
Array-Based Sensing of Normal, Cancerous, and Metastatic Cells Using Conjugated Fluorescent Polymers
Upload
khangminh22Category
view
2download
0
ANTI-CANCEROUS METABOLITES AND EXTRACELLULAR
ENZYME PRODUCTION BY ENDOPHYTIC PENICILLIUM AND
PAECILOMYCES STRAINS
Ph.D Thesis
By
Sajid Ali
CENTRE OF BIOTECHNOLOGY AND MICROBIOLOGY
UNIVERSITY OF PESHAWAR
2016

ANTI-CANCEROUS METABOLITES AND EXTRACELLULAR
ENZYME PRODUCTION BY ENDOPHYTIC PENICILLIUM AND
PAECILOMYCES STRAINS
By
Sajid Ali
A dissertation submitted to the University of Peshawar in partial
fulfilment of the requirements for the degree of
Doctor of Philosophy
In
Biotechnology and Microbiology
CENTRE OF BIOTECHNOLOGY AND MICROBIOLOGY
UNIVERSITY OF PESHAWAR




IN THE NAME OF ALLAH, THE BENEFICENT
THE MERCIFUL
Read! And thy Lord is Most Honorable and Most Benevolent, Who taught (to write) by pen, He taught man that which he knew not
(Surah Al-Alaq 30: 3-5) Al-Quran

Dedication
Dedicated to all humanity……

i
CONTENTS
Chapter Title Page No.
Contents i
List of Tables iii
List of Figures iv
Acknowledgement v
Abstract vi
1 INTRODUCTION AND LITRATURE REVIEW 1
1.1 Introduction to Endophytic Fungi 1
1.2 Role and Function of Endophytic Fungi 4
1.3 Essential Metabolites from Endophytic Fungi 5
1.3.1 Chemical Constituents from Endophytic Fungi 8
1.3.2 Phytohormones from Endophytic Fungi 11
1.3.3 Enzymes from Endophytic Fungi 11
1.4 Endophytic Biotechnology and Uses 13
1.5 Plant Species Selected 13
1.5.1 Ecology, Habitat and Traditional uses 14
1.6 Fungal Isolation, Identification and Diversity 15
1.7 Diversity of Endophytic Fungi by Using DGGE Analysis 20
1.8 Extracellular Enzymes, Indol Acetic Acid and ACC Deaminase
Production 21
1.9 Characterization of Bioactive Secondary Metabolites 24
1.10 Aims and Objectives of the Present Research Work 25
1.11 Study Benefits 26
2 MATERIALS AND METHODS 27
2.1 Plant Collection 27
2.2 Isolation of Fungal Endophytes 27
2.3 Morphological Characters and Molecular Identification of the
Fungal Endophytes
27
2.4 Genomic DNA Extraction 28
2.5 PCR Amplification 28
2.6 Nested PCR for DGGE analysis 29
2.7 Denaturing Gradient Gel Electrophoresis (DGGE) 30
2.7.1 Gel Casting Procedure 30
2.8 Phylogenetic Analysis 32
2.9 Diversity Analysis 33
2.10 ACC deaminase Activity of Endophytic Fungi 33
2.11 Quantification of Extracellular Enzymes 35
2.12 Reagents 36
2.12.1 Substrates 36
2.12.2 Buffer 37
2.12.3 Standards 37
2.12.4 Cultrul Filtrate 37
2.12.5 Microplate set-up 37
2.12.6 FluorescenceReadings 39
2.13 Indole Acetic Acid Quantification of Endophytic Fungi 40
2.14 Extraction and Purification of Bioactive Compounds 41
2.15 NMR Spectroscopy 41

ii
2.15.1 HSQC 41
2.15.2 HMBC 41
2.15.3 COSY 42
2.15.4 NOESY 42
2.15.5 Sample Preparation for NMR 42
2.16 Anticancer Activities 42
2.17 Statistical Analysis 43
3 RESULTS 44
3.1 Morphological Characteristics of Endophytic Fungi Colonies 44
3.1.1 Morphology of Endophytic Fungi Isolated from C. acutangula 44
3.1.2 Morphology of Endophytic Fungi Isolated from B. sacra 44
3.2 Diversity of Endophytic Fungi with Caralluma acutangula and
Boswellia sacra
45
3.3 Sequencing and Identification of Endophytes 47
3.4 Phylogenetic Analysis 48
3.5 ACC deaminase Activity of the Endophytic Fungi 52
3.6 Indole Acetic Acid Quantification of Endophytic Fungi 52
3.7 Extracellular Enzymes Production by Endophytes 53
3.7.1 α-glucosidase,Cellulases, Phospatases 55
3.8 Extraction and Purification of Compounds 57
3.9 Chromatographic and Spectroscopic Techniques 57
3.10 Characterization of Compounds 59
3.11 Enzyme Inhibitory Activities of Secondary Metabolites 65
3.12 MTT Assay 66
4 DISCUSSION 69
Section-1: Endophyte Diversity Assessment 69
Section-2: Potential Role of Endophytes 72
Section-3: Bioactive Metabolites from Endophytes 75
Section-4: Conclusion 81
5 REFRENCES 82
6 APPENDIXES 100

iii
LIST OF TABLES
Table No. Title Page No.
2.1 Constituents and their amounts in DGGE set up. 31
2.2 Ingredients of Stacking Gel 32
3.1 Endophytic fungi isolated from C. acutangula. 46
3.2 Endophytic fungi isolated from B. sacra 46
3.3 Sequence Similarities of Endophytic Fungi Isolated from
C.acutangula and Boswellia sacra
48
3.4 Gene Bank Numbers of Endophytic Fungi Isolated from C.
acutangula and B. sacra 49
3.5 Extracellular enzymes produced by endophytic fungi 54

iv
LIST OF FIGURES
Figure No. Title Page No.
1.1
Environmental continuum and endophytic interaction
with a plant during stress conditions (Adopted from
Khan et al., 2013).
4
1.2
Schematic diagrams of the research methods for
isolation and identification of endophyte community
(Adopted from; Sun and Guo, 2012). Xiang Sun and
Liang-Dong Guo (2012)
19
2.1 Schematic representation of a microplate set-up for the
study of kinetic parameters of β-glucosidase in two soils.
Adopted from (MC Marx, M Wood- 2001)
38
3.1 Evolutionary relationship endophytic fungal strains from
B. sacra
50
3.2 Evolutionary relationship endophytic fungal strains from
C. acutangula
51
3.3 ACC deaminase activity of the isolated endophytic fungi 52
3.4 Indole acetic acid production by endophytic fungi 53
3.5 Structures of Compounds 1-5 60
3.5a Demonstrate Compound 1; 11-Oxoursonic acid benzyl
ester
61
3.5b Demonstrate Compound 2; n-nonane 62
3.5c Demonstrate Compound 3;3-decene-1-ol 63
3.5d Demonstrate Compound 4; 2-Hydroxyphenyl acetic acid 64
3.5e Demonstrate Compound 5; Glochidacuminosides A 65
3.6 Enzyme inhibition activities of the secondary
metabolites isolated and characterized from the
endophytic fungi
66
3.7 Effect of cultural filtrates of endophytic fungi on the
viability of MCF-7 breast cancer cells in culture
67
3.8 Effect of pure compounds on the viability of MCF-7
breast cancer cells in culture
68

v
Acknowledgement
In the name of Allah, who has given me strength and courage to accomplish this PhD
work in the benefit of mankind.I bow my head on thanks and gratitude to Allah forhis
countless blessings.
My first debt of gratitude must go to my supervisor, Dr. Sumera Afzal. She patiently
provided the vision, encouragement and advice necessary for me to proceed through
the PhD program and completes my dissertation.
My deepest regards to my co-supervisor Dr. Muhammad Hamayun, Department of
Botany, Abdul Wali Khan University Mardan. I am especially thankful to Dr. Abdul
Latif Khan, Dr. Liaqat Ali Malik and Dr. Tania Rizvi, University of Nizwa, Chair
of Oman's Medicinal Plants and Marine Natural Products, for providing me laboratory
facilities to conduct analysis of my study.Their love, encouraging behavior and
support were always there where things didn‟t seem to work.
I am thankful to Prof. Dr. Bashir Ahmad, Dean, Faculty of Life and Environmental
Sciences, and Prof. Dr. Ghousia Lutfullah, Director, Center of Biotehnology and
Microbiology, University of Peshawar for their cooperation and facilitation of my
Ph.D research project.
Perhaps, I would not be able to present this work in present form without co-operation
of Higher Education Commission (HEC) Pakistan by funding me through
indigenous PhD fellowship programme.
Thanks to all teachers, students, friends and staff members of Center of
Biotechnology and Microbiology, University of Peshawar for sharing expertise and
for providing a friendly environment. Thanks to everybody who had contributed
directly or indirectly for the completion of this study. Special thanks to my sweet
brothers Asad Ali and Nasir Khan for their moral support and encouragement
throughout the studies.
In the last I wish to thank my parents, wife and daughters (Sofia and Rubab), their
love provided me inspiration and was my driving force. I owe them everything and
wish I could show them just how much I love and appreciate them.
Sajid Ali

vi
Abstract
Fungal endophytes colonize an important niche within the plants through secretion of
secondary metabolites. The metabolites and extracellular enzymes produced by
endophytic fungi regulate the growth of the host plant and contribute in defence
mechanisms.The medicinal plants Caralluma acutangula and Boswellia sacra were used
for the isolation of endophytic fungi. The endophytic fungi were identified as Penicillium
citrinum, Paecilomyces variotii, Aspergillus nidulans, Fusarium oxysporum, Epicucum
nigram, Penicillium purpurogenum, Penicillium spinulosum, Aspergillus caespitosus,
Phoma and Alternaria sp. and were assessed for their potential to produce anti-cancerous
metabolites by performing MTT assay and extracellular enzymes such as cellulases,
phosphatases and glucosidases in growth media. P. variotii, P. citrinum and F. oxysporum
showed significantly higher amount of phosphatases and glucosidases as compared to
other strains. Additionally, P. variotii and F. oxysporum showed significantly higher
potential of indole acetic acid production (tryptophan-dependent and independent
pathways). ACC (1-Aminocyclopropane-1-carboxylate) deaminase results showed that
P.citrinum, P. purpurogenum and P. Variotii had shown prominent ACC deaminase
activity (300 nmol α- ketobutyrate mg-1
h-1
). Fluorescence-based MUB (4-methyl
umbelliferone) standards were used to analyze the presence of extracellular enzymes
glucosidase, phosphatase and cellulase. The bioactive secondary metabolites from
endophytic P. citrinum also revealed some prominent results by performing MTT assay
on breast cancer cell line (MCF-7). The current study concludes that these fungi are
producing bioactive constituents that could provide unique niche of ecological adaptation
by symbiosis and greatly contributing to the healthy life of their host plant. However,
some of the endophytic fungi offer a great potential to produce anti-cancerous metabolites
and extracellular enzymes.

Chapter 1 Introduction and Literature Review
1
INTRODUCTION AND LITERATURE REVIEW
1.1 Introduction to Endophytic Fungi
In nature, almost all plants serve as a reservoir of asymptomatically occurring
microbial flora known as endophytes (Yang et al., 2014; Jain et al., 2015). The term
endophyte was first coined by De-Barry in 1866 (Verma et al., 2007). Hence
„endophyte‟ refers to „within the plant‟ and categorically used for microorganisms
which are inside the plant tissues (Mercado 2015). Microorganisms (bacteria/fungi)
occurring asymptomatically inside the plant tissues (leaves, stem, roots) without
causing any disease condition are classified as endophytes (Bacon and White, 2000;
Saikkonen et al., 2004; Schulz and Boyle, 2005). This definition excludes the
mycorrhizal fungi but does not imply that endophytic fungi are not cultivable on
synthetic media (Gallo et al., 2008). Most commonly these are the fungi that inhabit
in plant hosts for all or part of their life cycle, colonize the internal plant tissues
beneath the epidermal cell layers without causing any apparent harm or symptomatic
infection to their host and inhabit in all parts of the host plants (Selim et al., 2012).
The paleomycological evidences of endophytic symbioses with plants have been
estimated approximately 400 Million years old, thus placing it in the same geological
period as were mycorrhizal symbioses (Smith and Read, 1996; Sapp 2004; Krings et
al., 2007; Rodriguez and Redman, 2008; Heijdenet al., 2015). On the basis of
different parameters like evolution, classification, plant hosts and ecological niche
these endophytes are divided into two major groups i.e. (i) Calvicipitaceous (C) which
inhabit mostly in some grasses and (ii) Non-Calvicipitaceous (NC) which are
associated with tissues of nonvascular plants, ferns and allies, conifers and
angiosperms (Rodriguez et al., 2009; Harman 2011). C-endophytes are playing their

Chapter 1 Introduction and Literature Review
2
pivotal role mainly in stress condition and reported to extend benefits to the host
plants in biotic and abiotic stresses besides this also contributing in the increasing of
plant development (Bacon and White, 2000; Saikkonen et al., 2004; Faeth et al.,
2006; Rodriguez et al., 2009; Hamilton et al., 2010; Eaton et al., 2011; Harman
2011). These abilities of endophytic fungi are reliant on the species of host, its
genotype and environmental conditions (Faeth 2002; Redman et al., 2002; Waller et
al., 2005). NC-endophytes are more diverse in nature, may grow in both above and
below-ground tissues and can be recovered from almost every ecosystem of terrestrial
organisms. However, NC-endophytes represent at least three separate functional
groups (Class-I, II, III) which are based on their life style, features and their
ecological implication (Rodriguez et al., 2009). These endophytes asymptomatically
colonize and confer habitat-adapted fitness to plants (monocots and dicots/eudicots)
which have poor physiological capabilities to handle various environmental
conditions. Due to their adaptability between endophytic and free-living lifestyles,
they are explored with great interest (Selosse et al., 2004).
Since the last three decades of 20th
Century, most of the studies about endophytic
fungi were conducted on the population, habitat and classification. Endophytes are
isolated from variety of ecosystems ranging from hot deserts to tundra and temperate
forests (Hoffman and Arnold, 2008; Arnold and Lewis, 2005). Previously, some of
the studies estimated global diversity of fungi at 1.5 million species, drawing from a
ratio of six species of fungi per vascular plant species and only 7 % of the world‟s
fungi have so far been described. In the past century, many of the 0.1 million fungi
that have been described were those associated with various higher organisms as
either parasites or saprophyte on dead/ dying biological materials (Schulz et al.,2005;
Krings et al., 2007; Gallo et al., 2008). Thus, the question comes to our mind, where

Chapter 1 Introduction and Literature Review
3
are the remaining 0.9 million fungul species? Recently, Hibbett et al. (2011) and
Hawksworth (2001) suggested that this diversity could be more than 5.1 million
species. However, the global diversity of these endophytic fungi is limitless and still a
huge number of species and ecosystems need to be explored. In connection to this a
significant amount of work has also been carried out on the endophyte derived from
medicinal plants (Arnold and Lewis, 2005). The individual plant may serve as host to
more than one endophytes while plant and endophyte relationship is favoured by the
bioactive compounds, proposing that there may be many undiscovered endophytic
microbial flora (Strobel and Daisy, 2003; Wu et al., 2013). The identified strains of
endophytic microorganisms is very less in number which shows that there is
significant opportunities to explore the novel strains of endophytic microorganisms
from variety of ecosystems. Endophytic diversity and the symbiotic relationship of
host plant and endophyte greatly contribute to combat the adverse environmental
conditions and climatic change (Rodriguez et al., 2008; Khan et al., 2013; Yang et al.,
2014). This potential has also been considered for the ability of endophytes to produce
various kinds of biologically active metabolites and enzymes.
In plant microbe interacton the rhizospheric band also offer a place of safety to the microbe
in transportation, reproduction and accessibility to nutrients via the plant roots. Once the
switch over is successful and mutualism is established, it lasts for generations throughout
the plant‟s life. The plant provides a safe haven to the endophyte while facilitating it with
food. In return, it extends diverse benefits to the host plant ranging from an influx of
nutrients to regulating the essential biochemicals after exposure to abiotic stresses. Thus, the
effects on the environmental continuum are ameliorated by promoting the metabolism of
the phyllosphere involving the rhizosphere. The '+', '-' and '-/+' in the phyllosphere show the
increase, decrease and altered activities of various processes during plant development,
respectively, under stress conditions and endophytic association (Khan et al., 2013).

Chapter 1 Introduction and Literature Review
4
Figure 1.1:- Environmental continuum and endophytic interaction with a plant
during stress conditions (Adopted from Khan et al., 2013).
1.2 Role and Function of Endophytic Fungi
Endophytic fungi display a great diversity and are known to have a considerable
effect on their host (Strobel and Daisy, 2003). The microbial flora of the plant and
their diversity present a pivotal role mainly in stress conditions (Schulz and Boyle,
2005; Krings et al., 2007). A variety of relationships can exist between endophytes
and their host plants, ranging from mutualism or symbiosis to antagonism or slight
pathogenesis (Schulz and Boyle, 2005; Arnold 2007; Waqas et al., 2012; Khan et
al., 2013). Generally, the host-endophytes relationships can be described in three
ways: host-specificity, host-recurrence and host-selectivity (Zhou et al., 2006;
Cohen 2006). In host-specificity, a microorganism is limited to a single host or a
single species (Strobel 2003, Strobel and Daisy, 2003) whereas host-recurrence

Chapter 1 Introduction and Literature Review
5
refers to the frequent or predominant occurrence of endophytes in a particular host
or a range of plant hosts, although the endophytes may also be found rarely in
other host plants in the same habitat (Zhou and Hyde, 2001). A single endophytic
species may form relationships with two or many related host plants, but where
there is a preference for one particular host the phenomenon is defined as host
selectivity (Cohen 2006). The term host-preference is most frequently used to
indicate a common occurrence or uniqueness of occurrence of endophytes in a
particular host, and is also used to indicate the difference in endophytic
community composition and relation frequencies from different host plants
(Suryanarayanan and Kumaresan, 2000). Endophytes are also able to colonize
multiple host species of the same plant family within the same habitat, and the
distribution of some endophytes can be similar in closely related plant species (Hu
et al., 2008). The differences in endophytes in their metabolic profile, and hence in
their biological activity, even if between isolates of a species, might be related to
the chemical difference of host plants. This raises the importance of studying host-
endophytes relationships, and the effect of host plants on endophytic metabolite
production. A number of endophytic fungi with their important metabolites are
reported from different medicinal plants.
1.3 Essential Metabolites from Endophytic Fungi
More attention is now being given to study the biodiversity of fungal endophytes, the
chemistry and bioactivity of metabolites produced by endophytic fungi and the
relation between endophytes and their host plants (Schulz et al., 2002; Khan et al.,
2013). Hence, Endophytic fungi represent a significant section of fungal biodiversity,
effect on plant community diversity and structure by dint of producing certain
essential metabolites (Sanders 2004; Gonthier et al.,2006; Krings et al., 2007).

Chapter 1 Introduction and Literature Review
6
Endophytic fungi are an important and novel resource of natural bioactive compounds
with their potential applications in agriculture, medicine and food industry (Morath et
al., 2012; Chen et al., 2014). A variety of interesting molecules have been isolated
from endophytes, including flavonoids, peptides, alkaloids, steroids, terpenoids,
lignans, and volatile organic compounds, many of them are biologically active (Zhang
et al., 2006; Gallo et al., 2008; Kusari et al., 2012). In fact, the study of Schulz et al.,
(2005) showed that about 51% of biologically active metabolites are derived from
endophytes. In the past two decades, a lot of valuable bioactive compounds with
antimicrobial, insecticidal, cytotoxic, and anticancer activities have been successfully
discovered from these tiny endophytic factories. Some endophytes have the ability to
produce the same or similar bioactive compounds as those originated from their host
plants (Kusari et al., 2012). The remarkable discovery of bioactive compound
revealed from the endophytic fungus Taxomyces andreanae in 1993 (Stierle et al.,
1993) and the production of world‟s first billion-dollar anticancer drug, paclitaxel
(Taxol) from endophytic fungi Pestalotiopsis microspore, colonizing in Himalayan
yew tree without causing any disease condition to its host plant (Strobel et al., 1996;
Gallo et al., 2008; Kusari et al., 2012) shifted the attention of many scientists and
started research on the fungal endophytes as potential source of novel and biologically
active compounds. Presently, the wild Taxus plants have been used for the production
of paclitaxel. However, the amount of paclitaxel found in the various parts of Taxus
plant was extremely low. To normalise supply and demand of the market, the
alternative resource and potential strategy should be developed. In the last four
decades, many efficient approaches such as field cultivation, cell and tissue culture,
chemical synthesize for paclitaxel production have been developed, and much
progress has been achieved (Zhou and Wu, 2006). Kumaran et al., 2014 screen the

Chapter 1 Introduction and Literature Review
7
fungus Phoma betae for taxol production which was isolated from Ginkgo biloba,
demonstrated the production of „Taxol‟. Similarly, the production of taxol from an
endophytic fungus, Lasiodiplodia theobromae isolated from the medicinal
plant Morinda-citrifolia showed its cytotoxicity against human breast cancer cell
lines. The endophytic fungal taxol was tested for its bioactivity against human cancer
cell line and the results showed that the bioactive compounds from endophytic fungi
Lasiodiplodia theobromae possess anticancer activity (Pandi et al., 2013).
These discoveries allowed the scientist to explore a variety of endophytes for the
production of valuable compounds such as paclitaxel. Since then, many scientists
have been increasing their interests in studying fungal endophytes as potential source
for the production of a number of valuable compounds. In the past two decades, many
valuable bioactive compounds i.e. alkaloids, terpenoids, steroids, phenols, lactones
and quinones have also been isolated from endophytic fungi and showed
antimicrobial, insecticidal, cytotoxic and anticancer activities (Zhang et al., 2006, Xu
et al., 2008; Zhao et al., 2011).
That‟s why Endophytic fungi are considered as one of the most active group of
microorganisms for the production of biologically active secondary metabolites that
plays important biological roles for human life. The symbiotic relationship and co-
evolution paved the way for a friendly relationship between endophyte and their host
plants. The host provides nutritional requirements and habitation for the survival of its
endophytic flora. While, the endophytes would produce a number of bioactive
compounds for helping the host plants to resist to any stress condition and promoting
the host growth in return (Rodriguez et al., 2009; Silvia et al., 2007). Therefore, to
produce same or similar bioactive compounds as those originated from the host plant

Chapter 1 Introduction and Literature Review
8
is of great importance to understand and utilize this sort of relation in host and
endophyte (Schulz et al., 2002; Zhao et al., 2011; Yang et al., 2014). This will greatly
contribute in the production of rare and important biologically active compounds
(Gunatilaka 2006, Zhou et al., 2009).
The isolation and characterization of different therapeutic drugs from endophytic
fungi have shifted the focus of drug discovery from plants to these endophytes
(Tolulope et al., 2015). The recent advancement in biotechnological research paved
the way for the utilization of endophytes as a promising source of bioactive
compounds and has greatly contributed to preserve the endangered species of
medicinally valuable plants by confirming their activities in a short period of time
(Kharwar et al., 2011). Various endophytic fungi have been identified to yield several
modern medicines with main health cures.As Yang et al., (2003) reported six
endophytic fungi obtained from Sinopodophyllum hexandrum, Diphylleia sinensis and
Dysosma veitchii that have the ability to produce podophyllotoxin inevitable for warts
disease. In a simple way, more than 20% of the identified antibiotics and other drugs
have been produced by fungal and bacterial sources (Strobel and Daisy, 2003). While
the growing demand of human population for food, drug and effective agriculture
paved the way for the exploration of natural synthesizers of bioactive compounds.
1.3.1 Chemical Constituents from Endophytic Fungi
The biological diversity of endophytic fungi coupled with their capability to
biosynthesize bioactive secondary metabolites has provided the stimulus for a number
of investigations on endophytes and their chemical constituents. The endophytic
fungus Entrophosporain frequens obtained from Nothapodytes foetida has the ability
to produce camptothecin (CPT) (Amna et al., 2006). CPT and its analogue10-

