
Recent advances in Machining of Polymeric Composite Materials

ES
WOS7
h
••••
a
ARRAA
KSEC
1
mbtat
po[
(
0h
Journal of Hazardous Materials 261 (2013) 614– 620
Contents lists available at ScienceDirect
Journal of Hazardous Materials
jou rn al hom epage: www.elsev ier .com/ locate / jhazmat
xtracellular polymeric substances from copper-toleranceinorhizobium meliloti immobilize Cu2+
enjie Hou, Zhanqiang Ma, Liangliang Sun, Mengsha Han, Jianjun Lu, Zhenxiu Li,sama Abdalla Mohamad, Gehong Wei ∗
tate Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, College of Life Sciences, Northwest A&F University, Yangling, Shaanxi12100, China
i g h l i g h t s
EPS produced by Sinorhizobium meliloti CCNWSX0020 restricts uptake of Cu2+.We focused on the EPS, which is divided into three main parts.LB-EPS played a more important role than S-EPS and TB-EPS in Cu2+ immobilization.Proteins and carbohydrates were the main extracellular compounds which had functional groups such as carboxyl ( COOH), hydroxyl ( OH), and amide(N H), primarily involved in metal ion binding.
r t i c l e i n f o
rticle history:eceived 31 December 2012eceived in revised form 24 May 2013ccepted 18 June 2013vailable online 26 June 2013
eywords:
a b s t r a c t
The copper tolerance gene of wild-type heavy metal-tolerance Sinorhizobium meliloti CCNWSX0020 wasmutated by transposon Tn5-a. The mutant was sensitive up to 1.4 mM Cu2+. Production, components,surface morphology, and functional groups of extracellular polymeric substances (EPS) of the wild-typestrains were compared with sensitive mutant in immobilization of Cu2+. EPS produced by S. melilotiCCNWSX0020 restricts uptake of Cu2+. The cell wall EPS were categorized based on the compactness andfastness: soluble EPS (S-EPS), loosely bound EPS (LB-EPS), and tightly bound EPS (TB-EPS). LB-EPS played
2+
inorhizobium melilotixtracellular polymeric substances (EPS)opper tolerancea more important role than S-EPS and TB-EPS in Cu immobilization. Scanning electron microscopy(SEM) analysis LB-EPS had rough surface and many honeycomb pores, making them conducive to copperentry; therefore, they may play a role as a microbial protective barrier. Fourier transform-infrared (FT-IR)analysis further confirm that proteins and carbohydrates were the main extracellular compounds whichhad functional groups such as carboxyl ( COOH), hydroxyl ( OH), and amide (N H), primarily involvedin metal ion binding.
. Introduction
Wastewater from anthropogenic sources contains many heavyetal ions. Various chemical, physical, and biological methods have
een developed to remove heavy metals from wastewater. Duringhe last decade, research has focused on biological methods, whichre more economical and efficient. Biosorption is a cost-effectiveechnology for the treatment of high volumes of wastewater [1,2].
Certain types of microbial biomass secrete high molecular mass
olymers, which are released into the surrounding environmentr remain attached to cell surfaces (capsular polysaccharides)3]. A number of studies show that extracellular polymeric∗ Corresponding author. Tel.: +86 29 87091175; fax: +86 29 87091175.E-mail addresses: [email protected], [email protected]
G. Wei).
304-3894/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.jhazmat.2013.06.043
© 2013 Elsevier B.V. All rights reserved.
substances (EPS) are effective absorbents for removing heavymetals and organic pollutants such as dyes and pesticides [4–7].EPS are metabolic products that accumulate on the microbial cellsurface, causing the cells to aggregate and providing protectionby reducing chemical exposure, stabilizing the membrane againstthe harsh external environment, and serving as carbon and energyreserves during starvation [8]. This protective mechanism has beenobserved in comparisons of cells embedded in EPS and free cellswithout EPS. Cellular tolerance is partially due to binding and/orreaction of EPS components to/with heavy metal ions [9]. EPS areheterogeneous gel-like matrices of polymers composed of polysac-charides, proteins, nucleic acids, and lipids. Heavy metal ions areembedded through electrostatic and hydrophobic interactions. EPS
are classified as soluble (S-EPS) and bound (B-EPS). S-EPS are dis-solved in solution; B-EPS are bound to cells and are generallysubdivided into loosely bound EPS (LB-EPS) and tightly bound EPS(TB-EPS) [10,11]. LB-EPS may function as the primary surface for cell
dous M
aEbte
fiMkttosiesiEt
2
2
t2pmTBcrptW1
2
tpmw1(Ctwif
R
wt
2
v1rtw
W. Hou et al. / Journal of Hazar
ttachment and flocculation. However, most experimental work onPS has not specifically addressed the role of the three types EPS iniosorption in solution. Thus, little information is available to dis-inguish various compositions of the three types of EPS and theirffects on surface properties and biosorption.
Sinorhizobium meliloti CCNWSX0020 is a symbiotic nitrogen-xing bacteria tolerance to 1.4 mM copper; it was isolated fromedicago lupulina growing in mine tailings [12]. To the best of our
nowledge, the relative contributions of S-EPS, LB-EPS, and TB-EPSo copper immobilization have not been investigated. Therefore,he goal of this study was: (1) to investigate the main compositionsf three types of EPS from the wild-type strain and sensitive mutanttrain (2) and to identify the adsorption mechanisms, determine thendividual effects of the three types EPS on biosorption by scanninglectron microscopy (SEM) and Fourier transform-infrared (FT-IR)pectroscopy spectra analysis. We believe that a better understand-ng of the immobilization mechanisms of copper occurring withinPS and the factors that affect those mechanisms may contribute tohe development of more efficient biosorption treatment processes.
