
Besides adhesion: new perspectives of integrin functions in angiogenesis
Upload
independentCategory
view
1download
0
Inhibition of angiogenesis by Ab peptides
Daniel Paris1, Kirk Townsend1, Amita Quadros1, James Humphrey1, Jiazhi Sun2, Steven Brem3,Marguerite Wotoczek-Obadia3, Anthony DelleDonne1, Nikunj Patel1, Demian F. Obregon1, RobertCrescentini1, Laila Abdullah1, Domenico Coppola4, Amyn M. Rojiani4, Fiona Crawford1, Saı̈d M. Sebti2
& Michael Mullan11The Roskamp Institute, Sarasota, Florida, USA; 2Department of Interdisciplinary Oncology, Drug DiscoveryProgram, H. Lee Moffitt Cancer Center & Research Institute, Tampa, Florida, USA; 3Department of InterdisciplinaryOncology and Neurosurgery, Neuro Oncology Program, H. Lee Moffitt Cancer Center & Research Institute, Tampa,Florida, USA; 4Department of Interdisciplinary Oncology and Pathology, H. Lee Moffitt Cancer Center & ResearchInstitute, Tampa, Florida, USA
Received 20 October 2003; accepted in revised form 12 February 2004
Key words: Ab, adenocarcinoma, angiogenesis, b-amyloid, glioblastoma
Abstract
Ab peptides are naturally occurring peptides forming b-sheet aggregates that constitute an integral componentof senile plaques and vascular deposits in Alzheimer’s disease. Since several peptides adopting a b-sheetconformation have been shown to be anti-angiogenic, we investigated the effect of Ab on angiogenesis. Weshow that in vitro, Ab dose-dependently inhibits the formation of capillaries by human brain endothelial cellsplated on Matrigel and stimulates capillary degeneration at high doses. Preparations of Ab peptides containinga higher content of b-sheet structures are more potently anti-angiogenic in vitro. Ex vivo, Ab dose-dependentlyopposes angiogenesis in rat aortae and in human middle cerebral arteries. In vivo, Ab dose dependently inhibitsangiogenesis in the chick chorioallantoic membrane assay and suppresses bFGF-induced blood vessel formationin the corneal micropocket and Matrigel plug assays. Since angiogenesis is required for tumor growth, weexplored the effect of Ab on human glioblastoma (U87MG) and human lung adenocarcinoma (A549) tumors.We show that intra-tumoral injection of Ab potently inhibits the growth and vascularization of humanglioblastoma and human lung adenocarcinoma tumor xenografts in nude mice. Similarly to the intra-tumoralinjection regimen, Ab delivered intraperitoneally also suppressed the growth of human lung adenocarcinomatumor xenografts. Altogether our data show that Ab is an angiogenesis inhibitor.
Introduction
The 39–43 amino acids b-amyloid peptides (Ab),which are proteolytically derived from the amyloidprecursor protein (APP) [1] are thought to play a keyrole in the pathobiology of Alzheimer’s disease (AD)and are also known to form some deposits aroundblood vessels leading to cerebral amyloid angiopathy[2, 3]. With increasing concentration, changes in pHor the presence of chaperone proteins, Ab undergoesa conformational transition from a soluble form toaggregated fibrillary b-sheet structures [4, 5]. Recently,Anginex a designed peptide known to adopt a b-sheetconformation has been shown to be a potent inhibitorof angiogenesis [6, 7]. Several other b-sheet containing
peptides are also known to be anti-angiogenic includ-ing endostatin [8, 9], platelet factor-4 [10], TNF-a(depending on its concentration) [11] and bactericidal-permeability-increasing protein [12]. We thereforeinvestigated the potential impact of Ab on angiogen-esis using several quantitative assays. In vitro, theimpact of Ab was investigated on capillary morpho-genesis and regression assays. We analyzed theformation of microvessel outgrowths within Matrigel,from explants of rat aortae and human middlecerebral arteries in response to Ab. The impact ofAb on angiogenesis in vivo was evaluated using thechick chorioallantoic membrane (CAM) assay, the ratmodel of corneal neovascularization and the Matrigelplug assay [13]. Angiogenesis is required for thegrowth of tumors beyond a few mm3; we thereforeexplored the effect of Ab on the growth andvascularization of human glioblastoma and humanlung adenocarcinoma xenografts into nude mice.
