antigenic sites were identified in the sequence is unlikely to be a binding domain for IL-2.extracellular domain of IL-2Rb, using multiple over- The fact that non-inhibitory anti-IL-2Rb mAbs sharelapping peptides synthesized by the PEPSCAN meth- the same antigenic determinant indicates that theod. In order to confirm the antigenicity of the number of epitopes on the IL-2Rb extracellularidentified epitopes, polyclonal Abs were raised domain is small. In addition, three anti-IL-2Rb
against synthetic peptides with sequences corre- mAbs were tested, namely the TU27 (Takeshita etsponding to putative epitopes of the IL-2Rb. The al., 1989), Mikb1 (Tsudo et al., 1989) and HuMikb1possibility that the identified epitopes play a func- (Hakimi et al., 1993), which block binding of IL-2 to
H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 91
IL-2Rb. It was concluded that TU27 recognizes anon-linear epitope, since no crossreactivity wasobserved with any of the 239 overlapping peptideswith sequences spanning the entire IL-2Rb extracel-lular domain. The same results were obtained with6E8 and A41 mAbs (data not shown). On the otherhand, the antigenic profiles for Mikb1 (andHuMikb1) were relatively complex. Two major
106 148 170 202domains were identified, L–P and E–A ,which probably contain partial segments of a dis-continuous epitope recognized by Mikb1. The resultsobtained are consistent with the fact that PEPSCANis designed to search for antigenic determinantsrepresented by short continuous stretches on nativepolypeptide sequences, while most mAbs recognizediscontinuous epitopes (Jung and Beck-Sickinger,1992). As shown in Table 2, antigenic sequencesidentified by PEPSCAN as being recognized byMikb1 (HuMikb1) were also predicted as antigenicby the EPICON89 epitope prediction program (Bec-k-Sickinger et al., 1994; Gombert et al., 1994). TheTU11 linear epitope was not predicted byEPICON89, although it was clearly defined byPEPSCAN. Similarly, PEPSCAN successfully iden-tified linear antigenic sequences when compared tophage display (Williams et al., 1998).
Since Mikb1 (HuMikb1) inhibits binding of IL-2to IL-2Rb, sequences identified as partial segmentsof its conformational epitope are potential IL-2binding sites on the IL-2Rb. Our results indicate that
170 202the domain E–A is part of the Mikb1(HuMikb1) epitope consistent with previous ob-servations that cells bearing mutations in this motifhave lost their Mikb1 epitope (Miyasaki et al.,1991). This domain contains the ‘WS’ motif
194 198( WSPWS ) which is conserved among all mem-bers of the cytokine receptor superfamily and isconsidered essential for receptor folding required to
Table 2Fig. 4. Inhibition of activated T cell proliferation by synthetic
Comparison of results from PEPSCAN and EPICON89 analysispeptides. (a) Cell growth profiles for PBLs following a 48-h
85 96incubation with various concentrations of peptides ( R–Q , Antigenic sequences identified Antigenic sequences predicted107 118 126 140 178 199M–E , W–E , Y–Q , or negative control peptide: by PEPSCAN by EPICON89RRELFRQSLSQRETALV) and 5 mg/ml PHA. (b) IL-2 pro-
IL-2Rb 85–96 2duction by PBL cells following 48 h incubation with various
IL-2Rb 113–124 2107 118 178 199concentrations of peptides ( M–E , Y–Q , or negativeIL-2Rb 136–148 1
control peptide: RRELFRQSLSQRETALV) and 5 mg/ml PHA.IL-2Rb 172–183 1
CM1PHA, culture supernatants plus PHA; CM, culture super-IL-2Rb 191–202 1
natants without PHA.