Chapter 1 Introduction and Literature Review
9
hydroxy-camptothecin have been regarded as the most effective antineoplastic agents
and in clinical use against ovarian, small lung and refractory ovarian cancers
prevalently all over the world (Sirikantaramas et al., 2007). Camptothecine (CPT) is a
quinoline alkaloid and also known as a potent inhibitor of eukaryotic topoisomerase-I.
CPT is also produced by plant species belonging to the asteroid-clade and recently
efforts have been made to isolate endophytic fungi from some of these plants as
possible alternative sources of CPT. Shweta et al., (2013) documented the isolation of
endophytic fungi from fruit and seeds of Miqueliadentata that produce CPT, 9-
methoxy CPT and 10-hydroxy CPT from endophytic fungi Fomitopsis, Alternaria
alternate and Phomposis species (Shweta et al., 2013).
Similarly, xylarenic acid and xylarenones A and B were isolated from the endophytic
fungus Xylaria species NCY2. These compounds were evaluated for antitumor and
antimicrobial assays in which they showed antitumor activities against HeLa cells (Hu
et al., 2008). The ethyl acetate extract of endophytic fungus Periconiaspecies F-31
isolated from Annona muricata were evaluated for anti-tumor activities. Two new
terpenes named (3S, 6S, 7R, 8S)-periconone A and (1R, 4R, 6S, 7S)-2-caren-4,8-olide
were identified and compounds were pharmacologically evaluated for cytotoxic effect
against different human tumor cell lines. However, they exhibited low cytotoxic effect
(Han-Lin et al., 2011). According to Wu et al., (2013) the endophytic fungus of
Phomopsis species isolated from Aconitum carmichaeli showed very good results for
the production of steroids, more than four different types of steroids were isolated
from the culture broth and their inhibitory activities were evaluated against different
pathogenic fungi (Wu et al., 2013). Ying et al., 2014 used Huperzia serrate (toothed
clubmoss) for endophytic fungal isolation; they isolated and identified the novel types
of metabolites like norcyclocitrinol A, erythro-11α-hydroxy-neocyclocitrinol, and
pesudocyclocitrinol A, from fungal endophyte Penicillium chrysogenum P1X. These

Chapter 1 Introduction and Literature Review
10
compounds were identified by spectroscopic methods and revealed that it share the
C25steroid skeleton. In particular, norcyclocitrinol A, represents the first example of a
C25 steroid. All compounds were evaluated for their cytotoxic activities against HeLa
and HepG2 cell lines while showed no significant results (Ying et al., 2014).
Some of the earlier investigations have proved that Fusariumis a rich source of
biologically active secondary metabolites, including the anti-fungal agents
oxysporidinone and 6-epioxysporidinone which is an antimicrobial agents beauvericin
and bikaverin, fungal toxins fumonisin and sambutoxin, phosphatidylinositide 3-kinase
inhibitor wortmannin, and immunosuppressive agent cyclosporine A. In some of the
study, F. oxysporum culture led to the isolation of two new compounds, a new
oxysporidinone analogue and a new 3-hydroxyl-2-piperidinone derivative. The utilization
of endophytic fungi as a source agent in food industry, drug discovery and controlling
farm pests and pathogens is enormous. As the synthetic compounds showed high level of
toxicity toward environmental conditions the endophytes are the chemical synthesizers
inside plants could be greatly utilized in the synthesis of natural bioactive compounds
(Owen and Hundley 2004, Morath et al., 2012; Chen et al., 2014). Host plant itself has
functioned as a selection system for endophytic microbial flora and developed a
symbiotic association. There are number of examples of endophytes which produce the
same types of bioactive compounds as produce by their host plant. The endophytic fungus
Phomabetae isolated from Ginkgo biloba the endophytic fungus Lasiodiplodia
theobromae isolated from the medicinal plant Morinda citrifolia, the endophytic fungal
strain QJ18 from host plant Gentianama crophylla encodes forgentiopicrin like its host
plant G. macrophylla (Yin et al., 2009; Pandi et al., 2013; Kumaran et al., 2014). Thus,
the endangered species of medicinally important plants and other natural resources can be
protected by utilizing the endophytic microbial flora to satisfy the requirement of drugs
via production of plant-derived pharmaceutical leads by fermentation.

Chapter 1 Introduction and Literature Review
11
1.3.2 Phytohormones from Endophytic Fungi
Waqas et al., (2012) and Khan et al., (2013) suggested that endophytic fungi can
produce physiologically active phytohormones such as gibberellins and auxin, where
the ameliorative effects of endophytes is comparable with that of commercially
available plant growth regulators (Gibberellic Acid) during stressful conditions (Khan
et al., 2014; 2015). Initially, gibberellin was discovered in 1930s from the culture
filtrates of Gibberella fujikuroi (Ogas J, 2000). While, recently different types of GAs
have been recognised and a number of fungal species associated with plants has been
reported as GA producers (Kawaide H, 2006). Host-plants without endophyte-fungal
association are devastated by the waves of extreme temperature, drought, salinity and
pathogen attack (Saikkonen et al., 2010). Hence, the productivity is frequently
compromised in such situations. These endophytes get higher macro- and micro-
nutrients like phosphorus, sulfur, calcium, magnesium and potassium. This capability
has often been considered due to the potential of these endophytes to produce various
biologically active metabolites and enzymes (Yuan et al., 2010). Among metabolites,
plant hormones like GAs and auxin production is a new phenomenon in the endophytic
fungi. Both GAs and auxin have been reported to play an important role in plant
growth, reproduction, metabolism and respond to various environmental signals. In last
decade or so, it has been a known factor that these endophytic fungi, residing inside
host confer abiotic stress tolerance (Yuan et al., 2010; Arnold et al., 2007).
1.3.3 Enzymes from Endophytic Fungi
In addition to the role of endophytic fungi in the production of anticancer and
antimicrobial metabolites, the endophytic fungi have been also known for their potential
to produce different types of extracellular enzymes. The extracellular enzymes are
produced by endophytic fungi for penetrating the host cell wall, and their production
supplements the direct uptake of nutrients by microorganisms and is linked to nutrient
availability and environmental conditions (Latif Khan et al., 2016).

Chapter 1 Introduction and Literature Review
12
The enzymes derived from fungi and bacteria are often more stable than other sources
and are used in food, medicine, beverages, sweets, textiles and leather industries to
process raw materials (Sunitha et al., 2013; Castro et al., 2014). Fungal Kingdom
investigates the roles and importance of fungi play in the biosphere. Recently, only
five genera i.e. Aspergillus, Humicola, Penicillium, Rhizopus and Trichoderma account
for more than 50% fungal enzymes used in the different processes (Ostergaard and
Olsen, 2010).
Chimata et al., 2013 used the fungal strain Aspergillus sp. MK07 for the production of
extracellular amylases enzyme by utilizing wheat-bran as a substrate. Amylases are
important enzymes, mainly used in the starch processing industries to hydrolyse
polysaccharides into simple sugars, about 30% of the world‟s enzymes production is
based on amylases, being used in different processing like confectionary, baking, paper,
textile, detergent and pharmaceutical (Chimata et al., 2013).
Most of the amylases have been screened from soil fungi such as Aspergillus, Penicillum
and Rhizopus (Pandey et al., 2000) while the study of Zaferanloo et al., 2014 showed
that endophytic fungus also encodes for the production of amylases i.e. Preussia minima
can be isolated from Eremophilia longifolia, which can greatly contribute in the
production of α-amylase. The two strains of endophytic fungi Philaophora
finlandia and Philaophora fortinii isolated from the roots of alpine plant communities
were able to breakdown the polymeric forms of carbon, nitrogen and phosphorus found in
the plants (Caldwell et al., 2000). Similarly, Marlida et al., (2000) reported starch
degrading enzyme from endophytic fungi Gibberella pulicaris, Acremonium species
and Nodilusporium species. While, amylase production by few endophytic isolates from
mangrove angiosperm Acanthus ilicifolius and mangrove fern, Acrostichum aureum was
reported by Maria et al., in 2005.

Chapter 1 Introduction and Literature Review
13
1.4 Endophytic Biotechnology and Uses
Biotechnology has motivated the utilization of a distinctive group of plant-associated
microorganisms, known as endophytes. However, the biological and ecological roles of
fungal endophytes still totally unexplored. The European Cooperation in Science and
Technology intended to utilize endophytes in biotechnology and agriculture. There are
four working group activities started research on Ecology of endophytes, Identification of
new competent endophytes and new industrial products in life sciences.
The use of endophytic biotechnology to control plant-pathogenic bacteria and fungi is
receiving increasing attention as a sustainable alternative to synthetic pesticides and
antibiotics. Furthermore, these endophytic microorganisms are likely to be adapted to
the presence and metabolism of complex organic molecules and therefore, show
useful biodegradation activities. In order to reduce the input of pesticides and
fertilizers bring to eco-friendly agriculture, it will be important to develop inocula of
biofertilizers, stress protection and biocontrol agents.
The use of endophytic biotechnology to provide solutions for the economically and
ecologically compatible exploitation of endophytes will greatly contribute to our
present state of knowledge.
1.5 Plant Species Selected
In the present research work the endophytic fungi were isolated from Caralluma
acutangula and Boswellia sacra. Medicinally valuable plant, C. acutangula is found
in the northern and FATA region of Khyber Pakhtunkhwa, Pakistan. Boswellia sacra
a primary tree in the genus Boswellia from which frankincense, a resinous dried sap,
is harvested were collected from salalah region of Oman.

Chapter 1 Introduction and Literature Review
14
1.5.1 Ecology, Habitat and Traditional uses
Caralluma acutangula is a flowering plant belongs to family Asclepiadaceae, mostly
occurring in the northern and FATA region of Khyber Pakhtunkhwa, Pakistan. It is a
compactly branched, cactus-like, perennial stem succulent and having a height of 40-
100 cm with purple-black flowers. Caralluma acutangula is used in some of the
traditional medicine, while very little is known about the endophytic microorganism
from this plant.
Boswellia sacra a primary tree in the genus Boswellia from which frankincense is
harvested. Frankincense is an aromatic resin as an exuded gums obtained from trees of
the Burseraceae family. The resin has been used in incense and fumigants, as well as a
fixative in perfumes. The prominent chemical components in frankincense that provide
anti-inflammatory activities have been documented as boswellic acids. The traditional
applications of frankincense are very diverse - ranging from dental disease to skin
conditions, to respiratory complaints and digestive troubles. The resin was chewed to
stimulate the gums and treat dental infections and sore gums and to generally strengthen
the teeth. Buds and fruit provided a cleansing tonic for the digestive system. Modern
research has focused on frankincense anti-inflammatory properties, particularly in the
treatment of rheumatoid arthritis and soft tissue rheumatism for which it appears to be
extremely useful. Today, frankincense essential oil is used as a fixative and precious oil
not only in the perfume industry, but also lends its scent to soaps, detergents and
numerous cosmetic articles. Considerable amount of work has been attempted to
identify chemical composition of the plant, utilization of its component in various
medicines and isolation of different rhizospheric and endophytic microorganisms
associated with Boswellia sp. Boswellic acids have been identified as a major chemical
component in Boswellia sp. extracts that provide the anti-inflammatory activity.

Chapter 1 Introduction and Literature Review
15
In present research work the endophytic fungi were isolated from medicinally valuable
plants Caralluma acutangula and Boswellia sacra.
1.6 Fungal Isolation, Identification and Diversity
Endophytic fungi that asymptomatically reside in the internal tissues of plants beneath the
epidermal cell layer and contribute in the production of bioactive secondary metabolites
compounds, in addition provide defence to their host plant by producing a plethora of the
substances (D. Wilson, 2000; Strobel G, 2012; Kusari et al., 2014; Mercado-Blanco
2015). The occurrence of endophytic fungi inside plant tissues has been known since the
end of 19th century (Guerin, 1898). Their biological diversity is enormous in a variety of
ecosystems world-wide. The fungi are hosted in nearly 300,000 land plant species, with
each plant hosting one or more of these fungi. Endophytic strains have been isolated from
different parts of diverse plants species including trees, vegetables, fruits and other crops
(M. Rosenblueth and E. Martinez-Romero, 2006; Schulz et al., 2006).
Fungal endophytes have been isolated from different tissues of the plants and
representatives of all major lineages of land plants, that has been inspected (Carroll
and Petrini 1983; Schulz et al., 1999; Strobel G, 2012, Kusari and Spiteller, 2012b).
Previously, two major groups of endophytic fungi have been recognized which shows
dissimilarities on the basis of evolutionary relatedness, taxonomy, plant hosts and
ecological functions the clavicipitaceous endophytes (C-endophytes) and the
nonclavicipitaceous endophytes (NC-endophytes) (Clay & Schardl, 2002; Rodriguez
et al., 2009 ). To date, the best studied groups of fungal endophytes are species in the
family clavicipitaceae that form associations with some cool and warm season grass
species. These species are vertically transmitted, abundant in host tissue, and can
provide a wealth of fitness benefits to the host, for example, herbivore limitation

Chapter 1 Introduction and Literature Review
16
(Clay et al. 1985; De Battistaet al., 1990; Clay and Schardl, 2002). NC-endophytes
are representing a highly diverse assemblage of fungi and have been recovered from
every major lineage of land plants, most commonly from terrestrial ecosystems
(Arnold and Lutzoni, 2007). On the basis of host colonization patterns, mechanisms
of transmission and ecological function the NC-endophytes are classified into three
functional classes (Schardl et al., 2004; Rudgers and Clay 2007; Rodriguez et al.,
2009).
According to Xiang Sun and Liang-Dong Guo, (2012) endophytic fungal diversity can
be identified by two basic techniques i.e. direct observation (microscopy) and
cultivation-dependent methods. In the direct observation method, endophytic fungal
structures within living plant tissues are directly examined under a light and electron
microscope, which can show all endophytic mycobiota within the plant tissue,
particularly biotrophic fungi that cannot be cultured on standard growth media (Deckert
et al., 2001; Lucero et al., 2011). However, most endophytic fungi within plant tissue
have only a hyphal structure, and therefore cannot be identified to any taxonomic
category according to morphology due to lack of spore-producing structures and sexual
or asexual spores. In addition, endophytic isolates cannot be obtained as microbial
resources for further use with the direct observation method. Therefore, this is not
commonly used in endophyte diversity studies (Deckert et al., 2001).
In contrast to direct observation methods, cultivation dependent techniques have been
routinely employed in endophyte diversity studies (Petrini et al., 1982; Rodrigues and
Samuels 1990; Sun et al., 2011; Vieira et al., 2011). It is important to isolate
endophytic fungi for further detailed studies into their characterization, population
dynamics, species diversity to improve plant growth and health, or screening for novel
biologically active secondary metabolites (Zhang et al., 2010; Tejesvi et al., 2011).

Chapter 1 Introduction and Literature Review
17
With cultivation-dependent techniques, the isolation procedure is a critical and
important step in working with endophytic fungi. The living plant tissues are
subjected to a serial process of surface sterilization to remove all organisms from the
surface of the plant. Only internal fungi are isolated by means of incubation of the
plant samples onto nutrient plates.
Cultivation-dependent techniques generally include the following steps;
1) To remove adhering soil particles, debris and major epiphytes, the plant tissues
are washed under tap water.
2) The surface sterilization of plant tissue is important to remove any
epiphyticmicroorganisms on the host surface; a number of protocols are required
for different tissue types.
3) Isolation of endophytic fungi growing out from samples placed on nutrient agar
for further screening and analysis.
4) Purification and sporulation of endophytic isolates under various incubation
conditions.
5) Identification of the isolated endophytic fungi based on morphological
characteristics in the cultures. (Wei et al., 2007; Guo et al., 2008; Sun et al.,
2011; Sun and Guo, 2012).
The development of new techniques in molecular biology brings a novel perspective to
the study of endophyte diversity. Application of molecular techniques, such as DNA
fingerprinting and sequencing methods has the potential to overcome the obstacles in
traditional cultivation-dependent methods. Molecular methods are required for the
identification and understanding of the diversity of endophytic fungi. In a survey of
endophytic fungi from L. chinensis in Hong Kong, a large number of isolates (16.5% of

Chapter 1 Introduction and Literature Review
18
total) did not sporulate, remaining as Mycelia sterilia (Guo et al., 2012). These non-
sporulating isolates were grouped into 19 morphotypes based on their cultural
morphology and identified to different genera (Diaporthe, Mycosphaerella and Xylaria),
families (Pleosporaceae and Clypeosphaeriaceae), and order (Xylariales) based on ITS
sequence analyses. Sun et al. (2011) grouped 221 non-sporulating endophyte strains into
56 morphotypes, and placed these morphotypes into 37 taxa based on ITS sequence
similarity and phylogenetic analyses.
In our present work we did isolation, identification, extracellular enzymes assay and
anticancerous metabolites screening for endophytic fungi isolated from Caralluma
acutangula and Boswellia sacra.

Chapter 1 Introduction and Literature Review
19
Figure 1.2: Schematic diagrams of the research methods for isolation and
identification of endophyte community (Adopted fromGuo et al., 2012).

Chapter 1 Introduction and Literature Review
20
1.7 Diversity of Endophytic Fungi by Using Denaturing Gradient
Gel Electrophoresis (DGGE) Technique
The techniques used in molecular biology offer new opportunities for the analysis of the
structure and species composition of microbial communities. In particular, sequence
variation in rRNA has been exploited for deducing phylogenetic relationships among
variety of microbes and for designing specific nucleotide probes for the detection of
individual microbial taxa in natural environment (Woese 1987; Giovannoni et al.,
1988). All these techniques have also been applied to determine the genetic diversity of
microbial communities and to identifying a number of uncultured microbes. They
constitute the cloning of ribosomal copy DNA or polymerase chain reaction (PCR)-
amplified ribosomal DNA (rDNA) followed by sequence analysis of the resulting
clones (Giovannoni et al., 1990). Denaturing Dradient Gel Electrophoresis is an
approach for directly determining the genetic diversity of complex microbial
populations. The procedure is based on electrophoresis of PCR-amplified 18S rDNA
fragments in polyacrylamide gels containing a linearly increasing gradient of
denaturants. In denaturing gradient gel electrophoresis (DGGE), DNA fragments of the
same length but with different base-pair sequences can be separated. The separation in
DGGE is based on the electrophoretic mobility of a partially melted DNA molecule in
polyacrylamide gels, which is decreased, compared with that of the completely helical
form of the molecule. The melting of fragments proceeds in discrete so-called melting
domains: stretches of base pairs with an identical melting temperature.Once the melting
domain with the lowest melting temperaturereaches its melting temperature at a
particular positionin the DGGE gel, a transition of helical to partially meltedmolecules
occurs, and migration of the molecule will practicallyhalt. Sequence variation within
such domains causes their melting temperatures to differ. Sequence variants ofparticular

Chapter 1 Introduction and Literature Review
21
fragments will therefore stop migrating at different positions in the denaturing gradient
and hence can be separated effectively by DGGE (Lerman et al. 1984; Ward et al.,
1990). This technique has been successfully applied to identifying sequence variations
in a number of genes from several different organisms. PCR can beused to selectively
amplify the sequence of interest before DGGE is used. In a modification of the latter
method, GC-rich sequences can be incorporated into one of the primers to modify the
melting behaviour of the fragment ofinterest to the extent to which close to 100% of all
possible sequence variations can be detected. This procedure allows one for the first
time to directly identify the presence and relative abundance of different species and
thus, to profile microbial populations both qualitatively and semi-quantitatively.
1.8 Extracellular Enzymes, Indole Acetic Acid and ACC Deaminase
Production
Endophytic fungi are relatively unexplored producers of metabolites useful to
pharmaceutical and agricultural industries. A single endophyte produces several
bioactive metabolites. As a result, the role of endophytes in production of various
natural products with greater bioactivity has received increased attention over the last
three decades. Endophytes can influence soil stability directly by their mycelia in the
soil as well as indirectly altering roots and physical conditions of the host plants
(Patil, M. G et al., 2015).An endophytic fungus shows a complex interaction with host
plants and has been extensively studied over the last three decades as a productive
source of novel bioactive natural products. The enzymes derived from fungi and
bacteria are often more stable than other sources and are used in food, medicine,
beverages, sweets, textiles and leather industries to process raw materials (Sunitha et
al., 2013; Castro et al., 2014). The advantageous special effects of endophytic fungi
have been regarded for their potential to produce biologically active secondary

Chapter 1 Introduction and Literature Review
22
metabolites and other substances which can contribute in the defense, growth and
development of their host. Similarly, a variety of substances and enzymes have been
isolated from these endophytes (Khan, A. L et al., 2014).
The enzymes are produced by endophytic fungi in the host and their production
supplements the direct uptake of nutrients by microorganisms and is linked to nutrient
availability and environmental conditions (Khan, A.L et al., 2016). In connection to
this, extracellular enzymes are also produced by some endophytic fungi for
penetrating the host cell wall, as well as to contribute in biocontrol. The enzymes like
glucosidases and cellulases have been assessed from different endophytic microbial
flora. Some of the endophytes have also been known to produce various classes of
secondary metabolites and most of the work has been done on endophytic microbial
flora for screening of biologically active secondary metabolites. In case of bioactive
metabolites, phytoharmones are the natural substances mainly produced by plants to
regulate its growth.
The production of plant like hormones such as Indole acetic acid (IAA) and
Gibberellins is recently documented from different types of endophytes (Waqas et al.,
2012; Kusari et al., 2013; Khan, A. L et al., 2016). IAA plays a very important role in
plant development, growth and combating environmental stimuli while most of the
fungal endophytes are documented for encoding of IAA (Waqas et al., 2012; Khan,
A.L et al., 2016). Consequently, different biosynthetic pathways have been proposed
for IAA production: Tryptophan dependent and Tryptophan independent while the
proposed Trp-dependent pathways are of four types (indole-3-acetamide pathway
(IAM), Indole-3-acetaldoxime pathway (IAOX), Tryptamine pathway (TAM), and
Indole-3-pyruvic acid pathway (IPA)). In this way, a single endophytic strain

Chapter 1 Introduction and Literature Review
23
sometimes exploits more than one biosynthetic pathway by expressing the genes
present on plasmid and chromosome to contribute in the development of their host
(Idris et al., 2007).
In 1978, for the first time Honma and Shimomura reported ACC from Pseudomonas
(Honma and Shimomura, 1978).The enzyme 1-Aminocyclopropane-1-carboxylate
(ACC) deaminases have the capability to hydrolyze ACC which is the immediate
precursor of ethylene in plant. The lowering down of ethylene levels by ACC
deaminase is considered one of the major mechanisms employed by microbial flora of
the plant to facilitate plant growth. ACC deaminase has been found in various
bacteria, yeasts, and fungi, and it can convert ACC into α-ketobutyrate and ammonia
(Abeles, F. B et al., 2012; Yim et al., 2013).
The microbial flora of the plants are containing ACC deaminase and can take up and
degrade ACC from the plant and therefore decrease ethylene synthesis in the plants
(Glick, 2014). ACC deaminases are normally related to free-living soil
bacteria/rhizobacteria and some of the mycorrhizal. However, a very few endophytic
microbes have been known to produce ACC deaminase. The exuded ACC is of plant is
metabolized by fungi and bacteria possessing ability to produce ACC deaminase. This
stimulates plant ACC efflux, decrease the root‟s ACC and ethylene concentration, thus
increase root growth and development (Glick, 2014). The relationship of ACC and IAA
affect each other, the endogenous content of ACC increased the rate of ethylene
production in the presence of IAA, but failed to increase the ACC content in the
absence of IAA (Yoshii and Imaseki 1981). Therefore, it is suggested that the combine
effect of ACC deaminase and IAA are responsible for the pragmatic plant growth
promotion and development.