Correspondence to: Michael Mullan, The Roskamp Institute, 2040
Whitfield Avenue, Sarasota, FL 34243, USA. Tel: +1-941-752-2949;
Fax: +1-941-752-2948; E-mail: [email protected]
Angiogenesis 7: 75–85, 2004. 75� 2004 Kluwer Academic Publishers. Printed in the Netherlands.

Materials and methods
Tube formation by human middle cerebral artery (MCA)
Human MCA endothelial cells (5 · 104) were seeded onthe top of a Matrigel layer in 1 ml of EBM (CambrexBio Science, Maryland) containing 4% fetal calf serum.The cells were incubated for 24 h in the presence orabsence of Ab1–40, Ab1–42 or scrambled Ab (VIG-KYHGMSNLVGRSFEVHQGKGAEVDAHGLFDI-EAFVDV) at purity greater than 95% (Biosource,California). Cells were treated with freshly solubilizedAb1–40 and Ab1–42 and with preparations of Ab thatwere incubated for a week at 37 �C in PBS at aconcentration of 500 lM (Ab b-sheet content wasmonitored using a thioflavin T assay as previouslydescribed [14]) at time of plating. Briefly, 100 ll solutionof Ab peptides was added to 3 lM of thioflavin T in afinal volume of 3.0 ml of PBS. Fluorescence wasmonitored at excitation 450 nm and emission 482 nmusing a Dynex MFX microplate fluorometer. Fluores-cent values were averaged after subtracting the back-ground fluorescence of 3 lM of thioflavinT. Endothelial network experiments were performed inquadruplicate and at least two to three randomly chosenfields were photographed using a 4 · objective. Capillarylength was measured using the Image Pro Plus software(Media Cybernetic, Inc., Maryland).
Capillary degeneration assay and measurementof endothelial cell apoptosis
Capillary-like structures were formed as describedabove. The initial length of capillaries was determinedand capillaries were treated for three days with freshlysolubilized Ab1–40, Ab1–42, scrambled Ab or b-sheetbreaker peptide iAb5 at a dose equivalence of Ab : iAb5of 1 : 20 as suggested in the original manuscript [15]describing the use of this peptide or a combination ofAb1–42 and iAb5 as indicated. Results were expressed asthe percentage of initial capillary length. Endothelial cellapoptosis was quantified using the Cell Death DetectionELISA (Roche Molecular Biochemical, Germany) asper the recommendations of the manufacturer.
Arterial explant assays
Artery cultures were prepared as previously described[16, 17] using thoracic aortae excised from 9-month-oldSprague Dawley rats and human MCA obtained fromthree different human cases with no known history ofcardiovascular problem or vascular dementia (age rang-ing from 69 to 82 years, post-mortem delay of 4–5 h).Immediately following the embedding of arterial ex-plants in Matrigel, arterial explants were incubated inEBM containing freshly solubilized Ab1–40, scrambledAb1–40 peptide or the selective cyclooxygenase-2 (COX-
2) inhibitor NS-398 (Calbiochem, California). Mediumwas changed every 3 days. Microvessel outgrowth areawas quantified using the Image Pro Plus software.Briefly, ring cultures were photographed using a digitalvideo camera linked to an Olympus B · 60 microscope.The outgrowth area was delineated and measured withthe Image Pro Plus software by using a strategy ofmicrovessel outgrowth detection based on difference incolor intensities between the outgrowths, the Matrigeland the artery ring. The artery rings were manuallyselected and excluded from the area of measurement andthe color intensity threshold was adjusted to selectivelymeasure the area occupied by the microvessel out-growths. Results were expressed as a percentage of thearea occupied by microvessel outgrowths in controlconditions.
