
Hypersalinity enhances the production of extracellular polymeric substance (eps) in the texas brown...
7
71 J. Phycol. 36, 71–77 (2000) HYPERSALINITY ENHANCES THE PRODUCTION OF EXTRACELLULAR POLYMERIC SUBSTANCE (EPS) IN THE TEXAS BROWN TIDE ALGA, AUREOUMBRA LAGUNENSIS (PELAGOPHYCEAE) 1 Hongbin Liu 2 and Edward J. Buskey 3 Marine Science Institute, The University of Texas at Austin, 750 Channelview Drive, Port Aransas, TX 78373 Laboratory experiments with batch cultures showed that the Texas brown tide alga, Aureoumbra lagunensis Stockwell, DeYoe, Hargraves et Johnson, produced a large amount of extracellular polymeric substance (EPS) that formed an extracellular polysaccharide mucus layer. Both dissolved and particulate carbohy- drate and EPS concentrations increased as A. lagun- ensis cultures progressed from exponential growth phase, through stationary phase, to declining phase. Particulate carbohydrate and EPS concentrations per cell were more than three times higher during the declining phase than that in exponential growth phase, reflecting a large increase in the EPS mucus layer. The amounts of carbohydrate and EPS pro- duced by A. lagunensis were significantly higher un- der hypersaline conditions. The thicker EPS mucus layer surrounding A. lagunensis cells under hypersa- line conditions might be a protective adaptation that permits it to bloom under hypersaline conditions that most other phytoplankton cannot survive. This could be one of the reasons why the Texas brown tide persisted in the Laguna Madre, an often hyper- saline coastal lagoon, for 7 years. Key index words: Texas brown tide; extracellular poly- meric substance (EPS); carbohydrate; Aureoumbra la- gunensis; salinity Abbreviations: DDW, distilled and deionized water; EPS, extracellular polymeric substance; TEP, trans- parent exopolymer particles The Texas brown tide, a bloom formed by Aureoum- bra lagunensis Stockwell, DeYoe, Hargraves et Johnson, had persisted without interruption for .7 years in La- guna Madre, Texas, until October 1997, when intense rains flushed the Laguna Madre and dispersed the bloom. The primary causes of the initiation of the bloom are thought to be related to extreme hypersa- line conditions caused by a long period of drought, which decreased populations of planktonic and benthic grazers (Buskey and Stockwell 1993, Buskey and Hyatt 1995, Buskey et al. 1997) and released phy- toplankton from grazing controls. A severe freeze in December 1990 caused a massive die-off of fish and other marine organisms, which released a large pulse of nutrients into the system to fuel the initial bloom (Stockwell et al. 1993, DeYoe and Suttle 1994). Salin- ity in the Laguna Madre remained high (45–75 psu) throughout the bloom years (Buskey et al. 1996). Lab- oratory studies have revealed that A. lagunensis can grow in a wide range of salinity (10–90 psu) and grows rapidly under hypersaline conditions (Buskey et al. 1998). One important advantage that allows A. lagun- ensis to grow well under hypersaline conditions might be that it possesses an extracellular matrix composed of polysaccharide (DeYoe et al. 1997). The excretion of extracellular polymeric substance (EPS) or photosynthetic extracellular release (PER) by phytoplankton is well documented (e.g., Myklestad and Haug 1972, Lancelot 1983, Schuster and Herndl 1995, Hong et al. 1997, Philippis et al. 1998). The loose slime of exopolymers and particles formed from coagulation of dissolved secretions has recently been classified as a new class of invisible organic particles in the ocean, called transparent exopolymer particles (TEP; Alldredge et al. 1993). These exudates are pri- marily composed of acidic polysaccharides, but can contain many other types of organic components (Fogg 1962, Myklestad et al. 1972, Decho 1990). These exudates can create a microenvironment for the cell that may allow it to function more efficiently (Decho 1990); the exudates may also protect the cells from environmental changes in salinity and tempera- ture (Boyle and Reade 1983, Decho 1990). Several studies have revealed that, in general, phytoplankton EPS production appears to increase during stationary growth phase and under nutrient-limited conditions (Myklestad and Haug 1972, Kroen and Rayburn 1984, Myklestad et al. 1989, Obernosterer and Herndl 1995). However, little is known about the effects of other environmental conditions, such as salinity, on the production of EPS by phytoplankton. The only re- search to date that has addressed the effect of salinity on phytoplankton EPS production is that of Allan et al. (1972), who studied the characteristics and pro- duction of EPS by the diatom Nitzschia frustulum in media with salinities ranging from 0.6 to 41.6 psu. In this study we examine the effects of growth phase, sa- linity, and bacteria on EPS production of A. lagunensis and consider the potential role of EPS production in the persistence of the brown tide bloom in the hyper- saline Laguna Madre. 1 Received 29 April 1999. Accepted 6 October 1999. 2 Present address: Institute for Hydrospheric-Atmospheric Sciences, Nagoya University, Chikusa-ku, Nagoya 464-8601, Japan; e-mail hliu@ ihas.nagoya-u.ac.jp. 3 Author for reprint requests; e-mail [email protected].
Transcript of Hypersalinity enhances the production of extracellular polymeric substance (eps) in the texas brown...