Chapter 1 Introduction and Literature Review
24
The utilization of endophytes as a potential source of industrially relevant enzymes is
in queue. Hence, they occupy a relatively unexplored site and can represent a new
source in obtaining different enzymes. The present research was carried out to explore
new sources of valuable bioactive compounds and extracellular enzymes from
endophytic fungi of Caralluma acutangulaandBoswelliasacra also to understand their
functional role with the host.
1.9 Characterization of Bioactive Secondary Metabolites
There are more than 20,000 bioactive metabolites of microbial origin (Berdy 2005). In
eukaryotic organisms fungi are among the most important group that are well known
for producing many novel metabolites while many of endophytic fungi were recently
reported to produce bioactive metabolites such flavonoids, peptides, alkaloids,
steroids, phenolics, terpenoids and lignans with antimicrobial, anticancer and antiviral
potentiality. The discovery of taxol producing fungi increased the importance of
endophytes and shifted natural products research from plant to endophytic fungi
(Schulz et al., 2002; Chinet al., 2006).
Schulz et al., (2006) revealed that more than 50% of biologically active metabolites
originate from endophytes (Schulz et al., 2006) and many novel bioactive compounds
with antimicrobial, insecticidal, cytotoxic, and anticancer properties have been
successfully isolated and characterized from endophytic fungi (Berdy 2005; Khan et
al., 2011; Xiao et al., 2014).
The potential for the production of anticancer drug „taxol‟ (paclitaxel) from
Pestalotiopsis microspore and many other endophytic fungi such as Fusarium solani
isolated from Taxus chinensis, Pestalotiopsis guepini and Periconiaspecies encoding

Chapter 1 Introduction and Literature Review
25
for taxol (Strobel et al., 1997) Alternaria and Aspergillus isolated from Ginkgo biloba
and Podocarpus species respectively, has set the stage for increasing interest in fungal
endophytes (Liu et al., 2009).
In view of these potentialities of endophytic fungi, the present research work was
designed to isolate the endophytic fungi from Caralluma acutangula and Boswellia
sacra for the production of bioactive metabolites. The endophytic wealth is yet to be
explored from a variety of plants. The metabolomics produced at extracellular level
during the growth of these endophytic fungi were assessed using advanced
chromatographic and NMR spectroscopic techniques.
1.10 Aim and Objectives of the present research work
The aim of this research work is to explore biologically active secondary metabolites
from endophytic fungi isolated from medicinally valuable plant. New methodologies
and their utilization are important for the discovery of drugs from endophytes. As
most of free living fungi are pathogenic to human and causing a variety of diseases.
The self-medication, false practices and repeated use of antibiotics has led to the
increase resistant species of some existing available drugs in our country. Therefore, it
is very important to explore and identify biologically active secondary metabolites
from endophytic sources and preserve the endangered species of medicinally
important plants.
To isolate endophytic fungi from medicinally important plant Caralluma and
Boswellia species.
Screening and identification of isolated endophytic fungi by morphological
and molecular analysis.
Isolation and purification of secondary metabolites.

Chapter 1 Introduction and Literature Review
26
Structural elucidation of the purified metabolites.
Anti-cancerous and Enzyme inhibitory activities of isolated compounds.
1.11 Study Benefits
Plants and fungi are very important in health-care. Worldwide more than 80% of the
population relies on traditional medicine, much of which is based on plant remedies.
But recent trends and development in endophytic biotechnology focus on the isolation
of endophytic fungi and there bioactive compounds from medicinally important plants.
This will strengthen the importance of biotechnology and its products in our
country.
This will help in the utilization of bioactive compounds isolated from
endophytic fungi. Which may leads to result in the development of new
valuable pharmaceutical compounds.
This will greatly contribute in the study of extracellular enzymes from
endophytic fungi.
This can put up an association among the researchers and pharmaceutical
industries.
This will provide job opportunities in the field of biotechnology and
pharmaceutical industries.
Plants and microorganisms are used in a variety of medicine which is beneficial in
one way or the other. The isolation, identification and utilization of bioactive
compounds from endophytic fungi can greatly contribute to the research and
industries in a country like Pakistan.

Chapter 2 Materials and Methods
27
MATERIALS AND METHODS
2.1 Plant Collection
Different samples of Caralluma acutangula and Boswellia sacra were collected
from northern areas of Pakistan and salalah region of Oman, respectively. The
samples were subjected for the isolation of endophytic fungi and were brought to
laboratory in a sterilized zip bags (121°C for 20 min) in ice box (4°C). The
samples were labelled and stored till further processing.
2.2 Isolation of Fungal Endophytes
The samples were surface sterilized with sodium hypochlorite (2.5%; 30 min in a
shaking incubator at 120 rpm) and repeatedly washed with autoclaved distilled
water (DDW) to remove any epiphytic microbes and ecto-mycorrhizae (Bayman et
al., 1997). Isolation of fungi from bark/leaf were carried out on Hagem minimal
medium, containing 0.5% glucose, 0.05% KH2PO4, 0.05% MgSO4.7H2O, 0.05%
NH4Cl, 0.1% FeCl3, 100ppm streptomycin and 1.5% agar (pH 5.8±0.2). The newly
emerged fungal spots were separated and further grown and stored on potato
dextrose agar (PDA, 50 ppm). The efficiency of sterilization was monitored by
imprinting the tissues on Hagem and PDA plates. Upon contaminant growth, the
tree samples were again sterilized. The morphologically different (Arnold et al.,
2007) endophytic fungal strains were grown in Czapek broth medium (1%
Glucose, 1% Peptone, 0.05% KCl, 0.05% MgSO4.7H2O, and 0.001% FeSO4.7H2O;
pH 7.3±0.2) and incubated on shaking incubator (28ºC with 150 rpm for 8 days).
2.3 Morphological Charaters and Molecular Identification of the
Fungal Endophytes
The endophytic microbes were grouped into different groups on the basis of colony
shape, thickness, colour of aerial hyphae, colony reverse colour, growth rate and

Chapter 2 Materials and Methods
28
pattern, margin characteristics, surface texture, and growth depth into medium
(Arnold et al., 2007). The endophytes were identified by genomic DNA extraction,
PCR techniques, nucleotide sequencing, and phylogenetic analysis as described by
Khan et al., (2011).
2.4 Genomic DNA Extraction
In DNA extraction, two methods were applied in this study.gDNA was isolated
according to manufacturing instructions from fresh mycelial mates with a Solgent
Genomic DNA preparation kit and another efficient method was developed for the
isolation of genomic DNA from endophytic fungi, because usual CTAB extraction
method and mycelial grinding was causing DNA shearing. Rich mycelial culture was
obtained by growing fungus in Czapek culture broth (supplemented with 1% glucose
and peptone) for 7 days on rotary shaking incubator (120 rpm and 28°C), and
lyophilized for 24 hrs. A 0.5 g of lyophilized sample was broken carefully in 2 ml
eppendorf, with the blunt end spatula or with a glass rod. Double volume of lysis
buffer (20 mM Tris-HCL, pH8.0; 10 mM EDTA; 1% SDS) containing 1% of 2-
mercaptoethanol was added. The mixture was vortexed briefly (30 sec) to obtain
homogeneity and left to incubate for 2 hr in water bath set at 55°C. 250 μl/ml of pre-
heated 4% CTAB extraction buffer was added to lysed cells mixture and incubated
further at 65°C for 1 hr. Chloroform extraction followed by iso-propanol precipitation
yielded condensed strand of nucleic acid, which was cleaned from RNA using 10 µl
of RNase A for 2 hr of incubation at 37°C.The isolated DNA was suspended in 50µl
of autoclave deionized distilled water and tested for purity (Hamayun et al., 2009).
2.5 PCR Amplification
The fungal isolate was identified through sequence analysis of the internal transcribed
region (ITS) of 18S rDNA, using universal primers ITS-1 (5´-TCC GTA GGT GAA

Chapter 2 Materials and Methods
29
CCT GCG G-3´) and ITS-4 (5´-TCC TCC GCT TAT TGA TAT GC-3´) (Taylor and
Bruns 1999). A 25 µl of PCR mixture contained 2.5 µl of dNTPs and Ex-Taq buffer,
2 µl of each primer, 0.5 µl of DNA sample, and 0.2 µl of Ex-Taq polymerase. The
remaining volume was adjusted with 15.3 µl of autoclaved deionized distilled water.
For the amplification of ITS1 and ITS4 regions of 18S rDNA, the reaction cycle
consisted of initial denaturation (95°C) for 2 min, 35 cycles of denaturation (95°C) for
30 s, annealing (55°C) for 60 s, extension (72°C) for 30 s and a final extension time
for 5 min (72°C). The resultant products were gene cleaned using a Nucleogen gene
clean kit, ligated in T-vector using Takara Perfect T-cloning kit, and then inserted into
E. coli competent cells (RBC) by overnight incubation (37°C). Transformed cells
were selected, grown overnight (37°C) in LB broth and their plasmids were extracted
using SolGent Plasmid mini-prep kit, which were later sequenced.
2.6 Nested PCR for DGGE Analysis
Fungal 28S rDNA fragments fromsamples were amplified by nested PCR using specific
primers. In first round the primer set P1 (5P-ATCAATAAGCGGAGGAAAAG-3P) and
P2 (5P- CTCTGGCCTTCACCCCTATTC-3P) were used, yielding a PCR product of
approximately 800 bp. The 25-Wl PCR assays contained 2 W1 template, 5U PCR buffer
(335 mM Tris (pH 8.8), 83 mM (NH4)2SO4, 3.75 mM EGTA, 25% glycerol, 0.1%
Tween 20), 2.5 mM MgCl2, 200 WM of each dNTP, 10 pmol of each primer and 3 mg
ml31 BSA. After initial denaturation at 94°C for 4 min and cooling to 80°C, 2 U Tth-
Polymerase (Hybaid) were added. Thirty five cycles were performed by using 94°Cfor 1
min, 40°Cfor 1 min, 72°C for 2 min, followed by 72°C for 10 min. Negative controls
produced no PCR products. Nested PCR was performed using the U1 and U2 primers
with an additional 37-bp GC-clamp at the 5P-end of the primer U1. Forty PCR cycles
were performed as described above, except for the annealing temperature, which was set

Chapter 2 Materials and Methods
30
to 50³C. All amplifications were performed as hot start reactions in a PCR Express cycler
(Hybaid). DNA extracts of the fungal strains inoculated on the mortar blocks were used
as positive controls in the second PCR round following DGGE analysis. All PCR
products were analysed by electrophoresis in 1.5% agarose gels for 60 min at 100 V
before DGGE was carried out.
2.7 Denaturing Gradient Gel Electrophoresis (DGGE)
PCR products of fungal isolates were analysed by the DCode System (Bio-Rad) using
10% (w/v) acrylamide (37.5:1acrylamide: bisacrylamide) gels. Detailed procedure is
as follow.
2.7.1 Gel Casting Procedure
Before, gel casting, the gel casting plates were cleaned thoroughly, either with a
commercial solution for this purpose, or with detergent, DI water, and ethanol. The gel
casting plates were dried with a lab wipe. The blue gaskets were fixed to the glass plate
with curved corners and gaskets were sealed. To keep separate the glass plated we
inserted separators on the inside of the gasket‐covered glass. The plates were kept
together with the stronger casting clamps with two on each side and two on the bottom.
Frozen aliquot of 10% APS was used and 1000ul DI water with 0.1 g ammonium
persulfate. The following reagents and their respective amount were utilized.

Chapter 2 Materials and Methods
31
Table 2.1:- Constituents and their amounts in DGGE set up.
Reagents Chamber 1L Chamber 2R
Milli‐Q Water 10.832 mL
3.102 mL
Loading dye 500 uL 40% Acrylimide, 1.06 %
Bis‐Acrylimide 3.075 mL 3.075 mL 7 M urea, 40% formamide
6.183 mL 13.413 mL
50x TAE 410 uL 410 uL APS 100 uL 120 uL TEMED 6 uL 7.5 uL
Milli‐Q Water 10.832 mL
3.102 mL
Loading dye 500 uL 40% Acrylimide, 1.06 %
Bis‐Acrylimide 3.075 mL 3.075 mL 7 M urea, 40% formamide
6.183 mL 13.413 mL
50x TAE 410 uL 410 uL APS 100 uL 120 uL TEMED 6 uL 7.5 uL
Milli‐Q Water 10.832 mL
3.102 mL
Loading dye 500 uL 40% Acrylimide, 1.06 %
Bis‐Acrylimide 3.075 mL 3.075 mL 7 M urea, 40% formamide
6.183 mL 13.413 mL
50x TAE 410 uL 410 uL APS 100 uL 120 uL TEMED 6 uL 7.5 uL
Milli‐Q Water 10.832 mL
3.102 mL
Loading dye ‐‐‐ 500 uL 40% Acrylimide, 1.06 %
The addition of APS and TEMED resulted into polymerization. Immediately after the
addition of APS and TEMED and stirring, the valve was get opened on the mixing
chamber, as well as the in‐line valve on the tubing. The pump was switched on and
outlet needle was placed on one side of the gel casting rig. When the gel was 4‐5 cm
from the top of the casting plates the pump was turned off. The gel polymerized in
about 2 hours.

Chapter 2 Materials and Methods
32
The tank was filled with TAE buffer: 200 mL 50x TAE and 20 L DI water and heated
to 60°C. After the gel had polymerized water was poured off on top of the gel.
Stacking gel was prepared in a 15 mL conical tube as follow.
Table 2.2:- Ingredients of Stacking Gel
Reagents Volume
40% Acrylimide: 1.06 % Bis‐Acrylimide
3.075 mL
50x TAE 200 uL
APS 70 uL
TEMED 10 uL
Milli-Q Water 8.3 ml
The gels were run in 1UTAE (40 mM Tris, 20 mM acetate, 1 mM Na2EDTA (pH 7.8)
and a linear gradient of the denaturants urea and formamide increasing from 35 to
65%. One hundred percent denaturant is defined as 7 M urea and 40% formamide
(v/v; deionised). PCR products obtained from DNA extracts of fungal isolates and an
admixture of each PCR product from the isolates was used as positive control in
DGGE. The fingerprints of the inoculated samples were carried out with final PCR
products. Gels were run for 6 h at 150 V in 1UTAE at a temperature of 60³C. DGGE
bands were visualised by ethidium bromide staining and UV illumination. Digital
images acquired by CCD camera were inverted using the Easy Image Plus software.
2.8 Phylogenetic Analysis
The BLAST search program (http://blast.ncbi.nlm.nih.gov) was used to compare the
nucleotide sequence similarity of the ITS regions of related fungi. The closely related
sequences obtained were aligned through CLUSTAL W using MEGA version 6.0

Chapter 2 Materials and Methods
33
software (Tamura et al., 2013) and a neighbor-joining tree was constructed using the
same software. One thousand bootstrap replications were used as a statistical support
for the nodes in the phylogenetic tree. The aligned sequences were submitted to
GeneBank of NCBI for obtaining the accession numbers.
2.9 Diversity Analysis
The endophytic fungal diversity was estimated by using the Shannon diversity index
(H) and Simpson‟s diversity index (1-D) for both domesticated and wild types of
samples. The colonization density, colonization rates and isolation rates of fungal
diversity were calculated as the percentage of segments infected by one or more
isolates from the total number of segments of each plated tissue. All samples were
analyzed in triplicate. The data are presented as the mean ± standard error of the mean
(SEM) and differences were evaluated by using one-way analysis of variance
(ANOVA).
2.10 ACC Deaminase Activity of Endophytic Fungi
ACC deaminase activity was assayed according to a modification of the method of
Honma and Shimomura (1978) which measures the amount of α-ketobutyrate produced
upon the hydrolysis of ACC. The number of μmol of α-ketobutyrate produced by this
reaction was determined by comparing the absorbance at 540 nm of a sample to a
standard curve of α-ketobutyrate ranging between 10 and 200μmol. A stock solution of
100 mmol L-1 α-ketobutyrate was prepared in 0.1 mol L-1 Tris-HCl (pH 8.5) and
stored at 4°C. Just prior to use, the stock solution was diluted with the same buffer to
make 10 mmol L-1 solution from which a standard concentrations curve was generated.
In a series of known α-ketobutyrate concentrations, 2 mL of the 2, 4-dinitrophenyl-
hydrazine reagent (0.2% 2, 4-dinitrophenyl-hydrazine in 2 mol L-1 HCl) was added, the

Chapter 2 Materials and Methods
34
contents were vortexed and incubated at 30ºC for 30 min, during which time the α-
ketobutyrate was derivitized as a phenylhydrazine. The color of phenylhydrazine was
developed by the addition of 2 mL, a 2 mol L-1 NaOH, the absorbance of the mixture
was measured after mixing by using spectrophotometer at 540 nm.
For determining ACC deaminase activity, Endophytic fungal strains were grown in
rich medium (TSB) for 4 days at 28°C. The cells were then harvested by
centrifugation, washed with 0.1 M Tris-HCl (pH 7.5), and incubated for another 4
days in Dowkin and Foster minimal medium containing 5 mM ACC as the sole source
of nitrogen. The fungal cells were collected by centrifugation (Holguin and Glick
2001) and suspended in 5 mL of 0.1 mol L-1 Tris-HCl, pH 7.6, and transferred to
microcentrifuge tube. The contents of the tubes were centrifuged at 16000 rpm for 5
min and supernatant was removed. The pellets were suspended in 2 mL 0.1 mol L-1
TrisHCl, pH 8.5. Thirty μL of toluene were added to the cell suspension and vortexed
for 30 seconds. Two hunderedμL of the toluenized cells were placed in a fresh
microcentrifuge tube, 20 μL of 0.5 mol L-1 ACC were added to the suspension,
vortexed, and then incubated at 30°C for 15 min, following the addition of 1 mL of
0.56 mol L-1 HCl, the mixture was vortexed and centrifuged for 5 min at 16000 rpm
at room temperature. Two mL of the supernatant was vortexed together with 1 mL of
0.56 mol L-1 HCl. There upon, 2 mL of the 2, 4- dinitrophenylhydrazine reagents
(0.2% 2, 4-dinitrophenylhydrazine in 2 mol L-1 HCl) was added to the glass tube, and
the contents were vortexed and then incubated at 30°C for 30 min. Following the
addition and mixing of 2 mL of 2 mol L-1NaOH, the absorbance of the mixture was
measured by using spectrophotometer at 540 nm (Shaharoona et al., 2006).

Chapter 2 Materials and Methods
35
2.11 Quantification of Extracellular Enzymes
To quantify extracellular enzymes, the method of Marx et al., 2001 was adopted with
some modifications. Briefly, all the substrates were obtained from Sigma-Aldrich Co.
Ltd in crystalline form. Ten milliliters of a 10 mM stock solution of each 4-
methylumbelliferone (MUB) substrate was prepared, while the assay procedures were
the same for each substrate. Depending on the substrate, a 7-MUB standard was used.
A 10 mM stock solution of pure MUB was prepared in methanol (0.1762 g of 4-
methylumbelliferone in 100 mL). This stock solution was diluted in MES buffer to 1
μMand stored at 4°C.
The endophytic fungi grown in Czapek broth were harvested using centrifugation (4°C,
12,000 rpm for 10 min). The pure and fresh culture filtrates (CF) were syringe filtered
(0.22 μm) to remove traces of turbidity. For each type of enzyme analysis, a minimum
of three replicates for each substrate (CF + buffer + substrate), a quenched standard
(sample + buffer+ 4-MUB), and a substrate control (media + buffer + substrate) were
maintained. The total volume of liquid in the cuvette was 2 mL CF or buffer or media
and 100 μL substrate or 4-MUB with different types of CF obtained from endophytic
fungi. The pre-optimized fluorescence spectrophotometer (Shimadzo, Tokyo, Japan)
was used to read the absorbance with 360 nm excitation and 460 nm emission at time
zero and 30-minute intervals for 2 hours.
The Bradford reagents are more commonly used for protein assay in which under
acidic conditions the red form of the dye is converted into its bluer form to bind to the
protein being assayed. It is the simple and reliable method used for protein
determination (Bradford, 1976). The protein-dye complex formed has absorption
maxima at 595 nm, this complex is detected in the assay using spectrophotometer or

Chapter 2 Materials and Methods
36
micro plate reader (Reisner et al., 1975). Practical advantage of this method is that
reagent is simple to prepare and the colour is developed rapidly and is relatively
stable.Fluorescence-based MUB standards were used to analyze the presence of three
enzymes (β-1,4-glucosidase, 1,4-β-cellobiosidase, and phosphatase).
2.12 Reagents
2.12.1 Substrates
All substrate analogues were obtained from sigma-Aldrich Co. Ltd, in a crystalline
form. A list of substrates tested is given below (4-MUB = 4-methylumbel-liferone and
7-AMC = 7-amino-4-methyl coumarin). They allow the targeting of a wide range of
enzymes involved in the hydrolysis of C, N and P compounds. Ten ml of a 10 mM
stock solution of each MUB/AMC-substrate was prepared and assay procedures were
the same for each substrate.
4-MUB-β-D-glucoside
4-MUB-β-D-galactoside
4-MUB-7-β-D-xyloside
4-MUB-β-D-glucuronide
4-MUB-β-D-cellobioside
4-MUB-N-acetyl-β-glucosaminide
4-MUB-phosphate
L-leucine-7-AMC
L-tyrosine-7-AMC
L-arginine-7-AMC
The substrates were pre-dissolved in 1ml of ethylene glycol monomethylether
(methylcellosolve), except 4-MUB-β-D-glucuronide, 4-MUB-phosphate, L-leucine-7-

Chapter 2 Materials and Methods
37
AMC and L-arginine-7-AMC, which were dissolved in sterile deionised water. All
substrates were made up to 10ml with sterile deionised water and the resulting stock
solutions of 10 mM were kept at 4°C. New working solutions of the enzyme substrate
were prepared for each assay by diluting the stock solution to 1000, 100 and 10 µM in
sterile 25ml universal bottles. All dilutions were made in sterile (autoclaved) buffer.
2.12.2 Buffers
In order to standardize the method, all enzymes assays were carried out in buffered
conditions; this also stabilizes the fluorescence intensity of MUB, which is highly
dependent on pH (Chrost and Krambeck, 1986). The choice of buffer depended on the
particular enzyme. Glycosidases and acid phosphatase were assayed in 0.1M MES
buffer (2- [N-Morpholino] ethanesulfonic acid, pH 6.1) (sigma-Aldrich Co. Ltd), and
peptidase in 0.05M Trizma buffer, pH 7.8.
2.12.3 Standards
Depending on the substrate tested either a 4-MUBor a 7-MUB standard was used. A 10
mM stock solution of pure MUB was prepared in methanol (0.1762 g of 4-
methylumbelliferone in 100 ml). This stock solution was diluted in MES buffer to 1 µM.
For the AMC standard, 0.1752g of 7-amino-4-methyl coumarin was dissolved in 100 ml
methanol (to prepare a 10 mM stock solution), then diluted in Trizma buffer to 1 µM.
2.12.4 Cultrul Filtrate
For each sample, 2ml of fresh CF was taken into cuvette. A 100µl of substrate or 4-
MUB with different types of CF obtained from endophytic fungi.
2.12.5 Microplate set-up
The plate set-up varies according to the aim of the investigation (e.g. estimation of kinetic
parameters or simple measurement of enzyme activity rates under optimum substrate, pH
and temperature conditions, in different soils). Each plate therefore includes a minimum
of three replicates for each substrate at each substrate concentration (sample + buffer +

Chapter 2 Materials and Methods
38
substrate), a quenched standard (sample + buffer + 4-MUB/7-AMC), a substrate control
(sterile water + buffer + substrate) and an optional abiotic control (autoclaved soil +
buffer + substrate) (Sinsabaugh, personal communication). The total volume of liquid in
each well of the microplate was 200 µl. A typical plate set-up for kinetic studies of
glucosidase in two different soils is described below and represented schematically in
Figure 2.1.
Figure 2.1:- Schematic representation of a microplate set-up for the study of kinetic
parameters of β-glucosidase in the pure and fresh culture filtrates (CF) of endophytic
fungi grown in Czapek broth. Adopted from (MC Marx, M Wood- 2001).
1. For the enzyme assay and the standards 20 µl were withdrawn from the CF under
continuous stirring with a 4-channel pipette and dispensed into the microplate. At
the same time, the negative controls were set up, one in sterile water, to check for
contamination of the substrate or buffer, and another control in autoclaved soil
(optional), to investigate abiotic cleavage of the analogue.
2. Depending on the final substrate concentration in the wells, the appropriate amount
of sterile buffer was added into each well with a digital multi-channel pipette.

