
Habituation of Neurosecretory Responses to Extracellular ATP ...
8
The Journal of Neuroscience, August 1994, 14(8): 4831-4838 Habituation of Neurosecretory Responses to Extracellular ATP in PC 12 Cells Laura Cheever” and Daniel E. Koshland, Jr. Department of Molecular and Cell Biology, Division of Biochemistry and Molecular Biology, University of California at Berkeley, Berkeley, California 94720 Neuronally differentiated PC12 cells have been used as a model for habituation. These cells secrete norepinephrine in response to extracellular ATP, as well as other stimulants, and the response is diminished with repetitive stimulation. This loss of neurosecretory responsiveness displays char- acteristics commonly associated with habituative learning. These include an increase in both the rate and the relative degree of response loss with either increasing stimulation frequency or decreasing stimulus intensity, and selective generalization of the decreased responsiveness to heter- ologous stimuli. In PC12 cells, the loss of neurosecretory responsiveness appears to be entirely caused by stimula- tion-dependent inactivation of the ATP-gated cation chan- nel. Several unusual properties of this channel apparently allow its regulation to produce the behaviors associated with short-term habituation. Recovery of the ATP-gated channel requires several minutes, allowing habituation of responses to repetitive stimuli given at intervals of several minutes. Inactivation of the ATP-gated channel may be proportion- ately faster in response to lower ligand concentrations, al- lowing habituation to be more rapid in response to stimuli of weaker intensity. [Key words: receptor-operated channel, purinergic recep- tor, desensitization, inactivation, adaptation] Habituation is the process by which an organism learnsto di- minish its response to a stimuluswith repetition ofthat stimulus. Habituation is nearly ubiquitous (Harris, 1943) having been described in a wide variety of organisms including, but not limited to, protozoa (Wood, 1988) mollusks(cf. Pinsker et al., 1970), insects (cf. Corfas and Dudai, 1989), amphibians (cf. Fare1 et al., 1973),and mammals (cf. Thompson and Glanzman, 1976). A number of characteristic behaviors are common to habituative learning in a variety of behavioral studies (Thomp- sonand Spencer, 1966).These include an exponential decrease in the response to a stimulus as that stimulus is repeated and spontaneous recovery ofthe response when the stimulusis with- held. Following habituation and recovery of a response, the rate of habituation to a second series of repetitive stimuli is often increased, indicating a memory of prior habituation that may Received Sept. 8, 1993; revised Jan. 26, 1994; accepted Feb. 8, 1994. We thank Ann Fischer for her expert cell culturing. This work was supported by Grant DK09765 from the National Institutes of Health. Correspondence should be addressed to Daniel E. Koshland, Jr., at the above address. ,‘Present address: Howard Hughes Medical Institute, University of California at San Francisco, Third and Pamassus Avenue, Room U-426, San Francisco, CA 94 143-0724. Copyright 0 1994 Society for Neuroscience 0270-6474/94/144831-08$05.00/O endure long after the response has recovered (Peekeand Petri- novich, 1984). Both the rate and relative extent of response loss are typically increasedwith either increasing stimulation fre- quency or decreasing stimulus intensity. And, in some cases, the habituation of the response to one stimulus may be gener- alized to cause decreased responsiveness to heterologous stim- uli. In several systems (cf. Castellucci et al., 1970; Kupfermann et al., 1970; Fare1et al., 1973) habituation of behavioral re- sponses is correlated with decreased activity of neurons lying within the pathway that mediates the behavioral response. In some cases, it hasbeen shownthat reduction ofneurotransmitter release from the presynaptic cell is responsible for the decreased neuronal activity (Zucker, 1971; Castellucciand Kandel, 1974; Klein et al., 1980). PC12cells,rat pheochromocytoma cellsthat differentiate upon treatment with nerve growth factor to resembleclosely sym- pathetic neurons (Greene and Tischler, 1976; Dichter et al., 1977) are frequently used to study mechanisms of neuronal signal transduction and neurotransmitter release. PC12 cells take up radiolabeled norepinephrine (NE) and storeit in vesicles along with endogenous catecholamines (Greene and Rein, 1977b). The radiolabel is secretedin response to a variety of stimulants that elevate the cytosolic calcium concentration, in- cluding acetylcholine (Greene and Rein, 1977b) and depolar- ization (Greene and Rein, 1977a),permitting simple measure- ment of neurotransmitter release. Extracellular ATP has more recently been found to also stimulate norepinephrine release from PC12 cells (Inoue et al., 1989). ATP has long been pro- posedto act as a neurotransmitter (Holton and Holton, 1954) and evokes excitatory responses in both the CNS (Edwards et al., 1992)and PNS (cf. Yatani et al., 1982;Jahr and Jessel, 1983; Fyffe and Perl, 1984),including sympathetic neurons (cf. Siggins et al., 1977). In a previous study, we found that ATP-evoked NE release from PC12 cells is diminished with repetitive stimulation and displays several characteristics of both short- and long-term habituation (Cheever and Koshland, 1992). In particular, neu- rosecretory responsiveness decreases exponentially with re- peated application of ATP and recovers spontaneouslywhen stimulation is withheld. Following habituation and recovery of the response, .habituation occurs more rapidly to a second series of repetitive stimuli. Similar modulation of neurotransmitter release occurring in neurons mediating a behavioral response would cause habituation of that response. Sincethe mechanisms underlying neurosecretory modulation are likely to be similar in viva and in cultured cells, PC12 cells can provide a model systemfor investigating the molecular mechanisms underlying habituative processes (McFadden and Koshland, 1990a). Cell
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Habituation of Neurosecretory Responses to Extracellular ATP ...

The Journal of Neuroscience, August 1994, 14(8): 4831-4838
Habituation of Neurosecretory Responses to Extracellular ATP in PC 12 Cells
Laura Cheever” and Daniel E. Koshland, Jr.
Department of Molecular and Cell Biology, Division of Biochemistry and Molecular Biology, University of California at Berkeley, Berkeley, California 94720
Neuronally differentiated PC12 cells have been used as a model for habituation. These cells secrete norepinephrine in response to extracellular ATP, as well as other stimulants, and the response is diminished with repetitive stimulation. This loss of neurosecretory responsiveness displays char- acteristics commonly associated with habituative learning. These include an increase in both the rate and the relative degree of response loss with either increasing stimulation frequency or decreasing stimulus intensity, and selective generalization of the decreased responsiveness to heter- ologous stimuli. In PC12 cells, the loss of neurosecretory responsiveness appears to be entirely caused by stimula- tion-dependent inactivation of the ATP-gated cation chan- nel. Several unusual properties of this channel apparently allow its regulation to produce the behaviors associated with short-term habituation. Recovery of the ATP-gated channel requires several minutes, allowing habituation of responses to repetitive stimuli given at intervals of several minutes. Inactivation of the ATP-gated channel may be proportion- ately faster in response to lower ligand concentrations, al- lowing habituation to be more rapid in response to stimuli of weaker intensity.