71
J. Phycol.
36,
71–77 (2000)
HYPERSALINITY ENHANCES THE PRODUCTION OF EXTRACELLULAR POLYMERIC SUBSTANCE (EPS) IN THE TEXAS BROWN TIDE ALGA,
AUREOUMBRA LAGUNENSIS
(PELAGOPHYCEAE)
1
Hongbin Liu
2
and Edward J. Buskey
3
Marine Science Institute, The University of Texas at Austin, 750 Channelview Drive, Port Aransas, TX 78373
Laboratory experiments with batch cultures showedthat the Texas brown tide alga,
Aureoumbra lagunensis
Stockwell, DeYoe, Hargraves et Johnson, produced alarge amount of extracellular polymeric substance(EPS) that formed an extracellular polysaccharidemucus layer. Both dissolved and particulate carbohy-drate and EPS concentrations increased as
A. lagun-ensis
cultures progressed from exponential growthphase, through stationary phase, to declining phase.Particulate carbohydrate and EPS concentrationsper cell were more than three times higher duringthe declining phase than that in exponential growthphase, reflecting a large increase in the EPS mucuslayer. The amounts of carbohydrate and EPS pro-duced by
A. lagunensis
were significantly higher un-der hypersaline conditions. The thicker EPS mucuslayer surrounding
A. lagunensis
cells under hypersa-line conditions might be a protective adaptation thatpermits it to bloom under hypersaline conditionsthat most other phytoplankton cannot survive. Thiscould be one of the reasons why the Texas browntide persisted in the Laguna Madre, an often hyper-saline coastal lagoon, for 7 years.
Key index words:
Texas brown tide; extracellular poly-meric substance (EPS); carbohydrate;
Aureoumbra la-gunensis
; salinity
Abbreviations:
DDW, distilled and deionized water;EPS, extracellular polymeric substance; TEP, trans-
parent exopolymer particles
The Texas brown tide, a bloom formed by
Aureoum-bra lagunensis
Stockwell, DeYoe, Hargraves et Johnson,had persisted without interruption for
.
7 years in La-guna Madre, Texas, until October 1997, when intenserains flushed the Laguna Madre and dispersed thebloom. The primary causes of the initiation of thebloom are thought to be related to extreme hypersa-line conditions caused by a long period of drought,which decreased populations of planktonic andbenthic grazers (Buskey and Stockwell 1993, Buskeyand Hyatt 1995, Buskey et al. 1997) and released phy-
toplankton from grazing controls. A severe freeze inDecember 1990 caused a massive die-off of fish andother marine organisms, which released a large pulseof nutrients into the system to fuel the initial bloom(Stockwell et al. 1993, DeYoe and Suttle 1994). Salin-ity in the Laguna Madre remained high (45–75 psu)throughout the bloom years (Buskey et al. 1996). Lab-oratory studies have revealed that
A. lagunensis
cangrow in a wide range of salinity (10–90 psu) and growsrapidly under hypersaline conditions (Buskey et al.1998). One important advantage that allows
A. lagun-ensis
to grow well under hypersaline conditions mightbe that it possesses an extracellular matrix composedof polysaccharide (DeYoe et al. 1997).
The excretion of extracellular polymeric substance(EPS) or photosynthetic extracellular release (PER)by phytoplankton is well documented (e.g., Myklestadand Haug 1972, Lancelot 1983, Schuster and Herndl1995, Hong et al. 1997, Philippis et al. 1998). Theloose slime of exopolymers and particles formed fromcoagulation of dissolved secretions has recently beenclassified as a new class of invisible organic particles inthe ocean, called transparent exopolymer particles(TEP; Alldredge et al. 1993). These exudates are pri-marily composed of acidic polysaccharides, but cancontain many other types of organic components(Fogg 1962, Myklestad et al. 1972, Decho 1990).These exudates can create a microenvironment forthe cell that may allow it to function more efficiently(Decho 1990); the exudates may also protect the cellsfrom environmental changes in salinity and tempera-ture (Boyle and Reade 1983, Decho 1990). Severalstudies have revealed that, in general, phytoplanktonEPS production appears to increase during stationarygrowth phase and under nutrient-limited conditions(Myklestad and Haug 1972, Kroen and Rayburn 1984,Myklestad et al. 1989, Obernosterer and Herndl1995). However, little is known about the effects ofother environmental conditions, such as salinity, onthe production of EPS by phytoplankton. The only re-search to date that has addressed the effect of salinityon phytoplankton EPS production is that of Allan etal. (1972), who studied the characteristics and pro-duction of EPS by the diatom
Nitzschia frustulum
inmedia with salinities ranging from 0.6 to 41.6 psu. Inthis study we examine the effects of growth phase, sa-linity, and bacteria on EPS production of
A. lagunensis
and consider the potential role of EPS production inthe persistence of the brown tide bloom in the hyper-saline Laguna Madre.
1
Received 29 April 1999. Accepted 6 October 1999.
2
Present address: Institute for Hydrospheric-Atmospheric Sciences,Nagoya University, Chikusa-ku, Nagoya 464-8601, Japan; e-mail [email protected].
3
Author for reprint requests; e-mail [email protected].
72
HONGBIN LIU AND EDWARD J. BUSKEY
materials and methods
Texas brown tide alga,
Aureoumbra lagunensis
(strain TBA-2),was cultured in modified f/2 media at salinities ranging from10 to 90 psu (see Buskey et al. 1998). Low salinities wereachieved by mixing seawater with distilled water; high salinitieswere achieved by adding hypersaline water produced by evapo-rating seawater in a vacuum chamber. The cultures were main-tained in 250 or 500 mL polycarbonate flasks at 22
8
C in a 12:12L/D cycle at approximately 120
m
M photons·m
2
2
·s
2
1
. The firstexperiment was conducted with cultures grown at 10 differentsalinities (10, 15, 20, 30, 40, 50, 60, 70, 80, 90 psu).
