
Carbon nanomaterials as antibacterial and antiviral alternatives
Upload
khangminh22Category
view
0download
0
ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
A thesis submitted in fulfilment of the requirements for the degree
Of
Doctor of Philosophy
Chau Minh Khanh
B.Sc., Ho Chi Minh University of Sciences/Vietnam National University
M.Sc., Ho Chi Minh University of Sciences/Vietnam National University
School of Science
College of Science, Engineering and Health
RMIT University
March 2020

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
ii
DECLARATION
I certify that except where due acknowledgement has been made, the work is that of the author
alone; the work has not been submitted previously, in whole or in part, to qualify for any other
academic award; the content of the thesis is the result of work which has been carried out since
the official commencement date of the approved research program; any editorial work, paid or
unpaid, carried out by a third party is acknowledged; and, ethics procedures and guidelines
have been followed.
I acknowledge the support I have received for my research through the provision of an
Australian Government Research Training Program Scholarship.
Signed: ……………………….
Khanh Chau
Date: 27 March 2020

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
iii
ACKNOWLEDGEMENTS
I landed in Australia in November 2016 for PhD study and with much curiosity. Since then I
have experienced lots of happiness, also stress, during three years of study. I am so lucky to
have much help from many people that I met in Australia and Vietnam. So, I would like to
sincerely thank them all.
Firstly, I would like to send my deepest gratitude to my supervisor at RMIT, Prof. Robert. J.
Moore for his constant guidance and encouragement. I have always told my friends how lucky
I am to have a chance to be his PhD student. He is not only a knowledgeable supervisor, taking
me into the genetic world of antimicrobial peptides; but also, very patient to improve my
writing skill, because English is my second language. He encouraged and funded me to travel
to Europe to open my horizon on academic research in different continents. I started my
research work in Asia, inhaled the academic education in Australia, and experienced the
research environment during a visit to many research institutions in Europe. It has given me an
overview of academia worldwide. Also, he is very caring to his students with the lots of
celebrating, lab-lunches, and bowling entertainment. Of course, all these events always end
with our smiles.
My second grateful thanks go out to my co-supervisor in Australia, Dr. Hao Van. I felt her
kindness from the first day that I landed in Australia when she took me around the Bundoora
campus to complete administrative works for a newly arrived PhD student. Working with her
in the same lab, she also gave me considerable help and advice, particularly knowledge about
whole-bacterial genome sequencing and data analysis. She always cheered me up with many
invitations to her family for parties and dinners. And I really like her “Bun Bo”- the traditional
food speciality in her hometown and appreciated all the moments to feel at home with her
family members.
I also would like to express my heartfelt gratitude to my co-supervisor in Vietnam, Associate
Prof. Van Quyen Dong. I owe him a special debt of gratitude for the opportunity of the PhD
scholarship that he introduced to me. And he offered me valuable suggestions and advice in
my academic studies. Without Rob’s, Hao’s and Quyen’s consistent instruction, this thesis
could not have reached its present form.
My grateful thanks go to the Moore lab members, Bronwyn Campbell, Ben Vezina, Mian Cheer
Gor, Brydon Davidson, Canh Phung, and Chithralekha Murilidharan. I really enjoyed working

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
iv
in the same lab with them, particularly Bronwyn who gave me lots of technical help in the lab.
You were all always very kind and I appreciated all the happy times with you all.
Also, I would like to thank all my Vietnamese friends who I met at RMIT University: Huong
Nguyen, Loc Nguyen, Canh Phung, and Pop. We were working, eating, playing and sharing
during my PhD life. They were always willing to help me with smiles and we travelled together
to explore different cities in Australia.
I am greatly indebted to RMIT University for offering me my PhD scholarship via the RMIT-
VAST cooperative agreement.
I would like to send my great thanks to the Institute of Biotechnology (Vietnam Academy of
Science and Technology), where they set up the scholarship with RMIT University and
prepared a pile of administration works for my scholarship.
I also would like to thank my workplace, “NhaTrang Institute of Technology Research and
Application” (Vietnam Academy of Science and Technology) for supporting me during my
study abroad.
Last but not the least, my acknowledgement also extends to my parents who have given
unconditional assistance; supporting and caring for me all my life. I always love you.
All my best wishes to you.
Khanh Chau

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
v
TABLE OF CONTENT
Chapter 1 INTRODUCTION ..................................................................................................... 3
1.1. The rapid development of antibiotic-resistant bacterial pathogens and reduction in
antibiotic discoveries ................................................................................................................. 3
1.2. The marine environment: an alternative source for antibiotic discoveries ..................... 5
1.3. Marine Bacillus – promising hosts of novel antimicrobial peptides ............................... 6
1.4. Non-ribosomally synthesized peptides (NRPs)- the antimicrobial peptides exhibiting
both antifungal and bactericidal activity. ................................................................................... 6
1.5. The bacteriocins, a promising group of antimicrobial proteins ...................................... 8
1.5.1. Bacteriocins – definition .......................................................................................... 8
1.5.2. Bacteriocin – classification, biosynthesis ................................................................ 9
1.5.2.1. Class I bacteriocins......................................................................................... 10
1.5.2.2. Class II: unmodified bacteriocins ................................................................... 16
1.5.2.3 Class III: large antimicrobial proteins ............................................................ 16
1.5.3. Mode of action and bacteriocin resistance rate surveillance among bacterial
pathogens. ............................................................................................................................. 17
1.6. Current knowledge of marine-derived bacteriocins ...................................................... 18
1.7. Research questions and thesis objectives ...................................................................... 21
1.7.1. Research questions ................................................................................................ 21
1.7.2. Research aims and significance. ............................................................................ 21
1.7.3. Novel contributions ............................................................................................... 22
1.8. Thesis organization ....................................................................................................... 22
Chapter 2 RESEARCH METHODOLOGY ....................................................................... 25
2.1 Sampling and sample preparation for bacterial isolation .............................................. 25
2.2. Isolation of spore-forming bacteria from marine samples ............................................ 25
2.3. Assays to detect the antimicrobial activity. .................................................................. 26
2.3.1. Cross-streak assay for primary screening of marine isolates exhibiting antimicrobial
activity 26

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
vi
2.3.2. Well-diffusion assay .............................................................................................. 27
2.3.3. Spot-on-lawn assay ................................................................................................ 29
2.4. Bacterial identification based on 16S rRNA gene sequence and phylogenetic tree
construction .............................................................................................................................. 30
2.4.1. Extraction of genomic DNA by bead beating ....................................................... 30
2.4.2. Amplification of bacterial 16S rRNA gene sequences, Sanger sequencing and
phylogenetic identification ................................................................................................... 30
2.5. The sensitivity of antimicrobial activities to enzyme and heat. .................................... 30
2.6. Analysis of self-depression ability across marine Bacillus/Paenibacillus species ....... 31
2.7. Growth properties, antibiotic susceptibility testing, and enzyme production of isolates
31
2.8. Bacterial whole genome sequencing ............................................................................. 32
2.8.1. Extraction of genomic DNA (gDNA) for whole genome sequencing .................. 32
2.8.2. Qualification of the gDNA .................................................................................... 32
2.8.3. Preparation of DNA library and whole genome sequencing ................................. 33
2.9. Assembly of raw reads generated from whole genome sequencing ............................. 33
2.10. Calculation of Average Nucleotide Identities (ANI) across genomes of terrestrial and
marine Bacillus/Paenibacillus species ..................................................................................... 33
2.11. Estimation of a frequency distribution of CDS across genomes ............................... 34
2.12. Comparison between genomes of marine isolates and genomes of phylogenetically
related terrestrial strains. .......................................................................................................... 35
2.13. In silico prediction of putative antimicrobial peptides within genomes of marine
Bacillus species ........................................................................................................................ 37
2.14. Refinement of media and growth conditions for enhanced bacteriocin production . 39
2.15. Recovery of antimicrobial compounds after fermentation ........................................ 39
2.15.1. Recovery of antimicrobial peptides from the cell-free culture supernatant by
precipitation using ammonium sulphate .............................................................................. 39
2.15.2. Recovery of antimicrobial compounds from cell-free culture supernatants using
Diaion HP-20........................................................................................................................ 40

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
vii
2.15.3. Recovery of cell associated antimicrobial compounds from cell pellets by acidic
solvent extraction ................................................................................................................. 40
2.16. Ion exchange chromatography for partial purification of antimicrobial compounds 41
2.16.1. Identification of running buffer and types of resin for ion exchange
chromatography purification of antimicrobial compounds .................................................. 41
2.16.2. Determination of the NaCl concentration used in elution buffer ....................... 42
2.17. Hydrophobic interaction chromatography for partial purification of antimicrobial
compounds ............................................................................................................................... 42
2.18. Reverse phase high-pressure liquid chromatography (RP-HPLC) to obtain pure
antimicrobial compounds ......................................................................................................... 43
2.19. Determination of antimicrobial purity by Matrix-Assisted Laser
Desorption/Ionization time-of-flight mass spectrometry (MALDI-TOF MS) ........................ 43
2.20. Peptide sequencing techniques .................................................................................. 45
2.20.1. Peptide sequencing by de novo peptide sequencing .......................................... 45
2.20.2. Peptide sequencing by N-terminal sequencing (Edman degradation method) .. 45
2.21. Estimation of bacteriocin size by Tricine SDS-PAGE.............................................. 45
2.22. Zymogram assay ....................................................................................................... 47
2.23. Characterisation of the physicochemical properties of antimicrobial compounds ... 47
2.24. Overview of heterologous cloning and expression of bacteriocin ............................ 47
2.25. Extraction of the plasmids from E. coli host ............................................................. 48
2.26. Preparation of the plasmid backbone by enzymatic digestion .................................. 49
2.27. Preparation of the whole sactipeptide genome cluster by PCR amplification. ......... 49
2.28. Preparation of E. coli competent cell. ....................................................................... 49
2.29. Transformation of the fusion plasmid into E. coli by electroporation ...................... 49
2.30. Preparation of Bacillus competent cells .................................................................... 50
2.31. Transformation of the fusion plasmid into Bacillus subtilis BS34A ........................ 50
Chapter 3 BROAD SPECTRUM ANTIMICROBIAL ACTIVITIES OF SPORE-
FORMING BACTERIA FROM THE VIETNAM SEA .................................................... 51

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
viii
3.1. Introduction ................................................................................................................... 51
3.2. Results ........................................................................................................................... 52
3.2.1. Thermally resistant, spore-forming, bacteria derived from the marine environment
display antimicrobial activity against important pathogens ................................................. 52
3.2.2. Taxonomic analysis of antimicrobial isolates ....................................................... 57
3.2.3. Proteolytic digestion of antimicrobial culture revealed a dominance of
proteinaceous compounds .................................................................................................... 59
3.2.4. Determination of the growth depression within the short-listed group; showing the
production of potentially novel antimicrobial activities ...................................................... 60
3.2.5. Growth characteristics of antimicrobial-producing isolates .................................. 60
3.3. Summary of results and discussion ............................................................................... 63
Chapter 4 BIOINFORMATIC IDENTIFICATION OF PUTATIVE GENE CLUSTERS
ENCODING ANTIMICROBIAL PEPTIDE PRODUCTION .......................................... 66
4.1. Introduction ................................................................................................................... 66
4.2. Results ........................................................................................................................... 67
4.2.1. Overview of six draft genomes .............................................................................. 67
4.2.2. Calculation of ANI values across the genomes revealed a high degree of similarity
between marine species and terrestrial neighbour strains .................................................... 67
4.2.3. Analysis of the distribution frequency of genes across the species revealed a high
number of strain-specific genes in P. polymyxa genomes ................................................... 70
4.2.4. The pairwise comparison across genomes of B. halotolerans revealed the high
similarity in genome organisation and CDS. ....................................................................... 71
4.2.5. The pairwise comparison across genomes of B. amyloliquefaciens revealed high
similarity in genome organisation and CDS. ....................................................................... 72
4.2.6. The pairwise comparison across genomes of P. polymyxa revealed diversity in
genome organisation and CDS. ............................................................................................ 74
4.2.7. In silico prediction of antimicrobial compounds revealed high numbers of NRPs
and bacteriocins, including novel bacteriocins .................................................................... 76

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
ix
4.2.8. Prediction of gene organisation of biosynthetic gene clusters of characterised
bacteriocins........................................................................................................................... 83
4.2.8.1. The LCI gene clusters (found in the genomes of isolates # 06, #08, #11, #13)
83
4.2.8.2. The amylocyclicin gene clusters (found in the genomes of isolates #06, #08,
#11, and #13) .................................................................................................................... 84
4.2.8.3. The mersacidin gene cluster (found in the genome of isolate #08) ............... 85
4.2.8.4. The subtilosin A gene cluster (found in the genome of isolate # 01)............. 87
4.2.8.5. The plantazolicin gene cluster (found in the genome of isolate # 06) ........... 89
4.2.8.6. The paenicidin A gene cluster (found in the genome of isolate #23) ............ 90
4.2.9. Prediction of gene organisation of biosynthetic gene clusters of novel bacteriocins
92
4.2.9.1. The novel lantibiotic gene cluster (found in the genome of isolate #08) ....... 92
4.2.9.2. The novel lantibiotic gene cluster (found in the genome of isolate #23) ....... 94
4.2.9.3. The sactipeptide gene cluster (found in the genome of isolate # 23) ............. 96
4.2.9.4. The lassopeptide gene cluster (found in the genome of isolate #23) ............. 97
4.2.9.5. The thiopeptide gene clusters (found in both isolate #06 and isolate #13) .... 99
4.2.9.6. The thiopeptide gene cluster (found in the genome of isolate #11) ............. 102
4.2.9.7. The two-component lantibiotic gene cluster (found in the genome of isolate
#13) 103
4.3. Summary of results and discussion ............................................................................. 106
Chapter 5 PURIFICATION OF ANTIMICROBIAL PEPTIDES PRODUCED BY
BACILLUS AMYLOLIQUEFACIENS #11 ........................................................................ 111
5.1. Introduction ................................................................................................................. 111
5.2. Result .......................................................................................................................... 113
5.2.1. Analysis of the bacterial growth curve revealed to production of various
antimicrobial compounds ................................................................................................... 113
5.2.2. Purification of antimicrobial compounds from 12 hour-old culture elucidated the
presence of amylocyclicin bacteriocin ............................................................................... 114

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
x
5.2.2.1. Recovery of bacteriocins from cell-free culture and cell pellet. .................. 114
5.2.2.2. Purification of bacteriocins by ion exchange chromatography revealed the
presence of 1 antimicrobial peptide ................................................................................ 116
5.2.2.3. Purification of bacteriocin performed by analytical RP-HPLC elucidated the
presence of amylocyclicin .............................................................................................. 118
5.2.2.4. The amylocyclicin, a heat and pH stable bacteriocin exhibited activity against
only Gram-positive bacteria ........................................................................................... 121
5.2.3. Purification of antimicrobial substances from 36 hours-old culture showed the
presence of three antimicrobial compounds ....................................................................... 123
5.2.3.1. Recovery of antimicrobials compounds from the cell pellet........................ 123
5.2.3.2. RP-HPLC purification of antimicrobial compounds and peptide identification
125
5.3. Summary of results and discussion. ............................................................................ 126
Chapter 6 ANALYSIS OF ANTIMICROBIAL PEPTIDES PRODUCED BY
PAENIBACILLUS POLYMYXA #23 .................................................................................. 130
6.2. Result .......................................................................................................................... 131
6.2.1. Time course analysis of antimicrobial production during bacterial growth revealed
production of multiple antimicrobial substances. .............................................................. 131
6.2.2. Purification of antimicrobial compounds from 24 hour old PB culture revealed the
presence of three antimicrobial compound/s ...................................................................... 133
6.2.2.1. Recovery of antimicrobial compounds from cell-free culture and the cell pellet
133
6.2.2.2. Purification of the antimicrobial peptides in CFS fraction with hydrophobic
exchange chromatography elucidated the presence of two antimicrobial compounds. . 135
6.2.2.3. Purification of 30ACN fraction revealed the presence of polymyxin .......... 136
6.2.2.4. Purification of 40ACN fraction revealed the presence of paenicidin A, tridecaptin
139
6.2.4.1. The paenicidin A, a lantibiotic, displayed antimicrobial activity against Gram-
positive bacteria .............................................................................................................. 140

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xi
6.3. Summary of results and discussion ............................................................................. 141
Chapter 7 CLONING AND HETEROLOGOUS EXPRESSION OF SACTIPEPTIDE, A
NOVEL BACTERIOCIN FROM MARINE P. POLYMYXA #23, IN BACILLUS
SUBTILIS.............................................................................................................................. 145
7.1. Introduction. ................................................................................................................ 145
7.2. Methods and results .................................................................................................... 147
7.2.1. Successful isolation of intact sactipeptide whole gene cluster, and generation of the
expression plasmid (pdLL202/G23sac) ............................................................................. 147
7.2.2. Successful transformation of fusion plasmid into E. coli Top10 ........................ 150
7.2.3. Successful transformation of the fusion-plasmid into E. coli JIR702 for production
of multimeric plasmid. ....................................................................................................... 150
7.2.4. Successful expression of sactipeptide gene cluster in B. subtilis and optimization of
sactipeptide production ...................................................................................................... 152
7.2.5. Two-steps procedure to purify the sactipeptide from 24 hour LB culture .......... 153
7.2.6. Characterisation of sactipeptide revealed the proteinaceous nature and thermal
stability of the sactipeptide ................................................................................................. 155
7.2.7. N-terminal sequencing of the sactipeptide .......................................................... 156
7.3. Summary of result and discussion .............................................................................. 156
Chapter 8 CONCLUSION AND FUTURE WORK ......................................................... 160
8.1. Conclusion .................................................................................................................. 160
8.2. Future work ................................................................................................................. 165
APPENDIX ............................................................................................................................ 183

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xii
ABBREVIATIONS
aa Amino acid
ACN Acetonitrile
AMP(s) Antimicrobial peptide(s)
antiSMASH Antibiotics & secondary metabolite analysis shell
BAGEL Bacteriocin genome mining tool
BLIS Bacteriocin like inhibitory substance
CDS Coding DNA sequence
CFS Cell-free supernatant
CRE Carbapenem- resistant Enterobacteriaceae
CV Colum volume
EDTA Ethylenediaminetetraacetic acid
GES Guanidinium thiocyanate
LAP(s) Linear azol(in)e-containing peptides
LB Luria-Bertani
LPMA Lab prepared marine agar
LPMB Lab prepared marine broth
LP(s) Lipopeptide(s)
MALDI-TOF MS Matrix-assisted laser desorption/ionization mass spectrometry
MHA Muller Hilton Agar
MRKP Multidrug-resistant Klebsiella pneumonia
MRSA Methicillin-resistant Staphylococcus aureus
MSA Multiple sequence alignment
MW Molecular weight
NCBI National Centre for Biotechnology Information
NR Non-redundant
NRS(s) Non-ribosomal synthase(s)
NRSP(s) Non-ribosomally synthesized antimicrobial peptide(s)
ORF Open reading frame
PB Production broth
PBGCs Potential bacteriocin gene clusters

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xiii
PCR Polymerase chain reaction
PES Polyethersulfone
PK Polyketide
PKS Polyketide synthase
PTM Post translational modification
RAST Rapid annotation using subsystem technology
RP- HPLC Reverse-phase high-performance liquid chromatography
Sfp 4'-phosphopantetheinyl transferase
SSW Sterile seawater
TEMED Tetramethyl ethylenediamine
TOMM Thiazole/oxazole-modified microcins
TFA Trifluoroacetic acid
TMH Transmembrane helices
TSA Tryptic soy agar
TSB Tryptic soy broth
v/v Volume per volume
w/v Weight per volume
WGS Whole genome sequencing

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xiv
LIST OF FIGURES
Figure 1.1. Timeline of antibiotic resistance compared to antibiotic discovery. ....................... 4
Figure 1.2. Minimal domain structure of (A) nonribosomal peptide synthetase (NRPS) and (B)
polyketide synthase (PKS). ........................................................................................................ 7
Figure 1.3. Schematic representation of bacteriocin biosynthesis. ............................................ 9
Figure 1.4. Schematic representation of lantibiotic biosynthesis. ........................................... 11
Figure 1.5. Schematic representation of “head-to-tail linearized bacteriocin”. ....................... 12
Figure 1.6. Schematic presentative of sactipeptide. ................................................................. 13
Figure 1.7. Schematic presentative of LAP biosynthesis. ....................................................... 13
Figure 1.8. Schematic presentative thiopeptide biosynthesis. ................................................. 14
Figure 1.9. Schematic representation of glycocins biosynthesis. ............................................ 15
Figure 1.10. Schematic representation of lassopeptide biosynthesis. ...................................... 16
Figure 1.11.Bacteriocin’s mode of action. ............................................................................... 18
Figure 2.1.Locations of sampling trips within Nhatrang bay (Vietnam sea). .......................... 25
Figure 2.2. Cross-streak assay. ................................................................................................ 26
Figure 2.3. Well-diffusion assay. ............................................................................................. 28
Figure 2.4. Spot-on-lawn assay. ............................................................................................... 29
Figure 2.5. Schematic representation of pairwise genome comparison performed by the
“Sequence-Based Comparison Tool” (A) and MAUVE (B). .................................................. 36
Figure 2.6. Schematic representation of in silico prediction of antimicrobial peptides performed
by BAGEL and AntiSMASH tools. ......................................................................................... 38
Figure 2.7. The process of MALDI-TOF mass spectrometry. ................................................. 44
Figure 3.1. Diversity in the morphology of the marine isolates isolated from NhaTrang bay,
Vietnam Sea. ............................................................................................................................ 53
Figure 3.2. Phylogenetic tree of the 23 short-listed isolates. ................................................... 58
Figure 4.1 The heat map of the average nucleotide identity and genome-based phylogenetic
tree across Bacillus/Paenibacillus genomes ............................................................................ 69
Figure 4.2. Frequency distribution of CDS across the genomes.............................................. 70
Figure 4.3. Genome comparison between marine B. halotolerans #01 and terrestrial neighbour
B. halotolerans F41-3. ............................................................................................................. 71
Figure 4.4. Genome comparison between marine B. amyloliquefaciens and terrestrial B.
amyloliquefaciens .................................................................................................................... 74

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xv
Figure 4.5. Genome comparison between marine P. polymyxa #23 and terrestrial P. polymyxa
species. ..................................................................................................................................... 75
Figure 4.6. Multiple sequence alignment (A) and average distance tree (B) between the
bacteriocins precursors in this study and known precursors .................................................... 82
Figure 4.7 The LCI gene cluster found in marine B. amyloliquefaciens #06, #08, #11 and #13.
.................................................................................................................................................. 83
Figure 4.8. The amylocyclicin gene cluster found in marine B. amyloliquefaciens #06, #08, #11
and #13. .................................................................................................................................... 85
Figure 4.9. The mersacidin gene cluster found in the genome of isolate #08 ......................... 86
Figure 4.10. The subtilosin A gene cluster found in the genome of isolate #01 ..................... 88
Figure 4.11. The plantazolicin gene cluster found in genome of isolate #06 .......................... 89
Figure 4.12. The paenicidin A gene cluster found in the genome of isolate #23 .................... 91
Figure 4.13. The novel lantibiotic found in the genome of B. amyloliquefaciens # 08. .......... 93
Figure 4.14. The novel lantibiotic gene cluster found in the genome of isolate #23. .............. 95
Figure 4.15. The novel sactipeptide gene cluster found in the genome of P. polymyxa #23 and
the precursor sequence. ............................................................................................................ 96
Figure 4.16. The lassopeptide gene cluster found in the genome of isolate #23 ..................... 98
Figure 4.17. The novel thiopeptide gene cluster found in the genomes of B. amyloliquefaciens
#06 and #13. ........................................................................................................................... 100
Figure 4.18.The novel thiopeptide found in the genome of B. amyloliquefaciens #11 ......... 102
Figure 4.19. The two-component lantibiotic gene cluster in the genome of isolate #13. ...... 104
Figure 5.1. The seaweed from which B. amyloliquefaciens #11 isolated (A). Bacterial colony
morphology of #11 on Muller Hilton agar (B) ...................................................................... 112
Figure 5.2. Antimicrobial activity exhibited by marine isolate #11. ..................................... 113
Figure 5.3. Antimicrobial activity exhibited from cell-free supernatants of the cultures collected
at different time-points. .......................................................................................................... 114
Figure 5.4. Antimicrobial activity against L. plantarum exhibited by the fractions collected at
time-point of 12 hours. ........................................................................................................... 115
Figure 5.5. The experiments to select the resin, type of buffer, and NaCl concentration in
elution buffer for ion exchange chromatography ................................................................... 117
Figure 5.6. The MW of antimicrobial peptide presented in 12 hour old culture after partial
purification by cation exchange chromatography. ................................................................. 118
Figure 5.7. RP-HPLC to purify antimicrobial peptide presented in 12 hour old culture. ...... 119
Figure 5.8. Properties of amylocyclicin. ................................................................................ 121

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xvi
Figure 5.9. (A) Sequence alignment between amylocyclicin from isolate #11 genome
(11amyA) and FZB42 strains. (B) Maturation of amylocyclicin. ......................................... 122
Figure 5.10. Matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF) mass
spectrometry (MS) ................................................................................................................. 124
Figure 5.11. RP-HPLC used to separate the freshly dissolved aggregate. ............................ 125
Figure 6.1. Antimicrobial spectrum exhibited by marine P. polymyxa #23. ......................... 131
Figure 6.2. Antimicrobial activity exhibited by P. polymyxa #23 in culture media. ............. 132
Figure 6.3. Recovery of the antimicrobial compounds from the culture supernatant of P.
polymyxa #23 by absorption antimicrobial substances onto Diaion HP-20. ......................... 133
Figure 6.4. Nature of antimicrobial compounds from different fractions obtained during
purification. ............................................................................................................................ 134
Figure 6.5. Hydrophobic interaction chromatography (using Sep-Pak C18 cartridge) to purify
the antimicrobial “cell surface extract” (CFS). ...................................................................... 135
Figure 6.6. Zymogram of polymyxin against S. Enteritidis ................................................... 137
Figure 6.7 Sequencing result of the purified polymyxin. ..................................................... 138
Figure 6.8. The first round of RP-HPLC to separate the 40% ACN fraction.Two fractions
exhibited antimicrobial activity, with peaks at retention times of 23.443 minutes ([m/z] = 1616)
and 23.797 minutes ([m/z] = 3370)........................................................................................ 139
Figure 6.9. MALDI-TOF MS spectra of 40% ACN fraction under detection range of 800 Da –
2000 Da. ................................................................................................................................. 140
Figure 6.10. MW of purified paenicidin A by MALDI-TOF MS with the mass intensity of
[m/z] = 3370. .......................................................................................................................... 140
Figure 7.1. (A) Gene organisation on plasmid pDLL202; (B) The plasmid contains 4 antibiotic
resistance genes and 4 multi cloning sites (MCS); (C) the gene cluster of sactipeptide ....... 146
Figure 7.2. Construction of fusion plasmid. .......................................................................... 149
Figure 7.3. Gene organisation of fusion plasmid pDLL202/G23sac generated after Gibson
assembly. ................................................................................................................................ 150
Figure7.4.Confirmation by PCR for positive transformants after transformation. ................ 151
Figure 7.5. Antimicrobial activity exhibited by some B. subtilis transformants against MRSA
indicator. ................................................................................................................................ 152
Figure7.6. Reverse-phase (RP) high-performance liquid chromatography (HPLC)
chromatograms to separate the AMS fraction. ...................................................................... 154
Figure 7.7. MALDI-TOF MS of purified sactipeptide. ......................................................... 155
Figure 7.8. Properties of sactipeptide. ................................................................................... 155

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xvii
LIST OF TABLES
Table 1.1. List of marine bacteriocins published. .................................................................... 19
Table 2.1. List of indicators strains used in antimicrobial screening assays ........................... 27
Table 2.2. Electrode and gel buffers for Tricine–SDS-PAGE, ................................................ 46
Table 2.3. The composition of acrylamide mixtures used to cast the Tricine SDSD gel, ....... 46
Table 2.4. List of strains and plasmids used for cloning /expression ...................................... 48
Table 3.1. Antimicrobial producing bacteria identified from marine samples ........................ 52
Table 3.2. Antimicrobial activity of short-listed isolates against indicators strains. ............... 55
Table 3.3. Closest species of producers, identified by BLASTn of 16S rRNA against 16S rRNA
database (NCBI) (18/06/2019) ................................................................................................. 57
Table 3.4. Enzymatic sensitivity profile of antimicrobial activities. These shortlisted 19 isolates
showed antagonistic activities against C. perfringens. ............................................................ 59
Table 3.5. Antimicrobial activities amongst the short-listed isolates. ..................................... 61
Table 3.6. Characterization of short-listed bacterial isolates. .................................................. 62
Table 4.1.Overview of draft genomes of marine isolates ........................................................ 67
Table 4.2 List of CDS found in genome #01 relating to the biosynthesis of polyketide
compound. These CDS had low similarity with those found in the genome of F41-3 ............ 72
Table 4.3.Low identity CDSs in the genome of isolate #11 relating to the biosynthetic gene
cluster for bacteriocin production. ........................................................................................... 73
Table 4.4. The CDSs relating to unique bacteriocin biosynthetic gene clusters found in the
genome #23 .............................................................................................................................. 76
Table 4.5. List of putative antimicrobial peptides predicted within Bacillus/Paenibacillus draft
genomes. .................................................................................................................................. 78
Table 4.6. Amino acid sequences of precursor bacteriocins predicted within 6 Bacillus genomes
.................................................................................................................................................. 79
Table 4.7. Structural organisation of amylocyclicin gene cluster found in isolates #06, #08, #11
and #13 ..................................................................................................................................... 84
Table 4.8. Structural organisation of mersacidin gene cluster found in isolate #08. ............... 87
Table 4.9. Structural organisation of subtilosin A gene cluster found in the genome of isolate
#01............................................................................................................................................ 88
Table 4.10. Structural organisation of plantazolicin gene cluster found in isolate #06 .......... 90
Table 4.11. Structural organisation of paenicidin A gene cluster found in the genome of isolate
#23............................................................................................................................................ 92

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
xviii
Table 4.12. Structural organisation of novel lantibiotic gene cluster found in the genome of
isolate #08 ................................................................................................................................ 93
Table 4.13. Structural organisation of novel lantibiotic gene cluster found in the genome of
isolate #23 ................................................................................................................................ 95
Table 4.14. Structural organisation of novel sactipeptide gene cluster found in isolate #23 .. 97
Table 4.15. Structural organization of novel lassopeptide biosynthetic cluster found in isolate
#23............................................................................................................................................ 99
Table 4.16. Structural organization of the novel thiopeptide biosynthetic cluster in isolate #06
and #13 ................................................................................................................................... 101
Table 4.17. Structural organization of the novel thiopeptide biosynthetic cluster found in isolate
#11.......................................................................................................................................... 102
Table 4.18. Structural organization of the novel two-component lantibiotic in isolate #13 .. 105
Table 5.1. The antimicrobial spectrum of amylocyclicin produced by isolate #11 ............... 122
Table 5.2. Antimicrobial spectrum of purified thiopeptide supposed ................................... 126
Table 6.1. Antimicrobial spectra of fractions recovered from a 36-hours old culture ........... 134
Table 6.2. The antimicrobial spectra of 30ACN fraction, and 40ACN fraction. ................... 136
Table 6.3 The antimicrobial spectrum of polymyxin ............................................................. 139
Table 6.4. Antimicrobial activity of compounds present in the 40ACN fraction .................. 141
Table 7.1. List of primers used for cloning and confirmation ............................................... 148
Table 7.2. The growth phase of B. subtilis transformant in LB medium ............................... 153
Table 7.3. Antimicrobial spectrum of sactipeptide ................................................................ 156

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
1
ABSTRACT
The indiscriminate use of antibiotics in healthcare and agriculture over the past decades has led
to a high incidence of antibiotic-resistant pathogens worldwide. One of the solutions that could
be considered to combat this crisis is the identification of novel antibiotics or other alternatives.
However, the discovery pipeline for novel antimicrobials, identified from organisms in the
terrestrial environment, is sparse. Therefore, we have taken an alternative approach to this issue
by screening marine bacteria to seek novel bacteriocins for further antibiotic development.
Bacteriocins are a family of ribosomally synthesized antimicrobial peptides; some of which are
currently applied as safe food preservatives (nisin) and they are also gaining interest as
promising alternatives to conventional antibiotics. The Vietnam Sea has a variety of marine
ecosystems with diverse marine species, but there has been little exploration of this source for
novel active compounds. Therefore, we undertook a discovery program aimed at identifying
novel bacteriocins from bacterial isolates recovered from the Vietnam Sea.
In stage one, 64 spore-forming bacterial isolates that exhibited antimicrobial activity were
identified after sampling and bacterial isolation (Chapter 3). Inspection of their antimicrobial
spectra resulted in the short-listing of 23 isolates for further analysis. Based on 16S ribosomal
RNA sequences, these 23 isolates were identified as 22 Bacillus species and 1 Paenibacillus
species (Paenibacillus polymyxa), including ubiquitous species (Bacillus subtilis, Bacillus
amyloliquefaciens), and floral species (Bacillus halotolerans, Bacillus safensis, Bacillus
pacificus, Paenibacillus polymyxa, Bacillus licheniformis). They exhibited strong antibacterial
activity against a range of human and veterinary pathogens and food-poisoning bacteria with
dominance of proteinaceous antimicrobial substances. Inspection of the breadth and strength
of antimicrobial activities informed the selection of 6 isolates for whole genome sequencing in
the second stage of the study.
The six isolates subjected to whole genome sequencing were; four B. amyloliquefaciens
isolates (#06, #08, #11, #13), one B. halotolerans (#01) isolate, and one P. polymyxa (#23)
isolate (Chapter 4). Bioinformatic analysis identified genes encoding 61 putative antimicrobial
peptides within the genomes of the six isolates, including 41 mostly characterised non-
ribosomally synthesized peptides (lipopeptides, polyketides and bacilysin) and 20 bacteriocins.
Amongst the 20 putative bacteriocins found, there were 13 different types of bacteriocins. The
putative bacteriocin encoding genes clusters included those for 6 characterised bacteriocins
(mersacidin, paenicidin A, plantazolicin, LCI, amylocyclicin, subtilosin A), and 7

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
2
uncharacterised/ unique bacteriocins (2 thiopeptides, 1 two-component lantibiotic, 1
sactipeptide, 2 lantibiotic, and 1 lassopeptide).
In the third stage of the study (Chapters 5 and 6); two isolates were selected for antimicrobial
purification. Purification of antimicrobial peptides from isolate #11 resulted in the
identification of amylocyclicin ([m/z]= 6,381), iturin/surfactin ([m/z]= 1,058) and an undefined
peptide ([m/z]= 1,473). The undefined peptide ([m/z]= 1,473), was suspected to be a novel
thiopeptide due to the similarity of MW. However, the amino acid sequence of this peptide
couldn’t be elucidated by an N-terminal sequencing method. Antimicrobial purification from
isolate #23 identified polymyxin ([m/z]= 1168.7), tridecaptin ([m/z]= 1550.8), and paenicidin
A ([m/z]= 3290.4), but no novel bacteriocins.
In the fourth stage of the study, an attempt was made to heterologously produce the novel
sactipeptide which was one of three putative novel bacteriocins predicted from the genome of
isolate #23. The technique employed E. coli/B. subtilis shuttle plasmid, pDLL202, for plasmid
construction and a heterologous B. subtilis host for bacteriocin expression. A 15,487 bp fusion
plasmid (pDLL202/G23sac) was constructed, carrying an 8,556 bp–sactipeptide gene cluster
derived from isolate #23 and the 6,938 bp- plasmid backbone derived from plasmid pDL202.
The Bacilllus transformants successfully expressed the sactipeptide. The sactipeptide had a
molecular weight (MW) of 3,404 Da and depressed growth of Gram-positive bacteria.
Taken together, the marine Bacillus/Paenibacillus are abundant reservoirs of genes coding
novel bacteriocins for further discovery. These antimicrobial compounds produced by marine
Bacillus/Paenibacillus can be developed into therapeutic antimicrobial drugs against
antibiotic-resistant pathogens, particularly against antibiotic-resistant Gram-positive bacteria.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
3
Chapter 1 INTRODUCTION
1.1. The rapid development of antibiotic-resistant bacterial pathogens and reduction in
antibiotic discoveries
Antibiotics are natural or synthesized compounds that depress the growth of microorganisms.
The introduction of antibiotics is one of the greatest achievements in pharmacology since
antibiotics now play a vital role in the clinical treatment of many bacterial pathogens.
In 1928, the first antibiotic, Penicillin was discovered from Penicillium notatum by Alexander
Fleming, and in 1934 Penicillin was approved to use then rapidly became an important drug in
clinical treatments, in aquaculture, in veterinary treatments, and agriculture (1). Penicillin
became an important bulwark protecting humankind from numerous pathogenic infections,
decreasing mortality, increasing agricultural profitability, and initiated a wave of antibiotic
discoveries from natural resources [Fig 1.1]. Antibiotics vary greatly in nature, structure, and
mode of action. For example, antibiotics can act as inhibitors of cell wall synthesis
(cephalosporin, carbapenem), DNA synthesis inhibitors (fluoroquinolones), protein synthesis
inhibitors (macrolides, chloramphenicol, rifampin), mycolic acid synthesis inhibitors, and folic
acid synthesis inhibitors (2). Of these, the carbapenem group (meropenem, imipenem) are now
the most widely used antibiotics in the treatment of Gram-negative bacterial pathogens and are
considered as the last line of antibiotics to treat these kinds of pathogens.
However, the indiscriminate overuse of antibiotics for several decades has led to an antibiotic
resistance crisis (3). Bacterial pathogens rapidly form antibiotic-resistance after being exposed
to antibiotic treatment of insufficient dose. Dangerously, these resistance genes are frequently
located on movable genetic elements (transposons, insert sequences, plasmids, prophage), that
can facilitate the transfer of resistance genes to other bacterial recipients. This horizontal and
vertical transfer between pathogens has resulted in the complicated antibiotic resistance crisis
we now face (4-6). For example, in 2009, the first case of carbapenem-resistant Klebsiella
pneumonia, which carried the blaNDM-1 gene encoding a beta-lactamase, was found in India;
and now this resistance gene is found in all nosocomial Gram-negative pathogens
(Enterobacteriaceae, Pseudomonas aeruginosa, Acinetobacter baumannii) in all 5 continents
(7-9). The failure of carbapenem means there is a shortage of antibiotics to control Gram-
negative bacterial pathogens. High resistance rates have also been observed in nosocomial
Gram-positive pathogens. Vancomycin and methicillin have also lost effectiveness against

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
4
nosocomial Gram-positive bacterial pathogens, since the methicillin-resistant Staphylococcus
aureus and vancomycin-resistant Enterococcus sp. have rapidly spread worldwide (10, 11).
Therefore, antibiotic-resistant pathogens are pushing humans towards an age where there are
no effective antibiotics to fight pathogenic bacterial infections – a frightening and dangerous
situation.
Unfortunately, there is also a lack of research and few new alternative novel antibiotics have
been identified and brought to the market. Very few novel antimicrobial compounds have
recently been discovered from the terrestrial environment. The reason may be due to the
overexploitation of this traditional source for the last 90 years. Thus, the U.S. Food and Drug
Administration reported that the approval rate of new, medically important antibiotics has
decreased by 56% over the last few decades (1998 to 2002 versus 1983 to 1987) without any
evidence for an increase since that study (12). Although much effort has been carried out to
synthesize in vitro novel antibiotics, the natural world still has the potential to provide a rich
array of novel compounds
Figure 1.1. Timeline of antibiotic resistance compared to antibiotic discovery. Figure
from Zaman et al. (2017) (13).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
5
1.2. The marine environment: an alternative source for antibiotic discoveries
The marine environment is nowadays gaining interest as an alternative source for antibiotic
discovery. Marine microorganisms potentially serve as reservoirs of novel antimicrobial
compounds (14). However, the current traditional plating method only recovers about 0.001-
1% of total microorganisms from the marine environment (15). Through the recent
development of whole genome sequencing (WGS), numerous genes encoding novel
antimicrobial compounds from these cultivated bacteria have been identified, but the numbers
of such genes are only minor since studies on the marine environment have been limited, and
our knowledge on the marine species’ diversity is significantly less than their terrestrial
counterparts (16). Currently, a majority of marine bacteria are not able to be cultured. This can
lead to difficulties for drug discovery but also emphasizes the high potential of the marine
environment as a rich source of novel antimicrobial compounds for future exploitation.
In contrast to the terrestrial environment, the harsh marine conditions (i.e., limited living space
or nutrient sources; extreme chemo-physical conditions; high competition between marine
microorganisms) may act as a selective pressure driving marine bacteria to evolve increased
abilities for the production of diverse antimicrobial compounds for defence and environmental
adaptation. Notably, bacterial competition is more significant between sponges-associated
microorganism communities. Due to the typical structure with multiple pores, the sponges
provide many ideal ecological niches as sites for bacterial attachment. This leads to a high
density of microbial cells in sponge tissue with a high diversity of bacteria, microalgae, fungi,
and even viruses (17). Some sponges even have 40% of their tissue volumes contributed by
microorganism cells (17). The close cell-by-cell contact in a narrow space and limited nutrient
sources possibly leads to competition, in which bacteria tend to produce antimicrobials as
weapons against other competing bacteria. The plasticity of marine bacterial genomes may be
influenced by uptake of unique external nucleotides derived from the marine environment (18).
Sponges have proven to be a rich source of novel antimicrobials, with about 200 new bioactive
compounds published every year (17). The only way to fully explore the genomic variation of
marine species is by analysing the whole genome sequence in comparison with terrestrial
species.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
6
1.3. Marine Bacillus – promising hosts of novel antimicrobial peptides
The Bacillus genus are Gram-positive, spore-forming, facultative anaerobic bacteria, and
mostly non-pathogenic (exceptions include B. cereus and B. anthracis). They are safe for
human consumption as certified by the FDA; are fastidious growers; endogenous spore
formers; and well-known producers of various bioactive compounds including diverse
structural antimicrobial peptides (AMPs). The Bacillus derived AMPs exhibit antimicrobial
activity against a wide range of microorganisms, ranging from viruses, bacteria, and fungi to
parasites. Therefore, Bacillus species are widely used by humans in different applications,
including agriculture (as biocontrol agents, biofertilizers, probiotics), food industries (source
of enzymes, food preservatives, food additives), and pharmaceutical industries (antibiotics)
(19-21). In the marine environment, Bacillus species occupy various ecological niches, like
marine seaweeds, sponges, marine animals, invertebrates, sediments, and seawater. Members
of the marine Bacillus genus include both universal species (B. badius, B. subtilis, B. cereus,
B. licheniformis, B. firmus, B. pumilus, B, amyloliquefaciens, B. mycoides and B. lentus) which
are widely spread in both marine and terrestrial environments; and marine typical Bacillus
species (B. marinus, B. salexigens, B. dipsosauri, Halobacillus sp.). Marine Bacillus species
are considered a likely reservoir of novel AMPs, but they have not been widely studied.
The Bacillus-derived AMPs include both nonribosomal peptides (NRPs) and ribosomally-
synthesized peptides (bacteriocins) (21). They are typically small molecular-weight peptides,
amphipathic, membrane active, and exhibit various pharmaceutical activities, particularly
antimicrobial activity. The machinery to produce bacteriocins and NRPs involves a set of genes
responsible for various synthesis, processing and immunity functions, and are generally
clustered in a DNA region, called a biosynthetic gene cluster.
1.4. Non-ribosomally synthesized peptides (NRPs)- the antimicrobial peptides exhibiting
both antifungal and bactericidal activity.
The NRPs are produced by synthetase enzymes, via non-ribosomal biosynthetic pathways.
They include lipopeptides (LPs), polyketides (PKs), and bacilysin. The LPs are synthesized by
‘non-ribosomal peptide synthetase’ (NRPS) enzymes while PKs are produced by ‘polyketide
synthase’ enzymes (PKs). The NRPs are mostly produced by fungi and bacteria. In the bacterial
kingdom, production of NRPs is frequently observed in bacteria from the Actinobacteria and
Firmicutes phyla (Bacillus, Paenibacillus, Geobacillus, etc) (21-23)

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
7
The gene clusters of NRPs compounds are typically constituted of genes encoding; i) peptide
synthetase enzymes; ii) enzymes to generate monomers/acyl residues; and iii) enzymes
responsible for modification of the precursor (including processes of N-methylation, acylation,
glycosylation, tailing, and/or heterocyclic ring formation) (24). The synthase enzymes have
very large MW ranging from 100 kDa to more than 1600 kDa (25), and consist of multiple
modules. Each module has multiple domains which are fused covalently (26) [Figure 1.2]. For
details, typical NRPS and PKS enzymes include a “starting module” and multiple “extending
modules”. The starting module of PKS consists of; acyltransferase domain (AT); acyl carrier
protein domain (ACP), while each “extending module” has all above domains, with the
addition of; ketosynthase (KS); and either ketoreductase (KR) or dehydratase (DH) or enoyl
reductase (ER) or thioesterase (ET) domains. While, the NRPS enzymes consist of; peptidyl
carrier protein (PCP), adenylation domain (A), epimerase (E), methyltransferase (MT), 4’-
phosphopantetheine transferase domain (PPT), oxygenation domain (Oxy), and cyclization
domain (Cy). Of these domains, the ACP and PCP domains serve as places where specialized
amino acids/or monomers attach to each module, initiating the formation of chemical bonds
between these monomers to grow the compound chain. The ACP or PCP are usually translated
in the inactive forms and their activations are required for attachment of phosphopantetheine
(4'-PP) moiety derived from coenzyme A (CoA). This process is catalysed by the PPTase
enzyme encoded by the sfp gene. Therefore, the biosynthesis of lipopeptide and polyketide
compounds is dependent on the sfp gene (27).
Figure 1.2. Minimal domain structure of (A) nonribosomal peptide synthetase (NRPS)
and (B) polyketide synthase (PKS). C, condensation domain; PCP, peptidyl carrier
protein; A, adenylation domain; E, epimerase; MT, methyltransferase; PPT, 4’-

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
8
phosphopantetheine transferase domain; Oxy, oxygenation domain; Cy, cyclization
domain; ACP, acyl carrier protein; AT, acyltransferase domain; KS, ketosynthase
domain; KR, ketoreductase domain; DH, dehydratase domain; ER, enoyl reductase
domain; TE, thioesterase domain. The Figure is from Guzman- Chavez et al. (2018) (28)
The LPs are amphiphilic peptides, consisting of a hydrophilic polypeptide chain and a
hydrophobic acyl chain (lipid). Based on the variation of acyl chain (length, branch, linear and
saturation), LPs are distinguished into various LPs families, including three traditional families
(surfactin, iturin and fengycin), and a newly discovered family (locillomycins) (29, 30). The
LPs mainly exhibit antifungal activity, but some LPs also exhibit antibacterial activity
(paenibacterin and polymyxin) (21). The PKs are compounds that contain the typical carbonyl
methylene residue (-CO-CH2) or are derived from a precursor containing this residue. The
Bacillus-associated PKs include 3 traditional PKs (bacillaene, difficidin and macrolactin) and
newly discovered families (paenimacrolidin, basiliskamide). They mostly exhibit bactericidal
activity (zwittermicin A), but also antifungal activity (paenilamicin and paenilarvins),
antitumor and immunosuppressive activities. In contrast, bacilysin is a dipeptide and its
production is independent of 4′-phosphopantetheinyl transferase (PPTase). The bacilysin
comprises of an l-alanine residue and anonproteinogenic l-anticapsin. Bacilysin exhibits
antibacterial activity (against Erwinia amylovora, M. aeruginosa, A.
flosaquae, Nostoc sp., Anabaena sp), antifungal activity (against yeast) and also anti-algal
activity (against cyanobacteria and microalgae) (31, 32).
1.5. The bacteriocins, a promising group of antimicrobial proteins
1.5.1. Bacteriocins - definition
Bacteriocins are produced on ribosomes and are dependent on messenger RNA transcribed
from bacteriocin encoding genes. They are proteinaceous in nature, fewer than 100 amino acids
in length, and mostly depress the growth of bacterial species that are taxonomical neighbours
with the producing host. Numerous studies have been carried out to screen for bacteriocin
production from terrestrial resources, yielding various bacteriocins produced from various
bacteria (33). Bacteriocins have been most commonly found in Gram-positive bacteria, such
as Lactobacillus, Bacillus, Proteus and Actinobacteria, but some have also been found in
Gram-negative bacteria, such as E. coli, Pseudomonas sp., Vibrio sp. and Shigella sp (34, 35).
In bacterial communities, the lactic acid bacteria (LAB) group and Bacillus are two dominant