. Materials and methods
.1. Bacterial culture and mutation
S. meliloti CCNWSX0020 was cultivated in TY medium (5 g tryp-one, 3 g yeast extract, and 0.7 g CaCl2·2H2O per liter) for 72 h at8 ◦C with shaking at 150 rpm. Mutation was constructed usinglasmid pRL1063a, which harbors a Tn5 transposon with pro-oterless luxAB genes and a kanamycin resistance marker [13].
ri-parental mating was carried out as described by Chu [14].riefly, cells of the donor strain E. coli DH5a, harboring the sui-ide plasmid pRL1063a or the helper plasmid pRK2013, and theecipient strain, S. meliloti CCNWSX0020, were grown in LB sup-lemented with kanamycin and TY medium, respectively. Theransposon Tn5 was transferred into wild-type S. meliloti CCN-
SX0020, and the mutant strain was screened on plates containing.4 mM Cu2+.
.2. Copper accumulation
Cu2+ solution (1.4 mM) was prepared by dissolving CuCl2 in dis-illed water. The solution was sterilized by filtering with a 0.2 �more size filter [15]. Experiments were carried out using 100 mL TYedium in 150-mL Erlenmeyer flasks. The wild-type and mutantere cultured in Cu2+ medium at 28 ◦C for 72 h with shaking at
50 rpm. The cells were harvested from early stationary culturesOD600 = 1.2–1.5) by centrifugation at 5000 rpm for 10 min. Residualu2+ in the medium was measured by an atomic absorption spec-rophotometer – AAS every 6 h. A blank experiment was performedith Cu-free TY medium. Cu2+ tolerance was evaluated by compar-
son with the control. The removal rate of Cu2+ was evaluated asollows:
emoval rate (%) = (C0 − Ce)C0
× 100
here C0 was the initial metal ion concentration (mg L−1), Ce washe residual metal concentration in solution (mg L−1).
.3. Isolation, extraction, and characterization of EPS
After 3 days of pure cultivation in TY medium, cells were har-ested at room temperature by centrifugation at 5000 rpm for
0 min. The supernatant was dialyzed against Milli-Q water usingegenerated cellulose (RC) membranes (8000 MWCO from Spec-rum) to remove low-molecular-weight impurities. Milli-Q wateras changed every 3 h for 24 h dialysis [16]. S-EPS solution wasaterials 261 (2013) 614– 620 615
freeze-dried and reserved. To separate LB-EPS, cells were dissolvedin Milli-Q water and sonicated at 40 w for 1 min. The suspensionwas centrifuged at 7000 rpm for 20 min. Dialysis yielded the LB-EPSsolution, which was freeze-dried and reserved. Cells were dissolvedin Milli-Q water and heated at 80 ◦C for 10 min; the suspension wascentrifuged at 15,000 rpm for 20 min. Dialysis of the supernatantyielded TB-EPS solution [17].
The EPS carbohydrate content was measured by the phenol-sulfuric acid method with glucose standard [18]. Protein contentwas determined by the Bradford method with bovine albuminserum standard [19] and the nucleic acid content was determinedby the diphenylamine method [20].
Biosorption experiments were carried out in order to investi-gate Cu2+ removal by 10 mg EPS were placed in dialysis bags (8000MWCO) in the solution (at 28 ◦C, pH 7 and 1.4 M Cu2+).
2.4. Scanning electron microscopy (SEM)
The S-EPS, LB-EPS, TB-EPS of the wild strain and mutant sur-face structure and morphology were observed by SEM (JSM-6360LV; JEOL, Peabody, MA, USA). All EPS were dried in a critical-pointdryer using CO2 (K850; EMITECH, East Grinstead, UK). Metal-loadedsamples were mounted on a stainless steel slab with double-sidedadhesive tape and a thin layer of gold in a high vacuum. Specimenswere examined by SEM.
2.5. Fourier transform-infrared spectroscopy (FT-IR)
Infrared analysis was performed with a 330 FT-IR (Fouriertransform-infrared spectrometer, NICOLET AVATAR, USA NicoletCo.) at a range of 4000–400 cm−1. Powdered EPS (1 mg) was mixedand ground with 100 mg KBr (Spectrum pure) in an agate mortarto investigate the functional groups relevant to Cu2+ biosorption.
2.6. Experimental design and statistical analysis
All experiments were performed in triplicate and the mean val-ues are presented. Statistical analysis was performed with SPSS 13.0Bivariate Correlation Analysis. The Pearson rank order coefficientwas determined to compare Cu2+ tolerance between EPS types andthe removal of Cu2+ by the wild-type and mutant strains. Distinctdifferences were detected by Dunnett and Tukey grouping tests;p < 0.05 was considered statistically significant.