A. lagunensis
had been cultured at each of these salinities for more than 1year. During the experiment, cell densities were checked every1–2 days using a Z-1 Coulter Counter, and total carbohydratesand EPS concentration were measured every 2–3 days at ap-proximately the same time (10:00–12:00 h) during exponential,stationary, and declining growth phases. The second experimentwas carried out with cultures grown at 6 different salinities (20,30, 40, 50, 60, 70 psu). In addition to the measurements madein experiment 1, carbohydrates in dissolved and particulateforms were also determined. The cultures used in our experi-ments were not axenic. To assess the importance of bacteria onEPS release by
A. lagunensis
we compared our culture to anaxenic strain (CCMP1507, obtained from Bigelow Laboratoryfor Ocean Sciences) grown in the same media (salinity 32 psu)with growth conditions as described above.
Total carbohydrate concentration was determined spectro-photometrically with the phenol–sulfuric acid method usingglucose as a standard (Liu et al. 1973). Duplicate 2-mL culturesamples were each pipetted into colorimetric tubes, followed by1 mL of 10% phenol; then 5 mL of concentrated sulfuric acidwas added rapidly. The tubes were allowed to stand for 10 minat 25–30
8
C, shaken, then held for another 15 min. The absorp-tion at 485 nm was then determined against a reagent blank (2 mLdistilled and deionized water, DDW). Media blanks were alsodetermined, and results showed that carbohydrates in all mediawere not significantly higher than those in DDW. Duplicatesamples of 5–10 mL culture were filtered (0.4
m
m porosityPoretics polycarbonate membrane filter), and dissolved carbo-hydrates were analyzed in duplicate from 2-mL subsamples ofthe filtrates. Particulate carbohydrates were determined afterthe filter was soaked for 20 h in 5 mL of 80% sulfuric acid ac-cording to Passow et al. (1994). Comparison of a glucose stan-dard in water solution with hydrolysis procedures producedidentical values. We are aware that the so-called particulate car-bohydrates include both intracellular (storage) and extracellu-lar (EPS) carbohydrates. EPS was measured spectrophotometri-cally using alcian blue as a dye-binding assay (Passow andAlldredge 1995). Alcian blue binds to mucopolysaccharideswith carboxyl groups (Chamberlain 1976). Passow and All-dredge (1995) introduced this semiquantitative assay methodto measure the concentration of TEP. Because it stains bothcapsule and slime EPS, we use it here as an independent mea-surement of total EPS, although we are aware that there arecompositional differences between the two types of EPS.
results and discussion
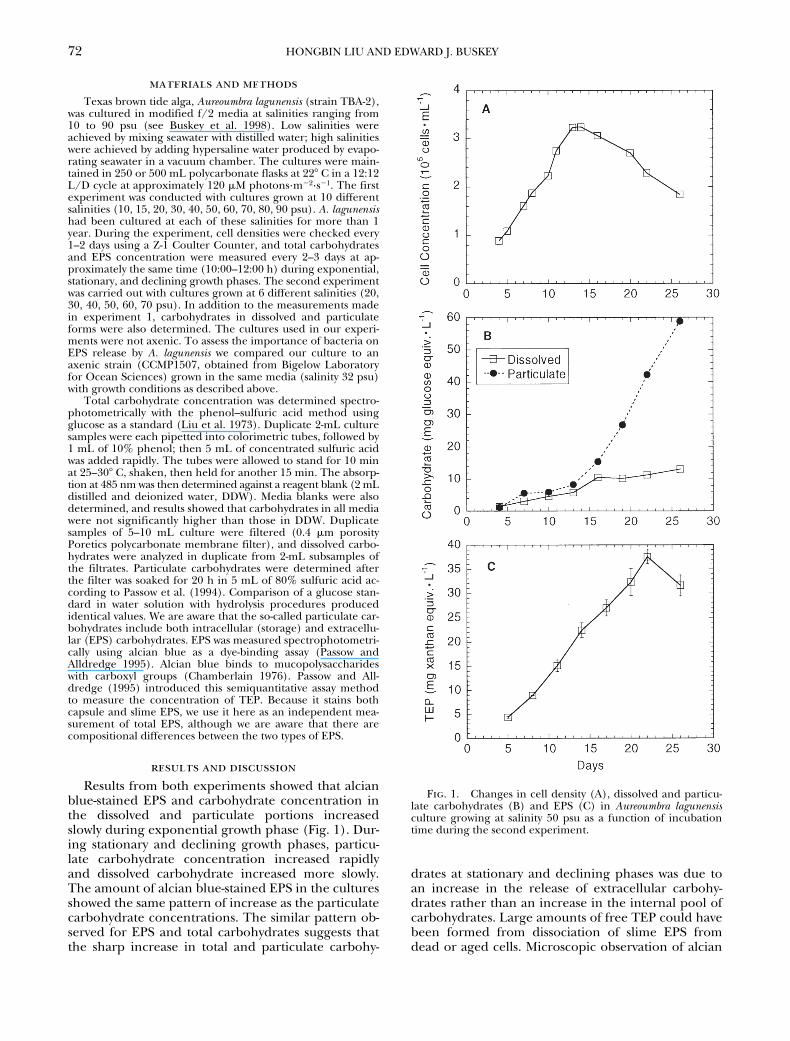
Results from both experiments showed that alcianblue-stained EPS and carbohydrate concentration inthe dissolved and particulate portions increasedslowly during exponential growth phase (Fig. 1). Dur-ing stationary and declining growth phases, particu-late carbohydrate concentration increased rapidlyand dissolved carbohydrate increased more slowly.The amount of alcian blue-stained EPS in the culturesshowed the same pattern of increase as the particulatecarbohydrate concentrations. The similar pattern ob-served for EPS and total carbohydrates suggests thatthe sharp increase in total and particulate carbohy-
drates at stationary and declining phases was due toan increase in the release of extracellular carbohy-drates rather than an increase in the internal pool ofcarbohydrates. Large amounts of free TEP could havebeen formed from dissociation of slime EPS fromdead or aged cells. Microscopic observation of alcian
Fig. 1. Changes in cell density (A), dissolved and particu-late carbohydrates (B) and EPS (C) in Aureoumbra lagunensisculture growing at salinity 50 psu as a function of incubationtime during the second experiment.