[Key words: receptor-operated channel, purinergic recep- tor, desensitization, inactivation, adaptation]
Habituation is the process by which an organism learns to di- minish its response to a stimulus with repetition ofthat stimulus. Habituation is nearly ubiquitous (Harris, 1943) having been described in a wide variety of organisms including, but not limited to, protozoa (Wood, 1988) mollusks (cf. Pinsker et al., 1970), insects (cf. Corfas and Dudai, 1989), amphibians (cf. Fare1 et al., 1973), and mammals (cf. Thompson and Glanzman, 1976). A number of characteristic behaviors are common to habituative learning in a variety of behavioral studies (Thomp- son and Spencer, 1966). These include an exponential decrease in the response to a stimulus as that stimulus is repeated and spontaneous recovery ofthe response when the stimulus is with- held. Following habituation and recovery of a response, the rate of habituation to a second series of repetitive stimuli is often increased, indicating a memory of prior habituation that may
Received Sept. 8, 1993; revised Jan. 26, 1994; accepted Feb. 8, 1994. We thank Ann Fischer for her expert cell culturing. This work was supported
by Grant DK09765 from the National Institutes of Health. Correspondence should be addressed to Daniel E. Koshland, Jr., at the above
address. ,‘Present address: Howard Hughes Medical Institute, University of California
at San Francisco, Third and Pamassus Avenue, Room U-426, San Francisco, CA 94 143-0724. Copyright 0 1994 Society for Neuroscience 0270-6474/94/144831-08$05.00/O
endure long after the response has recovered (Peeke and Petri- novich, 1984). Both the rate and relative extent of response loss are typically increased with either increasing stimulation fre- quency or decreasing stimulus intensity. And, in some cases, the habituation of the response to one stimulus may be gener- alized to cause decreased responsiveness to heterologous stim- uli. In several systems (cf. Castellucci et al., 1970; Kupfermann et al., 1970; Fare1 et al., 1973) habituation of behavioral re- sponses is correlated with decreased activity of neurons lying within the pathway that mediates the behavioral response. In some cases, it has been shown that reduction ofneurotransmitter release from the presynaptic cell is responsible for the decreased neuronal activity (Zucker, 197 1; Castellucci and Kandel, 1974; Klein et al., 1980).
PC 12 cells, rat pheochromocytoma cells that differentiate upon treatment with nerve growth factor to resemble closely sym- pathetic neurons (Greene and Tischler, 1976; Dichter et al., 1977) are frequently used to study mechanisms of neuronal signal transduction and neurotransmitter release. PC 12 cells take up radiolabeled norepinephrine (NE) and store it in vesicles along with endogenous catecholamines (Greene and Rein, 1977b). The radiolabel is secreted in response to a variety of stimulants that elevate the cytosolic calcium concentration, in- cluding acetylcholine (Greene and Rein, 1977b) and depolar- ization (Greene and Rein, 1977a), permitting simple measure- ment of neurotransmitter release. Extracellular ATP has more recently been found to also stimulate norepinephrine release from PC12 cells (Inoue et al., 1989). ATP has long been pro- posed to act as a neurotransmitter (Holton and Holton, 1954) and evokes excitatory responses in both the CNS (Edwards et al., 1992) and PNS (cf. Yatani et al., 1982; Jahr and Jessel, 1983; Fyffe and Perl, 1984), including sympathetic neurons (cf. Siggins et al., 1977).
In a previous study, we found that ATP-evoked NE release from PC12 cells is diminished with repetitive stimulation and displays several characteristics of both short- and long-term habituation (Cheever and Koshland, 1992). In particular, neu- rosecretory responsiveness decreases exponentially with re- peated application of ATP and recovers spontaneously when stimulation is withheld. Following habituation and recovery of the response, .habituation occurs more rapidly to a second series of repetitive stimuli. Similar modulation of neurotransmitter release occurring in neurons mediating a behavioral response would cause habituation of that response. Since the mechanisms underlying neurosecretory modulation are likely to be similar in viva and in cultured cells, PC 12 cells can provide a model system for investigating the molecular mechanisms underlying habituative processes (McFadden and Koshland, 1990a). Cell
4832 Cheever and Koshland * Habituation to Extracellular ATP
lines offer the advantage of being more amenable to biochemical analysis than more complex neural preparations. In this report, PC 12 cells stimulated with ATP are tested to determine whether they display the remaining characteristics associated with ha- bituative learning, and the mechanisms of neurosecretory ha- bituation are explored.
Materials and Methods Cell culture. PC I2 cells (American Tissue Culture Collection) were grown either on cvtodex 3 beads (Sigma. St. Louis, MO) as previously described (McFadden and Koshland, i996a) or in suspension. Cells were differ- entiated with 50 @ml NGF (Harlan Bioproducts for Science, India- napolis) in RPM1 1640 (Irvine Scientific, Santa Ana, CA) containing 5% fetal bovine serum (HyClone, Logan, UT), 10% horse serum (JRH Biosciences. Lenexa. KS), supplemental glutamine, penicillin, and strep- tomycin for 6-7 d before use (Greene and Tischler, 1976).
‘H-NE release. The time course of ‘H-NE release from PC12 cells was determined exactly as previously described (Cheever and Koshland, 1992). ‘H-NE released from perfused cells was measured every 0. I min, and release at each time point was expressed as a percentage of the total ‘H-NE remaining in the cells at that time, in order to correct for the ongoing loss of radiolabel. To determine the rates and extent of habit- uation, the total amount of ‘H-NE released during each stimulus was corrected for basal release as well as ongoing loss of radiolabel. Data were then fit by curves described in the text by nonlinear regression (KALEIDAGRAPH, Abelbeck Software).
For single measurements of ‘H-NE release (see Figs. 3, 5, 6) cells grown and differentiated in suspension were washed with low K+ buffer (I25 mM NaCI, 4.8 mM KCI, I.3 mM CaCI,, 1.2 mM MgCI,, 1.2 mM K?HPO,. 5.6 mM D-glucose, and 25 mM HEPES, pH 7.4) by gentle centrifugation and resuspension, and were then loaded with I &/ml ‘H-NE in low K + for 1 hr at 37°C. Unincorporated ‘H-NE was removed by extensive washing and cells were resuspended in low K ’ Fifty mi- croliters of stimulant were added to 50 ~1 aliquots ofcells and incubated at 25°C for the indicated stimulation times. Cells were removed by centrifugation and the amount of ‘H-NE released was determined by scintillation counting of an aliquot of the supernatant. The cells were then lysed with 1% Triton X- 100 in order to determine the total amount of ‘H-NE that had been present; released ‘H-NE is expressed as a per- centage of the total taken up by the cells. For each measurement of stimulated NE release, ‘H-NE in the supernatant of unstimulated cells treated in parallel was subtracted.
Intracellular calcium Ierels. Suspension cultures of differentiated PC1 2 cells were loaded with fura- by incubation for I hr at 37°C with I FM
fura-2-AM (Molecular Probes, Portland, OR) in 10 ml oflow K+ buffer. Free fura-2-AM was washed away, and cells were resuspended in 2 ml low K+ buffer. During calcium measurements, cells were maintained at 25°C with constant eentle stirring in the chamber of an SLM 4800s iuorimeter. For each time point,-the fluorescence intensity of the dye emitting at 5 IO nm was determined at excitation wavelengths of 340 and 380 nm in rapid succession. Three minutes after a stable basal recording was established, ATP in 0.5 ml low K+ was added. Where indicated, calcium-free low K+ containing 0.1 rnM EGTA was used. The intracellular calcium concentration at each time point was calcu- lated from the ratio of the fluorescent intensities excited at 340 and 380 nm according to the equation of Grynkiewicz et al. (1985) using a dissociation constant estimated at 200 nM for the binding of Cal+ to the dye. For each set of cells, the maximum ratio of 340:380 nm flu- orescence was determined after the stimulation period by lysing the cells with 1% Triton X-100 into low K+ buffer containing 1.3 ins CaCI,, and the minimum ratio of fluorescence was determined by subsequently adding EGTA to 80 mM. For each value, the autofluorescence of cells which had been similarly treated except for the absence of dye was subtracted.