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
9
producers of bacteriocins. Bacillus-derived bacteriocins are more diverse and exhibit broader
spectra of antimicrobial activity (36, 37).
Bacteriocins have been developed and used in a variety of applications, such as natural food
preservatives (nisin), food additives (nosiheptide, duramycin) and antibiotics (thiostrepton)
(19, 20). Bacteriocins are now of interest in pharmacology because they have; (i) strong activity
against antibiotic-resistant pathogens (e.g. MRSA, vancomycin-resistant E. faecalis); (ii) low
resistant rates; (iii) potential to scale up the production via cloning and heterologous
expression; (iv) exhibit a narrow antimicrobial spectrum when compared with conventional
antibiotics, therefore, ensuring the survival of harmless bacteria in human/animal gut.
1.5.2. Bacteriocin – classification, biosynthesis
The machinery to produce bacteriocins involves various genes that are usually located together
in a biosynthetic gene cluster. The gene composition of bacteriocin biosynthetic gene clusters
is significantly diverse, but generally comprises of; structural gene(s) encoding precursor
bacteriocin; gene(s) encoding modification enzyme(s), which are responsible for modify the
precursor (in case of class I bacteriocins); gene(s) encoding transport protein, that is necessary
for bacteriocin secretion; gene(s) encoding immunity protein to protect the host from a suicide-
effect; and regulatory genes to control transcription. The precursor of bacteriocin comprises an
N-terminal leader sequence (or leader sequence) and a C-terminal core sequence. Bacteriocin
maturation frequently involves the eliminating of the signal sequence followed by
modifications of the core sequence [Figure 1.3].
Figure 1.3. Schematic representation of bacteriocin biosynthesis. Firstly, genes within
the bacteriocin gene cluster are transcribed and translated. The modification enzymes

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
10
recognise the precursor, and subsequently post-translationally modify it (PTMs). The
PTM includes the elimination of signal sequence from a precursor and mediating core
sequence to mature. The matured bacteriocin is then secreted by a transmembrane
transporter. Figure from Ortega et al. (2016) (38).
Many bacteriocins have been described in the scientific literature. They are distinguished based
on structure, mode of action, thermal stability, molecular size and glycosyl tailing (36). The
Bacillus-derived bacteriocins are more diverse than those produced by Gram-negative bacteria
or other Gram-positive bacteria (LAB, Clostridium, etc.). Therefore, many classification
schemes have developed to distinguish Bacillus-derived bacteriocins. The Bacillus-derived
bacteriocins are classified into 3 groups:
Class I bacteriocins are small (<10 kDa), heat stable and are modified compounds.
Class II bacteriocins are small (<10 kDa) and heat stable peptides.
Class III bacteriocins are larger (>10 kDa) and heat sensitive proteins.
1.5.2.1. Class I bacteriocins
Class I bacteriocins are small MW proteins (≤10 kDa). The precursor bacteriocin is firstly
translated from the structural gene and subsequently undergoes post-translational
modifications (PTMs). Based on these PTMs, they are subdivided into 7 subclasses. The
subclass I1: The PTMs generate unusual amino acids (dehydrated amino acids, and cyclised
amino acids) on the core sequence; while for subclasses I2–I7, the precursor bacteriocin
undergoes a single typical modification (head-to-tail peptide, linear azole containing peptide,
lassopeptide, thiopeptide, sactipeptide, and glycosin).
Subclass 1: Lantibiotics
Lantibiotics contain some unusual amino acids residues, such as; 2,3-dehydroalanine (Dha);
2,3-dehydrobutyrine (Dhb); lanthionine (Lan); and methyllanthionine (MeLan) (39). These
amino acids are generated by enzymatic dehydration modification and cyclisation
modification. The hydratase enzyme catalyses the dehydration process by converting all
serine and/or threonine residues on the core sequence into Dha and/or Dhb; the cyclase enzyme
catalyses the cyclisation process by formation of thioether bonds between these dehydrated
amino acids and a nearby thiol group of a cysteine residue to yield Lan (from Dha) and MeLan
(from Dhb). There are two 4 modification enzymes responsible for these modifications,

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
11
including LanB (dehydratase), LanC (cyclase), LanM (having two domains of LanB and
LanC), LanKC, and LanL (40). Depending on the modification enzymes, lantibiotics are
subclassified into 4 subgroups. Lantibiotic subtype I is modified by LanB and LanC. They are
typical of elongated or screw shape and positively charged compounds. Lantibiotic class II is
modified by LanM. The lantibiotics class II are typical of small MW peptides, globular shape,
or two component peptides, negatively charged or no net charge compounds (mersacidin,
actagardine, and cinnamycin). The dehydration process by LanB and LanM are independent of
phosphorylation. The lantibiotic class III and IV are modified by LanKC, and LanL enzymes
respectively and their dehydration are dependent on phosphorylation. The LanL and LanKC
include 3 domains; an N-terminal lyase, a central kinase and a C-terminal cyclase/putative
cyclase domain. The central kinase domain catalyzes the phosphorylation of the
serine/threonine of the core sequence to generate dehydrated phosphoSer/phorphosphoTher.
Subsequently, the lyase domain removes the phosphate residues to yield Dha and Dhb
[Figure1.4].
The lantibiotics have been reported in B. subtilis, B. thuringinensis, B. cereus, B. megaterium,
B. mycoides, B. clausii, Bacillus sp., Geobacillus thermodenitrificans, Geobacillus
kaustophilus, Paenibacillus polymyxa, P. larvae, P. peoriae and .P. durus (36, 41-43).
Figure 1.4. Schematic representation of lantibiotic biosynthesis. (A) Dehydration and
cyclization to generate the lanthionine, methyl-lanthionine from threonine. B) Four
modification enzymes in lantibiotic biosynthesis. Figure from Yu et al. (2012) (40).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
12
Subclass 2: Head to tail cyclized peptides
Head-to-tail cyclized peptides are cyclic peptides, where the cyclization occurs between the N-
terminal amino acid and the C-terminal amino acid [Figure 1.5]. It is modified by a typical
serine protease that cleaves off the leader sequence from the precursor; subsequently an amide
bond forms between the terminal two amino acids on the core sequence strand to generate the
mature cyclic peptide. The head-to-tail cyclized peptides are reported in B. thuringiensis, B.
cereus, B. coagulans, B. pumilus, B. paralicheniformis, B. gobiensis, other Bacillus sp.,
Kyrpidia tusciae, Geobacillus stearothermophilus, Geobacillus kaustophilus, Geobacillus sp.,
Paenibacillus larvae and Paenibacillus mucilaginosus.
Figure 1.5. Schematic representation of “head-to-tail linearized bacteriocin”.The
cyclization formed from 2 amino acid residues at two termini under the catalysis of a
typical protease. Figure from Gabrielsen et al. (2014) (44).
Subclass 3: Sactipeptides
The sactipeptides are cyclic peptides, where cyclization occurs between unusual sulphur and
α-carbon on side chains [Figure 1.6]. This cross-linking bond is catalysed by a typical radical
S-adenosylmethionine (SAM). The sactipetides are known to be produced by B. subtilis, B.
atrophaeus, B. simthii (subtilosin A), B. atrophaeus, B. pumilus, B. subtilis (sporulation killing
factor (SKF)) (45), B. thuringiensis and B. cereus (thuricin H and thuricin CD, thuricin 7,
thuricin S) (46, 47).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
13
Figure 1.6. Schematic presentative of sactipeptide. (A) The structure of thuricin CD (as
reference) with sulphur cross-link; (B) Illustration of sulphur cross-link. Figure from
Poorinmohammad et al. (2014) (48).
Subclass 4: Linear azole-containing peptides (LAPs)
The LAPs contain heterocyclic azole/azoline rings, which are generated from enzymatic
modification of serine/threonine and cysteine residues. Three enzymes responsible for
modification include cyclodehydrataseenzyme (protein D), dehydrogenase enzyme (protein B),
and a “docking” enzyme (protein C) [Figure1.7] (49). Protein C and protein D, together, modify
the Cys, Ser, and Thr to generate azoline heterocycles. Subsequently, protein C oxidizes some
of these azolines to azoles. The LAP bacteriocins are mostly found in Lactobacillus, E. coli
(microcin) (50), B. amyloliquefaciens (plantazolicin A and B) (51).
Figure 1.7. Schematic presentative of LAP biosynthesis.(A) LAP biosynthetic gene
cluster comprises a structural bacteriocin gene encoding the precursor of LAP (black), 3
genes encoding 3 modifying enzymes - proteins B, C, D (yellow), a gene encoding
peptidase (brown), an immunity gene (blue), and an export gene (red). The maturation of
LAP involves posttranslational modification (cyclodehydration and dehydration)
catalysed by the BCD complex and proteolysis to remove the leader sequence. (B) The

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
14
formation of azoline ring is catalysed by protein C and protein D, and azole ring is formed
under catalysis of protein B. Figure from Burkhart et al. (2015) (49).
Subclass 5. Thiopeptides
Thiopeptides contain a nitrogen-containing six-membered ring (piperidine, dehydropiperidine,
or pyridine) which serves as a central ring for the tailing of azole rings and dehydrated amino
acid residues [Figure 1.8]. Thiopeptides are typically low MW (less than 2 kDa), rich in sulphur
residues, and contain heavily modified amino acids. They commonly depress the growth of
Gram-positive bacteria with little or no activity against Gram-negative bacteria. Currently,
there are approximately 100 known thiopeptides, varying in size, charge, and branches of azole
rings. They are found in Actinobacteria, B. cereus ATCC 14579 (thiocillins), B. subtilis,
Bacillus amyloliquefaciens and Lysinibacillus sphaericus. The biosynthesis of thiopeptides
occurs under various posttranslational modification processes, including; (i) dehydration and
cyclization to generate the dehydrated amino acid ;(ii) formation of azole rings; (iii) formation
of 6-membered nitrogen heterocycle; (iv) and elimination of the leader sequence; and variable
tailing processes.
Figure 1.8. Schematic presentative thiopeptide biosynthesis. Gene organisation of the
nosiheptide gene cluster (as an example). (B) Maturation processes of nosiheptile include
the formation of azole rings, dehydration and formation of the 6-membered nitrogen
heterocycle ring. Figure from Ortega et al. (2016) (38).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
15
Subclass 6: Glycocins
Glycocins are bacteriocins tailed with a glycosyl residue [Figure 1.9]. The glycocins are found
in B. subtilis (Sublancin 168 containing a β-S-linked glucose moiety), B. thuringiensis , B.
cereus, B. weihenstephanensis, B. lehensis, other Bacillus sp., Geobacillus sp. and
Paenibacillus sp. (52)
Figure 1.9. Schematic representation of glycocins biosynthesis. Sublancin as an example.
The bacteriocin is tailed with sugar moiety. Figure from Hseih et al. (2012) (52).
Subclass 7: Lassopeptides
Lassopeptides contain a macrolactam ring at the N-terminus. Two enzymes; a lasso cyclase
(protein C), and leader peptidase (protein B) are needed to generate the macrolactam ring by
forming a cross-link between either a glycine/alanine/serine or cysteine at the N-terminus and
the carboxylic acid residue of aspartate/glutamate on the side chain to cyclize [Figure 1.10].
The lassopeptides are found in Actinobacteria, Proteobacteria and Firmicutes, such as
Paenibacillus dendritiformis C454, Thermobacillus composti KWC4, Bacillus cereus VD115,
and Paenibacillus polymyxa CR1.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
16
Figure 1.10. Schematic representation of lassopeptide biosynthesis.Figure from Zhu et
al. (2016).
1.5.2.2. Class II: unmodified bacteriocins
In contrast to class I bacteriocins, class II bacteriocins undergo no posttranslational
modifications. They typically have an MW of <10 kDa, are heat-stable, and have linear
structure. The biosynthetic gene clusters of class II bacteriocins include a structural gene,
transport gene(s) and immunity gene(s). There are 4 subclasses within the class II bacteriocins,
distinguished based on the shape and ability to kill Listeria sp. Subclass IIa are pediocin-like
bacteriocins and exhibit anti-Listeria activity. The class IIb, class IIc and class IId are non-anti-
Listeria bacteriocins. Subclass IIb are two-component peptides, subclass IIc are circular, and
subclass IId are linear. The class II bacteriocins are found in B. coagulans I-4 (coagulin) (53),
Geobacillus sp. WCH70 (lactobin A, holins), B. subtilis, B. amyloliquefaciens, B. mycoides, B.
pseudomycoides, B. licheniformis, B. pumilus, B. thuringiensis (holin-like peptide BhlA), B.
amyloliquefaciens, Bacillus (LCI) and B. methylotrophicus (aureocin A53) (22, 54).
1.5.2.3 Class III: large antimicrobial proteins
Class III bacteriocins have larger MWs (>10 kDa) and are heat sensitive. The biosynthetic gene
clusters of class III bacteriocins include an immunity gene and a structural gene. The III
bacteriocins are found in B. megaterium ATCC 19213 (megacin), Escherichia coli (Colicin),
Bacillus sp. (M23 metallopeptidase family) (55-57).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
17
1.5.3. Mode of action and bacteriocin resistance rate surveillance among bacterial
pathogens.
The bacteriocins are membrane-active peptides, usually cationic-charged; therefore, they bind
to bacterial cell membranes which present mostly negatively charged components, initiating
different modes of action. Class I bacteriocins bind to the lipid II molecules which are main
transporters of cell wall components - the peptidoglycan subunits; therefore, preventing cell
wall formation. The binding results in a ‘bacteriocin-lipid II’ complex, serving as a pore
through the membrane, causing leakage of internal materials of cells. The class II bacteriocins,
which are typically have helical and/or amphiphilic structures, bind to phospholipid heads of
the membrane at hydrophobic regions and subsequently insert the hydrophilic part into the
membrane. The hydrophilic part binds to mannose phosphotransferase systems (PTS-Man)
proteins and together form a pore causing depolarization, resulting in cell death. Class III
bacteriocins can hydrolyse the cell wall components of Gram-positive bacteria, causing loss of
membrane integrity [Figure 1.11] (58).
There are several mechanisms by which bacteriocin-sensitive bacteria can develop resistance,
including; (i) modification of bacteriocins’ receptors on the membrane to prevent bacteriocin
binding; (ii) sequestering of bacteriocins; (iii) formation of efflux pumps to expel the
bacteriocins, or; (iv) production of bacteriocin-degrading enzymes. Different bacteria employ
different strategies, and even the same species can demonstrate different mechanisms.
Bacteriocin resistance frequencies in bacteria are low compared to the resistance rate of
conventional antibiotics, ranging from 10-9 to 10-2 (59, 60).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
18
Figure 1.11.Bacteriocin’s mode of action. Figure from Gražina et al. (2012) (58).
1.6. Current knowledge of marine-derived bacteriocins
Marine-derived bacteriocins have been much less studied than terrestrial-derived bacteriocins.
Marine bacteriocins have been mostly reported from lactic acid producing bacteria
(Lactobacillus, Carnobacterium), Bacillus sp. and, to a lesser extent, Enterococcus, Proteus,
Vibrio and Pseudomonas [Table 1.1]. These bacteria encoding bacteriocin production were
isolated mostly from seafood or aquatic animal (fish, prawn) intestines (61, 62), but also
sponges, seaweed and sediment (63, 64). These marine-derived bacteriocins were mostly small
(<10 kDa) and exhibited a variety of antimicrobial activities. Generally, Lactobacillus-
associated bacteriocins exhibited the broadest spectra of activity against both Gram-positive
and Gram-negative bacteria (against Vibrio, Shigella, E. coli, Salmonella sp); while the Vibrio
and Proteus derived bacteriocins commonly exhibited antimicrobial activity against Vibrio sp.
The bacteriocins produced by Carnobacterium or Bacillus possessed antimicrobial activity
mostly against Gram-positive bacteria, but also against Salmonella sp. and Vibrio sp. Most of
the bacteriocins are still not fully characterised, therefore they have been named BLIS
(bacteriocin-like-inhibitory substance) factors. BLIS factors are uncharacterised bacteriocins
that share similar activity and characteristics of typical bacteriocins, but the sequence is still to
be determined.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
19
Table 1.1. List of marine bacteriocins published.
Bacteriocins Producers strains MW Killing spectra Isolation source Reference
BLIS Lactobacillus pentosus 39 - A. hydrophila; L. monocytogenes salmon fillets (65)
BLIS Lactobacillus lactis 94 Bacillus, Staphylococcus, Enterococcus, E. coli, Pseudomonas, Shigella
sediment (64)
bacteriocin Lactobacillus fermentum 18 Vibrio, Listeria, Listeria sp., Staphylococcus fish gut, prawn muscle
(66)
SBS001 Lactobacillus fermentum 78 Klebsiella, Pseudomonas, E. coli seawater (67)
AP8 Lactobacillus casei 5 E. coli, Listeria spp., Salmonella sp., Staphylococcus, , Vibrio
fish gut (68)
bacteriocin Lactobacillus acidophilus 2.5 Lactobacillus, Salmonella, Staphylococcus, Bacillus, Salmonella, Escherichia, Klebsiella, Pseudomonas
prawn gut (62)
bacteriocin PSY2 Lactococcus lactis PSY2 Arthrobacter sp., Acinetobacter sp., Bacillus marine perch fish (69)
carnocin U149 Carnobacterium sp. 4.5 Lactobacillus, Pediococcus, Carnobacterium fish gut (70)
divergicin M35 Carnobacterium divergens M35 4,5 Carnobacterium, Listeria smoked mussel (71)
divergicin V41 Carnobacterium divergens V41 4.5 Carnobacterium, Listeria, Enterococcus, fish viscera (72)
carnobacteriocin Carnobacterium piscicola A9b 4.5 Listeria smoked salmon (73)
piscicocin CS526 Carnobacterium piscicola CS526
4.4 Tetragenococcus, Leuconostoc, Listeria, Enterococcus.
frozen surimi (74)
piscicoccin V1a Carnobacterium piscicola V1 4.5 Lactobacillus, Listeria, Enterococcus, Pediococcus. fish (75)
BLIS Enterococcus faecium CHG 2-1 - Enterococcus venus clams (76)
BLIS Enterococcus faecium C-K, C-S - Listeria fish fillet (76)

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
20
Bacteriocins Producers strains MW Killing spectra Isolation source Reference
enterococinB Enterococcus faeciumALP7 <6.5 Listeria, Staphylococcus, Bacillus, Lactobacillus. fermented shellfish (77)
BLIS Vibrio sp <5 Bacillus sp., Vibrio sp., Pseudomonas sp. marine fish gut (78)
BLIS Vibrio harveyi - - marine fish gut (79)
BLIS Vibrio spp. MMB2 - Aeromonas hydrophila marine microalgae (80)
BLIS Proteus sp.CT1.1, G1 - Vibrio marine fish, lobster (81)
BLIS Bacillus cereus - Vibrio marine fish (81)
BL8 Bacillus licheniformis <3 Staphylococcus, Bacillus sp. sediment (82)
bacteriocin Bacillus sp. NM12 <5 fish pathogens fish gut (78)
bacf3 Bacillus amyloliquefaciens ~3 Bacillus sp., Staphylococcus shark (83)
subtilomycin Bacillus subtilis MMA7 3.2 Bacillus, Listeria, Clostridium, Enterococcus marine sponge (84)
sonorensin Bacillus sonorensis MT93 6.3 Bacillus, Staphylococcus, Listeria, Pseudomonas, E. coli, Vibrio
sediment (85)
lichenicidin Bacillus licheniformis MRSA, Listeria, Enterococcus, Salmonella, E. coli seaweed (86)
pumiviticin Bacillus pumilus DR2 3.9 Salmonella, Proteus, Micrococcus, Lactobacillus seawater (87)
bacteriocin Bacillus sp. - Bacillus, Clostridium, Corynebacterium, Listeria fish gut (88)
bacteriocin Pseudomonas putida FStm2 Bacillus, Staphylococcus, Escherichia, Enterobacter, Serratia, Salmonella sp., Klebsiella, Vibrio, Pseudomonas
shark skin
(89)
BLIS: Bacteriocin-like-inhibitory substance

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
21
1.7. Research questions and thesis objectives
1.7.1. Research questions
Q1. Can Bacillus strains with a diverse range of antimicrobial activities be isolated from marine
environments?
Q2. Can these antimicrobial activities be harnessed to address a range of pathogenic microbial
threats to humans and animals?
Q3. Are there novel, previously uncharacterised, antimicrobials present within these isolates?
Q4. Can novel bacteriocin(s) be identified?
Q5. Can novel bacteriocin/s in marine isolates be heterologously cloned and expressed?
1.7.2. Research aims and significance.
The overarching aim of this project was to identify and characterise novel antimicrobial
compounds, particularly bacteriocins. This was to be achieved by:
Isolating marine-derived spore-forming bacteria within NhaTrang Bay, Vietnam Sea that
demonstrate inhibition against both Gram-positive and Gram-negative pathogens. The
diversity of antagonistic spore-forming bacteria within this environment was then evaluated.
Characterising isolates that show high antimicrobial activity by:
+ Testing for antimicrobial activity against a range of bacterial pathogens, including food
spoilage bacteria, and human and/or animal pathogens.
+ Bacterial identification based on 16S rRNA sequences.
+ Determining the nature of antimicrobial compounds presented in the media culture.
+ Selecting the isolates that were able to kill other marine antimicrobial producing species.
It likely indicates that these isolates produce novel antimicrobial compounds capable of
attacking other marine bacteria which harbour common immunity systems.
+ Characterising the nature of the antimicrobial products produced by these isolates to provide
evidence of bacteriocin production.
Genomics, via whole genome sequencing for promising isolates which displayed broad
spectra of inhibition. Various bioinformatics approaches were used to identify genes that are

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
22
likely to encode putative antimicrobials, with aims at understanding the mechanism of
antimicrobial production and identification of novel bacteriocin(s).
Purification of antimicrobial compounds for characterisation, using cation exchange
chromatography, hydrophobic interaction chromatography, reverse-phase high-pressure liquid
chromatography (RP-HPLC), matrix-assisted laser desorption ionization-time of flight mass
spectroscopy (MALDI-TOF MS) and other peptide sequencing techniques (N-terminal
sequencing and de novo sequencing).
Cloning and expression to produce the novel bacteriocins in a heterologous Bacillus subtilis
host.
1.7.3. Novel contributions
Elucidating the diversity of marine antagonistic Bacillus within NhaTrang bay, Vietnam Sea,
which has previously been unexplored
First employment of both genomic and protein approaches for antimicrobial compound
purification to understand the marine Bacillus genome and the potential of antimicrobial
compounds produced by marine Bacillus species.
Providing information about gene organisation of some novel bacteriocin biosynthetic gene
clusters (2 thiopeptides, 1 two-component lantibiotic, 1 sactipeptide and 1 lantibiotic, and 1
lassopeptide), which will be utilised for further cloning and heterologous expression to produce
these bacteriocins for both characterisation and commercialisation.
Successful cloning and expression of the sactipeptide bacteriocin.
1.8. Thesis organization
The project covers the following chapters:
Chapter 1: “Introduction”
Chapter 2: “Research methodology”.
Experimental chapter 3: “Broad antimicrobial spectra from marine spore forming bacteria
isolated from the Vietnam Sea”
This chapter includes the work of; sampling marine samples within the Vietnam Sea; the
isolation of the spore forming bacteria; screening to select candidates exhibiting antimicrobial
activity; selection of the promising candidates which may produce novel bacteriocins.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
23
This chapter aimed to understand the diversity of antibacterial marine isolates within NhaTrang
bay and select the isolates producing probable novel antimicrobial compounds. The findings
of this chapter will answer Research Questions 1 and 3.
Experimental Chapter 4: “Bioinformatic identification of biosynthesized gene clusters of
antimicrobial peptides within marine Bacillus genomes”.
Chapter 4 covers the work of; whole genome sequencing, genome annotation, genomic
comparisons between marine isolates and terrestrial neighbour species, to understand the
variation between genomes, and in silico prediction of all putative antimicrobial peptides
within 6 genomes. This chapter aims to understand the marine Bacillus genome characteristics
and select the isolates having biosynthetic genes clusters of novel bacteriocins for antimicrobial
purification. This chapter will answer research question 2.
Experimental chapter 5: “Purification and characterisation of antimicrobial peptides
produced by Bacillus amyloliquefaciens #11”
This chapter covers the purification of antimicrobial compounds produced by this bacterium.
The purification applies various techniques of separation and mass spectrometry.
This chapter aims to purify all the antimicrobial peptides that the bacterium produced to obtain
a novel bacteriocin (thiopeptide) which was predicted from the genome sequence. This chapter
will answer research question 4.
Experimental chapter 6: “Purification and characterisation of antimicrobial peptides
produced by Paenibacillus polymyxa #23”.
This chapter covers the purification of antimicrobial compounds produced by this bacterium.
This chapter aims to purify all the antimicrobial peptides that the bacterium produced to obtain
novel bacteriocins (sactipeptide, lantibiotic, lassopeptide) which were predicted from the
genome sequence. This chapter will answer research question 4.
Experiment chapter 7: “Cloning and heterologous expression of sactipeptide, a novel
bacteriocin from marine Paenibacillus polymyxa #23, in Bacillus subtilis”.
The biosynthetic gene cluster of a novel sactipeptide (8,556 bp) was identified by in silico
analysis of the isolate #23’s genome; however, this bacteriocin was purified (mentioned in
Chapter 6). Therefore, this chapter aims to produce this sactipeptide by a heterologous Bacillus
subtilis host for characterisation.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
24
The chapter includes work of; preparation of the sactipeptides gene clusters (8,556 bp) by
amplification from the genome of wild-type P. polymyxa #23 host, preparation of the plasmid
backbone originating from natural plasmid pDLL202, construction of the fusion plasmid from
these two materials by Gibson assembly, a transformation of the fusion plasmid to E. coli
TOP10 to control the plasmid copy number, transformation of fusion plasmid into E. coli
JIR702 to generate the multimeric fusion plasmid, transformation of the multimeric plasmid to
Bacillus subtilis BS34A for expression, optimization of sactipeptide production, purification
of sactipeptide, and characterisation of the sactipeptide. This chapter will answer research
question 2.
Chapter 8. Conclusion and future work.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
25
Chapter 2 RESEARCH METHODOLOGY
2.1 Sampling and sample preparation for bacterial isolation
Marine samples were collected by scuba-divers from 3 sites in NhaTrang bay (Vietnam Sea),
including Hon Mieu island (12.191837, 109.235086), Hon Mot island (12.173368, 109.271591)
and Hon Rua island (12.289501, 109.242413) [Figure 2.1]. The sites were chosen based on sea
sponge abundance. The sampling included sponges, seaweeds, sediments and seawater
samples. All the samples were transported under refrigerated conditions to the laboratory for
bacterial isolation.
Figure 2.1.Locations of sampling trips within Nhatrang bay (Vietnam sea).Three (X)
symbols indicate the three sites where marine samples were collected.
2.2. Isolation of spore-forming bacteria from marine samples
Solid samples were washed three times with sterile seawater (SSW) to remove externally
attached bacteria, 10 g of material was then homogenised in 90 ml of SSW using a sterile mortar
and pestle. The water samples were concentrated by centrifugation at 2500 x g for 20 minutes
to achieve 10 times concentration (50 mL in initial volume to 5mL). Sediments were dried by
heating at 50°C for 3 hours or air-dried overnight, and 10 g of each were vortexed in 90 mL of
SSW for 5 minutes, followed by centrifugation at 1000 x g for 1 minute. The supernatants were
collected, heated at 80°C for 20 minutes to kill non-spore-forming bacteria, serially diluted 10-

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
26
fold, and 100 µL aliquots were plated onto laboratory-prepared marine agar (LPMA) to isolate
the spore forming bacteria. Plates were incubated at 30°C for up to 3 days, and colonies were
picked daily based on morphology (size, colour, shape), and re-streaked onto LPMA plates to
obtain pure colonies. All pure isolates were stored at -80°C in LPMB supplemented with 20%
glycerol.
2.3. Assays to detect the antimicrobial activity.
2.3.1. Cross-streak assay for primary screening of marine isolates exhibiting
antimicrobial activity
Each bacterial isolate was streaked vertically onto an LPMA plate and incubated overnight at
30oC. The six indicator strains used in the initial screening were S. aureus, B. cereus, C.
albicans, E. coli, P. aeruginosa, and a methicillin-resistant S. aureus (MRSA). Each indicator
strain was streaked horizontally, from the edge of the plate to the pre-grown isolate streak. The
cross-streak assay plates were then incubated in the growth conditions appropriate for each
indicator strain, as detailed in Table 2.1. The gap between indicator strains and the test isolates’
growth indicated the presence or absence of antimicrobial activity [Figure 2.2]. After inspection
of the strength and spectrum of antimicrobial activity, a short-list of isolates with the strongest
antimicrobial activity was chosen for more detailed analysis (90). Their antagonistic activities
against 14 important pathogens were evaluated. The cross-streak assay was also used to detect
antimicrobial activities produced by the 23 short-listed isolates against the other members of
that group. Each marine isolate was streaked down the middle of LPMA plates, incubated
overnight at 30oC to allow growth and production of antimicrobial compounds, and then the
other 22 isolates were streaked from the edge of the plate to the central streak. The plate was
incubated overnight at 37oC and antimicrobial activity was determined based on the size of the
clear zone between test and indicator bacterial streaks.
Figure 2.2. Cross-streak assay.(A) Plates with antimicrobial-producing marine isolates as
there are zones of no growth between the marine isolate (vertical line), and indicators

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
27
(horizontal line). (B) Plate with a non-antimicrobial producing isolate without these
zones.
Table 2.1. List of indicators strains used in antimicrobial screening assays
Indicators Origins Media/temp./condition
(A) Gram-positive bacterial indicators
Lactobacillus plantarum Food origin, RMIT MRS/37°C/aerobic
Bacillus cereus ATCC 10876 MH/30°C/aerobic
Streptococcus faecalis ATCC9790 BA /37°C/aerobic
Staphylococcus aureus ATCC25923 BA /37°C/aerobic
MRSA Human pathogen, RMIT BA /37°C/aerobic
Listeria monocytogenes Human pathogen, RMIT BA /37°C/aerobic
Clostridium perfringens Chicken pathogen, RMIT BA /37°C/anaerobic
VRE Human pathogen, RMIT BA /37°C/aerobic
(B) Gram-negative bacterial indicators
Salmonella Enteritidis ATCC 13076 MH/37°C/aerobic
Escherichia coli ATCC 25922 MH/37°C/aerobic
Pseudomonas aeruginosa ATCC 15442 MH/37°C/aerobic
Campylobacter jejuni Chicken pathogen, RMIT BA/37°C/microaerophilic
Multidrug-resistant Klebsiella pneumonia
Human pathogen, RMIT MH/37°C/aerobic
(C) Yeast
Candida albicans ATCC 10231 MH/30°C/aerobic
2.3.2. Well-diffusion assay
The well-diffusion assay (WDA) was used to detect the antimicrobial compounds in a liquid
sample (91). This method relies on the diffusion of antimicrobial compounds through agar

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
28
plates, which then exhibit antimicrobial activity against sensitive bacteria, reducing or
preventing growth [Figure 2.3]. The diffusion of antimicrobial compounds creates a radial
concentration gradient outward from the well. Indicator strain sensitivity to antimicrobials can
then be quantified based on the measurement of the diameter of inhibition zones. The Muller
Hilton agar (MHA) (Oxoid) was used in the well diffusion assay as it supports the optimal
diffusion of antimicrobial products through the agar. MHA is a non-selective medium allowing
growth of all organisms plated and contains starch which can absorb possible toxins affecting
the bacterial growth, and also serves as a loose agar allowing better diffusion of the
antimicrobial compounds.
The well-diffusion assay was employed at this stage alongside the cross-streak method to
understand if the antimicrobial compounds were secreted into the culture supernatant. To make
the indicator lawn, several fresh colonies of indicator bacteria were suspended in buffered saline
to final turbidity of 0.5 McFarland units (OD60 0~0.08-0.1). A cotton bud was wet in this
bacterial suspension and used to inoculate a lawn of bacteria on MHA. A plug of the agar was
removed using a sterile glass Pasteur pipette (diameter of 6mm) and 100 µL of antimicrobial
samples (or media culture) was added to the wells then allowed to diffuse and incubated at the
indicators strains growth conditions. The diameter of the cell-free zone was measured.
Figure 2.3. Well-diffusion assay.The bacterial culture (or antimicrobial sample) was
added to the well. (+) presence of antimicrobial compounds with no bacterial growth;
(-) no antimicrobial compounds produced

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
29
2.3.3. Spot-on-lawn assay
The spot-on-lawn assay was also used to detect the presence of antimicrobial compounds in
liquid samples (92). It also relies on the diffusion of antimicrobial compounds across the agar
surface that consequently depresses the growth of indicator bacteria. The antimicrobial sample
is dropped directly onto the agar surface without the requirement of wells, therefore the method
uses less sample than other techniques (10 - 20 µL) and is suitable for enriched samples [Figure
2.4]
In this study, the spot-on-lawn assay was mainly used to detect the antimicrobial fractions
during purification. It was also used to identify marine bacteria exhibiting antimicrobial activity
via detection of inhibitory activity from fermented media culture. In practice, indicator colonies
were suspended in 20 mL of warm liquid soft media agar (0.75% agar) to a final cell density of
108 CFU/ mL, mixed by inversion and poured in a Petri plate and allowed to solidify. Next the
agar surface was dried in the fume hood for 10 minutes. Then 10-20 µL culture supernatant (or
antimicrobial liquid) was spotted directly onto the media surface, allowed to diffuse, and the
plate was incubated at indicators’ growth conditions. A zone of growth inhibition formed
around the bacterial spot indicated the presence of antimicrobial compounds.
Figure 2.4. Spot-on-lawn assay.The spot with the presence of a killing zone indicated the
presence of antimicrobial compounds.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
30
2.4. Bacterial identification based on 16S rRNA gene sequence and phylogenetic tree
construction
2.4.1. Extraction of genomic DNA by bead beating
Bacteria were grown in LB broth overnight (until stationary phase with an approximate of 2 x
109 cell/mL). Approximate 2 mL of bacterial cultures were centrifuged to collect the cell pellet.
It was washed twice with TE Buffer low EDTA (Tris-EDTA pH 8.0; 10 mM Tris base, 0.1 mM
EDTA) to remove the remaining culture, then resuspended in 500 µL of TE buffer low EDTA,
and used for DNA extraction by the bead beating method (93). The suspension was transferred
to a screw-cap polypropylene 1.5 ml vial containing 100 µL of 100 μm glass beads. The bead
beating was carried out in a Mini-Beadbeater in two cycles of 1 min at 3,450 oscillations/min
with a 1 min period of cooling on ice between cycles (94). After the bead beating, the cell debris
was removed by centrifugation at 14,000 g for 10 minutes at 4oC; and the supernatant was
collected in a fresh tube for PCR amplification.
2.4.2. Amplification of bacterial 16S rRNA gene sequences, Sanger sequencing and
phylogenetic identification
Bacterial 16S rRNA genes were amplified from the isolated gDNA. PCR was conducted using
primers with the sequences (5’–3’) GGCGTGCCTAATACATGCAA and
TACAAGGCCCGGGAACGT (95, 96). PCR conditions comprised initial denaturation at 98°C
for 30 seconds, 30 cycles of 98°C for 5 seconds, 56°C for 10 seconds and 72°C for 20 seconds,
with a final extension at 72°C for 2 minutes; and then storage at 4°C. PCR products were
checked by electrophoresis in 1% agarose gel, subsequently purified by QIAquick PCR
Purification Kit (Qiagen), and then Sanger-sequenced (Micromon, Monash University,
Australia). The raw reads were trimmed of unclear nucleotides at both 5’ and 3’ terminal ends,
and subsequently BLASTn was used to search for homologies on the Bacterial 16S rDNA
Database (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The highly homologous 16S rDNA
sequences were downloaded for phylogenetic tree construction. All the sequences were aligned
using ClustalW (97) and the phylogenetic tree was subsequently constructed in MEGA7 (98)
using the neighbour joining method with bootstrap tests performed 1000 times and pairwise
detection.
2.5. The sensitivity of antimicrobial activities to enzyme and heat.
Enzymatic treatments of cell-free supernatants (CFSs) were conducted for 3 hours at 37°C with
pronase-E (from Streptomyces griseus), proteinase, trypsin, and lipase, at final concentrations

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
31
of 2 mg/mL. All enzymes were purchased from Sigma Aldrich. The antimicrobial activities of
the treated CFS preparations were evaluated by well-diffusion assay, against Clostridium
perfringens. To evaluate the thermal stability of antimicrobial substances, the antimicrobial
sample was heated at 60oC, 80 oC, 100 oC for 30 minutes, 60 minutes respectively then chilled
on ice. The antimicrobial activities of the treated samples were evaluated by well-diffusion
assay or spot-on-lawn assays against indicators.
2.6. Analysis of self-depression ability across marine Bacillus/Paenibacillus species
This experiment was designed to demonstrate whether these marine isolates were able to
produce novel antimicrobial compounds. It was set up based on the hypothesis that
antimicrobial-producing bacteria always have an immunity system to protect themselves from
the activity of the antimicrobial compounds that they produce. If a bacterium is killed by another
closely related bacterium (particularly by a strain of same species), it indicates that the bacteria
doing the killing may be producing uncommon antimicrobial compounds. Therefore, the self-
depression observed between strains of the same species may indicate the production of a novel
antimicrobial compound. The experiment was carried out to select isolates for subsequent
whole genome sequencing (WGS), to search novel antimicrobial substances. In this study, the
self-depression experiment employed the cross-streak assay. In practice, each marine Bacillus
isolate was first streaked vertically and the other 22 isolates were streaked horizontally to serve
as indicators. The inhibitory gaps formed between the two biomasses were recorded. The assay
was replicated for all 23 marine isolates.
2.7. Growth properties, antibiotic susceptibility testing, and enzyme production of isolates
The ability of the short-listed isolates to grow on different media was evaluated by spotting 5
µL of bacterial cultures onto several different media, including low nutrition media, such as
marine agar (MA 2216) and LPMA, and rich nutritious media, such as LB agar and Muller
Hilton agar, and incubated at 30oC overnight. Aliquots of cultures were spotted on LB agar
plates, and incubated under microaerophilic, aerobic, and anaerobic conditions at 30°C, and
aerobically incubated at 40°C and 50°C. To measure pH tolerance, bacterial cultures were
spotted onto LB plates in which the media had been adjusted to pH 5.0, 6.0, 7.0, 8.0, and 9.0.
Sodium chloride tolerance was determined on LB plates supplemented with 0%, 1%, 2%, 4%,
6%, 8%, 10%, 12% and 15% (w⁄v) NaCl; while bile salt tolerance was carried out on LB plates
supplemented with bile salt to final concentrations of 0.1 M, 0.2 M, 0.3 M, 0.4 M, 0.5 M, 0.6
M, 0.7 M (Bile Salts Mixture No. 3; Neogen Corporation). For antibiotic susceptibility testing,

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
32
LB agar plates were prepared supplemented with tetracycline, ampicillin, nalidixic acid, and
kanamycin at a final concentration of 50 µg/mL. The production of proteases, cellulases, and
amylases was evaluated respectively by spotting of 5 µL of bacterial cultures onto skim milk
agar; carbon deficient media (CDM) supplemented with 1% carboxymethylcellulose (CMC),
and CDM supplemented with 1% soluble starch. CDM contained 0.1 g/L yeast extract; 0.5 g/L
peptone; 16.0 g/L agar. Plates were incubated at 30oC overnight, followed by flushing the CMC
plates and starch agar plates with Gram’s iodine solution (Sigma Aldrich) for 1 minute. Positive
reactions, indicating enzymatic activity, were noted via halo zones around the bacterial spots.
2.8. Bacterial whole genome sequencing
2.8.1. Extraction of genomic DNA (gDNA) for whole genome sequencing
Six isolates selected for whole genome sequencing were grown in LB broth overnight (to
stationary phase). The cultures were then centrifuged to collect the cell pellets which were
washed twice with milli-Q water and gDNAs were extracted by the ‘Guanidine thiocyanate
method’ (GES method) (99)
Cell pellets were suspended in 100 µL of 50 mg/mL lysozyme solution in TE buffer and
incubated at 37°C for 30 minutes. Then 500 µL of GES solution was added, gently inverted to
mix. These mixtures were then left at room temperature for 10 minutes. Next, 250 µL of cold
ammonium acetate (7.5 mol/L) was added to cell lysates; mixed by gently inverting the tubes
and left at room temperature for 10 minutes to precipitate proteins and large RNAs. 500 µl of
chloroform:phenol:isoamyl solution (26:24:1) was added; the tubes were inverted 6-8 times to
mix; followed by centrifugation at 25,000 g for 10 minutes. The top aqueous layers were
collected by gently pipetting using plastic transfer pipettes. Then 0.54 volumes of cold
isopropanol were added to these layers, inverted gently 6-8 times to precipitate gDNAs. The
gDNAs were collected by centrifugation at 10,000 g for 2 minutes at 5°C. The supernatant was
decanted and discarded. The DNA pellets were washed 5 times by gently adding 1 mL of cold
70% ethanol to tubes; centrifuging at 8,000 x g for 2 minutes; decanting the supernatants. The
washed DNA pellets were dried using a Speed-Vac at 40oC for 5 minutes and suspended in 100
µl nuclease-free water or 100 µl TE buffer low EDTA.
2.8.2. Qualification of the gDNA
The gDNAs’ quality was assessed with the aid of a NanoDrop One spectrophotometer
(ThermoFisher Scientific) and DNA electrophoresis. Thus, 1 µL of each gDNA was load onto
the NanoDrop; values of DNA concentration, A260/ A280 ratio (wavelengths of 260 nm:

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
33
wavelengths of 280 nm), and A260/A230 ratio were inspected to estimate the contamination
of RNA and proteins, as well as extraction solvents. The A260/ A280 ratio was expected to be
~1.8 while the A260/ A230 ratio was expected to be within the range of ~1.8-2.2.
Also, 1µL of each gDNA was loaded in an agarose gel (0.8% agarose gel) that was
supplemented with 1X Gelred nucleic acid gel stain (Biotum) during gel casting. The DNA
electrophoresis was performed alongside GeneRuler™ 1kb Plus DNA Ladder (Thermo
Scientific). The electrophoresis conditions were 100V for 40 minutes, followed by visualization
of gDNA bands on the agarose gel with the aid of gelDoc device (BioRad). The samples were
considered to contain gDNAs of suitable quality when there was a single band of DNA which
was higher than the top band of the DNA ladder (10,000bp). The gDNA was then adjusted to a
final concentration of 2.5 µg/mL by the aid of the Qubit dsDNA HS Assay Kit (Thermo Fisher
Scientific) to be ready for DNA library preparation
2.8.3. Preparation of DNA library and whole genome sequencing
The genomic library preparation was performed using the Nextera XT DNA Library
Preparation Kit (Nextera XT DNA Library Prep Reference Guide, Document # 15031942 v05
May 2019) following the manufacturer’s instruction. The libraries were denatured with NaOH,
diluted with hybridization buffer, pooled and sequenced using Illumina MiSeq with 2x300 bp
paired-end reads.
2.9. Assembly of raw reads generated from whole genome sequencing
The raw read data (fastq) were assembled using the A5-miseq pipeline under default
parameters for bacterial species (100). During assembly, the raw reads were automatically
trimmed, filtered by quality, corrected for errors, and finally assembled to generate contigs. The
outcome of the assembly process was draft genomes of bacteria, which contained multiple
contigs.
2.10. Calculation of Average Nucleotide Identities (ANI) across genomes of terrestrial and
marine Bacillus/Paenibacillus species
The genome similarity amongst the marine isolates versus the terrestrial closely related
Bacillus/Paenibacillus strains was estimated using Average Nucleotide Identity (ANI) and pair-
wise BLASTn for similarity search of nucleotide sequences. The terrestrial closely related
Bacillus/Paenibacillus were selected based on the similarity in 16S rDNA sequences with ones
of the marine isolates. All genome sequences were downloaded (from NCBI) as fasta files, and

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
34
then submitted to web-based ‘Rapid Annotation using Subsystem Technology’ (RAST) for
internal consistency in gene-calling using the Glimmer algorithm and amino acid translation
(101). The majority of the terrestrial genomes had been described and annotated on NCBI, but
their genomes were submitted again to RAST for consistency of annotation. The GENBANK
files generated by RAST were downloaded for calculation of the ANI value across the genomes
by the GET_HOMOLOGUES methodology (102). The distance matrix was visualized as a heat
map to show similarities and differences between genomes within the corresponding
phylogenetic tree.
2.11. Estimation of a frequency distribution of CDS across genomes
The frequency distribution of CDS across genomes were identified by the
GET_HOMOLOGUES tool employing the BLASTp for similarity search of amino acids
sequences (CDS), and the OrthoMCL algorithms (103). The outcome of this calculation was 4
directories; core genes, soft core, shell genes, and cloud genes. The 'core' catalogue contains
genes presented in all genomes. The soft-core catalogue contains genes found in 95% of
genomes. 'Shell' catalogue contains dispensable genes which remain moderately conserved and
present in several genomes. “Cloud’ catalogue contains strain-specific genes.
In practice, three analyses were calculated separately for 3 datasets; one dataset of 6 genomes
of B. amyloliquefaciens species, one dataset for 6 genomes of B. halotolerans species, and one
dataset for 6 genomes of P. polymyxa species. In each calculation, a pangenome matrix was
first obtained by calling ‘get_homologues.pl’ Perl script with option-t 0 reporting all the
computed CDS clusters, OMCL (-M), E < 1e-05 for BLASTp searches; and -C 75% as the cut
off for BLASTp (103). Next the cluster directories were a result of calculation by calling the
‘compare_clusters.pl’ Perl script with ‘pangenomes matrix’ as input generated from the above
script. Next the 'parse_pangenome_matrix.pl' was called to classify genes in these four
compartments of core genes, softcore, shell genes, and cloud.
Ratios between these compartments were compared across datasets (B. halotolerans, B.
amyloliquefaciens, P. polymyxa). This calculation aimed to understand which species having
higher degrees of diversity in genomes, which probably indicates a higher potential of novel
antimicrobial substances for discovery.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
35
2.12. Comparison between genomes of marine isolates and genomes of phylogenetically
related terrestrial strains.
The degree of similarity across genomes can be also confirmed by two bioinformatic tools;
“Sequence-based comparisons tool (SBCT)” of RAST annotation tool and MAUVE alignment
were employed to investigate diversity in the genomes across the bacilli species (104, 105).
The SBCT is used to determine the percentage of similar Coding sequences (CDS) found across
the genomes by using pair-wise BLASTp [Figure 2.5A]. The outcome is a circular graphic of
the chromosomes, in which each-by-each CDS was compared for similarity, then colour coded
based on the degree of similarity detected (from red to purple), as well as potentially being
marked as either unique or a unidirectional best hit or a bidirectional best hit [Fig.2.5A]. The
degree of similarity across genomes can be determined via a percentage of similar proteins
shared between genomes identified at a certain similarity threshold. In many similar studies, a
threshold of greater than 70% similarity has been used to evaluate the core proteins (106); while
in a recent publication a threshold of >50% similarity was used (107). In this study, the higher
threshold of greater than 70% similarity was used to obtain the percentage of similar protein
between related isolates. The higher the percentage of similar proteins (with a threshold of
greater than 70%) two bacteria shared the more homologous the two bacteria are.
The MAUVE was employed to compare the genome organisation across the genomes based on
the identification of ‘conserved gene regions’ visualized through ‘Locally Collinear Blocks’
(LCBs) shared between genomes [Figure 2.5B]. In MAUVE figures, two regions on two
genomes are shown as conserved when the LCB of them are the same in colour and are linked
by a thin coloured line. Therefore, the lesser numbers of LCB visualized the more homologous
the two genomes are. In this study, the draft genomes of marine isolates and completed genome
of terrestrial neighbour isolates were submitted to MAUVE software and the progressive-
Mauve mode of analysis was used. The software automatically reorders all contigs of draft
genome based on the gene composition of the completed genome, followed by pair-wise
alignment. The outcome was the organisation of multiple conserved colour blocks on two
genomes [Figure 2.5 B].