3. Results and discussion
3.1. Cu2+ accumulation
The wild-type strain grew well in medium supplemented with1.4 mM Cu2+, while the sensitive mutant strain grew extremelyslowly. The wild-type and mutant exhibited different Cu2+ removalabilities. At the end of the 3rd day, the sensitive mutant strain andwild-type adsorbed 9.8 and 20.6% of Cu2+ onto the cell surfaceand accumulated 6.1 and 3.9% of the Cu2+ into their intracellularspaces (Fig. 1). The wild-type absorbed more Cu2+ than the sen-sitive mutant strain. Bacteria, fungi, yeast, and algae are thoughtof as microbial biosorbents for removal of heavy metals. Previ-ous studies have shown that EPS has important roles in metalion biosorption by Shewanella species [21]. In order to cope withheavy metal exposure, strains have evolved several metal toler-ance strategies. Intracellular metal tolerance mechanisms includeefflux, complexation, or reduction of metal ions to a non-toxic form;
chelation of metal ions by certain extracellular polymeric sub-stances provide extracellular protection [22]. These observationsare consistent with the chemical nature of the released extracellu-lar compounds, which contain large amounts of siderophores and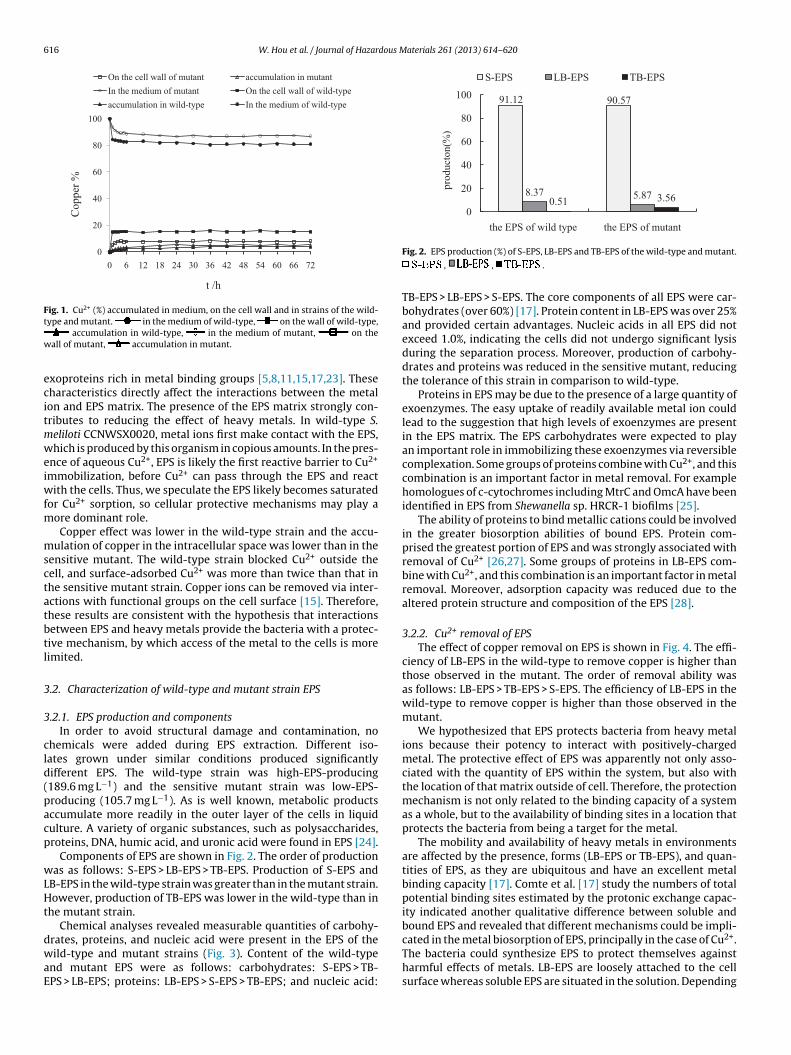
616 W. Hou et al. / Journal of Hazardous Materials 261 (2013) 614– 620
t /h
Co
pp
er %
0
20
40
60
80
100
0 6 12 18 24 30 36 42 48 54 60 66 72
On the cell wall of mutant accu mulation in mutant
In the medium of mutant On the cell wall of wi ld-ty pe
accu mulation in wild -ty pe In the medium of wild -ty pe
Fig. 1. Cu2+ (%) accumulated in medium, on the cell wall and in strains of the wild-t
w
ecitmweiwfm
msctatbtl
3
3
cld(pacp
wLHt
dwaE
91.12 90.57
8.37 5.870.51 3.56
0
20
40
60
80
100
the EPS of wild type the EPS of mutant
pro
duct
on(%
)
S-EPS LB-EPS TB-EPS
2+
ype and mutant. in the medium of wild-type, on the wall of wild-type,accumulation in wild-type, in the medium of mutant, on the
all of mutant, accumulation in mutant.
xoproteins rich in metal binding groups [5,8,11,15,17,23]. Theseharacteristics directly affect the interactions between the metalon and EPS matrix. The presence of the EPS matrix strongly con-ributes to reducing the effect of heavy metals. In wild-type S.eliloti CCNWSX0020, metal ions first make contact with the EPS,hich is produced by this organism in copious amounts. In the pres-
nce of aqueous Cu2+, EPS is likely the first reactive barrier to Cu2+
mmobilization, before Cu2+ can pass through the EPS and reactith the cells. Thus, we speculate the EPS likely becomes saturated
or Cu2+ sorption, so cellular protective mechanisms may play aore dominant role.Copper effect was lower in the wild-type strain and the accu-
ulation of copper in the intracellular space was lower than in theensitive mutant. The wild-type strain blocked Cu2+ outside theell, and surface-adsorbed Cu2+ was more than twice than that inhe sensitive mutant strain. Copper ions can be removed via inter-ctions with functional groups on the cell surface [15]. Therefore,hese results are consistent with the hypothesis that interactionsetween EPS and heavy metals provide the bacteria with a protec-ive mechanism, by which access of the metal to the cells is moreimited.