Results
ATP-stimulated norepinephrine release meets criteria .for habituation During habituative learning, the loss of response with repetitive stimulation is typically faster and more complete when the fre- quency of repetitive stimulation is increased. This effect of stim- ulation freauencv on habituation is always detected when re-
0 20 40 60 80 I”00
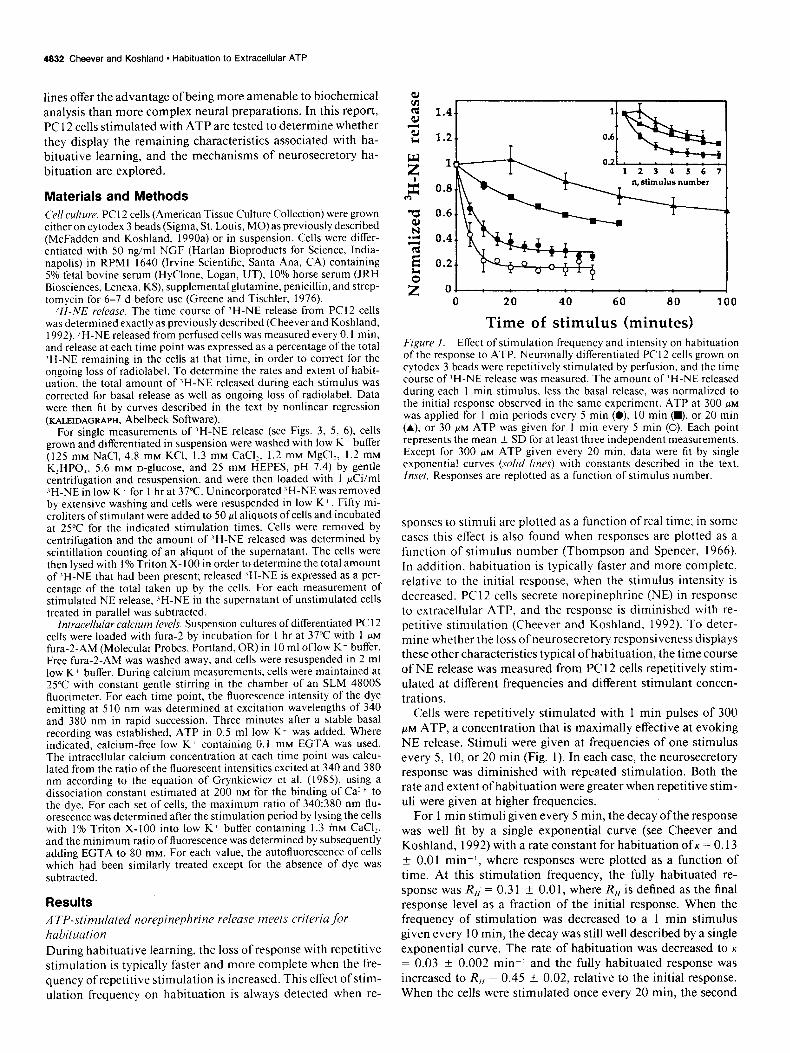
Time of stimulus (minutes) Figure 1. Effect of stimulation frequency and intensity on habituation of the response to ATP. Neuronally differentiated PC I2 cells grown on cytodex 3 beads were repetitively stimulated by perfusion, and the time course of ‘H-NE release was measured. The amount of ‘H-NE released during each I min stimulus, less the basal release, was normalized to the initial response observed in the same experiment. ATP at 300 PM
was applied for I min periods every 5 min (0) IO min (m), or 20 min (A). or 30 FM ATP was given for I min every 5 min (0). Each point represents the mean f SD for at least three independent measurements. Except for 300 PM ATP given every 20 min. data were fit by single exponential curves (so/id lines) with constants described in the text. Inset, Responses are replotted as a function of stimulus number.
sponses to stimuli are plotted as a function of real time; in some cases this effect is also found when responses are plotted as a function of stimulus number (Thompson and Spencer, 1966). In addition, habituation is typically faster and more complete, relative to the initial response, when the stimulus intensity is decreased. PC 12 cells secrete norepinephrine (NE) in response to extracellular ATP, and the response is diminished with re- petitive stimulation (Cheever and Koshland, 1992). To deter- mine whether the loss ofneurosecretory responsiveness displays these other characteristics typical ofhabituation, the time course of NE release was measured from PC 12 cells repetitively stim- ulated at different frequencies and different stimulant concen- trations.
Cells were repetitively stimulated with 1 min pulses of 300 PM ATP, a concentration that is maximally effective at evoking NE release. Stimuli were given at frequencies of one stimulus every 5, 10, or 20 min (Fig. 1). In each case, the neurosecretory response was diminished with repeated stimulation. Both the rate and extent of habituation were greater when repetitive stim- uli were given at higher frequencies.
For 1 min stimuli given every 5 min, the decay ofthe response was well fit by a single exponential curve (see Cheever and Koshland, 1992) with a rate constant for habituation of K = 0.13 + 0.01 min-I, where responses were plotted as a function of time. At this stimulation frequency, the fully habituated re- sponse was R,,‘= 0.31 4 0.01, where R,, is defined as the final response level as a fraction of the initial response. When the frequency of stimulation was decreased to a 1 min stimulus given every 10 min, the decay was still well described by a single exponential curve. The rate of habituation was decreased to K
= 0.03 ? 0.002 min’ and the fully habituated response was increased to R,, = 0.45 * 0.02, relative to the initial response. When the cells were stimulated once every 20 min. the second
A
I ATP
HK
40 60 a0 100 120 140
Perfusion time (minutes)
Perfusion time (minutes)
12 3 4 5
c HK
n
q .-g--o--Ef--El
12 3 4 5
Stimulus number
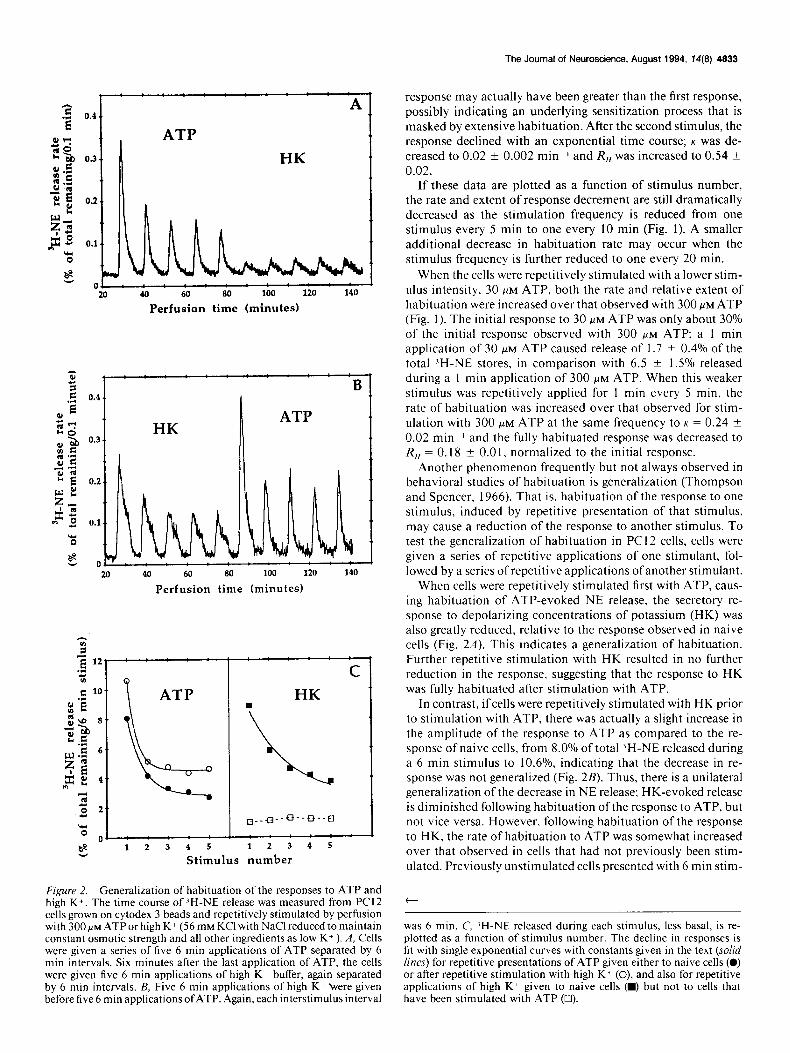
Figure 2. Generalization of habituation of the responses to ATP and high K+. The time course of ZH-NE release was measured from PC12 cells grown on cytodex 3 beads and repetitively stimulated by perfusion with 300 PM ATP or high K+ (56 mM KC1 with NaCl reduced to maintain constant osmotic strength and all other ingredients as low K+ ). A, Cells were given a series of five 6 min applications of ATP separated by 6 min intervals. Six minutes after the last application of ATP, the cells were given five 6 min applications of high K+ buffer, again separated by 6 min intervals. B, Five 6 min applications of high K+ were given before five 6 min applications ofATP. Again, each interstimulus interval
The Journal of Neuroscience, August 1994, 74(8) 4833
response may actually have been greater than the first response, possibly indicating an underlying sensitization process that is masked by extensive habituation. After the second stimulus, the response declined with an exponential time course; K was de- creased to 0.02 ? 0.002 minm’ and R,, was increased to 0.54 ? 0.02.