EFFECT OF SALINITY ON ALGAL EPS PRODUCTION
73
blue-stained slides (following the method of Passowand Alldredge 1994) confirmed the presence of abun-dant TEP, or slime EPS, in aged
A. lagunensis
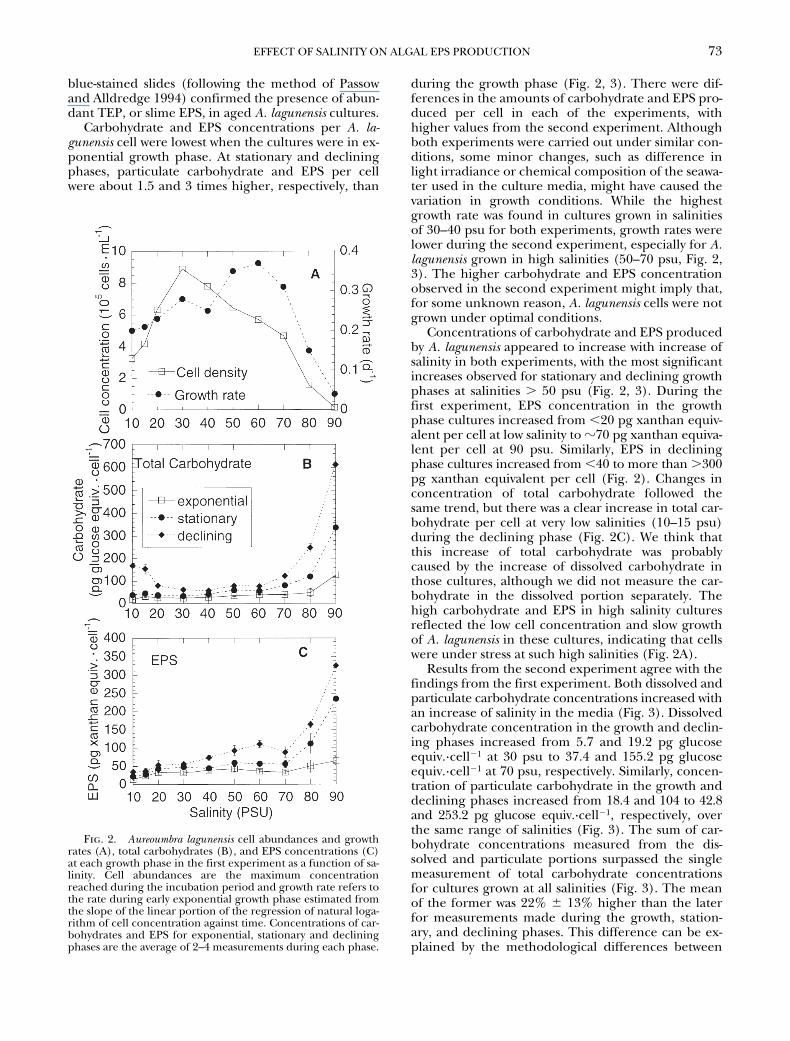
cultures.Carbohydrate and EPS concentrations per
A. la-gunensis
cell were lowest when the cultures were in ex-ponential growth phase. At stationary and decliningphases, particulate carbohydrate and EPS per cellwere about 1.5 and 3 times higher, respectively, than
during the growth phase (Fig. 2, 3). There were dif-ferences in the amounts of carbohydrate and EPS pro-duced per cell in each of the experiments, withhigher values from the second experiment. Althoughboth experiments were carried out under similar con-ditions, some minor changes, such as difference inlight irradiance or chemical composition of the seawa-ter used in the culture media, might have caused thevariation in growth conditions. While the highestgrowth rate was found in cultures grown in salinitiesof 30–40 psu for both experiments, growth rates werelower during the second experiment, especially for
A.lagunensis
grown in high salinities (50–70 psu, Fig. 2,3). The higher carbohydrate and EPS concentrationobserved in the second experiment might imply that,for some unknown reason,
A. lagunensis
cells were notgrown under optimal conditions.
Concentrations of carbohydrate and EPS producedby
A. lagunensis
appeared to increase with increase ofsalinity in both experiments, with the most significantincreases observed for stationary and declining growthphases at salinities
.