.2. Characterization of wild-type and mutant strain EPS
.2.1. EPS production and componentsIn order to avoid structural damage and contamination, no
hemicals were added during EPS extraction. Different iso-ates grown under similar conditions produced significantlyifferent EPS. The wild-type strain was high-EPS-producing189.6 mg L−1) and the sensitive mutant strain was low-EPS-roducing (105.7 mg L−1). As is well known, metabolic productsccumulate more readily in the outer layer of the cells in liquidulture. A variety of organic substances, such as polysaccharides,roteins, DNA, humic acid, and uronic acid were found in EPS [24].
Components of EPS are shown in Fig. 2. The order of productionas as follows: S-EPS > LB-EPS > TB-EPS. Production of S-EPS and
B-EPS in the wild-type strain was greater than in the mutant strain.owever, production of TB-EPS was lower in the wild-type than in
he mutant strain.Chemical analyses revealed measurable quantities of carbohy-
rates, proteins, and nucleic acid were present in the EPS of theild-type and mutant strains (Fig. 3). Content of the wild-type
nd mutant EPS were as follows: carbohydrates: S-EPS > TB-PS > LB-EPS; proteins: LB-EPS > S-EPS > TB-EPS; and nucleic acid:
Fig. 2. EPS production (%) of S-EPS, LB-EPS and TB-EPS of the wild-type and mutant.
, , .
TB-EPS > LB-EPS > S-EPS. The core components of all EPS were car-bohydrates (over 60%) [17]. Protein content in LB-EPS was over 25%and provided certain advantages. Nucleic acids in all EPS did notexceed 1.0%, indicating the cells did not undergo significant lysisduring the separation process. Moreover, production of carbohy-drates and proteins was reduced in the sensitive mutant, reducingthe tolerance of this strain in comparison to wild-type.
Proteins in EPS may be due to the presence of a large quantity ofexoenzymes. The easy uptake of readily available metal ion couldlead to the suggestion that high levels of exoenzymes are presentin the EPS matrix. The EPS carbohydrates were expected to playan important role in immobilizing these exoenzymes via reversiblecomplexation. Some groups of proteins combine with Cu2+, and thiscombination is an important factor in metal removal. For examplehomologues of c-cytochromes including MtrC and OmcA have beenidentified in EPS from Shewanella sp. HRCR-1 biofilms [25].
The ability of proteins to bind metallic cations could be involvedin the greater biosorption abilities of bound EPS. Protein com-prised the greatest portion of EPS and was strongly associated withremoval of Cu2+ [26,27]. Some groups of proteins in LB-EPS com-bine with Cu2+, and this combination is an important factor in metalremoval. Moreover, adsorption capacity was reduced due to thealtered protein structure and composition of the EPS [28].
3.2.2. Cu2+ removal of EPSThe effect of copper removal on EPS is shown in Fig. 4. The effi-
ciency of LB-EPS in the wild-type to remove copper is higher thanthose observed in the mutant. The order of removal ability wasas follows: LB-EPS > TB-EPS > S-EPS. The efficiency of LB-EPS in thewild-type to remove copper is higher than those observed in themutant.
We hypothesized that EPS protects bacteria from heavy metalions because their potency to interact with positively-chargedmetal. The protective effect of EPS was apparently not only asso-ciated with the quantity of EPS within the system, but also withthe location of that matrix outside of cell. Therefore, the protectionmechanism is not only related to the binding capacity of a systemas a whole, but to the availability of binding sites in a location thatprotects the bacteria from being a target for the metal.