Ab ELISA
Human Ab1–40 levels were determined by ELISA (Bio-source International, California) according to the rec-ommendations of the manufacturer in the plasma ofnude mice injected intraperitoneally with 50 mg/kg ofbody weight of human Ab1–40. Ab was extracted with80% formic acid as described [18]. Plasma from nudemice non-injected with human Ab was used as the blankin the ELISA. Results were expressed in pg of humanAb1–40/ml of plasma.
Chick CAM assay
This assay was performed as described previously [19,20] using methylcellulose disks impregnated withAb1–40, scrambled Ab (1, 10, 50, 150 and 250 lg) orthe vehicle alone that were implanted onto the outer2/3 of the CAM. Forty-eight hours later, vascularresponses were evaluated under an Olympus dissectingmicroscope and CAMs photographed followingperfusion with colloidal carbon and fixation withformalin.
Rat cornea assay
This assay was performed as previously described [19,20] using hydron pellets containing either bFGF(100 ng) alone, or in combination with Ab1–40 (0.1,0.5 and 1 lg), with Ab1–42 (0.1, 0.5 and 1 lg) or with1 lg of scrambled Ab peptide. Corneal vascularresponse was measured seven days post implantation:the lengths and widths of the vessels were measuredand the angiogenesis index (AI) calculated by theformula L · W¼AI. Rats were perfused with colloi-dal carbon, eyes enucleated and fixed in 10% bufferedformalin, corneas removed under an Olympus dissect-ing microscope and mounted on glass slides withCrystal Mount media.
76 D. Paris et al.

In vivo Matrigel plug angiogenesis assay
The mouse Matrigel plug assay was performed aspreviously described [21]. Nude mice (n¼ 10) wereinjected subcutaneously with 0.5 ml of Matrigelcontaining 100 ng/ml of bFGF. Mice were theninjected daily intraperitoneally (50 mg/kg/day) for aperiod of 6 days with either freshly dissolved Ab1–40(n¼ 5) or scrambled Ab (n¼ 5). (Levels of circulatingAb1–40 were evaluated by ELISA as described above).After 6 days, mice were humanely euthanitized andthe Matrigel plugs removed, fixed with 4% parafor-maldehyde/PBS and embedded in paraffin andsectioned (5 lm). Endothelial cells in the Matrigelplug sections were identified by PECAM-1 immuno-staining and total cell invasion was determined byimage analysis as previously described [21].
Tumor xenograft models
The human glioblastoma U87MG and human lungadenocarcinoma A549 cell lines were purchased fromAmerican Tissue Culture Type Collection (Manassas,Virginia) and were grown in DMEM containing 1 ·penicilline–streptomycine-fungizone and 10% fetalbovine serum at 37 �C in a humidified atmosphereof 5% CO2. Tumor cells (6 · 106) in 100 ll of PBSwere inoculated subcutaneously into both flanks of 8to 10-week-old female nude mice (Harland). Tumorvolume in mm3 was determined using the formula(length · width2)/2, where length was the longest axisand width the measurement at right angles to thelength [22]. When the tumor volumes reached approx-imately 150 mm3, animals were treated intratumorally(SC) everyday with 100 ll of PBS (vehicle group),50 mg/kg of body weight of Ab1–40 or scrambled Ab.Additionally, some animals were also treated intra-peritoneally with 100 ll of PBS (vehicle group) orwith 100 mg/kg of body weight of Ab1–40. Data wereexpressed as mean tumor volume ± S E for eachtreatment group.
Statistical analysis
Statistical analyses were performed using ANOVA withpost hoc comparisons using Scheffe’s or Bonferroni’smethod, or t-test for independent samples where appro-priate using SPSS for Windows release 10.1.