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
36
Figure 2.5. Schematic representation of pairwise genome comparison performed by the
“Sequence-Based Comparison Tool” (A) and MAUVE (B). (A) The output is a schematic of
chromosomes in which CDS are coloured based on the similarity in sequence. (B) Performed
by MAUVE. The output is an organisation of conserved regions on each genome. White colour
within LCB indicates genes loss/insertion caused by mobile DNA elements. The figures were
computed from example genomes provided.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
37
2.13. In silico prediction of putative antimicrobial peptides within genomes of marine
Bacillus species
Bacillus species isolates frequently produce antimicrobial peptides (including bacteriocins and
non-ribosomally synthesized antimicrobial peptides) (21, 36); therefore, genes encoding these
kinds of antimicrobial peptides were sought within the selected marine isolates’ genomes by
using two bioinformatic tools; web-based ‘BActeriocin GEnome mining tooL’ (BAGEL) and
web-based the ‘antibiotics and secondary metabolite analysis shell’ (AntiSMASH) (108, 109).
The BAGEL software is the tool of choice to predict putative bacteriocins because it can detect
the small ORFs coding precursors of bacteriocins. These precursors are frequently omitted by
similar software, because of their small size. Draft genomes (in fasta format) of marine isolates
were submitted to the web-based BAGEL tool (http://bagels.molgenrug.nl/) and predictions
were computed. The outcome of the BAGEL prediction was genomic ‘areas of interest’ (AOIs)
that comprises both multiple ORFs which may or may not be indicative of bacteriocin
biosynthesis [Figure 2.6 A]. In this study, manual annotation was subsequently performed to
determine the actual size of bacteriocin biosynthetic gene cluster (BBGC) based on; (i) presence
of proteins required for bacteriocin biosynthesis, including precursor bacteriocin, transporters,
modification enzymes, regulator and immunity proteins; (ii) the biosynthetic gene clusters of
bacteriocins are usually flanked by a promoter and a terminator; (iii) the gene organization of
known similar bacteriocin gene clusters. All ORFs suggested by BAGEL were used as queries
sequences to search against protein databases to attempt to predict function. The protein
databases used included ’uniref 90’ database’, ‘non-redundant’ database (NR) and
BACTIBASE database (for precursor CDS) (110). The BACTIBASE database is a bacteriocin
database which contains large numbers of characterised bacteriocins. Also, SignalP (v5.0) was
employed to predict the cleavage site within the precursor sequence, subsequently resulting in
the prediction of ‘signal sequence’ and ‘core sequence’ (111). The MW of mature bacteriocins
were predicted based on the core sequence using Expasy tool (at
https://web.expasy.org/compute_pi/) (112). The prediction of functional genes required for
bacteriocin processing, transport and immunity were carried out using other software, such as
the TMHMM server (v.3.0) (http://www.cbs.dtu.dk/services/TMHMM/) to predict helical
motifs that are always present in transporters and possibly in immunity proteins (113). The
whole gene cluster encoding bacteriocin production was also predicted based on the evidence
of the flanking promoters and terminators, which were also predicted by BAGEL version 4.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
38
The AntiSMASH is a tool to identify non-ribosomal peptides (NRPs) (lipopeptide, polyketide,
hybrids and bacilysin). The tool predicts NRPs compounds via detection of highly conserved
domains of synthase enzymes. In practice, the draft genomes of marine isolates were submitted
to AntiSMASH (at https://AntiSMASH.secondarymetabolites.org/#!/start). The outcome of
AntiSMASH prediction was predicted genomic ‘areas of interest’ (AOIs) encoding production
of NRPs and also bacteriocins, which frequently comprised ORFs with requirement/non-
requirement for biosynthesis [Figure 2.6 B].
Figure 2.6. Schematic representation of in silico prediction of antimicrobial peptides
performed by BAGEL and AntiSMASH tools. (A) The initial the genomic ‘areas of
interest’ containing the bacteriocin gene cluster was determined by the BAGEL. A
precursor of bacteriocin (green), enzymes for posttranslational modification of
bacteriocin (blue). (B) Example of a predicted non-ribosomally- synthesized
antimicrobial peptide encoding region predicted by AntiSMASH. Figure constructed
from the predictions using example genomes provided by the tools.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
39
2.14. Refinement of media and growth conditions for enhanced bacteriocin production
Growth curves of bacterial isolates in medium were investigated in association with quantitation
of antimicrobial activity. To determine the kinetics of bacterial growth, a starter culture was
first prepared by inoculating a single colony into 10 mL of selected broth, incubated at 30oC
with a shaking speed of 200 rpm overnight. One mL of this starter culture was inoculated into
100 mL of fresh media, grown at 30oC with a shaking speed of 150 rpm to produce the
antimicrobial compounds. Five mL of culture was removed every 6 hours, up to 36 hours, and
OD600 and pH measurements were taken before being centrifuged at 4,500 g for 20 minutes.
The supernatant was filtered through a 0.45 µm PES membrane to obtain cell-free supernatant.
These cell-free supernatants were utilised in a well-diffusion assay, or spot-on-lawn assay to
detect antimicrobial activity. This analysis was to determine the point in the growth curve at
which the bacteriocin production was maximal.
2.15. Recovery of antimicrobial compounds after fermentation
The antimicrobial compounds not only presented in the bacterial culture but also associated
with the bacterial cell surface. The antimicrobial substances from the bacterial culture
supernatant were recovered by either precipitation using ammonium sulphate or by an
absorbance/elution method using Diaion HP-20 resin. The antimicrobial compounds associated
with the cell surface were recovered by extraction in a polar solvent.
2.15.1. Recovery of antimicrobial peptides from the cell-free culture supernatant by
precipitation using ammonium sulphate
Bacteriocins, being proteinaceous, could be precipitated with the addition of ammonium
sulphate (AMS) to the culture supernatant (114). In principle, a protein’s solubility increases in
the presence of low concentrations of salt (<0.15 M), and decreases in high salt concentrations,
and can precipitate when the salt reaches its saturation concentration. AMS is commonly used
as a salt to precipitate proteins from a liquid suspension. Different proteins can be precipitated
at different saturation concentrations of AMS; therefore, the AMS can be used to partially
purify a protein of interest by judicious selection of AMS concentrations.
In practice, to determine the AMS concentration used to precipitate a bacteriocin, different
amounts of AMS was add to 100 mL samples of cell-free culture supernatant to a final
concentration of AMS of 30%, 40%, 50%, 60%, 70% and 80% at 5oC. The culture supernatants
with added AMS were stirred to dissolve and left at 5oC overnight without shaking. The
mixtures were centrifuged at 17,500 g for 90 minutes and the supernatants were gently

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
40
decanted. The pellets were dissolved in 2mL of 1X PBS pH7 and 100 µL of dissolved protein
was used in a well-diffusion assay against indicator bacteria of interest. The AMS concentration
at which the precipitated protein exhibited the strongest antimicrobial activity was selected for
large scale precipitation. After precipitation, the dissolved sample was desalted using either a
dialysis membrane (Sigma Aldrich) or centrifugal filter (Millipore) with a protein size cut off
(2 kDa), before ion-exchange chromatography.
2.15.2. Recovery of antimicrobial compounds from cell-free culture supernatants using
Diaion HP-20
The precipitation of bacteriocins with ammonium sulphate followed by dialysis was a time-
consuming process, requiring at least 2 days. Therefore, bacteriocins present in a liquid sample
were also recovered using absorption to Diaion HP-20 resin, as an alternative approach. This
resin is a non-ionic macro reticular; therefore, it can adsorb the amphiphilic antimicrobial
compounds like bacteriocins and NRPs through i) hydrophobic interaction, ii) polar interaction,
iii) pi-pi bonding, and iv) hydrogen bonding. Bacteriocins can be eluted in the presence of polar
solvents (methanol, acetonitrile, and isopropanol).
In practice, the bacterial culture was first adjusted to a pH of 6.5. The Diaion HP-20 resin was
washed with 99.9% HPLC-grade methanol, followed by washing with milli-Q water to
eliminate the organic solvent. Approximate 2% (w/v) of this conditioned resin was added to
cell-free culture supernatant and gently stirred overnight at 5oC. The resin was collected in a
fresh 50mL syringe, washed with milli-Q water (3CV) then with 30% methanol (3CV) and
eluted with 50% methanol (3CV), followed by 70% methanol (3CV). All three elutes which
came from the same methanol concentration were combined. Two elutes (at 50% methanol, and
70% methanol) were concentrated by using speed Vac to eliminate the solvent and used in a
well-diffusion assay or spot-on-lawn assay to determine which elutes exhibiting antimicrobial
activity. After identifying the concentration of methanol to elute the antimicrobial compounds,
this methanol concentration was used in large scale purification.
2.15.3. Recovery of cell associated antimicrobial compounds from cell pellets by acidic
solvent extraction
Antimicrobial peptides are found not only in the bacterial culture supernatant but also
associated with the bacterial cell pellet. This association with cell pellet, probably cell surface,
is based on the ion bonding between positively charged bacteriocins (or NRPs) and negatively
charged bacterial cells; therefore, an organic solvent could be used to release the antimicrobial

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
41
substances. This step facilitates latter chromatography to purify the antimicrobial substance by
selectively release the antimicrobial compound while remaining most other proteins in the cell
pellet.
In practice, the cell pellets was stirred in 1/20 (v/v) 70% isopropanol pH 2 (using concentrated
HCl to adjust the pH) at 5oC for at least 4 hours. Subsequently, the mixture was centrifuged at
4,500 g for 20 minutes, and the supernatant was then filtered through a syringe membrane
(0.45µm) to obtain a cell-free solvent fraction. The cell-free solvent was concentrated in a rotary
evaporator to 1/100 volume to remove the solvent and adjusted to pH 7 with NaOH (named the
CFS fraction). This fraction was desalted before use in ion exchange chromatography.
2.16. Ion exchange chromatography for partial purification of antimicrobial compounds
Ion exchange chromatography is a separation technique based on the net surface charge of a
protein. It is distinguished into ‘cation exchange chromatography’ and ‘anion exchange
chromatography’. Cation exchange chromatography uses negatively charged resins (SP
sepharose, CM sepharose) which can capture positively charged proteins; while anion exchange
chromatography employs positively charged resins (Q sepharose, DEAE sepharose) to bind to
negatively charged proteins. The protein’s charge varies depending on the protein’s isoelectric
point (pI) and the pH of the buffer that is used to dissolve the protein. The protein’s pI value is
calculated based on amino acid composition. Thus, protein has no charge when dissolved in a
buffer with the same pH as the protein’s pI, or a positive charge when dissolved in a buffer with
a pH higher than the pI value. The proteins (or peptides) that bind to the resin can be released
with elution buffer containing salt (NaCl is commonly used). For an unknown bacteriocin, it is
necessary to select the resin types, running buffer, and elution buffer for a successful separation.
2.16.1. Identification of running buffer and types of resin for ion exchange
chromatography purification of antimicrobial compounds
In practice, five sterile 2 mL-eppendorf tubes were prepared, to 4 of them 500 µL of SP
sepharose added and the fifth tube had 500 µL of Q sepharose. All resins were washed 3 times
with milli-Q water by adding of 1 mL milli-Q water, inverting tubes for 5 minutes, spinning
down, and decanting the milli-Q water. This step aimed to eliminate the ethanol which was used
to store the resin. Next, 1 mL of each of four buffers; 20 mM acetate buffers pH 3.6; 20 mM
acetate buffers pH 4.6; 20 mM acetate buffer pH 5.6; 20 mM phosphate buffer pH 7.0 were
respectively added to one of the tubes containing SP sepharose resin.While 1 mL of 20 mM
Tris HCL pH 8.0 was added to the tube containing Q sepharose. The tubes were inverted for 5

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
42
minutes, spun down and the buffer was discarded. These steps were replicated 3 times to
equilibrate the resin. One mL of each buffer was added to the corresponding tubes. One hundred
µL of dissolved protein (well-desalted) were aliquoted into the tubes and mixed overnight at
5oC. Next, the tubes were spun down and the supernatants collected (named as GT fraction).
The resins were washed (three times) by adding 1 mL of the same buffer (3 times), inverting
tubes for 5 minutes, and decanting the buffers. One mL of elution buffer (same buffer + 1M
NaCl) wase add to each tube, inverted tubes for 20 minutes, and spun down to collect the
supernatants (EL fraction). One hundred µL of GT fractions and EL fractions were used in a
well-diffusion assay against indicators of interest to detect the presence of antimicrobial
compounds. The most effective resin and buffer combinations were selected for large scale ion
exchange chromatography, based on the antimicrobial activity being concentrated in the EL
fractions.
2.16.2. Determination of the NaCl concentration used in elution buffer
Cation exchange chromatography was performed using the running buffer and resin identified
from the above experiment. The operation of cation exchange included 4 steps, i) equilibrating
resin (3CV) with running buffer, ii) loading the sample onto the resin, iii) washing the resin
with the running buffer (3CV), and iv) eluting the bound antimicrobial compounds with elution
buffer (CV). To determine the concentration of NaCl required for the elution buffer the elution
steps used the elution buffer supplemented with various concentration of NaCl (0.1 M, 0.2 M,
0.3 M, 0.4 M, 0.5 M and 1.0 M). All these elutes were collected and utilized in a well-diffusion
assay against indicators of interest. The elute exhibiting the highest antimicrobial activity with
the lowest concentration of NaCl was selected for large scale ion exchange chromatography.
2.17. Hydrophobic interaction chromatography for partial purification of antimicrobial
compounds
Hydrophobic interaction chromatography is a separation technique based on bacteriocin’s
hydrophobicity. This method employs a resin that is an immobilized hydrophobic ligand. This
ligand can capture hydrophobic compounds. The compounds can be released in the presence of
organic solvents. Therefore, the elution commonly utilises a linear concentration gradient of
organic solvent or a step-gradient process.
In this study hydrophobic interaction chromatography was carried out using a Sep-Pak C18
column (Waters). The Sep-Pak column was first conditioned by equilibrating the resin (3CV)
with 100% acetonitrile (+0.01% TFA) followed by washing (3CV) with 2% acetonitrile in water

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
43
(+0.01%TFA) to reduce the acetonitrile. Subsequently, the sample was loaded into the column
drop-by-drop and washed (3CV) with 2% acetonitrile in water (+0.01%TFA) to remove the
unbound protein. To determine the concentration of acetonitrile to elute the antimicrobial
compounds, the resin was eluted respectively with different concentration of acetonitrile (10%,
20%, 30%, 40%, 50%, 60%, 70%, 80% and 90%). Each concentration was used with three
column volumes. All elutes were collected, lyophilized, dissolved in milli-Q water and used to
detect the presence of antimicrobial compounds.
2.18. Reverse phase high-pressure liquid chromatography (RP-HPLC) to obtain pure
antimicrobial compounds
HPLC using a reverse-phase column (C4-C18) is used to separate compounds based on the
protein’s hydrophobicity. The sample is injected into the RP-HPLC device, mobile phase (a
mixture of two solvents) carries the sample through the column where the hydrophobic
components in the sample are captured by the ligands immobilized on the matrix. Depending
on the hydrophobicity, these compounds are washed out of the column at different retention
time (RT), corresponding to different concentration of organic solvent. The elution can be
conducted under isocratic conditions or a gradient of organic solvent concentrations. During
operation, the elution of antimicrobial peptide can be visualized by absorbance readings at 280
nm, 230 nm or 214 nm. This chromatography can be used to obtain compounds at high purity
for peptide sequencing.
In this study, RP-HPLC was performed using a Zobax 300SB C18 column (Agilent
Technology).The mobile phase were mixtures of two solutions including solution A (milli-Q
water + 0.1% TFA) and solution B (acetonitrile + 0.1% TFA). The running parameters were
modified after consideration of the compounds’ physicochemical properties. Elutes were
collected every two minutes or based on the peak visualized on the monitor. All elutes were
lyophilized and the antimicrobial activity was evaluated. Some cycles of RP-HPLC were
performed until the elute fractions containing 1 peak visualized on the monitors, and 1 peak
visualized by MALDI-TOF MS.
2.19. Determination of antimicrobial purity by Matrix-Assisted Laser
Desorption/Ionization time-of-flight mass spectrometry (MALDI-TOF MS)
MALDI-TOF MS can be used to detect the presence of antimicrobial peptides in samples,
estimate the purity of antimicrobial samples, and can be utilised for protein identification (115).
The MALDI-TOF MS device consists of a “matrix-assisted laser desorption/ionization

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
44
(MALDI)” part, a “time-of-flight (TOF)” part containing a mass analyser detector, and a
monitor. A sample is dropped onto a metal plate, allowed to dry and then loaded into the
machine, the MALDI part shoots an ultraviolet laser beam at the sample spot to convert all
molecules present into ions. These ions vapourize into the TOF part where they are separated
according to their velocity under an electrostatic field, due to the charge and mass. The ions are
detected by a mass analyser detector which converts the ions into electrical signals through the
value of mass-to-charge (m/z). These signals are then transmitted to a computer for calculation
[Figure 2.7]. The m/z value of a protein is also the MW of that protein in Dalton.
Figure 2.7. The process of MALDI-TOF mass spectrometry.The sample is mixed with
the matrix which is commonly α-Cyano-4-hydroxycinnamic acid matrix (HACCA). The
mixture is dropped onto the metal plate, dried and the plate is load into the device. The
MALDI part shoots the laser beam to the dry spot of the sample to generate multiple ions
of charged molecules. These ions are separated in the TOF tube and detected by an MS
detector, which is analysed to generate the MS profile. The figure is from Clark et al.
(2013) (116).
In practice, 2 µL of both antimicrobial sample and protein standards were mixed separately
with 2 µL saturated α-Cyano-4-hydroxycinnamic acid matrix (HACCA). In case of
bacteriocins, two types of protein standards kit were selected including “Peptide calibration
standard II” (Burker) which included known peptides with a mass range of ~700 Da – 3,200
Da; and “Protein calibration standard I” (Burker) which consisted of known peptides covering
a mass range of ~4,000 Da - 20,000 Da. Next, 1 µL of each mixture was dropped onto a metal
plate at two different spots and dried at room temperature. This metal plate with sample spots
was load into the Bruker AutoFlex MALDI-TOF MS (Bruker). The operation started with a
calibration process on the protein standard first, followed by the measurement of the sample.
The calibration process was performed by shooting the UV beam at the standard spot multiple
times to obtain the mass range for calculation. After calibration, the sample was measured by

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
45
shooting at the sample spot until clear protein peaks were obtained. The spectrometry of the
protein sample was visualized through the monitor.
2.20. Peptide sequencing techniques
The bacteriocin consisted of 20 amino acid residues and was sequenced by de novo peptide
sequencing or N-terminal sequencing methods.
2.20.1. Peptide sequencing by de novo peptide sequencing
The HPLC purified bacteriocin samples were first fragmented by proteolytic digestion
(trypsin), which cleaves exclusively at arginine and lysine residues in the amino acid chain.
After desalting to remove residues of enzyme and salt, the pure fragmented bacteriocins were
load into a mass spectrometer, an LC-MS/MS instrument, to obtain the m/z value of all
fragments. The m/z values are searched against the Mascot database
(http://www.matrixscience.com/) to determine the amino acid sequence of each of the
fragments. The method is highly effective for known bacteriocins when the multiple digested
bacteriocins fragments have been assigned into the MASCOT database (117).
2.20.2. Peptide sequencing by N-terminal sequencing (Edman degradation method)
The method is frequently used to detect the amino acid sequence of novel bacteriocins by
identifying each amino acid residue, starting from N-terminus (118). In principle, the pure
peptide is mixed with phenylisothiocyanate (PITC) to form a complex of phenylthiohydantoin
(PTH)-bacteriocin. This mixture is loaded onto the device. Each cycle of the operation removes
an amino acid at the N-terminus until the sequence is fully identified. In this study, the N-
terminal sequencing technique was employed to identify the amino acid residues of
antimicrobial peptides using a Perkin-Elmer Precise Sequencer (Perkin-Elmer).
2.21. Estimation of bacteriocin size by Tricine SDS-PAGE
Tricine SDS-PAGE was used to estimate the bacteriocin size (119). In this study, the
antimicrobial samples were separated on a 16% Tris-Tricine gel. The gel consisted of a 16%
resolving gel layer at the bottom and 4% stacking gel layer at the top (described in table 2.3).
ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
46
Table 2.2. Electrode and gel buffers for Tricine–SDS-PAGE, followed the protocol
suggested by Schagger H, 2006 (119)
Gel components Anode
buffer (10×)
Cathode buffer
(10×)
Gel buffer
(3×)
Tris (M) 1.0 1.0 3.0
Tricine (M) - 1.0 -
HCl (M) 0.225 - 1.0
SDS (%) - 1.0 0.3
pH 8.9 ~8.25 8.45
Table 2.3. The composition of acrylamide mixtures used to cast the Tricine SDSD gel,
followed the protocol suggested by Schagger H, 2006 (119)
Gel component Stacking gel
(mL)
16% resolving gel
(mL)
dH2O 1.10 1.56
40% (w/v) acrylamide mix 0.27 4.00
3X Gel buffer 0.62 3.33
Glycerol (g) - 1.06
10% (w/v) APS 0.01 0.05
TEMED 0.002 0.005
Samples were prepared by adding 32 µL of antimicrobial sample to 8 µL of 4X tricine sample
buffer (Bio-Rad) to give a total of 40 µL of a mixture and boiled at 100 °C for 5 min. Then, 15
µL of the mixture was loaded into two separated wells on a16% Tris-Tricine gel, along with
Dual Xtra Precision Plus Protein™ Prestained Standards (Bio-Rad). The running buffers used
a cathode buffer and an anode buffer (described in table 2.2). The running condition was set at
80 V for 15 minutes to allow the sample in the stacking gel layer to completely migrate into the
resolving gel layer and then increased to 100 V and run for 3.5 hours. The gel was cut into two
halves. One half was stained with Coomassie Blue staining solution or Silver staining solution
(if protein stained by Coomassie Blue was faint), followed by destaining in “destaining buffer”
to visualize the protein bands; the other half of the gel was used to conduct a zymogram assay.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
47
Size and location of bacteriocin on the gel was estimated from results of both the stained gel
and zymogram assay.
2.22. Zymogram assay
Half of the Tris-Tricine gel was first fixed in ‘fixing solution’ for 2 hours, was then washed
with milli-Q water for 4 hours (with 4 water changes) and placed on soft media agar (0.75%
agar). This medium agar was initially prepared by mixing 15 mL of molten media agar (at 50
°C and 0.75% agar) with 200μL of an overnight culture of a bacterial indicator, pouring into
the plate to solidify. When the gel was placed on this agar, another 15 mL of seeded molten soft
agar was poured to overlay the gel and allowed to solidify. The plate was incubated at optimal
conditions for the bacterial indicator. Bacteriocin size was then identified by comparing the
position of the killing zone that appeared on zymogram half gel to the stained half gel in
association with the protein ladder (120).
2.23. Characterisation of the physicochemical properties of antimicrobial compounds
To determine enzymatic degradation susceptibility of the antimicrobial compounds, 50 µL of a
sample (or culture) were mixed with various enzymes (proteinase K, pronase E, trypsin,
catalase, and lipase) used at a final concentration of 2 µg/L. The mixtures were incubated at
37oC for 2 hours, followed by heating at 80oC for 5 minutes to inactivate the enzymes. The
antimicrobial activity of the treated samples was evaluated using a well-diffusion assay or spot-
on-lawn assay against indicators of interest. Thermal stability was determined by incubating
purified peptide or culture at various temperatures (40oC, 50oC, 60oC, 80oC, 90oCand 100oC)
for 1 hour, followed by cooling on ice before being used in spot-on-lawn or a well-diffusion
assay.
2.24. Overview of heterologous cloning and expression of bacteriocin
The heterologous cloning and expression employed an E. coli/Bacillus shuttle plasmid,
pDLL202, as the carrier of a whole bacteriocin gene cluster, and B. subtilis BS54A as host for
expression of bacteriocins. This customized plasmid and the bacterium was provided by
Monash University that is designed for cloning a large nucleotide fragment and expression the
gene of interest. The plasmid pDLL202 contains two compatible origins of replication
(colE1rep and repA) enabling replication in both E. coli and B. subtilis hosts, four multi-
cloning-sites (MCS) which include recognition sites of various restriction enzymes, and
antibiotic resistance genes (kanR, ermB, catP, and ampR) encoding resistance to kanamycin,
erythromycin, chloramphenicol and ampicillin. During cloning and transformation, E. coli

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
48
DH5α cells were used to maintain the native pDLL2002 plasmid. E. coli Top10 served as host
to control the plasmid copy number, E. coli JIR702 served as a host to induce a multimeric
fusion plasmid, described in Table 2.4. The B. subtilis BS54A strain was incubated in Trypton
soy agar (TSA) or broth (TSB) at 30°C while the E. coli strains were propagated aerobically in
Luria−Bertani broth at 37°C. Chloramphenicol was used as a selective antibiotic at a final
concentration of 5 μg/mL for B. subtilis and 30 μg/mL for E. coli strains, while kanamycin was
used at a concentration of 50 μg/mL.
Table 2.4. List of strains and plasmids used for cloning /expression
Strains or plasmids Relevant properties Origins
Strains
E. coli DH5α host for copy control pD202L vectors RMIT university
E. coli Top10 host for copy control pD202L/ insert vectors RMIT university
P. polymyxa #23 Wild type sactipeptide producing strain This study
E. coli JIR702 Host for induction multimeric DNA Monash university
B. subtilis BS34A Host for heterologous expression of sactipeptide Monash university
MRSA indicator strain for an antimicrobial test, Cm5R RMIT university
Plasmids
pDLL202 E. coli and B. subtilis shuttle cloning vector; pCC1BAC origin and pAMbeta1 origin, EmR, CmR, AmpR (10,153 bp)
Monash university
pDLL202/G23sac Fusion plasmid harbouring whole sactipeptide gene cluster (15,479 bp)
This study
2.25. Extraction of the plasmids from E. coli host
To extract the plasmid (pDLL202 and the fusion plasmid), the E. coli host (DH5α, Top10,
JIM702) was sub-cultured in 10mL of selective LB broth. Of that culture,, 2mL was centrifuged
at 5,000 g for 20 minutes and the pellet was collected to extract the plasmid. The plasmid was

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
49
extracted using the Monarch® Plasmid Miniprep Kit (New England Biolabs). The procedure
followed the manufacturers’ instructions.
2.26. Preparation of the plasmid backbone by enzymatic digestion
The native plasmid pDLL202 was digested by MluI-HF and SpeI-HF (New England Biolabs).
The mixture was then loaded onto a 1% agarose gel and electrophoresed for 40 minutes at 100V.
The band of the plasmid backbone was identified based on the size, excised and purified using
the Monarch® DNA Gel Extraction Kit (NEB).
2.27. Preparation of the whole sactipeptide genome cluster by PCR amplification.
The P. polymyxa #23 genomic DNA was extracted by the GES method and used as a template
in PCR. The whole sactipeptide gene cluster was amplified from P. polymyxa #23’s genome
using the designed primers. Due to the requirement of many optimization steps, the details of
PCR reaction was mentioned in chapter 7.
2.28. Preparation of E. coli competent cell.
To prepare the E. coli competent cells (Top 10, JIR702) for electroporation, a starter culture
was grown by inoculating a single fresh bacterial colony into 10 mL of LB and incubated at
37oC overnight at 200 rpm. All the starter culture was inoculated into 1L of fresh LB media and
incubated at 37oC with shaking until the OD600 reached ~0.35-0.4. The culture was centrifuged
at 1,000 g for 20 minutes at 4oC to collect the bacterial pellet. The pellet was then washed twice
with 250 mL of cold milli-Q water followed with two washes with 100 mL of cold 10% glycerol
before resuspending in 2 mL of 10% glycerol (OD600~200-250). The bacterial suspension (100
µL) was aliquoted to prechilled 1.5 mL Eppendorf tubes and stored at -80 oC.
2.29.Transformation of the fusion plasmid into E. coli by electroporation
The transformation of the fusion plasmid into E. coli (Top10/ JIM702) was carried out by
electroporation. Briefly, the competent E. coli cells (100 μl) were mixed with the given plasmid
vector and held on ice for 5 min. The mixture was transferred to a 0.2 cm electroporation cuvette
(Bio-Rad). After pulsing under specific conditions (field strength 2.1 kV/cm, capacitance 25
μF, resistance 100 Ω), the cell suspension was immediately diluted with 1 ml of pre-warmed
SOC media (see the appendix for recipe), then incubated at 37oC for 1 hour with shaking speed
of 250 rpm. A portion of the cell suspension (100 μl) was placed on selective LB and incubated
at 37°C overnight. The colonies which formed on the plates were patched onto a fresh selective
plate for subsequent confirmation.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
50
2.30. Preparation of Bacillus competent cells
The multimeric fusion plasmid was transformed into B. subtilis BS34A via natural
transformation. To prepare the competent Bacillus subtilis BS34A cells, a fresh Bacillus colony
on a TSA plate was suspended in 10 mL of spC media in a 50 mL falcon tube to a final OD600
of~0.2 the suspension was incubated at 37oC at 275-300 rpm until cells reached early stationary
phase. The best indicator for reaching this stage is that the OD increases less than 20% in 30
minutes. Then 2 mL of the culture was diluted by adding it to 10 mL of SpT medium (see the
appendix for recipe) in a fresh flask and incubating at 37oC for 70-90 minutes at 250 rpm and
1 mL of the SpT culture was transferred to a fresh epperdorf tube.
2.31. Transformation of the fusion plasmid into Bacillus subtilis BS34A
To transform B. subtilis, 1-3 μL of plasmid was added to a tube containing the suspension of
Bacillus competent cells. One to three uL of sterile milli-Q water was added to another tube as
a negative control. These tubes were then shaken at 37oC for 30 minutes at 200 rpm. The cells
were harvested by low-speed centrifugation at 4,500 x g for 10 minutes, resuspended in 1-2 mL
of LB broth and incubated with shaking at 37oC for 90 minutes. The bacterial pellet was
centrifuged, resuspended in 0.5 mL of LB, and 0.1 mL of samples were plated onto Tryptone
soya agar (TSA) (Oxoid) plates supplemented with chloramphenicol to the final concentration
of 5 µg/mL. Presence of fusion plasmid in the transformant was confirmed by PCR of the gene
encoding the structural gene from the bacteriocin gene cluster, and by sequencing of the PCR
product.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
51
Chapter 3 BROAD SPECTRUM ANTIMICROBIAL ACTIVITIES OF SPORE-
FORMING BACTERIA FROM THE VIETNAM SEA
3.1. Introduction
The emergence and spread of bacterial pathogens resistant to last-line antibiotics, for example,
carbapenem-resistant Gram-negative pathogens, methicillin-resistant Staphylococcus aureus
(MRSA), and vancomycin-resistant Enterococci (VRE), has been of great concern in both
human and veterinary medicine. Antibiotic resistance genes are frequently located on mobile
elements such as conjugative plasmids and transposons which facilitate horizontal and vertical
transmission, leading to increasing numbers of multi-resistant bacteria worldwide. The high
incidence of antibiotic-resistant bacteria has many implications for pathogen control and there
is an urgent need for alternatives to currently available antibiotics to re-control these life-
threating antibiotic-resistant pathogens. The discovery of novel antibiotics from terrestrial
environments over the last few decades has been challenging, due to exhaustion of these
traditional antibiotic sources. According to the FDA, approval of new medically important
antibiotics had decreased by 56% over the last few decades and there has been no evidence for
an increase in discovery rate since that study (12).
The marine environment has been identified as a promising alternative source for antibiotic
discovery, due to the high abundance of antibiotics produced by various members of diverse
marine microbial communities. It has been hypothesized that marine bacteria are under
unusually rigorous selection pressures because of the environmental conditions with which they
must contend. They are typically exposed to low levels of nutrition and rapid changes in
nutrition and physical conditions due to wave and tidal action. These harsh chemo-physical
conditions have selected bacteria that can employ adaptations to out-compete other bacteria.
Diversification of antimicrobial production is one such adaptation that can be essential for
occupying and defending an ecological niche. It is hypothesized that marine environments may
harbour species producing novel antimicrobial compounds which could be effective against a
range of bacteria, including antibiotic-resistant bacteria. Among the diverse marine bacterial
communities, members of the Bacillus genus characterised as spore formers, have been shown
to produce an array of structurally diverse antibiotics, including ribosomally synthesized
peptides (bacteriocins), non-ribosomal secondary metabolites and bacilysin. Bacteriocins are of
interest. These small cationic peptides commonly have a narrow spectrum of activity, affecting
a limited range of bacteria and a low resistance rate; making them attractive antimicrobials for
application against bacteria that have acquired resistance to the current range of available

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
52
antibiotics. Many terrestrially derived bacteriocins have been characterised, but little is known
of marine-derived bacteriocins. Vietnam covers more than 1 million square kilometres of
coastal areas, incorporating a variety of tropical marine ecosystems with abundant marine
species. These characteristics indicate that the Vietnam Sea is a promising source to explore for
novel antimicrobials.
The objective of this study was to evaluate a diverse collection of spore-forming bacteria from
the Vietnam Sea for antimicrobial molecules with activity against a range of important
pathogens, including multi-antibiotic-resistant pathogens. The bacterial screening was
restricted to spore-forming bacteria because such bacteria have previously proven to be a richer
source of antimicrobials than general bacterial populations, and facilitates for antibiotic
production at the industrial level (121, 122).
3.2. Results
3.2.1. Thermally resistant, spore-forming, bacteria derived from the marine
environment display antimicrobial activity against important pathogens
A total of 389 heat resistant, spore-forming, bacterial isolates were cultured from 50 marine
samples. They demonstrated a range of colony morphologies (Figure 3.1, Table 3.1).
Table 3.1. Antimicrobial producing bacteria identified from marine samples
Number of marine samples collected
Number of isolated spore-forming bacteria
Number of isolates with antimicrobial activity
Percentage of antimicrobial isolates *
Sponges 16 183 28 71.97%
Seaweeds 13 92 15 3.86%
Sediments 13 81 16 4.11%
Seawaters 8 33 6 1.54%
Total 50 389 64 16.45%
* Percentage of antimicrobial isolates was calculated based on the number of isolates
with antimicrobial activity and total number of isolated spore-forming bacteria

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
53
Figure 3.1. Diversity in the morphology of the marine isolates isolated from NhaTrang
bay, Vietnam Sea.(A) Examples of three of the four kinds of samples collected for
bacterial isolation; sponge, seaweed, sediment. (B) Colony morphologies of some pure
isolates. (C) Plate with colonies isolated from seaweed.
A total of 389 heat resistant, spore-forming, bacterial isolates, were cultured from 50 marine
samples (Table 3.1). They demonstrated a range of colony morphologies (Figure 3.1). Primary
screening against six indicator strains identified 64 isolates (16.4%) with antimicrobial activity
(Figure 3.2.B). The proportion of isolates that exhibited activity against Gram-positive indicator
strains, B. cereus (93.7%), S. aureus (84.3%), and S. faecalis (87.5%), was higher than the
proportion that exhibited activity against the Gram-negative indicators, E. coli (50%) and P.
aeruginosa (4.6%), and the yeast, C. albicans (21.4%).
The spectra of antimicrobial activities, exhibited by a select group of the 23 most potent isolates
from the primary screen, were determined against an expanded panel of 14 indicator strains,
including important multidrug-resistant pathogens, were performed using two assays; a cross-

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
54
streak assay and a well-diffusion assay. There was considerable variation in the strength and
spectra of antimicrobial activity across the 23 short-listed isolates. The pathogenic indicator
strains most commonly affected by the antimicrobial compounds expressed by the screened
isolates were the Gram-positive species (Table 3.2). Of the bacterial indicators, C. perfringens,
B. cereus and S. aureus were inhibited by 83% (19/23) of the test isolates, whereas the
proportion with activity against the Gram-negative indicators; Campylobacter jejuni, 70%
(16/23), Campylobacter coli, 61% (14/23), was lower. Two antibiotic-resistant Gram-positive
pathogens; MRSA and VRE were inhibited by respectively 83% (19/23) and 57% (13/23) of
isolates, while an antibiotic-resistant Gram-negative pathogen, multidrug-resistant Klebsiella
pneumonia (MRKP), was inhibited by only one isolate, P. polymyxa #23. The Gram-negative
bacteria P.aeruginosa was also inhibited by P. polymyxa #23. The growth of food-borne
pathogens; L. monocytogenes, E. coli and Salmonella Enteritidis were depressed by,
respectively, 78%, 44%, and 57% of the isolates. None of the selected isolates had inhibitory
activity against C. albicans.
The cross-streak assay was found to be more sensitive than the well-diffusion assay. In most
cases, antimicrobial activity was detected in both assays but occasionally the activity was less
or absent in the well-diffusion assay. This difference was particularly apparent when S. aureus,
MRSA and most of the Gram-negative pathogens; C. jejuni, C. coli, E. coli and S. Enteritidis.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
55
Table 3.2. Antimicrobial activity of short-listed isolates against indicators strains. The antimicrobial activities were evaluated by well diffusion
assay (WDA) and cross streak assay (CSA) methods
#01 #02 #03 #04 #05 #06 #07 #08 #09 #10 #11 #12
CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CS WD
C. perfringens ++ ++ ++ + - - + + ++ +++ ++ ++ - - +++
++ - - + + +++ ++ ++ +
S. aureus +++ ++ ++ - + - - - + + +++ - - - +++ - - - ++ ++ ++ - ++ -
B. cereus ++ ++ ++ ++ - - + - + ++ +++ ++ - - +++ +++ ++ - + + +++ ++ + +
L. monocytogenes ++ ++ ++ + - - + - - - +++ ++ + - ++ + - - - - ++ + - -
L. plantarum ++ + + + + + - - - - ++ ++ - - ++ ++ - - + + ++ ++ - -
C. albicans - - - - - - - - - - - - - - - - - - - - - - - -
E. coli + - + - - - - - + - - - - - + + - - - - + + + -
S. Enteritidis + - - - - -
- - - ++ - + - ++ + - - - - ++ + + -
C. jejuni ++ + ++ + - - - - ++ - ++ ++ + - ++ ++ - - - - +++ ++ - -
C. coli + +++ ++ ++ - - ++ +++
+
+ - + +++ + - + +++
+
- - - - ++ +++ ++ ++
P. aeruginosa - - - - - - - - + - - - - - - - - - - - - - - -
MRSA +++ ++ ++ - + - - - + + +++ - - - +++ - - - ++ ++ ++ - ++ -
VRE ++ ++ ++ ++ - - + + - - + - - - +++ +++ - - ++ ++ ++ ++ + ++
MRKP - - - - - - - - - - - - - - - - - - - - - -

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
56
#13 #14 #15 #16 #17 #18 #19 #20 #21 #22 #23 Percent (%)
CSA WDA CSA WDA CSA WDA CSA WDA CSA WD CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA CSA WDA
C. perfringens +++ +++ ++ + ++ + ++ + ++ ++ ++ + ++ ++ ++ + +++ +++ ++ - ++ + 86.7 82.6
S. aureus +++ - +++ - +++ - +++ - ++ - ++ - ++ +++ +++ - - - +++ - +++ + 82.6 21.7
B. cereus ++ +++ +++ ++ +++ ++ +++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ 82.6 82.6
L. monocytogenes ++ ++ +++ +++ +++ ++ +++ ++ ++ ++ ++ +++ ++ - +++ +++ ++ ++ ++ ++ ++ - 78.2 60.8
L. plantarum ++ +++ - - - - - - - - - - ++ ++ - - ++ ++ - - + + 47.8 47.8
C. albicans - - - - - - - - - - - - - - - - - - - - - - 0 0
E. coli ++ + - - - - - - - - - - ++ - ++ - - - - - ++ ++ 43.5 17.3
S. Enteritidis ++ - - - + - + - + - + - ++ - ++ - +++ + + - +++ ++ 56.5 17.3
C. jejuni + - ++ - + - + - ++ - ++ - - - - - + - ++ - ++ ++ 69.5 26
C. coli + ++++ - - - - - - ++ - + - ++ - + - ++ ++++ + - +++ ++ 60.8 43.5
P. aeruginosa - - - - - - - - - - - - - - - - - - - - ++ ++ 4.3 4.3
MRSA +++ - +++ ++ +++ ++ +++ ++ ++ ++ ++ +++ ++ + +++ ++ - - +++ ++ +++ ++ 82.6 21.7
VRE ++ +++ - - - - - - - - + + + ++ - - ++ ++ - - + + 56.5 52.2
MRKP - - - - - - - - - - - - - - - - - - - - ++ ++ 4.3 4.3
+: zone of inhibition observed with a clear halo of growth inhibition in at least one time point; ++ and +++: increased activity as assessed visually by the
increased diameter of the inhibition zone; -: no inhibition. The percentage values were calculated based on the ratio between the number of isolates with
antimicrobial activity against the indicator strain and the total 23 isolates tested. MRKP: Multidrug-resistant Klebsiella pneumonia; VRE: Vancomycin-
resistant Enterococcus faecalis; MRSA: Methicillin-resistant Staphylococcus aureus.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
57
3.2.2. Taxonomic analysis of antimicrobial isolates
Of the 23 isolates selected as expressing the most antimicrobial activity, there were 22 Bacillus
species isolates and 1 Paenibacillus polymyxa isolate. Their 16S rRNA sequences shared 99%
- 100% identity with, mostly, terrestrially derived species (Table 3.3, Figure 3.2). The most
commonly identified species amongst the short-listed isolates were B. subtilis (10 isolates) and
other members of the B. subtilis group, such as B. amyloliquefaciens (5 isolates), B.
licheniformis (1 isolate), and B. safensis (1 isolate). Besides, members of other Bacillus groups
were also identified including B. pacificus (2 isolates), belonging to the B. cereus group; B.
halotolerans (3 isolates), and P. polymyxa (1 isolate)
Table 3.3. Closest species of producers, identified by BLASTn of 16S rRNA against 16S
rRNA database (NCBI) (18/06/2019)
Isolates Closed species Degree of identity (%)
#01 Bacillus halotolerans / Bacillus mojavensis/ Bacillus subtilis subsp. 100 #02 Bacillus subtilis 100 #03 Bacillus licheniformis/ Bacillus haynesii 99.92 #04 Bacillus subtilis 100 #05 Bacillus subtilis 100 #06 Bacillus amyloliquefaciens 99.92 #07 Bacillus pacificus/ Bacillus paranthracis / Bacillus cereus 100 #08 Bacillus amyloliquefaciens 99.92 #09 Bacillus pacificus/ Bacillus paranthracis / Bacillus cereus 100 #10 Bacillus safensis/ Bacillusaustralimaris/ Bacillus pumilus 100 #11 Bacillus amyloliquefaciens 99.92 #12 Bacillus subtilis 100 #13 Bacillus amyloliquefaciens 99.92 #14 Bacillus subtilis 100 #15 Bacillus subtilis 99.92 #16 Bacillus subtilis 99.87 #17 Bacillus subtilis 99.92 #18 Bacillus subtilis 99.76 #19 Bacillus halotolerans/ Bacillus mojavensis 100 #20 Bacillus tequilensis/ Bacillus subtilis 100 #21 Bacillus amyloliquefaciens subsp. plantarum/ Bacillus siamensis 99.84 #22 Bacillus subtilis 100 #23 Paenibacillus polymyxa 99.45

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
58
Figure 3.2. Phylogenetic tree of the 23 short-listed isolates.The neighbour-joining
phylogenetic tree was constructed using the maximum composite likelihood method,
bootstrap method of 1000 replication and pairwise deletion by MEGA &. In this tree, the
23 marine isolates were displayed by assigned number, while all reference strains bacteria
were closely related bacteria identified and downloaded from NCBI.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
59
3.2.3. Proteolytic digestion of antimicrobial culture revealed a dominance of
proteinaceous compounds
The proteinaceous nature of the anti-C. perfringens activity of the isolates was determined by
digesting cell culture supernatants used in the well-diffusion assay with proteases. In 16 of the
19 isolates tested the antimicrobial activities against C. perfringens were reduced or lost after
treatment with at least one proteolytic enzyme (Table 3.4). The antimicrobial activity of each
preparation, treated with each of the enzymes, were directly compared to the untreated control
on the same plate and therefore the zones of clearing are directly comparable and small
differences in zone diameter are informative. The pronase-E enzyme eliminated anti-C.
perfringens activity from 10 isolates, while proteinase-K removed the activity from 4 isolates.
Some antimicrobials (produced by isolates #06, #11, and #21) were affected by both proteinases
and a lipase. Other isolates showed reduced, but not eliminated, antimicrobial activity following
protease treatment (#01, #08, #20, #21). The antimicrobial activities produced by isolates #5,
#18, #19 were not affected by any of the enzymes used. Of the isolates in which antimicrobial
activity was not affected by proteases, isolates #05 and #19 were also resistant to heat treatment
whereas anti- C. perfringens activity was abolished when #18 supernatant was heated.
Table 3.4. Enzymatic sensitivity profile of antimicrobial activities. These shortlisted 19
isolates showed antagonistic activities against C. perfringens.
Isolates Killing
zone (mm)
Proteinase
K
Pronase
E
Trypsin Lipase Sensitive to
#01 14 11 11 14 14 Pronase, proteinase K #02 11 9 - 11 11 Pronase, proteinase K #04 9 0 - - 9 Pronase, proteinase K trypsin #05 24 24 24 24 24 - #06 15 13 11 - - Pronase, trypsin, lipase #08 14 12 12 14 14 Pronase, proteinase K #10 9 - - - 9 Pronase, proteinase K trypsin #11 17 15 - 15 - Pronase, proteinase K lipase #12 12 9 - - 12 Pronase, proteinase K, trypsin #13 16 16 16 13 18 Trypsin #14 9 9 - - - Pronase, proteinase K trypsin, #15 9 0 - 9 9 Pronase, trypsin #16 12 0 - - 12 Pronase, proteinase K, trypsin #17 12 10 - - 9 Pronase, proteinase K, trypsin #18 10 10 10 10 10 - #19 14 14 14 14 13 - #20 11 11 9 11 11 Pronase #21 15 11 12 15 12 Pronase, proteinase K, lipase #23 13 13 - 13 13 Pronase

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
60
(*): The diameter of the killing zone (in millimetres). The value included the diameter of
wells (~6mm)
3.2.4. Determination of the growth depression within the short-listed group;
showing the production of potentially novel antimicrobial activities
It is hypothesised that bacteria produce antimicrobial compounds to compete within an
ecological niche. We, therefore, investigated whether this group of isolates exhibited any
antimicrobial activities against each other [Table 3.5]. Growth inhibition was detected, using
the cross-streak assay, between strains of the same species for isolates B. amyloliquefaciens
#06, #08, and B. subtilis #05. Cross-species antimicrobial activities were detected for many of
the isolates, including, B. amyloliquefaciens #06, #08, #11, #13, B. licheniformis #03; B.
safensis #10; B. halotolerans #01, and P. polymyxa #23. Particularly, P. polymyxa #23
depressed growth of the entire marine Bacillus species.
3.2.5. Growth characteristics of antimicrobial-producing isolates
The basic growth parameters of the 23 isolates were qualitatively evaluated (Table 3.6). All
isolates could grow on various kinds of media. Growth was better in rich nutritious medias such
as Luria-Bertani (LB), brain heart infusion (BHI), Muller Hinton agar (MH), and blood agar
(BA), than growth in less nutritious media such as; lab-prepared marine agar (LPMA) and
marine broth (Difco 2216). All 23 isolates grew vigorously under both aerobic and
microaerophilic conditions but not in anaerobic condition, under incubations temperatures of
30°C, 37oC, 40°C, 50°C, and under pHs ranging from 6.0 to 9.0. Some isolates had reduced
growth at pH5. All isolates could grow in media supplemented with 5% NaCl, and all but isolate
#23 grew in 7% NaCl. No isolate could tolerate 12% NaCl. The marine environment, from
which the isolates were derived, typically has a salt content of 3.5%. No isolates grew in LB
media supplemented with bile salts, even at the lowest concentration tested (0.1M). All isolates
were sensitive to 3 antibiotics; nalidixic acid (50 µg/mL), kanamycin (50 µg/mL) and
tetracycline (50 µg/mL), but 14 of the 23 isolates were resistant to ampicillin (50 µg/mL). All
the isolates had detectable levels of cellulolytic, proteolytic or amylolytic activity