The mobility and availability of heavy metals in environmentsare affected by the presence, forms (LB-EPS or TB-EPS), and quan-tities of EPS, as they are ubiquitous and have an excellent metalbinding capacity [17]. Comte et al. [17] study the numbers of totalpotential binding sites estimated by the protonic exchange capac-ity indicated another qualitative difference between soluble andbound EPS and revealed that different mechanisms could be impli-
cated in the metal biosorption of EPS, principally in the case of Cu .The bacteria could synthesize EPS to protect themselves againstharmful effects of metals. LB-EPS are loosely attached to the cellsurface whereas soluble EPS are situated in the solution. Depending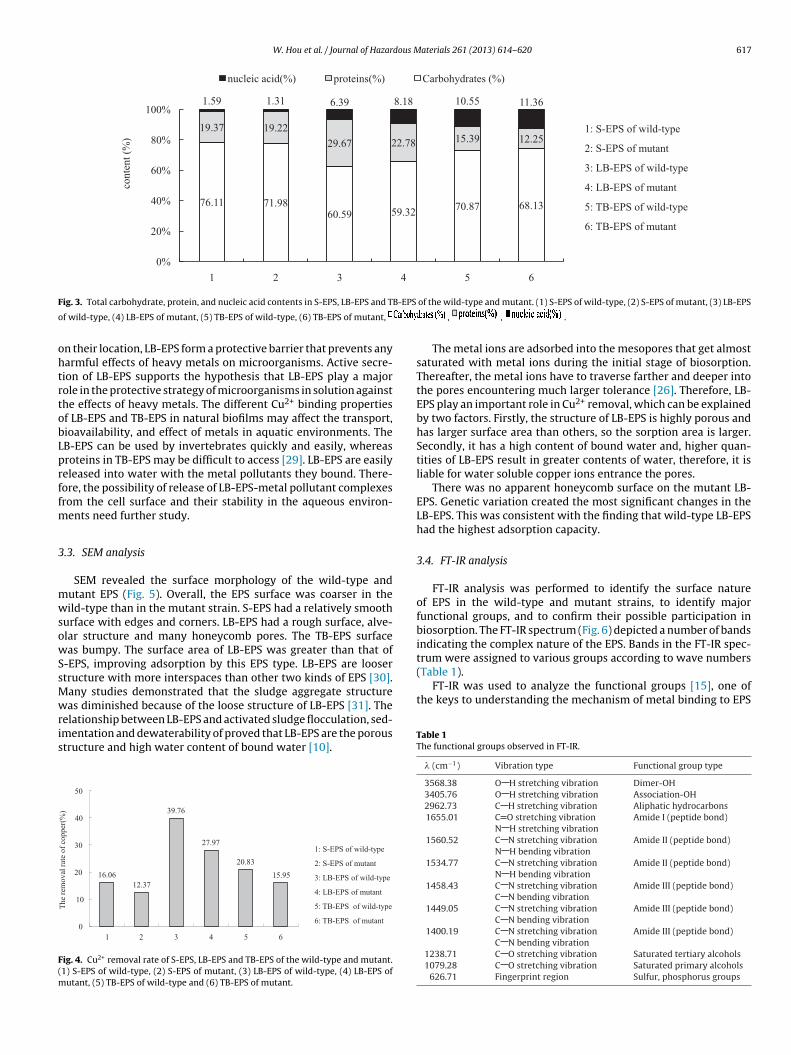
W. Hou et al. / Journal of Hazardous Materials 261 (2013) 614– 620 617
1: S-EPS of wild -type
2: S-EPS of mut ant
3: LB -EPS of wild -type
4: LB -EPS of mut ant
5: TB -EPS of wild -type
6: TB -EPS of mut ant
76.11 71.9860.59 59.32
70.87 68.13
19.37 19.22
29.67 22.78 15.39 12.25
1.59 1.31 6.39 8.18 10.55 11.36
0%
20%
40%
60%
80%
100%
1 2 3 4 5 6
con
ten
t (%
)
nucleic ac id( %) proteins (%) Carbohydrates (%)
F B-EPS
o
ohtrtobLprffm
3
mwsowSsMwris
F(m
trum were assigned to various groups according to wave numbers(Table 1).
FT-IR was used to analyze the functional groups [15], one ofthe keys to understanding the mechanism of metal binding to EPS
ig. 3. Total carbohydrate, protein, and nucleic acid contents in S-EPS, LB-EPS and T
f wild-type, (4) LB-EPS of mutant, (5) TB-EPS of wild-type, (6) TB-EPS of mutant,
n their location, LB-EPS form a protective barrier that prevents anyarmful effects of heavy metals on microorganisms. Active secre-ion of LB-EPS supports the hypothesis that LB-EPS play a majorole in the protective strategy of microorganisms in solution againsthe effects of heavy metals. The different Cu2+ binding propertiesf LB-EPS and TB-EPS in natural biofilms may affect the transport,ioavailability, and effect of metals in aquatic environments. TheB-EPS can be used by invertebrates quickly and easily, whereasroteins in TB-EPS may be difficult to access [29]. LB-EPS are easilyeleased into water with the metal pollutants they bound. There-ore, the possibility of release of LB-EPS-metal pollutant complexesrom the cell surface and their stability in the aqueous environ-
ents need further study.
.3. SEM analysis
SEM revealed the surface morphology of the wild-type andutant EPS (Fig. 5). Overall, the EPS surface was coarser in theild-type than in the mutant strain. S-EPS had a relatively smooth
urface with edges and corners. LB-EPS had a rough surface, alve-lar structure and many honeycomb pores. The TB-EPS surfaceas bumpy. The surface area of LB-EPS was greater than that of
-EPS, improving adsorption by this EPS type. LB-EPS are loosertructure with more interspaces than other two kinds of EPS [30].any studies demonstrated that the sludge aggregate structureas diminished because of the loose structure of LB-EPS [31]. The
elationship between LB-EPS and activated sludge flocculation, sed-mentation and dewaterability of proved that LB-EPS are the poroustructure and high water content of bound water [10].
1: S-EPS of wild -type
2: S-EPS of mu tant
3: LB-EPS of wild -type
4: LB-EPS of mu tant
5: TB-EPS of wild-type
6: TB-EPS of mutant
16.06
12.37
39.76
27. 97
20. 83
15.95
0
10
20
30
40
50
1 2 3 4 5 6
The
rem
ov
al r
ate
of
cop
per
(%)
ig. 4. Cu2+ removal rate of S-EPS, LB-EPS and TB-EPS of the wild-type and mutant.1) S-EPS of wild-type, (2) S-EPS of mutant, (3) LB-EPS of wild-type, (4) LB-EPS of
utant, (5) TB-EPS of wild-type and (6) TB-EPS of mutant.
of the wild-type and mutant. (1) S-EPS of wild-type, (2) S-EPS of mutant, (3) LB-EPS
, , .