50 psu (Fig. 2, 3). During thefirst experiment, EPS concentration in the growthphase cultures increased from
,
20 pg xanthan equiv-alent per cell at low salinity to
z
70 pg xanthan equiva-lent per cell at 90 psu. Similarly, EPS in decliningphase cultures increased from
,
40 to more than
.
300pg xanthan equivalent per cell (Fig. 2). Changes inconcentration of total carbohydrate followed thesame trend, but there was a clear increase in total car-bohydrate per cell at very low salinities (10–15 psu)during the declining phase (Fig. 2C). We think thatthis increase of total carbohydrate was probablycaused by the increase of dissolved carbohydrate inthose cultures, although we did not measure the car-bohydrate in the dissolved portion separately. Thehigh carbohydrate and EPS in high salinity culturesreflected the low cell concentration and slow growthof
A. lagunensis
in these cultures, indicating that cellswere under stress at such high salinities (Fig. 2A).
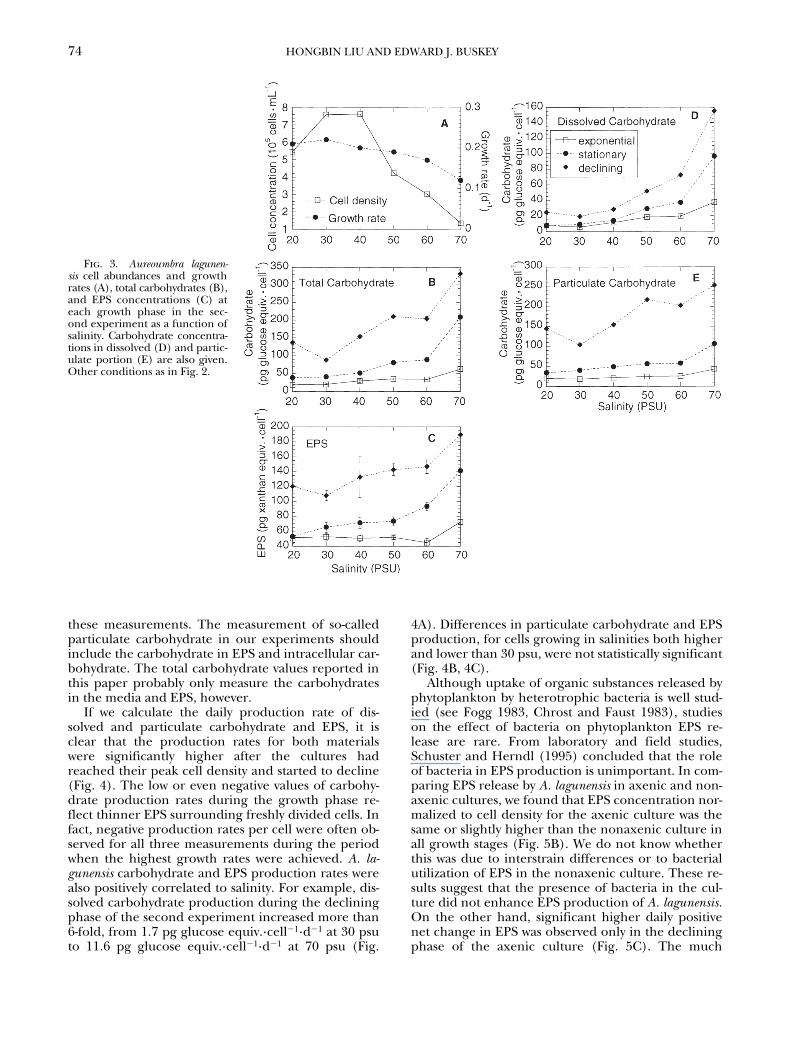
Results from the second experiment agree with thefindings from the first experiment. Both dissolved andparticulate carbohydrate concentrations increased withan increase of salinity in the media (Fig. 3). Dissolvedcarbohydrate concentration in the growth and declin-ing phases increased from 5.7 and 19.2 pg glucoseequiv.·cell
2
1
at 30 psu to 37.4 and 155.2 pg glucoseequiv.·cell
2
1
at 70 psu, respectively. Similarly, concen-tration of particulate carbohydrate in the growth anddeclining phases increased from 18.4 and 104 to 42.8and 253.2 pg glucose equiv.·cell
2
1
, respectively, overthe same range of salinities (Fig. 3). The sum of car-bohydrate concentrations measured from the dis-solved and particulate portions surpassed the singlemeasurement of total carbohydrate concentrationsfor cultures grown at all salinities (Fig. 3). The meanof the former was 22%
6
13% higher than the laterfor measurements made during the growth, station-ary, and declining phases. This difference can be ex-plained by the methodological differences between
Fig. 2. Aureoumbra lagunensis cell abundances and growthrates (A), total carbohydrates (B), and EPS concentrations (C)at each growth phase in the first experiment as a function of sa-linity. Cell abundances are the maximum concentrationreached during the incubation period and growth rate refers tothe rate during early exponential growth phase estimated fromthe slope of the linear portion of the regression of natural loga-rithm of cell concentration against time. Concentrations of car-bohydrates and EPS for exponential, stationary and decliningphases are the average of 2–4 measurements during each phase.