Results
Inhibition of capillary network formation by Ab
We assessed the effect of Ab on the capillary-likenetwork formation of human brain endothelial cells on
Matrigel. 5 lM of freshly solubilized Ab1–40 or Ab1–42significantly reduced the formation of capillary-likenetworks (Figures 1a, b). No difference was observed(for a given dose) between the anti-angiogenic proper-ties of freshly solubilized Ab1–40 and aged Ab1–40.However, the effect of aging of a given dose of Ab1–42was to increase its ability to prevent network formation(500 nM of aged Ab1–42 having the same potency as5 lM of freshly solubilized Ab1–42). Ab1–42 is known tobe more amyloidogenic than Ab1–40, and aging ofamyloidogenic peptides is known to increase their b-sheet content, which is also dose dependent. Weestimated the b-sheet content of the Ab preparationsemployed in the network length experiments by usingthioflavin T, a dye that specifically binds to beta-sheetstructures. We found that network length or thioflavinT measurements were both statistically dependent onAb type, dose and age by ANOVA. (For networklength, ANOVA revealed significant main effects of Abspecies (P < 0.001), Ab dose (P < 0.001) and Ab age(P < 0.001) as well as an interactive term between Abspecies and Ab dose (P < 0.04) and between Ab typeand Ab age (P < 0.001)). Not surprisingly then,network length and thioflavin T values for the Abpreparations used are highly correlated, which suggeststhat there is a biological relationship between degree ofb-sheet content and the propensity to inhibit theformation of endothelial cell networks. In fact, inpreparations of Ab showing very little b-sheet content(500 nM of freshly solubilized Ab1–40 or Ab1–42)capillary morphogenesis is enhanced. Higher concen-trations of Ab1–40 (10, 20 and 30 lM) inhibitedcapillary morphogenesis to a level similar to the5 lM concentration of Ab1–40 (data not shown).Control experiments using scrambled Ab peptide (con-centrations ranging from 100 nM to 30 lM) revealedno impairment in capillary formation (data notshown). The effect of Ab peptides was also investigatedon established capillary networks to determine whethercertain preparations could cause degeneration of pre-existing capillaries. Primary cultures of human brainendothelial cells were differentiated on Matrigel for20 h. Capillary-like structures were then incubated withdifferent concentrations of Ab1–40 for 3 days. Concen-trations of Ab1–40 ranging from 500 nM to 5 lM didnot significantly induce capillary degeneration, howeverhigher doses of Ab1–40 (10, 20 and 30 lM) promotedthe degeneration of capillary-like structures (data notshown). Scrambled Ab doses ranging from 500 nM to30 lM did not promote capillary degeneration showingthe specificity of Ab’s effects in this assay (data notshown). Similarly, 30 lM of Ab1–42 significantly in-duced capillary degeneration, which appears to beprevented by the b-sheet breaker iAb5 (an inhibitor ofAb fibrillogenesis) [15] suggesting that capillary degen-eration induced by Ab is dependent upon the forma-tion of b-sheet structures (Figures 1c, d). Capillarydegeneration was associated with apoptosis of humanbrain endothelial cells following 3 days of incubation
Ab and angiogenesis 77

with 30 lM Ab1–42 (Figure 1e). The b-sheet breakerpeptide iAb5 also partially prevented endothelial cellapoptosis triggered by 30 lM Ab1–42. Lower doses ofAb (500 nM to 5 lM) were unable to significantlypromote the apoptosis of human brain endothelial cells(data not shown) which is in agreement with previouspublished reports showing that apoptosis of cerebralendothelial cells occurs only in response to high doses
of Ab (30–50 lM) and is dependent on the b-sheetconformation adopted by the Ab peptides [23, 24].