The metal ions are adsorbed into the mesopores that get almostsaturated with metal ions during the initial stage of biosorption.Thereafter, the metal ions have to traverse farther and deeper intothe pores encountering much larger tolerance [26]. Therefore, LB-EPS play an important role in Cu2+ removal, which can be explainedby two factors. Firstly, the structure of LB-EPS is highly porous andhas larger surface area than others, so the sorption area is larger.Secondly, it has a high content of bound water and, higher quan-tities of LB-EPS result in greater contents of water, therefore, it isliable for water soluble copper ions entrance the pores.
There was no apparent honeycomb surface on the mutant LB-EPS. Genetic variation created the most significant changes in theLB-EPS. This was consistent with the finding that wild-type LB-EPShad the highest adsorption capacity.
3.4. FT-IR analysis
FT-IR analysis was performed to identify the surface natureof EPS in the wild-type and mutant strains, to identify majorfunctional groups, and to confirm their possible participation inbiosorption. The FT-IR spectrum (Fig. 6) depicted a number of bandsindicating the complex nature of the EPS. Bands in the FT-IR spec-
Table 1The functional groups observed in FT-IR.
� (cm−1) Vibration type Functional group type
3568.38 O H stretching vibration Dimer-OH3405.76 O H stretching vibration Association-OH2962.73 C H stretching vibration Aliphatic hydrocarbons1655.01 C O stretching vibration Amide I (peptide bond)
N H stretching vibration1560.52 C N stretching vibration Amide II (peptide bond)
N H bending vibration1534.77 C N stretching vibration Amide II (peptide bond)
N H bending vibration1458.43 C N stretching vibration Amide III (peptide bond)
C N bending vibration1449.05 C N stretching vibration Amide III (peptide bond)
C N bending vibration1400.19 C N stretching vibration Amide III (peptide bond)
C N bending vibration1238.71 C O stretching vibration Saturated tertiary alcohols1079.28 C O stretching vibration Saturated primary alcohols
626.71 Fingerprint region Sulfur, phosphorus groups

618 W. Hou et al. / Journal of Hazardous Materials 261 (2013) 614– 620
F wild-to
iwtcs(rtccc
vTa1gc
ig. 5. SEM of S-EPS, LB-EPS and TB-EPS of the wild-type and mutant. (1) S-EPS of
f wild-type, (6) TB-EPS of mutant.
n the wild-type and mutant strains. Our finding is in accordanceith others studies. Chowdhury et al. [32] used FT-IR to analyze
he biosorption of basic green 4 from aqueous solution by Ananasomosus leaf powder. The peaks were particularly evident, corre-ponding to the strong band of proteins and polysaccharides groupsTable 1). Zhang et al. [23] reported that the polysaccharides wereesponsible for metal removal. Guibaud et al. [5] demonstrated thathe protein was a part of the EPS and played a major role in theomplexation of metal ions and the number of binding sites; theomplexation constant was strongly linked to the protein, polysac-haride, and humic substance content.
The absorption band at 1655 cm−1 was assigned to the strongibration of C O and C N groups associated with peptide bonds.he 1540 cm−1 band represented the stretching vibration of C N
nd the deformation vibration N H of peptide bonds. Bands at410 cm−1 and 1240 cm−1 were assigned to the vibration of C Oroup of carboxylates and carboxylic acids. These groups areharacteristic of bacterial cell walls that are commonly found inype, (2) S-EPS of mutant, (3) LB-EPS of wild-type, (4) LB-EPS of mutant, (5) TB-EPS
biological wastewater treatment. EPS was polyanionic at circum-neutral pH and formed organometallic complexes with multivalentcations through electrostatic interactions.
The main differences between EPS samples from the wild-typeand mutant strains were associated with the amide II (N H bond-ing) vibrations. Shifts of the absorption banding from 1670 cm−1
to 1050 cm−1 were observed. The shifts were likely due to confor-mational changes associated with changes in H-bonds due to thegenetic mutation. Based on extensive infrared studies and results,we suggest the carboxylate, carboxylic, hydroxyl, and amino groupions are involved in EPS removal of heavy metal. The O H stretch-ing group and the C O stretching group were involved in metalbiosorption [15]. Some of the strength of the weaker groups indi-cated the presence of acidic or alkaline salts in the carboxyl group,
combined with some bands that suggest the minimal presence ofuronic acid (sugar notable band) and humic acid (CH2 and phe-nol); lipids and acid in the FT-IR spectrum was very difficult todetect [16,33]. Some bands in the fingerprint region of the nucleic
W. Hou et al. / Journal of Hazardous Materials 261 (2013) 614– 620 619
F wild
o
aTc
dst1uBtgcatpatp
t
ig. 6. FT-IR of S-EPS, LB-EPS and TB-EPS of the wild-type and mutant. (1) S-EPS off wild type and (6) TB-EPS of mutant.
cid composition might indicate the presence of phosphate groups.here was a possibility that the polysaccharides differed in chemi-al composition and hence had different affinities for Cu2+.