Chapter 2 Materials and Methods
39
3. The standards were added to give final amount of 0, 10, 20, 30, 40, 50, 60 and 70
pmol 4-MUB/7-AMC well-1
.
4. Various aliquots of the 10, 100 and 1000 µM working substrate solutions were
added into the wells in order to establish the substrate saturation curves. The final
substrate concentrations in the wells were 2, 8, 20, 50, 80, 200, 300, 500, 700 and
900 µM.
Inhibition of the reaction and maximization of the fluorescence intensity through
alkalinisation of the mediumas suggestedby Freeman et al., (1995), was found to be
unnecessary because of the high sensitivity of the analytical equipment used to
measure fluorescence intensity of MUB is very low (Chrost and Krambeck, 1986), a
change in fluorescence was detected. The elimination of the alkalinisation and
purification step makes the microplate method very quick and offers potential for
routine soil analysis.
2.12.6 Fluorescence Readings
The fluorescence intensity was measured by a computerized microplate fluorimeter
(BioLuminTM
960, Kinetic fluorescence/absorbance, Molecular Dynamics, Inc).
This analytical equipment allows incubation of the microplate at a constant
temperature and automatically repeats measurements after a given time interval
without having to interrupt the reaction. For the measurements described here the
fluorimeter was programmed to shake the microplate for 5 S in order to homogenise
the reaction medium, then to pre-incubate the microplate for 10 min at 30°C (bring
microplate and its content up to temperature), before starting the first reading cycle.
From then on measurements were taken every minute for 35 min at 30°C. After
finishing the readings, the computer software automatically fitted a regression line
to the data and calculated the rate of fluorescence increase in each well. Since the

Chapter 2 Materials and Methods
40
rate of fluorescence was measured rather than the absolute amount of fluorescence
at the of the incubation period, there was no need to incubate a „time zero blank‟
into the microplate for background fluorescence. Initial experiments demonstrated
that background fluorescence did not change during the incubation time.
2.13 Indole Acetic Acid Quantification of Endophytic Fungi
Estimation of indole-3-acetic acid (IAA) in the culture broth was done using
colorimetric assay. The endophytes cultured in 20 mL LB broth without (0 g/L) and
with 0.1 g/L of L-tryptophan and incubated at 30±2°C in shaking incubator at 200 rpm
for seven days. The fungal cultures were then centrifuged at 10,000xg for 10 min at 4°C
and the cell free cultures were filtered through 0.45µm cellulose acetate filter
(DISMIC®, Denmark). The filtrates were acidified to pH 2.8 with 1 N HCland extracted
3 times with 20 mL ethyl acetate. The ethyl acetate fractions were combined and
evaporated under vacuum at 45°C in a rotary evaporator. The residue was re-suspended
in 3 mL 50% methanol. One milliliter of supernatant was mixed with 2 mL Salkowski
reagent (12 g FeCl3/L of 7.9 M H2SO4) and kept in dark for 30 min. The resultant
reddish color was read after 30 min at 535 nm in ELISA Spectrophotometer (BioRad,
USA). The amount of IAA was calculated with standard of pure IAA (Sigma-Aldrich.,
Korea, Ltd) prepared separately.Indol-Acetic-acid may be produced by two pathways;
(1) Tryptophan dependent and (2) tryptophanindependent pathways. One set of the
fungal strains were grown in the Czapek broth without supplementing any precursor
(Tryptophan) for IAA production to check IAA production via tryptophan-independent
pathway. For the investigation of tryptophan dependent production of IAA endophytic
fungi were grown in Czapek medium containing different concentration of
L-tryptophan. This precursor was added to the media in 500, 1000, 1500 and 2000µg
50mL-1
.Strains were harvested after seven days of growth through filtration.

Chapter 2 Materials and Methods
41
2.14 Extraction and Purification of Compounds
The cultural filtrates were extracted by implying the equal volume of ethyl acetate
(EtOAc) three times. Both the extracts were compiled and concentrated in vacuo to get
the crude extract. The extracts were compiled and dried over sodium sulfate
(anhydrous) and concentrated in vacuo to afford the crude extract. The EtOAc extract
was then subjected to silica gel column chromatography using gradients of ethyl
acetate/n-hexane system to afford five fractions (Fr.A to Fr.E). Fr.E was further
subjected to recycling preparative HPLC (JAI) analysis. Compound (mg) was purified
at a retention time of 23 min by using ethyl acetate/n-hexane (6:4) in a silica gel
column with the flow rate 3.5 mL/min after five recycles.
2.15 NMR Spectroscopy
Nuclear magnetic resonance spectroscopy is a powerful tool for structure elucidation
of different natural products. The NMR spectra (1H and 13C) were recorded on
Bruker spectrometer operating at 600-MHz (150-MHz for 13C). The δ-values on the
chemical shift scale are reported in ppm, while the J values of the coupling constants
were noted in Hz.
2.15.1 HSQC
Hetero-nuclear Multiple Quantum Correlation. It is 2D experiment which correlates the
chemical shift of proton with the chemical shift of the directly bonded carbon. On the
bottom axis is a proton spectrum and on the other is a carbon. The cross peaks give the
shift of the corresponding proton and carbon with each other. This experiment utilizes
the one-bond coupling between carbon and proton (J=120-215 Hz).
2.15.2 HMBC
Hetero-nuclear Multiple Bond Correlation. This experiment utilizes multiple bond
couplings over two or three bonds (J=215Hz. The Cross peaks are between protons and

Chapter 2 Materials and Methods
42
carbons that are two or three bonds away (and sometimes up to four or five bonds away).
Direct one-bond cross-peaks are suppressed. This experiment is analogous to the proton-
proton COSY experiment in that it provides connectivity information over several bonds.
2.15.3 COSY
This is the correlation spectroscopy which is good for determining basic connectivity
via J- couplings, through-bond.
2.15.4 NOESY
Nuclear Overhauser Effect Spectroscopy: This allows one to see through-space
effects, useful for investigating conformation and for determining proximity of
adjacent spin systems.
2.15.5 Sample Preparation for NMR
NMR instruments require 5 mm tubes (5mm outer diameter of the tube) whereas
length of NMR tube is usually 7 or 8 inches. The samples were dissolved in 0.6 mL of
CDCl3 genetely mixed and transferred into NMR tube. The solution height in the
NMR tube must be within 3.5-4.5 cm. Marker pen were used for a short code written
at the top of the NMR tube.For 1H spectra of organic compounds with a molecular
mass less than 500, the quantity of material required is usually 3-10 mg. More
samples were needed for 13
C, which is ~6000 times less sensitive than 1H. Bruker 600
mhz instrument with a cryo-probe optimised for 13
C measurements.
2.16 Anticancer Activities
Breast cancer cell line (MCF-7) was used for thescreening of cytotoxicity of the crude
extracts and bioactive pure compounds obtained from endophytic fungi. The cell lines
were culturedin Advanced DMEM with 10% NBCS (inactivated) and5mM l-
glutamine, and then grown at 37C in a humidatmosphere with 5% CO2 in air. The 3-

Chapter 2 Materials and Methods
43
(4,5-dimethylthiazol-2-yl)-2,5diphenyltetrazolium bromide(MTT) colorimetric assay
developed by Mosmann, 1983.
2.17 Statistical Analysis
All samples were analyzed in triplicate. The data are presented as the mean ± standard
error of the mean (SEM). Differences were evaluated using one-way analysis of
variance (ANOVA). Differences were considered significant at P < 0.05 and were
calculated by GraphPad Prism Version 5.0 for Windows (GraphPad Software, San
Diego, CA, USA). The mean values were compared using Duncan‟s multiple range
tests at P < 0.05 (SAS 9.1, Cary, NC, USA). The Unscrambler version 9.0 by Camo
was used for principal component analysis to understand the correlation of enzyme
production abilities by the endophytic fungi.

Chapter 3 Results
44
RESULTS
3.1 Morphological Characteristics of Endophytic FungiColonies
3.1.1 Morphology of Endophytic Fungi Isolated from C. acutangula
The fungal endophytes shows “circular” form of colony on agar plates, and all
colonies were flat with entire margins displaying off white color with the passage of
time the color changed into yellow and brownish-black. F1 (Paecilomyces variotii)
colony was raised showing filamentous fungi strongly resembles to Penicillium but
can be differentiated by its loose branches and cylindrical-conidiogenous cells with
tapering-tips on PDA plates. The colour of P. variotii colony initially white, and
becomes yellow, yellow-brown. The FEF6B (Alternaria sp) flat colony and was
covered by grayish, short aerial hyphae. Initially, the surface was grayish white and
converted into dark-brown due to the production of pigments. FEF7 (Aspergillus
nidulans), were showing septate hyphae with a woolly-colony texture and white
mycelia. The green colour of colonies is due to pigmentation of the spores. BSR
(Epicoccum nigram), colonies were bright shades of yellow, orange and often with
brown or black and fast growth were noted on PDA.
FEF5A (Penicillium purpurogenum) were showing white to light green mycelia while
the back side of the plate were light pink. FEF1 (Fusarium oxysporum), initially, the
mycelia of the isolates on PDA plates were delicate, white to creamy and pink, the
manual of Booth 1971 was used for the identification of these fungi and were having
dispersed form of colony with black sporos organization.
3.1.2 Morphology of Endophytic Fungi Isolated from B. sacra
The endophytic fungi isolated from B. sacra were characterized on the basis of their
morphological characteristics. FEF1 (Fusarium oxysporum) were also isolated from

Chapter 3 Results
45
B.sacra, the mycelia of the isolates on PDA plates were delicate, white to creamy and
silightly pink in colour. The Strain FEF2 (Penicillium spinulosum) was white in
colour and forming a circular and fragile body, While it‟s a difficult task to
differentiate most of penicillium species phenotypically. The FEF3 (Aspergillus
caespitosus) colonies of Aspergillus caespitosus varying on different media, in
Czapek solution agar were slow growing. The mycelium was largely submerged and
very tough to tear. Aspergillus caespitosus was characterized particularly by clusters
of irregular ovoid structure, initially, colorless becoming reddish purple with the
passage of time. FEF4 were recorded as uncultured species. FEF5 (Alternaria sp) was
common in both B. sacra and C. acutangula. These were also having flat colony.
Initially, the surface was grayish white and converted into dark-brown due to the
production of different pigments. FEF6 (P. citrinum), the colonies of P. citrinum were
floccose, showing slow growth. The mycelium was white to greyish-orange in colour.
3.2 Diversity of Endophytic Fungi with C. acutangula and B. sacra
For sampling, a total of 80 plants parts from C. acutangula and 67 different parts from
B. sacra were selected for the isolation of endophytic fungi; the different pieces were
sterilized for isolation of endophytic fungi. A total of 25 endophytic fungal strains
were isolated and only 12 were selected for further processing (Table 3.1 and 3.2).
The selected endophytes were grouped on the basis of different parameters: colour of
aerial hyphae, surface texture, colony, shape, height from the medium, base colour,
growth rate and pattern (Arnold et al., 2007). The overall morphological analysis
suggested that all the selected stains possessed different characteristics.

Chapter 3 Results
46
Table 3.1:- Endophytic fungi isolated from C. acutangula.
Strain
Code
Part used for
Isolation
Fungal endophytes Plant species
F1 Stem Paecilomyces variotii C. acutangula
FEF6B Root Alternaria sp. C. acutangula
FEF7 Stem Aspergillus nidulans C. acutangula
BSR Root Epicoccum nigram C. acutangula
FEF5A Stem Penicillium
purpurogenum
C. acutangula
FEF1 Leaf Fusarium oxysporum C. acutangula
Table 3.2:- Endophytic fungi isolated from B. sacra
The colonization rate in C. acutangula shows the significant results and is much
higher in stem as compare to root. The colonization rate for the stem part was 2.28
whilst for root it was 0.681. Similarly, the isolation rate was significantly higher in
stem (1.46) as compared to roots (0.435). The diversity indices for fungal endophytes
as analysed by Shannon-Weiner indices (𝐻1= 0.9077) and Simpson indices (1/𝑙=
1.778) indicated differences in parts variation and species richness. The rate of
Strain Code Part used for
Isolation
Fungal endophytes Plant species
FEF1 Leaf Fusarium oxysporum B. sacra
FEF2 Leaf Penicillium spinulosum B. sacra
FEF3 Stem Aspergilluscaespitosus B. sacra
FEF4 Leaf Uncultured B. sacra
FEF5 Leaf Alternaria sp. B. sacra
FEF6 Leaf Penicillium citrinum B. sacra

Chapter 3 Results
47
colonization also much higher in the leaves (3.12%) of B. sacra compared to the
stems (1.7%), and isolation rate was higher in leaves (4.11%) compared to stems
(1.22%).The diversity indices for endophytic fungi from B.sacrawere also analysed
by Shannon-Weiner indices (𝐻1= 0.8877) and Simpson indices (1/𝑙= 1.06) indicated
differences in parts variation and species richness.
The fungal species did not differ significantly between plant species, whereas they are
different from each other in their habitation. These endophytic fungi were further
identified by extracting the gDNA, amplification of PCR, sequencing of 18S rRNA
region and bioinformatics tools (BLASTn search).
3.3 Sequencing and Identification of Endophytes
The ITS sequences of the rRNA gene region from the 10 endophytic fungi revealed
that the fragment lengths ranged from 600–900 bp. The sequences were aligned in
MEGA 6.0 and BLASTn searched to correlate them with the highly homologous
fungal strains. Most of the fungal sequences showed 95–100% homology with related
fungi (Table 3.3). The phylogenetic analysis of these strains showed 94–99%
homology with ITS sequences of rRNA genes of related species. The evolutionary
history was inferred using the Maximum Parsimony method.

Chapter 3 Results
48
Table 3.3:- Sequence Similarities of Endophytic Fungi Isolated from C. acutangula
and B. sacra
ID
Name
Length
Identities
Match Total (%)
F1 Paecilomyces variotii 997 722 756 100%
FEF2 Penicillium spinulosum 568 567 571 98%
FEF6B Alternaria sp. 701 620 691 99%
FEF7 Aspergillus nidulans 820 768 801 98%
BSR Epicucum nigram 924 561 564 98%
FEF5A Penicillium purpurogenum 920 776 810 100%
FEF6 Penicillium citrinum 867 781 809 100%
FEF1 Fusarium oxysporum 823 756 780 98%
FEF4 Uncultured. 742 690 710 94%
FEF3 Aspergilluscaespitosus 802 735 792 95%
3.4 Phylogenetic Analysis
The 18S rDNA sequences from these endophytic fungi revealed that the fragment
lengths range from 520 – 997 bp. The sequences were aligned in MEGA 6.0 and then
were BLASTn searched to correlate them with the highly homologous fungal strains.
Most of the fungal sequences showed 94-99% homology with related fungi (Table
3.3). The isolated fungi belonged to Paecilomyces, Fusarium, Penicillium, Aspergillus
and Alternaria species. Based on 94 – 99% sequence similarity, we identified the
fungal strains as Aspergillus nidulans, Aspergillus caespitosus, Paecilomyces variotii,
Fusarium oxysporum, Alternaria sp. Penicillium purpurogenum, Penicillium

Chapter 3 Results
49
spinulosum, Phoma sp. Epicucum nigram and Penicillium citrinum (Table 3.1 and
Table 3.2). The phylogenetic analysis of these strains showed 94 – 99% homologies
with ITS sequences of related species. The sequences were deposited in the GenBank,
NCBI for accession numbers.
Table 3.4:- Gene Bank Numbers of Endophytic Fungi Isolated from C. acutangula
and B. sacra
Strain
Code
Gene Bank
Number
Fungal endophytes Plant species
F1 KY921608 Paecilomyces variotii C. acutangula
FEF6B KY921609 Alternaria sp. C. acutangula
FEF7 KY921610 Aspergillus nidulans C. acutangula
BSR KY921611 Epicoccum nigram C. acutangula
FEF5A KY921612 Penicillium
purpurogenum
C. acutangula
FEF1 KY921613 Fusarium oxysporum C. acutangula
FEF1 KY474347 Fusarium oxysporum B. sacra
FEF2 KY474348 Penicillium spinulosum B. sacra
FEF3 KY474349 Aspergilluscaespitosus B. sacra
FEF4 KY474350 Uncultured endophyte B. sacra
FEF5 KY474351 Alternaria sp. B. sacra
FEF6 KY474352 Penicillium citrinum B. sacra

Chapter 3 Results
50
Evolutionary Relationship of Endophytic Fungal Strains from B. sacra
Figure 3.1:- The evolutionary history was inferred using the Neighbor-Joining method
(Saitou and Nei 1987). The optimal tree with the sum of branch length = 7.97184890 is
shown. The percentage of replicate trees in which the associated taxa clustered together
in the bootstrap test (2000 replicates) are shown next to the branches (Felsenstein J,
1985). The tree is drawn to scale, with branch lengths in the same units as those of the
evolutionary distances used to infer the phylogenetic tree. The evolutionary distances
were computed using the Maximum Composite Likelihood method (Tamura et al.,
2004) and are in the units of the number of base substitutions per site. The analysis
involved 59 nucleotide sequences. All positions containing gaps and missing data were
eliminated. There were a total of 94 positions in the final dataset. Evolutionary analyses
were conducted in MEGA6 (Tamura et al., 2013).

Chapter 3 Results
51
Evolutionary Relationship of Endophytic Fungal Strains from C. acutangula
Figure 3.2:- Evolutionary relationships of taxa of endophytic fungal strains isolated from
Caralluma-acutangula. A neighbour joining tree was constructed of homologous ITS
sequences using MEGA-6.0 with 1K-bootstrap replication. Botrybasidium-subcoronatum
was used as an out group.
Aspergillus sp. BAB-3919
Aspergillus nidulans strain BPPTCC 6038
Aspergillus nidulans strain NRRL 2395
Aspergillus sp. BAB-4426
LF1
SF2
Paecilomyces variotii strain LXM5
Paecilomyces variotii strain GF59
Paecilomyces variotii strain SCSAAF0011
Paecilomyces variotii isolate D SC2
LF3
Fusarium proliferatum isolate Fp-1
Fusarium proliferatum strain PA3
Fusarium proliferatum strain MA84
Epicoccum nigrum strain A168
Epicoccum nigrum strain A152
Epicoccum nigrum strain A52
Epicoccum nigrum strain A49
Penicillium purpurogenum strain KCTC6820
Penicillium purpurogenum isolate Tian1
Penicillium purpurogenum strain KCTC16073
Penicillium purpurogenum strain HS-A82
SF5
6B
Alternaria sp. B13
Alternaria sp. CPCC 480375
Alternaria sp. GE
Alternaria sp. SPS-04
SF3
Botryobasidium subcoronatum voucher KHL s.n.
97
97
100
83
56
100
96
100
100
100
83
100
93
100
100
100
0.1

Chapter 3 Results
52
3.5 ACC Deaminase Activity of the Endophytic Fungi
ACC deaminase was observed in endophytic fungal strains by their growth in
Dworking and Foster (DF) minimal medium containing ACC (Figure 3.3), P. citrinum
(FEF6) strain had the highest value of deamination of ACC (358nmol α- ketobutyrate
mg-1
h-1
) followed by P. variotii (F1). In this way, the endophytic fungal strain
Epicucum nigram (BSR) had the lowest value (62nmol α- ketobutyrate mg-1
h-1
) of
ACC deaminase activity.
Figure 3.3:-ACC deaminase activity by the isolated endophytic fungi in (DF)
minimal medium containing ACC. Different letters in the column shows that values
are significantly different (p<0.05) from each other as evaluated from DMRT
(Duncan's Multiple Range Test) test.
3.6 Indole Acetic Acid Quantification of Endophytic Fungi
Our results showed that P. citrinum (FEF6) and P. variotii (F1) have the highest
results (1.64 and 1.54 nM/mL) for IAA production, while Epicucum nigram (BSR)
and Phoma Sp. (BSL), produced a little amount (0.21 and 0.30 nM/mL) (Figure 3.4).
0
50
100
150
200
250
300
350
400
F1 FEF2 FEF6B FEF7 BSR FEF5A FEF6 FEF1 FEF4 FEF3

Chapter 3 Results
53
Similarly, the endophytic fungi P. spinulosum, A. caespitosus and Alternaria
speciesalso revealed some prominent results for IAA production. The production of
IAA is dependent on the type of pathway utilized by endophyte. In our studies, for
the estimation of IAA in culture broth sophisticated methodologies were used as
colorimetric assay in addition with L-tryptophan which favoured the high amount of
IAA. Then endophytic fungal IAA was also quantified by using Waters Ultra
Performance Liquid Chromatography (UPLC) system.
Figure 3.4:- Indole acetic acid production by endophytic fungi. Different letters in the
column shows that values are significantly different (p<0.05) from each other as
valuated from DMRT (Duncan's Multiple Range Test) test.
3.7 Quantification of Extracellular Enzymes from Endophytic Fungi
To recognise the extracellular enzymes producing capability of isolated endophytic
fungi during axenic conditions. All the endophytic strains were grown in broth
medium for 14 days and were centrifuged to obtain pure culture. Fluorescence-based
MUB standards were used to analysed the presence of three enzymes (β-1,4-
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
1.8
F1 FEF2 FEF6B FEF7 BSR FEF5A FEF6 FEF1 FEF4 FEF3