Effect of Ab on angiogenesis using explants of rat aortaeand human cerebrovessels
To further study the effect of Ab on angiogenesis, weused the rat aorta model, which is a self-limited
Figure 1. Modulation of capillary network formation by freshly solubilized (fs) and aged forms of Ab1–40 and Ab1–42 in Matrigel reconstituted
basement membrane. (a) Human brain endothelial cells were grown without Ab (control), with 5 lM fs Ab1–40, with 5 lM fs Ab1–42, and with 5
lM aged Ab1–42. (b) Quantification of network length by image analysis. The numbers in parenthesis on the x-axis represent the number of 4 ·fields analyzed. ANOVA revealed significant main effects of Ab dose (P < 0.001) as well as significant difference between groups (P < 0.001).
Post-hoc testing showed significant difference between control and 500 nM fs Ab1–40 (P < 0.001), control and 5 lM fs Ab1–40 (P < 0.005),
control and 500 nM aged Ab1–40 (P < 0.001), control and 5 lM aged Ab1–40 (P < 0.02), control and 500 nM fs Ab1–42 (P < 0.001), control and
5 lM fs Ab1–42 (P < 0.05), control and 5 lM aged Ab1–42 (P < 0.001) but no significant difference between control and 500 nM aged Ab1–42(P¼ 0.28). Post hoc testing also revealed significant differences between 5 lM aged Ab1–40 and 5 lM aged Ab1–42 (P < 0.003). (c) Effects of Ab1–42 and the b-sheet breaker peptide (iAb5) on capillary regression. (Similar results were also observed with Ab1–40 and iAb5 (data not shown)). (d)
Quantification of capillary length by image analysis. ANOVA revealed significant main effects for Ab1–42 (P < 0.001), iAb5 (P < 0.001) as well
as an interactive term between them (P < 0.001) and significant differences between groups (P < 0.001). Post-hoc analysis reveals significant
difference between control and Ab1–42 (P < 0.001) but not between control and iAb5 (P¼ 0.66) and between control and Ab1–42+iAb5(P¼ 0.54). (e) Effect of the b-sheet breaker peptide (iAb5) on the apoptosis induced by 30 lMAb1–42 in human brain endothelial cells. Apoptosis
was quantified by measuring the amount of cytoplasmic oligonucleosomes (by ELISA) following 3 days of incubation of confluent human brain
endothelial cells with either culture medium alone (control condition), or 30 lM of Ab1–42 or a combination of iAb5 and Ab1–42. ANOVA
revealed a significant main effect of Ab (P < 0.001) as well as a significant interactive term between iAb5 and Ab (P < 0.003). Post hoc analysis
demonstrated significant differences between control and Ab (P < 0.001), Ab and iAb5+Ab (P < 0.03) and between control and iAb5 + Ab(P < 0.003) showing a partial protection of iAb5 against endothelial cell apoptosis triggered by Ab.
78 D. Paris et al.

angiogenic process, triggered by injury and regulated bywell-defined autocrine/paracrine mechanisms [16].Normally, the rat aortic endothelium exposed to athree-dimensional matrix switches to a microvascularphenotype generating branching networks of microves-sels [16, 25]. We observed that high concentrations ofAb1–40 (5 lM and above) added to the covering mediumresulted in inhibition of the vascular outgrowth from rataortic rings (Figures 2a, b). By contrast, low local
concentrations diffusing from 1 or 2 lM Ab1–40 addedto the covering medium resulted in stimulation of theformation of microvessel outgrowths from rat aorticrings. In this rat aortic ring model, scrambled Ab (doserange from 500 nM to 30 lM) added to the coveringmedium failed to affect angiogenesis indicating a pep-tide-specific effect of Ab on angiogenesis (Figure 2c).NS-398 (20 lM), a selective cyclooxygenase-2 (COX-2)inhibitor known to inhibit the angiogenic process [26]
Figure 2. (a) Effect of Ab1–40 on microvessel outgrowths from explanted rat aortic rings. (Rat aortic rings were photographed on day 5.)