92 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
generate the IL-2 binding site (Miyasaki et al., structure of the growth hormone–receptor complex1991). According to Bazan (1990), the sequence (Bazan, 1990). This model shows that both se-178 199Y–Q lies on the loop connecting the F9 and G9 quences are located in the hormone binding domainstrands of the IL-Rb. It was shown that the corre- when superimposed with its homologous domain onsponding loop of the human growth hormone–human the growth hormone receptor. Interestingly, the linear
85 96prolactin receptor complex also contains hormone epitope R–Q recognized by TU11, is located farbinding domains (Somers et al., 1994). In fact, away from the putative hormone binding domain. Itcomputer-assisted primary sequence comparisons was shown that TU11 inhibits IL-15-dependentwith the GenBank/EMBL databases revealed that proliferation of PHA-stimulated PBMCs withoutthis region displays homology to rat and mouse-IL- affecting the physiological function of IL-2 which2Rb (data not shown). In addition, the sequence suggests that IL-15 binds to IL-2Rb by a different170 181E–Q constitutes a highly conserved domain mechanism (Grabstein et al., 1994; DeJong et al.,among human and murine IL-2Rb chains (Kono et 1996). Antibodies such as TU11 do not block IL-2al., 1990). Our results from in vitro IL-2 binding binding to IL-2Rb, but they do partially inhibit
170 181assays indicated that the sequence E–Q binds IL-15-stimulated proliferation, and Mikb1, which125I-IL-2 and, therefore, it is probably part of the completely blocks IL-2 binding to IL-2Rb, onlynative IL-2 binding domain. Interestingly, this do- partially inhibits proliferation stimulated by IL-15.
178main contains the Y conserved residue, a structur- Our data support these speculations. The possibility85 96ally important component of the ligand–receptor that the TU11 epitope R–Q is a binding domain
binding interfaces for class I and class II cytokine for IL-15 interactions with IL-2Rb is very intriguingreceptor superfamilies, as confirmed from the X-ray since IL-15 is the only cytokine other than IL-2structure of the IFNg/ IFNgRa complex (Walter et which shares the IL-2Rb chain for the formation ofal., 1995). its high-affinity receptor complex (Giri et al., 1994).
126 140 178 199Our results show that the sequence W–E is We have shown that peptides Y–Q and107 118probably an antigenic site, since an antibody could M–E added to mitogen-stimulated PBLs sig-
126 140be raised against a W–E synthetic peptide nificantly inhibited their proliferation rates in a dose-which crossreacted with the IL-2Rb. The sequence dependent manner. Failure of the production of126 140 106W–E contained in the IL-2Rb domain L– either IL-2, or its high affinity receptor, or binding of
148P identified by PEPSCAN as epitopic, could be IL-2 to IL-2R upon antigenic stimulation, results in132 134functionally significant. It contains the SHY failure of the T-cell immune response (Waldmann,
motif which lies between the B9 and C9 strands in all 1989; Hatakeyama and Taniguchi, 1991; Smith,members of the cytokine receptor superfamily and 1992; Taniguchi and Minami, 1993), a concept thatseems to be essential for IL-2 binding (Imler et al., is being exploited beneficially in transplantation1992). Based on X-ray structures, this domain con- medicine. Therapeutic strategies to control grafttains binding determinants for complexes of the versus host disease (GVHD) have focused in the pasthuman growth hormone–growth hormone receptor on the use of therapeutic regimens combining subth-(DeVos et al., 1992) and the human growth hor- erapeutic doses of immunosuppressive drugs andmone–prolactin receptor (Somers et al., 1994). anti-interleukin-2 receptor (IL-2R) mAbs in order toMoreover, this domain contributes to binding of IL-6 eliminate activated T-cells only and, thus, removeas shown by site-directed mutagenesis of the corre- graft versus host reactive T-cells without globallysponding region of IL-6R (Kalai et al., 1996). impairing T-cell functions (Mouzaki et al., 1987;However, this sequence was not shown to bind IL-2 DiStefano et al., 1988; Sykes, 1993). Nowadays,in vitro and did not affect proliferation of stimulated humanized Abs are used in experimental models inPBLs. order to avoid anti-idiotypic responses in the recipi-
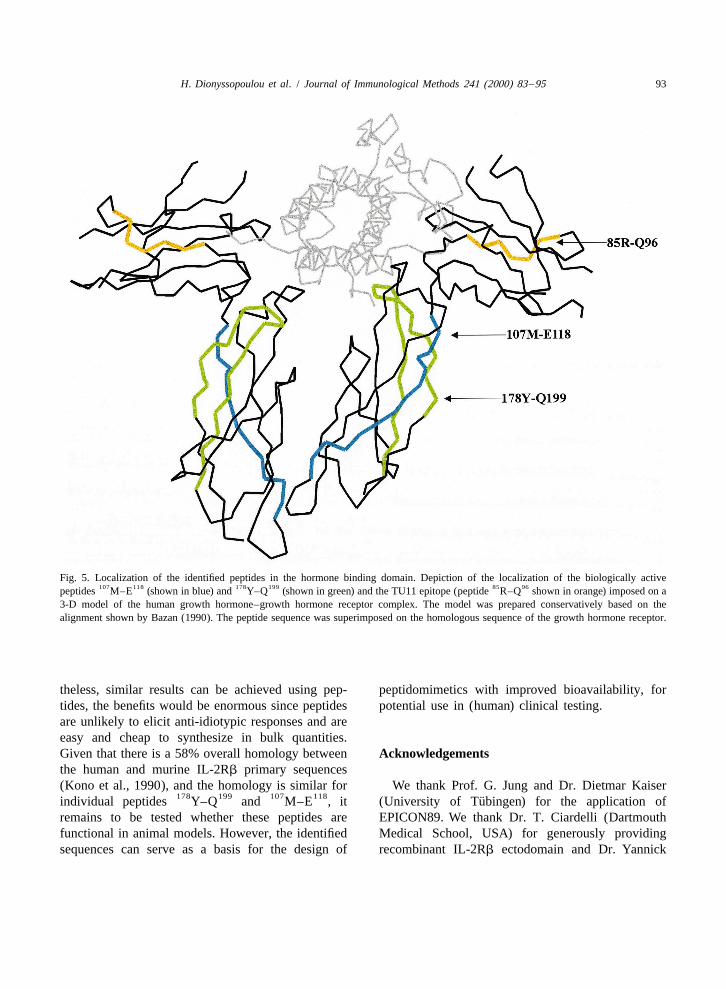
Fig. 5 depicts an alignment of the IL-2Rb se- ent while maintaining a high enough affinity for the107 118 178 199quences M–E and Y–Q identified as Abs to be effective (Kenyon et al., 1999; Kirk et al.,
putative IL-2 binding segments, with the X-ray 1999; Pescovitz, 1999; Woodle et al., 1999). Never-
H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 93
Fig. 5. Localization of the identified peptides in the hormone binding domain. Depiction of the localization of the biologically active107 118 178 199 85 96peptides M–E (shown in blue) and Y–Q (shown in green) and the TU11 epitope (peptide R–Q shown in orange) imposed on a
3-D model of the human growth hormone–growth hormone receptor complex. The model was prepared conservatively based on thealignment shown by Bazan (1990). The peptide sequence was superimposed on the homologous sequence of the growth hormone receptor.
theless, similar results can be achieved using pep- peptidomimetics with improved bioavailability, fortides, the benefits would be enormous since peptides potential use in (human) clinical testing.are unlikely to elicit anti-idiotypic responses and areeasy and cheap to synthesize in bulk quantities.Given that there is a 58% overall homology between Acknowledgementsthe human and murine IL-2Rb primary sequences(Kono et al., 1990), and the homology is similar for We thank Prof. G. Jung and Dr. Dietmar Kaiser
178 199 107 118 ¨individual peptides Y–Q and M–E , it (University of Tubingen) for the application ofremains to be tested whether these peptides are EPICON89. We thank Dr. T. Ciardelli (Dartmouthfunctional in animal models. However, the identified Medical School, USA) for generously providingsequences can serve as a basis for the design of recombinant IL-2Rb ectodomain and Dr. Yannick

94 H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95
S., Fung, V., Beers, C., Richardson, J., Schoenborn, M.,Jacques (INSERM, France) for providing the mono-Ahdieh, M., Johnson, L., Alderson, M., Watson, J., Anderson,clonal antibodies: 6E8, CF1, 6E10 and A41. TU11D., Giri, J., 1994. Cloning of a T-cell growth factor that
and HuMikb1 were kindly given to us by Drs. K. interacts with the beta-chain of the interleukin-2 receptor.Sugamura (Tohoku University, Japan) and J. Hakimi Science 264, 965.
Hakimi, J., Ha, V.C., Lin, P., Campbell, E., Gately, M.K., Tsudo,(Hoffmann-LaRoche, New Jersey).M., Payne, P.W., Waldmann, T.A., Grant, A.J., Tsien, H. et al.,1993. Humanized Mik beta 1, a humanized antibody to theIL-2 receptor beta-chain that acts synergistically with human-ized anti-Tac. J. Immunol. 151, 1075.References
Halimi, H., Eisenstein, M., Oh, J.W., Revel, M., Chebath, J., 1995.Epitope peptides from interleukin-6 receptor which inhibit the
Bazan, F., 1990. Structural design and molecular evolution of a growth of human myeloma cells. Eur. Cytokine Netw. 6, 135.cytokine receptor superfamily. Proc. Natl. Acad. Sci. USA 97, Hanisch, U., Quiron, R., 1996. Interleukin-2 as a neuroregulatory6934. cytokine. Brain Res. Rev. 21, 246.