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
61
Table 3.5. Antimicrobial activities amongst the short-listed isolates. The analysis was conducted by cross streak method. Columns with highlight
colour were results from growth depression by isolates of the same species
Marine isolates
#02 #04 #05 #12 #14 #15 #16 #17 #18 #20 #22 #06 #08 #11 #13 #21 #07 #09 #03 #10 #19 #01 #23 #02 B. subtilis - - - - - - - - - - - - + - - - - + - - - - - #04 B. subtilis - - - - - - - - - - - - + - - - - - - - - - - #05 B. subtilis + + - + - + + + + + + + + + + + + + + + - - + #12 B. subtilis - - - - - - - - - - - - - + - - - - - - - - #14 B. subtilis - - - - - - - - - - - - - - - - - - - - - - - #15 B. subtilis - - - - - - - - - - - - - - - - - - - + - - - #16 B. subtilis - - - - - - - - - - - + + - + ++ - - - ++ + + + #17 B. subtilis - - - - - - - - - - - + + - ++ ++ - - - ++ ++ - + #18 B. subtilis - - - - - - - - - - - + + - ++ ++ - + - + ++ - ++ #20 B. subtilis - - - - - - - - - - - - - - ++ + - - - - + - - #22 B. subtilis - - - - - - - - - - - - - - ++ ++ - + - - ++ - ++ #06 B. amyloliquefaciens + + + + - - ++ ++ ++ ++ ++ - + + + ++ + + + + ++ + ++ #08 B. amyloliquefaciens + + + ++ + + + + + ++ ++ + - + ++ + + - + - + + + #11 B. amyloliquefaciens - - - - - - - - - - - - - - - - - - - + - - - #13 B. amyloliquefaciens - - - ++ - - + + + ++ ++ - - - - - ++ ++ - + + - ++ #21 B. amyloliquefaciens - - - + - + - + + + + - - - - - + + - - ++ - - #07 B. pacificus - - - - - - - - - - - - - - - - - - - + - - - #09 B. pacificus + + + + + + - + + + + + + + + + - - + + + + + #03 B. licheniformis + + + + ++ + ++ ++ ++ ++ ++ + + + ++ ++ + + - + ++ + + #10 B. safensis - - - - - - - - - - - + + - + + - - - - + - + #01 B. halotolerans + + - + + + + + + + + + + + + + + + ++ + - - + #19 B. halotolerans + + - + ++ ++ + + ++ + + + + + + + + + ++ + - - + #23 P. polymyxa ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ -
+: zone of inhibition observed with a clear halo of growth inhibition; -: no inhibition. The isolates’ names were temporarily identified based on the name
of first species mentioned 16S rRNA analysis in Table 3. The isolates are arranged to group the isolates of the same species together.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
62
Table 3.6. Characterization of short-listed bacterial isolates.
Isolates # #01 #02 #03 #04 #05 #06 #07 #08 #09 #10 #11 #12 #13 #14 #15 #16 #17 #18 #19 #20 #21 #22 #23 Salinity tolerance (NaCl) 3%-5% ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ 6%-7% ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ - 8% ++ ++ + ++ ++ + - + ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ + - 9% + ++ - ++ ++ - - - ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ - - 10% - + - + + - - - + + + + + + + + + + ++ + + - - 12% - - - - - - - - - - - - - - - - - - - - - - - Bile salt tolerance (mole/L) 0.1M -0.7M - - - - - - - - - - - - - - - - - - - - - - - Antibiotic susceptibility Kanamycin - - - - - - - - - - - - - - - - - - - - - - - Tetracycline - - - - - - - - - - - - - - - - - - - - - - - Ampicillin + + + - + + + + + - - - - + + + + - - - + + - Nalidixic acid - - - - - - - - - - - - - - - - - - - - - - - pH tolerance 5 ++ ++ + + + + + + + + + + + + + + ++ ++ ++ ++ ++ ++ + 6, 7, 8, 9 ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ Thermal tolerance 30°C, 40oC 50oC ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ oxygen requirement for growth
Aerobic, Microaerophillic ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ Anaerobic - - - - - - - - - - - - - - - - - - - - - - - Enzymes production Protease +++ + - + - +++ + ++ ++ + + + ++ - + + ++ ++ +++ - ++ + ++ cellulose degradation ++ ++ ++ ++ ++ ++ + ++ + - ++ ++ ++ ++ ++ ++ ++ ++ ++ - - ++ ++ starch degradation + + - + + + + + + - + + + + + + + + + + + + + Haemolysis β β α β β α ϓ α ϓ β β β α β β β β β β β ϓ β ϓ Growth on media LPMA, MA + + + + + + + + + + + + + + + + + + + ++ ++ ++ ++ LBA, MHA, BHI ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++ ++

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
63
3.3. Summary of results and discussion
There is growing interest in the marine environment as a potential source of bacteria that
produce novel antimicrobial compounds, particularly bacteriocins. The antimicrobial peptides
have been gaining interest as potential drug candidates for clinical treatment of antibiotic-
resistant pathogens (19). In this study, spore-forming bacteria isolated from the coastal marine
environment of Nha Trang (Vietnam Sea) were screened to identify Bacillus isolates producing
antimicrobial compounds. Members of the Bacillus genus were targeted because they are well-
recognized as producers of structurally diverse. Of the various types of marine samples
collected, the antimicrobial producing isolates were most frequently recovered from sponges,
followed by sediments, seaweeds and sea-water samples. Strains with pigments were regularly
detected with low/no antimicrobial activity. Marine sponges have multi-porous structures that
may trap and maintain high bacterial densities, leading to higher recovery rates of antimicrobial
producers. The recovery rate in marine sponges, at 7.6%, was at the lower end of the range
reported in previous studies (5.5% to 50.0%) (123). A lower recovery rate of antimicrobial
producing isolates was also seen from seaweeds, at 3.6%, whereas previous studies had
identified them from seaweeds at 11.0%–16.0% (63). The differences between this study and
previous studies could be due to geographical differences, marine conditions, or heat treatment
to select the spore-formers that may eliminate the metabolically active vegetative cells.
The methods used to detect antimicrobial activity had different levels of sensitivity. The well-
diffusion assay was less sensitive than the cross-streak assay. This effect was most obvious with
the failure to detect activity against Gram-negative bacteria, S. aureus and MRSA when the
latter assay was used. This may occur because of the production of multiple antimicrobial
compounds, with variable relative expression levels in liquid and solid media-based cultivation.
Many studies have reported the influence of various factors on bacteriocin production in liquid
culture such as type of culture media, pH, temperature, growth phase, and quorum sensing
regulation (124). Therefore, the production conditions for some bacteriocins may require
optimization, like varing the concentrations of medium components, screening of incubation
condition (temperature, shaking speed).
The most commonly identified species amongst the 23 shortlisted isolates that exhibited the
strongest antimicrobial activities, were members of the B. subtilis group. These species have
been widely reported in both marine environments and terrestrial environments and can tolerate
the broad environmental conditions (nutrients, pHs, chemo-physical conditions) that are
typically found in marine environments. For example, a study by Ivanova et al. (1999) reported

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
64
that 55.0% (11/ 20) of endospore-forming bacteria isolated from different areas of the Pacific
Ocean were B. subtilis species (125). While B. subtilis, B. amyloliquefaciens, B. pumilus and
P. polymyxa were all identified amongst aerobic spore-forming isolates from marine sources
from the Gulf of Mexico (126), or isolated from seaweed samples collected from the Irish Sea
(86). Other typical marine species, for example, B. aquimaris, B. algicola and B. hwajinpoensis
may require additional special nutrients or salts to recover, or could have been eliminated during
screening and shortlisting based on the strength of antimicrobial activity.
The marine Bacillus/Paenibacillus isolates that we have characterised had broad antimicrobial
activity against a range of human, veterinary and foodborne pathogens, including three
antibiotic-resistant pathogens (MRKP, MRSA and VRE). B. amyloliquefaciens #06, #08, #11,
#13, B. halotolerans #01, #19, B. licheniformis #03, B. safensis #10, P. polymyxa #23 had
broad antimicrobial activity against both Gram-positive and Gram-negative pathogen indicator
strains and other marine Bacillus of different species. This indicated production of either
multiple antimicrobial compounds by a single Bacillus, or a broad-spectrum antimicrobial
compound. Members of the Bacillus genus are known to produce various types of antimicrobial
compounds including polyketides, lipopeptides, bacteriocins, bacilysin, and volatile
compounds. The synergistic effects of these antimicrobials could result in a broad spectrum of
antimicrobial activity, such as noted for a number of the isolates in this study. Also, production
of broad-spectrum bacteriocin was recently reported for a marine Bacillus; sonorensin,
identified from a marine B. sonorensis isolate, exhibited broad-spectrum antibacterial activity
towards both Gram-positive and Gram-negative bacteria (85). Amongst Gram-positive bacteria,
such as Bacillus sp., the expression of bacteriocins and other antimicrobial compounds with
activity against other Gram-positive bacteria is widespread and extensively studied. However,
the production of compounds with activity against Gram-negative bacteria is less common,
therefore, the isolates that have antimicrobial activity against Gram-negative bacteria are of
particular interest. Rarely observed antimicrobial activities such as that observed against
Campylobacter, P. aeruginosa, and even multidrug-resistant K. pneumonia, represent
potentially novel compounds. The P. polymyxa #23 isolate had the strongest and broadest
activity and the activity against Gram-negative bacteria likely results from the expression of
polymyxin and tridecaptin, which has long been known to have such activity (42, 127). The
other isolates with activity against some of the Gram-negative bacteria tested, for example, B.
amyloliquefaciens #11, which had significant activity against both Campylobacter species but
lesser activity against E. coli and Salmonella, appears to indicate a spectrum of activity that has

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
65
not previously been reported and so the compound responsible may be novel and hence warrants
further investigation.
The finding that many of the compounds that had activity against C. perfringens were
inactivated by proteases, indicated that the antimicrobial compounds produced probably
included bacteriocins. Interestingly, the production of large quantities of these antimicrobial
compounds for drug development in the future could likely be achieved as it was demonstrated
that most of the isolates were well adapted to a broad range of growth conditions (various kinds
of nutrients, pHs, salt concentration, temperature, etc). These characteristics of the isolates
represent advantages that could facilitate manufacturing processes, products storage, and the
potential harnessing of these isolates for in vivo use in animal or food applications. The culture
conditions may affect the level of bacteriocin production. Besides, these marine
Bacillus/Paenibacillus isolates were also shown to have proteolytic, cellulolytic, and
amylolytic activity and hence may represent a promising source of important industrial enzymes
such as proteases, cellulases, and amylases. Marine-derived enzymes have been noted to have
significant advantages in manufacturing because they commonly have high adaptability to high-
salt concentration, and fluctuating temperature, pH, organic solvents, and ions (128).
Based on the results of the chapter, it can be confirmed that the Vietnam Sea region harbors an
abundance of marine Bacillus isolates expressing antimicrobial activities against a wide range
of bacterial pathogens, particularly antimicrobial-resistant bacterial pathogens. Some of the
antimicrobials may represent novel compounds that warrant further studies. This supports the
hypothesis raised in the introduction.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
66
Chapter 4 BIOINFORMATIC IDENTIFICATION OF PUTATIVE GENE CLUSTERS
ENCODING ANTIMICROBIAL PEPTIDE PRODUCTION
4.1. Introduction
The recent development of affordable whole genome sequencing (WGS) and various
bioinformatic tools for genome analysis are speeding up antibiotic discovery from natural
resources. Nowadays, the elucidation of antimicrobial potential of bacteria is facilitated by in
silico prediction of genes encoding all putative antimicrobial substances. The method allows
the identification of candidates which harbour genes of novel substances for latter purification,
therefore, i) omitting the unnecessary purification processes of replicated compounds. In
addition, ii) the prediction of antimicrobial peptides from genomes can provide information on
gene components of bacteriocins biosynthetic gene cluster for further cloning/ expression for
mass production; and iii) can provide information on properties of antimicrobial substance
which are key factors to determine appropriate purification procedures. Therefore, genome
analysis is a powerful tool that can be applied before the direct purification, to save time and
reduce the range of purifications that need to be trialed.
In this study 6 marine isolates including 4 B. amyloliquefaciens (#06, #08, #11 and #13), 1 B.
halotolerans (#01), and 1 P. polymyxa (#23) were selected for genome sequencing using the
Illumina MiSeq sequencing platform. They were selected because they exhibited strong
antimicrobial activity against a range of important hospital-acquired pathogens and foodborne
pathogens such as Gram-negative bacteria (Salmonella, E. coli, Campylobacter coli,
Campylobacter jejuni, Pseudomonas aeruginosa), (129) and a majority of the Gram-positive
bacteria tested. In practice, the genome analysis included the steps of (i) generation of raw
sequence data, (ii) assembly the raw reads using the A5-miseq pipeline to generate draft
genomes; (iii) annotation of the draft genomes using web-based RAST for gene calling and
amino acid translation; iv) calculation of ANI values to estimate the genome similarity within
the marine species and versus the genomes of terrestrial species; (v) confirmation of the genome
similarity by pairwise comparison using the SBCT tool (of RAST) and MAUVE, to visualize
the percentage of similar CDS shared between genomes, and genome patterns; (vi) in silico
prediction of putative antimicrobial peptide encoding genes, including bacteriocins and NRPs,
by BAGEL and AntiSMASH; and vii) determination of gene components of bacteriocin gene
clusters. The final aim is to identify candidate biosynthetic gene clusters of putative novel
bacteriocins and to select strains for purification of bacteriocins

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
67
4.2. Results
4.2.1. Overview of six draft genomes
After assembly, the size of the draft genomes varied based on the species [Table 4.1]. Thus,
five Bacillus species had genome sizes ranging from 3.87 Mbps to 4.14 Mbps, and the genome
of P. polymyxa #23 was ~6.01 Mbps. The G+C contents of all genomes were similar, around
43.8% - 46.5%. There were no plasmids found. The draft genome of P. polymyxa #23 consisted
of 68 contigs while the number of contigs in genomes of Bacillus species ranged from 17 to 33
contigs.
Table 4.1.Overview of draft genomes of marine isolates
Marine isolates
#01 # 06 #08 # 11 #13 #23
Identification B. halotolerans
B.
amyloliquefaciens
P. polymyxa
Source Sediment sponge sponge seaweed sponge sponge
Numbers of contigs 31 17 23 33 19 68
Genome size 4.14 3.87 3.95 4.01 3.90 6.01
RNAs 119 114 102 118 102 187
No. CDSs 4033 3720 3839 3900 3776 5236
Subsystem coverage 48% 49% 48% 49% 48% 42%
N50 1077099 988071 991768 339244 1024489 469875
G+C (%) 43.8 46.5 46.3 46.2 46.5 45.2
Subsystems quantity 467 462 459 469 465 452
4.2.2. Calculation of ANI values across the genomes revealed a high degree of similarity
between marine species and terrestrial neighbour strains
To evaluate the genetic similarity of genomes between marine strains and closely related
bacteria isolated from terrestrial sources, an average nucleotide identity (ANI) plot (Figure 4.1)
was generated using GET-HOMOLOGOUS. The Bacillus/Paenibacillus species used for this
calculation included 6 B. amyloliquefaciens subsp. plantarum, 4 of which were isolated in this
study and 2 were from the NCBI genome database (#06, #08, #11, #13, FZB42, CAUB419),; 6
P. polymyxa (#23, CR1, E681, SC2, SQR21, HY962); 6 B. halotolerans (#01, ZB201701,

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
68
F413, NRRLB41617, ZB201702, ATC25096). These terrestrial strains were selected because
they had high similarity in 16S rRNA sequences with those of the marine isolates.
The phylogenetic tree included three distinct clades; clades A (comprising 6 B.
amyloliquefaciens species); clade B (comprising 6 B. halotolerans), and clade C (comprising 6
P. polymyxa species) [Figure 4.1]. The ANI values across genomes ranged from 50.27% to
96.03% where the lowest ANI value was observed between the genomes of B.
amyloliquefaciens #08 and P. polymyxa CR1; the highest ANI value was seen between genomes
of B. amyloliquefaciens #6 and B. amyloliquefaciens #08. The ANI was highest across genomes
of B. amyloliquefaciens species (94.08%-96.03%), followed by B. halotolerans (77.79% -
78.83%), and lowest ANI was observed across P. polymyxa (50.02% - 51.17%). Within the B.
amyloliquefaciens species, B. amyloliquefaciens #11 was closer to B. amyloliquefaciens FZB42
(ANI=95%) while B. amyloliquefaciens #06, #08 #13 were closer to B. amyloliquefaciens
CAUB419 (95.73%, 95.71%, 95.65%).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
69
Figure 4.1 The heat map of the average nucleotide identity and genome-based phylogenetic tree across Bacillus/Paenibacillus genomes. These
genomes included 6 B. amyloliquefaciens (Ba) (#06, #08, #11, #13, FZB42, CAUB41, 6 P. polymyxa (Pp) (#23, CR1, E681, SC2, SQR21) and 6
B. halotolerans (Bh) (#01, ZB201701, F413, NRRLB41617). The graphic was generated by calculation of GET-HOMOLOGOUS software using
BLASTn for gene clustering and determination of similarity across the genomes dataset. The highest similarity across genomes was observed at B.
amyloliquefaciens, followed by B. halotolerans, and the P. polymyxa genomes displayed the lowest similarity.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
70
4.2.3. Analysis of the distribution frequency of genes across the species revealed a high
number of strain-specific genes in P. polymyxa genomes
The frequency distribution of orthologous coding sequence (CDS) across the genomes of 6 B.
amyloliquefaciens, 6 P. polymyxa, and 6 B. halotolerans genomes were calculated separately
and then compared. The result showed that 6 B. amyloliquefaciens genomes contained 5172
orthologous proteins comprising 1337 cloud CDS (25.85%), 312 CDS in the shell (6.03%) and
3523 CDS in the cloud (68.12%), 3320 in the core (64.19%). The B. halotolerans contained
6063 orthologous proteins including 2071 CDS in clouds (34.16%), 467 CDS in shells (7.70%),
3525 CDS in soft-core (58.14%), and 2395 CDS in the core (39.50%). The 6 P. polymyxa
genomes contained 9206 orthologous CDS, comprising of 4444 CDS in the cloud (48.27%),
and 707 in the shell (7.68%), 4055 soft-core (44.05%), and 3549 genes in the core (38.55%).
Therefore, numbers of CDS in the cloud catalogue was highest at P. polymyxa, followed by B.
halotolerans and B. amyloliquefaciens. The ratio between the number of CDS presented in 1
genome (‘strict cloud’) and the number of genes presented in 6 genomes ('strict' cores') was ~1
in P. polymyxa, higher than that in B. halotolerans, and B. amyloliquefaciens (about ~1/2 X).
This indicated a sightly enrichment of strains-specific genes in P. polymyxa genomes [Table
4.2]
Figure 4.2. Frequency distribution of CDS across the genomes.. (A) Of 6 B. halotolerans
genomes; (B) 6 B. amyloliquefaciens genomes; (C) 6 P. polymyxa. Bars from the left to
right illustrated a number of CDS presented in 1 genome (named as 'strict' cloud); 2
genomes, 3 genomes, 4 genomes 5 genomes, and 6 genomes (called as ‘strict’ core)
respectively. The ratio of 'strict' cloud: 'strict' core were ~1/2 in B. halotolerans and B.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
71
amyloliquefaciens, but there was a slight enrichment of unique CDS in P. polymyxa when
this ratio was ~1.
4.2.4. The pairwise comparison across genomes of B. halotolerans revealed the high
similarity in genome organisation and CDS.
The similarity across the genomes was confirmed by MAUVE and SBCT tools. Among the
isolates of B. halotolerans, the genome of #01 was closer to B. halotolerans F41-3 (ANI =
94.85%). Genomes alignment performed by MAUVE showed that the genome patterns between
B. halotolerans #01 and B. halotolerans F41-3 had 31 LCBs, most of the genomes were
collinear, no translocations, no inversions, but the presence of some deleted block, indicating
the integration of mobile nucleotide elements (probably phage, transposons and/or integrases)
into the genomes [Figure 4.3 A]
Pairwise comparison between genomes of the isolate #01 and F41-3 resulted in 90.7% similar
CDS (3822/ 4210) (amino acid similarity of ≥70%) [Figure 4.3 B]. There were approximated
2.5% CDS (106/4210) with low similarity found in the only genome of isolate #01 but not in
genome of B. halotolerans F41-3, which were then annotated as; transporters responsible to
transport maltose/maltodextrin and relating to multidrug resistance, transcription regulators,
phage related proteins, hypothetical proteins, and a biosynthetic gene cluster of unknown
polyketide [Table 4.2].
Figure 4.3. Genome comparison between marine B. halotolerans #01 and terrestrial
neighbour B. halotolerans F41-3. (A) Performed by MAUVE to visualize the

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
72
organisation of conserved DNA regions on genome. (B) And by SBCT to identify the
similar proteins. The comparison was conducted between B. halotolerans F41-3 (bottom)
and B. halotolerans #01 as reference (top).
Table 4.2 List of CDS found in genome #01 relating to the biosynthesis of polyketide
compound. These CDS had low similarity with those found in the genome of F41-3
Gene ID Function
fig|6666666.473256.peg.3879 Polyketide synthase modules and related proteins
fig|6666666.473256.peg.3881 Polyketide synthase modules and related proteins
fig|6666666.473256.peg.3881 Polyketide synthase modules and related proteins
fig|6666666.473256.peg.3711 Polyketide biosynthesis 3-hydroxy-3-methylglutaryl-ACP
synthase PksG
fig|6666666.473256.peg.3704 Malonyl CoA-acyl carrier protein transacylase (EC 2.3.1.39)
fig|6666666.473256.peg.3710 Polyketide biosynthesis malonyl-ACP decarboxylase PksF
fig|6666666.473256.peg.3717 Modular polyketide synthase
fig|6666666.473256.peg.1975 Uncharacterized transcriptional response regulator YcbL
fig|6666666.473256.peg.1978 Bacitracin transport ATP-binding protein bcrA
fig|6666666.473256.peg.1673 Two-component system sensor histidine kinase
fig|6666666.473256.peg.3712 Probable polyketide biosynthesis enoyl-CoA hydratase
PksH
fig|6666666.473256.peg.3713 Modular polyketide synthase
fig|6666666.473256.peg.3709 Polyketide biosynthesis acyl-carrier-protein AcpK
fig|6666666.473256.peg.3710 Polyketide biosynthesis malonyl-ACP decarboxylase PksF
fig|6666666.473256.peg.3020 3-oxoacyl-[acyl-carrier-protein] synthase, KASII (EC
2.3.1.179)
fig|6666666.473256.peg.2003 ABC-type Na+ transport system not coupled with H+ or K+
uptake, ATPase component NatA
4.2.5. The pairwise comparison across genomes of B. amyloliquefaciens revealed high
similarity in genome organisation and CDS.
Both MAUVE and SBCT concluded a high degree of similarity between the genomes of marine
B. amyloliquefaciens species and those of terrestrial B. amyloliquefaciens. Of these species,
isolate #11 was closer to B. amyloliquefaciens FZB42 while isolates #06, #08 and #13 were
closer to B. amyloliquefaciens CAUB946.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
73
The MAUVE arrangement showed that there were only 3 LCBs between isolate #11 and FZB42
without genes inversion; 11 LCBs between genomes of #13 and FZB42 with some inversions
and translocations [Figure 4.4]. The high similarity in genomes between #11 and FZB42 was
again confirmed by SBCT when the tool detected that the two genomes had 97.51% similar
proteins, which was significantly higher than the 89.33% similarity between FZB42 and isolate
#13 [Figure 4.4B]. The genome of #13 was closer to B. amyloliquefaciens CAUB946; they
shared 93.83% similar proteins. The comparison performed between marine species indicated
the genome of #13 was similar to that of isolate #08 (had 100% similar proteins), followed by
#06 (had 93.19% similar proteins). Besides, all these marine B. amyloliquefaciens were
‘amyloliquefaciens’ sub-species, as the proportion of similar CDS sharing between marine B.
amyloliquefaciens and strains of ‘plantarum’ subspecies (B. amyloliquefaciens FZB42 or B.
amyloliquefaciens CAUB946) was 90.64%, therefore a higher degree of similarity than that
shared between marine isolates and strain of ‘amyloliquefaciens’ subspecies (B.
amyloliquefaciens DSM 07); 86.67%. There were 4.19% (166/3961) CDS found in the genome
of isolate #11 but presented as low similarity CDS or non-present in the genome of FZB42
[Table 4.3]. These CDS were annotated as hypothetical proteins (65.06%) with unknown
function, phage-relating proteins, and a gene cluster responsible for the biosynthesis of
thiopeptide, a class I bacteriocin.
Table 4.3.Low identity CDSs in the genome of isolate #11 relating to the biosynthetic
gene cluster for bacteriocin production.
Gene code Annotation
fig|6666666.242001.peg.370 Bacitracin ABC transporter, permease protein, putative
fig|6666666.242001.peg.372 Sensory box histidine kinase
fig|6666666.242001.peg.373 Positive regulation of late competence genes and surfactin production (comS, srfA, rapA)
fig|6666666.242001.peg.374 Hypothetical protein
fig|6666666.242001.peg.375 Lanthionine biosynthesis protein, LanM
fig|6666666.242001.peg.376 methionine ABC transporter ATP-binding protein
fig|6666666.242001.peg.377 hypothetical protein
fig|6666666.242001.peg.378 lanthionine biosynthesis protein, LanM

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
74
Figure 4.4. Genome comparison between marine B. amyloliquefaciens and terrestrial B.
amyloliquefaciens (A) Performance by MAUVE between the genome of isolate #11 and
genome FZB42 (A1), and between the genome of isolate #13 and FZB42 (A2). (B)
Performance by SBCT. From the outer ring to inner ring were genomes of FZB42, #08,
#06, DSM7, CAUB946, CBMB205, #11. The comparison was performed against the
genome of isolate #13 as reference.
4.2.6. The pairwise comparison across genomes of P. polymyxa revealed diversity in
genome organisation and CDS.
Both MAUVE and RAST revealed an average degree of similarity between genomes of marine
P. polymyxa #23 and terrestrial neighbour P. polymyxa [Figure 4.3]. The genome of isolate #23
shared 87.20% similar CDS with P. polymyxa SQR21, 81.72% with P. polymyxa SC2, and
78.47% CDS with P. polymyxa CR1 [Table 4.5]. Therefore, the genome of #23 was closer to
the genome of P. polymyxa SQR21. The MAUVE analysis confirmed this similarity between

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
75
these two isolates when there was less number of LCBs between the genome of isolate #23 and
SQR21 than that observed between isolate #23 and P. polymyxa CR1.
Figure 4.5. Genome comparison between marine P. polymyxa #23 and terrestrial P.
polymyxa species. (A) The performance was carried out by MAUVE analysis between
genomes of isolate #23 and the genome of terrestrial P. polymyxa SQR21 (A1); and
between isolate # 23 and terrestrial P. polymyxa CR1 (A2). (B) The performance was
carried out by SBCT between marine P. polymyxa # 23 (used as reference) and genomes
of P. polymyxa SQR21, P. polymyxa CR1, P. polymyxa SC2 (from the inner ring to outer
ring).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
76
There were 12.71% (680/ 5352) CDSs found in the genome of isolate #23 which had low
similarity to those found in the genome of SQR21. These low similarity CDSs were located
mostly in contigs #33 and #34. They were annotated as hypothetical proteins (64.85%),
transcriptional regulators (4.41%) including HXlR, XRE, ADA, AraC, HcpR, MerR, TetR,
LysR, ArsR, AsnC, LrpA, AbrB, PadR which were known to regulate carbon-storage, sugar
metabolism, stress response; transmembrane transporters (2.05%) which including multidrug-
efflux transporter, bacitracin ABC transport, multidrug resistant transporter, Bcr/cflA family,
methionine transporters which may be responsible for antibiotic resistance; and proteins
relating a bacteriocin biosynthetic gene cluster [Table 4.4].
Table 4.4. The CDSs relating to unique bacteriocin biosynthetic gene clusters found in
the genome #23 but had low similarity to those found in the genome of isolate CAUB419
Gene code Functions suggested by RAST
Bacteriocin gene cluster
fig|6666666.242000.peg.4718 Methionine ABC transporter ATP-binding protein
fig|6666666.242000.peg.4719 Lipid A export ATP-binding/permease protein MsbA
fig|6666666.242000.peg.4720 Glycosyl transferase, family 8
fig|6666666.242000.peg.4721 Glycosyltransferase, group 2 family protein
fig|6666666.242000.peg.4722 hypothetical protein
fig|6666666.242000.peg.4723 UDP-4-amino-4-deoxy-L-arabinose--oxoglutarate aminotransferase (EC 2.6.1.-)
fig|6666666.242000.peg.4724 hypothetical protein
fig|6666666.242000.peg.4725 Radical SAM
fig|6666666.242000.peg.4726 hypothetical protein
4.2.7. In silico prediction of antimicrobial compounds revealed high numbers of NRPs
and bacteriocins, including novel bacteriocins
In silico prediction of putative antimicrobial peptides from these six genomes by both BAGEL
and antiSMASH bioinformatic tools resulted in the detection of a total of 61antimicrobial
peptides (AMPs) including 20 bacteriocins and 41 NRPs compounds (lipopeptides, polyketides,
hybrids, bacilysin) [Table 4.5]. The B. amyloliquefaciens and P. polymyxa harboured higher
numbers of AMPs than those found in the genome of B. halotolerans #01. Types of NRPs
predicted from genomes from the Bacillus taxon were different to the types predicted to be

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
77
present in the P. polymyxa taxon, which included lipopeptides (tridecaptin, polymyxin,
paenibacterin, and an unknown LP), polyketides (fusaricidin and an unknown polyketide).
Although 41 NRSPs were predicted, there were only 14 different types of substances. This was
because many Bacillus derived polyketides (bacillaene and macrolactin), lipopeptides
(surfactin, fengycin, and bacillomycin D), siderophores (bacillibactin) and a dipeptide bacilysin
were presented in all members of the Bacillus taxon. However, the genes encoding the difficidin
were found in all B. amyloliquefaciens species but were not found in the genome of B.
halotolerans #01. The NRPs gene clusters were large in size (commonly larger than 12 kDa),
including multiple replicated modules of synthetases enzymes (28), which would give many
difficulties for further cloning/ expression to characterise. Most of the NRPs gene clusters were
well characterised, the gene determinants of some gene clusters of undefined compounds were
separated on different contigs due to multiple gaps presented within the incompleted
chromosome after miseq sequencing, therefore the identification of full gene clusters would
have many challenges, therefore gene organisation of the NRPs gene cluster was not further
annotated in this study. Also, they were not types that were of interest for this study, because
they were predicted with less novelty of substances.
Although 20 bacteriocins were predicted within six genomes there were only 13 different types
[Table 4.5, Table 4.6]. This was because of the replication of several bacteriocins (LCI,
amylocyclicin) in all the B. amyloliquefaciens genomes; and 1 novel thiopeptide cluster
presented in genomes of both isolates #06 and #13. Of these 13 bacteriocins, 6 bacteriocins
were found to be characterised from earlier studies including subtilosin A (isolate # 01),
plantazolicin (# 06), mersacidin (# 08), paenicidin A (#23), LCI and amylocyclicin (all #06,
#08, #11, #13); and 7 uncharacterised bacteriocins including 2 thiopeptides (one presented in
both #08 and #13, and one found in #11), 1 two-component lantibiotic (#13), 1 sactipeptide
(#23), 1 lantibiotic (#23), 1 lantibiotic (#08) and 1 uncharacterised lassopeptide (#23). A
bacteriocin, in this study, was considered as a unique or uncharacterised bacteriocin when the
precursor’s amino acid sequence had a low identity (< 90% of similarity), or was unmatched to
any characterised bacteriocins listed from the bacteriocin database (BACTIBASE), and uniref
90 database (UNIPROT).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
78
Table 4.5. List of putative antimicrobial peptides predicted within Bacillus/Paenibacillus
draft genomes.
Similar known peptides
Class Similarity Marine isolates
#01 #06 #08 #11 #13 #23
Bacteriocins
LCI class II 100% • • • • •
amylocyclicin head to tail cyclized peptide
100% • • • • •
subtilosin A sactipeptide 100% •
paenicidin A lantibiotic type 1 - •
mersacidin lantibiotic type 1 100% •
plantazolicin LAPs 100% •
unknown thiopeptide - • •
unknown thiopeptide - •
unknown sactipeptide - •
paeninodin lassopeptide 60% •
unknown lantibiotic type 1 - •
unknown lantibiotic type 2 - •
fengycin lipopeptide 100% • • • • •
bacillibactin lipopeptide 100% • • • • •
surfactin lipopeptide 100% • • • • •
fusaricidin lipopeptide 100% •
penibacterin lipopeptide 60% •
tridecaptin lipopeptide 100% •
polymyxin lipopeptide 100% •
unknown lipopeptide - •
bacillaene polyketide 100% • • • • •
macrolactin polyketide 100% • • • • •
difficidin polyketide 100% • • • •
unknown polyketide •
bacilysin other 100% • • • • •

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
79
Table 4.6. Amino acid sequences of precursor bacteriocins predicted within 6 Bacillus genomes
. Hosts
Class Amino acid sequence MW (kDa)
Similar known bacteriocin
Similarity (%)
Characterized bacteriocins
1 #01 sactipeptide MKKAVIVENKGCATCSIGAACLVDGPIPDFEIAGATGLFGLWG
3.44 subtilosin A 100%
2 #06; #08; #11; #13
unmodified MKFKKVLTGSALSLALLMSAAPAFAASPTASASVENSPISTKADAGINAIKLVQSPNGNFAASFVLDGTKWIFKSKYYDSSKGYWVGIYESVDK
5 LCI 100%
3 #06 LAPs MNSLSINFMEEVTIMTQIKVPTALIASVHGEGQHLFEPMAARCTCTTIISSSSTF
1.34 plantazolicin 100%
4 #06; #08; #11; #13
head to tail cyclized peptide
MMNLVKSNKKSFILFGAALAAATLVYALLLTGTELNVAAAHAFSANAELASTLGISTAAAKKAIDIIDAASTIASIISLIGIVTGAGAISYAIVATAKTMIKKYGKKYAAAW
6.38 amylocyclicin 100%
5 #08 lantibiotic MSQEAIIRSWKDPFSRENSTQNPAGNPFSELKEAQMDKLVGAGDMEAACTFTLPGGGGVCTLTSECIC
1.98 mersacidine 100%
6 #23 lantibiotic MAENLFDLDIQVNKSQGSVEPQVLSIVACSSGCGSGKTAASCVETCGNRCFTNVGSLC
3.38 paenicidin A 100%
7 #23 lassopeptide MAAINPAKDHYRITYGEEVYVSDMDGEKVMMSIHTGKYYNLGFTGGRIWELAEFSPSIEDIVTVLTNEYEVDEEQCRQQVHTFVAELEREGLLRLLRELD
paeninodine 60%
Uncharacterized bacteriocins
8 #06, #13 thiopeptide MKKEKNDLLNNLEITGLDVTEITDSISIPETGATSGSGGHSTCGSSSCCSSCCS
1.88 unique -

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
80
. Hosts
Class Amino acid sequence MW (kDa)
Similar known bacteriocin
Similarity (%)
9 #08 lantibiotic MSEKQIERTIRAWKDPEFRKTLSDVADHPSGSIEARQLSMLGSEDNVNPMTTPVSPAPTVVIRVSLKVCPTFKIKCKV
lichenicidinVK21A2
37%
10 #11 thiopeptide MEKMMSQNVKEKEDSLDLTSILDEIESIDIMEVSESMLLSETGATSGSSSSSSTSCCGSCSCASCGACTSCG
- unique -
11 #13 lantibiotic MKKDFQALTPMTEEELKNLAGGSDATPMTVTPTTITIPISLAGCPTTKCASIVSPCND
3.61 haloduracin β 41%
12 #23 sactipeptide MRKLVKRSTNVGDTIEAFGCGCSCYCPCSCYCAGSLTRSSNTSRESDGSYRRDNGTGIGNY
4.4.6 unique -
13 #23 lantibiotic MNSPELVFFEQEDTLDLDLQINDLTLKQAKNPCTSTVTCSVSRCLGTHVTCECWC
2.82 unique -
The bold letters indicated the position of core sequence on precursor, normal letters denoted the leader sequence. It was predicted by SingalP.v5.0
prediction (http://www.cbs.dtu.dk/services/SignalP/). The MW was predicted based on the core sequence using expasy tool
(https://web.expasy.org/compute_pi/).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
81
A1 A2
A3
0BAC014_|_mersacidin 4008lan 2423lan1 913lan 1023lan2 2BAC215_|_Paenicidin_A 2BAC197_|_Lichenicidin_A2 0BAC196_|_Lichenicidin_A1
Consensus
.M......
.S......
.Q......
.E......
.A......
.I......
.I......
.R......
.S......
.W......
.K......
.D......
.P......
.F......
.S......
.R......
.EM.....
.NA.....
.SE.....
.TN.....
.QL.....
.NF.....
.PD.....
.AL.....
.GD.....
.NI.....
.PQ.....
.FV.....
.SN.....
.EK.....
.LS.M...
.KQMN...
.EGKS...
.ASKP...
.QVDE...
.MEFL...
.DPQV...
.KQAF...
.LVLFVT.
.VLTELT.
19BAC014_|_mersacidin 6708lan 5123lan1 4913lan 4423lan2 29BAC215_|_Paenicidin_A 28BAC197_|_Lichenicidin_A2 28BAC196_|_Lichenicidin_A1
Consensus
.GSPQSP.
.AIMEIAT
.GVTDVTI
.DAETATT
.MCELCSLs
.ESEDSSS
.ASLLSWT
.AGKDGTCc
CCCNLCCA
TTGLQGII
FFSAISTL
TTGGNGAS
LLKGDKGK
PP.SL..P
.
.
.DT..L
.
.
.A....
.
.
.T....
.
.
.P....
.
.
.M....
.
.
.T....
.
.
.V....
.
.
.TL...
.
.
.PK...
.
.
.TQ...
.
.
.TA...
.
.
.IK.V.t
GGTTNTTG
GGAIPAVN
GGAPCASN
GGSITSAG
.
.CSSCSYv
VVVLTVLL
CCEAVE.Ct
TTTGTT.Tc
LLCCCCCV
TTGPSGPT
SSNTVNTK
EERTSRTEc
CCCKRCKC
IIFCCFCM
20BAC014_|_mersacidin 6808lan 5823lan1 5813lan 5423lan2 36BAC215_|_Paenicidin_A 32BAC197_|_Lichenicidin_A2 32BAC196_|_Lichenicidin_A1
Consensus
.
.TALTTP
.
.NSGN..
.
.VITV..
.
.GVHG..s
.
.SSVSS.
.
.LPTLRSc
CCCCCCCC
.
.
.NE..N
.
.
.DC...
.
.
.
.W...
3101sac 4023sac 31BAC098_|_subtilosin 22BAC217_|_Thuricin_CD_beta 20BAC201_|_Thurincin_H 31BAC166_|_Propionicin-F
Consensus
.M....
.R....
.K....
.L....
.V....
.K....
.R....
.S...W
MTM..F
KNK..Y
KVK..Q
AGA..G
VDV..M
ITI..N
VIV..Ie
EEE..A
NANGDI
KFKWWYg
GGGVTAc
CCCACN
.G....
ACACWI
TSTVSGc
CCCGCG
SYSALV
ICICVAg
GPGGCNa
ACATAIa
ASAVAIc
CCCCCGl
LYLLSYv
VCVAVT
DADSEEg
GGGG.A
PSP..A
ILIGLV
PTPVLA
DRDGNT
FSFTLLe
ESEE..
4301sac 6023sac 43BAC098_|_subtilosin 30BAC217_|_Thuricin_CD_beta 31BAC201_|_Thurincin_H 43BAC166_|_Propionicin-F
Consensus
INIFVLa
ATAATGa
GSGAAAa
ARAAAVt
TETSTVg
GSGYGA
LDLFAV
FGFLSA
GSG.TP
LYL.AV
WRW.SV
GRG..P
.D....
.N....
.G....
.T....
.G....
.I....
.G....
.N....
26nosM 25nocM 33tsrH 36sioH 29cltA 21tpdA 22tpaA 2913thioA 4011thioA
Consensus
.
.
.
.
.
.
.
.M
.
.
.
.
.
.
.
.E
.
.
.
.
.
.
.
.K
.
.MM....M
.
.SS....M
.
.NT....S
.
.AA....Q
.
.AA....N
.
.LI....V
.
.
.V....K
.
.
.GM..ME
.
.
.QE..KK
.
.EEK..KE
M.IIE..ED
DMGGL..KS
ASVVV..NL
AAEDLM.DD
HDGGDDMLL
LLLLLLNLT
SSTTASKNS
DAGGDDDNI
LLLLLLILL
DNDDSPDED
.
.
.
.
.
.
.
.E
IIVVVMLII
DDDDDDSTE
ASTAEVAGS
LLLLLFILI
EEEEDEEDD
IIIIVLIVI
SSSSLASTM
EEDDPDDEE
FFYYT.LIV
LLMMS.ITS
DDDDPDSDE
EDEEGGESS
SSTTAVTIM
RRLLGAESL
LLLLLVQIL
EEDDEESPS
50nosM 49nocM 58tsrH 61sioH 60cltA 49tpdA 48tpaA 5413thioA 7111thioA
Consensus
DDGGSSDEE
SSEEILDTT
EEDDNTAGG
VVLLVALAA
VVTSGGSTT
AAVVHHQSS
KKTTAGVGG
VVMMMMMSS
MMIVVTAGS
SSASEEAGS
.
.SSIVSHS
.
.
.
.GG..S
.
.
.
.
.
.
.
.S
.
.
.
.A...T
AAAASACSS
SSSSNSTTC
CCCCCCTCC
TTTTTNTGG
TTTTS.GSS
CCCCTCCSC
EEIIGFASS
CCCCTCCCC
CSTTPYSCA
.
.
.
.AI..S
.
.
.
.S....
CCCCCCSSC
.
.
.
.CC..G
SSSSSSSSA
CCCCCCSCC
SSSSCSSCT
SSSSCSSSS
.
.
.
.CAT.C

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
82
B3
B2
B1
Figure 4.6. Multiple sequence alignment (A) and average distance tree (B) between the bacteriocins precursors in this study and known
precursors . The calculations were performed on DNAman software using Clustal W (130). (1) lantibiotic precursors (13lan1, 13lan2 were
novel precursors), (2) lassopeptide precursors (23sacA was novel precursor); (3) thiopeptides precursors (06thioA, 13thioA were novel
precursor). The letters in black (100% of identity), pink (≥70%), and blue (50%). The thiopeptides for comparison were acquired from
THIOBASE (131) while sactipeptides and lantibiotic were downloaded from BACTIBASE database (110).
06thioA
13thio
11thioA
ZP_00237834.1
ZP_00237835.1
ZP_00237837.1
ZP_00237830.1
ZP_00237832.1
NP_834755.1
NP_834756.1
NP_834757.1
NP_834758.1
nosM
sioH
YP_003974670.1
YP_003974672.1
cltA
100%
100%
100%
100%
100%
100%
100%
95%
61%
53%
45%
33%
26%
20%
17%
11%
100% 0%80% 60% 40% 20%
01sac
BAC098_|_subtilosin
BAC201_|_Thurincin_H
23sac
BAC217_|_Thuricin_CD_beta
BAC166_|_Propionicin-F
100%
32%
17%
15%
4%
100% 0%80% 60% 40% 20%
BAC014_|_mersacidin
08lan
BAC196_|_Lichenicidin_A1
23lan1
BAC215_|_Paenicidin_A
13lan
BAC197_|_Lichenicidin_A2
23lan2
100%
100%
59%
35%
24%
16%
11%
100% 0%80% 60% 40% 20%

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
83
4.2.8. Prediction of gene organisation of biosynthetic gene clusters of characterised
bacteriocins
4.2.8.1. The LCI gene clusters (found in the genomes of isolates # 06, #08, #11, #13)
The biosynthetic gene clusters of LCI were found in all 4 B. amyloliquefaciens genomes (#06,
#08, #11, #13) [Figure 4.7]. The gene clusters were the same in all 4 species, and also had
100% identity to the LCI gene cluster found ubiquitously in the B. subtilis/B.
amyloliquefaciens group. The LCI is an unmodified bacteriocin (class II), pI of 9.85,
molecular weight (MW) of 5 kDa and exhibits antimicrobial activity against Gram-negative
bacteria (Xanthomonas sp., P. solanacearum) but is not active against E. coli (54).
The gene organisation of the LCI biosynthetic gene clusters found in these marine isolate
genomes were the same as the well-characterised ones (54, 132). It consisted of a structural
gene encoding a precursor bacteriocin and an ABC transport gene that was responsible for
immunity/transportation. The precursor LCI sequence had 94 amino acids consisting of a
leader sequence and a 46 aa core sequence. The core sequence had a molecular mass of 5.46
kDa and a pI of 10.25.
Figure 4.7 The LCI gene cluster found in marine B. amyloliquefaciens #06, #08, #11
and #13. (A) The gene organisation of the LCI gene cluster. Green colour shows
structural gene. (B) The multiple sequence alignment (MSA) between LCI of #11, #06,
#08, #13 and B. amyloliquefaciens FZB42.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
84
4.2.8.2. The amylocyclicin gene clusters (found in genomes of #06, #08, #11 and#13)
The biosynthetic gene clusters of amylocyclicin were also found in all marine B.
amyloliquefaciens species (#06, #08, #11 and #13) (named 11amy) [Figure 4.8]. The gene
clusters were the same in all 4 species, and also had 100% identity to the amylocyclicin gene
cluster found in B. amyloliquefaciens FZB42 (133). This bacteriocin is a head-to-tail cyclised
bacteriocin (class Ib) with a pI of 9.8, MW of 6.38 kDa and is active against Gram-positive
bacteria (Bacillus, Staphylococcus, MRSA, Lactobacillus Listeria and Enterococcus).
The gene organisation of these amylocyclicin biosynthetic gene clusters were the same as the
well characterised one found in B. amyloliquefaciens FZB42 genome (133). It was 4,490 bp
in length, flanked by 2 terminators and consists of 6 ORFs encoding 6 proteins, 11amyA-F
[Figure 4.8, Table 4.7]. The 111 aa precursor bacteriocin (11amyA) had a 47 aa leader
sequence at the C-terminus and a 64 aa core sequence at the N-terminus. The maturation of
head-to-tail bacteriocin commonly relates to three steps: enzymatic cleavage of the leader
sequence and circularization (44). In this gene cluster, the 11amyC was 100% identical to
acnC (found in the one of FZB42) which were responsible to cleavage of the leader sequence
and cyclise two amino acids at two terminal ends of core sequence (133). Two ORFs; 11amyD
(ATP binding domain) and 11amyE (permease domain), which were homology to acnD and
acnE, formed an ATP-dependant two-component ABC transport for secreting the bacteriocin
(134). The 11amyF was an immunity protein for protecting the host from the bacteriocin
effect, and the 11amyB was a hypothetical protein with unknown function
Table 4.7. Structural organisation of amylocyclicin gene cluster found in isolates #06,
#08, #11 and #13
Protein Description Function Similar proteins*
11amyB hypothetical protein unknown function acnB
11amy A precursor precursor acnA
11amy C FIG01241261: hypothetical protein modification enzyme acnC
11amy D ABC transporter, ATP-binding protein ABC Transport
ABC permease
acnD
11amy E uncharacterized protein MJ0793 acnE
11amy F hypothetical protein immunity acnF

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
85
* The similar proteins were observed from the amylocyclicin gene cluster of Bacillus
amyloliquefaciens FZB42 (133).
Figure 4.8. The amylocyclicin gene cluster found in marine B. amyloliquefaciens #06,
#08, #11 and #13. (A) The gene organisation of the gene cluster. (C) The multiple
sequence alignment (MSA) between precursors of #11, #06, #08, #13 and B.
amyloliquefaciens FZB42. The precursor sequences were 100% identical.
4.2.8.3. The mersacidin gene cluster (found in the genome of isolate #08)
The biosynthetic gene cluster of mersacidin was found within scaffold #01 of B.
amyloliquefaciens #08’s draft genome [Figure 4.9]. The mersacidin gene cluster was 100%
similar to that found in Bacillus sp. HIL-Y85/54728 (GenBank AJ250862.2, or BGC0000527)
(135). The mersacidin has an MW of 1,825 Da, globular structure, pI of 4.2, and displays
antimicrobial activity against Gram-positive bacteria including MRSA (41).
The genome organisation of the mersacidin biosynthetic gene cluster in the genome of isolate
08 (named as 08mrs) was the same as that of Bacillus sp. HIL-Y85/54728 (135). It was 12,300
bp in length and consisted of 10 ORFs encoding 10 proteins, 08mrsA-K [Figure 4.9, Table
4.8]. The precursor bacteriocin (08mrsA) comprised a deduced 48 aa leader sequence at the

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
86
C-terminal end and a 20 aa core sequence at the N-terminal end. The lantibiotics commonly
mature via two modification steps; i) removal of the signal sequence, ii) enzymatic
dehydration and cyclisation to convert serine and/or threonine residues to yield unusual amino
acids that are typical for lantibiotic (Dha, Dhb, Lan, MeLan) (39). The 08mrsD was annotated
as a protease, therefore, it was responsible to cleave off the leader sequence. The 08mrsM was
annotated as a lanM-type enzyme which was a typical modification enzyme to modify the
class Ib lantibiotic (39, 40, 136). The matured form of bacteriocin was secreted by 08mrsT
(lanT-type transporter). Three ORFs, mrsF, mrsG, and mrsH formed a group II type ABC
transporter system serving as an immune system to protect host (136-138). Besides, the gene
cluster also harbours a two-component regulatory system formed from 08mrsI and 08mrsJ
(139).
Figure 4.9. The mersacidin gene cluster found in the genome of isolate #08 . (A) Gene
organisation of the gene cluster. (B) MSA between the mersacidin precursors of #08
(08mrsA) and Bacillus sp. HIL-Y85/54728. (C) Multiple sequence alignment at the
gene cluster level (MSAG) between the mersacidin gene cluster of isolate #08 and
Bacillus sp. HIL- Y85/ 54728 (135). The multiple sequences alignment at gene level
(MSAG) was performed by ‘MultiGeneBlast’ tool indicated 100 % similarity between
two gene clusters (140).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
87
Table 4.8. Structural organisation of mersacidin gene cluster found in isolate #08.
Proteins Description Functions Similar protein*
08mrsJ protein putative histidine-kinase regulatory mrsK2
08mrsI protein putative response-regulator regulatory mrsR2
08mrsH ABC-transporter immunity mrsF
08mrsG ABC-transporter integral membrane protein immunity mrsG
08mrsF ABC-transporter integral membrane protein immunity mrsE
08mrsA mersacidin precursor precursor mrsA
08mrsE protein putative response-regulator regulator mrsR1
08mrsD peptidase maturation mrsD
08mrsC lanM- lantibiotic modification enzyme maturation mrsM
08mrsB lanT transportation protein transportation mrsT
* The similar proteins were observed from the mersacidin gene cluster of Bacillus sp. HIL-
Y85/54728.
4.2.8.4. The subtilosin A gene cluster (found in the genome of isolate # 01)
The biosynthetic gene cluster of subtilosin A was found in the genome of isolate #01 [Figure
4.10]. The whole gene cluster had a high degree of similarity (100%) with that of B. subtilis
subsp. spizizenii ATCC 6633 (AJ430547.1, or BGC0000602) (141). The subtilosin A is a
sactipeptide with an MW of 3398.9 Da, pI of 3.88, and exhibits antimicrobial activity against
Gram-positive bacteria (Bacillus sp., Enterococcus sp. and Listeria sp.) as well as Gram-
negative bacteria at high concentration (142).
The biosynthetic gene cluster of subtilosin A found in #01’s genome (named 01sbt) was 8,250
bp in length, flanked by two terminators, and consisted of 8 genes encoding proteins, 01sblA-
H [Figure 4.10, Table 4.9]. The bacteriocin precursor (01sbtA) contained 42 amino acids. The
sactipeptide commonly matures via modification steps; removal of the leader sequence and
the core sequence is modified by a radical SAM that catalyses to form a cross-linking bond
between two amino acid residues (143). In this gene cluster, the putative zinc protease
(01sbtG) and peptidase (01sbtH) were responsible for the elimination of the leader peptide,
modification on core peptide under the catalysis of SAM-radical-enzyme (01sbtB) to mature.
Next, a two-component transport system (clustered by 01stbE and 01stbG) was responsible
for secreting the bacteriocin. The immunity protein (01stbD) was found within the cluster that