The FT-IR also showed that the major functional groupsiffered between the wild-type and mutant strains. This demon-trated the composition of S-EPS and TB-EPS had changed dueo genetic mutant. Strong absorption at 3427 cm−1, 1633 cm−1,067 cm−1, and 657 cm−1 were assigned to O H, peptide bond, sat-rated primary alcohols, sulfur, and phosphorus groups in LB-EPS.iosorption investigations demonstrated protonation or interac-ion between cations (H) and negative charges of acidic functionalroups in the polysaccharide. Alternatively, protein moieties of EPSould be the main macromolecules involved in metal ion binding,nd lack of binding by S-EPS and TB-EPS could be due to the facthat these EPS types are mainly composed of polysaccharides. Therotein/carbohydrate ratios of the EPS greatly influenced sorptionbility. The ratios suggest that extracellular proteins could pro-
ect cells against heavy metal substances. The results implied thatolyanionic groups in EPS were the primary binding sites for Cu2+.FT-IR analysis of EPS adsorption of copper ions confirmedhat, adsorption was mediated by proteins and polysaccharides.
type, (2) S-EPS of mutant, (3) LB-EPS of wild type, (4) LB-EPS of mutant, (5) TB-EPS
Polysaccharides and other extracellular proteins and polymers aregenerally negatively charged; therefore, the metal ions played abridging role. Previous analysis has demonstrated that proteins andpolysaccharides are the main components in EPS. In the subsurfaceenvironment, where nutrients are usually limited, microorganismstend to produce more EPS [21,34]. On a biomass basis, EPS in a sub-surface biofilm has a higher reactive surface area than cells becauseit is mostly a highly hydrated open matrix. Hence, EPS could bemuch more reactive than cells in terms of sorption, depending onthe relative density of sorption sites.
4. Conclusions
The results from this study facilitate a better understandingabout the roles of EPS in protecting microbial cells against heavymetals. We compared Cu2+ immobilization by three principaltypes of EPS of wild-type heavy metal-tolerance strain S. meliloti
CCNWSX0020 with sensitive mutant strain and demonstrated (1)LB-EPS played a more important role than S-EPS and TB-EPS inCu2+ immobilization; (2) SEM and FT-IR analysis further con-firm that proteins and carbohydrates were the main extracellular
6 dous M
cnmt
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
20 W. Hou et al. / Journal of Hazar
ompounds associated with tolerance and exhibited more pro-ounced copper binding properties. LB-EPS had rough surface andany honeycomb pores, making it conducive to copper entry;
herefore, they may play a role as a microbial protective barrier.
eferences
[1] B. Volesky, Z.R. Holan, Biosorption of heavy-metals, Biotechnol. Prog. 11 (1995)235–250.
[2] R. Nadeem, M.A. Hanif, A. Mahmood, M.S. Jamil, M. Ashraf, Biosorption of Cu(II)ions from aqueous effluents by blackgram bran (BGB), J. Hazard. Mater. 168(2009) 1622–1625.
[3] H. Liu, H.H.P. Fang, Characterization of electrostatic binding sites of extracel-lular polymers by linear programming analysis of titration data, Biotechnol.Bioeng. 80 (2002) 806–811.
[4] Y. Liu, M.C. Lam, H.H.P. Fang, Adsorption of heavy metals by EPS of activatedsludge, Water Sci. Technol. 43 (2001) 59–66.
[5] G. Guibaud, N. Tixier, A. Bouju, M. Baudu, Relation between extracellular poly-mers’ composition and its ability to complex Cd, Cu and Pb, Chemosphere 52(2003) 1701–1710.
[6] D. Zhang, J. Wang, X. Pan, Cadmium sorption by EPSs produced by anaer-obic sludge under sulfate-reducing conditions, J. Hazard. Mater. 138 (2006)589–593.
[7] X.F. Sun, S.G. Wang, X.M. Zhang, J.P. Chen, X.M. Li, B.Y. Gao, Y. Ma, Spectroscopicstudy of Zn2+ and Co2+ binding to extracellular polymeric substances (EPS) fromaerobic granules, J. Colloid Interface Sci. 335 (2009) 11–17.
[8] V.I. Slaveykova, N. Parthasarathy, K. Dedieu, D. Toescher, Role of extracellularcompounds in Cd-sequestration relative to Cd uptake by bacterium Sinorhizo-bium meliloti, Environ. Pollut. 158 (2010) 2561–2565.
[9] I.D. Henriques, N.G. Love, The role of extracellular polymeric substances in thetoxicity response of activated sludge bacteria to chemical toxins, Water Res. 41(2007) 4177–4185.
10] X.Y. Li, S.F. Yang, Influence of loosely bound extracellular polymeric substances(EPS) on the flocculation, sedimentation and dewaterability of activated sludge,Water Res. 41 (2007) 1022–1030.
11] G.P. Sheng, H.Q. Yu, X.Y. Li, Extracellular polymeric substances (EPS) ofmicrobial aggregates in biological wastewater treatment systems: a review,Biotechnol. Adv. 28 (2010) 882–894.
12] L.M. Fan, Z.Q. Ma, J.Q. Liang, H.F. Li, E.T. Wang, G.H. Wei, Characterization of acopper-resistant symbiotic bacterium isolated from Medicago lupulina growingin mine tailings, Bioresour. Technol. 102 (2011) 703–709.
13] C.P. Wolk, Y. Cai, J. Panoff, Use of a transposon with luciferase as a reporterto identify environmentally responsive genes in a cyanobacterium, Proc. Natl.Acad. Sci. U.S.A. 88 (1991) 5355–5359.
14] T.P.T. Chu, J.E. Kreutz, F.A. Neacsu, J.N. Shepherd, Investigation of the biosyn-thetic pathway of rhodoquinone by complementation of the Rhodospirillumrubrum F11 mutant through tri-parental mating, Abstr. Pap. Am. Chem. Soc.231 (2006), 735-CHEN.