I f these data are plotted as a function of stimulus number, the rate and extent of response decrement are still dramatically decreased as the stimulation frequency is reduced from one stimulus every 5 min to one every 10 min (Fig. 1). A smaller additional decrease in habituation rate may occur when the stimulus frequency is further reduced to one every 20 min.
When the cells were repetitively stimulated with a lower stim- ulus intensity, 30 FM ATP, both the rate and relative extent of habituation were increased over that observed with 300 WM ATP (Fig. 1). The initial response to 30 FM ATP was only about 30% of the initial response observed with 300 HM ATP; a 1 min application of 30 FM ATP caused release of 1.7 + 0.4% of the total ‘H-NE stores, in comparison with 6.5 ? 1.5% released during a 1 min application of 300 FM ATP. When this weaker stimulus was repetitively applied for 1 min every 5 min, the rate of habituation was increased over that observed for stim- ulation with 300 FM ATP at the same frequency to K = 0.24 k 0.02 min-I and the fully habituated response was decreased to R,, = 0.18 + 0.0 1, normalized to the initial response.
Another phenomenon frequently but not always observed in behavioral studies of habituation is generalization (Thompson and Spencer, 1966). That is, habituation of the response to one stimulus, induced by repetitive presentation of that stimulus, may cause a reduction of the response to another stimulus. To test the generalization of habituation in PC12 cells, cells were given a series of repetitive applications of one stimulant, fol- lowed by a series of repetitive applications of another stimulant.
When cells were repetitively stimulated first with ATP, caus- ing habituation of ATP-evoked NE release, the secretory re- sponse to depolarizing concentrations of potassium (HK) was also greatly reduced, relative to the response observed in naive cells (Fig. 2A). This indicates a generalization of habituation. Further repetitive stimulation with HK resulted in no further reduction in the response, suggesting that the response to HK was fully habituated after stimulation with ATP.
In contrast, ifcells were repetitively stimulated with HK prior to stimulation with ATP, there was actually a slight increase in the amplitude of the response to ATP as compared to the re- sponse of naive cells, from 8.0% of total ‘H-NE released during a 6 min stimulus to 10.6%, indicating that the decrease in re- sponse was not generalized (Fig. 2B). Thus, there is a unilateral generalization of the decrease in NE release; HK-evoked release is diminished following habituation ofthe response to ATP, but not vice versa. However, following habituation of the response to HK, the rate of habituation to ATP was somewhat increased over that observed in cells that had not previously been stim- ulated. Previously unstimulated cells presented with 6 min stim-
t
was 6 min. C, ‘H-NE released during each stimulus, less basal, is re- plotted as a function of stimulus number. The decline in responses is fit with single exponential curves with constants given in the text (solid lines) for repetitive presentations of ATP given either to naive cells (0) or after repetitive stimulation with high K + (0), and also for repetitive applications of high K+ given to naive cells (w) but not to cells that have been stimulated with ATP (0).

4834 Cheever and Koshland * Habituation to Extracellular ATP
Table 1. Rate constants for decay of Ca; after application of the indicated ATP concentrations
450 IA t-l [ATP], pM: I
[ATPI, UM
Decay constant, min-l SD n
10 1.76 0.20 3 30 0.92 0.19 4
100 0.58 0.05 4 300 0.42 0.06 4
1000 0.41 0.09 3
uli of 300 PM ATP separated by 6 min intervals habituated with a rate constant of K = 0.11 f 0.0 1 mini. Following repetitive stimulation with HK, the rate constant for habituation was in- creased to K = 0.19 + 0.03 mini (Fig. 2C). This suggests a different sort of generalization in which habituation of the re- sponse to one stimulus does not reduce the response to another stimulant but does cause the response to that stimulant to de- crease more rapidly with repetitive stimulation (see Peeke and Petrinovich, 1984).
Thus, in PC12 cells as in animal studies, habituation of the response to one stimulant may be generalized to cause either a reduction in the response or an increased rate of habituation to another stimulant. The degree and type of generalization seen depend on the stimulants used. These interactions of the effects ofdifferent stimulants may offer clues as to the molecular mech- anisms causing decreases in the response and increases in the habituation rate.
.4 TP receptors mediating norepinephrine release
In PC12 cells, extracellular ATP exerts its effects by opening a receptor-operated cation channel that allows influx of calcium (moue, 1989) causing exocytotic release ofnorepinephrine. Thus, addition of ATP to the cells causes a dose-dependent increase in intracellular calcium concentrations (Ca,); Ca, rises rapidly upon addition of ATP and then decays slowly toward the basal level (Fig. 3A). The decay of Ca, is exponential with the rate constants described in Table 1. In the absence of extracellular calcium, no calcium influx can occur and ATP does not cause any detectable increase in Ca, (Fig. 3A). Similarly, ATP stim- ulates a dose-dependent release of NE from the cells (Fig. 3B). In the absence of extracellular calcium, ATP-stimulated NE release is also undetectable. These data suggest that ATP-evoked NE release is entirely mediated by calcium influx through ATP- gated channels.
Others working with a different strain of undifferentiated PC 12 cells have suggested that a second PzY-type ATP receptor is also present on the surface of PC 12 cells (Fasolato et al., 1990). PZ,- type receptors (for nomenclature, see Gordon, 1986; Williams and Cusack, 1990) are expected to act via G-proteins to stim- ulate phospholipase C activity (Dubyak et al., 1988; Boyer et al., 1989; Sasakawa et al., 1989) producing inositol triphosphate and diacylglycerol and thereby causing release of calcium from intracellular stores (Berridge and Irvine, 1984) and activation of protein kinase C (cf. Nishizuka, 1992; but see Gonzalez et al., 1989). In contrast to this strain of PC 12 cells, our PC 12 cells (from ATCC) display no increase in Ca, in the absence of ex- tracellular calcium (Fig. 3A), making it unlikely that ATP is
-1 10 100 1000
[ATPI @MI
Figure 3. ‘H-NE release and Ca, elevations evoked by ATP. In order to quantify the relationship between ATP-evoked increases in Ca, and NE release, elevations in Ca, evoked by various concentrations of ATP (A) and ‘H-NE release evoked with similar treatment(B) were measured. A, Cells preloaded with fura- were resuspended in low K+ (0, 0, 0, n ) or calcium-free low K+ (A), and intracellular calcium levels were measured. Three minutes after establishing a baseline Ca, level, ATP was added to a concentration of 10 (0), 30 (O), 100 (Cl), or 300 (w, A) /IM. B, Cells that had been preloaded with ‘H-NE were incubated with the indicated concentrations of ATP in low K+ (0) or calcium-free low K+ (0) for 6 min and the percentage of ZH-NE that was released into the supematant was determined. Points represent the mean ? SD of at least six independent measurements, each made in duplicate.
acting via a P,, receptor to stimulate release of calcium from intracellular stores.