Chapter 3 Results
54
glucosidase , phosphatase and 1,4- β –cellobiosidase . Standard curve readings were
taken for the florigenic substrate excitation and emission in the presence of buffer and
in combination with MUB standard. A curve fitting values for Cellulases (R2=0.98; y
= 6.35), phosphatases (R2=0.96; y = 113.04) and glucosidases (R
2=0.98; y = 36.4)
was recorded on fluorescence spectrophotometer.
The different fungal strains showed varying concentrations of cellulases,
phosphatases and glucosidases in their pure culture filtrates. The endophytic fungal
strain P. citrinum and P. variotii displayed very prominent results in the culture media
while P. spinulosum, A. caespitosus strain and A. nidulans strain also encoded for a
higher amount of extracellular enzymes as compare to F. oxysporum, P. spinulosum
and Alternaria sp.
Table 3.4:- Extracellular enzymes produced by endophytic fungi
ID NAME OF
ENDOPHYTIC FUNGI
Cellulases (μM
-1min
1mL)
Phosphatases (μM
-1min
-1mL)
Glucosidases (μM
-1min
-1mL)
F1 Paecilomyces variotii 5.65±0.00 0.41±0.00 35.22±0.01
FEF2 Penicillium spinulosum 0.41±0.00 2.12±0.00 46.82±0.00
FEF6B Alternaria sp. 0.64±0.00 0.02±0.00 16.66±0.00
FEF7 Aspergillus nidulans 21.12±0.00 0.68±0.00 18.73±0.00
BSR Epicucum nigram 0.67±0.00 1.22±0.00 31.23±0.00
FEF5A Penicillium purpurogenum 35.66±0.00 0.53±0.00 38.33±0.00
FEF6 Penicillium citrinum 0.38±0.00 3.31±0.00 42.24±0.16
FEF1 Fusarium oxysporum 15.24±0.00 0.70±0.00 03.23±0.00
FEF4 Uncultured endophyte 05.64±0.00 0.35±0.00 52.44±0.00
FEF3 Aspergilluscaespitosus 15.33±0.00 0.72±0.00 32.13±0.00

Chapter 3 Results
55
3.7.1 Glucosidases, Cellulases and Phosphatases
In case of glucosidases A. nidulans, P. variotii, P. purpurogenum and Alternaria has
significantly higher concentration where P. spinulosum, A. caespitosus and P. citrinum
were encoding for very little amount of glucosidases. The phosphatase production in P.
citrinum, P. variotii and P. spinulosum was significantly higher. The P. purpurogenum,
A. caespitosus and Alternaria sp. also produce significantly higher amount of enzymes.
While A. nidulans, Fusarium oxysporum and Alternaria sp. encodes for a very little
amount of phosphates enzymes (Figure 3.5). The cellulases activity showed significant
results for most of the isolated endophytic fungi. The production of cellulases enzymes
was much higher in P. variotii followed by F. oxysporum, A. caespitosus and P.
spinulosum (Figure 3.6). While the endophytic fungal strains A. nidulans and P.
purpurogenum encodes for a very little amount of these enzymes.
The capability to produce extracellular enzymes, indol-acetic acid and proficiency of
ACC deaminase by endophytic fungal species from Caralluma acutangula and
Boswellia sacra was the central part of the present research work. The isolated
endophytes from the different parts (root, leaf and stem) of Caralluma acutangula and
Boswellia sacra showed very prominent results in different biological activities.
Extracellular enzymes are encoded by endophytic fungi for penetrating the host cell
wall, as well as contribute in biocontrol.
The enzymes like glucosidases, cellulases and phosphatases were estimated by using
florigenic substrates and standard curve readings were taken in combination with
MUB standard. The endophytic fungi A. nidulans, P. variotii, P. purpurogenum and
Alternaria has significantly higher concentration where P. spinulosum, A. caespitosus
and P. citrinum were encoding for very little amount of glucosidases. The previous
data also described the purification, crystallization and different properties of α-

Chapter 3 Results
56
glucosidase from the mycelia of Mucor (Wei et al., 2013) α-glucosidases from
Aspergillus niger and Asp. Nidulans have so far been isolated from culture filtrate.
While α-glucosidase is also reported from the different species of Penicillium (Hsu et
al., 2013; Liu et al., 2014).
Even though, the diversity of fungal species estimated about 1.5 million and a very
little number is identified (Hawksworth, 1991) only few genera like Aspergillus,
Penicillium, Rhizopus and Trichoderma etc. produce more than 50% fungal enzymes
used in various industrial processes (Ostergaard and Olsen, 2010). The wide variety
uses of these enzymes enhance the interests of many researchers for the exploration of
a low coast and sustainable resource. In connection to this, the endophytic microbial
floras have the potential to encode for these enzymes. In our research, the isolated
endophytic fungi showed very noticeable results for phosphatase production by
endophytic fungi. The productions of phosphatase were significantly higher in P.
citrinum, P. variotii and P. spinulosum. The strain P. purpurogenum, A. caespitosus
and Alternaria sp. also produce significantly higher amount of these enzymes. While
A. nidulans, F. oxysporum and Alternaria sp. comparatively encodes for a very little
amount of phosphates enzymes.
There are different physical, chemical and enzymatic processes known for the
hydrolysis of cellulose. In addition to, the cellulases enzymes produced by endophytic
fungi Pycnoporus sanguineus isolated from Baccharisdra cunculifolia also showed
very proficient results in the hydrolysis of cellulose (Onofreet et al., 2015). While in
our results the isolated endophytic strains revealed significant results for most of the
strains. The production of cellulases enzymes was much higher in P. variotii followed
by F. oxysporum, A. caespitosus and P. spinulosum. While the endophytic fungal
strains A. nidulans and P. purpurogenum encodes for a very little amount of these
enzymes. It was also observed that by keeping the pH at 5.5 subsidise in the greater
amount of cellulase production in liquid media.

Chapter 3 Results
57
3.8 Extraction and Purification of Compounds
The cultural filtrates were extracted by implying the equal volume of ethyl acetate
(EtOAc) three times. Both the extracts were compiled and concentrated in vacuo to
get the crude extract. The extracts were compiled and dried over sodium sulfate
(anhydrous) and concentrated in vacuo to afford the crude extract. The EtOAc extract
was then subjected to silica gel column chromatography using gradients of ethyl
acetate/n-hexane system to afford five fractions (Fr.A to Fr.E). Fr.E was further
subjected to recycling preparative HPLC (JAI) analysis. Compound (mg) was purified
at a retention time of 23 min by using ethyl acetate/n-hexane (6:4) in a silica gel
column with the flow rate 3.5 mL/min after five recycles.
3.9 Chromatography and Spectroscopic Techniques for Identification
The mycelial mats and the culture filtrate were extracted completely with ethyl acetate
(EtOAc). Both the extracts were compiled and dried over sodium sulfate (anhydrous)
and concentrated in vacuo to afford the crude extract (7.5 g). The ethyl acetate extract
(7.5 g) of the endophytic fungus Penicillium citrinum was then subjected to repeated
column chromatography (silica gel, n-hexane, DCM/n-hexane, and MeOH/DCM) to
get various sub-fractions (PC1-PC14). The sub-fraction PC12, which was obtained at
5% MeOH/ DCM were subjected to recycling preparative High Performance Liquid
Chromatography (HPLC) by JAI for the final purification. Compound 1 (3.8 mg) was
obtained at a retention time of 27 min through a 1H/2H column by using chloroform
at a flow rate of 3.5 mL/min. Along with some semi-pure compounds.These semi-
pure compounds were loaded on preparative TLC plates and the known compounds-
4(12.5 mg) and 5 (14.1 mg) were purified at DCM/n-hexane (90:10) and DCM/n-
hexane (95:5), respectively. The sub-fraction PC5 afforded compound 2 (3.3 mg) and
3 (2.7 mg) by repeated silica gel column chromatography at DCM/n-hexane (15:85)
and DCM/n-hexane (20:80), respectively.

Chapter 3 Results
58
Compound 1: Amorphous powder, 1H-NMR (600 MHz, CDCl3), δ 7.34-7.31 (5H,
overlap signals, H-33 to H-37), 5.54 (1H, br s, H-12), 5.14 (1H, d, J = 12.7 Hz, H-
31a), 5.04 (1H, d, J = 12.7 Hz, H-31b), 2.31 (1H, s, H-9), 1.51 (1H, br s, H-18), 1.37
(3H, s, CH3-23), 1.24 (3H, s, CH3-27), 1.15 (6H, s, CH3-25 and CH3-26), 0.92 (3H,
overlap signal, CH3-30), 0.80 (3H, s, CH3-28), 0.76 (3H, d, J = 6.3 Hz, CH3-29). 13
C-
NMR (150 MHz, CDCl3), δ 208.4 (C-3), 198.9 (C-11), 173.1 (C-24), 165.5 (C-13),
130.3 (C-12), 128.6-128.5 (C-32 to C-37), 66.9 (C-31), 59.9 (C-9), 59.0 (C-18), 58.5
(C-5), 57.7 (C-4), 53.4 (C-4), 44.8 (C-14), 43.8 (C-8), 41.0 (C-22), 40.9 (C-1), 39.3
(C-19 and C-20), 37.1 (C-10), 36.7 (C-2), 33.9 (C-17), 32.7 (C-7), 28.8 (C-28), 21.1
(C-23 and C-30), 20.4 (C-27), 18.3 (C-26), 17.4 (C-29), 13.4 (C-25). ESI-MS m/z:
581.06 [M + Na]+for C37H50O4Na (Appendix- I).
Compound 2: Colorless oil, 1H-NMR (600 MHz, CDCl3), δ 1.40-1.23 (14H, overlap
signal, H-2 to H-8), 0.86 (6H, t, J = 7.1 Hz, CH3-1 and CH3-9). 13
C-NMR (150 MHz,
CDCl3), δ 31.9-29.4 (C-2 to C-8), 14.1 (C-1 and C-9). ESI-MS mlz: 150.79 [M +
Na]+for C9H20Na (Appendix- II).
Compound 3: Colorless lequid, 1H-NMR (600 MHz, CDCl3), δ 5.32-5.19 (2H,
overlap signal, H-3 and H-4), 3.63 (2H, m, H-1), 1.53 (2H, m, H-2), 0.83 (3H, t, J =
4.8 Hz, H-10). 13
C-NMR (150 MHz, CDCl3), δ 130.9 (C-4), 128.9 (C-3), 66.5 (C-1),
32.8 (C-2), 29.3-29.0 (C-5 to C-9), 14.1 (C-10). ESI-MS mlz: 178.94 [M + Na]+for
C10H20ONa(Appendix- III).
Compound 4: Gummy solid, 1H-NMR (600 MHz, CDCl3), δ7.18 (1H, t, J = 8.6 Hz,
H-4), 7.12 (1H, d, J = 7.3 Hz, H-6), 6.90 (1H, d, J = 8.8 Hz, H-3), 6.89 (1H, t, J = 7.4
Hz, H-5). 13
C-NMR (150 MHz, CDCl3), δ 173.1 (C-8), 154.5 (C-2), 131.2 (C-6),

Chapter 3 Results
59
129.3 (C-4), 123.1 (C-1), 121.2 (C-5), 117.3 (C-3), 36.8 (C-7). ESI-MS mlz: 150.79
[M + Na]+for C8H8O3Na (Appendix- IV).
Compound 5: Colorless powder, 1H-NMR (600 MHz, CDCl3), δ7.48 (2H, dd, J =
8.1, 11.2 Hz, H-4 and H-5), 7.23 (2H, d, J = 7.4 Hz, H-3 and H-6), 5.64 (1H, d, J =
7.3 Hz, H-1 of glu), 4.41-3.42 (6H, overlap signal, H-2 to H-6 of glu). 13
C-NMR (150
MHz, CDCl3), δ 174.5 (C-8), 150.1 (C-2), 127.8 (C-3 and C-6), 126.4 (C-4 and C-5),
124.1 (C-1), 101.5 (C-1 of glu), 77.2-70.5 (C-2 to C-5 of glu), 63.4 (C-6 of glu), 39.4
(C-7). ESI-MS mlz: 336.84 [M + Na]+for C14H18O8Na (Appendix- V).
3.10 Characterization of Compounds
The ethyl acetate extract (7.5 g) of the endophytic fungus Penicillium was subjected
to repeated column chromatography followed by the final purification through
recycling HPLC to afford five known compounds: 11-oxoursonic acid benzyl ester
(1), n-nonane (2), 3-decene-1-ol (3), 2-Hydroxyphenyl acetic acid (4), and
Glochidacuminosides A (5).

Chapter 3 Results
60
O
O
O
CH3
CH3 CH3
H3C
CH3
CH3
H3C
H
H
O
OH
OH
O
OGlu
OH
O
OH
1
2
3
1211
10
9
8
76
54
19
18
17
16
13
1514
2021
22
23 24
25 26
27
28
29
30
1
4
3132
33
34
35
36
37
5
2
3
12
38
7
654
12
38
7
654
Figure 3.5:- Structures of Compounds 1-5: 11-oxoursonic acid benzyl ester (1), n-nonane
(2), 3-decene-1-ol (3), 2-Hydroxyphenyl acetic acid (4), Glochidacuminosides A (5)
Compound 1 was isolated in the form of amorphous powder which gave characteristic
pink color on the TLC plates after spraying with ceric sulphate reagent (Figure 3.5a).
This indicates the terpenoid skeleton in the molecule. The 1H NMR spectrum of 1
displayed seven methyl signals at 1.37, 1.24, 1.15, 1.15, 0.92, 0.80, and 0.76, which
were indicative of ursane skeleton. The olefinic proton H-12 appeared as broad singlet

Chapter 3 Results
61
at δ5.54, indicating the presence of trisubstituted double bond. The downfield 1H
NMR absorptions at δ5.14 and 5.05 (1H each, d, J = 12.7 Hz, H-31) and HMBC
correlation of H-31 to C-28 (δ173.1), and the aromatic carbons (C-32 to C-37),
showed the presence of benzyl ester at C-28. The presence of conjugated ketone
moiety at C-11 was evident by the 13
C NMR signals and the HMBC interactions of H-
12 with C-11 (δ198.9) and C-13 (δ165.5). Another downfield signal at δ208.4 was
assigned to the ketone functionality at C -3. The study of 13
C NMR (BB and DEPT)
data showed signals for seven methyls, nine methylenes, eleven methines and ten
quaternary carbons for a triterpene skeleton having O-benzyl substitution. Further
comparison of the spectral data with the literature valuesestablished the structure of
compound 1 as benzyl ester of 3,11-dioxo-ursolic acid. This compound has earlier
been reported in a Chinese patent on ursolic acid derivatives.
Figure 3.5a: Demonstrate Compound 1; 11-Oxoursonic acid benzyl ester
Compound 2 was isolated as a colorless oil and showed pseudo molecular ion peak at
m/z 150.79 corresponding to the molecular formula C9H20Na for the sodium adduct.
The 1H NMR spectrum of 2 showed a triplet for six protons due to two terminal
methyl groups (CH3-1 and CH3-9) at δ 0.86 (6H, t, J = 7.1 Hz). The methylenes

Chapter 3 Results
62
moieties associated with the long chain of 2 appeared as overlap signal at δ 1.40-1.23.
The 13
C NMR spectrum of compound 2 showed signals due to two terminal methyl
groups in the aliphatic hydrocarbon chains at δ 14.1, whereas the remaining
methylenes of the chain appeared at a range from δ 31.9 to 29.4. The final structure of
2 as n-nonane was confirmed through comparison with the literature data.
Figure 3.5b: Demonstrate Compound 2; n-nonane
Compound 3 was isolated as a colorless liquid. The molecular formula C10H20O was
established on the basis of MS, 1D and 2D NMR spectra. The ESI-MS showed the
pseudo molecular ion peak at m/z 178.94, suggesting the molecular formula
C10H20ONa in the form of sodium adduct. The 1H NMR spectrum displayed a triplet
at δ 0.83 (3H) and a broad singlet at δ 1.23 corresponding to the straight chain
hydrocarbon. The presence of hydroxyl group at C-1 was also inferred from 1H NMR
spectrum which displayed signals at δ 3.63 (αH, H-1) which showed the presence of
hydroxyl at C-1 position. The olefinic double bond at C-3 was indicated by the
presence of overlap signal between δ 5.32-5.19, which was further supported through
13C NMR spectrum. The
13C NMR spectrum (BB and DEPT) showed characteristics
signals due to oxygenated methylene at δ 66.5, whereas the down field signals at δ
130.9 and 128.8 were assigned to the double bond at C-3 position. The signals
observed between δ 29.3 and 29.0 indicated the presence of a long chain hydrocarbon.
The position of the double bond and the hydroxyl group was further confirmed
through mass fragmentation as well as from the HMBC correlations. Thus, on the
basis of above discussion coupled with the literature value, compound 3 was
confirmed as 3-decene-1-ol.

Chapter 3 Results
63
Figure 3.5c: Demonstrate Compound 3;3-decene-1-ol
Compound 4 was obtained as colorless powder. The molecular formula C8H8O3 was
deduced from MS and 13
C NMR spectral data. The ESI-MS showed the pseudo
molecular ion peak at m/z 175 [M + Na]+, suggesting the molecular formula C8H8O3 for
compound 4. The analysis of 1H NMR spectrum revealed the presence of a singlet for a
methylene moiety at δ 3.69 and four signals for one proton each in the downfield
aromatic region (δ 7.18 to 6.89). These aromatic signals showed splitting in the form of
two doublets (δ 7.12, J = 7.3 Hz and 6.90, J = 8.8 Hz) and two triplets (δ 7.18, J = 8.6
Hz and 6.89, J = 7.4), which indicated the presence of ortho-disubstituted benzene ring
in compound 4. The 13
C NMR spectrum (BB and DEPT) indicated the presence of eight
signals which were resolved into one methylene, four methane and three quaternary
carbons. The downfield signal at δ 173.1 was assigned to the carboxylic functionality
whereas the methylene carbon appeared at δ 36.8. The sp2 methine carbons of the
aromatic ring appeared at δ 131.2, 129.3, 121.2, and 117.3, whereas the signals at
δ154.5 and 123.1 were assigned to C-2 and C-1 respectively. The position of individual
groups in the molecule was further confirmed through HMBC interactions. Thus, the
structure was finally confirmed as 2-Hydroxyphentlacetic acid which was in complete
agreement with the reported values.

Chapter 3 Results
64
Figure 3.5d: Demonstrate Compound 4; 2-Hydroxyphenyl acetic acid
Compound 5 was isolated as a gummy solid and the structure was assigned on the
basis of comparison of the MS and NMR data in the reported literature. The ESI-MS
showed the pseudo molecular ion peak at m/z 336.84 [M + Na]+, which suggested the
molecular formula C14H18O8 for compound 5. The spectral data of compound 5 was
similar to that of compound 4 with the additional signals for a sugar moiety in the
molecule. The aromatic signals showed splitting in the form of a doublet at δ 7.23
(2H, d, J = 7.4 Hz, H-3 and H-6) and a doublet of doublets at δ7.48 (2H, dd, J = 8.1,
11.2 Hz, H-4 and H-5), which indicated the presence of ortho-disubstituted benzene
ring in compound 5. The anomeric proton was observed at δ 5.64 as doublet (J = 7.3
Hz), indicating the beta linkage of the sugar. The remaining protons of the sugar
moiety were observed at δ 4.41 to 3.42. The assignments of the sugar moiety were
further confirmed through 13
C NMR spectra which indicated the presence of anomeric
carbon at δ 101.5, a methylene carbon at δ 63.4 and four methine signals at δ 70.5 to
77.2. The identity of the sugar moiety was thus indicated as β-D-glucopyranoside
which was further confirmed through co-TLC with the authentic sample. The position
of the sugar moiety and the side chain was further confirmed through HMBC
interactions. The structure was finally established as 2-Hydroxyphentlacetic acid-2-O-
β-D-glucopyranoside, previously isolated from the leaves of Glochidionacuminatum
and commonly known as glochidacuminosides A.