74
HONGBIN LIU AND EDWARD J. BUSKEY
these measurements. The measurement of so-calledparticulate carbohydrate in our experiments shouldinclude the carbohydrate in EPS and intracellular car-bohydrate. The total carbohydrate values reported inthis paper probably only measure the carbohydratesin the media and EPS, however.
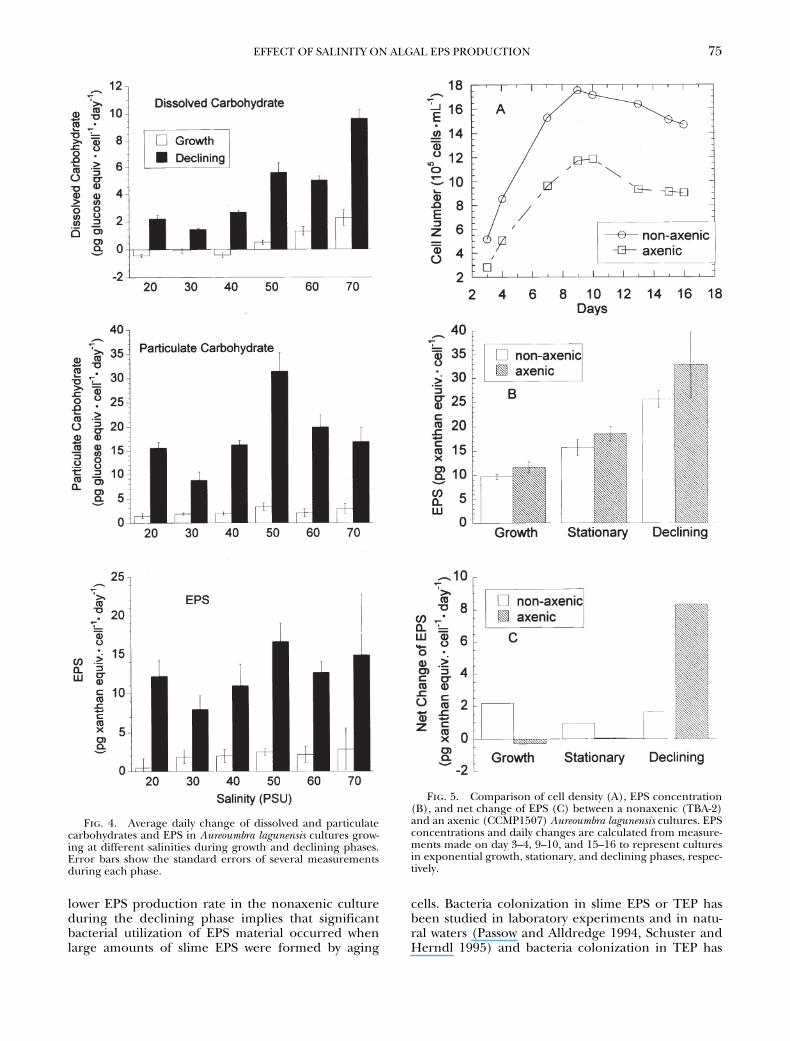
If we calculate the daily production rate of dis-solved and particulate carbohydrate and EPS, it isclear that the production rates for both materialswere significantly higher after the cultures hadreached their peak cell density and started to decline(Fig. 4). The low or even negative values of carbohy-drate production rates during the growth phase re-flect thinner EPS surrounding freshly divided cells. Infact, negative production rates per cell were often ob-served for all three measurements during the periodwhen the highest growth rates were achieved.
A. la-gunensis
carbohydrate and EPS production rates werealso positively correlated to salinity. For example, dis-solved carbohydrate production during the decliningphase of the second experiment increased more than6-fold, from 1.7 pg glucose equiv.·cell
2
1
·d
2
1
at 30 psuto 11.6 pg glucose equiv.·cell
2
1
·d
2
1
at 70 psu (Fig.
4A). Differences in particulate carbohydrate and EPSproduction, for cells growing in salinities both higherand lower than 30 psu, were not statistically significant(Fig. 4B, 4C).
Although uptake of organic substances released byphytoplankton by heterotrophic bacteria is well stud-ied (see Fogg 1983, Chrost and Faust 1983), studieson the effect of bacteria on phytoplankton EPS re-lease are rare. From laboratory and field studies,Schuster and Herndl (1995) concluded that the roleof bacteria in EPS production is unimportant. In com-paring EPS release by
A. lagunensis
in axenic and non-axenic cultures, we found that EPS concentration nor-malized to cell density for the axenic culture was thesame or slightly higher than the nonaxenic culture inall growth stages (Fig. 5B). We do not know whetherthis was due to interstrain differences or to bacterialutilization of EPS in the nonaxenic culture. These re-sults suggest that the presence of bacteria in the cul-ture did not enhance EPS production of
A. lagunensis.
On the other hand, significant higher daily positivenet change in EPS was observed only in the decliningphase of the axenic culture (Fig. 5C). The much
Fig. 3. Aureoumbra lagunen-sis cell abundances and growthrates (A), total carbohydrates (B),and EPS concentrations (C) ateach growth phase in the sec-ond experiment as a function ofsalinity. Carbohydrate concentra-tions in dissolved (D) and partic-ulate portion (E) are also given.Other conditions as in Fig. 2.