(b) Quantification of microvessel outgrowths from Ab1–40 and NS-398-treated rat aortic rings. Error bars represent standard error and the
number in bracket represents the number of aortic rings analyzed. ANOVA revealed significant main effects of Ab dose (P < 0.001) as well as
significant differences between groups (P < 0.001). Post hoc testing showed significant differences between control and 1 lM Ab1–40 (P < 0.02),
between control and 2 lM Ab1–40 (P < 0.001), between control and 5 lM Ab1–40 (P < 0.001), between control and 20 lM Ab1–40 (P < 0.001)
and between control and NS-398 (P < 0.001). c) Quantification of microvessel outgrowths from scrambled Ab-treated on aortic rings. ANOVA
revealed no significant main effect of scrambled Ab doses (P¼ 0.976) suggesting that scrambled Ab does not impact microvessel outgrowth
formation in the rat aortic ring model of angiogenesis. (d) Anti-angiogenic effect of Ab1–40 on human middle cerebral artery explants. Human
middle cerebral artery rings were photographed on day 9 using a 2· objective (or a 10· objective where indicated) revealing the capillary like
structures attached to the artery wall of human cerebral middle artery explants. (e) Quantification of microvessel outgrowths in response to Ab1–40, scrambled Ab and NS-398 treatment on human middle cerebral artery rings. ANOVA revealed significant main effects of Ab dose (P < 0.001)
and NS-398 (P < 0.01). Post-hoc comparison showed a significant difference between control and 5 lMAb1–40 (P < 0.005) and between control
and NS-398 (P < 0.05), but not between control and 1 lM Ab1–40 (P ¼ 0.179), control and 1 lM scrambled Ab (P ¼ 0.783) and control and
5 lM scrambled Ab (P ¼ 0.845).
Ab and angiogenesis 79

was used as a positive control in this assay andeffectively inhibited outgrowths.We extended these findings using human MCA
explants in a similar assay. Inhibition of microvesseloutgrowth was observed when human MCA ringsembedded in Matrigel were covered with culturemedium containing 5 lM Ab1–40 for 9 days (Figures 2d,e). To assess the specificity of the Ab1–40 effect onangiogenesis, we tested the impact of a scrambled Abpeptide and found that it did not affect the formation ofoutgrowths from human middle cerebral artery rings. Inthis human cerebrovascular model of angiogenesis, NS-398 also partially inhibited the microvessel outgrowths.
Effect of Ab on angiogenesis in vivo
We examined the in vivo activity of Ab in a variety ofmodel systems used in drug discovery of inhibitors ofangiogenesis. Chick embryo chorioallantoic membrane(CAM) assays were performed for 48 h with or withoutaddition of a dose range of Ab1–40 or scrambled Abvarying from 1 to 250 lg per disk. Dose dependentinhibition of angiogenesis was observed with Ab1–40 butnot with scrambled Ab showing the specificity of theAb1–40 angiosuppressive effect in this assay (Figure 3a,b). The antiangiogenic activity of Ab was also tested inthe avascular rat cornea assay. After 7 days, both Ab1–40and Ab1–42 dose dependently inhibited neovasculariza-tion stimulated by bFGF in the rat cornea whereas thescrambled Ab peptide showed no effect (Figures 3a, c).The anti-angiogenic activity of Ab1–40 was also
investigated in the in vivo Matrigel plug assay. Matrigel
containing 100 ng/ml of bFGF was injected subcutane-ously and mice were either treated for 6 days with anintraperitoneal injection of scrambled Ab or humanAb1–40. Histologic sections of the Matrigel plugs indi-cated a significant inhibition of the angiogenic responsein the Ab1–40-treated group compared to the group ofanimals treated with scrambled Ab. In particular,endothelial cell invasion (detected by PECAM-1 immu-nostaining) and vessel formations were inhibited as aconsequence of the Ab1–40 treatment (Figure 4).