Chapter 3 Results
65
Figure 3.5e: Demonstrate Compound 5; Glochidacuminosides A
3.11 Enzyme Inhibitory Activities of Secondary Metabolites
Our aforementioned screening for α-glucosidase inhibition assay showed that the
highest α-glucosidase inhibitory activity was revealed from ethyl acetate extract of
endophytic fungi P. citrinum (FEF6).The α-glucosidase inhibitory activities of the
isolated bioactive compounds from endophytic fungus P. citrinum FEF6 demonstrates
that the characterized compounds 2-Hydroxyphenyl acetic acid (4), 11-oxoursonic acid
benzyl ester (1) and Glochidacuminosides A (5) endure high level of potential for the
inhibition of α-glucosidase (Figure 3.6). Among these compounds, compound 1 showed
anIC50 273.87±1.59 μg/mL. The urease inhibition assay showed that compound 2, 3 and
4 have a very weak role in urease enzyme inhibition while compound1 (11-oxoursonic
acid benzyl ester) and 5 (Glochidacuminosides A) shows high level of urease inhibitory
activities (Figure 4).
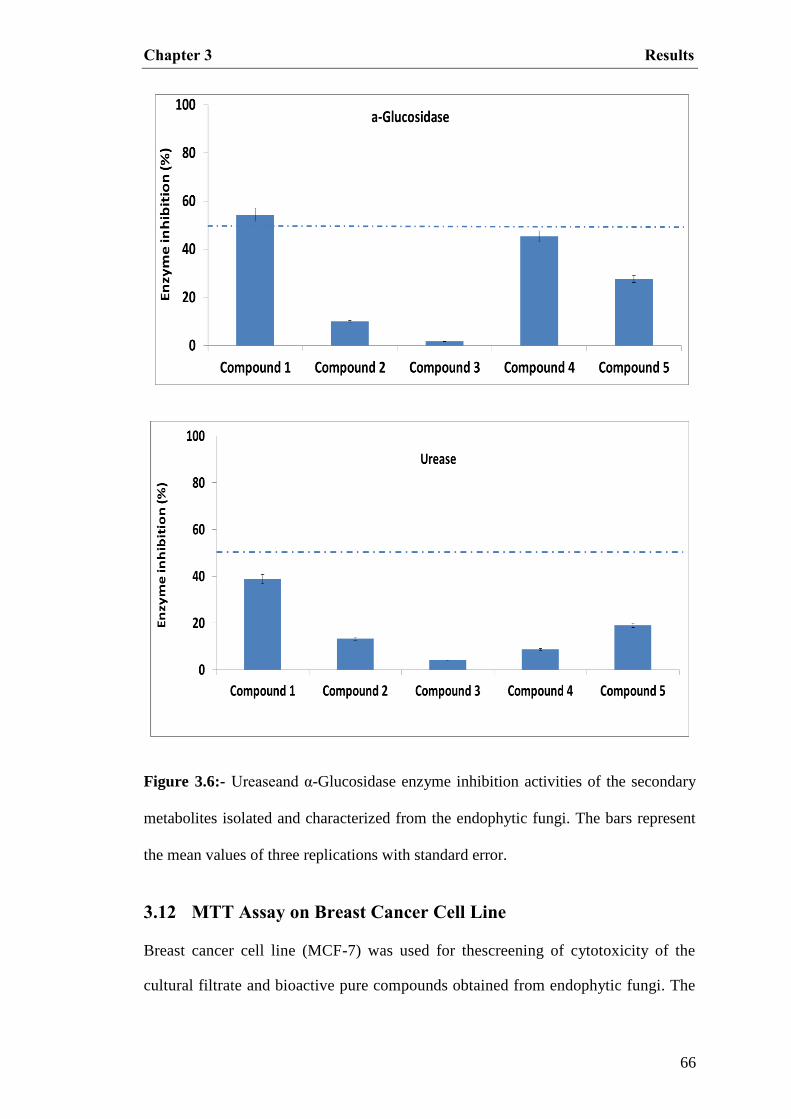
Chapter 3 Results
66
Figure 3.6:- Ureaseand α-Glucosidase enzyme inhibition activities of the secondary
metabolites isolated and characterized from the endophytic fungi. The bars represent
the mean values of three replications with standard error.
3.12 MTT Assay on Breast Cancer Cell Line
Breast cancer cell line (MCF-7) was used for thescreening of cytotoxicity of the
cultural filtrate and bioactive pure compounds obtained from endophytic fungi. The

Chapter 3 Results
67
cell lines were cultured in advanced DMEM with 10% NBCS (inactivated) and5mM
l-glutamine, and then grown at 37Ċ in a humidatmosphere with 5% CO2 in air. The
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium-bromide(MTT) colorimetric
assay developed by Mosmann, 1983. Anticancer activities of endophytic fungal
cultural filtrate and pure compounds were assessed by using breast cancer cell
(MCF-7) lines. Various low concentrations of CF were prepared and tested against
the grown cell cultures. The screening results showed that P. citrinum, P.
purpurogenum and P. variotii extract hasgreatly reduced the cancer cell viability as
compared to other extracts (Figure 3.7).
Figure 3.7:- Effect of cultural filtrate of endophytic fungi on the viability of MCF-7
breast cancer cells in culture
Breast cancer cell line, MCF-7, was incubated with indicated concentrations of the
samples from endophytic fungi for 24 hours. The effect on cell proliferation was
detected by performing MTT assay as described in „Methods‟ section. All results are
expressed as percentage of control ± SE of triplicate determinations.
0
10
20
30
40
50
60
70
80
90
F1 FEF1 FEF5A FEF6 FEF6B FEF7 FEF2
150µg/ml
300µg/ml
Ce
ll V
iab
ility
(%
)

Chapter 3 Results
68
Our aforementioned screening for MTT assay showed that the highest activity was
shown in the ethyl acetate extract of endophytic fungi P. citrinum (FEF6), P.
purpurogenum (FEF5A), and P. variotii (F1).
The activities of the isolated bioactive compounds from endophytic fungus P.
citrinum (FEF6) demonstrate that the characterized compounds 2-Hydroxyphenyl
acetic acid (4), 11-oxoursonic acid benzyl ester (1) and Glochidacuminosides A (5)
endure a significantlyhigher potential to inhibit cell proliferation.The compound1 (11-
oxoursonic acid benzyl ester) and 5 (Glochidacuminosides A), show significantly
higher level of cells growth inhibition (Figure 3.8).
Figure 3.8:- Effect of pure compounds on the viability of MCF-7 breast cancer cells
in culture.
0
20
40
60
80
100
120
Control C1-PBLT C4-PCF-5D C5-PCF11
100. 50 μg/ml
100 μg/ml

Chapter 4 Discussion
69
DISCUSSION
Section 1: Endophyte Diversity Assessment
In present research work, a range of endophytic fungi were evaluated by using
denaturing gradient gel electrophoresis (DGGE) for the identification of various
endophytic fungi. The medicinally valuable plants „Caralluma acutangula and
Boswellia sacra’ offered an array of endophytic fungi. The surface-sterilized samples
were assessed by means of isolation techniques and phylogenetic analysis. With these
approaches we were able to detect differences in the relative abundance of endophytic
fungi.The rate of colonization was higher in the stem part of C. acutangula while
leaves of B. sacra showed higher colonization rate.
Mycorrhizal fungi colonize in near vicinity of plant roots or penetrate into plant root
tissues in case of endomycorrhiza, whereas endophytic fungi reside within all tissues
of the plants and are symptomless (Khan et al.,2013; Stone et al., 2004). Historically,
endophytic fungi were divided into two major groups „Clavicipitaceous and
Nonclavicipitaceous endophytes‟ on the basis of their hosts, taxonomy, ecological
function and phylogenetic analysis. However, nonclavicipitaceous endophytes are
highly diverse in nature and now further grouped into three different classes, based on
their life history and ecological consequences (Arnold, 2007). Endophytic fungi
greatly contribute in stress conditions and favours host growth regardless of the
habitat of origin. Diverse kinds of fungal endophytes establishes important niche with
plants through secretion of bioactive constituents, regulating the growth of host even
underharsh environmental conditions. Endophytic microbes associated with
medicinally important plants have been shown to enhance host plant growth and
development. This could possibly be owing to the production of various constituents

Chapter 4 Discussion
70
during the symbiotic processes with the host plants (Khan et al., 2013; Kusari et al.,
2013). Endophytes are also able to colonize more than one host species of the same
plant family within the same habitat. Endophytic fungi often shows single host-
specificity at plant species level which could be also influenced by environmental
conditions (Cohen 2006).
To further understand the interactions in in-vitro environments of culturable
endophytes from medicinal plants, we isolated and identified endophytic fungal
species from C. acutangula. To our knowledge, there have beenno previous studies on
this plant and its associated endophytic microorganisms. In the current study, we
grouped endophytic fungi on the basis of colony shape, thickness, colour of aerial
hyphae, growth rate and pattern, margin characteristics, surface texture, and growth
depth into medium (Arnold et al., 2007). The endophytes were identified by genomic
DNA extraction, PCR techniques, nucleotide sequencing, and phylogenetic analysis
as described by Khan et al., (2011).
We isolated and identified various strains of Paecilomyces, Epicucum, Fusarium,
Penicillium, Aspergillus, and Alternaria species using molecular techniques. Based
on ITS sequence homology, we identified these fungal strains as A. nidulans, P.
variotii, F. oxysporum, E. nigram, P. purpurogenum and Alternaria sp. These
strains were assessed for their potential to produce ACC deaminase, extra-cellular
enzymesand IAA during their growth. The advantageous effects of endophytic fungi
and bacteria have been regarded for their potential to produce biologically active
secondary metabolites and other substances. The production of such substances can
maintain their own metabolism as well as supports the growth of their hosts
(Saxena, 2014). These substances can range from secondary metabolites to various

Chapter 4 Discussion
71
enzymes. In case of metabolites, endophytic microbes have recently been also
known to produce different plant hormones (Hardoim et al., 2008). The distribution
of identified endophytic fungi in plant organs differs in diversity and abundance. As
our results showed that higher colonization rate was observed in the stem (2.41) of
C. acutangula, whereas for leaves and roots was 1.26 and 1.11, respectively. The
isolation rate was much higher (1.82) for the stem of C. acutangula. The diversity
indices analyzed by Shannon-Weiner (𝐻1= 0.8424) and Simpson (1/𝑙= 1.662)
indices indicated differences in plant parts and species richness (Wei et al., 2013).
While the leaves of B. sacra showed higher colonization rate. The study of Maria et
al., (2005) also reported fungal endophytes from medicinal plants and isolated 18
endophytic fungi from the stem and leaf segments of host plants. The dominant
species were Curvularia clavata, C. lunata, C. pallescens and F. oxysporum. The
highest species richness as well as frequency of colonization of endophytic fungi
was found in the leaf segments rather than the stem and bark segments of the host
plant species. The greatest numbers (11 species) of endophytic fungi were found
within Callicarpato mentosa, whereas Lobeliani cotinifolia harbored the lowest
number of fungal endophytes (5 species). The study also provides evidence that
fungal endophytes are host and tissue specific (Mariaet al., 2005).
However, the biodiversity of fungal species is estimated at about 1.5 million, a limited
number of species have been identified, includingonly fewgenera, such as Aspergillus,
Penicillium, Rhizopus, and Trichoderma, which produce >50% of the fungal enzymes
used in various industrial processes (Rytioja etal., 2015; Gawas-Sakhalkar etal.,
2012).The wide variety of uses of these enzymes has enhanced the interest of many
researchers to explorethese as a low cost, sustainable resource.

Chapter 4 Discussion
72
Section 2: Potential Role of Endophytes
The endophytic fungi from Caralluma and Boswellia species are rarely employed for
endophytic fungal diversity, extracellular enzymes production and their bioactive
compounds. The genus Penicillium is most prevalent followed by Aspergillus, Alternaria
and Paecilomyces in C. acutangula, while Phoma, Fusarium and Aspergillus were
screened from B. sacra. Recently, Penicillim species has been also reported from
different medicinal plants (Nagerabi et al., 2014). Penicillium is an important genus of
ascomycetous fungi which greatly contribute in the production variety of drugs and a
large number of its species produce mycotoxins (Diblasi et al., 2015).
The most famous and economically important being penicillin is produced by Penicillium
chrysogenum. Mycophenolic acid produced by Penicillium brevicompactum and
Compactins produced by Penicillium solitum, while several other secondary metabolites
from Penicillium species showed potential anticancer activities including alternariol,
fumitremorgin C, hadacidin, paclitaxel, penicillic acid, PR-toxin, and viridicatumtoxin.
Fumitremorgin C and paclitaxel appear to show most promise in cancer treatment
(Samson and Frisvad 2004; Houbraken et al., 2011). In 2011 five new di- and tri-citrinols
were added to the known citrinin family from P. citrinum all of them showed cytotoxic
activity against human leukemia HL-60, colon cancer HCT-116, and cervical cancer KB
cell lines. The endophytic fungus Penicillium chrysogenum was reported by Zhou et al.,
in 2011 obtained from Lycopodium serratum have the ability to produce huperzine A, the
drug which is used for Alzheimer's disease, memory and age-related memory impairment
(Zhou et al., 2011). The P. citrinum as an endophyte with bioactive compounds in the
bark of Taxus cuspidate has been also reported by Yong et al.,in 2003.
The genus Paecilomyces comprises a number of species that are able to produce a wide
collection of bioactive secondary metabolites of different chemical classes and with
different biological activities, such as antimicrobial and cytotoxic activities (Kyong et

Chapter 4 Discussion
73
al., 2001; Wang et al., 2002). Initially, the production of bioactive secondary
metabolite polygalactosamine from Paecilomyces sp. with anti-tumor activity was
reported by Ishitani et al., in 1988. The species of Paecilomyces marquandii was
reported by Radics et al. (1987) for the production of leucinostatins A, D and K as
peptide antibiotics. Similarly, the antitumor antibiotics; saintopin and UCE1022 with
topoisomerase- dependent DNA cleavage activity were described in last ten to fifteen
years (Long and Balasubramanian 2000; Fan et al., 2001). In some of the reported
studies Paecilomyces sp. have been used for the production of leu-cinostatins A, D, H
and K as peptide antibiotics, polygalactosamine which showed antitumor activities;
saintopin as antitumor antibiotics with topoisomerase dependent DNA cleavage
activity. Similarly, Aspergillus has usually been found as a saprophytic fungus, and
producing a variety of mycotoxins such as patulin and cytochalasin E (Liu et al., 2009;
Lopez-Diaz, T.M. and Flannigan 1997). The apoptotic antitumor activity antioxidant
and immune-stimulating activities of Paecilomyces japonica were reported by Fujji et
al., (1994), while the Paecilomyces variotii has been exploited by sachan et al., (2006)
for the biotransformation of p-coumaric acid (Nam, 2001). Recently, a new compound
3H-oxepine-containing alkaloid, varioxepine A, was isolated from endophytic
fungus Paecilomyces variotii. This compound has the ability to inhibit the growth of
some pathogenic fungus (Zhang et al., 2014; Zhang et al., 2015).
The relationships exist between endophytes and their host plants, ranging from
mutualism or symbiosis to antagonism or slight pathogenesis (Schulz and Boyle,
2005; Khan et al., 2013).Fungal endophytes have also a varied relationship between
pathogen and host plant and alsoinfer how they limit pathogen damage to the host
(tropical trees). Endophytes represent a ubiquitous yetcryptic component of terrestrial
plant communities. The fundamental aspects of endophytic interactions with hosts are

Chapter 4 Discussion
74
unknown. In contrast to vertically transmitted endophytes, horizontally transmitted
endophytes of woody angiosperm are thought to contribute little to host defence
mechanism. The fungal endophytes helpto decrease both leaf necrosis and leaf
mortality when T. cacao seedlings are challenged with pathogen (Phytophthora sp.).
Further endophyte mediated protection was greater in mature leaves, which bear less
intrinsic defence against fungal pathogens than do young leaves (Arnold et al., 2003).
Endophytes may have developed friendly relationships with their hosts during
evolution and may be host or tissue specific. Host or tissue specificity of endophytic
fungi must depends on certain factors i.e. endophyte colonization or active ingredients
within host tissues (Arnold, 2007). Fungal endophytes produce bioactive metabolites
that mediate in the plant–endophyte interaction (Strobel, 2003). In addition, fungal
endophytic metabolites are useful resources for natural products which effectively
have wide range of application in medicine, agriculture, and industry (strobel and
Daisy 2003; Selim et al., 2012). Fungal endophytes have the ability to produce
numerous extracellular enzymes; such as pectinases, cellulases, lipases, amylases,
laccases, and proteinases. These fungal enzymes play the key role in biodegradation
and hydrolysis processes which are significantly important mechanisms against
pathogenic infection and to obtain their nutritional need from the host plants (Sunitha
et al., 2013).
Many bioactive metabolites are originated from microbial organisms, fungi are the
core important groups of eukaryotic organisms that have wide capacity to produce
numerous metabolites with antimicrobial activities and possess potential application
as drugs. Phytohormones such as auxins are produced in the different parts of the
plant and these are mainly responsible for the regulation of plant developmental

Chapter 4 Discussion
75
processes. The array of IAA within various parts of the plant is a key factor for
plant growth and at the same time its additional supply can support the host in
stress condition.
In our study, P. variotii and A. nidulans showed considerable IAA production. The
production of IAA is dependent on the type of pathway utilized by endophytes;
therefore, IAA in the culture broth was estimated by the presence and absence of L-
tryptophan. Similar results for IAA were found by Mei and Flinn, where IAA
producing endophytic fungi improved plant growth under stress conditions. Thus,
previous findings about IAA-producing endophytic fungi strongly support our current
results. The results conclude that endophytic fungi associated with medicinally
important plants possess a unique potential to produce bioactive metabolites, which
might improve the growth of the host in severe environmental conditions. Such
bioactive fungal strains may have applications for crop improvement and industrial
production of important constituents (Mei and Flinn 2010; Fu et al., 2015).
The functional role of IAA in plant growth in addition to the capacity of fungal
endophytes to produce IAA has gained great attention due to their impact on the
concentration and distribution of IAA in plant tissues. Little is known about the
biology and ecology of fungal endophytes; subsequently, isolation and
characterization of fungal endophytes that colonize different plant species of various
habitats and ecosystem is potentially useful.
Section 3: Bioactive Metabolites from Endophytes
Endophytes may yield a plethora of bioactive secondary metabolites that may be
involved in the host-endophyte relationship and also have the capability to encode for
the same type of metabolites as produced by their host plant (Strobel 2004). Natural

Chapter 4 Discussion
76
products from endophytic microbial flora have a broad spectrum of biological
activities i.e. antifungal, antibacterial, antidiabetic and anticancer etc. Therefore,
bioactive secondary metabolites from endophytic fungi can be grouped into several
categories like steroids, alkaloids, terpenoids, phenol and lignin (Qawasmeh et al.,
2012; Xiao et al., 2014). There are different Endophytic fungi which are reported
from genus Boswellia. The genus Alternaria is the most prevalent followed by
Aspergillus and Rhizopusstolonifer. Recently, Penicillim species has been also
reported from Boswellia (Nagerabi et al., 2014).
In our study we quantified the endophytic fungal extracts and tested for α-glucosidase
inhibition which revealed significant inhibition for α-glucosidase enzymes.The ethyl
acetate extract of P. citrinum (FEF6) and P. spinulosum (FEF2) showed prominent
results. While in urease inhibitory activity of endophytic fungi extracts the isolated
endophytic fungi showed more than 60% rate of inhibition. The endophytic fungus P.
citrinum (FEF6) revealed increased rate of inhibition.Previously, Singh et al., (2015)
evaluated the maximum inhibitory activity of an endophyte Cladosporium sp. isolated
from T. cordifolia (TN-9S) they observed thatpurified inhibitor is a phenolic
compound with amine groups and it can also inhibited the activity of α-glycosidase in
vivo condition. There is a variety of organisms which encodes for the production of
urease enzymes including plants, fungi, bacteria and invertebrates, and it also occurs
in soils as a soil enzyme (Krajewska 2009).Khan et al., (2015) isolated the endophytic
fungus Bipolaris sorokiniana LK12 from the leaves of Rhazyastricta. They
characterized bipolarisenol from B. sorokiniana evaluated for its potential role in
urease inhibition. Their results suggest that Bipolarisenolinhibit urease in a dose-
dependent manner with high IC50 (81.62 ± 4.61 µg·mL−1).

Chapter 4 Discussion
77
In addition, cellulase enzymes produced bythe endophytic fungi, Pycnoporus
sanguine isolated from Baccharis dracunculifolia effectively hydrolyzed cellulose
(Onofreet al., 2015). The production of cellulase enzymes was considerable higher in
F. oxysporum, whereas it was the lowest inAlternaria sp. P. variotii and P.
purpurogenum produced high amounts of cellulases, followed by A. nidulans and
Alternaria sp. It was also observed that by keeping the pH at 5.5, a greater amount of
cellulase was produced in liquid media.
It has been reported that endophytic microbial flora can significantly contribute to the
production of phytohormones (Marwah et al., 2007; Kuldau G and Bacon C, 2008). In
our study, P. variotii and A. nidulans showed considerable IAA production. The
production of IAA is dependent on the type of pathway utilized by endophytes;
therefore, IAA in the culture broth was estimated by the presence and absence of L-
tryptophan.Similar results for IAAwere foundby Mei and Flinn (Mei and Flinn, 2010),
Where IAA producing endophytic fungi improved plant growth under stressconditions.
Thus, previous findings about IAA-producing endophytic fungi strongly support our
current results (Fu et al., 2015).The results conclude that endophytic fungi associated
with medicinally important plants possess a unique potential to produce bioactive
metabolites, which might improve the growth of the host in severe environmental
conditions. Such bioactive fungal strains may have applications for crop improvement
and industrial production of important constituents.
The production of enzymes such as ACC (1-aminocyclopropane-1-carboxylate)
deaminase is common in bacteria while some of fungal strains also encoding genes for
ACC deaminase. The fungal stain P. citrinum and a very few other fungal strains are
documented for the production of ACC (Jiaet al., 1999, Yim et al., 2013). The

Chapter 4 Discussion
78
endophytic microbial flora also contain the enzyme 1-aminocyclopropane-1-
carboxylate (ACC) deaminase and this enzyme can cleave the ethylene precursor ACC
to α-ketobutyrate and ammonia and thereby lower the level of ethylene in developing or
stressed plants (Hontzeas et al., 2005). ACC deaminase-producing endophytes promote
plant growth under stress conditions and also playing a very important role in the
process of nodulation, specific to plant species.
Our results suggested a significantly higher amount of ACC deaminase production by
P. purpurogenum and P. variotii strains. Of the endophytic fungal strains grown in DF
minimal medium containing ACC, P. purpurogenum showed the highest productionof
ACC at 355nmol α- ketobutyrate mg-1
h-1
among the isolated strains. Previously, P.
citrinum and some other fungal strains were found to produce ACC (Yim et al., 2013;
Jia et al., 1999). The endophytic microbial flora also contained the enzyme 1-
aminocyclopropane-1-carboxylate (ACC) deaminase which was confirmed by the
total protein content inthe pure growth cultures of P. purpurogenum and P. variotii.
However, extracellular enzyme production by endophytes isessential to help the
microorganism penetrate the host cell wall, and can contribute to the defence
responses and mineral uptake in host plants (Weiet al., 2013). Enzymes, such as β-
glucosidases, cellulases, and phosphatases, were determined using florigenic
substrates and standard curve readings were taken in combination with MUB
standards. In our study, the extracellular enzyme, β-glucosidase, encoded by
endophytic fungi isolated from C. acutangula revealed significant results; P. variotii
and P. purpurogenum produced higher amounts of β-glucosidases than other strains.
The previous data also described the purification, crystallization, and different
properties of α-glucosidase from the mycelia of Mucor (Kato et al., 202), andα-

Chapter 4 Discussion
79
glucosidases from Aspergillus niger and A. nidulans (Hsu et al., 2013) have been
isolated from culture filtrates;Since α-glucosidase has also been reported from the
different species of Penicillium (Liu etal., 2010; Qureshietal., 2013).
In our study the ethyl acetate extract of endophytic fungus P. citrinum (FEF6) was
subjected to repeated column chromatography followed by recycling HPLC to givethe
compounds: 11-oxoursonic acid benzyl ester (1), n-nonane (2), 3-decene-1-ol (3), 2-
Hydroxyphenyl acetic acid (4), and Glochidacuminosides A (5). Compound 1 was
isolated in the form of amorphous powder which gave characteristic pink colour on
the TLC plates after spraying with ceric sulphate reagent. Compound 2 and 3were
isolated as a colourless liquids while Compound 4 and 5 were obtained as colourless
powder and as a gummy solid respectively. The α-glucosidase inhibitory activities of
the isolated bioactive compounds from endophytic fungus P. citrinum FEF6
demonstrates that the characterized compounds 2-Hydroxyphenyl acetic acid (4), 11-
oxoursonic acid benzyl ester (1) and Glochidacuminosides A (5) exhibited high level
of potential for the inhibition of α-glucosidase. While assay for urease inhibition
showed that compound 2, 3 and 4 have a very weak role in urease enzyme inhibition
as compare to compound 1 (11-oxoursonic acid benzyl ester) and 5
(Glochidacuminosides A) which shows a substantial level of urease inhibitory
activities. The anticancer activities for bioactive secondary metabolites showed that
the three compounds, 11-oxoursonic acid benzyl ester (1), 2-Hydroxyphenyl acetic
acid (4), and Glochidacuminosides A (5) from endophytic fungi Penicillium citrinum
bears a considerable level of inhibition of cancer cell proliferation and apoptosis using
human breast cancer cell line (MDA-MB-231). In our results the cell viability was
greatly arrested by 11-oxoursonic acid benzyl ester (1) followed by 2-Hydroxyphenyl
acetic acid (4). Similarly Hu et al., (2008) isolated xylarenones A, Band xylarenic

Chapter 4 Discussion
80
acid from endophytic fungus Xylaria sp. NCY2, obtained from Torreya jackii CHUN
and were evaluated for antitumor and antimicrobial assays in in-vitrocondition which
demonstrated that these compounds exhibit moderate antitumor activities against
HeLa cells line (Hu et al., 2008).