EFFECT OF SALINITY ON ALGAL EPS PRODUCTION
75
lower EPS production rate in the nonaxenic cultureduring the declining phase implies that significantbacterial utilization of EPS material occurred whenlarge amounts of slime EPS were formed by aging
cells. Bacteria colonization in slime EPS or TEP hasbeen studied in laboratory experiments and in natu-ral waters (Passow and Alldredge 1994, Schuster andHerndl 1995) and bacteria colonization in TEP has
Fig. 4. Average daily change of dissolved and particulatecarbohydrates and EPS in Aureoumbra lagunensis cultures grow-ing at different salinities during growth and declining phases.Error bars show the standard errors of several measurementsduring each phase.
Fig. 5. Comparison of cell density (A), EPS concentration(B), and net change of EPS (C) between a nonaxenic (TBA-2)and an axenic (CCMP1507) Aureoumbra lagunensis cultures. EPSconcentrations and daily changes are calculated from measure-ments made on day 3–4, 9–10, and 15–16 to represent culturesin exponential growth, stationary, and declining phases, respec-tively.

76
HONGBIN LIU AND EDWARD J. BUSKEY
been found to be positively correlated to the age ofTEP (Schuster and Herndl 1995). We have observednumerous bacteria attached to slime EPS or TEP in
A.lagunensis
cultures during declining phase.EPS production is common in many phytoplank-
ton groups, such as diatoms (Myklestad 1974), mi-croflagellates (Guillard and Wangersky 1958, Marker1965), and cyanobacteria (De Philippis et al. 1998). Ingeneral, release of EPS occurs at the highest rate dur-ing the stationary phase of growth (Myklestad 1974,Chrost and Faust 1983, Obernosterer and Herndl1995). Our findings that
A. lagunensis
produces moreEPS during stationary and declining phases agreeswith previous observations that phytoplankton usuallyrelease more EPS under stress, such as nutrient limita-tion. There are, however, few studies that address theinfluence of salinity on phytoplankton EPS releaseand therefore it is not well understood why
A. lagunen-sis
produces more EPS under hypersaline conditions.We think that enhanced EPS production by
A. lagun-ensis
under hypersaline conditions may be a protectiveadaptation. An increased EPS layer may help the cellsby creating a microenvironment surrounding the cellwall that buffers the osmotic disequilibrium across thecell membrane and protects the cell from dehydra-tion under hypersaline condition. Besides offeringprotection against high salinity environments, theEPS layer may also provide protection against grazers.The thicker EPS layer surrounding
A. lagunensis
cellsunder hypersaline condition may prevent it from be-ing grazed, either through inhibition of nano- and mi-crozooplankton feeding by clogging their feeding ap-paratus, or through resisting digestion and remainingviable after passage through a grazer’s gut. Furtherstudies are needed to test these hypotheses.
This study was supported by NSF grant (OCE-9529750). Wethank Tracey McDonnell for assisting with preliminary EPSstudies and Brian Wysor for establishing cultures at various sa-linities. Comments from Patricia Wheeler and two anonymousreviewers improved the manuscript considerably. University ofTexas Marine Science Institute Contribution number 1120.
Allan, G. G., Lewin, J. & Johnson, P. G. 1972. Marine polymers. IV.diatom polysaccharides.
Bot. Mar.
15:102–8.Alldredge, A. L., Passow, U. & Logan, B. E. 1993. The abundance
and significance of a class of large, transparent organic parti-cles in the ocean.
Deep-Sea Res. I.
40:1131–40.Boyle, D. C. & Reade, A. E. 1983. Characterization of two extracellu-
lar polysaccharides from marine bacteria.
Appl. Environ. Micro-biol.
46:392–9.Buskey, E. J. & Hyatt, C. J. 1995. Effects of the Texas (USA) “brown
tide” alga on planktonic grazers.
Mar. Ecol. Prog. Ser.
126:285–92.
Buskey, E. J., Montagna, P. A., Amos, A. F. & Whitledge, T. E. 1997.Disruption of grazer populations as a contributing factor to theinitiation of the Texas brown tide bloom.
Limnol. Oceanogr.
42:1215–22.Buskey, E. J., Stewart, S., Peterson, J. & Collumb, C. 1996.
Currentstatus and historical trends of brown tide and red tide phytoplanktonblooms in the Corpus Christi Bay National Estuary Program studyarea.
Corpus Christi Bay National Estuary Program, CCBNEP-1,90 pp.
Buskey, E. J. & Stockwell, D. A. 1993. Effects of a persistent “brown
tide” on zooplankton populations in the Laguna Madre ofsouth Texas.
In
Smayda, T. J. & Shimizu, Y. [Eds.]
Toxic Phy-toplankton Blooms in the Sea: Proceedings of the 5th Conference onToxic Marine Phytoplankton.
Elsevier, Amsterdam and New York,pp. 659–66.
Buskey, E. J., Wysor, B. & Hyatt, C. 1998. The role of hypersalinityin the persistence of the Texas “brown tide” in the LagunaMadre.
J. Plankton Res.
20:1553–65.Chamberlain, A. H. L. 1976. Algal settlement and secretion of adhe-
sive materials.
In
Sharpley, J. M. & Kaplan, A. M. [Eds.]
Proc.3rd Int. Biodegrad. Symp.
Applied Science Publishers, London,pp. 417–32.
Chrost, R. H., & Faust, M. A. 1983. Organic carbon release by phy-toplankton: its composition and utilization by bacterioplank-ton.
J. Plankton Res.
5:477–93.Decho, A. W. 1990. Microbial exopolymer secretions in ocean envi-
ronments: their roles in food webs and marine processes.