Beck-Sickinger, A., Rotering, H., Wiesmuller, K., Dorner, F., Hatakeyama, M., Taniguchi, T., 1991. Interleukin-2. In: Sporn,Jung, G., 1994. Mapping of antigenic and immunogenic sites M.B., Roberts, A.B. (Eds.), Peptide Growth Factors and Theirof Haemophilus influenzae outer membrane protein P6 using Receptors I. Springer-Verlag, New York, p. 523.synthetic lipopeptides. Biol. Chem. Hoppe-Seyler 375, 173. Herzberg,V.L., Smith, K.A., 1987. T-cell growth without serum. J.
Cullen, B., Podlaski, F., Peffer, N., Hosking, J., Greene, W., 1988. Immunol. 139, 998.Sequence requirements for ligand binding and cell surface Higo, J., Umeyama, H., 1997. A theoretical approach to theexpression of the Tac antigen, a human interleukin-2 receptor. bell-shaped dependency of cell proliferation on the hormoneJ. Biol. Chem. 263, 4900. concentration. J. Theor. Biol. 186, 477.
DeJong, J., Farner, N., Widmer, M., Giri, J., Sondel, P., 1996. Imler, J.-L., Miyajima, A., Zurawski, G., 1992. Identification ofInteraction of IL-15 with shared IL-2 receptor b and g three adjacent aa of interleukin-2 receptor b chain whichc
subunits. The IL-15/b /g receptor–ligand complex is less control the affinity and specificity of the interaction withc
stable than the IL-2 /b /g receptor–ligand complex. J. Im- interleukin-2. EMBO J. 11, 2047.c
munol. 156, 1339. Jung, G., Beck-Sickinger, A., 1992. Multiple peptide synthesisDeVos, A., Ultsch, M., Kossiakoff, A., 1992. Human growth methods and their applications. Angew. Chem. Int. Ed. Engl.
hormone and extracellular domain of its receptor: crystal 31, 367.structure of the complex. Science 255, 306. Kalai, M., Montero-Julian, F., Grotzinger, J., Wollmer, A., Morel-
DiStefano, R., Mouzaki, A., Araneda, D., Diamanstein, T., Tilney, le, D., Brochier, J., Rose-John, S., Heinrich, P.C., Brailly, H.,N.L., Kupiec-Weglinski, J.W., 1988. Anti-interleukin 2 re- Content, J., 1996. Participation of two Ser–Ser–Phe–Tyrceptor monoclonal antibodies spare phenotypically distinct T repeats in interleukin-6 (IL-6)-binding sites of the human IL-6suppressor cells in vivo and exert synergistic biological receptor. Eur. J. Biochem. 238, 714.effects. J. Exp. Med. 167, 1981. Kenyon, N.S., Chatzipetrou, M., Masetti, M., Ranuncoli, A.,
Fink, R., Ehrhardt, R., Dancygier, H., 1988. Bombensin and its Oliveira, M., Wagner, J.L., Kirk, A.D., Harlan, D.M., Burkly,analogues inhibit interleukin-2-induced proliferation of CTLL- L.C., Ricordi, C., 1999. Long-term survival and function of2 cells. Regul. Pept. 23, 323. intrahepatic islet allografts in rhesus monkeys treated with
Francois, C., Sorel, M., Cherel, M., Brailly, H., Minvielle, S., humanized anti-CD154. Proc. Natl. Acad. Sci. USA 96, 8132.Blanchard, D., Jaques, Y., 1995. Immunological characteriza- Kimura, Y., Takeshita, M., Kondo, N., Ishii, N., Nakamura, M.,tion of antigenic domains on human IL-2 receptor b subunit: Van Snick, J., Sugamura, K., 1994. Sharing of the IL-2epitope-function relationships. Int. Immunol. 7, 1173. receptor g chain with the functional IL-9 receptor complex.