Inactivation of the ATP-gated channel is responsible for decreased NE release
Reversible, stimulation-dependent inactivation of the machin- ery mediating the response to stimulation may be the simplest mechanism that could cause short-term habituation (Cheever and Koshland, 1992). For ATP-stimulated NE release, this sort of feedback inactivation could potentially occur at several points. The ATP-gated channel allowing influx of calcium could be inactivated during presentation of ATP, or some component of the secretory machinery that mediates norepinephrine release in response to cytosolic calcium might be inactivated. Alter- natively, ATP might increase the activity of pumps or buffers
The Journal of Neuroscience, August 1994, 74(8) 4335
k u “0 5 10 15 20 25 Incubation time (minutes)
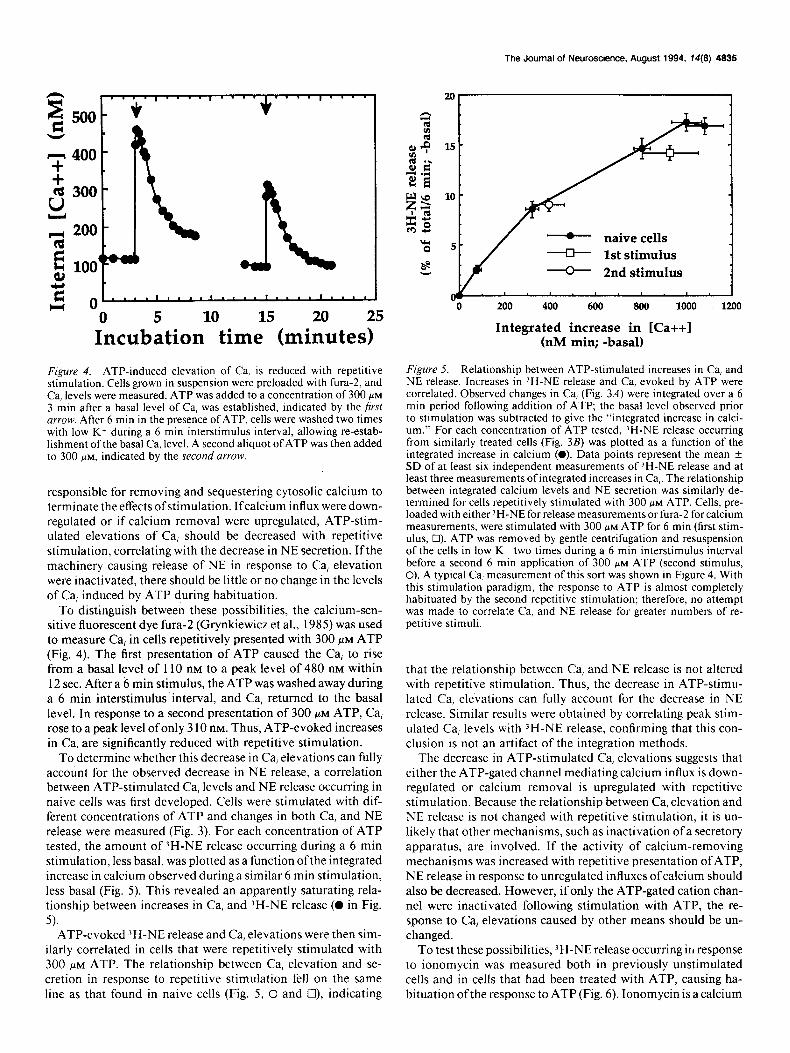
Figure 4. ATP-induced elevation of Ca, is reduced with repetitive stimulation. Cells grown in suspension were preloaded with fura-2, and Ca, levels were measured. ATP was added to a concentration of 300 PM
3 min after a basal level of Ca, was established, indicated by the first urrow. After 6 min in the presence of ATP, cells were washed two times with low K+ during a 6 min interstimulus interval, allowing re-estab- lishment of the basal Ca, level. A second aliquot of ATP was then added to 300 PM, indicated by the second arrow.
responsible for removing and sequestering cytosolic calcium to terminate the effects ofstimulation. Ifcalcium influx were down- regulated or if calcium removal were upregulated, ATP-stim- ulated elevations of Ca, should be decreased with repetitive stimulation, correlating with the decrease in NE secretion. Ifthe machinery causing release of NE in response to Ca, elevation were inactivated, there should be little or no change in the levels of Ca, induced by ATP during habituation.
To distinguish between these possibilities, the calcium-sen- sitive fluorescent dye fura- (Grynkiewicz et al., 1985) was used to measure Ca, in cells repetitively presented with 300 FM ATP (Fig. 4). The first presentation of ATP caused the Ca, to rise from a basal level of 110 nM to a peak level of 480 nM within 12 sec. After a 6 min stimulus, the ATP was washed a)vay during a 6 min interstimulus interval, and Ca, returned to the basal level. In response to a second presentation of 300 FM ATP, Ca, rose to a peak level of only 3 10 nM. Thus, ATP-evoked increases in Ca, are significantly reduced with repetitive stimulation.
To determine whether this decrease in Ca, elevations can fully account for the observed decrease in NE release, a correlation between ATP-stimulated Ca, levels and NE release occurring in naive cells was first developed. Cells were stimulated with dif- ferent concentrations of ATP and changes in both Ca, and NE release were measured (Fig. 3). For each concentration of ATP tested, the amount of )H-NE release occurring during a 6 min stimulation, less basal, was plotted as a function ofthe integrated increase in calcium observed during a similar 6 min stimulation, less basal (Fig. 5). This revealed an apparently saturating rela- tionship between increases in Ca, and JH-NE release (0 in Fig.
5). ATP-evoked XH-NE release and Ca, elevations were then sim-
ilarly correlated in cells that were repetitively stimulated with 300 FM ATP. The relationship between Ca, elevation and se- cretion in response to repetitive stimulation fell on the same line as that found in naive cells (Fig. 5, 0 and Cl), indicating
- naive cells -[)- 1st stimulus + 2nd stimulus
200 400 600 800 1000 1200
Integrated increase in [Ca++l (nM min; -basal)
Figure 5. Relationship between ATP-stimulated increases in Ca, and NE release. Increases in ‘H-NE release and Ca, evoked by ATP were correlated. Observed changes in Ca, (Fig. 3A) were integrated over a 6 min period following addition of ATP; the basal level observed prior to stimulation was subtracted to give the “integrated increase in calci- um.” For each concentration of ATP tested, ‘H-NE release occurring from similarly treated cells (Fig. 3B) was plotted as a function of the integrated increase in calcium (0). Data points represent the mean i SD of at least six independent measurements of ‘H-NE release and at least three measurements ofintegrated increases in Ca,. The relationship between integrated calcium levels and NE secretion was similarly de- termined for cells repetitively stimulated with 300 FM ATP. Cells, pre- loaded with either ‘H-NE for release measurements or fura- for calcium measurements, were stimulated with 300 PM ATP for 6 min (first stim- ulus, Cl). ATP was removed by gentle centrifugation and resuspension of the cells in low K+ two times during a 6 min interstimulus interval before a second 6 min application of 300 MM ATP (second stimulus, 0). A typical Ca, measurement of this sort was shown in Figure 4. With this stimulation paradigm, the response to ATP is almost completely habituated by the second repetitive stimulation; therefore, no attempt was made to correlate Ca, and NE release for greater numbers of re- petitive stimuli.
that the relationship between Ca, and NE release is not altered with repetitive stimulation. Thus, the decrease in ATP-stimu- lated Ca, elevations can fully account for the decrease in NE release. Similar results were obtained by correlating peak stim- ulated Ca, levels with 3H-NE release, confirming that this con- clusion is not an artifact of the integration methods.