Chapter 4 Discussion
81
Section 4: Conclusion
In conclusion, diverse fungal endophytes have established important niches with plants
through the secretion of bioactive constituents, regulating host growth during harsh
environmental conditions. Endophytic fungi were isolated from medicinally important
plants i.e.Caralluma acutangula and Boswellia sacra. The isolated endophytic fungi
were identified on the basis of their morphological traits and by using genomic DNA
extraction, PCR amplification and sequencing the internal transcribed spacer regions,
whereas a detailed phylogenetic analysis of the same gene fragment was made with
homologous sequences. The endophytic fungi were identified as Penicillium citrinum;
Paecilomyces variotii, Aspergillus nidulans, Fusarium oxysporum, Epicucum nigram,
Penicillium purpurogenum, Penicillium spinulosum, Aspergillus caespitosus, Phoma
and Alternaria sp. Furthermore, the endophytic fungi P. citrinum, P. variotii,
Aspergillus nidulans, Fusarium oxysporum, Penicillium purpurogenum and Alternaria
sp. were assessed for their potential to produce anti-cancerous metabolites by
performing MTT assay and extracellular enzymes such as cellulases, phosphatases and
glucosidases in growth media. The P. variotii, P. citrinum and F. oxysporum showed
significantly higher amount of phosphatases and glucosidases as compared to other
strains. The NMR spectra were recorded on Bruker spectrometer operating at 600-MHz
(150-MHz for 13C) which give singals for five different bioactive secondary
metabolites from endophytic fungi. Additionally, endophytic P. citrinum, P. variotii
and F. oxysporum showed significantly higher potential of indole acetic acid
production.The above mentioned activities were for the first time elucidated for our
isolated endophytic fungi. We also concluded that the isolated endophytic fungi
produce bioactive constituents that could provide a unique niche of ecological
adaptation by symbiosis and greatly contribute to healthy life of their host plantand can
be utilized for future drug discovery.

References
82
REFERENCES
1. Abeles, F. B. Morgan, P. W and Saltveit Jr, M. E (2012). Ethylene in plant
biology. Academic press.
2. Amna, T. Puri, S. Verma, V. Sharma, J. P. Khajuria, R. K. Musarrat, J. Qazi, G. N
(2006). Bioreactor studies on the endophytic fungus Entrophosporain frequens for
the production of an anticancer alkaloid camptothecin. Canadian journal of
microbiology. Vol. 52(3). Pp. 189-196.
3. Arnold, A. E and Lutzoni, F (2007). Diversity and host range of foliar fungal
endophytes: are tropical leaves biodiversity hotspots?.Ecology. Vol. 88(3). Pp.
541-549.
4. Arnold, A.E and L.C. Lewis (2005). Evolution of fungal endophytes, and their
roles against insects. Ecological and Evolutionary Advances in Insect-Fungus
Associations (F. Vega and M. Blackwell, eds.). Oxford University Press, Oxford.
Pp. 74-96.
5. Bacon, C.W and White, J.F (2000). Microbial Endophytes. Marcel Dekker, New
York, Pp. 341-388.
6. Bayman, P. Lebron, L. L. Tremblay, R. L and Lodge, D (1997). Variation in
endophytic fungi from roots and leaves of Lepanthes (Orchidaceae).
NewPhytologist. Vol. 135(1). Pp 143-149.
7. Berdy, J. (2005). Bioactive microbial metabolites. Journal of Antibiotics, 58(1), 1.
8. Bladt, T. T. Frisvad, J.C. Knudsen, P. B. Larsen, T. O (2013). Anticancer and
antifungal compounds from Aspergillus, Penicillium and other filamentous
fungi. Molecules. Vol. 18(9). Pp. 11338-11376.
9. Bradford, M. M (1976). A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding.
Analytical biochemistry, Vol. 72(1-2), Pp. 248-254.
10. Caldwell, B.A. Jumpponen, A. Trappe, J.M (2000). Utilization of major detrital
substrates by dark septate, root endophytes. Mycologia. Vol. 92. Pp. 230–232.

References
83
11. Castro, R. A. Quecine, M. C., Lacava, P. T., Batista, B. D., Luvizotto, D. M.,
Marcon, J and Azevedo, J. L (2014). Isolation and enzyme bioprospection of
endophytic bacteria associated with plants of Brazilian mangrove ecosystem.
SpringerPlus, Vol. 3(1), 121-131.
12. Chen, H. Yang, C. Ke, T. Zhou, M. Li, Z. Zhang, M. and Hou, T (2015).
Antimicrobial activity of secondary metabolites from Streptomyces sp. K15, an
endophyte in Houttuynia cordata Thunb. Natural product research. Vol. 29(23).
Pp. 2223-2225.
13. Chen, L. Zhang, Q. Y. Jia, M. Ming, Q. L. Yue, W. Rahman, K and Han, T
(2014). Endophytic fungi with antitumor activities: Their occurrence and
anticancer compounds. Critical reviews in microbiology. Pp. 11-20.
14. Chevrier, M R. Ryan, AE. Lee, DY. Zhongze, M. Wu-Yan (2005). Boswellia
carterii extract inhibits TH1 cytokines and promotes TH2 cytokines in vitro.
ClinDiagn Lab Immunol. Vol. 12. Pp.575-580.
15. Chimata, N. K. Sasidhar, P. Challa, S (2013). Production of extracellular amylase
from agricultural residues by a newly isolated Aspergillus species in solid state
fermentation. African Journal of Biotechnology. Vol. 9(32). Pp. 5162-5169.
16. Chrost, R. J. and Krambeck, H. J (1986). Fluorescence correction for
measurements of enzyme activity in natural waters using methylumbelliferyl-
substrates. ArchivfürHydrobiologie, Vol. 106(1), Pp. 79-90.
17. Clay, K. and Schardl, C (2002). Evolutionary origins and ecological consequences
of endophyte symbiosis with grasses. The American Naturalist. Vol. 160(S4). Pp.
S99-S127.
18. Clay, K. Hardy, T. N and Hammond Jr, A. M (1985). Fungal endophytes of
Cyperus and their effect on an insect herbivore. American Journal of Botany. Pp.
1284-1289.
19. Cohen, S. D. (2006). Host selectivity and genetic variation of Discula umbrinella
isolates from two oak species: analyses of intergenic spacer region sequences of
ribosomal DNA. Microbial ecology, 52 (3), 463-469.

References
84
20. De Battista, J. P. Bouton, J. H. Bacon, C. W and Siegel, M. R (1990). Rhizome
and herbage production of endophyte-removed tall fescue clones and populations.
AgronomyJournal. Vol.82 (4). Pp. 651-654.
21. De Cesare, D. and Sassone-Corsi, P (2000). Transcriptional regulation by cyclic
AMP-responsive factors. Progress in nucleic acid research and molecular biology,
Vol.64, Pp. 343-369.
22. Deckert, RJ. Melville, LH. Peterson, RL (2001). Structural features of a
Lophodermiumendophyte during the cryptic life-cycle phase in the foliage of
Pinus strobus. Mycol Res. Vol. 105. Pp. 991–997.
23. Diblasi, L. Arrighi, F. Silva, J. Bardón, A. Cartagena, E (2015). Penicillium
commune metabolic profile as a promising source of antipathogenic natural
products. Natural product research. Pp.1-7.
24. Eaton, C.J. Cox, M.P. Scott, B (2011). What triggers grass endophytes to switch
from mutualism to pathogenism? Plant Sci. Vol. 180. Pp. 190–195.
25. El-Nagerabi SA, Elshafie AE, Alkhanjari SS (2014). Endophytic fungi associated
with endogenous Boswellia sacra. Biodiversitas. J Biol Div. Vol. 15(1). Pp. 24-30.
26. El-Nagerabi, S. A. Elshafie, A. E and Alkhanjari, S (2014). Endophytic fungi
associated with endogenous Boswellia sacra. Biodiversitas. Pp. 24-30.
27. Faeth, S.H (2002). Are endophytic fungi defensive plant mutualists? Oikos 98. Pp.
25–36.
28. Faeth, S.H. Gardner, D.R. Hayes, C.J. Jani, A. Wittlinger, S.K and Jones, T.A
(2006). Temporal and spatial variation in alkaloid levels in Achnatherumrobustum,
a native grass infected with the endophyte Neotyphodium. J. Chem. Ecol.Vol.32.
Pp. 307–324.
29. Fan, Y. Shi, L. M. Kohn, K. W. Pommier, Y and Weinstein, J (2001).
Quantitative structure-antitumor activity relationships of camptothecin analogues:
cluster analysis and genetic algorithm-based studies. Journal of medicinal
chemistry. Vol. 44(20). Pp. 3254-3263.
30. Felsenstein, J (1985). Confidence limits on phylogenies: an approach using the
bootstrap. Evolution. Pp. 783-791.

References
85
31. Fu SF, Wei JY, Chen HW, Liu YY, Lu HY, Chou JY (2015). Indole-3-acetic
acid: A widespread physiological code in interactions of fungi with other
organisms. Plant Sig Beh. Vol. 10(8). Pp.1048-1056.
32. Fujii, N. Yamashita, Y. Ando, K. Agatsuma, T. Saitoh, Y. Gomi, K. Nishiie, Y.
Nakano, H (1994). UCE1022, a new antitumor antibiotic withtopoisom erase I
mediated DNA cleavage activity, from Paecilomyces. J. Antibiot. (Tokyo). Vol.
47. Pp. 949-951.
33. Gallo, M.B.C, Guimaraes, D.O. Momesso, Saikai, R. Bezbaruah, R.L. Bora, T. C
and Pupo, M.T (2008). Natural Products from Endophytic Fungi. Microbial
Biotechnology. New India Publishing Agency, New Delhi, India, pp. 139-168.
34. Gallo. Guimarães, D.O. Momesso, L.S and Pupo, M.T (2008). Natural Products
from Endophytic Fungi. In: Saikai, R., Bezbaruah, R.L., Bora, T. C. (Eds.).
Microbial Biotechnology. New India Publishing Agency, New Delhi, India. Pp.
139-168.
35. Gawas-Sakhalkar P, Singh SM, Simantini N, Ravindra R (2012). High-
temperature optima phosphatases from the cold-tolerant Arctic fungus
Penicilliumcitrinum. Polar Res. Vol.31.
36. Giovannoni, S. J. E. F. Delong, G. J. Olsen, and N. R. Pace(1988). Phylogenetic
group-specific oligodeoxynucleotide probes for identification of single microbial
cells. J. Bacteriol. Vol. 170, Pp. 3584-3592.
37. Giovannoni, S. J. T. B. Britschgi, C. L. Moyer and K. G. Field (1990). Genetic
diversity in Sargasso Sea bacterioplankton. Nature (London), Vol. 345, Pp. 60-63.
38. Glick, B. R (2014). Bacteria with ACC deaminase can promote plant growth and
help to feed the world. Microbiological Research, Vol. 169(1), Pp. 30-39.
39. Guerin, P (1898). Sur la presence d‟un champignon dans l‟ivraie. J. Botanique.
Vol. 12, Pp. 230–238.
40. Gunatilaka AAL (2006). Natural products from plant-associated microorganisms:
distribution, structural diversity, bioactivity, and implications of their occurrence.
Journal of Natural Products. Vol. 69. Pp. 505-526.

References
86
41. Guo, B. Wang, Y. Sun, X and Tang, K (2008). Bioactive natural products from
endophytes: a review. Applied Biochemistry and Microbiology. Vol. 44(2). Pp.
136-142.
42. Hamayun, M. Khan, S. A. Ahmad, N. Tang, D. S. Kang, S. M and Kim, J. G
(2009). Cladosporium sphaerospermum as a new plant growth-promoting
endophyte from the roots of Glycine max (L.) Merr. World Journal of
Microbiology and Biotechnology. Vol. 25(4). Pp. 627-632.
43. Hamilton, C.E. Dowling, T.E and Faeth, S.H (2010). Hybridization in Endophyte
Symbionts Alters Host Response to Moisture and Nutrient Treatments. Microb.
Ecol. Vol. 59. Pp. 768–775.
44. Hardoim PR, van Overbeek LS, Elsas JD (2008). Properties of bacterial
endophytes and their proposed role in plant growth. Trends Microbiol. Vol. 16.
Pp. 463–71.
45. Harman, G.E (2011). Multifunctional fungal plant symbionts: new tools to
enhance plant growth and productivity. New Phytol. Vol. 189. Pp. 647– 649.
46. Hawksworth, D. L. (2001). The magnitude of fungal diversity: the 1.5 million
species estimate revisited. Asia-Pacific Mycological Congress on Biodiversity and
Biotechnology, and held at the University of Hong Kong on 9-13 July 2000.
Mycological research, 105(12), 1422-1432.
47. Heijden, M. G. Martin, F. M. Selosse, M. A and Sanders, I. R (2015). Mycorrhizal
ecology and evolution: the past, the present, and the future. New Phytologist.
Vol. 205(4). Pp. 1406-1423.
48. Hibbett, D.S. Ohman, A. Glotzer, D. Nuhn, M. Kirk, P and Nilsson, R.H (2011).
Progress in molecular and morphological taxon discovery in Fungi and options for
formal classification of environmental sequences. Fungal Biol. Rev. Vol. 25. Pp.
38-47.
49. Hoffman, M and Arnold, A.E (2008). Geography and host identity interact to
shape communities of endophytic fungi in cupressaceous trees. Mycol. Res. Vol.
112. Pp. 331-344.

References
87
50. Holguin, G. and Glick, B. R (2001). Expression of the ACC Deaminase Gene
fromEnterobacter cloacae UW4 in Azospirillum brasilense. Microbial Ecology,
Vol. 41(3), Pp. 281-288.
51. Honma, M and Shimomura, T (1978). Metabolism of 1-aminocyclopropane-1-
carboxylic acid. Agricultural and Biological Chemistry, Vol. 42(10), Pp. 1825-
1831.
52. Honma, M., & Shimomura, T. (1978). Metabolism of 1-aminocyclopropane-1-
carboxylic acid. Agricultural and Biological Chemistry, 42(10), 1825-1831.
53. Houbraken, J. Frisvad, J. C and Samson, R. A (2011). Fleming's penicillin
producing strain is not Penicillium chrysogenum but P. rubens. IMA fungus.
Vol.2 (1). Pp. 87-95.
54. Hsu WK, Hsu TH, Lin FY, Cheng YK and Yang JPW (2013). Separation,
purification, and α-glucosidase inhibition of polysaccharides from Coriolus
versicolor LH1 mycelia. Carbohydrate Polymers. Vol. 92(1). Pp. 297-306.
55. Hu ZY, Li YY, Huang YJ, Su WJ, Shen YM (2008). Three new sesquiterpenoids
from Xylaria sp. NCY2. HelvChimActa. Vol. 91. Pp. 46-52.
56. Hu, Z.Y. Li, Y. Huang, Y. J. Su, W.J. Shen, Y.M (2008). Three new
sesquiterpenoids from Xylaria sp. NCY2. Helv. Chim. Acta. Vol. 91. Pp. 46-52.
57. Idris, E. E. Iglesias, D. J. Talon, M and Borriss, R (2007). Tryptophan-dependent
production of indole-3-acetic acid (IAA) affects level of plant growth promotion
by Bacillus amyloliquefaciens FZB42. Molecular Plant-Microbe Interactions,
Vol.20 (6), Pp. 619-626.
58. Ishitani, K. Suzuki, S. Suzuki, M (1988). Antitumor activity of polygalactosamine
isolated from Paecilomyces sp. I-1 strain. J. Pharmacobiodyn. Vol. 11. Pp. 58-65.
59. Jain, P and Pundir, R. K (2015). Diverse Endophytic Microflora of Medicinal
Plants. In Plant-Growth-Promoting Rhizobacteria (PGPR) and Medicinal Plants.
Springer International Publishing. Pp. 341-357.

References
88
60. Jia YJ, Kakuta Y, Sugawara M, Igarashi T, Oki N, Kisaki M, Shoji T, Kanetuna
Y, Horita T, Matsui H, Onma M (1999). Synthesis and degradation of 1-
aminocyclopropane-1-carboxylic acid by Penicillium citrinum.Biosci Biotechnol
Biochem. Vol. 63, Pp. 542–549.
61. Kandeler, E. Tscherko, D. and Spiegel, H (1999). Long-term monitoring of
microbial biomass, N mineralisation and enzyme activities of a Chernozem under
different tillage management. Biologyandfertilityof soils, Vol. 28(4), Pp. 343-351.
62. Kato N, Suyama S, Shirokane M, Kato M, Kobayashi T, Tsukagoshi N (2002).
Novel α-glucosidase from Aspergillus nidulans with strong transglycosylation
activity. Applied Env Microbiol. Vol.68 (3). Pp. 1250-1256.
63. Kawaide. H (2006). Biochemical and molecular analysis of gibberellin
biosynthesis in fungi. Biosci Biotechnol Biochem. Vol. 70. Pp. 583-590.
64. Khan AL, Hussain J, Al-Harrasi A, Al-Rawahi A, Lee IJ (2013). Endophytic
fungi: a source of gibberellins and crop resistance to abiotic stress. Critical Rev
Biotech; Vol. 35(1), Pp. 62-74.
65. Khan AL, Hussain J, Al-Harrasi A, Al-Rawahi A, Lee IJ (2015). Endophytic
fungi: a source of gibberellins and crop resistance to abiotic stress. Crit Rev
Biotech. 1-13.
66. Khan, A. L. Al-Harrasi, A. Al-Rawahi, A. Al-Farsi, Z. Al-Mamari, A. Waqas, M
and Lee, I. J (2016). Endophytic Fungi from Frankincense Tree Improves Host
Growth and Produces Extracellular Enzymes and Indole Acetic Acid. PloS one,
Vol. 11(6), 0158207.
67. Khan, A. L. Hamayun, M. Kim, Y. H. Kang, S. M and Lee, I. J (2011).
Ameliorative symbiosis of endophyte (Penicillium funiculosum LHL06) under
salt stress elevated plant growth of Glycine max L. Plant Physiology and
Biochemistry. Vol.49 (8). Pp. 852-861.
68. Khan, A. L. Waqas, M. Kang, S. M. Al-Harrasi, A. Hussain, J. Al-Rawahi, A.
Lee, I. J (2014). Bacterial endophyte Sphingomonas sp. LK11 produces
gibberellins and IAA and promotes tomato plant growth. Journal of Microbiology,
Vol. 52(8), Pp. 689-695.

References
89
69. Kharwar, R. N. Strobel, G. A and Dubey, N. K (2011). Fungal endophytes: an
alternative source of bioactive compounds for plant protection. Natural products
in plant management.CAB International, Oxfordshire. Pp. 218-241.
70. Krajewska B (2009). Ureases: Functional, catalytic and kinetic properties: A
review. J MolCatal. Vol. 59. Pp. 9–21.
71. Krings, M. Taylor, T. N. Hass, H. Kerp, H. Dotzler, N and Hermsen, E. J (2007).
Fungal endophytes in a 400 million‑yr‑old land plant: infection pathways,
spatial distribution and host responses. New Phytol. Vol. 174. Pp. 648‑57.
72. Kumaran, R. S. Choi, Y. K. Lee, S. Jeon, H. J. Jung, H and Kim, H. J (2014).
Isolation of taxol, an anticancer drug produced by the endophytic fungus,
Phomabetae. African Journal of Biotechnology. Vol. 11(4). Pp. 950-960.
73. Kusari, P., Spiteller, M., Kayser, O and Kusari, S (2014). Recent advances in
research on Cannabis sativa L. endophytes and their prospect for the
pharmaceutical industry. Microbial Diversity and Biotechnology in Food Security.
Springer India. Pp. 3-15.
74. Kusari, S. and Spiteller, M (2012b). Metabolomics of endophytic fungi producing
associated plant secondary metabolites: progress, challenges and opportunities. In
Metabolomics. Pp. 241–266.
75. Kusari, S. Hertweck, C and Spiteller, M (2012). Chemical ecology of Endophytic
Fungi: origins of Secondary metabolites. Chemistry Biology. Vol. 19. Pp. 792-798.
76. Kusari, S. Pandey, S. P and Spiteller, M (2013). Untapped mutualistic paradigms
linking host plant and endophytic fungal production of similar bioactive
secondary metabolites. Phytochemistry, Vol. 91, Pp. 81-87.
77. Kyong, S.N. Young, S.J. Yong, H.K. Jin, W.H. Ha, W.K (2001). Cytotoxic
activities of acetoxyscirpenediol and rgosterol peroxide from Paecilomyces
tenuipes. Life Sci. Vol. 69. Pp. 229-237.
78. Latif Khan, A., Ahmed Halo, B., Elyassi, A., Ali, S., Al-Hosni, K., Hussain, J., &
Lee, I. J. (2016). Indole acetic acid and ACC deaminase from endophytic bacteria
improves the growth of Solarium lycopersicum. Electronic Journal of
Biotechnology, 19(3), 58-64.

References
90
79. Lerman, L. S. S. G. Fischer, I. Hurley, K. Silverstein, and N. Lumelsky (1984).
Sequence-determined DNA separations. Annu. Rev. Biophys. Bioeng. Vol. 13,
Pp. 399-423.
80. Liu C, Liu T, Yuan F, and Gu Y (2010). Isolating endophytic fungi from
evergreen plants and determining their antifungal activities. Africa J
Microbiol.Vol. 4. Pp. 2243-2248.
81. Liu, Y. J. Zhai, C. Y. Liu, Y and Zhang, K (2009). Nematicidal activity of
Paecilomyces spp. and isolation of a novel active compound. The Journal of
Microbiology. Vol. 47(3). Pp. 248-252.
82. Long, B. H and Balasubramanian, B (2000). Non-camptothecin topoisomerase I
active compounds as potential anticancer agents. Expert Opinion on Therapeutic
Patents. 10(5). Pp. 635-666.
83. Lopez-Diaz, T. M and Flannigan, B (1997). Production of patulin and
cytochalasin E by Aspergillus clavatus during malting of barley and wheat.
International journal of food microbiology. Vol. 35(2). Pp. 129-136.
84. Lucero, ME. Cooke, P. Dowd, S. Sun, SL (2011). Endophyte microbiome
diversity in micropropagated Atriplex canescensand Atriplex torreyi,griffithsii.
PLoS ONE. Vol. 6. Pp. 176-93.
85. Maria, G. L., Sridhar, K. R and Raviraja, N. S (2005). Antimicrobial and enzyme
activity of mangrove endophytic fungi of southwest coast of India. J Agric
Technol. Vol. 1(1). Pp. 67-80.
86. Maria, G.L. Sridhar. K.R. Raviraj, N.S (2005). Antimicrobial and enzyme activity
of mangrove endophytic fungi of southwest coast of India. J. Agric. Technol. Vol.
1. Pp. 67–80.
87. Marlida, Y. Saari, N. Hassan, Z. Radu, S (2000). Raw Starch degrading enzyme
from isolated strains of endophytic fungi. World J. Microbiol. Vol. 16. Pp. 573–
578.
88. Marwah RG, Fatope MO, Al Mahrooqi R, Varma GB, Al Abadi H, Al-Burtamani
SKS. (2007). Antioxidant capacity of some edible and wound healing plants in
Oman. Food Chemistry. Vol. 101. Pp. 465-470.