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
88
protected the host from bacteriocin activity. The role of two hypothetical proteins; 01slbI,
01sbtB were unknown.
Figure 4.10. The subtilosin A gene cluster found in the genome of isolate #01. (A) Gene
organisation of the subtilosin A gene cluster. (B) MSA of the mersacidin precursors of
#01 ((01-sbtA), and Bacillus sp. HIL-Y85/54728 indicated 100% of similarity. (C) The
MSAG between the subtilosin A gene cluster of isolate #01 and B. subtilis subsp.
spizizenii ATCC 6633. Two gene clusters shared 100% sequence similarity.
Table 4.9. Structural organisation of subtilosin A gene cluster found in the genome of
isolate #01
Protein Description Functions Similar proteins*
01sbtA precursor pepetide precursor sboA
01sbtB unknown unknown sboX
01sbC radical SAM maturation Alba
01sbtD immunity immunity albB
01sbtE ABC transport (ATP binding) transportation albC
01sbtG ABC transport permease transportation albD
01sbtG putative zinc protease maturation able
01sbtH peptidases maturation albF
01sblI unknown unknown albG
* The proteins were observed from the subtilosin A gene cluster of B. subtilis subsp. spizizenii
ATCC 6633.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
89
4.2.8.5. The plantazolicin gene cluster (found in the genome of isolate # 06)
The plantazolicin gene cluster was found in the genome of B. amyloliquefaciens #06. The
gene cluster shared 91% similarity to that of B. amyloliquefaciens FZB42 [Figure 4.11] (51).
The plantazolicin is a ‘linearized-azole-peptide’ bacteriocin. It has MW of 1,336 Da and
displays antimicrobial activity only against Gram-positive bacteria.
The plantazolicin gene cluster (06plz) was 13.3 kb in length and comprised 10 ORFs encoding
10 proteins, 06plzA-N [Figure 4.11, Table 4.10]. The 40aa precursor bacteriocin (06plzA)
was composed of a 26 aa leader sequence at the C-terminus and a 14 aa core sequence at the
N-terminus. The maturation of linearized azole peptides (LAPs) are common via typical
modification steps; enzymatic removal of the leader sequence and modification on core
peptide under the catalysis of 3 enzymes; cyclodehydratase, dehydrogenase, a
docking/scaffolding protein (51, 144). Therefore, these genes were searched in this gene
cluster. Four ORFs were annotated as modification enzymes found including zinc protease
(06plzK) for the elimination of the signal peptides, and cyclodehydratase (06plzH),
dehydrogenase (06plzJ) and a docking/scaffolding protein (06plzI) for modification of the
core sequence (51, 144). Besides, a methyltransferase (06plzL) was found in the gene cluster
that may carry and attach additional methyl moieties to the side chain (51, 145). The
bacteriocin was secreted by a two-component ABC transporter (clustered by 06plzD and
06plzE). The hypothetical protein (06plzF) was also included within the gene cluster but the
function was unknown. A transmembrane protein (06plzB) served an immunity function for
protection.
Figure 4.11. The plantazolicin gene cluster found in genome of isolate #06. (A) Gene
organisation of the plantazolicin gene cluster. (B) MSA of the mersacidin precursors of

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
90
#06 (06plzA) and B. amyloliquefaciens FZB42 indicated 100% of similarity. (C) The
MSAG between the subtilosin A gene cluster of isolate #01 and the one of B. subtilis
subsp. spizizenii ATCC 6633. Two gene clusters shared 91% sequence similarity.
Table 4.10. Structural organisation of plantazolicin gene cluster found in isolate #06
Protein Description Functions Similar proteins*
06plzB hypothetical protein unknown plzF
06plzC transcriptional regulators, marR/emrR family regulation plzK
06plzD TOMM export ABC transporter, ATP-binding transportation plzG
06plzE TOMM export ABC transporter, permease protein transportation plzH
06plzF Similar to Uncharacterized low-complexity proteins
unknown plzI
06plzA precursor bacteriocins precursor plzA
06plzG hypothetical protein (FIG01245785) maturation plzJ
06plzH TOMM biosynthesis cyclodehydratase (protein C) maturation plzC
06plzI TOMM biosynthesis docking scaffold (protein D) maturation plzD
06plzJ TOMM biosynthesis dehydrogenase (protein B) maturation plzB
06plzK TOMM biosynthesis zinc protease removal of leader sequence
plzE
06plzL SAM-dependent methyltransferase (EC 2.1.1) tailing plzL
* The proteins were from the plantazolicin gene cluster of B. amyloliquefaciens FZB42.
4.2.8.6. The paenicidin A gene cluster (found in the genome of isolate #23)
The biosynthetic gene cluster of paenicidin A was found in the genome of P. polymyxa #23
[Figure 4.12]. The whole genome cluster was similar to that of P. polymyxa NRRL B-30509
(146). The paenicidin A has an MW of 3376.5 Da, and displays antibacterial activity against
Gram-positive bacteria (Bacillus sp., Lactobacillus lactis, L. monocytogenes and E. faecium).
The paenicidin A gene cluster (23pae) was 9,667 bp in length and consisted of 6 ORFs
encoding 6 proteins, 23paeA-23paeF [Figure 4.12, Table 4.11]. The precursor (23paeA) was
57 amino acids, consisting of a 21 aa leader sequence at the C- terminus and a 36 aa core
sequence at the N-terminus. The sequence was 100% identical to the paenicidin A precursor

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
91
of P. polymyxa NRRL B-30509. The maturation of paenicidin A occurred under catalysis of
3 enzymes; LanB-type enzyme (23paeB); LanC-type enzyme (23paeD); and LanT-type ABC
transporter (23paeC). The LanB and lanC were well known as two typical modification
enzymes of subclass Ia which catalysed the dehydration and cyclisation of amino acid residues
to convert them to Dha, Dhb amino acids (40, 147). The lanT- type ABC transport had
conserved domains (peptidase, ATPase, permease), therefore it had both functions of removal
of the leader sequence and secretion of the matured peptide (39, 138, 148). A cluster of 3
proteins (23PaeF, 23PaeG, 23PaeH) formed an ABC transport type II that was an immunity
system (137).
Figure 4.12. The paenicidin A gene cluster found in the genome of isolate #23. (A) Gene
organisation of the gene cluster. (B) MSA of the mersacidin precursors of #23 (23paeA)
and P. polymyxa NRRL B-30509. Two precursors were 100% identical. (C) The MSAG
between the paenicidin A gene cluster of isolate #23 and P. polymyxa NRRL B-30509.
Two gene clusters shared 100% sequence similarity.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
92
Table 4.11. Structural organisation of paenicidin A gene cluster found in the genome of
isolate #23
Protein Description Functions Similar proteins*
23paeA paenicidin precursor precursor paeA
23paeH transporter ATP binding protein immunity paeF
23pae G ABC transporter membrane-bound subunit
transportation paeE
23pae F ABC transporter membrane-bound subunit
transportation paeG
23pae D lantibiotic dehydratase modification enzymes
paeB
23paeC lantibiotic ABC transporter transport paeT
23paeB lantibiotic cyclase modification enzyme
paeC
* The proteins were from the paenicidin A gene cluster of P. polymyxa NRRL B-30509.
4.2.9. Prediction of gene organisation of biosynthetic gene clusters of novel bacteriocins
4.2.9.1. The novel lantibiotic gene cluster (found in the genome of isolate #08)
The biosynthetic gene cluster of a novel lantibiotic (named 08lan) was found within the isolate
#08’s genome [Figure 4.13]. Presence of the lanM-type enzyme within the gene cluster
indicated that this bacteriocin was a lantibiotic subclass Ib (40).
The 08lan lantibiotic gene cluster was 5,977 bp in length, flanked by 2 terminators and
consisted of 4 ORFs encoding 4 proteins 08lanA-D [Figure 4.13, Table 4.12]. The precursor
bacteriocin (08lanA) had 78 aa, comprising a 48 aa leader sequence and a 35 aa core sequence
at the N-terminal. The amino acid sequence was similar to lichenidin A1 (37% similarity) and
lichenidin VK21A2 (43% similarity), but had 100% identity to an uncharacterized
mersacidin/lichenicidin family type 2 lantibiotic (WP_052587026.1) in the NR database
[Figure 4.6]. This novel lantibiotic possibly matured under the modifications processes under
the catalysis of a protease (08lanD) for cleavage of the signal sequence and a lanM-type
enzyme (08lanB) to modify the core sequence. The 08lanC shared 42.42% similarity to a
multidrug resistance ABC transporter ATP-binding/ permease protein, BmrA, and possessed

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
93
5 transmembrane helices in the structure suggesting a possibility of either secretion and/or an
immunity function (149, 150)
Figure 4.13. The novel lantibiotic found in the genome of B. amyloliquefaciens # 08.(A)
Gene organisation of the gene cluster. (B) Precursor bacteriocin coding sequence and
amino acid translation. The amino acid sequence of precursor bacteriocin was
highlighted in green colour. The grey box indicated the presence of the ribosomal
binding site (RBS) and start codon (methionine).
Table 4.12. Structural organisation of novel lantibiotic gene cluster found in the genome
of isolate #08
Proteins TMH* Description Function Similar proteins in uniref90 database
08lanD 0 serine protease removal of leader sequence
28.22%
08lanC 5 ABC transporter ATP-binding/permease protein
secretion 42.42%.
08lanB 0 mersacidin modifying enzyme
modification 35.32%
08lanA 0 bacteriocin precursor precursor -
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
94
4.2.9.2. The novel lantibiotic gene cluster (found in the genome of isolate #23)
The biosynthetic gene cluster of a novel lantibiotic (named as 23lan) was found within
scaffold #1 of P. polymyxa #23’s genome. The whole gene cluster included two parts; short
DNA regions containing 3 ORFs that inserted in-between the S-layer glycan biosynthetic gene
cluster, and a part containing the ORFs borrowed from S-glycan gene cluster [Figure 4.14].
The S-layer glycan gene cluster is responsible to produce transmembrane glycans that serve
as antigens on the bacterial surface and exhibits no antimicrobial activity (151). The presence
of two typical lantibiotic modification enzymes; lanB and lanC within the gene cluster
indicated that this bacteriocin was a lantibiotic of subclass Ia (40)
The insert region including 3 ORFs; a precursor bacteriocin (23lanA), a lanB-like enzyme
(23lanB) and a lanC-like enzyme (23lanC), was flanked by 2 terminators. The precursor
(23lanA) was unable to be detected by the BAGEL, only elucidated by manual annotation by
reducing the threshold value (cut off at least 50 amino acids) for gene calling. The precursor
bacteriocin had 53 amino acids, consisting of a 29 aa signal sequence at the C-terminus and
24 aa core sequence at the N-terminus with a theoretical MW of 2375.2 Da. The sequence
was unique on the BACTIBASE database and shared low identity (45%) with a hypothetical
protein, PAC_2c01280, found in Paenibacillus alvei DSM23 (EJW18364.1). Two proteins;
23lanC, 23lanB were annotated as lanB-like dehydratase and lanC-like cyclase which
modified the core sequence.
Other functional proteins were found on the S-layer gene cluster. Two ORFs (23lanE and
23lanF) located at upstream of the insert region formed a two-component ABC transport for
bacteriocin secretion. The 23lanD containing 15 transmembrane helicases was possibly an
immunity protein (149, 152). However, it contained two conserved domains (Wzy_C
(IPR007016) and TPR-like helical_dom_sf (IPR011990)) which were annotated without the
evidence of immunity function. There was no ORF at the surrounding of the insert regions
functioning in the removal of the leader sequence.
The gene cluster was flanked by two terminators; one was at upstream of the ABC transport
and one was at downstream of 23lanA. Based on these terminators, the gene cluster was
temporarily predicted with a length of 12,147 bp, consisting of 6 ORFs encoding 6 proteins
(23lanA-F) [Table 4.13].

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
95
Figure 4.14. The novel lantibiotic gene cluster found in the genome of isolate #23.(A)
Gene organisation of the gene cluster. (B) The MSAG between the lantibiotic gene
cluster of isolate #23 and S-layer glycan. It indicated the insert regions in-between of
S-layer glycan gene cluster (in the box). (C) Precursor coding sequence and amino acid
translation after manual prediction on Snapgene. The grey box indicates the presence of
the ribosomal binding site (RBS) and start codon (methionine).
Table 4.13. Structural organisation of novel lantibiotic gene cluster found in the genome
of isolate #23
Proteins TMH Description Function Similar proteins in uniref90 database
23lanA 0 precursor bacteriocin precursor -
23lanB 0 lanB-type enzyme (PF04738, PF14028)
maturation V9W9P1_9BACL (65.4%); P. larvae DSM 2543
E3EGY4_PAEPS (60.2%); - P. polymyxa (strain SC2)
23lanC 0 lantibiotic biosynthesis protein (PF05147)
maturation V9WAJ5_9BACL (63.1% ); P. larvae DSM 25430
23lanD 15 polymerase immunity E3EAL6_PAEPS (88.4%); P. polymyxa (strain SC2)
23lanE 6 transport permease protein (PF1061)
transportation A0A0M0WD78_9BACI (85.3%); Bacillus sp. FJAT-21945
23lanF 0 ABC transporter, ATP-binding protein (PF00005; PF14524)
transportation C6J2R6_9BACL (68.15%) Paenibacillus sp. oral 786 str. D14

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
96
4.2.9.3. The sactipeptide gene cluster (found in the genome of isolate # 23)
The biosynthetic gene cluster of a novel sactipeptide (23sac) was found within scaffold #7 of
#23’s genome [Figure 4.15]. The presence of a radical SAM enzyme within the gene cluster
indicated that this gene cluster related to a sactipeptide (143).
The 23sac gene cluster was 8556 bp in length, consisting of 8 ORFs encoding 8 proteins,
23sacA-H [Figure 4.15, Table 4.14]. It was flanked by 2 terminators, one was downstream of
23sacG and one was at upstream of 23sacH. The precursor bacteriocin (23sacA) had 61 aa,
comprising a 19 aa leader sequence and a 42 aa core sequence. The precursor sactipeptide
was 100% identical to an uncharacterized putative bacteriocin, CLI_3235 family
(A0A378XQ17-P. polymyxa) but unique on BACTISBASE database. The 42 aa core
sequence was cysteine-rich with a theoretical MW of 4,455 Da and a pI of 7.73.
The maturation of the sactipeptide was probably initiated by a subtilase-family serine protease
(23sacH) which was responsible for cleavage of the signal sequence (153), followed by
modification of the core sequence by 2 enzymes; the radical SAM protein (23sacD) and an
‘acyl carrier protein homolog’ (TIGR04069) (23sacF) (154-156). The matured peptide was
then secreted by a putative ABC transporter (23sacC). The 23sacE was a hypothetical protein
belonging to IGR04066 family. This protein family contained subunits of an H+-transporting
two-sector ATPase, suggesting a function in immunity. The 23sacG was annotated as ‘muscle
M-line assembly protein unc-89 uncoordinated protein with unknown function for bacteriocin
biosynthesis.
Figure 4.15. The novel sactipeptide gene cluster found in the genome of P. polymyxa #23 and
the precursor sequence.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
97
Table 4.14. Structural organisation of novel sactipeptide gene cluster found in the
genome of isolate #23
Proteins TMH*
Description Function Similar proteins in uniref90 database
23sacA Precursor bacteriocin precursor -
23sacB ECF subfamily RNA polymerase sigma-24 subunit
unknown A0A378XN73 (99.3%); P. polymyxa
23sacC 6 Multidrug ABC transporter protein
transportation A0A378XMM5 (100%); P. polymyxa
23sacD 0 Cys-rich peptide radical SAM maturase TIGR04068
modification A0A2S6NY22 (95.4%); P. peoriae
23sacE 0 Protein with domain TIGR04066
immunity UPI000D2F5CC2 (10%); P. polymyxa
23sacF 0 Peptide maturation system acyl carrier-related protein (TIGR04069)
modification A0A378XN50 (100%); P. polymyxa
23sacG 0 Muscle M-line assembly protein unc-89 Uncoordinated protein 89
unknown A0A378XQ26 (100%); P. polymyxa
23sacH 0 Subtilin-like serine protease removal of leader sequence
A0A378XQL5 (100%); P. polymyxa
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).
4.2.9.4. The lassopeptide gene cluster (found in the genome of isolate #23).
This gene cluster of a lassopeptide (named as 23las) was located within scaffold 0 of #23’s
draft genome (namely 23las) [Figure 4.16]. Bioinformatic analysis using whole 23las amino
acid sequence 40% identical to the paeninodin gene cluster found in P. dendritiformis C454
(157). This biosynthetic gene cluster was 9233 bp in length and consisted of 8 ORFs encoding
8 proteins; 23lasA-H [Figure 4.16, Table 4.15]. The gene cluster was flanked by a promoter
locating upstream of 23lasI and a terminator at downstream of 23lasB. The precursor
lassopeptide (23lasA) comprised a deduced 21 aa signal sequence and a 21 aa core sequence
that has a predicted MW of 2413.1 Da. The 23lasA precursor bacteriocin was found to be

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
98
unique on the BACTIBASE database, but highly identical (difference in 1 amino acid) to an
uncharacterized lassopeptide ‘papoA’ of P. polymyxa CR1 (158, 159).
The lassopeptide maturation frequently relates to the modification steps; removal of leader
sequence by an ‘ATP-dependent cysteine protease’ (named as B protein), and a lassopeptide
cyclase (C protein) fold the amino acid residue at N-terminus of the core sequence and
attached to an Asp/Glu residue on the side chain to form a macrolactam ring (38, 160, 161).
The protein B can present at a “split” form; B1, B2 (162). In this gene cluster, B1-protein
(23lasE); B2 protein (23lasD) and C protein (23lasG) were found. The 23lasF was an HPr
kinase/phosphorylase that was responsible for the regulation (159). It regulated the operon by
first recognition of the precursor, binding and then activating the modification processes.
Additionally, a sulfotransferase protein (23lasH) transferred and attached sulphur moieties to
the side chain (159). The lassopeptide was secreted by an ABC transport (23lasB) which also
served as an immunity protein. A kinase (23lasD) was responsible to regulate the operon's
transcription.
Figure 4.16. The lassopeptide gene cluster found in the genome of isolate #23. (A) Gene
organisation of the gene cluster. (B) MSA of the lassopeptide precusor of #23 (23paeA)
and known lassopeptide precursors (baceA from B. cereus VD115, thcoA from
Thermobacillus composti KWC4 (163), padeA from P. dendritiformis C454, papoA
from P. polymyxa CR1(157)). (C) The homology tree across the precursor sequences.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
99
Table 4.15. Structural organization of novel lassopeptide biosynthetic cluster found in
isolate #23
Proteins TMH*
Description Function Similar proteins in uniref90 database
23lasA Precursor bacteriocin precursor -
23lasB 5 ABC transporter ATP-binding protein/ Permease (PF00005.27; PF00664.23)
Transportation, immunity.
A0A378XVV8 (97.7%); P. polymyxa
23lasC 0 Uncharacterized nucleotidyltransferase
tailing A0A378XWG9 (43.1%); Paenibacillus sp. BC26
23lasD 0 Lasso peptide biosynthesis B2 protein
maturation A0A3G8R0N7 (95.7%); P. polymyxa 152
23lasE 0 Coenzyme PQQ synthesis protein D (PqqD) (protein B1)
maturation A0A378XW98 (98.0%); P. polymyxa
23lasF 0 HPr kinase/phosphorylase regulation A0A378XTM6 (98.5%); P. polymyxa
23lasG 0 Asparagine synthase (protein C)
maturation A0A378XUA7 (98.7%) P. polymyxa
23lasH 0 Protein-tyrosine sulfotransferase (PF09037.5 )
tailing A0A378XVA8 (97.2%); P. polymyxa
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).
4.2.9.5. The thiopeptide gene clusters (found in the genomes of isolates #06 and #13).
The thiopeptide gene cluster was located within the genomes of both #06 and #13 [Figure
4.17; Table 4.16]. The thiopeptide gene clusters in #06 and #13 were the same. Therefore it
was named as 13thio gene cluster.
This 13thio gene cluster was 12 kbp in length and was comprised of 13 ORFs encoding 13
proteins, 13thioA-K [Figure 4.17, Table 4.16]. The precursor (13thioA) had 51 amino acids,
comprising a 31 aa leader sequence at the C-terminus and a 21 aa core sequence at the N-
terminus. The sequence was unique on BACTIBASE database but 100% identical with an

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
100
“uncharacterized thiopeptide" of Bacillus species” on the non-redundant (NR) database. The
core sequence had a theoretical MW of 1884.97 Da and theoretical pI of 6.39.
Thiopeptide commonly matures via heavy modification processes; i) cyclization, dehydration,
or oxidation of Ser, Thr, and Cys residues on the precursor to form oxazole ring; ii) conversion
of the Ser and Thr to Dha, Dhb, Lan and MeLan residues; iii) formation of the central six-
membered ring along with the elimination of the signal sequence; iv) additional modifications
on side chain like oxidations, cyclizations, methylations, tailing of indolic or quinaldic acid
moieties (164). To find these highly conserved proteins in the genome of isolate #13, these
proteins were acquired from the ‘nosiheptide genes clusters’ (165), then searched against the
genome of isolate #13 to locate homology CDS. The proteins included a pair of NosG (SpaB
C-terminal domain, PF14028) and nosF (Lant_dehyd_C, PF04738), a pair of NosD (YcaO
domain, PF02624) and nosE (PF00881), and the pair of nosO and nosH. After searching, two
ORFs (13thio G, and 13thioF) were found to be homology with NosG and nosF which were
responsible to convert amino acid to lantibiotic-typical amino acids through dehydration and
cyclisation. A pair of (11thioD and 11thiE) were homology to (NosD and nosE) catalysing to
form the azole rings; and (13thioO, 13thioH) were homology to (nosO and nosH) that
functioned in folding the peptide chain to form a six membered ring and also elimination of
the leader sequence. The thiopeptide was secreted by an ABC transport system (formed from
13thioI, and 13thioJ). There was a transcriptional regulator (13thioK) locating at end the of
the gene cluster. However, there was no immunity gene found so it was unclear if this cluster
had the potential to produce an antimicrobial.
Figure 4.17. The novel thiopeptide gene cluster found in the genomes of B. amyloliquefaciens
#06 and #13.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
101
Table 4.16. Structural organization of the novel thiopeptide biosynthetic cluster in
isolate #06 and #13
Proteins TMH*
Description Function Similar proteins in uniref90 database
13thioA Precursor bacteriocin precursor -
13thioB 0 Lantibiotic dehydratase domain protein (protein B) PF04738
dehydration of
aa residue
UPI0006246BFE (99.7%); B. amyloliquefaciens
13thioC 0 Lantibiotic biosynthesis dehydratase C-term (PF14028)
cyclisation of
aa residue
A0 A1G6Y285; (24.8%); Aquimonas voraii
13thioD 0 Hypothetical protein unknown UPI00084A0EF5 (100.0%); B. amyloliquefacines
13thioE 0 TOMM precursor leader sequence-binding protein (PF02624)
formation of azole
UPI0006246BFE; (99.7%); B. amyloliquefacines
13thioF 0 Lantibiotic biosynthesis dehydratase C-term (PF14028)
formation of azole
J8HJV6 (51.3%); B. mycoides
13thioG 0 SagB/ThcOx family dehydrogenase (PF00881); putative nitroreductase MJ1384
formation of azole
UPI00052AFFE7 (100%); B. amyloliquefacines
13thioH 0 TOMM precursor leader sequence-binding protein (TGR02603)
formation of pyrine ring.
UPI00104D7783 (25.7%); Tumebacillus sp
13thioI 0 ABC transporter ATP-binding protein
transportation UPI000653513B (100%); Bacillus sp.
13thioJ 6 ABC transporter permease transportation UPI00052AA050 (100%); Bacillus sp.
13thioK 0 Transcriptional regulator regulator I2C213 (93.8%); B. amyloliquefaciens
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
102
4.2.9.6. The thiopeptide gene cluster (found in the genome of isolate #11).
This novel thiopeptide gene cluster (namely 11thio) was located within scaffold 8 of isolate
#11’s draft genome [Figure 4.18]. The gene organization was like the thiopeptide gene cluster
found in #13 and #06 but with some variance.
Figure 4.18.The novel thiopeptide found in the genome of B. amyloliquefaciens #11
Table 4.17. Structural organization of the novel thiopeptide biosynthetic cluster found
in isolate #11
Proteins TMH*
Description Function Similar proteins in uniref90 database
11thioA precursor bacteriocin precursor
11thioB 0 lantibiotic dehydratase protein (PF04738)
maturation A0A369C8Z1 (100%); B. amyloliquefaciens
11thioC 0 unknown unknown -
11thioD 0 unknown unknown -
11thioE 0 bacteriocin biosynthesis docking scaffold, SagD family (PF02624)
maturation J8I0L0 ( 34.5%); B. mycoides
11thioF 0 sagB/ThcOx family dehydrogenase (PF00881)
maturation A0A418UV33; Actinomyces sp. 2119
11thioG 0 lantibiotic biosynthesis dehydratase C-term (PF14028)
maturation A0A1M5UR24 (39.9%) Caloranaerobacter azorensis DSM 13643
11thioH 0 sagB-type dehydrogenase enzyme
maturation A0A369CBC6 (100%); B. amyloliquefaciens
11thioI 0 hypothetical protein unknown
11thioJ 6 ABC-2 type transport system ATP-binding protein (PF00005)
transportation A0A369C905 (100%); B. amyloliquefaciens
11thioK 0 ABC transporter permease transportation A0A2I5JX77 (100%); B. amyloliquefaciens

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
103
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).
The 11thio gene cluster was 13,386 bp in length and consisted of 11 ORFs, encoding 11
proteins 11thioA-K [Figure 4.18, Table 4.17]. The precursor (11thioA) had 72 aa, comprised
of a signal sequence at the C-terminus and a 16aa core sequence at the N-terminus. The core
sequence had a theoretical MW of 1777.98 Da and pI of 5.23, was unique on BACTIBASE
database but identical with an “uncharacterized thiopeptide-like bacteriocin” found in B.
amyloliquefaciens (H57 Ga0082361_104/ KACC 18228) on the NR database.
Similar annotation like the 13thioeptpide gene cluster, the pair (11thio G, and 11thioF) were
found to be homology with (Nos G and nosF), pair of (11thioD and 11thiE) were homology
to (NosD and nosE); and (11thioO, 11thioH) were homology to (nosO and nosH). A two-
component ABC transport (clustered from 11thioJ and11thioK) was responsible to secrete the
bacteriocin (150). Two hypothetical proteins (11thioC, 11thioI) were unknown functions,
possibly serving as immunity proteins because this kind of functional protein was still not
found within the gene cluster.
The 11thio gene cluster was flanked by a promoter upstream of 11thioA and a terminator
located downstream of 11thioK. Within the gene cluster, 11thioA itself was flanked by a
promoter and Rho-independent terminator; there was a second promoter detected upstream of
11thioB. Therefore, it suggested that the 11thioA was first transcribed, subsequently serving
as an enhancer to promote the expression of the whole operon using the second promoter.
4.2.9.7. The two-component lantibiotic gene cluster (found in genome of isolate #13).
This lantibiotic gene cluster was found in #13’s draft genome (namely 13lan) [Figure 4.19].
It was 15,526 bp in length, was comprised of 11 ORFs; 13lanA-J [Figure 4.19, Table 4.18].
There were 2 precursors (13lanA1; 13lanA2) within the gene cluster. The 13lanA1 precursor
was found to be 100% identical to ‘Plantaricin C family lantibiotic’ from Bacillus species
(WP_003154891.1). The 13lanA2 had low identical (27%) to ‘lichenicidinVK21A2’ (B.
licheniformis strain DSM 13). The 13lanA2 had 58 aa, comprising a leader sequence at the
C-terminus and a core sequence at the N-terminus via a conserved processing site of two
glycine residues (166, 167).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
104
There were two lanM-like enzymes (13lanH, 13lanJ) that were responsible to modify the
precursor bacteriocins; therefore, each of the enzymes may modify one precursor. This kind
of gene distribution was also typical in the biosynthetic gene cluster of two-component
lantibiotics (168). A LanT-type ABC transport (13lanI), which contained a C39 peptidase
domain (PF03412) and ABC_tran (PF00005) functioned in cleaving off the leader sequence
and secreting the mature bacteriocin. Another ABC transport (clustered from 13lanD and
13lanE) may serve as an immunity system that protected the host from bacteriocin activity
(169). Besides, a lanKR-type two-component signal transduction system (clustered from
13lanB and 13lanC) regulated the transcription of the bacteriocin gene cluster (170). Another
two-component regulatory system (13lanF, 13lanG) was also found that also played a role in
the regulation of bacteriocin production.
Figure 4.19. The two-component lantibiotic gene cluster found in the genome of isolate
#13. (A) Gene organisation of the gene cluster. (B) MSA of the two precursors of #23
(23lanA1, and 23lanA2) and lichenicidine A2. (C) The homology tree across the
precursor sequences. Tree and MSA were calculated by DNAman software.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
105
Table 4.18. Structural organization of the novel two-component lantibiotic in isolate
#13
Protein TMH*
Description Function Similar proteins (on Uniref90 database)
13lanB 0 Transcriptional regulatory protein YvrH (PF00486, PF00072)
transcription P94504 (36.56%); Bacilli
13lanC 2 Sensor histidine kinase YbdK (PF00512)
transcription O31433 (25.42%); Bacillii
13lanD 0 Lantibiotic ABC transporter ATP-binding protein (PF00005)
transportation Q5WIQ7_BACSK (46.01%); B. clausii KSM-K16
13lanE 6 Lantibiotic ABC transporter/ permease (PF12730.7)
modification A0A1W6HGX2 (97.1%); B. venezensis
13lanF 2 Sensor histidine kinase ComP (PF02518)
transcription Q99027 (29.49%); Bacillaceae
13lanG 0 Transcriptional regulatory protein ComA (PF00072)
transcription P14204 (41.23%); Bacilli
13lanA1 Precursor precursor -
13lanH 0 Lantibiotic modifying enzyme (PF05147)
modification Q65DC3_BACLD; (32.07%); B. licheniformis DSM 13
13lanI 5 MrsT protein (PF00005; PF03412)
transportation and cleaving off the leader sequence
Q9RC21_BACSY (35.49%); Bacillus sp. strain HIL-Y85/54728
13lanA2 Structural bacteriocin (PF04604, PF16934)
precursor -
13lanA3 Precursor precursor -
13lanJ 0 Lantibiotic modifying enzyme (PF05147.13, PF13575.6)
modification Q62NU6_BACLD (27.32%); B. licheniformis DSM 13
* TMH, number of transmembrane helices predicted by the TMHMM server, v.2.0
(http://www.cbs.dtu.dk/services/TMHMM/).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
106
4.3. Summary of results and discussion
There is an urgent need for novel antimicrobial substances to be identified for new antibiotic
development when the majority of antibiotics currently used in the treatment of hospital-
acquired infection are found to be less effective with the development of resistance. Much
effort has been made to synthesize in vitro novel antimicrobial agents; nevertheless, nature is
still a rich and cheap source to search for novel antimicrobials substances. In this
investigation, 6 marine Bacillus/Paenibacillus isolates were selected to search for novel
bacteriocins, including; 1 P. polymyxa #23; 4 B. amyloliquefaciens (#06, #08, #11, #13); and
1 B. halotolerans (#01). These bacteria were chosen because they displayed antimicrobial
activity against a wide range of problematic pathogens, including Gram-positive bacteria, and
some serious hospital-acquired Gram-negative bacteria such as Campylobacter, Klebsiella,
and Pseudomonas.
First, genomes were sequenced, assembled and annotated and then comparisons were
performed to visualize the difference in genomes within marine isolates versus closely related
bacteria derived from terrestrial sources. All genome comparison calculations (first
employing calculation of ANI across the genomes and later confirming the finding from ANI
calculation by SBCT tool and MAUVE) were in agreement that genomes of B.
amyloliquefaciens were significantly homologous, followed by B. halotolerans, and the P.
polymyxa displayed the lowest similarity in genomes. Also, analysis of the frequency
distribution of CDS across the genomes of each of species (including both marine strains and
terrestrial strains), verified by GET-HOMOLOGOUS, indicated the highest diversity of the
gene repertoire of P. polymyxa when compared with B. amyloliquefaciens, and B.
halotolerans, emphasizing the potential of this species in the discovery of novel antimicrobial
agents. The high numbers of strains-specific CDS in P. polymyxa genomes can be derived
from prophage sequences or external movable DNA that infected to the host by horizontal
transfer event (171, 172). The infection can be observed via the presence of the deleted blocks
within LCBs of the genome from the MAUVE analysis. Due to the larger chromosome, P.
polymyxa can carry high numbers of prophages sequences that often possess unique genes
and genes relating biosynthetic gene clusters of ribosomally synthesized peptides (microcins,
lantibiotic) (141, 172).
In silico prediction of antimicrobial peptide encoding genes from these 6 genomes, by both
BAGEL and antiSMASH, elucidated the presence of 61 gene clusters relating to the

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
107
biosynthesis of lipopeptides (LPs), polyketides (PKs), bacilysin and bacteriocins. The number
of predicted lipopeptides encoding regions was twice the number of polyketide, hybrid, or
bacteriocin encoding regions. This finding is in agreement with earlier studies on drug
discoveries from bacilli. It was reported that 31% of the Firmicutes had gene clusters predicted
to encode antimicrobial peptides, with 70% of the total predictions being for NRPs synthesis
and 30% for hybrid LPs/PKS or PKS (173). The presence of both NRPs and bacteriocin agents
mark a difference between the antimicrobial potential of Bacillus and Lactobacillus isolates,
with the Lactobacillus hosts only carrying genes for bacteriocin synthesis (37)
There were 41 NRPs gene clusters predicted. They related to the biosynthesis of 14 different
types of NRPs compounds. This is because some NRPs were replicated in all Bacillus species,
including LPs (bacillaene, and macrolactin), lipopeptides (surfactin, fengycin, and
bacillomycin D), and siderophores (bacillibactin). These NRPs types of Bacillus were
different from those of Paenibacillus genomes (tridecaptin, polymyxin, putative
paenibacterin, fusaricidin), suggesting that the NRPs are taxon associated. During searches
for the presence of these derived NRPs from other species, we found that these NRPs were
also found in other members of B. subtilis groups (174-176) but not in other members of the
Bacillus group, probably suggesting that they are associated with the B. subtilis group only.
This may indicate that these NRPs gene clusters were acquired in a common ancestor, and
then distributed and maintained within the members of the group as it diversified. However,
the lipopeptide difficidin was only found in genomes of B. amyloliquefaciens but not in the
B. halotolerans genome, suggesting that this lipopeptide is only associated with B.
amyloliquefaciens. This observation is similar to earlier studies which also reported that the
distribution of the fengycin gene cluster was in a narrow subset of the B. subtilis group, or
bacitracin cluster presented in some species of B. subtilis group (such as B. licheniformis, B.
subtilis) (172, 177). The potential novelty of completely new NRPs compounds is less
reported, although the variants of traditional agents are seldom mentioned due to addition/loss
of acyl residue attached on main chains (127). To conclude, the presence of these NRPs
compounds in marine Bacillus/Paenibacillus emphasizes the potential application of these
marine species in pharmaceuticals and agriculture. Several Bacillus-derived NRPs are
approved for use in various treatment, such as immunosuppressive (rapamycin), anti-cancer
(bleomycin, daunorubicin, mitomycin) and even antibiotics (amphotericin B, erythromycin,
vancomycin, polymyxin, tridecaptin) (24, 56). In agriculture, AMPs genes are widely used as
markers to evaluate the application ability of a bacterium as biocontrol. For example, bmyB

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
108
(coding bacillomycin D synthetase B), and fenD genes (fengycin synthetaseD) were used as
markers to evaluate the B. amyloliquefaciens FZB42 (178); or bmyB, fenD, and ituC (iturin
synthetase D) are used to evaluate strains QST713, RGAF51, UMAF6639, RLID 12.1 (179)
The study has shown that marine Bacillus/Paenibacillus are a rich resource for the discovery
of novel bacteriocins, with 53.8% of all the predicted bacteriocins identified as novel or
uncharacterised. The six genomes investigated contained 20 bacteriocin gene clusters, with
each genome encoding at least 3 types of bacteriocins, excepting the genome of B.
halotolerans that contained only a single subtilosin A encoding region. This emphasizes the
high potential of marine B. amyloliquefaciens, and Paenibacillus as abundant pools of
bacteriocins, including novel bacteriocins. Although 20 bacteriocin encoding gene clusters
were found, they were predicted to only encode 13 different types of bacteriocins and these
belonged mostly to class I bacteriocins (lantibiotics, lassopeptide, thiopeptide, sactipeptide,
LAP). There was 1 predicted class II bacteriocin, and no class III. Lantibiotics were the most
common type (4/12) (including mersacidin, paenicidin A, two unknown lantibiotics).
The P. polymyxa #23 derived bacteriocins were more diverse; including 4 bacteriocins
belonging to 3 different subclasses of class I bacteriocins (sactipeptide, lassopeptide, 2
lantibiotics). Three of 4 bacteriocins were novel/uncharacterised and one bacteriocin that has
been previously characterised (paenicidin A), marking this species as the one with the most
potential for seeking novel bacteriocins in the marine environment (at least in this
investigation). This is an unsurprising result as the genome of this species was found to be
highly variable, indicating great flexibility in adapting to environmental changes (last chapter,
mentioned in the study of comparative genomics). Of three novel bacteriocins, the predicted
lassopeptide encoding gene cluster had 40% similarity to a paeninodin encoding gene cluster
found in the genome of P. dendritiformis C454, and the precursor sequence had 1 amino acid
differing to the lassopeptide precursor in P. polymyxa CR1; however, this predicted peptide
is yet to be characterised.
Amongst the 5 sequenced Bacillus isolates, there were 8 gene clusters of bacteriocins found,
including 5 previously characterised ones (mersacidin, subtilosin A, plantazolicin, LCI, and
amylocyclicin). The bacteriocins found in the Bacillus isolates were also different from those
of P. polymyxa. Three bacteriocins (LCI, amylocyclicin and thiopeptide) were found in all B.
amyloliquefaciens isolates, but not in the sequenced B. halotolerans isolate. The LCI
bacteriocin has been also reported in many studies, not only associated with B.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
109
amyloliquefaciens but also found in B. subtilis, suggesting that the presence of this bacteriocin
is associated with the related species of the B. subtilis group. Both amylocyclicin and
thiopeptides are found only in genomes of marine B. amyloliquefaciens, indicating strain-
specific bacteriocins. Therefore, it appears that some of the Bacillus derived bacteriocins may
be specific/restricted to certain species. There were 3 novel bacteriocins (two thiopeptides,
one two-component lantibiotic). All the predicted thiopeptides found in this investigation are
novel. The thiopeptides are highly modified peptides of low MW (< 2 kDa), and their
biosynthesis uses both non-ribosomal synthesis, and ribosomal synthesis. The thiopeptides
are recognised as a promising bacteriocin group for drug development; some of them are
already in clinical trials for the treatment of human infections. Besides, thiopeptides also
exhibit anti-cancer, anti-plasmodial, immunosuppressive, antifungal, and even anti-malaria
activity which address important areas of therapeutic application.
It is interesting to review a number of the performance of the bioinformatic tools that were
used to predict the antimicrobial peptide encoding regions; BAGEL and AntiSMASH.
BAGEL is currently the tool of choice for prediction of putative bacteriocins. It has been
designed to better perform the task when compared with other similar software because it can
detect small ORFs of structural genes which are frequently omitted by other software, and it
predicts the bacteriocins cluster by also interrogating surrounding genes to identify other
genes that may participate in bacteriocin biosynthesis, processing, and export. In this
investigation, it was found that BAGEL was unable to detect the thiopeptide gene clusters,
but AntiSMASH could. AntiSMASH is the tool of choice for prediction of NRPs compounds
in bacterial genomes. However, both software was unable to reveal in detail the gene
organisation of thiopeptide gene clusters and locate the structural gene within the gene cluster.
Therefore, the attempt to locate gene components was supplemented by manual annotation
by i) reducing the minimum peptide length cut off to 50 amino acids for calling genes
encoding precursor. The sequence of precursor peptide comprises of a core sequence of 12 to
17 residues at C- terminus, and a leader sequence of 34 to 55 residues at the N-terminus (164).
ii) using highly conserved sequences of modification enzymes typical for thiopeptide
biosynthesis (like nosF, nosG, nosE, nosD, nosF in nosiheptide cluster) as reference (or driver
sequences) to locate homologous CDS within the cluster. Therefore, prediction of
antimicrobial peptides from Bacillus/Paenibacillus may require a combination of BAGEL,
AntiSMASH, and manual annotation, to fully explore all the gene clusters. However, due to
incompleteness of genomes (draft genomes, with gaps) generated from Illumina sequencing,

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
110
some clusters lack the context of some surrounding gene-determinants, particularly
loss/division of gene components across multiple gaps, into different contigs, is commonly
observed from large gene clusters such as those used to produce non-ribosomally synthesized
peptides. This can cause overprediction of the number of antimicrobial peptide encoding
regions. To avoid this it would be desirable to close genomes and generate complete
assembled genome sequences (e.g., by adding data from long sequence read technologies such
as Pacbio or MinIon) (180, 181), to give a conclusive picture of antimicrobial potential from
marine Bacillus/Paenibacillus. Taken together, the results emphasized that the marine
Bacillus/Paenibacillus from the Vietnam Sea are abundant sources of non-ribosomally
synthesized antimicrobial peptides and bacteriocins, and novel bacteriocins. Although the list
of the compounds and their biosynthetic gene clusters were well-analysed in this study, the
overall picture of antimicrobial potential of these marine Bacillus/Paenibacillu is still unclear
until in vitro characterisation is carried out to validate the findings. Therefore, two of the
bacterial isolates were used to purify the antimicrobial substances produced. That work is
detailed in the next chapters.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
111
Chapter 5 PURIFICATION OF ANTIMICROBIAL PEPTIDES PRODUCED BY
Bacillus amyloliquefaciens #11
5.1. Introduction
Bacillus amyloliquefaciens is a Gram-positive, spore-forming non-pathogenic bacterium. It is
ubiquitous in both the terrestrial and marine environment. This species has two subspecies;
plantarum (commonly associated with plants) and amyloliquefaciens (non-plant associated).
Terrestrially derived B. amyloliquefaciens are well-known producers of enzymes, and
antimicrobial peptides (particularly bacteriocins) (36, 182, 183). The potential of bacteriocins
produced by marine-derived B. amyloliquefaciens is still yet to be exploited in any detail and
therefore they may prove to be a promising reservoir for the discovery of novel bacteriocins.
In the last Chapter 4, B. amyloliquefaciens #11 was isolated from a seaweed sample and
exhibited strong antibacterial activity. The antibacterial activity exhibited by isolate #11
encompassed both Gram-positive pathogens (Staphylococcus sp., Enterococcus sp., Bacillus
sp., Streptococcus sp, Clostridium sp., MRSA) and Gram-negative bacteria (E. coli, S.
Enteritidis, C. coli). Therefore, it was selected for whole genome sequencing to search for
novel bacteriocins. The bacterium was identified based on the 16S rDNA sequence sharing
99.9% of similarity to one of B. amyloliquefaciens FZB42. The FZB42 which is a plant
growth-promoting rhizobacteria, and serves as a Gram-positive bacterium model for
biocontrol in agriculture (184). The comparison of the in silico predicted antimicrobials
peptides from the genome of #11, with the prediction of those encoded by FZB42, indicates
significant similarity in antimicrobial potential between these two bacteria. Both bacteria have
gene clusters predicted to encode the same 8 - 10 types of antimicrobial peptides with high
homology, including non-ribosomally synthesized peptides (siderophore bacillibactin,
difficidin, macrolactin, surfactin, fengycin, bacillomycin D) and two bacteriocins
(amylocyclicin, and LCI). They are only different due to the presence of plantazolicin
bacteriocin and an uncharacterised LPs (nrs gene cluster), which are found only in FZB42,
and a novel thiopeptide which is found only in the genome of #11. The plantazolicin belongs
to the linear azol(in)e peptides (LAP) subclass; both LAP and thiopeptide are two of members
of the TOMM class (thiazole/oxazole-modified microcins). This group is typically small
bacteriocins (MW of less than 2 kDa), highly modified structures containing azoline rings
generated from enzymatic modification of serine/threonine and cysteine residues. By
comparing with LAP bacteriocins, the thiopeptides have the addition of both Dha/Dhb

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
112
residues and a nitrogen-containing six-membered ring; therefore, the gene cluster of
thiopeptide comprised more genes than LAP. Therefore, the heterologous cloning and
expression of a thiopeptide gene cluster is likely to be more challenging than cloning and
expression of other classes of bacteriocins. Thiopeptides have applications in
pharmaceuticals; with some of them used in the treatment of vaginal infections (185).
This chapter aimed to purify the thiopeptide from #11, for characterisation in vitro. To
achieved this aim, a purification procedure was developed including; i) the growth curve
analysis of the bacterium in LPMA to identify at which time-point the maximal antimicrobial
activity against two indicators, L. plantarum and B. cereus, was present; ii) chromatography
(ion exchange chromatography, hydrophobic interaction chromatography, reverse phase-
HPLC) to partially purify the compound, and iii) mass spectrometry (MALDI-TOF MS, N-
Terminal sequencing) to identify the composition. The work presented in this chapter
describes the bacterium, to underpin its use for pharmaceutical, industrial and agricultural
applications.
Figure 5.1. The seaweed from which B. amyloliquefaciens #11 isolated (A). Bacterial
colony morphology of #11 on Muller Hilton agar (B).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
113
Figure 5.2. Antimicrobial activity exhibited by marine isolate #11. (A) Image
demonstrated the antimicrobial activities against some Gram-positive bacteria including
S. aureus (1), MRSA (2), S. faecalis (3), B. cereus (4). (B) Image demonstrated the
activities against some Gram-negative bacteria such as S. Enteritidis (1), E. coli (2), L.
monocytogenes (3); and P. aeruginosa (4).
5.2. Result
5.2.1. Analysis of the bacterial growth curve revealed to production of various
antimicrobial compounds
The growth of B. amyloliquefaciens in LPMB broth was monitored up to 48 hours (30oC, 150
rpm) and sampled over that period. The samples were tested in an antimicrobial assay against
two indicators; L. plantarum and B. cereus [Figure 5.3]. The first indication of antimicrobial
activity in the culture was observed at 8 hours, with inhibition of L. plantarum. The strongest
activity was noted at 12 hours and activity was lost after 24 hours. Activity against B. cereus
was first observed after 12 hours and continued to be observed over all time points, up to 48
hours. It was obvious from these results that the bacterium produced at least two antimicrobial
compounds that peaked in activity, within the culture, after different fermentation times. To
further explore the antimicrobial peptides produced, 12 hour old culture and 24 hour old
culture were collected and purified separately.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
114
Figure 5.3. Antimicrobial activity exhibited from cell-free supernatants of the cultures
collected at different time-points. (A) The spot-on lawn assay was used to detect
antimicrobial activity against L. plantarum on MRS agar; (B) the well diffusion assay
was used to detect the antimicrobial activity against B. cereus on Muller Hilton agar.
cultures at 12 hours (4), at 18 hours (5); at 24 hours (6), 36 hours (7); 42 hours (8), 48
hours (9); (-) uninoculated media was used as a negative control. The activity against L.
plantarum was strongly observed at 12 hours old culture and was lost after 24 hours,
while the activity against B. cereus was first observed at 12 hour-old culture and from
then on continuously observed.
5.2.2. Purification of antimicrobial compounds from 12 hour-old culture elucidated the
presence of amylocyclicin bacteriocin
5.2.2.1. Recovery of bacteriocins from cell-free culture and cell pellet.
The culture lost the antimicrobial activity following proteolytic treatment (by proteinase K or
pronase E), indicating the presence of a proteinaceous antimicrobial compound, probably a
bacteriocin. During purification, we found that both cell-free culture and extract from cell
pellet exhibited similar antimicrobial activity [Figure 5.4]. Therefore, two separate procedures
were set up to recover the antimicrobial compounds.
The bacteriocin present in the cell-free culture was precipitated using ammonium sulphate.
An experiment was first carried out to determine the concentration of ammonium sulphate to
precipitate this bacteriocin. It was found that the bacteriocin precipitated when the ammonium
sulphate was at 40% (w/v) at 5oC. After centrifugation, the protein pellet was collected. The
protein pellet was dissolved in 1X phosphate-buffered saline (PBS) and desalted against milli-