15] S. Ozturk, B. Aslim, Z. Suludere, Evaluation of chromium(VI) removal behaviourby two isolates of Synechocystis sp. in terms of exopolysaccharide (EPS) produc-
tion and monomer composition, Bioresour. Technol. 100 (2009) 5588–5593.16] L. Domínguez, M. Rodríguez, D. Prats, Effect of different extraction methodson bound EPS from MBR sludges. Part I. Influence of extraction methods overthree-dimensional EEM fluorescence spectroscopy fingerprint, Desalination261 (2010) 19–26.
[
aterials 261 (2013) 614– 620
17] S. Comte, G. Guibaud, M. Baudu, Biosorption properties of extracellular poly-meric substances (EPS) resulting from activated sludge according to their type:soluble or bound, Process Biochem. 41 (2006) 815–823.
18] B. Zhang, B.S. Sun, M. Jin, T.S. Gong, Z.H. Gao, Extraction and analysis ofextracellular polymeric substances in membrane fouling in submerged MBR,Desalination 227 (2008) 286–294.
19] N. Ceyhan, G. Ozdemir, Extracellular polysaccharides produced by coolingwater tower biofilm bacteria and their possible degradation, Biofouling 24(2008) 129–135.
20] S.S. Adav, D.J. Lee, Extraction of extracellular polymeric substances from aer-obic granule with compact interior structure, J. Hazard. Mater. 154 (2008)1120–1126.
21] B. Cao, B. Ahmed, D.W. Kennedy, Z. Wang, L. Shi, M.J. Marshall, J.K. Fredrickson,N.G. Isern, P.D. Majors, H. Beyenal, Contribution of extracellular polymeric sub-stances from Shewanella sp. HRCR-1 biofilms to U(VI) immobilization, Environ.Sci. Technol. 45 (2011) 5483–5490.
22] P.M. Joshi, A.A. JuwarkarIn, In vivo studies to elucidate the role of extracellu-lar polymeric substances from Azotobacter in immobilization of heavy metals,Environ. Sci. Technol. 43 (2009) 5884–5889.
23] D.Y. Zhang, J.L. Wang, X.L. Pan, Cadmium sorption by EPSs produced by anaer-obic sludge under sulfate-reducing conditions, J. Hazard. Mater. 138 (2006)589–593.
24] H. Wang, H. Deng, L. Ma, L. Ge, Influence of operating conditions on extracellularpolymeric substances and surface properties of sludge flocs, Carbohydr. Polym.92 (2013) 510–515.
25] B. Cao, L. Shi, R.N. Brown, Y. Xiong, J.K. Fredrickson, M.F. Romine, M.J. Marshall,M.S. Lipton, H. Beyenal, Extracellular polymeric substances from Shewanella sp.HRCR-1 biofilms: characterization by infrared spectroscopy and proteomics,Environ. Microbiol. 13 (2011) 1018–1031.
26] Q.Q. Tao, Z.R. Liu, Y. Dai, X.X. Zhan, Biosorption properties of extracellular poly-meric substances towards Zn(II) and Cu(II), Desalin. Water Treat. 45 (2012)40–47.
27] G.P. Sheng, J. Xu, H.W. Luo, W.W. Li, W.H. Li, H.Q. Yu, Z. Xie, S.Q. Wei, F.C. Hu,Thermodynamic analysis on the binding of heavy metals onto extracellularpolymeric substances (EPS) of activated sludge, Water Res. 47 (2013) 607–614.
28] R.D. Lins, T.P. Straatsma, Computer simulation of the rough lipopolysaccharidemembrane of Pseudomonas aeruginosa, Biophys. J. 81 (2001) 1037–1046.
29] A.W. Decho, G.R. Lopez, Exopolymer microenvironments of microbial floramultiple and interactive effects on trophic relationships, Limnol. Oceanogr. 38(1993) 1633–1645.
30] G.P. Sheng, H.Q. Yu, X.Y. Li, Stability of sludge flocs under shear conditions:roles of extracellular polymeric substances (EPS), Biotechnol. Bioeng. 93 (2006)1095–1102.
31] H. Li, Y. Wen, A. Cao, J. Huang, Q. Zhou, P. Somasundaran, The influence ofadditives (Ca2+, Al3+, and Fe3+) on the interaction energy and loosely boundextracellular polymeric substances (EPS) of activated sludge and their floccu-lation mechanisms, Bioresour. Technol. 114 (2012) 188–194.
32] S. Chowdhury, S. Chakraborty, P. Saha, Biosorption of Basic Green 4 from aque-ous solution by Ananas comosus (pineapple) leaf powder, Colloids Surf. B 84(2011) 520–527.
33] P. D’Abzac, F. Bordas, E. Hullebusch, P.N.L. Lens, G. Guibaud, Extraction ofextracellular polymeric substances (EPS) from anaerobic granular sludges:
comparison of chemical and physical extraction protocols, Appl. Microbiol.Biotechnol. 85 (2009) 1589–1599.34] K. Myszka, K. Czaczyk, Characterization of adhesive exopolysaccharide (EPS)produced by Pseudomonas aeruginosa under starvation conditions, Curr. Micro-biol. 58 (2009) 541–546.
Copyright © 2022 FDOKUMEN




