Geysen, M., Barteling, S., Meloen, R., 1984. Use of peptide- Int. Immunol. 7, 115.synthesis to probe viral-antigens for epitopes to a resolution of Kirk, A.D., Burkly, L.C., Batty, D.S., Baumgartner, R.E., Berning,a single amino-acid. Proc. Natl. Acad. Sci. USA 81, 3998. J.D., Buchanan, K., Fechner, Jr. J.H., Germond, R.L., Kampen,
Geysen, M., Rodda, S., Mason, T., Tribbick, G., Schoofs, P., 1987. R.L., Patterson, N.B., Swanson, S.J., Tadaki, D.K., TenHoor,Strategies for epitope analysis using peptide synthesis. J. C.N., White, L., Knechtle, S.J., Harlan, D.M., 1999. TreatmentImmunol. Methods 102, 259. with humanized monoclonal antibody against CD154 prevents
Giri, J., Ahdieh, M., Eisenman, J., Shanebeck, K., Grabstein, K., acute renal allograft rejection in nonhuman primates. Nat.Kumaki, S., Namen, A., Park, L., Cosman, D., Anderson, D., Med. 5, 686.1994. Utilization of the beta-chain and gamma-chain of the Kondo, M., Takeshita, T., Ishi, N., Nakamura, M., Watanabe, S.,IL-2 receptor by the novel cytokine IL-15. EMBO J. 13, 2822. Arai, K., Sugamura, K., 1993. Sharing of the interleukin-2
Gombert, F., Wertz, W., Schluter, M., Bayer, A., Werner, R., (IL-2) receptor gamma chain between receptors for IL-2 andBerthold, W., Jung, G., 1994. Pentapeptide identified as a IL-4. Science 262, 1874.monoclonal antibody binding site in the serine-protease do- Kono, T., Doi, T., Yamada, G., Hatakeyama, M., Minamoto, S.,main of t-PA. Biol. Chem. Hoppe-Seyler 375, 471. Tsudo, M., Miyasaka, M., Miyata, T., Taniguch, T., 1990.
Grabstein, K., Eisenman, J., Shanebeck, K., Rauch, C., Srinivasan, Murine interleukin 2 receptor beta chain: desregulated gene

H. Dionyssopoulou et al. / Journal of Immunological Methods 241 (2000) 83 –95 95
expression in lymphoma line EL-4 caused by promoter Schneider, G., Schrodl, W., Wallukat, G., Muller, J., Nissen, E.,insertion. Proc. Natl. Acad. Sci. USA 87, 1806. Ronspeck, W., Wrede, P., Kunze, R., 1998. Peptide design by
´Langeveld, J.P.M., Casal, J.I., Osterhaus, A.D.M.E., Cortes, E., de artificial neural networks and computer-based evolutionarySwart, R., Vela, C., Dalsgaard, K., Puijk, W.C., Schaaper, search. Proc. Natl. Acad. Sci. USA 95, 12179.W.M.M., Meloen, R.H., 1994. First peptide vaccine providing Smith, K.A., 1992. Interleukin-2. Curr. Opin. Immunol. 4, 271.protection against viral infection in the target animal: studies Somers, W., Ultsch, M., DeVos, A., Kosslakoff, A., 1994. Theof canine parvovirus in dogs. J. Virol. 68, 4506. X-ray structure of a growth hormone–prolactin receptor
Leonard, W., 1996. The molecular basis of X-linked severe complex. Nature 372, 478.combined immunodeficiency: defective cytokine receptor sig- Steward, J.M., Young, J.D., 1984. In: Solid Phase Peptide Syn-naling. Annu. Rev. Med. 47, 229. thesis. Pierce Chemical Company, Rockford, p. 76.
Livnah, O., Stura, E., Johnson, D., Middleton, S., Mulcahy, L., Sugamura, K., Asao, H., Kondo, M., Tanaka, N., Ishii, N.,Wrighton, N., Dower, W., Jolliffe, L., Wilson, I., 1996. Nakamura, M., Takeshita, T., 1995. The common g-chain forFunctional mimicry of a protein hormone by a peptide agonist: multiple cytokine receptors. Adv. Immunol. 59, 225.the EPO receptor complex at 2.8 A. Science 273, 464. Suzuki, J., Takeshita, T., Ohbo, K., Asao, H., Tada, K., Sugamura,
Meloen, R., Amerongen, A., Hage-Van Noort, M., Langedijk, J., K., 1989. IL-2 receptor subunit p75: direct demonstration of itsPosthumus, W., Puijk, W., Plasman, H., Lenstra, J., Langeveld, IL-2 binding ability by using a novel monoclonal antibody.J., 1991. The use of peptides to reconstruct conformational Int. Immunol. 1, 373.determinants; a brief review. Ann. Biol. Clin. 49, 231. Sykes, M., 1993. Novel approaches to the control of graft versus
Miyasaki, T., Maruyama, M., Yamada, G., Hatakeyama, M., host disease. Curr. Opin. Immunol. 5, 774.Taniguchi, T., 1991. The integrity of the conserved ‘WS Takeshita, T., Goto, Y., Tada, K., Nagata, K., Asao, H., Sugamura,motif’ common to IL-2 and other cytokine receptors is K., 1989. Monoclonal antibody defining a molecule possiblyessential for ligand binding and signal transduction. EMBO J. identical to the p75 subunit of interleukin-2 receptor. J. Exp.10, 3191. Med. 169, 1323.