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
115
Q water using a dialysis tubing membrane (protein cut off 2 kDa) (Sigma Aldrich), to yield a
desalted ‘AMS fraction’. The remaining supernatant was also retained, adjusted to pH 2 by
concentrated HCl, and left overnight at 5oC without stirring. The protein precipitate was
collected by centrifugation at 17,500 g for 90 minutes, dissolved in 1X PBS to yield ‘LP
fraction’.
The cell pellet was also collected, then stirred in 70% isopropanol pH 2 at 5oC overnight
(acidified by concentrated HCl). The volume of isopropanol used was 20% of the volume of
bacterial culture. The cell-free solvent was collected by; centrifugation at 4,500 g for 20
minutes; filtered through a syringe membrane (0.45 µm, PES membrane), and then
concentrated in a rotary evaporator to 1/100 volume, to remove the solvent. The concentrate
was adjusted to pH 7 using NaOH, desalted against milli-Q water overnight at 5oC to yield a
desalted ‘fraction CFS’.
All three fractions exhibited antimicrobial activity against L. plantarum [Figure 5.4]. Of them,
the AMP yielded larger inhibitory diameter than LP or CFS. The antimicrobial compounds
presented in LP faction was considered due to the remaining of protein pellet during decanting
of the supernatant after centrifugation
Figure 5.4. Antimicrobial activity against L. plantarum exhibited by the fractions
collected at time-point of 12 hours. AMP: AMP fraction; LP: LP fractions, SF: CFS
fraction; CR: crude bacterial culture; (-) 1X PBS as negative control.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
116
To estimate the size of the antimicrobial peptides, two of the fractions; AMP and CFS, were
loaded on a 16% Tricine-SDS-PAGE and zymography were then performed. After
electrophoresis, one half of the gel was stained with Coomassie blue staining solution, but the
protein bands on stained gel appeared too faint to visualize; although antimicrobial band
appeared clearly on zymogram gel [Figure 5.6 B]. Therefore, the half gel was stained in silver
staining solution to increase the sensitivity of detection. This showed there was only a single
protein band on the gel. This single protein band, visualised by silver staining was at the same
position as the region of activity identified by zymogram. The band and activity derived from
AMP and CFS fractions were similar, with an estimated MW of ~5-10 kDa [Figure 5.6 A].
However, the AMP fraction contained lots of contaminating proteins when compared with
the CFS fraction. Therefore, CFS would be more suitable for use in future experiments for
quick purification of bacteriocin. The CFS fraction was then desalted by either dialysis (cut
off of 2 kDa) (Sigma Aldrich) or Amicon® Ultra Centrifugal Filters with MW cut off of
approximately 3 kDa (Merck Millipore). The desalted sample was then further purified by ion
exchange chromatography.
5.2.2.2. Purification of bacteriocins by ion exchange chromatography revealed the
presence of 1 antimicrobial peptide
To purify the unknown bacteriocin by ion exchange chromatography, two experiments were
first carried out to select the most suitable types of running buffer, types of resin and
concentration of NaCl for elution.
The first experiment showed that the antimicrobial substance was absorbed onto SP sepharose
in either of 20 mM acetate buffer pH 3.6 or pH 4.6 or pH 5.6 or 20 mM phosphate buffer pH
6.6. In contrast, it was not absorbed onto Q sepharose in 20 mM Tris-HCl buffer pH 8 [Figure
5.5A]. This indicated cation exchange chromatography was suitable to purify the bacteriocin,
and this bacteriocin had a high pI value, which may be more than 9. Subsequently, the second
experiment was carried out to determine the concentration of NaCl in the elution buffer. In
this stage, the cation exchange chromatography using SP sepharose was set up using 20 mM
acetate buffer pH 3.6 as running buffer. The result showed that the bacteriocin was eluted in
the presence of 0.2 M NaCl [Figure 5.5B]. Three elutes with 0.2 M NaCl were combined,
desalted by using Amicon® Ultra Centrifugal Filters (cut off of approximately 3 kDa).
MALDI-TOF MS identified two peaks; one with [m/z] = 6.390; one with [m/z]= 3.056 [Figure
5.6 C]. The desalted sample was then separated by RP-HPLC.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
117
Figure 5.5. The experiments to select the resin, type of buffer, and NaCl concentration
in elution buffer for ion exchange chromatography (A) The experiments to select the
resin and buffer. Well #1 to well # 4: SP sepharose and running buffer at pH 3.6, 4.6,
5.6, 6.6 respectively; well # 5: Q sepharose and buffer pH 8. (a) Antimicrobial sample
collected after mixing with the resin for 20 minutes. (b) Elutes of resin by elution buffer
supplemented with 0.5M NaCl. (B) The experiment to determine the concentration of
NaCl in the elution buffer. The cation exchange chromatography was performed in this
experiment using SP-sepharose and 20 mM acetate buffer pH 3.2. (S) The sample
collected after running through the resin, (w) the buffers collected after washing the
resin (twice), and antimicrobial fractions after eluting the resin using buffer
supplemented with 0.1 M (0.1), 0.2 M (0.2), 0.3 M (0.3) to 1M NaCl respectively. The
experiment indicated the antimicrobial was able to be eluted by elution buffer
supplemented with 0.2M NaCl.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
118
Figure 5.6. The MW of antimicrobial peptide presented in 12 hour old culture after
partial purification by cation exchange chromatography. (A) Estimated by Tricine-SDS
PAGE from AMP fraction (1), CFS fraction (2), purified bacteriocin (3) in the
visualization of (B) zymogram gel against L. plantarum (the same antimicrobial bands
presented in AMP and CFS samples indicated the presence of the same peptide). (C)
MALDI-TOF MS detected two protein peaks from the desalted sample obtained from
cation exchange chromatography. The peptide had mass intensity at m/z 6,399.27.
5.2.2.3. Purification of bacteriocin performed by analytical RP-HPLC elucidated the
presence of amylocyclicin
The desalted sample was injected in a C18 column and was operated under a 60 min-
acetonitrile-water gradient. The operation consisted of; 2% buffer B for 5 minutes; 2%-100%
solution B from minute 6-55; 100% solution B from minutes 56-60. Solution A was milli-Q
water + 0.1% trifluoroacetic acid (TFA) and solution B was 100% HPLC-grade acetonitrile
+ 0.1% TFA. The operation was monitored under light wavelengths of 230 nm and 280 nm
[Figure 5.7 A]. The active fraction was eluted at a retention time of 43.9 minutes (against L.
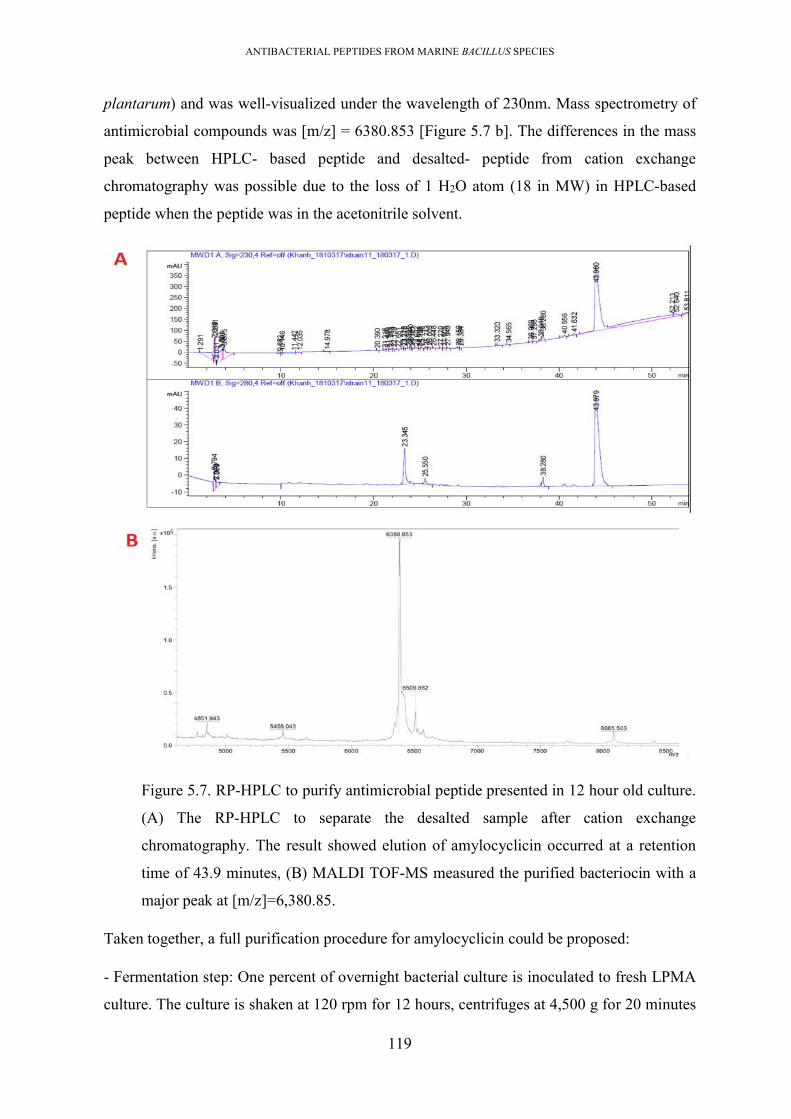
ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
119
plantarum) and was well-visualized under the wavelength of 230nm. Mass spectrometry of
antimicrobial compounds was [m/z] = 6380.853 [Figure 5.7 b]. The differences in the mass
peak between HPLC- based peptide and desalted- peptide from cation exchange
chromatography was possible due to the loss of 1 H2O atom (18 in MW) in HPLC-based
peptide when the peptide was in the acetonitrile solvent.
Figure 5.7. RP-HPLC to purify antimicrobial peptide presented in 12 hour old culture.
(A) The RP-HPLC to separate the desalted sample after cation exchange
chromatography. The result showed elution of amylocyclicin occurred at a retention
time of 43.9 minutes, (B) MALDI TOF-MS measured the purified bacteriocin with a
major peak at [m/z]=6,380.85.
Taken together, a full purification procedure for amylocyclicin could be proposed:
- Fermentation step: One percent of overnight bacterial culture is inoculated to fresh LPMA
culture. The culture is shaken at 120 rpm for 12 hours, centrifuges at 4,500 g for 20 minutes

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
120
to collect both bacterial pellet and supernatant. The bacterial pellet is washed twice with milli-
Q water. The supernatant is filtered through a filter membrane (cut off 0.45 um, PES) to obtain
cell-free supernatant.
- Recovery step: bacteriocin can be recovered from either cell-free supernatant or cell pellet.
However, the cell pellet source contains less contaminating proteins and required less time
for purification than recovery from cell-free supernatant. Precipitating the bacteriocin from
cell-free supernatant can be achieved by using 40% saturated ammonium sulphate (at 5oC
overnight), subsequently centrifuging (17,500 x g, 90 minutes, 5oC) to collect the protein
pellet which is dissolved in 20 mM acetate buffer pH 3.6 (volume of the buffer: volume of
supernatant = 1: 50). Stirring the bacterial cell pellet in 1/5 volume of isopropanol (adjusted
to pH 2 by concentrated HCl) at 5oC overnight, subsequently centrifuging (4500 x g, 20
minutes) followed by filtering of supernatant through a syringe membrane (0.45um, PES) and
concentrating to 1/10 volume. These two fractions can be combined and dialysed against 20
mM acetate buffer pH 3.6 overnight at 5oC using dialysis tubing (cut off 2 kDa).
- Purification step: The desalted sample is passed through SP sepharose column for a cation
exchange chromatography, using 20 mM acetate buffer pH 3.6 as running buffer. The
chromatography can be operated automatically or manually. The automatic chromatography
uses prepacked HiTrap SP HP cation exchange chromatography column (GE Healthcare Life)
and with the aid of ÄKTA pure chromatography system (GE Healthcare Life). The operation
is under a gradient of solution A (20 mM acetate buffer pH 3.6) and solution B (20 mM acetate
buffer pH 3.6 + 1 M NaCl), visualized under the wavelength of 230 nm. The manual
chromatography used SP Sepharose Fast Flow cation exchange chromatography resin (Sigma
Aldrich). The operation included some steps; (i) packing the SP sepharose into an empty
column; (ii) washing the resin by adding milli-Q water (3CV) to remove ethanol; (iii)
equilibrating the resin by adding 20 mM acetate buffer pH 3.6 to column (3 CV); (iv) loading
the sample by dripping the sample into the resin; (v) washing resin (3CV) with 20 mM acetate
buffer pH 3.6; (vi) washing resin (3CV) with 20 mM acetate buffer pH 3.6 supplemented with
0.1 M NaCl to eliminate the weakly- bound proteins; and (iv) eluting the bacteriocin by adding
20 mM acetate buffer pH 3.6 supplemented with 0.2 M NaCl (3CV).
- All antimicrobial fractions are combined, desalted and concentrated using Amicon® Ultra
Centrifugal Filters (MW cut off of approximately 3 kDa)

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
121
- Injecting desalted sample into an RP-HPLC C18 column. Operation is carried out using a
C18 Column with solution A (milli-Q water +0.1% TFA) and solution B (100% HPLC-grade
acetonitrile + 0.1%TFA). Operation condition includes 2% solution B for 5 minutes; 2%-
100% solution B from minute 6-55 minutes; 100% solution B from minutes 56–60 minutes
with visualization under a wavelength of 230 nm. The pure bacteriocin is eluted at a retention
time of 43.9 minutes.
5.2.2.4. The amylocyclicin, a heat and pH stable bacteriocin exhibited activity against
only Gram-positive bacteria
The purified bacteriocin was used to determine antimicrobial activity against a panel of
bacterial pathogens. The result showed that it could depress the growth of only Gram-positive
bacteria [Figure 5.8 C, Table 5.1]. It lost activity after proteolytic treatment with protease E,
proteinase K, but was resistant to trypsin, lipase and catalase treatment. The bacteriocin was
thermo-stable to 80oC for 1 hour, only losing activity at 100oC [Figure 5.8 A, B]. From these
finding, along with inspection of the bacteriocins predicted from the genomic data, it was
determined that this bacteriocin was amylocyclicin (133).
Figure 5.8. Properties of amylocyclicin. Thermal stability; activity remained after being heat
at 80oC for 1 hour. (B) Enzymatic degradability against pronase E (proE) and proteinase K
(proK). (C) antimicrobial activity of amylocyclicin against some indicators; from left to
right and from top to bottom was C. perfringens, S. aureus, MRSA, L. plantarum, E.
faecalis, E. coli, C. jejuni, S. Enteritidis, P. aeruginosa.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
122
Figure 5.9. (A) Sequence alignment between amylocyclicin from isolate #11 genome
(11amyA) and FZB42 strains. (B) Maturation of amylocyclicin. The amylocyclicin
precursor included a 48 aa N-terminal leader sequence, the core sequence. The
maturation related to the cleavage of the signal sequence at the cleavage site between
E−1 and L1 (vertical arrow), and circulation of core sequence between W64 and L1.
Six tryptic fragments and their mass were detected after digestion by trypsin including
the cyclization site WL (bold letters). Image B was obtained from Scholz et al. (2014)
(133)
Table 5.1. The antimicrobial spectrum of amylocyclicin produced by isolate #11
Indicators Medium used Antagonistic activity
S. aureus MHA ++
MRSA MHA ++
B. cereus MHA +
L. monocytogenes MHA +
C. perfringens MHA ++
L. plantarum MRS +++
E. faecalis (VRE) MRS ++

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
123
Indicators Medium used Antagonistic activity
S. Enteritidis MHA -
E. coli MHA -
C. jejuni MHA -
C. albicans MHA -
5.2.3. Purification of antimicrobial substances from 36 hours-old culture showed the
presence of three antimicrobial compounds
5.2.3.1. Recovery of antimicrobials compounds from the cell pellet
The novel thiopeptide was not found from 12 hour culture; therefore, the 36 hour culture was
investigated alternatively. The antimicrobial activity against B. cereus in the culture
supernatant remained after proteolytic digestion by trypsin, pronase E, proteinase K, but no
antimicrobial band(s) could be detected on zymogram plates against B. cereus.
The antimicrobial substances were collected from 36 hour cell pellet. The cell pellet collected
from 36 hours old culture was stirred in acetonitrile (70% pH 2 (acidified by TFA)) overnight.
Acetonitrile and TFA were used instead of isopropanol and acid HCl because they were
volatile solvents, therefore, easily eliminated during rotary evaporation. The mixture was
centrifuged at 5,000 g for 20 minutes and the supernatant was retained. A cell-free solvent
fraction was obtained by filtering the supernatant through a PES syringe membrane (0.45µm).
MALDI-TOF was used to identify peptides presented in the cell-free solvent fraction,
detecting three peak clusters from this sample [Figure 5.9 A]. Based on the mass identity of
NRPs compounds mentioned in the literature (186), one peak cluster with mass of m/z 1002
-1068 indicated the presence of an iturin/surfactin with its iso-protons; a peak with a mass
intensity of m/z 800 was an unknown compound; and one cluster of peaks at a mass range of
m/z 1,500 -1,543 was consistent with fengycin or possibly thiopeptide as both fengycin and
thiopeptide have similar MW.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
124
Figure 5.10. Matrix-assisted laser desorption ionization time-of-flight (MALDI-TOF)
mass spectrometry (MS) of (A) CFS fraction from 24-hour culture, showing compounds
in the MW range of 800 – 2000 Da. The m/z 1002 -1068 peaks related to iturin, while
the peaks at m/z 1500 -1543 indicated the presence of an unidentified peptide, with an
MW similar to fengycin and thiopeptide; (B) MALDI-TOF trace obtained from the
purified compound with a mass of m/z 1500 -1543.
A
B

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
125
5.2.3.2. RP-HPLC purification of antimicrobial compounds and peptide identification
The cell-free solvent fraction was concentrated to 1/20 volume under reduced pressure in a
rotary evaporator. During concentration, a precipitate appeared. This precipitate was
separated from the supernatant by centrifugation, washed twice with milli-Q water and
dissolved in 50% acetonitrile (it was insoluble in water or PBS buffer). This dissolve material
had antimicrobial activity against Gram-positive bacteria (but not Staphylococcus species).
During storage at 5oC or -20oC this dissolve formed aggregated again and was insoluble in
50% acetonitrile. Therefore, the freshly-dissolved aggregate was injected into an RP-HPLC
(C18 column) and the separation was carried out under a 60 minutes acetonitrile-water
gradient cycle including; 30% buffer B for 5 minutes, 30%-50% of buffer B from minute 6 to
minute 0; 50%-100% buffer B from minute 11–minute 55; 100% buffer B for minutes 55-60.
The chromatography elutes were visualized under wavelengths of 230 nm, of 214 nm and 280
nm. The antimicrobial compound was collected at a retention time of 44.618 minutes,
corresponding to approximately 80% acetonitrile. The MALDI-TOF MS revealed a major
peak cluster at an intensity of m/z 1417-1663; indicating the MW of the compound and its
isoprotons [Figure 5.9 B]. This peptide was only detected under UV wavelength of 214nm
and 230 nm but not under the wavelength of 280 nm, indicating a peptide [Figure 5.10]
Taken together these findings suggest that this antimicrobial peptide was highly hydrophobic
and exhibited an antimicrobial spectrum against Gram-positive bacteria. We failed to
determine the peptide sequence using Edman degradation N-terminal sequencing.
Figure 5.11. RP-HPLC used to separate the freshly dissolved aggregate.The antimicrobial
was eluted at a retention time of 44.6 minutes

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
126
Table 5.2. Antimicrobial spectrum of purified thiopeptide supposed
Indicators Media Antagonistic activity
Staphylococcus aureus MHA -
MRSA MHA -
Bacillus cereus MHA +
Listeria monocytogenes MHA -
Clostridium perfringens MHA +
Lactobacillus plantarum MRS -
Enterococcus faecalis (VRE) MRS -
Salmonella Enteritidis MHA +
Escherichia coli MHA +
Campylobacter jejuni Blood agar +
Candida albicans MHA -
5.3. Summary of results and discussion.
The B. amyloliquefaciens #11 was isolated from seaweed Sargasum which was collected in
NhaTrang bay, Vietnam Sea. Genome analysis revealed it possessed gene clusters encoding
synthesis of non-ribosomally synthesized lipopeptides (macrolactin, surfactin, fengycin,
bacillomycin D), two polyketides (bacillibactin), dipeptide bacilysin, and three bacteriocins
(amylocyclicin, LCI, and 1 thiopeptide). Of three bacteriocins, thiopeptide bacteriocin was
identified as novel bacteriocin as its sequence was unmatched to any bacteriocin previously
characterised. The questions arose as to which antimicrobial substances were produced in
vitro, and if the production of the novel thiopeptide occurs. The answer is only clear when all
the antimicrobial compounds produced by this isolate are characterised. Therefore, different
culture fractions and different purification methods were employed to purify all the
antimicrobial substances produced by this bacterium.
First, the growth curve of bacteria in LPMA was observed in association with the ability of
antimicrobial exhibition against two indicators, L. plantarum and B. cereus. From the studies
carried out during the course of this whole research program it was found that L plantarum is
consistently one of the most sensitive indicators for detection of bacteriocin activity, while B.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
127
cereus is more sensitive to NRPs, and also bacteriocins. One of the key factors possibly
contributing to the high sensitivity of L. plantarum to the Bacillus-derived bacteriocins is
probably due to usage of the MRS medium for well-diffusion assay or spot-on-lawn assay.
This media contains the detergent, Tween 80, which is known as an agent that stabilizes highly
modified compounds like bacteriocins, and consequently can enhance bacteriocin activity
(187). This media is known as the best media for production of Lactobacillus-derived
bacteriocins (188, 189). Besides, B. cereus may require higher killing dosages of bacteriocins
(high Minimum inhibitory concentration’ (MIC)) than L. plantarum. Therefore, selection of
appropriate indicators for bacteriocin detection is also important.
Harvesting cultures grown in LMPB, at different points in culture growth, demonstrated that
the antimicrobial compounds produced by B. amyloliquefaciens #11 varied, such that one type
of activity peaked at about 12 hours of growth whereas other activity peaked after 36 hours
of growth, identified by assessment of antimicrobial activity against B. cereus, and L.
plantarum. This indicated the production of multiple antimicrobial substances. Thus, the
culture broth during log-phase growth (< 12 hours of cultivation) was able to kill L.plantarum,
but not B. cereus. While the broths collected during the latter growth phase inhibited the
growth of B. cereus but not L. plantarum. The 12 hour old broth lost antimicrobial activity
when it was incubated with protease enzymes but was resistant to lipase, indicating that the
antimicrobial peptide contains no lipid moiety for its biological activity, therefore, it could be
a bacteriocin. The 36 hours-culture still retained antimicrobial activity when it was incubated
with proteolytic enzymes and also lipase, indicating the unknown nature of the antimicrobial
substances. The growth curve analysis indicated the possible production of different
antimicrobial peptides with different properties at the two time points of 12 hours and 36
hours. Therefore, these broths were collected to purify the antimicrobial peptides.
Purification of the antimicrobial peptides from the 12 hour culture was successful via a three-
step procedure, including precipitating the antimicrobial compounds with ammonium
sulphate, then partial purification via cation exchange chromatography, followed by
purification on an RP-HPLC C18 column. The purity and MW of compound were monitored
by MALDI-TOF MS. Purification resulted in the isolation of an antimicrobial substance
having MW of 6,381 Da; It displayed strong activity against Gram-positive bacteria,
particularly against MSRA, but no activity against Gram-negative bacteria; it was sensitive to
proteinase K and pronase E but resistant to trypsin and lipase, it was very heat-stable, still
retaining activity after being heated to 80°C for 60 min. Based on the properties of this

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
128
compound and properties of the bacteriocins predicted from genome, this antimicrobial
substance is likely to be amylocyclicin. It has the same physiochemical properties as
amylocyclicin, isolated from Bacillus amyloliquefaciens FZB42 (133). In the literature, heat
stable, pH stable bacteriocins are also reported for other Bacillus bacteriocins, like pumilicin
and laterosporulin (190, 191)
The putative novel thiopeptide was not seen in the 12 hour culture, therefore it was searched
in 36 hour cultures. Antimicrobial peptides were recovered from the bacterial cell pellet of 36
hour cultures using a solvent extraction method. A 2-step process was used; i) the bacterial
cells were extracted by stirring in 70% acetonitrile (adjusted to pH 2 by TFA) to release the
antimicrobial peptides into the solvent, and the cell-free solvent was then collected after
concentration of cell-free solvent under reduced pressure (Speed-Vac, or Rotary evaporator);
ii) chromatography on an RP-HPLC C18 column to separate and purify the antimicrobial
peptides from the concentrated solvent extract. After extraction from cell pellets, the cell-free
solvent presented 3 peptide peaks detected using MALDI-TOF under a detective range of
800-2,000 Da. They were (([m/z] = 1,058 Da, (([m/z] = 800), ([m/z] = 1,473) with isoforms.
In the literature, the Bacillus-derived antimicrobial peptides derived from liquid culture-
grown supernatants are reported as mostly falling into three mass ranges of; 850–950 m/z
(kurstakins); 1000–1100 m/z (surfactins, iturins); and 1450–1550 m/z (fengycins,
polymyxins, and bacitracins) (186). Based on this data, the 1058 Da substance could be an
iturin/surfactin. The 800Da-compounds is of similar MW as kurstakins, but this LPs was not
predicted from the genome and therefore, it is an unknown peptide. The 1,473 Da-substance
may be a fengycin or possibly a thiopeptide, as both peptides have similar MWs and
physicochemical properties (high hydrophobicity). To elucidate, this solvent extract was
concentrated in a rotary evaporator for separation using RP-HPLC. During concentration
under low pressure of rotary evaporator, the 1,473 Da compound was precipitated, collected
by centrifugation, and washed several times with milli-Q water, and dissolved in 50%
acetonitrile, as it is unable to dissolve in water or aqueous buffer. The material exhibited
antimicrobial activity against B. cereus. However, when it was stored at chilled temperatures
(5oC or -20oC), the antimicrobial peptide aggregated again and did not redissolve in water or
acetonitrile. Purification of freshly prepared aggregate, using an RP-HPLC C18 column,
resulted in an antimicrobial peptide at a retention time of 44.618 minutes, indicating elution
at approximately 85% acetonitrile. This suggested that this antimicrobial peptide was highly
hydrophobic. However, attempts to elucidate the amino acid sequence of this peptide by N-

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
129
terminal sequencing failed, suggesting its structure may be blocked at N and/or C terminus,
either by some form of chemical modification/protection or cyclization.
Some bacteriocin and NRPs compounds which were predicted from the genome analysis were
not found in any of the culture fraction. These included LCI and difficin. There are some
possible reasons why these compounds were not identified, including i) limitation in the
extraction procedures, such as usage of bacterial indicators that were not sensitive; ii) loss of
peptides during dialysis (using a membrane with cut off 3 kDa), or iii) the media used for
fermentation is inappropriate. This last hypothesis is raised because bacteriocin production
is known to be highly affected by media components, and other tress response factors.
Alternatively, the gene clusters predicted to encode synthesis of these undetected compounds
may be inactive due to repression or mutations. Activation of expression may require stress-
responding factors (nutrients stress, or factors for quorum sensing).
To conclude, the study has identified two antimicrobial compounds that appear to be novel,
one with an MW of 800 Da and the other, which may be a thiopeptide, with an MW of 1,473
Da. The 1,473 Da compound is suspected to be a novel thiopeptide because such a molecule
was predicted as a novel bacteriocin from the genome. However, a full structural analysis of
the peptide is required to finally resolve its identity. Therefore, the therapeutic application of
this bacterium is still unclear, but the B. amyloliquefaciens #11 isolate clearly has extensive
antimicrobial activity and could potentially be used as a biocontrol agent in various
applications, including in agriculture for control of plant and animal pathogens.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
130
Chapter 6 ANALYSIS OF ANTIMICROBIAL PEPTIDES PRODUCED BY
PAENIBACILLUS POLYMYXA #23
6.1. Introduction
Paenibacillus polymyxa is a non-pathogenic, spore-forming, Gram-positive bacterium. It is
ubiquitous in both terrestrial and marine environments (192). Terrestrial P. polymyxa is
known to exhibit various bioactivities like; nitrogen fixation, leading its use as a biofertilizer
in agriculture (193); production of phytohormones (194), and hence used as a biocontrol agent
in agriculture and aquaculture application (193, 195); lipid, cellulose, and starch degradation
which gives it value for the production of important enzymes (protease, lipase, cellulase,
amylases) (196). These enzymes are widely used in food and waste treatment industries.
Isolates of P. polymyxa also, typically produce a range of antimicrobial compounds with
activity against a wide range of fungi and bacteria. Such isolates have the potential to be
developed as probiotics or the purified antimicrobials used as pharmaceuticals. There are still
only a few limited studies conducted on marine-derived isolates of this species, to understand
the antimicrobial potential of this species.
In this study, P. polymyxa #23, isolated from a marine sponge collected in the NhaTrang Sea,
Vietnam, was examined. The bacterium exhibited antimicrobial activity against a wide range
of bacterial indicator strains, including Gram-positive bacteria (S. aureus, C. perfringens, B.
cereus, L. plantarum, L. monocytogenes, MRSA and VRE), Gram-negative bacteria (C.
jejuni, P. aeruginosa, E. coli, S. Enteritidis and even multidrug-resistant K. pneumonia) and
all of the marine antimicrobial producing Bacillus [Figure 6.1, Table 6.1]. It indicates that the
bacterium may produce multiple antimicrobial compounds. Therefore, it was selected for
sequencing and analysis to predict the putative antimicrobial peptides encoded within the
genome. In Chapter 4, in silico prediction of antimicrobial peptides potentially produced by
this isolate uncovered the presence of LPs (1 paenibacterin, 1 tridecaptin, 1 polymyxin, 1
unknown LPs), PKs (1 unknown polyketide), and 4 bacteriocins (paenicilin A, 1 novel
lantibiotic, 1 novel sactipeptide, 1 uncharacterised lassopetide). The work presented in this
chapter aimed to purify some of these novel bacteriocins for in vitro characterisation. The
bacterium was fermented and the antimicrobial peptides present in the culture were purified,
employing various separation techniques and in aid of mass spectrometry for identification of
peptides. The procedures included; i) selection of media for enhancement of antimicrobial
production, and identification of the time-point during bacterial growth for best yield of

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
131
antimicrobial products; ii) extraction of antimicrobial peptides from the cell surface using
acidic polar solvent; iii) RP-HPLC. The result in this chapter describes the antimicrobial
potential to underpin applications in pharmaceuticals.
Figure 6.1. Antimicrobial spectrum exhibited by marine P. polymyxa #23. (A) Several
representative Gram-positive bacteria inhibited. (1) S. aureus, (2) MRSA, (3) S.
faecalis, (4) B. cereus. (B) Several representative Gram-negative bacteria and yeast
inhibited such as P. aeruginosa (Ps), S. Enteriditis (Sal), E. coli (E), C. albicans (Ca).
The test was performed by cross streak assay with a duplicate of indictors.
6.2. Result
6.2.1. Time course analysis of antimicrobial production during bacterial growth
revealed production of multiple antimicrobial substances.
To compare the effect of media composition on bacteriocin production, six different media
formulations were tested, including production media (PB) (51); marine broth (Difco 2216)
(MB); lab-prepared marine broth (LPMB); nutrient broth (NB) (Oxoid); tryptone soy broth
(TSB) (Oxoid), and TSB supplemented with 0.6% yeast extract (TSBY). See the appendix for
the recipes of all defined media. An overnight starter bacterial culture, grown in LB broth was
added (1 mL) to 100 mL of each of the above media. All inoculated media were incubated at
30oC under a shaking speed of 150 rpm. Five mL aliquots of each culture were collected after
12, 18, 24, 36, 42, 48, 60, and 66 hours of incubation, filtered through a 0.45µm PES
membrane and the antimicrobial activity in the cell-free supernatant was evaluated against S.
Enteritidis and B. cereus as indicators.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
132
The results showed that growth in PB medium yielded the highest levels of antimicrobial
production, compared with other media [Figure 6.2 A]. In the PB grown culture, the
antimicrobial activity against S. Enteritidis was first observed after 6 hours of fermentation,
and the strongest inhibitory effect was seen from the 24 hour culture, and activity remained
at later times. The killing zones (from the well-diffusion assay) against Salmonella were
characterised by sharp boundaries at the edge of the zone in the majority of time-points, but
only 48 hours of culture, resulted in a more diffuse, blurred edge of the killing zone. The
activity against B. cereus was first observed after 18 hours of cultivation, reached peak
activity after 24 hours of cultivation, and started reducing after 36 hours [Figure 6.2 B]. Taken
together, it was clear that the antimicrobial activity against both indicator strains peaked in 24
hour culture; therefore, the 24 hour culture was collected for and used for antimicrobial
purification.
Figure 6.2. Antimicrobial activity exhibited by P. polymyxa #23 in culture media. (A)
Media selection for antimicrobial production. Six media were selected for antimicrobial
production including PB (1), LPMB (2), MB (3), NB (4), TSB (5). The cultures were
collected after 12 hours (a), 24 hours (b) and 36 hours (c) for testing against Salmonella
(left) and B. cereus (right). (B) The antimicrobial activity exhibited across the growth
phases in PB medium. Culture aliquots were collected after 6 hours to 66 hours and

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
133
tested against two indicators. Resulted showed that the activity against Salmonella was
started being observed from 6 hours culture. Activity against B. cereus was started being
observed from 18 hours old culture. The blurred edge of killing zone from 48 hours old
culture indicated the production of additional antimicrobial substances at this time
point.
6.2.2. Purification of antimicrobial compounds from 24 hour old PB culture revealed
the presence of three antimicrobial compound/s
6.2.2.1. Recovery of antimicrobial compounds from cell-free culture and the cell pellet
In the 24 hour old culture, the antimicrobial activity was observed from both cell-free culture
supernatant and cell pellet extracts. Therefore, two separate procedures were designed to
recover the antimicrobial compounds from these two sources. The antimicrobial presented in
the cell-free culture supernatant was absorbed onto Diaion HP-20, and the antimicrobial
substance was eluted with 70% acetonitrile [Figure 6.3]
Figure 6.3. Recovery of the antimicrobial compounds from the culture supernatant of
P. polymyxa #23 by absorption antimicrobial substances onto Diaion HP-20. Elutions
were carried out with; 30%, 50%,70%, and 100% ACN. The antimicrobial compound
could be eluted at least 70% acetonitrile.
The elute was then centrifuged at 5,000 g to collect the supernatant, filtered through a syringe
membrane (0.45µm PES) to obtain the cell-free solvent containing the antimicrobial
compound. The cell-free solvent was concentrated by a rotary evaporator under reduced
pressure. During concentration, a precipitate appeared which was separated from clear
supernatant (fraction CFS) by centrifugation. The precipitate was washed (twice) with milli-
Q water and dissolved in 40% acetonitrile (named SFP). Both SFP and CFS exhibited
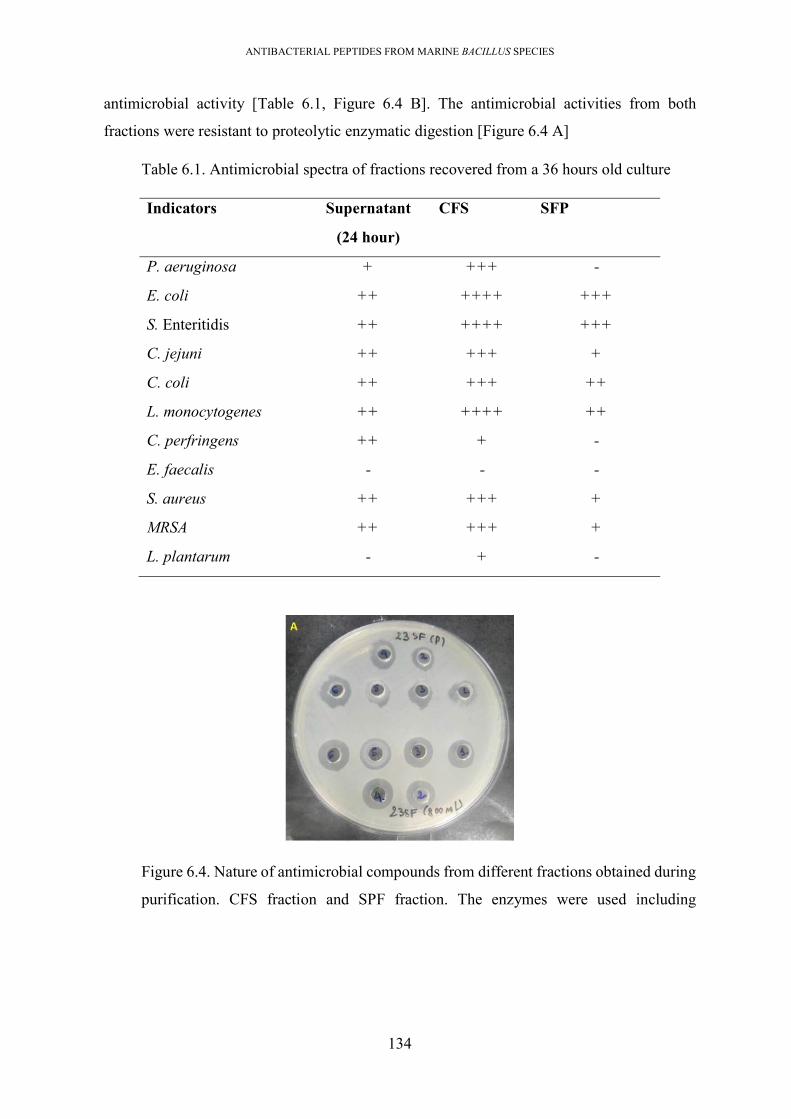
ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
134
antimicrobial activity [Table 6.1, Figure 6.4 B]. The antimicrobial activities from both
fractions were resistant to proteolytic enzymatic digestion [Figure 6.4 A]
Table 6.1. Antimicrobial spectra of fractions recovered from a 36 hours old culture
Indicators Supernatant
(24 hour)
CFS SFP
P. aeruginosa + +++ -
E. coli ++ ++++ +++
S. Enteritidis ++ ++++ +++
C. jejuni ++ +++ +
C. coli ++ +++ ++
L. monocytogenes ++ ++++ ++
C. perfringens ++ + -
E. faecalis - - -
S. aureus ++ +++ +
MRSA ++ +++ +
L. plantarum - + -
Figure 6.4. Nature of antimicrobial compounds from different fractions obtained during
purification. CFS fraction and SPF fraction. The enzymes were used including

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
135
proteinase K (1), pronase E (2), trypsin (3), catalase (4), lipase (5), and fraction without
treatment as a positive control (6).
6.2.2.2. Purification of the antimicrobial peptides in CFS fraction with hydrophobic
exchange chromatography elucidated the presence of two antimicrobial
compounds.
The CFS sample was loaded into a C18 cartridge (Waters), and the column was then eluted
with a linear concentration gradient, ranging from 30% to 100% ACN (+0.01%TFA) to
recover the antimicrobial compounds. All elutes were collected and evaluated for
antimicrobial activity against both Bacillus and Salmonella. Antimicrobial activity was
observed in two fractions; at 30% acetonitrile (named as 30ACN elute) and at 40% acetonitrile
(named as 40ACN elute). The 30ACN elute displayed activity against S. Enteritidis while the
40ACN exhibited activity against both S. Enteritidis and B. cereus [Figure 6.5, Table 6.2].
Therefore, these two elutes were lyophilized, redissolved in milli-Q water to inject separately
into an RP-HPLC using C18 column.
Figure 6.5. Hydrophobic interaction chromatography (using Sep-Pak C18 cartridge) to
purify the antimicrobial “cell surface extract” (CFS).Two elutes with antimicrobial
activity were detected with elution at 30% acetonitrile (30ACN) and 40% acetonitrile
(40%ACN).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
136
Table 6.2. The antimicrobial spectra of 30ACN fraction, and 40ACN fraction.
Indicators 30%ACN 40%ACN
B. cereus - +
L. monocytogenes - +
S. aureus - +
MRSA - +
L. plantarum - +
E. faecalis ++ +
E. coli ++ +
S. Enteritidis - +
C. jejuni ++ -
K. pneumonia ++ -
6.2.2.3. Purification of 30ACN fraction revealed the presence of polymyxin
Separating this 30ACN elute using RP-HPLC (C18 column) resulted in an antimicrobial
fraction at 25.6 minutes. MALDI-TOF MS showed that this fraction had a single peak at [m/z]
=1,168.772. It was thermally stable, as its activity was unaffected after being heated at 100oC
for 1 hour; and it was stable under various enzymatic digestions (proteinase K, protein E,
trypsin, lipase, catalase and lysozyme). It exhibited antimicrobial activity against most Gram-
negative bacteria (but not against C. jejuni) and displayed a circle of inhibition on a zymogram
gel at a range of 2-10 kDa against S. Enteritidis [Figure 6.6].

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
137
Figure 6.6. Zymogram of polymyxin against S. Enteritidis
The determination of the amino acid sequence of this compound failed with N-terminal
sequencing (Edman degradation technique). This indicated that the peptide’s N-terminal was
blocked. As an alternatively, tandem MS was employed. During the cycle, the compound
presented in three charged states. The single charged version also demonstrated some gas-
phase dimerization, most likely due to the high intensity [Figure 6.7]. Although strong tandem
MS spectra were acquired for all three charge states, no primary sequence was forthcoming
from automated de novo sequencing from the PEAKS software. Additionally, wildcard
searches in Byonic, of Bacillus proteins (197), where unknown masses may be added to
peptides, did not yield reasonable sequences. Therefore, manual de novo sequencing was
performed. Numerous water losses were observed, and most mass differences did not
correspond to proteinogenic amino acid residues, though there was potentially some evidence
for threonine and isoleucine/leucine. Several instances of mass differences that could
correspond to non-proteinogenic amino acids occurred in the spectra, including;
diaminopropanoic acid, diaminobutanoic acid (like shorter chain lysines) and
hydroxynorvaline. Using the most intense ions and water losses as a reference, a potential
sequence based on the doubly charged fragment spectrum was vLLvTvvTvu, where v is 2,4-
diaminobutanoic acid (or equivalent, C4H10N2O2) and u is something akin to Amino(1H-
pyrrol-2-YL) acetic acid (C6H8N2O2).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
138
Figure 6.7 Sequencing result of the purified polymyxin. (A) Peptide sequencing of the
purified polymyxin by modified tandem mass spectrometry. (B) Structure of known
polymyxin. The image was obtained from Poirel et al, 2017 (198).