The decrease in ATP-stimulated Ca, elevations suggests that either the ATP-gated channel mediating calcium influx is down- regulated or calcium removal is upregulated with repetitive stimulation. Because the relationship between Ca, elevation and NE release is not changed with repetitive stimulation, it is un- likely that other mechanisms, such as inactivation ofa secretory apparatus, are involved. If the activity of calcium-removing mechanisms was increased with repetitive presentation of ATP, NE release in response to unregulated influxes of calcium should also be decreased. However, if only the ATP-gated cation chan- nel were inactivated following stimulation with ATP, the re- sponse to Ca, elevations caused by other means should be un- changed.
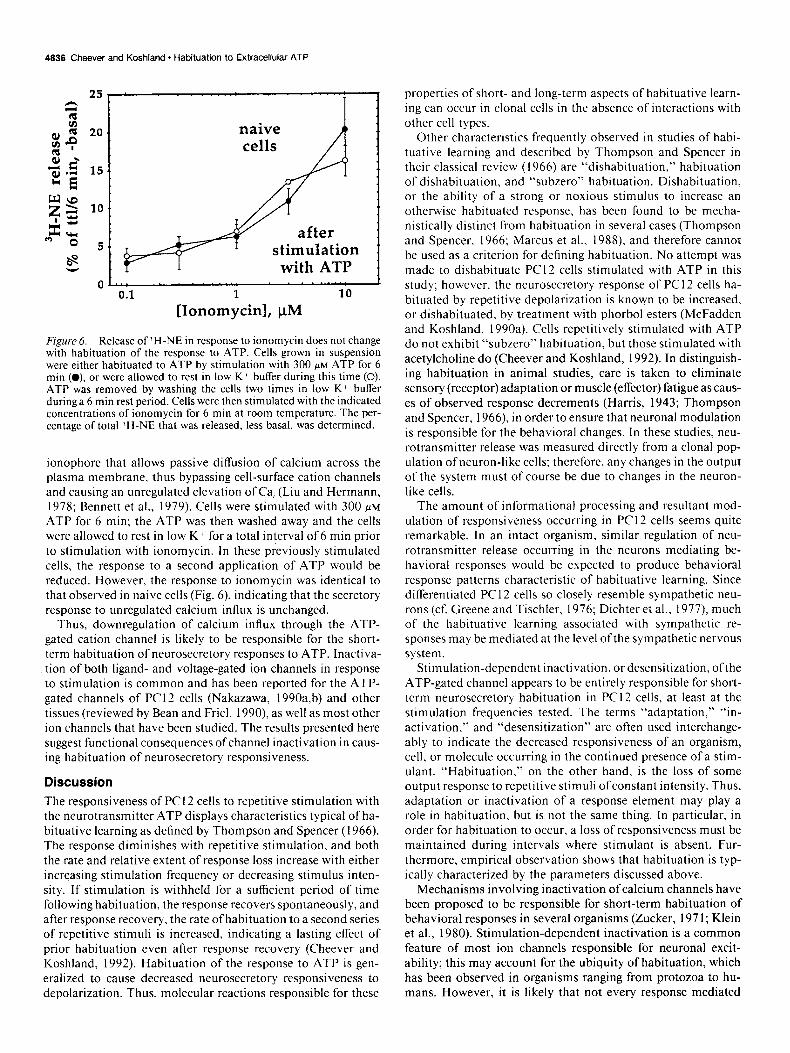
To test these possibilities, ?H-NE release occurring in response to ionomycin was measured both in previously unstimulated cells and in cells that had been treated with ATP, causing ha- bituation ofthe response to ATP (Fig. 6). Ionomycin is a calcium
4836 Cheever and Koshland l HabituaOon to Extracellular ATP
naive
stimulation with ATP
0.1 1 10
[Ionomycinl, p-M
Figure 6. Release of ‘H-NE in response to ionomycin does not change with habituation of the response to ATP. Cells grown in suspension were either habituated to ATP by stimulation with 300 FM ATP for 6 min (0). or were allowed to rest in low K * buffer during this time (0). ATP was removed by washing the cells two times in low K+ buffer during a 6 min rest period. Cells were then stimulated with the indicated concentrations of ionomycin for 6 min at room temperature. The per- centage of total ‘H-NE that was released, less basal, was determined.
ionophore that allows passive diffusion of calcium across the plasma membrane, thus bypassing cell-surface cation channels and causing an unregulated elevation of Ca, (Liu and Hermann, 1978; Bennett et al., 1979). Cells were stimulated with 300 PM
ATP for 6 min; the ATP was then washed away and the cells were allowed to rest in low K’ for a total interval of 6 min prior to stimulation with ionomycin. In these previously stimulated cells, the response to a second application of ATP would be reduced. However, the response to ionomycin was identical to that observed in naive cells (Fig. 6), indicating that the secretory response to unregulated calcium influx is unchanged.
Thus, downregulation of calcium influx through the ATP- gated cation channel is likely to be responsible for the short- term habituation of neurosecretory responses to ATP. Inactiva- tion of both ligand- and voltage-gated ion channels in response to stimulation is common and has been reported for the ATP- gated channels of PC12 cells (Nakazawa, 1990a,b) and other tissues (reviewed by Bean and Friel, 1990), as well as most other ion channels that have been studied. The results presented here suggest functional consequences of channel inactivation in caus- ing habituation of neurosecretory responsiveness.
Discussion
The responsiveness of PCI 2 cells to repetitive stimulation with the neurotransmitter ATP displays characteristics typical of ha- bituative learning as defined by Thompson and Spencer (1966). The response diminishes with repetitive stimulation, and both the rate and relative extent of response loss increase with either increasihg stimulation frequency or decreasing stimulus inten- sity. If stimulation is withheld for a sufficient period of time following habituation, the response recovers spontaneously, and after response recovery, the rate of habituation to a second series of repetitive stimuli is increased, indicating a lasting effect of prior habituation even after response recovery (Cheever and Koshland, 1992). Habituation of the response to ATP is gen- eralized to cause decreased neurosecretory responsiveness to depolarization. Thus, molecular reactions responsible for these
properties of short- and long-term aspects of habituative learn- ing can occur in clonal cells in the absence of interactions with other cell types.
Other characteristics frequently observed in studies of habi- tuative learning and described by Thompson and Spencer in their classical review (I 966) are “dishabituation,” habituation of dishabituation, and “subzero” habituation. Dishabituation, or the ability of a strong or noxious stimulus to increase an
otherwise habituated response, has been found to be mecha- nistically distinct from habituation in several cases (Thompson and Spencer, 1966; Marcus et al., l988), and therefore cannot be used as a criterion for defining habituation. No attempt was made to dishabituate PC12 cells stimulated with ATP in this study; however, the neurosecretory response of PC I2 cells ha- bituated by repetitive depolarization is known to be increased, or dishabituated, by treatment with phorbol esters (McFadden and Koshland, 1990a). Cells repetitively stimulated with ATP do not exhibit “subzero” habituation, but those stimulated with acetylcholine do (Cheever and Koshland, 1992). In distinguish- ing habituation in animal studies, care is taken to eliminate sensory (receptor) adaptation or muscle (effector) fatigue as caus- es of observed response decrements (Harris, 1943; Thompson and Spencer, 1966), in order to ensure that neuronal modulation is responsible for the behavioral changes. In these studies, neu- rotransmitter release was measured directly from a clonal pop- ulation of neuron-like cells: therefore, any changes in the output of the system must of course be due to changes in the neuron- like cells.