In
Bar-nes, M. [Ed.]
Oceanography and Marine Biology Annual Review
(Vol. 28). Aberdeen University Press, pp. 73–153.De Philippis, R., Margheri, M. C., Materassi, R. & Vincenzini, M.
1998. Potential of unicellular cyanobacteria from saline envi-ronments as exopolysaccharide producers.
Appl. Environ. Mi-crobiol.
64:1130–2.DeYoe, H. R., Stockwell, D. A., Bidigare, R. R., Latasa, M., Johnson,
P. W., Hargraves, P. E. & Suttle, C. A. 1997. Description andcharacterization of the algal species
Aureoumbra lagunensis
gen.et sp. nov. and referral of
Aureoumbra
and
Aureococcus
to thePelagophyceae.
J. Phycol.
33:1042–8.DeYoe, H. R. & Suttle, C. A. 1994. The inability of the Texas “brown
tide” alga to use nitrate and the role of nitrogen in the initia-tion of a persistent bloom of this organism.
J. Phycol.
30:800–6.Fogg, G. E. 1962. Extracellular products.
In
Lewin, R. A. [Ed.]
Phys-iology and Biochemistry of Algae.
Academic Press, New York. pp.475–89.
Fogg, G. E. 1983. The ecological significance of extracellular prod-ucts of phytoplankton photosynthesis.
Bot. Mar.
26:3–14.Guillard, R. R. L., & Wangersky, P. J. 1958. The production of extra-
cellular carbohydrates by some marine flagellates.
Limnol.Oceanogr.
3:449–54.Hong, Y., Smith, W. O., Jr. & White, A.-M. 1997. Studies on trans-
parent exopolymer particles (TEP) produced in the Ross Sea(Antarctica) and by
Phaeocystis antarctica
(Prymnesiophyceae).
J. Phycol.
33:368–76.Kroen, W. K. & Rayburn, W. R. 1984. Influence of growth status and
nutrients on extracellular polysaccharide synthesis by the soil alga
Chlamydomonas mexicana
(Chlorophyceae).
J. Phytol.
20:253–7.Lancelot, C. 1983. Factors affecting phytoplankton extracellular re-
lease in the Southern Bight of the North Sea.
Mar. Ecol. Prog.Ser.
12:115–21.Liu, D., Wong, P. T. S. & Dutka, B. J. 1973. Determination of carbo-
hydrate in lake sediment by a modified phenol–sulfuric acidmethod.
Water Res.
7:741–6.Marker, A. F. H. 1965. Extracellular carbohydrate liberation in the
flagellates
Isochrysis galbana
and
Prymnesium parvum.
J. Mar. Biol.Assoc. U.K.
45:755–72.Myklestad, S. 1974. Production of carbohydrates by marine plank-
tonic diatoms. I. Comparison of nine different species in cul-ture.
J. Exp. Mar. Biol. Ecol.
15:261–74.Myklestad, S. & Haug, A. 1972. Production of carbohydrates by the
marine diatom
Chaetoceros affinis
var.
willei
(Gran) Hustedt. I.Effect of the concentration of nutrients in the culture me-dium.
J. Exp. Mar. Biol. Ecol.
9:125–36.Myklestad, S., Haug, A. & Larsen, B. 1972. Production of carbohy-
drates by the marine diatom
Chaetoceros affinis
var.
willei
(Gran)Hustedt. II. Preliminary investigation of the extracellularpolysaccharide.
J. Exp. Mar. Biol. Ecol.
9:137–44.Myklestad, S., Holm-Hansen, O., Varum, K. M. & Volcani, B. E.
1989. Rate of release of extracellular amino acids and carbohy-drates from the marine diatom
Chaetoceros affinis.
J. PlanktonRes.
11:763–73.Obernosterer, I. & Herndl, G. J. 1995. Phytoplankton extracellular
release and bacterial growth: dependence on the inorganicN:P ratio.
Mar. Ecol. Prog. Ser.
116:247–57.

EFFECT OF SALINITY ON ALGAL EPS PRODUCTION
77
Passow, U. & Alldredge, A. L. 1994. Distribution, size and bacterialcolonization of transparent exopolymer particles (TEP) in theocean.
Mar. Ecol. Prog. Ser.
113:185–98.Passow, U. & Alldredge, A. L. 1995. A dye-binding assay for the
spectrophotometric measurement of transparent exopolymerparticles (TEP).
Limnol. Oceanogr.
40:1326–35.Passow, U., Alldredge, A. L. & Logan, B. E. 1994. The role of partic-
ulate carbohydrate exudates in the flocculation of diatomblooms.
Deep-Sea Res. I.
41:335–57.
Schuster, S. & Herndl, G. J. 1995. Formation and significance oftransparent exopolymeric particles in the northern AdriaticSea.
Mar. Ecol. Prog. Ser. 124:227–36.Stockwell, D. A., Buskey, E. J. & Whitledge, T. E. 1993. Studies on
conditions conducive to the development and maintenance ofa persistent “brown tide” in Laguna Madre, Texas. In Smayda,T. J. & Shimizu, Y. [Eds.] Toxic Phytoplankton Blooms in the Sea:Proceedings of the 5th Conference on Toxic Marine Phytoplankton.Elsevier, Amsterdam and New York, pp. 693–8.




