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
139
Table 6.3 The antimicrobial spectrum of polymyxin
Indicators organisms Activity
B. cereus -
L. monocytogenes -
S. aureus -
MRSA -
L. plantarum -
E. faecalis ++
E. coli ++
S. Enteritidis +
C. jejuni -
K. pneumonia ++
+ and ++ indicate the relative strength of antimicrobial activity.
6.2.2.4. Purification of 40ACN fraction revealed the presence of paenicidin A,
tridecaptin
Purification of antimicrobial peptides from the 40ACN elute was performed using an RP-
HPLC using a C18 column, yielding two fractions exhibiting the antimicrobial activity at
retention times of 23.443 minutes ([m/z] = 1616) and 23.797 minutes ([m/z] = 3370) [Figure
6.9]. The 3,370 Da antimicrobial peptide displayed antagonistic activity against only B.
cereus, while the 1,616 Da compound was able to kill both Salmonella and B. cereus.
Figure 6.8. The first round of RP-HPLC to separate the 40% ACN fraction.Two
fractions exhibited antimicrobial activity, with peaks at retention times of 23.443
minutes ([m/z] = 1616) and 23.797 minutes ([m/z] = 3370)

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
140
Figure 6.9. MALDI-TOF MS spectra of 40% ACN fraction under detection range of
800 Da – 2000 Da. The high mass intensity in the m/z range of 1,400–1,700 indicated
the presence of tridecaptin.
6.2.4.1. The paenicidin A, a lantibiotic, displayed antimicrobial activity against Gram-
positive bacteria
The 3,370 Da antimicrobial compound was sensitive to proteolytic treatment (pronase E and
proteinase K) and demonstrated antimicrobial activity against only Gram-positive bacteria
(but not against L. plantarum and E. faecalis) [Table 6.8]. Based on the antimicrobial
spectrum, MW, and list of antimicrobial peptides predicted by in silico analysis of the
genomic sequence, this antimicrobial peptide is likely to be paenicidin A [Figure 6.10; Table
6.4]
Figure 6.10. MW of purified paenicidin A by MALDI-TOF MS with the mass intensity
of [m/z] = 3370.
6.2.4.2. The tridecaptin, a lipopeptide with anti-Campylobacter activity
The 1,616Da-compound was resistant to proteolytic treatment and exhibited antimicrobial
activity against only Gram-negative bacteria (C. jejuni, S. Enteritidis and E. coli but not P.
0940.383 1614.894
977.096
983.091
1660.637
1668.588
1570.349
1683.232
1690.817
1004.225
1150.014
1706.565
1483.229 1789.3721848.7771038.312
1010.264
1729.125
1544.438
1529.0891114.570 1251.774
Native23_60%eluteACN_22min 0:D5 MS Raw
0.5
1.0
1.5
5x10In
ten
s. [a
.u.]
900 1000 1100 1200 1300 1400 1500 1600 1700 1800 1900m/z

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
141
aeruginosa and K. pneumonia) [Table 6.4]. Based on the antimicrobial spectrum, MW, and
list of antimicrobial peptides predicted from the genome, this compound was identified as
tridecaptin.
Table 6.4. Antimicrobial activity of compounds present in the 40ACN fraction
Indicators PaenicidinA Tridecaptin
B. cereus ++ -
L. monocytogenes ++ -
S. aureus + -
MRSA + -
L. plantarum + -
E. faecalis + -
E. coli - ++
S. Enteritidis - ++
C. jejuni - +
K. pneumonia - -
P. aeruginosa - -
6.3. Summary of results and discussion
Attempt to extract the predicted novel thiopeptide bacteriocin from B. amyloliquefaciens #11
culture failed. Therefore, as an alternative approach, P. polymyxa #23 was used to attempt
isolation of novel bacteriocins. The P. polymyxa #23 genome had 3 novel bacteriocins gene
clusters (1 lassopeptide, 1 lantibiotic, 1 sactippetide) along with paenicidin A and various
NRPs substances (tridecaptin A, polymyxin, and fusaricidin, paenibacterin). The
antimicrobial potential of #23 is similar to that of P. polymyxa NRRL B-30509, which is
known as a producer of polymyxin, tridecaptin, and paenicidin A (146). To determine if the
predicted novel bacteriocins were produced in vitro, #23 was grown in liquid culture, and
various purification methods were applied to obtain these novel bacteriocins for
characterisation.
First, six media types (PB, MB, LMPB, NA, TSB, and TSBY) were selected to culture the
bacterium, to evaluate the yield of antimicrobial compounds. These media were selected
because they were used in earlier studies to ferment for bacteriocin production (51, 84, 199,

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
142
200). The selection of culture conditions for enhancement of bacteriocin production is a
complex process, which medium formulations and growth conditions are two factors highly
influencing bacterial growth and bacteriocin production. The result showed that cultivation of
bacterium in PB media resulted in the strongest antimicrobial activity against two indicators
(S. Enteritidis, and B. cereus) across growth-phases. The PB is a completely defined media
which is formulated with rich nutrients (carbon and nitrogens source) and minerals and can
produce high cell densities and in consequence high bacteriocin production. This observation
is in agreement with earlier studies which reported that completely defined medium frequently
increased the yield of bacteriocin production (201).
The bacterium was cultured in PB medium, and samples were collected every 6 hours and
antimicrobial activities were evaluated against two indicators; S. Enteritidis and B. cereus.
The two indicators were killed by cultures collected at different time-points, indicating the
production of multiple antimicrobial substances. Thus, activity against Salmonella was
observed starting at 6 hours of cultivation, reached a maximum at 24 hours, and was
maintained throughout the rest of the culture monitoring period (66 hours). Interestingly, the
culture collected at 60 hours exhibited a strange diffuse edge to the killing zone, suggesting
the production of additional antimicrobial substances. The activity against B. cereus was first
observed from the 12 hour sample and steadily reduced from 36 hours. Because the 24 hour
broth displayed strong activity against both Salmonella and Bacillus, the culture at this time-
point was collected for antimicrobial purification, to characterise all the putative antimicrobial
peptides produced
The purification of antimicrobial compounds from 24hour old PB culture resulted in the
presence of 3 antimicrobial peptides, which were; polymyxin ([m/z]= 1168.7), paenicidin A
([m/z]= 3290.4), and tridecaptin ([m/z]= = 1550.8). It is assumed that the activities against
Gram-negative bacteria are due to polymyxin and tridecaptin. Of them, the activity against C.
jejuni is mainly by tridecaptin, and ability to inhibit P. aeruginosa and K. pneumonia is a
result of polymyxin activity. The activity against Gram-positive bacteria is due to paenicidin
A, which is commonly found in Paenibacillus species (43, 146). In the literature, polymyxin
is found and extracted from various terrestrial Paenibacillus species (202, 203). Eight variants
of polymyxin (A to E, M, S, and T) are described in works of literature, but only polymyxin
B and E are widely used in the treatment of serious Gram-negative bacterial infections with
another name as colistin (colistin E, colistin B) (204, 205). The tridecaptin is a lipopeptide
previously extracted from P. jamilae M-152, P. terrae NRRL B-30644 and P. polymyxa

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
143
NRRL B-30509, with the presence of some variants (tridecaptin A1, newly tridecaptin M)
(127, 146). Polymyxin, and tridecaptin display different killing modes against the targeted
bacteria, therefore they can be used in combination to cure various infectious diseases caused
by Gram-negative bacteria, even carbapenem-resistant Enterobacteriaceae (8, 206). Due to
the difference in the mode of action, tridecaptin may be a good compound for further
development in treatment against colistin-resistant pathogens. The identification of
tridecaptin and polymyxin as antimicrobials produced from isolate #23 emphasizes the
potential therapeutical value of this bacterium.
Some other NRPs compounds and novel bacteriocins were not observed during purification.
There are several possible explanations for the inability to purify these antimicrobials; i) the
novel bacteriocins were not produced under the culture conditions used, or ii) loss of the novel
bacteriocin during purification.
Production of bacteriocin by bacteria is known to vary greatly, depending on types of
bacteriocin, type of host, and types of media used for fermentation, and the growth phase
(124, 207). Refining media for production is a complicated process, but it is necessary for the
enhancement of compounds of interest. Thus, some bacteriocin production was enhanced
when media was supplemented with surfactant (like Tween), NaCl, or ethanol (187, 207-210).
Of them, tween may stabilize the bacteriocin structure by preventing the aggregation of
bacteriocin molecules. Tween 80 is a common supplement in MRS medium, which is reported
as the best medium for high yield of bacteriocins deriving from Lactobacillus (211).
Supplementation of NaCl and ethanol can also increase bacteriocin yield, but in some cases,
high concentrations of NaCl and ethanol can also result in inhibition of bacteriocin
production, such as curvacin A (210). Besides, media with fewer nutrients is reported to
enhance bacteriocin yield; such conditions may activate bacteriocin transcription in response
to nutrient stress (212). However, some bacteriocin productions are only maximized in media
with which supply appropriate levels of sugars, vitamins, and nitrogen (207). High nutrient
media can yield a high density of cells, therefore improving bacteriocin yield; however, an
oversupply of these nutrient sources can lead to inhibition of both bacterial growth and
bacteriocin production.
While, loss of novel bacteriocins during the purification process is also possible, probably due
to limitation of this purification procedure or inactivation during the processing steps used.
Thus, the bacteriocin may be produced at low concentration in an unsuitable media, which

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
144
may lower the ability to detect the antimicrobial activity against indicator strains. This may
result in an inability to detect fractions containing these bacteriocins, during purification.
Solutions that could be considered to overcome this failure include heterologous cloning and
expression of the bacteriocin gene cluster to produce the antimicrobial in an engineered
system, or inactivation the genes predicted to be required for NRPs production (sfp genes) to
stop NRPs production and identify the change in the antimicrobial activity of the mutated
strain (27).
Three solutions can be suggested to overcome purification failure based on the above reasons.
Firstly, selecting an appropriate bacterial indicator may be a critical factor in determining
success in the detection of novel bacteriocins production. For instance, use of mutant strain
HB0042 of B. subtilis as an indicator. This bacterium was used as an indicator to detect the
presence of novel plantazolicin and amylocyclicin produced by Bacillus amyloliquefaciens
FZB42 (51, 133). This strain contains the gene encoding an extracytoplasmic sigma factor
(SigW) that provides the resistance against the NRPs produced by bacilli (213). Alternatively,
the use of a marine Bacillus species among the shortlisting of 23 marine isolates in this study
as an indicator. The marine strain selected has a known genome sequence which contains the
genes responsible for the biosynthesis of NRPs compounds, but deficient genes of novel
bacteriocins. For some novel bacteriocins that their productions appear to be inactive with
unknown reasons, dead indicator cells can be added to the fermentation culture. The cells can
serve as inducing molecules to activate the “silent” bacteriocin production via a quorum
sensing mechanism. Secondly, deficient in the genes relating to NRPs of wide-type bacteria
by genetic modifications, then fermentation of this mutant for production of novel bacteriocin.
Thirdly, heterologous cloning and expression of the gene clusters of novel peptides
Taken together, the main aim of this chapter, to purify the putative novel antimicrobials
predicted to be produced by P. polymyxin, was not achieved. However, the study has provided
information to understand the isolate, particularly the antimicrobial activities that could be
developed to address issues in therapeutics and agriculture.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
145
Chapter 7 CLONING AND HETEROLOGOUS EXPRESSION OF SACTIPEPTIDE,
A NOVEL BACTERIOCIN FROM MARINE P. POLYMYXA #23, IN BACILLUS
SUBTILIS
7.1. Introduction.
Genomic analysis of P polymyxa #23 identified 13 biosynthetic gene clusters that were
predicted to direct the synthesis of antimicrobial compounds. Three were predicted to encode
uncharacterised bacteriocins; 1sactipeptide, 1 lantibiotic, 1 lassopeptide. However, these three
novel bacteriocins were not observed during the purification procedures outlined in Chapter
6, even though the gene clusters appear to have full complements of all the accessory genes
required for biosynthesis. Therefore, heterologous cloning and expression of sactipeptide
were attempted, to provide an alternative source from which to purify the bacteriocin for in
vitro characterisation. This technique was expected to enable sactipeptide production by using
strong promoters that are unlikely to be responsive to suppression by inhibitors which may
have interfered with biosynthesis from the native gene cluster.
Cloning and expression techniques employ a variety of host-vector systems which are selected
depending on the type of gene(s) to be expressed and the size of gene clusters used. For
bacteriocin expression, it is often necessary to clone the whole gene cluster carrying the
structural gene for bacteriocin and all the accessory genes. Therefore, the bacterial host for
expression must be able to receive large vectors, be deficient in other bacteriocin genes, and
contains accessories genes supporting expression. Examples of early studies to heterologous
express bacteriocins include the production of mersacidin (214), pediocin PA-1 (215),
plantaricin ZJ5 (216), and nisin (217). In the literature, the majority of reported studies used
E. coli as the host for expression and variable vector types.
In this study, the cloning and expression of sactipeptide were processed using plasmid
pDLL202 (kindly provided by Monash University) and B. subtilis cells for expression. The
plasmid pDLL202 contains two compatible origins of replication (colE1 rep and repA),
enabling replication in both E. coli and B. subtilis hosts [Figure 7.1 A]. The plasmid contains
four multi-cloning-sites (MCS) which include recognition sites of various restriction
enzymes, for genetically engineering, and 4 antibiotic resistance genes; kanR, ermB, catP,
and ampR, which confer on the host resistance to kanamycin, erythromycin, chloramphenicol,
and ampicillin. The sactipeptide biosynthetic gene cluster that was cloned is 8,556 bp in length
and consisted of a precursor bacteriocin, 3 modification enzymes, a transporter for bacteriocin

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
146
secretion, and a putative immunity protein [Figure 7.1B]. The precursor bacteriocin is 61 aa,
comprising of a 19 aa leader sequence and a 42 aa core sequence. The core sequence was
cysteine-rich, and was predicted with a theoretical MW of 4,455.81 Da, and a pI of 7.73. The
MW of sactipeptide was estimated from core sequence, which was predicted by SignalP
software.
Figure 7.1. (A) Gene organisation on plasmid pDLL202; (B) The plasmid contains 4
antibiotic resistance genes and 4 multi cloning sites (MCS), and (C) the gene cluster of
sactipeptide;. The figure of plasmid was provided from Monash University. The gene
organisation of the sactipeptide gene cluster is derived from the annotation of the P.
polymyxa #23 genome sequence.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
147
Transformation into B. subtilis cells is more successful with plasmids in multimeric form
rather than ones in monomeric form (218, 219); therefore, the procedure included an
intermediate step of transformation into E. coli JIR702 cells that produce fusion plasmids to
generate the multimeric plasmid. In practice, the whole procedure consisted of the following
steps; (i) amplification of the sactipeptide gene cluster from the genome of P. polymyxa #23
using designed primers and high fidelity polymerase; ii) preparation of the plasmid backbone
by double restriction enzyme digestion of plasmid pDLL202, to accept the amplified gene
cluster; (iii) construction to generate the fusion plasmid, pDLL202/G23sac, by Gibson
assembly (220) ; (iv) transformation of the fusion plasmid into E. coli Top 10 cell to recover
the constructed clone; (v) extraction of plasmid DNA from E. coli Top 10 cells and
transformed into the E. coli JIR702 to generate the multimeric form of the plasmid; (vi)
extraction and transformation of this multimeric fusion plasmid into B. subtilis BS34A for
expression of sactipeptide; (vii) selection of culture media for bacteriocin expression by B.
subtilis transformants; and (vii) purification of the sactipeptide for characterisation.
7.2. Methods and results
7.2.1. Successful isolation of intact sactipeptide whole gene cluster, and generation of
the expression plasmid (pdLL202/G23sac)
The whole sactipeptide gene cluster was amplified from P. polymyxa #23’s genome using the
designed primers [Table 7.1]. The P. polymyxa #23 genomic DNA was extracted by the GES
method and used as a template in PCR. The 50 μL PCR reaction included 2.5 μL of each 10
mM forward and reverse primer (G23sac_F/ G23sac_R), 1.5 μL of DMSO, 25 μL of
Phusion® High-Fidelity PCR Master Mix (Thermo Fisher Scientific), 1 μL of HF buffer, 2
μL of intact gDNA (100 ng) and 15.5 μL of water. The PCR conditions were; 98oC for 30
seconds, 35 cycles of; 98oC for 10 seconds, 56oC for 30 seconds, 72oC for 4.5 minutes, and a
final extension at 72oC for 10 minutes. The PCR product was purified using the Monarch PCR
clean kit (NEB). The primers (G23sac_F/ G23sac_R) were designed to generate a PCR
product containing the sactipeptide gene cluster flanked by ends regions of 20 nucleotides,
which overlapped with the terminal regions on the plasmid vector backbone.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
148
Table 7.1. List of primers used for cloning and confirmation
Primers 5’-3’ sequence Nucleotide Tm
(to)
Purpose Source
G23sac_F CAGCGAGATCTGATCACGCGCAGTTTACCAGTAGCTGCTC
40 58 amplification of 8,556 bp- sactipeptide gene cluster from wide-type P. polymyxa #23
This study
G23sac_R GCTAGCGACGTCTAGACTAGCGGGAATTAAGTAGGGGTATAGG
43 59
23Sac-test_F
ATAATTACCAATTCCCGTTGC
21 51 amplification of 249 bp- DNA region inbetween sactipeptide gene cluster for confirmation
This study
23Sac-test_R
GTAAGGAATTAGTATGGGTGAAC
23 51
Both the purified plasmid vector and the purified whole sactipeptide gene cluster were
checked to be of the expected approximate size by electrophoresis on a 0.8% agarose gel. The
1 kbp DNA ladder (Thermo Scientific) was used as a standard to estimate size [Figure 7.2].
The plasmid and amplicon bands were of the expected size. The PCR product of sactipeptide
gene cluster was further confirmed by PCR amplification using a second pair of primers
(23Sac-test_F/ 23sac_test_R). The 20 μL PCR reaction included; 0.8 µL of each 10 mM
primers (23sac-test_F and 23sac_test_R), 10 µL of MyTaq Red HS master mix (Bioline), 1
µL of purified sactipeptide gene cluster, and 8.4 µL of molecular water. The PCR conditions
were 95oC for 1 minute, 30 cycles of; 95oC for 15 seconds; 49oC for 20 seconds; 72oC for 10
seconds, and a final extension at 72oC for 10 minutes, and then held at 5oC. This primer pair
was designed to amplify a 249 bp- DNA region locating within the sactipeptide gene cluster
but did not amplify any DNA regions within the plasmid pDLL202 or genome of the B.
subtilis BS34A host.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
149
Figure 7.2. Construction of fusion plasmid. Band of 8,556 bp gene cluster of
sactipeptide amplified from #23 genome (A); the band of 6,938 bp of the plasmid vector,
from restriction enzyme digestion of plasmid pdLL202 (B); the band of 15,497 bp
fusion plasmid generated after Gibson assembly (C); 1 kbp DNA ladder (M)
The fusion plasmid was generated from the purified plasmid, pDLL202, and the purified PCR
product of the sactipeptide gene cluster using the Gibson assembly method. Briefly, a 20 µL
reaction was prepared including; 10 µL of NEBuilder HiFi DNA Assembly Master Mix
(NEB); ~0.15 pmol of insert and plasmid backbone (ratio of 1: 3); and water. The Gibson
reaction was incubated at 50oC for 60 minutes and immediately used for transformation or
stored at -20oC until usage. The gene content of the fusion plasmid shown in Figure 7.3.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
150
Figure 7.3. Gene organisation of fusion plasmid pDLL202/G23sac generated after
Gibson assembly. The figure was prepared in Snap-gene software.
7.2.2. Successful transformation of fusion plasmid into E. coli Top10
After assembling the mixture was transformed into E. coli Top10 by electroporation. The
transformation mix was plated on selective LB (chloramphenicol) and all colonies forming
on the plate were picked and purified by streaking onto another fresh selective plate.
Successful cloning was confirmed by both PCR amplification and restriction enzyme
digestion of the plasmid. The colony PCR was carried out on all the transformant colonies
using the primer pair (sacti-test_F/ sacti-test_R). Results showed the PCR product presented
from some colonies when compared with positive control [Figure 7.4]. The plasmid DNA
from the PCR-positive transformants was isolated using a Monarch® DNA Gel Extraction
Kit (NEB) and confirmed again by restriction enzyme digestion, analysed on an agarose gel.
7.2.3. Successful transformation of the fusion-plasmid into E. coli JIR702 for
production of multimeric plasmid.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
151
The fusion plasmid DNA was extracted from an overnight culture of E. coli Top10
transformants using the Monarch® DNA Gel Extraction Kit (NEB). It was transformed into
E. coli JIR702 by electroporation to induce the multimeric form of this plasmid. The
multimeric plasmid was required for transformation into the B. subtilis BS34A host. The
transformation procedure was similar to that performed into the E. coli Top10. The positive
transformants were also confirmed by PCR primer pair G23sac_test_F/ G23sac_test_R via
size confirmation and enzymatic digestion. The multimeric form of the pDLL202/G23sac
plasmid was extracted by the Monarch® DNA Gel Extraction Kit (NEB) and subsequently
transformed into B. subtilis BS34A cells under the natural transformation method. The outline
of the method was mentioned in the methodology of chapter 2.
Figure7.4. Confirmation by PCR for positive transformants after transformation.(A) On
E. coli JIR702 transformants. #01 to #14: used transformants’ DNA as templates, well
#15: positive control using genomic DNA of P. polymyxa #23 as a template, #16:
negative control (water as a template). (B) For B. subtilis BS34A transformants; using
transformants’ DNA as templates, #14: positive control using genomic DNA of P.
polymyxa #23 as a template.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
152
7.2.4. Successful expression of sactipeptide gene cluster in B. subtilis and optimization
of sactipeptide production
The Bacillus transformants which were confirmed for the presence of fusion plasmid were
fermented in two types of culture media including Production broth (PB); and LB broth, to
observe the antimicrobial production. The production of sactipeptide was only seen in LB
broth [Figure 7.5]. During purification of the sactipeptide, all cultures and solid media were
supplemented with 5 µg/mL chloramphenicol to maintain the fusion plasmid. The methicillin-
resistant S. aureus (MRSA), which was resistant to chloramphenicol, was used as an indicator
in antimicrobial detection assays (the strain was provided by Microlab-RMIT University)
Analysis on bacterial growth in LB medium in association with a strength of antimicrobial
exhibition showed that the sactipeptide was first produced after 6 hours of fermentation, and
exhibited the strongest activity against MRSA at 24 hours of incubation (corresponding to an
of OD600 of ~1.4) and reduced and lost activity after 32-hours of incubation [Figure 7.5 B,
Table 7.2]. Therefore, the 24hour-old culture was collected for purification.
Figure 7.5. Antimicrobial activity exhibited by some B. subtilis transformants against
MRSA indicator.(A) Cultivation of 3 B. subtilis transformants in Production media (PB)
and Lueria media (LB). The antimicrobial activity was only seen in LB media. Culture
of the B. subtilis harbouring native pDLL202 was used as a negative control. (B)

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
153
Antimicrobial activity exhibited by LB cultures collected at different growth phases.
The production of sactipeptide was initially observed after 6 hours of cultivation.
Table 7.2. The growth phase of B. subtilis transformant in LB medium
Time-points OD600 Killing diameter
against MRSA(cm)
2 hours 0.020 0
4 hours 0.056 0
6 hours 0.134 11
8 hours 0.306 13
10 hours 0.802 12
12 hours 0.872 12
24 hours 1.405 16
26 hours 1.306 14
28 hours 1.738 12
30 hours 1.955 13
32 hours 2.01 10
7.2.5. Two-steps procedure to purify the sactipeptide from 24 hour LB culture
The purification procedure included two steps; precipitation of sactipeptide in the culture at
70% saturated concentration of ammonium sulphate; and separation under two cycles of
HPLC using a reverse-phase column (C18). In practice, the ammonium sulphate was added
to the cell-free LB supernatant to final 70% saturation concentration. The mixture was
centrifuged at 17,500 g for 90 minutes, and the protein pellet was dissolved in a minimum
volume of milli-Q water. Next, the material was purified by RP-HPLC (C18 column) with
two cycles, until the pure compound could be collected. During purification, MRSA was used
as an indicator to detect the fractions exhibiting the antimicrobial activity using the spot-on-
lawn method. In the first round of RP-HPLC, the chromatography was operated with a
solution A (milli-Q water + 0.1% (TFA)) and solution B (100% HPLC-grade acetonitrile +
0.1% TFA). The cycle was operated at a flow rate of 1 mL per minute under 55 minutes cycles
of; 2% B in 5 minutes, a gradient of 2%- 95% B in minute #6 to #45, and isocratic elution of
95% eluent B for 10 min. The elution was visualized under UV wavelengths of 214 nm, 230

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
154
nm, and 280 nm. Elutes were collected every 2 minutes and used in the antimicrobial testing
assay. The first RP-HPLC cycle resulted in an antimicrobial fraction at a retention time of 30–
32 minutes [Figure7.6 A]. This fraction was concentrated by SpeedVac (at 37oC) and loaded
into the RP-HPLC as input for the 2nd operation. The second round of RP-HPLC resulted in
the elution of an antimicrobial compound at a retention time of 32.7 minutes [Figure7.6 B].
The MALDI-TOF-MS revealed a major peak with m/z 3404 Da [Figure 7.7].
Figure7.6. Reverse-phase (RP) high-performance liquid chromatography (HPLC)
chromatograms to separate the AMS fraction.(A) The first round of RP-HPLC resulted
in antimicrobial elution at tR of 30–32 minutes. (B) The second round of RP-HPLC
resulted in antimicrobial elution at tR of 32.77 minutes.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
155
Figure 7.7. MALDI-TOF MS of purified sactipeptide.The measurement showed a major
peak of the sactipeptide at m/z 3404.
7.2.6. Characterisation of sactipeptide revealed the proteinaceous nature and thermal
stability of the sactipeptide
The thermal and enzymatic stability of the antimicrobial substance was identified.
Consequently, it displayed thermal stability, because the antimicrobial activity remained after
heat-treatment of 100 °C for 1 hour, although the activity was observed to be reduced. It was
sensitive to pronase E, proteinase K, and trypsin but was resistant to lipase and catalase. The
antimicrobial spectrum of sactipeptide was assayed using the HPLC-based, purified material,
resulting in activity against only Gram-positive bacteria. It strongly depressed the growth of
MRSA [Figure 7.8, Table 7.3].
Figure 7.8. Properties of sactipeptide. Enzymatic degradability: catalase (C), lipase (L),
pronase E (E), proteinase K (K) and trypsin (T). The result showed that the activity was
reduced after digestion with trypsin, pronase E and proteinase K. The thermal stability

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
156
determined by heating peptide at 40oC, 50oC, 60oC, 70oC, 80oC, 90oC; 100oC for 1 hour.
The result showed that activity remained after thermal treatment at 100oC
Table 7.3. Antimicrobial spectrum of sactipeptide
Indicators Media Antagonistic activity
S. aureus MHA +
MRSA MHA ++
B. subtilis MHA +
B. cereus MHA +
L. monocytogenes MHA -
C. perfringens MHA -
L. plantarum MRS -
E. faecalis (VRE) MRS +
S. Enteritidis MHA -
E. coli MHA -
C. jejuni MHA+ 5% blood -
C. albicans MHA -
7.2.7. N-terminal sequencing of the sactipeptide
The bacteriocin sequence was unable to be determined by N-terminal sequencing, indicating
a blocked N-terminus.
7.3. Summary of result and discussion
The sactipeptide is one of three uncharacterised bacteriocins predicted from the genome of P.
polymyxa #23 with all of the genes expected to be required for biosynthesis, but it was not
observed during purification. Therefore, heterologous cloning and expression were employed
to enhance sactipeptide production.
In this study, the whole sactipeptide gene cluster was isolated from P. polymyxa #23 and
cloned into the pDLL202 plasmid vector, and B. subtilis BS34A was used as the expression
host. It is an E. coli/Bacillus shuttle plasmid enabling the transformation into both E. coli and
B. subtilis. The heterologous cloning and expression of sactipeptide were successfully
performed but each step had to be refined and modified to achieve success. (a) The PCR to

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
157
amplify the large sactipeptide gene cluster had to meet some demands such as; intact PCR
product with few or no nonspecific bands; high yield; high purity; no mutations during
amplification. These requirements were achieved through; i) selection of a gDNA extraction
method that gave long genomic DNA fragments from P. polymyxa #23; ii) selection of a high
fidelity polymerase enzyme, iii) optimisation of PCR mixture components (e.g. primer
concentration), iv) testing of a variety of PCR running conditions to reduce the nonspecific
bands, v) selection of appropriate kits for the tasks of DNA plasmid extraction, gel
purification, cleaning-up for high efficiency of recovery. The GES DNA extraction method
was shown to be suitable to extract intact genomic DNA from P. polymyxa. It may be due to
the activity of guanidine thiocyanate in extraction buffer, which served as an inhibitory agent
of endonuclease abundantly released from P. polymyxa cells after cell breakage. Also, all kits
(plasmid extraction kit, gel purification, PCR product purification) purchased from NEB
(Monarch kit) always gave good DNA yields compared to kits purchased from several other
manufacturers). The Phusion ® High-Fidelity polymerase (NEB) was the most suitable for
amplification of large DNA fragments and resulted in intact and high yields of PCR product
(when compared with Q5 polymerase), and HF buffer supplemented into the PCR mixture
enhanced PCR efficiency.
- (b) There was no success when standard ligation conditions were used to attempt to construct
the fusion expression plasmid. Construction was only successful when Gibson assembly was
used. For Gibson assembly, the PCR amplification primers were redesigned to generate
nucleotide fragments which contained the sactipeptide gene cluster flanked by two regions of
20-nucleotide sequences at the termini. These two regions were designed to be
complementary to sequences at the ends of the linearized plasmid vector.
- (c) Transformation was difficult because of the large size of the fusion plasmid (>10 kbp).
It was achieved by selection of appropriate E. coli types, and transformation methods
(electroporation or natural transformation).
- (d) Purification of sactipeptide from Bacillus transformants was achieved by selection of a
medium in which expression occurred and by the use of sensitive indicator cells for detection
of antimicrobial activity during purification. In our investigation, we found that the
sactipeptide was only produced in LB media, not PB media. And an MRSA strain which can
resistant to chloramphenicol was used as the indicator strain.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
158
The analysis of the growth curve found that the antimicrobial substance was produced at a
stationary growth phase, similar to earlier studies on bacteriocin production (84, 221).
Purification of the sactipeptide from LB liquid culture was achieved via a 2 steps-procedure;
(i) precipitation of the peptide by adding ammonium sulphate to 70% saturation concentration
at 5oC; (ii) purification of the sactipeptide by RP-HPLC employing a C18 column. The pure
sactipeptide was identified with an MW of 3.4 kDa; was heat-stable; sensitive to proteinase
K, pronase E and trypsin; and displayed antimicrobial activity against only Gram-positive
bacteria. However, the amino acid sequence of sactipeptide was not elucidated because N-
terminal sequencing was not successful, suggesting that the N-terminal of the protein is
blocked. The failure of N-terminal sequencing has been frequently reported for highly
modified bacteriocins such as Enterocin AS-48 (222), or subtilomyxin (84), subtilosin A
(223), circularin A and closticin 574 (224)
The MW of the purified sactipeptide was 3.4 kDa, while the MW predicted, based on the
amino acid sequence of the core sequence, was 4,455.81 Da. Based on multiple sequence
alignment between this sactipeptide and previously characterised sactipeptides (obtained from
BACTIBASE database), the #23 sactipeptide sequence is closest to subtilosin A. Both these
two bacteriocins have the same numbers of amino acids residues and have similar MWs of
~3-4 kDa. Therefore, this discrepancy in MW may be a result of a misprediction of the SignalP
software for identifying the signal sequence cleavage position on the precursor, subsequently
causing misprediction of MW of core sequence. To confirm, we submitted sequences of well-
characterised bacteriocin precursors (like amylocyclicin, mersacidin and paenicidin A) to this
software for calculation of cleavage site, but the software suggested incorrect recognition
sites, leading to an incorrect prediction of the MW of core sequence. Although SignalP is
widely used for this purpose, it is clear that the software is sometimes ineffective in the case
of small peptides like bacteriocins. The #23 sactipeptide only depressed growth of Gram-
positive bacteria, while subtilosin A has antimicrobial activity extending to both Gram-
positive bacteria and Gram-negative bacteria (142). This indicates the diversity in
antimicrobial spectra of sactipeptide-type bacteriocins.
The use of a synthetic biology approach to isolate operons from uncultivable strains for
improvement on already isolated operons may be interesting. Although there is no example
of this sort of approach in the literature it may have the potential to greatly expand access to
novel antimicrobial operons from uncultivable bacteria. Metagenomic analysis of
environmental samples could be used as a tool to achieve this purpose. However, for smany

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
159
antimicrobial compound, the bioactive antimicrobial compounds need the expression of many
genes for accessory protein products that participate in the various functions of immunity,
secretion, posttranslational modification, and precursor processing. These genes are often
located together in one operon but are sometimes dispersed and hence difficult to identify.
Due to the ineffectiveness of current genetic systems, metagenomics may be a difficult task
to manipulate large size antimicrobial gene cluster. Currently, many studies mainly
concentrate on developing different heterologous expression systems to enhance the
efficiency to capture the antimicrobial gene clusters derived from cultivable bacteria, such as
TAR System, IR System, Red/ET recombineering (225-227). Alternatively, the studies are
performed to improve heterologous cloning/expression by evaluating the impact of different
media and media components on bacteriocin production, i.e., Garvicin KS (228).
Taken together, the findings in this chapter confirms the usefulness of plasmid pDLL202 for
cloning of the bacteriocin from bacilli and facilitated successful expression to produce the
recombinant bacteriocin. The pure novel #23 sactipeptide can now be further characterised,
both in terms of structural analysis and antimicrobial activity.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
160
Chapter 8 CONCLUSION AND FUTURE WORK
8.1. Conclusion
In the circumstance of emerging antibiotics resistance of current pathogens, humankind is
seeking insights outside of the traditional bacterial reservoirs and increased investment in
other classes of antimicrobial peptides for antibiotic development, such as bacteriocins.
Hypothesizing that there is an abundance of novel bacteriocins unidentified in marine Bacillus
since marine Bacillus/Paenibacillus species have been under-utilised, we therefore selected
an unexplored sea region, the NhaTrang bay (Vietnam Sea) to seek novel bacteriocins, for
application in pharmaceuticals, aquaculture, agriculture and food industries.
From the results of all the experimental chapters, five research questions raised in Chapter 1
have been clarified. Bacillus bacteria with diverse antimicrobial activities could be isolated
from marine environments. The sampling and bacterial isolation work detailed in Chapter 3
resulted in 16.4% of the total spore-forming bacteria isolated from the Vietnam showing clear
antimicrobial activity. Of the isolates, the Bacillus and Paenibacillus, with a diversity of
species, were found, including members of the B. subtilis group (including B. licheniformis,
B. safensis and ubiquitous isolates like B. subtilis, B. amyloliquefaciens), members of the B.
cereus group (B. pacificus) and Paenibacillus (P. polymyxa). The activities against Gram-
positive bacterial indicators were most commonly observed, including against antibiotic-
resistant pathogens (VRE, MRSA); and activities against Gram-negative bacteria, although
limited, were found in the isolate collection. Six isolates (B. halotolerans 01, B.
amyloliquefaciens #06, #08, #11, #13 and P. polymyxa #23) killed 11 or more of the 14
indicators strains, covering a range of bacterial pathogens including food spoilage bacteria,
and human and/or animal pathogens. A high percentage of the antimicrobial substances were
found to be sensitive to proteolytic enzymes and the dominance of antimicrobial activity
against Gram-positive bacteria indicated production of bacteriocins. Therefore, antimicrobial
compounds derived from marine Bacillus/Paenibacillus could potentially be used to address
a range of pathogenic microbial threats to humans and animals. The aims of Chapter 3, to
understand the antimicrobial spectra of spore-forming bacteria in Vietnam Sea, and to select
the promising candidates for genome sequencings, were achieved.
Discovery of novel bacteriocins (and other antimicrobial peptides) is now facilitated by
analysis of bacterial genomes to identify genes encoding these novel compounds. Therefore,
6 marine isolates (4 B. amyloliquefaciens, 1 B. halotolerans #01, 1 P. polymyxa #23), that

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
161
were selected in Chapter 3 were sequenced and the genomic information was used to all the
encoded putative antimicrobial peptides. The in silico screen resulted in the identification of
an abundance of known and novel antimicrobials (bacteriocins, secondary metabolites) from
the 6 genomes. A total of 61 gene clusters of various AMPs were identified, including 41
NRPs compounds (mostly LPs), 20 bacteriocins that comprised 13 different types of
bacteriocins. Of these 13 bacteriocins, 54% (7/13) were novel and belonged to different
subclasses of bacteriocin class I (1 sactipeptide, 1 lassopeptide, 2 thiopeptides, and 3
lantibiotics). The gene organisation of every bacteriocin gene cluster found in this study was
analysed to understand the mechanism of bacteriocin production. Therefore, the aims of the
chapter, to understand the mechanism of antimicrobial production and identification of novel
bacteriocin(s) and select the candidates harbouring novel bacteriocins for in vitro
characterisation, were achieved. Seven gene clusters of novel bacteriocins were identified;
two potentially novel compounds, produced by isolates #23 and isolate #11, were targeted for
purification. The results of the chapter showed that marine B. amyloliquefaciens and P.
polymyxa contained a rich source of novel bacteriocins gene clusters for further in vitro
characterisation
In addition, in this chapter genome comparisons were performed within these marine species
and versus the terrestrially derived closely related species. The findings demonstrated the high
degree of similarity across the genomes, as judged by, ANI values, and the similarity of the
coding sequence (CDS) potential shared between the genomes. Of the three species
investigated, B. amyloliquefaciens had a significantly higher degree of similarity in genomes,
followed by B. halotolerans, but genomes of P. polymyxa isolates were found to be highly
variable and P. polymyxa had a significantly high diversity of gene repertoire, which may
indicate that future studies could efficiently focus on these sorts of isolates to increase the
discovery rate of novel antimicrobial compounds. The high potential for discovery of novel
bacteriocins from P. polymyxa was confirmed by the in silico prediction of putative
antimicrobial peptides encoded by P. polymyxa #23. The analysis predicted the presence of
many gene clusters of various antimicrobial peptides, including two gene clusters relating to
novel NRPs compounds and three clusters of uncharacterised bacteriocins (1 sactipeptide, 1
lassopeptide, 1 lantibiotic), alongside the presence of gene clusters for NRPs and bacteriocin
characterised previously (tridecaptin, polymyxin, fursacidin, paenicidin A). The marine B.
amyloliquefaciens also serves as a second promising pool of novel bacteriocins, with 3 gene
clusters of novel bacteriocins found in 4 genomes (2 thiopeptides, and 1 two component

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
162
lantibiotic). The B. halotolerans contained genes for production of 1 subtilosin A, therefore,
the potential of novel bacteriocin production from this species is limited.
The purification of antimicrobial peptides was carried out for two bacteria; isolate #11 which
contained genes of novel thiopeptide, and isolate #23 which contained genes of 3
uncharacterised bacteriocins (lasso peptide, 1 lantibiotic, and 1 sactipeptide). The aim was to
obtain the novel bacteriocins for in vitro characterisation; however, the novel bacteriocins are
yet to be identified. The purification of antimicrobial peptides produced by isolate #11 in
LPMA elucidated the presence of amylocyclicin bacteriocin ([m/z] = 6,381], a surfactin
([m/z] =1,058 Da], and [m/z= 1,473]) in the culture. The 1,473 Da substance was suspected
to be a novel thiopeptide, or it could also be a fengycin, because these two compounds have
similar MW to this peptide mass. However, the N-terminal sequencing failed to determine the
amino acid sequence of this compound, therefore, production of novel thiopeptide in culture
is yet to be fully confirmed. Purification of antimicrobial peptides from culture of P. polymyxa
identified the presence of polymyxin ([m/z] = 1168.7), paenicidin A ([m/z]= 3290.4), and
tridecaptin ([m/z]= 1550.8). Attempts to purify novel bacteriocins directly from the isolates
was difficult because various lipopeptides and polyketides were co-produced, and these
exhibited strong antimicrobial activities which hide the presence of other antimicrobial
compounds. The production of NRPs (fengycin, tridecaptin, surfactin), excepting polymyxin,
were frequently observed during the latter phase of culture growth. Bacteriocins
(amylocyclicin, paenicidin) production tended to occur at earlier stages of growth. Therefore,
it is necessary to select the correct time-point for harvesting cultures for purification of
bacteriocins. All the Bacillus/Paenibacillus-derived antimicrobial peptides that were purified
could not be analysed by N-terminal sequencing. This probably indicated that the
Bacillus/Paenibacillus derived AMPs were highly modified peptides which contained either
blocked N-terminus, unusual amino acid residues, and/or various cross-linkages resulting in
complex tertiary structure. Therefore, characterisation and sequencing of Bacillus-derived
AMPs were more challenging compared to that generally found with the characterisation of
Lactobacillus bacteriocins. As an alternative, the peptides could be identified by nuclear
magnetic resonance (NMR) techniques, which have been widely used to identify bacteriocins
(229, 230).
The absence of some of the novel bacteriocins and other NRPs compounds, which were
predicted in silico from genomes, during purification could be due to many reasons.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
163
i) Gene clusters relating to these compounds are not transcribed in the conditions produced
by growth in LPMA, or PB medium. It may be activated in other media or requires the
supplement of some factors that can induce expression. For example, the presence of dead
cells of indicators in the culture can activate the production of some bacteriocin via the
quorum sensing system (231, 232). The selection of media for enhancement of bacteriocin
production is a complex process requiring a balance between the most efficient
macronutrients (carbon, nitrogen, phosphate) and the growth-essential micronutrients (zinc,
iron, and manganese) (233, 234). Apart from nutrient effects, the bacteriocin production is
also dependent on the pH, dissolved oxygen level, growth phase (235, 236). The production
of novel sactipeptide only in LB media but not LPMA or PB media can confirm it the media
dependence of sactipeptide production.
ii) The loss of these compounds during purification is possible due to the low detection of
novel bacteriocin. This can happen if the novel bacteriocins are produced at low
concentration; lower than MICs required to kill the indicators. The MIC values of bacteriocins
against indicators are variable depending on types of bacteriocin and types of indicator cells.
And due to the coproduction of non-ribosomally synthesized peptides that commonly exhibit
strong antimicrobial activities B. cereus, the weaker antimicrobial activities of novel
bacteriocin are possibly omitted during purification by accident, particularly when
inappropriate indicators selected.
iii) Loss of compounds during purification is also possible due to the limitation of the
extraction procedure. Bacteriocin (with MWs of less than 2 kDa) can be lost during the
dialysis using 3 kDa cut-off membrane. Therefore, it would be advisable to apply MALDI-
TOF MS analysis at the beginning of the procedure to check the presence/loss of all peptides
during purification.
However, some approaches could be modified to obtain the pure novel bacteriocins for
characterisation; including, a) generation of a mutant which is deficient in NRPs production,
which can mask the presence of other AMPs, and fermentation of this mutant for bacteriocin
production, and; b) heterologous cloning and expression of the gene clusters of novel peptides.
a) The first approach could be achieved by deleting the sfp gene and bacA from the wild-type
isolates. The sfp gene encodes the sfp - 4'-phosphopantetheinyl transferase, required for
activation of synthase enzymes in the biosynthesis of lipopeptide and polyketides. The bacA

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
164
gene is a structural gene encoding bacilysin. This technique was used previously to
characterise the antimicrobial peptides produced by B. amyloliquefaciens FZB42 (237)
b) The second approach can be an ideal method to produce, heterologously, the novel
bacteriocins, when the gene clusters of these bacteriocins were well annotated, as in the last
chapter. However, the selection of specific expression vectors and host for successful
bacteriocin production must be carefully considered. Successful expression of bacteriocin
requires cloning of the whole bacteriocin biosynthetic gene cluster into a vector to generate
the large expression plasmid. The larger plasmid can be challenging to construct, manipulate
and transform into different hosts for expression analysis. Particularly, in the case of
thiopeptide, of which the biosynthetic gene clusters are large (>12 bps), and required a large
number of optimization steps and trials
In chapter 07, the heterologous cloning and expression methods were employed to produce
one of three uncharacterised bacteriocins found in the genome of P. polymyxa #23. This
bacteriocin was predicted in silico from the genome, and its biosynthesis was predicted to be
encoded by an 8.5 kbp gene cluster containing all accessories genes required for biosynthesis;
however, it was not observed during purification from the wild-type isolate. The heterologous
cloning and expression of the sactipeptide gene cluster employed a shuttle E. coli/Bacillus
pDLL202 plasmid vector and was expressed in B. subtilis BS54B. Attempts with lots of
optimization steps were made and finally led to the successful production of sactipeptide
(question 5). Optimization included i) Selection of an appropriate method to extract the intact
gDNA from wild-type P. polymyxa #23. In this investigation, the genomic DNA of wild-type
P. polymyxa was extracted using a guanidine thiocyanate method (GES). ii) Amplification of
the sactipeptide gene cluster from intact gDNA needs a specific pair of primers, Phusion High-
Fidelity Master-Mix (Thermo Scientific), and HF buffer as a supplement to improve amplicon
yield. iii) Preparation of plasmid backbone from native pDLL202 plasmid by enzymatic
digestion of native plasmid. iv) Joining the gene cluster and plasmid vector DNA molecules
were only achieved with the aid of the Gibson assembly method. v) Transformation of fusion
plasmid into B. subtilis cell required intermediate transformations employing E. coli hosts (E.
coli Top10 and E. coli JIR 702 cells), which can receive a large size fusion plasmid to control
the plasmid copies as well as produce the multimeric form of the fusion plasmid. vi) Finally,
the expression of a novel sactipeptide in Bacillus transformant cells was successful in LB
medium, and MRSA (resistant to chloramphenicol) was used as an indicator. The sactipeptide
had MW of 3,404 Da, was sensitive to protease, proteianase K, trypsin, but not lipase. It had