References
91
89. Marx, M. C. Wood, M. and Jarvis, S. C (2001). A microplate fluorimetric assay
for the study of enzyme diversity in soils. Soil Biology and Biochemistry, Vol.
33(12), Pp. 1633-1640.
90. Mathan, S., Subramanian, V., & Nagamony, S. (2013). Optimization and
antimicrobial metabolite production from endophytic fungi Aspergillus terreus
KC 582297. Eur J Exp Biol, 3, 138-44.
91. Mei C, Flinn BS (2010). The use of beneficial microbial endophytes for plant
biomass and stress tolerance improvement.Recent Pat Biotechnol. Vol.4. Pp. 81–
95.
92. Mercado-Blanco, J (2015). Life of Microbes inside the Plant. In Principles of
Plant-Microbe Interactions. Springer International Publishing. Pp. 25-32.
93. Mercado-Blanco, J (2015). Life of microbes inside the plant. In Principles of
Plant-Microbe Interactions. Springer International Publishing. Pp. 25-32.
94. Morath, S. U. Hung, R and Bennett, J. W (2012). Fungal volatile organic
compounds: a review with emphasis on their biotechnological potential. Fungal
Biology Reviews. Vol. 26(2). Pp 73-83.
95. Mosmann, T. (1983). Rapid colorimetric assay for cellular growth and survival:
application to proliferation and cytotoxicity assays. Journal of immunological
methods, 65(1-2), 55-63.
96. Nam, K. S. Jo, Y. S. Kim, Y. H. Hyun, J. W and Kim, H. W (2001). Cytotoxic
activities of acetoxyscirpenediol and ergosterol peroxide from Paecilomyces
tenuipes. Life sciences. Vol. 69(2). Pp. 229-237.
97. Ogas. J (2000). Gibberellins. Current Biology. Vol. 10. Pp. 48-58.
98. Onofre SB, Santos ZM, Kagimura FY, Mattiello SP (2015). Cellulases produced
by the endophytic fungus Pycnoporussanguineus (L.)Murrill. Afr J Agr Res. Vol.
10(13). Pp. 1557-1564.
99. Owen. NL, Hundley N (2004). Endophytes-the chemical synthesizers inside
plants. Science Progress. Vol. 87. Pp.79–99.

References
92
100. Pandey, A. Nigam, P. Soccol, C.R. Soccol, V.T. Singh, D. Mohan, R (2000).
Advances in microbial amylases. Biotechnol. Appl. Biochem. Vol. 31. Pp. 135–
152.
101. Pandi, M. Kumaran, R. S. Choi, Y. K. Kim, H. J and Muthumary, J (2013).
Isolation and detection of taxol, an anticancer drug produced from
Lasiodiplodiatheobromae, an endophytic fungus of the medicinal plant
Morindacitrifolia. African Journal of Biotechnology. Vol. 10(8). Pp. 1428-1435.
102. Patil, M. G. Pagare, J., Patil, S. N and Sidhu, A. K (2015). Extracellular
Enzymatic Activities of Endophytic Fungi Isolated from Various Medicinal
Plants. Int. J. Curr. Microbiol. App. Sci, Vol. 4(3). Pp. 1035-1042.
103. Petrini, O. Stone J, Carroll FE (1982). Endophytic fungi in evergreen shrubs in
western Oregon. A preliminary study. Can J Bot. Vol. 60, Pp. 789–796.
104. Prabavathy D, Valli Nachiyar C(2012). Screening for Extracellular Enzymes
and Production of Cellulase by an Endophytic Aspergillus sp, Using
Cauliflower Stalk as substrate. Int J App Bioeng. Vol. 6(2), Pp. 454-485.
105. Qawasmeh A, Obied HK, Raman A, Wheatley W (2012). Influence of fungal
endophyte infection on phenolic content and antioxidant activity in grasses:
interaction between Loliumperenne and different strains of
Neotyphodiumlolii. J AgriculFoodChem. Vol. 60(13). Pp. 3381-3388.
106. Qureshi AS, Marium S, Khushk I, Bhutto MA (2013). Production of alkaline
phosphatase from newly isolated Aspergillusfumigatus EFRL05. Top Class J
Microbiol. Vol.1. Pp. 67-73.
107. Radics, L. Kajtar-Peredy, M. Casinovi, C.G. Rossi, C. Ricci, M. Tuttobello, L
(1987). Leucinostatins H and K, two novel peptide antibiotics withtertiary
amine-oxide terminal group from Paecilomyces marquandii isolation, structure
and biological activity. J. Antibiot. (Tokyo). Vol. 40. Pp. 714-716.
108. Redman, R.S. Sheehan, K.B. Stout, R.G. Rodriguez, R.J and Henson, J.M
(2002). Thermo-tolerance conferred to plant host and fungal endophyte during
mutualistic symbiosis. Science Pp. 290-298.

References
93
109. Reisner, A. H., Nemes, P and Bucholtz, C (1975). The use of Coomassie
Brilliant Blue G250 perchloric acid solution for staining in electrophoresis and
isoelectric focusing on polyacrylamide gels. AnalyticalBiochemistry, Vol.64
(2), Pp. 509-516.
110. Rodrigues, KF. Samuels, GJ (1990). Preliminary study of endophytic fungi in a
tropical palm. Mycol Res. Vol. 94. Pp. 827–830.
111. Rodriguez, R and Redman, R (2008a). More than 400 million years of evolution
and some plants still can‟t make it on their own: plant stress tolerance via fungal
symbiosis. J. Exp. Bot. Vol. 59.
112. Rodriguez, R. J, White, Jr, J. F, Arnold, A. E and Redman, R. S (2009). Fungal
endophytes: diversity and functional roles. New Phytologist. Vol. 182(2). Pp.
314-330.
113. Rodriguez, R.J. White, J.F. Jr, Arnold, A.E and Redman, R.S (2009). Fungal
endophytes: diversity and functional roles. New Phytologist. Vol. 182, Pp. 314-30.
114. Rosenblueth, M. and Martinez-Romero, E (2006). Bacterial endophytes and their
interactions with hosts. Acta Pharmacologica Sinica, Vol. 19 (8). Pp. 827–837.
115. Rudgers, J. A and Clay, K (2007). Endophyte symbiosis with tall fescue: how
strong are the impacts on communities and ecosystems? Fungal Biology
Reviews. Vol. 21(2). Pp. 107-124.
116. Rytioja J, Hildén K, Mäkinen S, Vehmaanperä J, Hatakka A, Mäkelä MR (2015).
Saccharification of Lignocelluloses by Carbohydrate Active Enzymes of the
White Rot Fungus Dichomitus squalens. PloS one. Vol. 10(12). Pp. 145-166.
117. Sachan, A., Ghosh, S., Mitra, A (2006). Biotransformation of p‐coumaric acid by
Paecilomyces variotii. Letters in applied microbiology. Vol. 42(1). Pp. 35-41.
118. Saikkonen, K. Wali, P. Helander, M and Faeth, S.H (2004). Evolution of
endophyte–plant symbioses. Trends Plant Sci. Vol. 9, Pp. 1360-1385.
119. Saikkonen. K, Saari. S, Helander. M (2010). Defensive mutualism between
plants and endophytic fungi? Fungal Divers. Vol. 41. Pp. 101–113.

References
94
120. Saitou, N and Nei, M (1987). The neighbor-joining method: a new method for
reconstructing phylogenetic trees. Molecular biology and evolution. Vol. 4(4).
Pp. 406-425.
121. Samson, R. A. Hoekstra, E. S and Frisvad, J. C (2004). Introduction to food-and
airborne fungi. Centraalbureau voor Schimmelcultures (CBS).
122. Sapp, J (2004). The dynamics of symbiosis: an historical overview. Can. J. Bot.
Vol. 82, Pp. 1046/56.
123. Saxena S. Microbial metabolites for development of ecofriendly agrochemical.
Allelopath J (2014). Vol. 33. Pp.1–24.
124. Schardl, C. L., Leuchtmann, A and Spiering, M. J (2004). Symbioses of grasses
with seedborne fungal endophytes. Annu. Rev. Plant Biol. Vol.55. Pp. 315-340.
125. Schulz, B and Boyle, C (2005). The endophytic continuum. Mycolog. Res. Vol.
109. Pp. 661–686.
126. Schulz, B. Boyle, C. Draeger, S. Römmert, A. K and Krohn, K (2002).
Endophytic fungi: a source of novel biologically active secondary metabolites.
Mycological Research. 106 (09). Pp. 996-1004.
127. Schulz, B. Mmert, R. Dammann, AK. Aust, U. Strack, D (1999). The
endophyte-host nteraction: a balanced antagonism. Mycol. Res. Vol. 103. Pp.
1275–1283.
128. Selim, K. A, El-Beih, A. A, Abdel-Rahman, T. M and El-Diwany (2012).
Biology of endophytic fungi. Vol. 2(1), Pp. 31-82.
129. Selosse, M.A. Baudoin, E. Vandenkoornhuyse, P (2004). Symbiotic
microorganisms, a key for ecological success and protection of plants. Comptes
Rendus Biologies Vol. 327. Pp 639–648.
130. Shaharoona, B. Arshad, M. Zahir, Z. A and Khalid, A (2006). Performance of
Pseudomonas spp. containing ACC-deaminase for improving growth and yield
of maize (Zea mays L.) in the presence of nitrogenous fertilizer. Soil Biology
and Biochemistry, Vol. 38(9), Pp. 2971-2975.

References
95
131. Shweta, S. Gurumurthy, B. R. Ravikanth, G. Ramanan, U. S and Shivanna, M.
B (2013). Endophytic fungi from Miquelia dentata. Produce the anti-cancer
alkaloid, camptothecine. Phytomedicine. Vol. 20(3). Pp. 337-342.
132. Silvia F, Sturdikova M, Muckova M (2007). Bioactive secondary metabolites
produced by microorganisms associated with plants. Biologia. Vol. 62. Pp. 251-
257.
133. Singh B, Kaur T, Kaur S, Manhas RK, Kaur A(2015). An alpha-glucosidase
inhibitor from an endophytic Cladosporium sp. with potential as a biocontrol
agent. Appl Biochem Biotechnol. Vol. 51. Pp. 29-37.
134. Sirikantaramas, S. Asano, T. Sudo. H, Yamazaki. M, Saito. K(2007).
Camptothecin: therapeutic potential and biotechnology. Current Pharmaceutical
Biotechnology. Vol. 8, Pp. 196-202.
135. Smith, S. E and Read, D. J (1996). Mycorrhizal symbiosis. Academic press.
136. Stemmer, M. Gerzabek, M. H and Kandeler, E (1998). Invertase and xylanase
activity of bulk soil and particle-size fractions during maize straw
decomposition. Soil Biology and Biochemistry.Vol. 31(1). Pp. 9-18.
137. Stierle A, Strobel G, Stierle D (1993). Taxol and taxane production by
Taxomycesandreanae, an endophytic fungus of Pacific yew. Science. Vol. 260.
Pp. 214-216.
138. Strobel G, Daisy B, Castillo U, Harper J (2004). Natural products from
endophytic microorganisms. J Nat Prod Vol. 67. Pp. 257–68.
139. Strobel, G. A (2003). Endophytes as sources of bioactive products. Microbes
and infection. Vol. 5(6). Pp. 535-544.
140. Strobel, G. Daisy, B (2003). Bioprospecting for microbial endophytes and their
natural products. Microbiol Mole Biol Rev. Vol. 67. Pp. 491–502.
141. Subramanian, R. Asmawi, M. Z, Sadikun, A (2008). In vitro alpha-glucosidase
and alpha-amylase enzyme inhibitory effects of Andrographispaniculata extract
and andrographolide. ActaBiochim Pol. Vol. 55(2). Pp. 391-398.
142. Sun, X and Guo, L. D (2012). Endophytic fungal diversity: review of traditional
and molecular techniques. Mycology. Vol. 3(1). Pp. 65-76.

References
96
143. Sun, X. Guo, L. D and Hyde, K. D (2011). Community composition of
endophytic fungi in Acer truncatum and their role in decomposition.
FungalDiversity. Vol.47 (1). Pp. 85-95.
144. Sunitha, V. H. Devi, D. N and Srinivas, C (2013). Extracellular enzymatic
activity of endophytic fungal strains isolated from medicinal plants. World
Journal of Agricultural Sciences. Vol. 9(1). Pp. 01-09.
145. Suryanarayanand, T. S., & Kumaresan, V. (2000). Endophytic fungi of some
halophytes from an estuarine mangrove forest. Mycological Research, 104(12),
1465-1467.
146. Tamura, K. Stecher, G. Peterson, D. Filipski, A and Kumar, S (2013). MEGA6:
molecular evolutionary genetics analysis version 6.0. Molecular biology and
evolution. Vol. 30(12). Pp. 2725-2729.
147. Tan RX, Zou WX (2001) Endophytes: a rich source of functional metabolites.
Nat Product Reports Vol. 18. Pp. 448-459.
148. Tejesvi, M. V. Kajula, M. Mattila, S and Pirttila, A. M (2011). Bioactivity and
genetic diversity of endophytic fungi in Rhododendron tomentosum Harmaja.
Fungal Diversity. Vol. 47(1). Pp. 97-107.
149. Tolulope, R. A., Adeyemi, A. I., Erute, M. A and Abiodun, T. S (2015).
Isolation and screening of endophytic fungi from three plants used in traditional
medicine in Nigeria for antimicrobial activity. International Journal of Green
Pharmacy. Vol. 9(1). Pp. 50-58.
150. Verma, V. C, Gond. S. K, Kumar. A, Kharwar. R. N and Strobel, G (2007). The
endophytic mycoflora of bark, leaf, and stem tissues of Azadirachtaindica A.
Juss (Neem) from Varanasi (India). Microbial Ecology. Vol. 54(1). Pp 119-125.
151. Vieira, PDS. de Souza Motta, CM. Lima, D. Torres, JB. Quecine, MC.
Azevedo, JL. de Oliveira (2011). Endophytic fungi associated with transgenic
and non-transgenic cotton. Mycology. Vol. 2. Pp. 91–97.
152. Waller, F. Achatz, B. Baltruschat, H. Fodor, J. Becker, K. Fischer, M. Heier, T.
Huckelhoven, R. Neumann, C. Von-Wettstein, D. Franken, P and Kogel, K.H
(2005). The endophytic fungus Piriformisindica reprograms barley to salt-stress
tolerance, disease resistance and higher yield. PNAS, Vol. 102. Pp. 13386-
13391.

References
97
153. Wang, J. Huang, Y. Fang, M. Zhang, Y. Zheng, Z. Zhao, Y. Su, W (2002).
Brefeldin A, a cytotoxin produced by Paecilomyces sp. and Aspergillus clavatus
isolated from Taxusmairei and Torreyagrandis. FEMS Immunology & Medical
Microbiology. Vol. 34(1). Pp. 51-57.
154. Waqas, M. Khan, A. L. Kamran, M. Hamayun, M. Kang, S. M. Kim, Y. H and
Lee, I. J (2012). Endophytic fungi produce gibberellins and indoleacetic acid
and promotes host-plant growth during stress. Molecules, Vol. 17(9), Pp.
10754-10773.
155. Ward, D. M. R. Weller and M. M. Bateson (1990). 16S rRNA sequences reveal
numerous uncultured microorganisms in a natural community. Nature (London)
Vol. 344, Pp. 63-65.
156. Wei H, Wang W, Yarbrough JM, Baker JO, Laurens L, Van Wychen S, Zhang
M (2013). Genomic, proteomic, and biochemical analyses of oleaginous
Mucorcircinelloides: Evaluating its capability in utilizing cellulolytic substrates
for lipid production. PlOS ONE. Vol. 8(9). Pp. 710718.
157. Wei, Y. K. Gao, Y. B. Zhang, X. Su, D. Wang, Y. H. Xu, H and Nie, L. Y
(2007). Distribution and diversity of Epichloe Neotyphodium fungal endophytes
from different populations of Achnatherum sibiricum (Poaceae) in the Inner
Mongolia Steppe, China. FungalDivers. Vol. 24. Pp. 329-345.
158. Wilson, D (2000). Ecology of woody plant endophytes. Microbial endophytes.
Pp. 389-420.
159. Woese, C. R(1987). Bacterial evolution. Microbiol. Rev. 51, Pp. 221- 271.
160. Wu, S. H. Huang, R. Miao, C. P and Chen, Y. W (2013). Two new steroids
from an endophytic fungus Phomopsis sp. Chemistry & biodiversity. Vol. 10(7).
Pp. 1276-1283.
161. Xiang Sun and Liang-Dong Guo (2012). Endophytic fungal diversity: review of
traditional and molecular techniques, Mycology. An International Journal on
Fungal Biology. Vol. 3:1. Pp. 65-76.
162. Xiao J, Zhang Q, Gao YQ, Tang JJ, Zhang AL, Gao JM (2014). Secondary
metabolites from the endophytic Botryosphaeriadothidea of Melia azedarach
and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J Agric
Food Chem. Vol. 62(16). Pp. 3584-3590.

References
98
163. Xu L, Zhou L, Zhao J, Jiang W (2008). Recent studies on the antimicrobial
compounds produced by plant endophytic fungi. Natural Product Research and
Development. Vol. 20. Pp. 731-740.
164. Yang, X. Guo, S. Zhang, L and Shao H (2003). Selection of producing
podophyllotoxin endophytic fungi from podophyllin plant. Natural Product
Research and Development. Vol. 15. Pp. 419-422.
165. Yang, Y. Yan, M and Hu, B (2014). Endophytic fungal strains of soybean for
lipid production. BioEnergy Research. Vol. 7(1). Pp. 353-361.
166. Yim W, Seshadri S, Kim K, Lee G, Sa T (2013). Ethylene emission and PR
protein synthesis in ACC deaminase producing Methylobacterium spp.
inoculated tomato plants, challenged with Ralstoniasolanacearum under
greenhouse conditions. Plant Physiol Bioch. Vol.67, Pp. 95-104.
167. Yim, W. Seshadri, S. Kim, K. Lee, G and Sa, T (2013). Ethylene emission and
PR protein synthesis in ACC deaminase producing Methylobacterium spp.
inoculated tomato plants (Lycopersicon esculentum Mill.) challenged with
Ralstonia solanacearum under greenhouse conditions. Plant Physiology and
Biochemistry, Vol. 67, Pp. 95-104.
168. Yin. H, Zhao. Q, Sun. FM (2009). Gentiopicrin-producing endophytic fungus
isolated from Gentianamacrophylla. Phytomedicine. Vol. 16. Pp. 793–797.
169. Ying, Y. M. Zheng, Z. Zhang, L.W. Shan, W.G. Wang, J.W and Zhan, Z.J
(2014). Rare C25Steroids Produced by Penicilliumchrysogenum P1X, a Fungal
Endophyte of Huperziaserrata. HCA. Vol. 97 Pp. 95–101.
170. Yong, X. An-guo, L. Wen-fang, W (2003). Identification of
Taxuscuspidatasieb. Et Zucc. Endophytic fungi-new species, species known and
their metabolite. Journal of Forestry Research. Vol. 14(4). Pp. 290-294.
171. Yoshii, Hand Imaseki, H (1981). Biosynthesis of auxin-induced ethylene.
Effects of indole-3-acetic acid, benzyladenine and abscisic acid on endogenous
levels of 1-aminocyclopropane-l-carboxylic acid (ACC) and ACC synthase.
Plant and Cell Physiology.Vol. 22(3), Pp. 369-379.

References
99
172. Zhang, J. Y. Tao, L. Y. Liang, Y. J. Chen, L. M. Mi, Y. J. Zheng, L. S and Fu,
L. W (2010). Anthracenedione derivatives as anticancer agents isolated from
secondary metabolites of the mangrove endophytic fungi. Marinedrugs. Vol.
8(4). Pp. 1469-1481.
173. Zhang, J.H. Liu, Y.P. Pan, Q.H. Zhan, J.C. Wang, X.Q and Huang, W.D (2006).
Changes in membrane-associated H+-ATPase activities and amounts in young
grape plants during the cross adaptation to temperature stresses. Plant Sci. Vol.
170. Pp. 768–777.
174. Zhang, P. Li, X. M. Wang, J. N. Li, X and Wang, B. G (2015). New butenolide
derivatives from the marine-derived fungus Paecilomyces variotii with DPPH
radical scavenging activity. Phytochemistry Letters. Vol. 11. Pp. 85-88.
175. Zhang, P. M ndi, A. Li, . M. Du, F. . Wang, J. N. Li, . Wang, B. G (2014).
Varioxepine A, a 3 H-Oxepine-Containing Alkaloid with a New Oxa-Cage from
the Marine Algal-Derived Endophytic Fungus Paecilomyces variotii. Organic
letters. Vol. 16(18). Pp. 4834-4837.
176. Zhao, J. Shan, T. Mou, Y and Zhou, L (2011). Plant-derived bioactive
compounds produced by endophytic fungi. Mini reviews in medicinal
chemistry. Vol. 11. Pp. 159-168.
177. Zhao, J. Shan, T. Mou, Y and Zhou, L (2011). Plant-derived bioactive
compounds produced by endophytic fungi. Mini reviews in medicinal
chemistry. Vol. 11(2). Pp. 159-168.
178. Zhou, D., & Hyde, K. D. (2001). Host-specificity, host-exclusivity, and host-
recurrence in saprobic fungi. Paper presented at the Asian Mycological
Congress 2000 (AMC 2000) incorporating the 2nd Asia-Pacific Mycological
Congress on Biodiversity and Biotechnology, and held at the University of
Hong Kong on 9-13 July 2000. Mycological Research, 105(12), 1449-1457.
179. Zhou, L. Wu, J (2006). Development and application of medicinal plant tissue
cultures for production of drugs and herbal medicinal in China. Natural Product
Reports. Vol. 23. Pp. 789-810.
180. Zhou, L. Zhao, J. Xu, L. Huang, Y. Ma, Z. Wang. J, Jiang, W (2009).
Antimicrobial compounds produced by plant endophytic fungi. In: De Costa P,
Bezerra P, eds. Fungicides: Chemistry, Environmental Impact and Health
Effects. New York: Nova Science Publishers. Pp. 91-119.

APPENDIXES

Appendixes
100
APPENDIX- IA

Appendixes
101
APPENDIX-IB

Appendixes
102
APPENDIX-IC

Appendixes
103
APPENDIX-ID

Appendixes
104
APPENDIX-IE

Appendixes
105
APPENDIX-IF

Appendixes
106
APPENDIX-IG

Appendixes
107
APPENDIX-IH

Appendixes
108
APPENDIX-IJ

Appendixes
109
APPENDIX-IK

Appendixes
110
APPENDIX-IIA

Appendixes
111
APPENDIX-IIB

Appendixes
112
APPENDIX-IIC

Appendixes
113
APPENDIX-IID

Appendixes
114
APPENDIX-IIE

Appendixes
115
APPENDIX-IIF

Appendixes
116
APPENDIX-IIG

Appendixes
117
APPENDIX-IIIA

Appendixes
118
APPENDIX-IIIB

Appendixes
119
APPENDIX-IIIC

Appendixes
120
APPENDIX-IIID

Appendixes
121
APPENDIX-IIIE

Appendixes
122
APPENDIX-IIIF

Appendixes
123
APPENDIX-IIIG

Appendixes
124
APPENDIX-IIIH

Appendixes
125
APPENDIX-IVA

Appendixes
126
APPENDIX-IVB

Appendixes
127
APPENDIX-IVC

Appendixes
128
APPENDIX-IVD

Appendixes
129
APPENDIX-IVE

Appendixes
130
APPENDIX-IVF

Appendixes
131
APPENDIX-VA

Appendixes
132
APPENDIX-VB

Appendixes
133

Appendixes
134

Appendixes
135

Appendixes
136
APPENDIX-VC

Appendixes
137
APPENDIX-VD

Appendixes
138
APPENDIX-VE

Appendixes
139
APPENDIX-VF

Appendixes
140
APPENDIX-VG
Copyright © 2022 FDOKUMEN




