Mosmann, T.R., 1996. The expanding universe of T-cells subsets Taniguchi, T., Minami, Y., 1993. The IL-2 / IL-2 receptor system: a— Th1, Th2 and more. Immunol. Today 17, 138. current overview. Cell 73, 5.
Mouzaki, A., Volk, H.D., Osawa, H., Diamanstein, T., 1987. Tsudo, M., Kitamura, F., Miyasaka, M., 1989. Characterization ofBlocking of IL-2 binding to the IL-2R is not required for the the interleukin-2 receptor beta chain using three distinctin vivo action of anti-IL-2R monoclonal antibody. I. The monoclonal antibodies. Proc. Natl. Acad. Sci. USA 86, 1982.production, characterization and in vivo properties of a new Tzartos, S., Remoundos, M., 1990. Fine localization of the majormouse anti-rat IL-2R mAb that reacts with an epitope different a-bungarotoxin binding site to residues a189–195 of theto the one that binds to IL-2 and the mAb ART-18. Eur. J. torpedo acetylcholine receptor. J. Biol. Chem. 265, 21462.Immunol. 17, 335. Voss, S., Leary, T., Sondel, P., Robb, J., 1993. Identification of a
Mouzaki, A., Rungger, D., Tucci, A., Doucet, A., Zubler, R.H., direct interaction between interleukin 2 and the p64 interleukin1993. Occurrence of a silencer of the interleukin-2 gene in 2 receptor g chain. Proc. Natl. Acad. Sci. USA 90, 2428.naive but not in memory resting T-helper lymphocytes. Eur. J. Waldmann, T., 1989. The multi-subunit interleukin-2 receptor.Immunol. 23, 1469. Annu. Rev. Biochem. 58, 875.
Noguchi, M., Nakamura, Y., Russell, S., Ziegler, S., Tsang, M., Walker, B., 1994. Solid-phase peptide synthesis. In: Wisdom, G.B.Cao, X., Leonard, W., 1993. Interleukin-2 receptor gamma (Ed.), Peptide Antigens: a Practical Approach. IRL Press,chain: a functional component of the interleukin-7 receptor. Oxford, p. 75.Science 262, 1877. Walter, M., Windsor, W., Nagabhushan, T., Lundell, D., Lunn, C.,
Pescovitz, M.D., 1999. New usage paradigms in antibody therapy: Zauodny, P., Narula, S., 1995. Crystal structure of a complexsafety of the new agents. Transplant. Proc. 31, 1201. between interferon-g and its soluble high-affinity receptor.
Plaue, S., Briand, J.P., 1988. Solid-phase peptide synthesis. In: Nature 376, 230.Van Regenmortel, M.H.V., Briand, J.P., Muller, S., Plaue, S. Williams, S.C., Badley, R.A., Davis, P.J., Puijk, W.C., Meloen,(Eds.), Synthetic Polypeptide as Antigens. Elsevier, Amster- R.H., 1998. Identification of epitopes within beta lactoglobulindam, p. 87. recognised by polyclonal antibodies using phage display and
Robb, R., Rusk, C., Neeper, M., 1988. Structure-function relation- PEPSCAN. J. Immunol. Methods 213, 1.ships for the interleukin 2 receptor: localization of ligand and Woodle, E.S., Xu, D., Zivin, R.A., Auger, J., Charette, J.,antibody binding sites on the Tac receptor chain by mutational O’Laughlin, R., Peace, D., Jollife, L.K., Haverty, T., Blues-analysis. Proc. Natl. Acad. Sci. USA 85, 5654. tone, J.A., Thistlethwaite, Jr. J.R., 1999. Phase I trial of
Sana, T., Wu, Z., Smith, K., Ciardelli, T., 1994. Expression and humanized, Fc receptor nonbinding OKT3 antibody,ligand binding characterization of the b-subunit (p75) ectodo- huOKT3gamma1(Ala-Ala) in the treatment of acute renalmain of the interleukin-2 receptor. Biochemistry 33, 5838. allograft rejection. Transplantation 68, 608.




