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
165
antimicrobial activity against most Gram-positive bacteria. Unfortunately, the amino acid
sequence of the sactipeptide could not be confirmed by N-terminal sequencing, suggesting
the presence of a block at the N-terminus.
The therapeutic potential of isolate #11 is unclear as the production of the predicted
thiopeptide in vitro could not be confirmed. However, the potential to use isolate #23 in
therapeutic applications is clear, with tridecaptin and polymyxin both produced by in vitro
grown cultures. These antimicrobial compounds are currently proven as useful AMPs to cure
hospital-acquired Gram-negative bacteria, particularly against carbapenem-resistant
Enterobacteriaceae. Also, the novel sactipeptide produced heterologously exhibits activity
against MRSA and VRE, therefore, it displayed potential as an antimicrobial for the treatment
of antibiotic-resistant pathogens. Nevertheless, this potential requires more studies in future
to characterise the molecules and properties such as crystallisation of molecules,
determination of toxicity to human cells, stability analysis under different physical condition.
They could be used to protect plants against the invasion and growth of various phyto-fungi
and/or pathogenic bacteria or to maintain the microbiota in soils and water without usage of
antibiotics or pesticides. These isolates could be effective solutions to promote
environmentally friendly, and sustainable, agriculture and aquaculture. In addition, the
isolates were observed to have the ability to degrade different organic substances, like starch,
cellulose, and protein, indicating that these bacteria can produce enzymes like amylase,
cellullase and protease. These enzymes are currently used in many manufacturing processes
and the waste treatment industry. The marine-derive enzymes identified in this study are
effective enzymes with some unusual properties, that have potential for a wide range of
application. This may be, because of the highly variable environment from which they are
derived. A further analysis of the collection of marine isolates established at the start of this
study can be expected to yield additional candidates for application in health and industry
8.2. Future work
There are many avenues for future research, including:
Sequencing the #23’s novel sactipeptide and an un-identified antimicrobial peptide
produced by isolate #11 that was considered a thiopeptide. Attempts could be made using de
novo sequencing techniques or NMR methods.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
166
The deletion of the structural genes of the thiopeptide from the genome to confirm the above
antimicrobial peptide as a thiopeptide.
Characterisation of novel bacteriocins, including; bacteriocin crystallization for structural
analysis, mode of action, MIC against selected pathogens, and optimizing media culture for
optimal novel bacteriocin production.
Testing the marine isolates to develop into probiotic products for animal-food supplements
or for aquaculture since the marine Bacillus can tolerate high salt concentration.
Testing the marine bacteriocins to develop into food preservatives or antibiotics.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
167
REFERENCE
1. Lobanovska M, Pilla G. Penicillin's discovery and antibiotic resistance: lessons for the future? The Yale Journal of Biology and Medicine. 2017;90:135-45.
2. Davies J, Davies D. Origins and evolution of antibiotic resistance. Microbiology and Molecular Biology Reviews 2010;74:417-33.
3. Llor C, Bjerrum L. Antimicrobial resistance: risk associated with antibiotic overuse and initiatives to reduce the problem. Therapeutic Advances in Drug Safety. 2014;5:229-41.
4. Babakhani S, Oloomi M. Transposons: the agents of antibiotic resistance in bacteria. Journal of Basic Microbiology. 2018;58:905-17.
5. Bennett PM. Plasmid encoded antibiotic resistance: acquisition and transfer of antibiotic resistance genes in bacteria. British Journal of Pharmacology and Chemotherapy. 2008;153:347-57.
6. Partridge S, Kwong S, Firth N, Jensen S. Mobile genetic elements associated with antimicrobial resistance. Clinical Microbiology Reviews. 2018;31.
7. Evans BA, Hamouda A, Amyes SG. The rise of carbapenem-resistant Acinetobacter baumannii. Current Pharmaceutical Design. 2013;19:223-38.
8. Perez F, Van Duin D. Carbapenem-resistant Enterobacteriaceae: a menace to our most vulnerable patients. Cleveland Clinic Journal of Medicine. 2013;80:225-33.
9. Walters MS, Grass JE, Bulens SN, Hancock EB, Phipps EC, Muleta D, et al. Carbapenem-Resistant Pseudomonas aeruginosa at US emerging infections program sites, 2015. Emerging Infectious Diseases. 2019;25:1281-8.
10. Bouchiat C, Curtis S, Spiliopoulou I, Bes M, Cocuzza C, Codita L, et al. MRSA infections among patients in the emergency department: a European multicentre study. Journal of Antimicrobial Chemotherapy. 2017;72:372-5.
11. Kafil HS, Asgharzadeh M. Vancomycin-resistant Enteroccus faecium and Enterococcus faecalis isolated from education hospital of Iran. Maedica (Buchar). 2014;9:323-7.
12. Spellberg B, Powers JH, Brass EP, Miller LG, Edwards JE, Jr. Trends in antimicrobial drug development: implications for the future. Clinical Infectious Diseases. 2004;38:1279-86.
13. Zaman SB, Hussain M, Nye R, Mehta V, Mamun T, Hossain N. A review on antibiotic resistance: alarm bells are ringing. Cureus. 2017;9:e1403.
14. Tortorella E, Tedesco P, F. PE, January GG, Fani R, Jaspars M, et al. Antibiotics from deep-sea microorganisms: current discoveries and perspectives. Marine drugs. 2018;16:355.
15. Kogure K, Simidu U, Taga N. A tentative direct microscopic method for counting living marine bacteria. Canadian Journal of Microbiology. 1979;25:415-20.
16. Rezaei Javan R, van Tonder AJ, King JP, Harrold CL, Brueggemann AB. Genome sequencing reveals a large and diverse repertoire of antimicrobial peptides. Frontiers in Microbiology. 2018;9(2012).
17. Taylor MW, Radax R, Steger D, Wagner M. Sponge-associated microorganisms: evolution, ecology, and biotechnological Potential. Microbiology and Molecular Biology Reviews. 2007;71:295.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
168
18. Anderson RE, Reveillaud J, Reddington E, Delmont TO, Eren AM, McDermott JM, et al. Genomic variation in microbial populations inhabiting the marine subseafloor at deep-sea hydrothermal vents. Nature Communications. 2017;8:1114.
19. Behrens HM, Six A, Walker D, Kleanthous C. The therapeutic potential of bacteriocins as protein antibiotics. Emerging Topics in Life Sciences. 2017;1:65-74.
20. Silva CCG, Silva SPM, Ribeiro SC. Application of bacteriocins and protective cultures in dairy food preservation. Frontiers in Microbiology. 2018;9:594-.
21. Sumi CD, Yang BW, Yeo LC, Hahm YT. Antimicrobial peptides of the genus Bacillus: a new era for antibiotics. Canadian Journal of Microbiology. 2015;61:93-103.
22. Zhao X, Kuipers OP. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genomics. 2016;17:882.
23. Egan K, Field D, Ross RP, Cotter PD, Hill C. In silico prediction and exploration of potential bacteriocin gene clusters within the bacterial genus Geobacillus. Frontiers in Microbiology. 2018;9:2116.
24. Hancock RE. Peptide antibiotics. Lancet. 1997;349:418-22.
25. Stachelhaus T, Marahiel MA. Modular structure of genes encoding multifunctional peptide synthetases required for non-ribosomal peptide synthesis. FEMS Microbiology Letter. 1995;125:3-14.
26. Challis GL, Naismith JH. Structural aspects of non-ribosomal peptide biosynthesis. Current Opinion in Structural Biology. 2004;14:748-56.
27. Chen XH, Vater J, Piel J, Franke P, Scholz R, Schneider K, et al. Structural and functional characterization of three polyketide synthase gene clusters in Bacillus amyloliquefaciens FZB 42. Journal of Bacteriology. 2006;188:4024-36.
28. Guzmán-Chávez F, Zwahlen RD, Bovenberg RAL, Driessen AJM. Engineering of the filamentous fungus Penicillium chrysogenum as cell factory for natural products. Frontiers in Microbiology. 2018:2768.
29. Meena K, Kanwar S. Lipopeptides as the antifungal and antibacterial agents: applications in food safety and therapeutics. BioMed Research International. 2015;2015:1-9.
30. Luo C, Liu X, Zhou X, Guo J, Truong J, Wang X, et al. Unusual biosynthesis and structure of locillomycins from Bacillus subtilis 916. Applied and Environmental Microbiology. 2015;81:6601-9.
31. Loeffler W, Tschen JSM, Vanittanakom N, Kugler M, Knorpp E, Hsieh TF, et al. Antifungal effects of bacilysin and fengymycin from Bacillus subtilis F-29-3 a comparison with activities of other Bacillus antibiotics. Journal of Phytopathology. 1986;115:204-13.
32. Rogers HJ, Newton GG, Abraham EP. Production and purification of bacilysin. The Biochemical Journal. 1965;97:573-8.
33. Nishie M, Nagao J, Sonomoto K. Antibacterial peptides "bacteriocins": an overview of their diverse characteristics and applications. Biocontrol Science. 2012;17:1-16.
34. Jack RW, Tagg JR, Ray B. Bacteriocins of Gram-positive bacteria. Microbiological Reviews. 1995;59:171-200.
35. Es B, Bhat SG. Marine bacteriocins: a review. Journal of Bacteriology and Mycology Open Access. 2016;2.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
169
36. Abriouel H, Franz CM, Ben Omar N, Gálvez A. Diversity and applications of Bacillus bacteriocins. FEMS Microbiology Reviews. 2011;35:201-32.
37. Mokoena M. Lactic acid bacteria and their bacteriocins: classification, biosynthesis and applications against uropathogens: A mini-review. Molecules. 2017;22:1255.
38. Ortega MA, van der Donk WA. New insights into the biosynthetic logic of ribosomally synthesized and post-translationally modified peptide natural products. Cell Chemical Biology. 2016;23:31-44.
39. McAuliffe O, Ross RP, Hill C. Lantibiotics: structure, biosynthesis and mode of action. FEMS Microbiology Reviews. 2001;25:285-308.
40. Yu Y, Zhang Q, van der Donk WA. Insights into the evolution of lanthipeptide biosynthesis. Protein Science. 2013;22:1478-89.
41. Chatterjee S, Chatterjee S, Lad SJ, Phansalkar MS, Rupp RH, Ganguli BN, et al. Mersacidin, a new antibiotic from Bacillus. Fermentation, isolation, purification and chemical characterization. The Journal of Antibiotics. 1992;45:832-8.
42. He Z, Kisla D, Zhang L, Yuan C, Green-Church KB, Yousef AE. Isolation and identification of a Paenibacillus polymyxa strain that coproduces a novel lantibiotic and polymyxin. Applied and Environmental Microbiology. 2007;73:168-78.
43. Lohans CT, Huang Z, van Belkum MJ, Giroud M, Sit CS, Steels EM, et al. Structural characterization of the highly cyclized lantibiotic paenicidin A via a partial desulfurization/reduction strategy. Journal of the American Chemical Society. 2012;134:19540-3.
44. Gabrielsen C, Brede DA, Nes IF, Diep DB. Circular bacteriocins: biosynthesis and mode of action. Applied and Environmental Microbiology. 2014;80:6854-62.
45. Westers H, Braun PG, Westers L, Antelmann H, Hecker M, Jongbloed JD, et al. Genes involved in SkfA killing factor production protect a Bacillus subtilis lipase against proteolysis. Applied and Environmental Microbiology. 2005;71:1899-908.
46. Cherif A, Ouzari H, Daffonchio D, Cherif H, Ben Slama K, Hassen A, et al. Thuricin 7: a novel bacteriocin produced by Bacillus thuringiensis BMG1.7, a new strain isolated from soil. Letters in Applied Microbiology. 2001;32(4):243-7.
47. Rea MC, Sit CS, Clayton E, O'Connor PM, Whittal RM, Zheng J, et al. Thuricin CD, a posttranslationally modified bacteriocin with a narrow spectrum of activity against Clostridium difficile. Proceedings of the National Academy of Sciences of the United States of America. 2010;107:9352-7.
48. Poorinmohammad N, Bagheban-Shemirani R, Hamedi J. Genome mining for ribosomally synthesised and post-translationally modified peptides (RiPPs) reveals undiscovered bioactive potentials of actinobacteria. Antonie Van Leeuwenhoek. 2019;112:1477-99.
49. Burkhart BJ, Hudson GA, Dunbar KL, Mitchell DA. A prevalent peptide-binding domain guides ribosomal natural product biosynthesis. Nature Chemical Biology. 2015;11:564-70.
50. Budič M, Rijavec M, Petkovšek Z, Zgur-Bertok D. Escherichia coli bacteriocins: antimicrobial efficacy and prevalence among isolates from patients with bacteraemia. PloS one. 2011;6:e28769-e.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
170
51. Scholz R, Molohon KJ, Nachtigall J, Vater J, Markley AL, Süssmuth RD, et al. Plantazolicin, a novel microcin B17/streptolysin S-Like natural product from Bacillus amyloliquefaciens FZB42. Journal of Bacteriology. 2011;193:215.
52. Hsieh YSY, Wilkinson L, O’Connell MR, Mackay JP, Matthews JM, Payne RJ. Synthesis of the bacteriocin glycopeptide sublancin 168 and S-glycosylated bariants. Organic Letters. 2012;14:1910-3.
53. Hyronimus B, Le Marrec C, Urdaci MC. Coagulin, a bacteriocin-like inhibitory substance produced by Bacillus coagulans I4. Journal of Applied Microbiology. 1998;85:42-50.
54. Gong W, Wang J, Chen Z, Xia B, Lu G. Solution structure of LCI, a novel antimicrobial peptide from Bacillus subtilis. Biochemistry. 2011;50:3621-7.
55. Holland IB. The purification and properties of megacin, a bacteriocin from Bacillus megaterium. The Biochemical Journal. 1961;78:641-8.
56. Braun V, Schaller K, Wabl MR. Isolation, characterization, and action of colicin M. Antimicrobial Agents and Chemotherapy. 1974;5:520-33.
57. Yang JY, Wang P, Li CY, Dong S, Song XY, Zhang XY, et al. Characterization of a new M13 metallopeptidase from deep-sea Shewanella sp. E525-6 and mechanistic insight into its catalysis. Frontiers in Microbiology. 2016;6:1498-.
58. Gražina J, Schleining G, Viskelis P, Urbonaviciene D, Eidukonyte D, Bobinas C. Fermentation processes using lactic acid bacteria producing bacteriocins for preservation and improving functional properties of food products. Advances in Applied Biotechnology. Advances in Applied Biotechnology2012.
59. Bastos Mdo C, Coelho ML, Santos OC. Resistance to bacteriocins produced by Gram-positive bacteria. Microbiology. 2015;161:683-700.
60. Gravesen A, Jydegaard Axelsen AM, Mendes da Silva J, Hansen TB, Knøchel S. Frequency of bacteriocin resistance development and associated fitness costs in Listeria monocytogenes. Applied and Environmental Microbiology. 2002;68:756-64.
61. A R S, Lipton A, Aishwarya Ms, Dhivya R. Isolation of a bacteriocin-producing Lactococcus lactis and application of its bacteriocin to manage spoilage bacteria in high-value marine fish under different storage temperatures. Applied Biochemistry and Biotechnology. 2012;167:1280-9.
62. Natesan S, Muthuraman S, Gopal S. Probiotic effect of Lactobacillus acidophilus against vibriosis in juvenile shrimp (Penaeus monodon). African Journal of Biotechnology. 2012;11:15811-8.
63. Lemos ML, Toranzo AE, Barja JL. Antibiotic activity of epiphytic bacteria isolated from intertidal seaweeds. Microbial Ecology. 1985;11:149-63.
64. Rajaram G, Manivasagan P, Thilagavathi B, Saravanakumar A. Purification and characterization of a bacteriocin produced by Lactobacillus lactis isolated from marine environment. Advance Journal of Food Science and Technology. 2010;2:138-44.
65. Anacarso I, Messi P, Condò C, Iseppi R, Bondi M, Sabia C, et al. A bacteriocin-like substance produced from Lactobacillus pentosus 39 is a natural antagonist for the control of Aeromonas hydrophila and Listeria monocytogenes in fresh salmon fillets. Lebensmittel-Wissenschaft & Technologie Food Science and Technology. 2014;55:604–11.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
171
66. Ringø E, Hoseinifar SH, Ghosh K, Doan HV, Beck BR, Song SK. Lactic acid bacteria in finfish-An update. Frontiers in Microbiology. 2018;9:1818-.
67. Singh R, Kandasamy S, Jayalakshmi S, Kumar S, Selvi C. Isolation and production of bacteriocin by marine Lactobacillus fermentum SBS001. International Journal of Current Microbiology and Applied Sciences. 2013;2:67-73.
68. Andryukov BG, Mikhaylov VV, Besednova NN, Zaporozhets TS, Bynina MP, Matosova EV. The bacteriocinogenic potential of marine microorganisms. Russian Journal of Marine Biology. 2018;44:433-41.
69. Sarika AR, Lipton AP, Aishwarya MS, Dhivya RS. Isolation of a bacteriocin-producing lactococcus lactis and application of its bacteriocin to manage spoilage bacteria in high-value marine fish under different storage temperatures. Applied Biochemistry and Biotechnology. 2012;167:1280-9.
70. Stoffels G, Nes IF, Guthmundsdóttir A. Isolation and properties of a bacteriocin-producing Carnobacterium piscicola isolated from fish. Journal of Applied Microbiology. 1992;73:309-16.
71. Tahiri I, Desbiens M, Benech R, Kheadr E, Lacroix C, Thibault S, et al. Purification, characterization and amino acid sequencing of divergicin M35: a novel class IIa bacteriocin produced by Carnobacterium divergens M35. International Journal of Food Microbiology. 2004;97:123-36.
72. Métivier A, Pilet MF, Dousset X, Sorokine O, Anglade P, Zagorec M, et al. Divercin V41, a new bacteriocin with two disulphide bonds produced by Carnobacterium divergens V41: primary structure and genomic organization. Microbiology. 1998;144 2837-44.
73. Nilsson L, Huss HH, Gram L. Inhibition of Listeria monocytogenes on cold-smoked salmon by nisin and carbon dioxide atmosphere. International Journal of Food Microbiology. 1997;38:217-27.
74. Yamazaki K, Suzuki M, Kawai Y, Inoue N, Montville TJ. Purification and characterization of a novel class IIa bacteriocin, piscicocin CS526, from surimi-associated Carnobacterium piscicola CS526. Applied and Environmental Microbiology. 2005;71:554.
75. Bhugaloo VP, Dousset X, Metivier A, Sorokine O, Anglade P, Boyaval P, et al. Purification and amino acid sequences of piscicocins V1a and V1b, two class IIa bacteriocins secreted by Carnobacterium piscicola V1 that display significantly different levels of specific inhibitory activity. Applied and Environmental Microbiology. 1996;62:4410-6.
76. Umemura E, Wakiyama Y, Kumura K, Ueda K, Masaki S, Watanabe T, et al. Synthesis of novel lincomycin derivatives and their in vitro antibacterial activities. The Journal of Antibiotics. 2013;66:195-8.
77. Pinto AL, Fernandes M, Pinto C, Albano H, Castilho F, Teixeira P, et al. Characterization of anti-Listeria bacteriocins isolated from shellfish: potential antimicrobials to control non-fermented seafood. International Journal of Food Microbiology. 2009;129:50-8.
78. Sugita H, Matsuo N, Hirose Y, Iwato M, Deguchi Y. Vibrio sp. strain NM 10, isolated from the intestine of a Japanese coastal fish, has an inhibitory effect against Pasteurella piscicida. Applied and Environmental Microbiology. 1997;63:4986-9.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
172
79. Bergh Ø. Bacteria associated with early life stages of halibut, Hippoglossus hippoglossus L., inhibit growth of a pathogenic Vibrio sp. Journal of Fish Diseases. 1995;18:31-40.
80. Selvendran M, B. MM. Studies on novel bacteriocin like inhibitory substance (BLIS) from microalgal symbiotic Vibrio spp MMB2 and its activity against aquatic bacterial pathogens. Journal of Applied Pharmaceutical Science. 2013;3:169-75.
81. Nguyen VD, Pham TT, Nguyen TH, Nguyen TT, Hoj L. Screening of marine bacteria with bacteriocin-like activities and probiotic potential for ornate spiny lobster (Panulirus ornatus) juveniles. Fish & Shellfish Immunology. 2014;40:49-60.
82. Smitha S, Bhat SG. Thermostable bacteriocin BL8 from Bacillus licheniformis isolated from marine sediment. Journal of Applied Microbiology. 2013;114:688-94.
83. Bindiya ES, Tina KJ, Sasidharan RS, Bhat SG. BaCf3: highly thermostable bacteriocin from Bacillus amyloliquefaciens BTSS3 antagonistic on food-borne pathogens. 3 Biotech. 2019;9:136.
84. Phelan RW, Barret M, Cotter PD, O'Connor PM, Chen R, Morrissey JP, et al. Subtilomycin: a new lantibiotic from Bacillus subtilis strain MMA7 isolated from the marine sponge Haliclona simulans. Marine Drugs. 2013;11:1878-98.
85. Chopra L, Singh G, Choudhary V, Sahoo DK. Sonorensin: an antimicrobial peptide, belonging to the heterocycloanthracin subfamily of bacteriocins, from a new marine isolate, Bacillus sonorensis MT93. Applied and Environmental Microbiology. 2014;80:2981-90.
86. Prieto ML, O'Sullivan L, Tan SP, McLoughlin P, Hughes H, O'Connor PM, et al. Assessment of the bacteriocinogenic potential of marine bacteria reveals lichenicidin production by seaweed-derived Bacillus spp. Marine Drugs. 2012;10:2280-99.
87. K. DRS, Gurunathan J. Isolation and partial characterisation of new bacteriocin from Bacillus pulmius DR2 isolated from sea water. CIBTech Journal of Microbiology 2012;1:33-41.
88. Rezaei M, Ghanbari M, Soltani M, Shahhosseini G, Abedian Kenari A. Production of bacteriocin by a novel Bacillus sp. strain RF 140, an intestinal bacterium of Caspian Frisian Roach (Rutilus frisii kutum). Journal of Biotechnology. 2008;136.
89. Ahmad A, Hamid R, Dada AC, Usup G. Pseudomonas putida strain FStm2 isolated from shark skin: a potential source of bacteriocin. Probiotics and Antimicrobial Proteins. 2013;5:165-75.
90. Velho-Pereira S, Kamat NM. Antimicrobial screening of actinobacteria using a modified cross-streak method. Indian Journal of Pharmaceutical Sciences. 2011;73:223-8.
91. Magaldi S, Mata-Essayag S, Hartung de Capriles C, Perez C, Colella MT, Olaizola C, et al. Well diffusion for antifungal susceptibility testing. International Journal of Infectious Diseases. 2004;8:39-45.
92. Harris LJ, Daeschel MA, Stiles ME, Klaenhammer TR. Antimicrobial activity of lactic acid bacteria against Listeria monocytogenes. Journal of Food Protection. 1989;52:384-7.
93. Fujimoto S, Nakagami Y, Kojima F. Optimal bacterial DNA isolation method using bead-beating technique. Memoirs of Kyushu University School of Health Sciences. 2004;3.
94. de Bruin OM, Birnboim HC. A method for assessing efficiency of bacterial cell disruption and DNA release. BMC Microbiology. 2016;16:197-.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
173
95. Parker D, Martin FJ, Soong G, Harfenist BS, Aguilar JL, Ratner AJ, et al. Streptococcus pneumoniae DNA Initiates Type I Interferon Signaling in the Respiratory Tract. mBio. 2011;2:e00016-11.
96. Sillanpää J, Nallapareddy SR, Singh KV, Ferraro MJ, Murray BE. Adherence characteristics of endocarditis-derived Streptococcus gallolyticus sp. gallolyticus (Streptococcus bovis biotype I) isolates to host extracellular matrix proteins. FEMS Microbiology Letters. 2008;289:104-9.
97. Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research. 1994;22:4673-80.
98. Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution. 2016;33:1870-4.
99. Pitcher DG, Saunders NA, Owen RJ. Rapid extraction of bacterial genomic DNA with guanidium thiocyanate. Letters in Applied Microbiology. 1989;8:151-6.
100. Coil D, Jospin G, Darling AE. A5-miseq: an updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics. 2015;31:587-9.
101. Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, et al. The RAST server: rapid annotations using subsystems technology. BMC Genomics. 2008;9:75.
102. Contreras-Moreira B, Vinuesa P. GET_HOMOLOGUES, a versatile software package for scalable and robust microbial pangenome analysis. Applied and Environmental Microbiology. 2013;79:7696-701.
103. Li L, Stoeckert C, Roos D. OrthoMCL: Identification of ortholog groups for eukaryotic genomes. Genome Research. 2003;13:2178-89.
104. Darling AC, Mau B, Blattner FR, Perna NT. MAUVE: multiple alignment of conserved genomic sequence with rearrangements. Genome Research. 2004;14:1394-403.
105. Overbeek RA, Olson R, Pusch GD, Olsen GJ, Davis JJ, Disz T, et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Research. 2014;42:D206-D14.
106. Hiller NL, Janto B, Hogg JS, Boissy R, Yu S, Powell E, et al. Comparative genomic analyses of seventeen Streptococcus pneumoniae strains: insights into the pneumococcal supragenome. Journal of Bacteriology. 2007;189:8186-95.
107. Tettelin H, Masignani V, Cieslewicz MJ, Donati C, Medini D, Ward NL, et al. Genome analysis of multiple pathogenic isolates of Streptococcus agalactiae: implications for the microbial "pan-genome". Proceedings of the National Academy of Sciences of the United States of America. 2005;102:13950-5.
108. Medema MH, Blin K, Cimermancic P, de Jager V, Zakrzewski P, Fischbach MA, et al. antiSMASH: rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Research. 2011;39(Web Server issue):39-46.
109. van Heel AJ, de Jong A, Song C, Viel JH, Kok J, Kuipers OP. BAGEL4: a user-friendly web server to thoroughly mine RiPPs and bacteriocins. Nucleic Acids Research. 2018;46:W278-w81.
110. Hammami R, Zouhir A, Ben Hamida J, Fliss I. BACTIBASE: a new web-accessible database for bacteriocin characterization. BMC Microbiology. 2007;7:89.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
174
111. Nielsen H. Predicting secretory proteins with SignalP. Methods in Molecular Biology. 2017;1611:59-73.
112. Gasteiger E, Gattiker A, Hoogland C, Ivanyi I, Appel RD, Bairoch A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Research. 2003;31:3784-8.
113. Krogh A, Larsson B, von Heijne G, Sonnhammer EL. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. Journal of Molecular Biology. 2001;305:567-80.
114. Wingfield P. Protein precipitation using ammonium sulfate. Current Protocols In Protein Science. 2001;Appendix 3:Appendix-3F.
115. Counterman AE, Thompson MS, Clemmer DE. Identifying a protein by MALDI-TOF mass spectrometry: an experiment for the undergraduate laboratory. Journal of Chemical Education. 2003;80:177.
116. Clark AE, Kaleta EJ, Arora A, Wolk DM. Matrix-assisted laser desorption ionization-time of flight mass spectrometry: a fundamental shift in the routine practice of clinical microbiology. Clinical Microbiology Reviews. 2013;26:547-603.
117. Frank AM, Savitski MM, Nielsen ML, Zubarev RA, Pevzner PA. De novo peptide sequencing and identification with precision mass spectrometry. Journal of Proteome Research. 2007;6(1):114-23.
118. Reim DF, Speicher DW. N-terminal sequence analysis of proteins and peptides. Current Protocols in Protein Science. 2001;Chapter 11:Unit 11.0.
119. Schägger H. Tricine-SDS-PAGE. Nature Protocols. 2006;1:16-22.
120. Losteinkit C, Uchiyama K, Ochi S, Takaoka T, Nagahisa K, Shioya S. Characterization of bacteriocin N15 produced by Enterococcus faecium N15 and cloning of the related genes. Journal of Bioscience and Bioengineering. 2001;91:390-5.
121. Elshaghabee FMF, Rokana N, Gulhane RD, Sharma C, Panwar H. Bacillus as potential probiotics: status, concerns, and future perspectives. Frontier in Microbiology. 2017;8:1490.
122. Hong HA, Duc le H, Cutting SM. The use of bacterial spore formers as probiotics. FEMS Microbiology Reviews. 2005;29:813-35.
123. Laport MS, Santos OC, Muricy G. Marine sponges: potential sources of new antimicrobial drugs. Current Pharmaceutical Biotechnology. 2009;10:86-105.
124. Gutowski EZ, Klein C, Siegers K, Bohm K, Hammelmann M, Entian KD. Growth phase-dependent regulation and membrane localization of SpaB, a protein involved in biosynthesis of the lantibiotic subtilin. Applied and Environmental Microbiology. 1994;60:1-11.
125. Ivanova EP, Vysotskii MV, Svetashev VI, Nedashkovskaya OI, Gorshkova NM, Mikhailov VV, et al. Characterization of Bacillus strains of marine origin. International Microbiology. 1999;2:267-71.
126. Siefert JL, Larios-Sanz M, Nakamura LK, Slepecky RA, Paul JH, Moore ER, et al. Phylogeny of marine Bacillus isolates from the gulf of Mexico. Current Microbiology. 2000;41:84-8.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
175
127. Jangra M, Kaur M, Tambat R, Rana R, Maurya SK, Khatri N, et al. Tridecaptin M, a new variant discovered in mud bacterium, shows activity against colistin- and extremely drug-resistant Enterobacteriaceae. Antimicrobial Agents and Chemotherapy. 2019;63:e00338-19.
128. Fernandes P. Marine enzymes and food industry: insight on existing and potential interactions. Frontiers in Marine Science. 2014;1.
129. Ravi RK, Walton K, Khosroheidari M. MiSeq: A next generation sequencing platform for genomic analysis. Methods in Molecular Biology. 2018;1706:223-32.
130. Song Y, Sun H, Du J. Identification and tissue distribution of chemosensory protein and odorant binding protein genes in Tropidothorax elegans Distant (Hemiptera: Lygaeidae). Scientific Reports. 2018;8:7803.
131. Li J, Qu X, He X, Duan L, Wu G, Bi D, et al. ThioFinder: a web-based tool for the identification of thiopeptide gene clusters in DNA sequences. PloS One. 2012;7:e45878-e.
132. Chen XH, Koumoutsi A, Scholz R, Eisenreich A, Schneider K, Heinemeyer I, et al. Comparative analysis of the complete genome sequence of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Nature Biotechnology. 2007;25:1007-14.
133. Scholz R, Vater J, Budiharjo A, Wang Z, He Y, Dietel K, et al. Amylocyclicin, a novel circular bacteriocin produced by Bacillus amyloliquefaciens FZB42. Journal of Bacteriology. 2014;196.
134. K. TK, Kikuchi Y, Sano K, Y., Shintani S, Yakushiji M, Kuramitsu HK, et al. Characterization of a potential ABC-type bacteriocin exporter protein from Treponema denticola. BMC Oral Health. 2016;17:18-.
135. Altena K, Guder A, Cramer C, Bierbaum G. Biosynthesis of the lantibiotic mersacidin: organization of a type B lantibiotic gene cluster. Applied and Environmental Microbiology. 2000;66:2565-71.
136. Higgins CF. ABC transporters: from microorganisms to man. Annual Review of Cell and Developmental Biology. 1992;8:67-113.
137. Tanaka KJ, Song S, Mason K, Pinkett HW. Selective substrate uptake: The role of ATP-binding cassette (ABC) importers in pathogenesis. Biochimica et Biophysica Acta Biomembranes. 2018;1860:868-77.
138. Håvarstein LS, Diep DB, Nes IF. A family of bacteriocin ABC transporters carry out proteolytic processing of their substrates concomitant with export. Molecular Microbiology. 1995;16:229-40.
139. Tierney ARP, Rather PN. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiology. 2019;14:533-52.
140. Medema MH, Takano E, Breitling R. Detecting sequence homology at the gene cluster level with MultiGeneBlast. Molecular Biology and Evolution. 2013;30:1218-23.
141. Stein T, Düsterhus S, von Renthe-Fink A, Entian KD. Subtilosin production by two Bacillus subtilis subspecies and variance of the sbo-alb cluster. Applied and Anvironmental Microbiology. 2004;70:2349-53.
142. Shelburne CE, An FY, Dholpe V, Ramamoorthy A, Lopatin DE, Lantz MS. The spectrum of antimicrobial activity of the bacteriocin subtilosin A. Journal of Antimicrobial Chemotherapy. 2007;59:297-300.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
176
143. Flühe L, Marahiel MA. Radical S-adenosylmethionine enzyme catalyzed thioether bond formation in sactipeptide biosynthesis. Current Opinion in Chemical Biology. 2013;17:605-12.
144. Dunbar KL, Melby JO, Mitchell DA. YcaO domains use ATP to activate amide backbones during peptide cyclodehydrations. Nature Chemical Biology. 2012;8:569-75.
145. Banala S, Ensle P, Süssmuth RD. Total synthesis of the ribosomally synthesized linear azole-containing peptide plantazolicin A from Bacillus amyloliquefaciens. Angewandte Chemie. 2013;52:9518-23.
146. van Belkum MJ, Lohans CT, Vederas JC. Draft genome sequences of Paenibacillus polymyxa NRRL B-30509 and Paenibacillus terrae NRRL B-30644, strains from a poultry environment that produce tridecaptin A and paenicidins. Genome Announcements. 2015;3:e00372-15.
147. Lee H, Kim H. Lantibiotics, class I bacteriocins from the genus Bacillus. Journal of Microbiology and Biotechnology. 2011;21:229-35.
148. Vidya Prabhakar K, Lingala V, Peele A, Mikkili I, Vekateswarulu TC, Dulla J. Biosynthesis and potential applications of bacteriocins. Journal of Pure and Applied Microbiology. 2013;7.
149. Deng Y, Li CZ, Zhu Y, Wang P, Qi Q, Fu J, et al. ApnI, a transmembrane protein responsible for subtilomycin immunity, unveils a novel model for lantibiotic immunity. Applied and Environmental Microbiology. 2014;80:6303-15.
150. Nes IF, Diep DB, Håvarstein LS, Brurberg MB, Eijsink V, Holo H. Biosynthesis of bacteriocins in lactic acid bacteria. Antonie van Leeuwenhoek. 1996;70:113-28.
151. Novotny R, Pfoestl A, Messner P, Schäffer C. Genetic organization of chromosomal S-layer glycan biosynthesis loci of Bacillaceae. Glycoconjugate Journal. 2004;20:435-47.
152. Britton AP, van der Ende SR, van Belkum MJ, Martin-Visscher LA. The membrane topology of immunity proteins for the two-peptide bacteriocins carnobacteriocin XY, lactococcin G, and lactococcin MN shows structural diversity. Microbiology Open. 2020;9:e00957-e.
153. van der Meer JR, Polman J, Beerthuyzen MM, Siezen RJ, Kuipers OP, De Vos WM. Characterization of the Lactococcus lactis nisin A operon genes nisP, encoding a subtilisin-like serine protease involved in precursor processing, and nisR, encoding a regulatory protein involved in nisin biosynthesis. Journal of Bacteriology. 1993;175:2578.
154. Grove T, Himes P, Hwang S, Yumerefendi H, Bonanno J, Kuhlman B, et al. Structural insights into thioether bond formation in the biosynthesis of sactipeptides. Journal of the American Chemical Society. 2017;139.
155. Flühe L, Burghaus O, Wieckowski BM, Giessen TW, Linne U, Marahiel MA. Two [4Fe-4S] clusters containing radical SAM enzyme SkfB catalyze thioether bond formation during the maturation of the sporulation killing factor. Journal of the American Chemical Society. 2013;135:959-62.
156. Byers D, Gong H. Acyl carrier protein: structure-function relationships in a conserved multifunctional protein family. Biochemistry and Cell Biology 2008;85:649-62.
157. Zhu S, Hegemann JD, Fage CD, Zimmermann M, Xie X, Linne U, et al. Insights into the unique phosphorylation of the lasso peptide paeninodin. The Journal of Biological Chemistry. 2016;291:13662-78.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
177
158. Eastman AW, Weselowski B, Nathoo N, Yuan Z. Complete genome sequence of Paenibacillus polymyxa CR1, a plant growth-promoting bacterium isolated from the corn rhizosphere exhibiting potential for biocontrol, biomass degradation, and biofuel production. Genome Announcements. 2014;2(1):e01218-13.
159. Zhu S, Hegemann JD, Fage CD, Zimmermann M, Xie X, Linne U, et al. Insights into the unique phosphorylation of the lasso peptide paeninodin. Journal of Biological Chemistry. 2016;291:jbc.M116.722108.
160. Pan SJ, Rajniak J, Cheung WL, Link AJ. Construction of a single polypeptide that matures and exports the lasso peptide microcin J25. ChemBioChem. 2012;13(3):367-70.
161. Yan K, Li Y, Zirah S, Goulard C, Knappe TA, Marahiel MA, et al. Dissecting the maturation ateps of the lasso peptide microcin J25 in vitro. ChemBioChem. 2012;13:1046-52.
162. Zhu S, Fage CD, Hegemann JD, Mielcarek A, Yan D, Linne U, et al. The B1 protein guides the biosynthesis of a lasso peptide. Scientific Reports. 2016;6:35604-.
163. Watanabe K, Nagao N, Yamamoto S, Toda T, Kurosawa N. Thermobacillus composti sp. nov., a moderately thermophilic bacterium isolated from a composting reactor. International Journal of Systematic and Evolutionary Microbiology. 2007;57:1473-7.
164. Just-Baringo X, Albericio F, Álvarez M. Thiopeptide antibiotics: retrospective and recent advances. Marine Drugs. 2014;12:317-51.
165. Yu Y, Duan L, Zhang Q, Liao R, Ding Y, Pan H, et al. Nosiheptide biosynthesis featuring a unique indole side ring formation on the characteristic thiopeptide framework. ACS Chemical Biology. 2009;4:855-64.
166. van Belkum MJ, Worobo RW, Stiles ME. Double-glycine-type leader peptides direct secretion of bacteriocins by ABC transporters: colicin V secretion in Lactococcus lactis. Molecular Microbiology. 1997;23:1293-301.
167. Aucher W, Lacombe C, Héquet A, Frère J, Berjeaud J. Influence of amino acid substitutions in the leader peptide on maturation and secretion of mesentericin Y105 by Leuconostoc mesenteroides. Journal of Bacteriology. 2005;187:2218.
168. McAuliffe O, Hill C, Ross RP. Each peptide of the two-component lantibiotic lacticin 3147 requires a separate modification enzyme for activity. Microbiology. 2000;146:2147-54.
169. Tanaka-Kumazawa K, Kikuchi Y, Sano-Kokubun Y, Shintani S, Yakushiji M, Kuramitsu HK, et al. Characterization of a potential ABC-type bacteriocin exporter protein from Treponema denticola. BMC Oral Health. 2016;17:18-.
170. Azevedo AC, Bento CBP, Ruiz JC, Queiroz MV, Mantovani HC. Distribution and genetic diversity of bacteriocin gene clusters in rumen microbial genomes. Applied and Environmental Microbiology. 2015;81:7290.
171. Croucher NJ, Mostowy R, Wymant C, Turner P, Bentley SD, Fraser CM. Horizontal DNA transfer mechanisms of bacteria as weapons of intragenomic conflict. PLoS Biology. 2016;14:e1002394-e.
172. Du Y, Ma J, Yin Z, Liu K, Yao G, Xu W, et al. Comparative genomic analysis of Bacillus paralicheniformis MDJK30 with its closely related species reveals an evolutionary relationship between B. paralicheniformis and B. licheniformis. BMC Genomics. 2019;20:283.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
178
173. Aleti G, Sessitsch A, Brader G. Genome mining: Prediction of lipopeptides and polyketides from Bacillus and related Firmicutes. Computational and Structural Biotechnology Journal. 2015;13:192-203.
174. Abdel-Mawgoud A, Hassouna N. Characterization of surfactin produced by Bacillus subtilis isolate BS5. Applied Biochemistry and Biotechnology. 2008;150:289-303.
175. Chen H, Wang L, Su CX, Gong GH, Wang P, Yu ZL. Isolation and characterization of lipopeptide antibiotics produced by Bacillus subtilis. Letters in Applied Microbiology. 2008;47:180-6.
176. Lawton EM, Cotter PD, Hill C, Ross RP. Identification of a novel two-peptide lantibiotic, Haloduracin, produced by the alkaliphile Bacillus halodurans C-125. FEMS Microbiology Letters. 2007;267:64-71.
177. Awais M, Shah A, Hameed A, Hasan F. Isolation, identification and optimization of bacitracin produced by Bacillus sp. Pakistan Journal of Botany. 2007;39:1303-12.
178. Chowdhury SP, Hartmann A, Gao X, Borriss R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42 – a review. Frontiers in Microbiology. 2015;6:780.
179. Mora I, Cabrefiga J, Montesinos E. Antimicrobial peptide genes in Bacillus strains from plant environments. International Microbiology : The Official Journal of The Spanish Society for Microbiology. 2011;14:213-23.
180. Rhoads A, Au KF. PacBio sequencing and its applications. Genomics, Proteomics & Bioinformatics. 2015;13:278-89.
181. Lu H, Giordano F, Ning Z. Oxford nanopore MinION sequencing and genome assembly. Genomics, Proteomics & Bioinformatics. 2016;14(5):265-79.
182. Shibata T, Ando T. The restriction endonucleases in Bacillus amyloliquefaciens N strain. substrate specificities. Biochimica et Biophysica Acta. 1976;442:184-96.
183. Deb P, Talukdar SA, Mohsina K, Sarker PK, Sayem SMA. Production and partial characterization of extracellular amylase enzyme from Bacillus amyloliquefaciens P-001. SpringerPlus. 2013;2:154.
184. Fan B, Wang C, Song X, Ding X, Wu L, Wu H, et al. Bacillus velezensis FZB42 in 2018: the gram-positive model strain for plant growth promotion and biocontrol. Frontiers in Microbiology. 2018;9:2491.
185. Donia MS, Cimermancic P, Schulze CJ, Wieland Brown LC, Martin J, Mitreva M, et al. A systematic analysis of biosynthetic gene clusters in the human microbiome reveals a common family of antibiotics. Cell. 2014;158:1402-14.
186. Price NPJ, Rooney AP, Swezey JL, Perry E, Cohan FM. Mass spectrometric analysis of lipopeptides from Bacillus strains isolated from diverse geographical locations. FEMS Microbiology Letters. 2007;271:83-9.
187. Huot E, Barrena-Gonzalez C, Petitdemange H. Tween 80 effect on bacteriocin synthesis by Lactococcus lactis subsp. cremoris J46. Letters in Applied Microbiology 1996;22:307-10.
188. De Man JC, Rogosa M, Sharpe ME. A medium for the cultivation of Lactobacilli Journal of Applied Bacteriology. 1960;23:130-5.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
179
189. Pal A, Ramana KV, Bawa AS. Simplification and optimization of deMan Rogosa Sharpe (MRS) medium for enhanced production of bacteriocin by Weissella paramesenteroides DFR-8. Journal of Food Science and Technology. 2010;47:258-65.
190. Aunpad R, Na-Bangchang K. Pumilicin 4, a novel bacteriocin with anti-MRSA and anti-VRE activity produced by newly isolated bacteria Bacillus pumilus strain WAPB4. Current Microbiology. 2007;55:308-13.
191. Singh PK, Chittpurna, Ashish, Sharma V, Patil PB, Korpole S. Identification, purification and characterization of laterosporulin, a novel bacteriocin produced by Brevibacillus sp. strain GI-9. PLoS One. 2012;7:e31498.
192. Grady EN, MacDonald J, Liu L, Richman A, Yuan Z. Current knowledge and perspectives of Paenibacillus: a review. Microbial Cell Factories. 2016;15:203-.
193. Tang Q, Puri A, Padda KP, Chanway CP. Biological nitrogen fixation and plant growth promotion of lodgepole pine by an endophytic diazotroph Paenibacillus polymyxa and its GFP-tagged derivative. Botany. 2017;95(6):611-9.
194. Lebuhn M, Heulin T, Hartmann A. Production of auxin and other indolic and phenolic compounds by Paenibacillus polymyxa strains isolated from different proximity to plant roots. FEMS Microbiology Ecology. 1997;22:325-34.
195. Guemouri-Athmani S, Berge O, Bourrain M, Mavingui P, Thiéry JM, Bhatnagar T, et al. Diversity of Paenibacillus polymyxa populations in the rhizosphereof wheat (Triticum durum) in Algerian soils. European Journal of Soil Biology. 2000;36:149-59.
196. Alvarez VM, von der Weid I, Seldin L, Santos AL. Influence of growth conditions on the production of extracellular proteolytic enzymes in Paenibacillus peoriae NRRL BD-62 and Paenibacillus polymyxa SCE2. Letters in Applied Microbiology. 2006;43:625-30.
197. Bern M, Kil Y, Becker C. Byonic: advanced peptide and protein identification software. Current Protocols in Bioinformatics. 2012;Chapter 13:Unit13.20.
198. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clinical Microbiology Reviews. 2017;30:557.
199. Halim M. Stability of Bacteriocin-like inhibitory substance (BLIS) produced by Pediococcus acidilactici kp10 at different extreme conditions. BioMed Research International. 2018;2018:1-11.
200. Luti K, Al-Saeedi B. Bacteriocin from Streptococcus salivarius optimized statistically by response surface methodology active against different clinical oral pathogenic Streptococci. Iraqi Journal of Science. 2018;59.
201. Niu CYR, Zhang L, Yi H. Development of a chemically defined medium for better yield and purification of Enterocin Y31 from Enterococcus faecium Y31. Journal of Food Quality. 2017;2017:1-8.
202. Jangra M, Randhawa HK, Kaur M, Srivastava A, Maurya N, Patil PP, et al. Purification, characterization and in vitro evaluation of polymyxin A from Paenibacillus dendritiformis: an underexplored member of the polymyxin family. Frontiers in Microbiology. 2018;9.
203. Yu Z, Sun Z, Yin J, Qiu J. Enhanced production of polymyxin E in Paenibacillus polymyxa by replacement of glucose by starch. BioMed Research International. 2018;2018:1934309-.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
180
204. Shoji J, Kato T, Hinoo H. The structure of polymyxin S. (Studies on antibiotics from the genus Bacillus. XXI). The Journal of Antibiotics. 1977;30:1035-41.
205. Storm DR, Rosenthal KS, Swanson PE. Polymyxin and related peptide antibiotics. Annual Review of Biochemistry. 1977;46:723-63.
206. Gelband H, Miller PM, Pant S, Gandra S, Levinson J, Barter D, et al. The state of the world's antibiotics 2015. Wound Healing Southern Africa. 2015;8:30-4.
207. De Vuyst L, Vandamme EJ. Influence of the carbon source on nisin production in Lactococcus lactis subsp. lactis batch fermentations. Journal of General Microbiology. 1992;138:571-8.
208. Iyapparaj P, Maruthiah T, Ramasubburayan R, Prakash S, Kumar C, Immanuel G, et al. Optimization of bacteriocin production by Lactobacillus sp. MSU3IR against shrimp bacterial pathogens. Aquatic Biosystems. 2013;9:12.
209. Terraf MC, Juárez Tomás MS, Nader-Macías ME, Silva C. Screening of biofilm formation by beneficial vaginal lactobacilli and influence of culture media components. Journal of Applied Microbiology. 2012;113:1517-29.
210. Neysens P, Messens W, De Vuyst L. Effect of sodium chloride on growth and bacteriocin production by Lactobacillus amylovorus DCE 471. International Journal of Food Microbiology. 2003;88:29-39.
211. Malheiros PS, Sant'Anna V, Todorov SD, Franco BDGM. Optimization of growth and bacteriocin production by Lactobacillus sakei subsp. sakei2a. Brazilian Journal of Microbiology. 2015;46:825-34.
212. Parlindungan E, May BK, Jones OAH. Metabolic insights into the effects of nutrient stress on Lactobacillus plantarum B21. Frontiers in Molecular Biosciences. 2019;6.
213. Butcher BG, Helmann JD. Identification of Bacillus subtilis σW-dependent genes that provide intrinsic resistance to antimicrobial compounds produced by Bacilli. Molecular Microbiology. 2006;60:765-82.
214. Kayalvizhi N, Rameshkumar N, Gunasekaran P. Cloning and characterization of mersacidin like bacteriocin from Bacillus licheniformis MKU3 in Escherichia coli. Journal of Food Science and Technology. 2016;53:2298-306.
215. Marugg JD, Gonzalez CF, Kunka BS, Ledeboer AM, Pucci MJ, Toonen MY, et al. Cloning, expression, and nucleotide sequence of genes involved in production of pediocin PA-1, and bacteriocin from Pediococcus acidilactici PAC1.0. Applied and Environmental Microbiology. 1992;58:2360-7.
216. Song D, Li P, Gu Q. Cloning and heterologous expression of plantaricin ZJ5, a novel bacteriocin from Lactobacillus plantarum ZJ5, in Escherichia coli. Natural Product Communications. 2018;13:1934578X1801301231.
217. Kong W, Lu T. Cloning and optimization of a nisin biosynthesis pathway for bacteriocin harvest. ACS Synthetic Biology. 2014;3:439-45.
218. Canosi U, Morelli G, Trautner TA. The relationship between molecular structure and transformation efficiency of some S. aureus plasmids isolated from B. subtilis. Molecular and General Genetics. 1978;166:259-67.
219. Voss C, Schmidt T, Schleef M, Friehs K, Flaschel E. Production of supercoiled multimeric plasmid DNA for biopharmaceutical application. Journal of Biotechnology. 2003;105:205-13.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
181
220. Li L, Jiang W, Lu Y. A modified Gibson assembly method for cloning large DNA fragments with high GC Contents. Methods in Molecular Biology. 2018;1671:203-9.
221. Bizani D, Brandelli A. Characterization of a bacteriocin produced by a newly isolated Bacillus sp. Strain 8A. Journal of Applied Microbiology. 2002;93(3):512-9.
222. Perez RH, Zendo T, Sonomoto K. Circular and leaderless bacteriocins: biosynthesis, mode of action, applications, and prospects. Frontiers in microbiology. 2018;9:2085-.
223. Babasaki K, Takao T, Shimonishi Y, Kurahashi K. Subtilosin A, a new antibiotic peptide produced by Bacillus subtilis 168: isolation, structural analysis, and biogenesis. Journal of Biochemistry. 1985;98:585-603.
224. Kemperman R, Kuipers A, Karsens H, Nauta A, Kuipers O, Kok J. Identification and characterization of two Novel clostridial bacteriocins, circularin A and closticin 574. Applied and Environmental Microbiology. 2003;69:1589.
225. Larionov V, Kouprina N, Eldarov M, Perkins E, Porter G, Resnick MA. Transformation-associated recombination between diverged and homologous DNA repeats is induced by strand breaks. Yeast. 1994;10:93-104.
226. Du D, Wang L, Tian Y, Liu H, Tan H, Niu G. Genome engineering and direct cloning of antibiotic gene clusters via phage ϕBT1 integrase-mediated site-specific recombination in Streptomyces. Scientific Reports. 2015;5:8740.
227. Bian X, Huang F, Stewart FA, Xia L, Zhang Y, Müller R. Direct cloning, genetic engineering, and heterologous expression of the syringolin biosynthetic gene cluster in E. coli through Red/ET recombineering. Chembiochem. 2012;13:1946-52.
228. Telke AA, Ovchinnikov KV, Vuoristo KS, Mathiesen G, Thorstensen T, Diep DB. Over 2000-fold increased production of the leaderless bacteriocin garvicin KS by increasing gene dose and optimization of culture conditions. Frontiers in Microbiology. 2019;10.
229. van De Ven FJM, Jung G. Structures of lantibiotics studied by NMR. Antonie van Leeuwenhoek. 1996;69:99-107.
230. Acedo JZ, van Belkum MJ, Lohans CT, Towle KM, Miskolzie M, Vederas JC. Nuclear magnetic Resonance solution structures of lacticin Q and aureocin A53 reveal a structural motif conserved among leaderless bacteriocins with broad-spectrum activity. Biochemistry. 2016;55:733-42.
231. Rojo-Bezares B, Sáenz Y, Navarro L, Zarazaga M, Ruiz-Larrea F, Torres C. Coculture-inducible bacteriocin activity of Lactobacillus plantarum strain J23 isolated from grape must. Food Microbiology. 2007;24:482-91.
232. Shanker E, Federle MJ. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes. 2017:15.
233. Coisne S, Béchet M, Blondeau R. Actinorhodin production by Streptomyces coelicolor A3(2) in iron-restricted media. Letters in Applied Microbiology. 1999;28:199-202.
234. Owen GA, Pascoe B, Kallifidas D, Paget MSB. Zinc-responsive regulation of alternative ribosomal protein genes in Streptomyces coelicolor involves zur and sigmaR. Journal of Bacteriology. 2007;189:4078-86.
235. Desai RP, Leaf T, Woo E, Licari P. Enhanced production of heterologous macrolide aglycones by fed-batch cultivation of Streptomyces coelicolor. Journal of Industrial Microbiology and Biotechnology. 2002;28:297-301.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
182
236. Rønnest NP, Stocks SM, Eliasson Lantz A, Gernaey KV. Introducing process analytical technology (PAT) in filamentous cultivation process development: comparison of advanced online sensors for biomass measurement. Journal of Industrial Microbiology & Biotechnology. 2011;38(10):1679-90.
237. Chen XH, Scholz R, Borriss M, Junge H, Mögel G, Kunz S, et al. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. Journal of Biotechnology. 2009;140:38-44.

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
183
APPENDIX
General media, solutions, and buffers
Acrylamide- bisacrylamide mixture (49.5% T, 3% C): 48% (w/v) acrylamide, 1.5% (w/v)
bisacrylamide
Coomassie blue stain for SDS-PAGE: 40% (v/v) methanol, 10% (v/v) acetic acid, 0.05%
Coomassie brilliant blue R-250 in Milli-Q water
Destaining solution for SDS-PAGE: 40% (v/v) methanol and 10% (v/v) glacial acetic acid.
Electrophoresis buffers for Tricine-SDS-PAGE:
+ Cathode buffer (inside the cassette): 0.1 M Tris, 0.1 M Tricine, 0.1% SDS
+ Anode buffer (outside the cassette, inside the chamber): 0.2 M Tris-HCl, pH 8.9
Fixing solution: 20% (v/v) isopropanol; 10% (v/v) acetic acid
GES solution: 0.5% v/v Sarkosyl (GES reagent), 5 mol/l Guanidium thiocyanate, and 100mM
EDTA)
LPMB medium: 2.5 g/L yeast extract, 5 g/L peptone, 1 g/L dextrose, 0,2 g/L K2HPO4, 0.05
g/L MgSO4.7H2O, 750 mL/L aged sea water, 250 mL/L water, pH= 7.5). The aged seawater
was prepared by storing the fresh seawater in dark for 2-4 weeks, aiming to stabilize/ or
neutralize the chemicals (heavy metals, toxic compounds)
PB medium: 40 g soy peptone, 40 g dextrin 10, 1.8 g KH2PO4, 4.5 g K2HPO4, 0.3 g
MgSO4·7H2O, and 0.2 ml KellyT trace metal solution per litter was used. KellyT trace metal
solution contains 25 mg EDTA disodium salt dihydrate, 0.5 g ZnSO4·7H2O, 3.67 g
CaCl2·2H2O, 1.25 g MnCl2·4H2O, 0.25 g CoCl2·6 H2O, 0.25 g ammonium molybdate, 2.5 g
FeSO4·7H2O, and 0.1 g CuSO4·5H2O adjusted to pH 6 with NaOH, 500 mL H2O (51)
Saline water: 0.85% (w/v) sodium chloride (NaCl) in Milli-Q water.
SpC medium: 0.1 ml /L 50% glucose; 0.1 ml/L 2% MgSO4; 0.25 ml/L 1% casamino acids;
0.2 mL /L 10% nutrient broth; 0.05 ml /L 1% required amino acid*
SpT medium: 0.1 mL /L 50% glucose; 0.41mL/L 2% MgSO4; 0.1 mL /L 1% casamino acids;
0.1 ml /L 10% nutrient broth; 0.05mL/L 1% required amino acid*

ANTIBACTERIAL PEPTIDES FROM MARINE BACILLUS SPECIES
184
* Because these are defined media, cultures of auxotrophs should be supplemented with any
required nutrients (amino acids, vitamins, purines, etc.). The Bacillus subtilisBS34A growth
requires addition of filter-sterilised L-Tryptophan (1% in dH20 – should be clear solution)
made fresh on day of experiment, which may need to be shaken at 37°C for 30 min to dissolve
TE buffer (10mmol Tris-HCl; 1mmol/L EDTA, pH 8) for 30 minutes at 37°C. 500µL of GES
solution (0.5% v/v Sarkosyl (GES reagent), 5 mol/l Guanidium thiocyanate, and 100mM
EDTA)
T-base media: 2 g/L (NH4)2SO4; 14 g/L K2HPO4; 6 g/L KH2PO4; 1g/L Na3-citrate-6H2O;
dH2O to 1L
TAE buffer (50×): Tris-base (24.2% w/v), glacial acetic acid (5.17% w/v) and EDTA (1.86%
w/v) were dissolved in Milli-Q water. This was diluted to 1× with Milli-Q water before use.
Copyright © 2022 FDOKUMEN




