The amount of informational processing and resultant mod- ulation of responsiveness occurring in PCI 2 cells seems quite remarkable. In an intact organism, similar regulation of neu- rotransmitter release occurring in the neurons mediating be- havioral responses would be expected to produce behavioral response patterns characteristic of habituative learning. Since differentiated PC1 2 cells so closely resemble sympathetic neu- rons (cf. Greene and Tischler, 1976; Dichter et al., 1977), much of the habituative learning associated with sympathetic re- sponses may be mediated at the level ofthe sympathetic nervous system.
Stimulation-dependent inactivation, or desensitization, ofthe ATP-gated channel appears to be entirely responsible for short- term neurosecretory habituation in PC I2 cells, at least at the stimulation frequencies tested. The terms “adaptation,” “in- activation,” and “desensitization” are often used interchange- ably to indicate the decreased responsiveness of an organism, cell, or molecule occurring in the continued presence of a stim- ulant. “Habituation,” on the other hand, is the loss of some output response to repetitive stimuli ofconstant intensity. Thus, adaptation or inactivation of a response element may play a role in habituation, but is not the same thing. In particular, in order for habituation to occur, a loss of responsiveness must be maintained during intervals where stimulant is absent. Fur- thermore, empirical observation shows that habituation is typ- ically characterized by the parameters discussed above.
Mechanisms involving inactivation of calcium channels have been proposed to be responsible for short-term habituation of behavioral responses in several organisms (Zucker, I97 I; Klein et al., 1980). Stimulation-dependent inactivation is a common feature of most ion channels responsible for neuronal excit- ability; this may account for the ubiquity of habituation, which has been observed in organisms ranging from protozoa to hu- mans. However, it is likely that not every response mediated

The Journal of Neuroscience, August 1994, 14(B) 4837
by ion channels that inactivate during stimulation will produce habituation. Therefore, it is useful to consider the properties of the ATP-gated channel that may allow it to produce response patterns associated with habituation.
One feature of the ATP-gated channel that allows it to cause
short-term habituation is its slow recovery from stimulation- induced inactivation. Following inactivation with a brief pulse of ATP. the ATP-gated channels of PC12 cells (Nakazawa et al., 1990b) and other tissues (Bean and Friel, 1990) require about 5 min to regain the ability to respond to stimulation. The data shown here indicate that recovery ofthe neurosecretoryresponse is even slower following 1 min applications of ATP. This pro- longed inactivation is expected to cause habituation when re- petitive stimuli are presented at frequencies in the minute-’ time range. Channels that recover more rapidly would not be expected to give rise to habituation at these stimulation fre- quencies.
A second feature typical of habituation is a decrease in the rate and extent of habituation with decreasing stimulation fre- quencies. Any mechanism involving reversible, stimulation-de- pendent inactivation of the response machinery could theoret- ically account for this property (Cheever and Koshland, 1992). Therefore, it is not surprising to observe this phenomenon in PC1 2 cells in which reversible, stimulation-dependent inacti- vation of the ATP-gated channel causes reduction of the neu- rosecretory response to ATP.
Generalization of the loss of response may also be easily ex- plained by this mechanism. If stimulation with ATP produces cellular conditions that cause inactivation of voltage-gated cal- cium channels (VGCCs), which mediate NE release in response to depolarization, then repetitive stimulation with ATP would be expected to cause inactivation of the response to depolar- ization, as was observed here. Most likely, inactivation of the L-type VGCCs that are present on PC12 cells requires depo- larization of the cell membrane (Fox, 1981; Lux and Brown, 1984) and/or elevation of Ca, (cf. Greenberg et al., 1985; Arm- strong, 1989); stimulation with ATP causes both of these con- ditions (Inoue, 1989). In contrast, if depolarization does not provide the cellular conditions that cause inactivation of the ATP-gated channel, then habituation of depolarization-evoked NE release would not be generalized to ATP. Since previous depolarization does not cause inhibition ofthe response to ATP, the membrane depolarization and Ca, elevation induced by treatment with potassium must not be sufficient to cause in- activation of the ATP-gated channel; possibly ATP must si- multaneously be bound to the channel or the channel must be in the open state for inactivation to occur.
It is somewhat surprising that the rate and extent of habitu- ation, relative to the initial response, are increased with de- creasing stimulus intensity in PC12 cells. In cases of ligand- induced inactivation of receptor-operated channels or other re- ceptors, the relative rate of inactivation is typically slower with lower ligand concentrations. With repetitive stimulation, this would be expected to cause a decreased rate of habituation. However, the ATP-gated channel of PC 12 cells may actually inactivate more rapidly in response to lower ligand concentra- tions, as suggested by the time course of changes in Ca, in re- sponse to varying concentrations of ATP (Fig. 3A). Following application of ATP, Ca, rises to a peak level and then decays following an exponential time course; the rate constant for Ca, decay is increased with lower ATP concentrations (Table I), suggesting a faster rate of channel inactivation. This may rep-
resent an unusual property of ATP-gated channels that results in an increased rate of habituation with decreased stimulus in- tensity. Interestingly. the rate of Ca, decay is also somewhat increased upon repetitive application of a constant concentra- tion of ATP. For 6 min presentations of 300 PM ATP separated by a 6 min interstimulus interval, the mean increase in the Ca, decay rate constant is 0. I9 t 0.1 1 min I, which is statistically significant with p = 0.02 (paired t test, II = 5 separate experi- ments). This increased decay rate probably does not contribute strongly to the decreased NE release with repetitive stimulation, since changes in the peak level of Ca, evoked by ATP correlate well with the changes in neurosecretion.
Thus, there seem to be several features of ATP-gated channel regulation that allow it to produce modulation ofresponsiveness typical of short-term habituative learning. It will be interesting to see whether other neural systems that utilize ATP as a neu- rotransmitter display similar habituation with repetitive stim- ulation. Further studies will be required to determine whether modulation of this channel also accounts for the long-term ha- bituation of ATP-induced responses that have been observed in PC1 2 cells (Cheever and Koshland, 1992), or whether other reactions are involved.
References Armstrong DL (I 989) Calcium channel regulation by calcineurin, a
Ca’+-activated phosphatase in mammalian brain. Trends Neurosci 12:l 17-122.
Bean BP, Friel DD (1990) ATP-activated channels in excitable cells. Ion Channels 2: 169-203.
Bennett JP, Cockroft S, Gomperts BD (1979) Ionomycin stimulates mast cell histamine secretion by forming a lipid-soluble calcium com- nlex. Nature 282:851-853.
Betridge MJ, Irvine RF (I 984) Inositol triphosphate, a novel second messenger in cellular signal transduction. Nature 3 I2:3 IS-32 I.
Boyer JL,-Downes CP, Harden TK (1989) Kinetics of activation of phospholipase C by P,, purinergic receptor agonists and guanine nu- cleotides. J Biol Chem 264:894-890.
Castellucci VF, Kandel ER (I 974) A quanta1 analysis of the svnaptic depression underlying habituation of the gill-withdrawal reflex in Aplvsia. Proc Nat1 Acad Sci USA 71:5004-5008.
Castellucci V, Pinsker H, Kupfermann I, Kandel ER (I 970) Neuronal mechanisms of habituation and dishabituation of the gill-withdrawal reflex in Ap/wia. Science 167: 1745-l 748.
Cheever L, Koshland DE Jr (1992) Retention of habituation in PCI 2 cells. Proc Natl Acad Sci USA 88: 10084-10088.
Corfas G, Dudai Y (I 989) Habituation and dishabituation of a clean- ing reflex in normal and mutant Dmsophilu. J Neurosci 9:56-62.
Dichter MA, Tischler AS, Greene LA (1977) Nerve growth factor- induced increase in electrical excitability and acetylcholine sensitivity of a rat pheochromocytoma cell line. Nature 268:501-504.
Dubyak GR, Cowen DS, Meuller LM (1988) Activation of inositol phospholipid breakdown in HL60 cells by PZ-purinergic receptors for extracellular ATP: evidence for mediation bv both oertussis toxin- sensitive and pertussis toxin-insensitive mechanisms. J Biol Chem 263:18108-18117.
Edwards FA. Gibb AJ. Colquhoun D (I 992) ATP receptor-mediated synaptic currents in the central nervous system. Nature 359:144-147.
Fare1 PB, Glanzman DL, Thompson RF (1973) Habituation of a monosynaptic response in vertebrate central nervous system: lateral column-motoneuron pathway is isolated from spinal cord. J Neu- rophysiol 36: I I 17-l 130.
Fasolato C, Pizzo P, Pozzan T (I 990) Receptor-mediated calcium influx in PC12 cells. J Biol Chem 265:20351-20355.
Fox AP (1981) Voltage-dependent inactivation of a calcium channel. Proc Natl Acad Sci USA 78:953-956.
Fyffe REW, Perl ER (1984) Is ATP a central synaptic mediator for certain primary afferent fibers from mammalian skin? Proc Natl Acad Sci USA 81:6890-6893.
Gonzalez FA, Rozengurt E, Heppel LA (I 989) Extracellular ATP in- duces the release of calcium from intracellular stores without the

4838 Cheever and Koshland * Habituation to Extracellular ATP
activation of protein kinase C in Swiss 3T6 mouse tibroblasts. Proc Nat1 Acad Sci USA 86:4530-4534.
Gordon JL (1986) Extracellular ATP: effects, sources, and fates. Bio- them J 233:309-319.
Greenberg DA, Carpenter CL, Messing RO (1985) Inactivation of 45Ca++ uptake by prior depolarization of PC12 cells. Neurosci Lett 62:377-381.
Greene LA, Rein G (1977a) Release, storage and uptake of catecho- lamines by a clonal cell line of nerve growth factor (NGF) responsive pheochromocytoma cells. Brain Res 129:247-263.
Greene LA, Rein G (1977b) Release of [ZH]norepinephrine from a clonal line ofpheochromocytoma cells (PC1 2) by nicotinic cholinergic stimulation. Brain Res 13852 l-528.
Greene LA, Tischler AS (1976) Establishment of a noradrenergic clon- al line of rat adrenal pheochromocytoma cells which respond to nerve growth factor. Proc Nat1 Acad Sci USA 73:2424-2428.
Grynkiewicz G, Poenie M, Tsien RY (1985) A new generation of Ca’+ indicators with greatly improved fluorescence properties. J Biol Chem 260:3440-3450.
Harris JD (1943) Habituatory response decrement in the intact or- ganism. Psychoi Bull 40:385222.-
Holton FA. Holton P (1954) The caoillarv dilator substances in drv powders of spinal roots: a possible role ofadenosine triphosphate in chemical transmission from nerve endings. J Physiol (Lond) 126: 124- 140.
Inoue K, Nakazawa K, Fujimori K, Takanaka A (1989) Extracellular adenosine 5’-triphosphate-evoked norepinephrine secretion not re- lating to voltage-gated Ca’+ channels in pheochromocytoma PC12 cells. Neurosci Lett 106:294-299.
Jahr CE, Jesse1 TM (1983) ATP excites a subpopulation of rat dorsal horn neurones. Nature 304:730-733.
Klein M, Shapiro E, Kandel ER (1980) Synaptic plasticity and the modulation of the Ca’+ current. J Exp Biol 89: 117-I 57.
Kupfermann I, Castellucci V, Pinsker H, Kandel E (1970) Neuronal correlates of habituation and dishabituation of the gill-withdrawal reflex in Apl.vsia. Science 167: 1743-I 745.
Liu C, Hermann TE (1978) Characterization of ionomycin as a calcium ionophore. J Biol Chem 253:5892-5894.
Lux HD, Brown AM (1984) Single channel studies on inactivation of calcium currents. Science 225:432-434.
Marcus EA, Nolen TG, Rankin CH, Carew TJ (1988) Behavioral dissociation of dishabituation, sensitization, and inhibition in Aply- siu. Science 24 I:2 IO-2 13.
McFadden PN, Koshland DE Jr (I 990a) Habituation in the single cell:
diminished secretion ofnorepinephrine with repetitive depolarization of PC12 cells. Proc Nat1 Acad Sci USA 87:2031-2035.
McFadden PN, Koshland DE Jr (1990b) Parallel pathways for habit- uation in repetitively stimulated PC 12 cells. Neuron 4:6 15-62 1.
Nakazawa K, Inoue K, Fujimori K, Takanaka A (1990a) ATP-acti- vated single-channel currents recorded from cell-free patches of pheo- chromocytoma PC12 cells. Neurosci Lett 119:5-8.
Nakazawa K, Fujimori K, Takanaka A, Inoue K (1990b) An ATP- activated conductance in pheochromocytoma cells and its suppres- sion by extracellular calcium. J Physiol (Land) 428:257-272.
Nishizuka Y (I 992) Intracellular sianalina bv hvdrolvsis of ohospho- lipids and activation of protein kiiase CT Science 258:607-614:
Peeke HVS, Petrinovich L (1984) Approaches, constructs, and ter- minology for the study of response change in the intact organism. In: Habituation, sensitization, and behavior (Peeke HVS, Petrinovich L, eds). New York: Academic.
Pinsker H, Kupfermann I, Castelhtcci V, Kandel E (1970) Habituation and dishabituation of the gill-withdrawal reflex in Aplysia. Science 167:1740-1742.
Sasakawa N, Nakaki T, Yamamoto S, Kato R (1989) Stimulation by ATP of inositol triphosphate accumulation and calcium mobilization in cultured adrenal chromaffin cells. J Neurochem 52:441-447.
Siggins GR, Groul DL, Padjen AL, Forman DS (1977) Purine and pyrimidine mononucleotides depolarize neurones of explanted am- phibian sympathetic ganglia. Nature 270:263-265.
Thompson RF, Glanzman DL (1976) Neural and behavioral mech- anisms ofhabituation and sensitization. In: Habituation: perspectives from child development, animal behavior, and neurophysiology (Tighe TJ, Leaton RN, eds). New York: Erlbaum.
Thompson RF, Spencer WA (1966) Habituation: a model phenom- enon for the study of neuronal substrates of behavior. Psycho1 Rev 73: 16-43.
Williams M, Cusack NJ (1990) Neuromodulatory roles of purine nu- cleosides and nucleotides: their receptors and ligands. Neurotrans- missions 6: l-6.
Wood DC (1988) Habituation in Sfentor: a response-dependent pro- cess. J Neurosci 8:2248-2253.
Yatani A, Tsuda Y, Akaike N, Brown AM (1982) Nanomolar con- centrations of extracellular ATP activate membrane Ca+ + channels in snail neurons. Nature 296: 169-l 7 I.
Zucker RS (197 1) Crayfish escape behavior and central synapses. II. Physiological mechanisms underlying behavioral habituation. J Neu- rophysiol 35:62 l-637.




