Effect of Ab on tumor growth and vascularization
Since the development of tumors beyond a few mm3
requires the formation of new blood vessels, we inves-tigated the effect of Ab1–40 on the growth and vascular-ization of human lung adenocarcinoma (A549) andhuman glioblastoma (U87MG) tumors.We implanted A549 cells subcutaneously in the right
and left flanks of nude mice and when the tumors
Figure 3. (a) Effect of Ab1–40 and scrambled Ab peptides in the chick
CAM and rat cornea models of angiogenesis. Methylcellulose disks
impregnated with different doses of Ab1–40 or scrambled Ab (ranging
from 1 to 250 lg) were applied on 6-day-old CAMs. After 48 h
incubation, CAMs were perfused with colloidal carbon, fixed and
photographed. Representative pictures of CAMs of chick embryo
incubated for 48 h with a methylcellullose disk impregnated with
vehicle alone (distilled water) (I), with 50 lg of Ab1–40 (II). Repre-
sentative flat-mount photomicrographs of rat corneas (original mag-
nification · 100) 7 days after implantation of hydron pellets. (III)
bFGF alone and (IV) bFGF and 1 lg Ab1–40. (b) Effect of Ab1–40 andscrambled Ab peptides in the chick CAM model of angiogenesis. No
difference in vascular response was discernable between control
conditions (n¼ 7), and scrambled Ab for the dose range used (1–
250 lg), 1 lg (n¼ 13) or 10 lg of Ab1–40 (n¼ 13). 22% of the CAMs
treated with 50 lg of Ab1–40 (n¼ 23) and 38% of the CAMs treated
with Ab1–40 doses ranging from 150 to 250 lg (n¼ 13) showed
inhibition of angiogenesis. Effect of Ab1–40, Ab1–42 and scrambled Abin the rat corneal model of angiogenesis. (c) Summary data of the
in vivo angiogenic response to bFGF, and bFGF plus Ab1–40, Ab1–42and scrambled Ab in the rat cornea assay. Angiogenesis indexes
are expressed as mean ± SE. ANOVA revealed significant main
effects of Ab1–40 dose (P < 0.001) and Ab1–42 dose (P < 0.003) but
no significant main effect of scrambled Ab (P¼ 0.577). Post-hoc
analysis showed significant differences between control and 1 lg of
Ab1–40 (P < 0.001), between control and 0.5 lg of Ab1–40 (P < 0.01),
between control and 1 lg of Ab1–42 (P < 0.007), between control and
0.5 lg of Ab1–42 (P < 0.05) but no significant difference between
control and scrambled Ab (P¼ 0.997).
c
80 D. Paris et al.
reached a volume of approximately 150 mm3, Ab1–40,scrambled Ab (control peptide) or vehicle alone wereinjected intra-tumorally as described in the material andmethod section. Figure 5a shows that human lungadenocarcinoma tumors from vehicle treated andscrambled Ab treated animals grew to an average
size of 235 ± 28 and 208 ± 21 mm3 respectively. Bycontrast, lung adenocarcinoma tumors from Ab1–40treated animals grew to an average size of only110 ± 18 mm3. These data show that Ab1–40 inhibitsthe growth of human lung adenocarcinoma xenograftscompared to tumors treated with the vehicle only or thescrambled Ab peptide. Tumors were then collected andprocessed for CD31 immunostaining to evaluate theeffect of Ab1–40 on tumor angiogenesis. A reduction oftumor vascularization by approximately 60% wasobserved in the Ab1–40-treated tumors compared to thetumors treated with the vehicle only or the scrambledAb peptide suggesting that Ab1–40 inhibits angiogenesisin vivo in A549 tumor xenografts (Figure 5b).We next implanted U87MG cells in the right and left
flanks of nude mice and when the tumors reached avolume of approximately 150 mm3, tumors were in-jected subcutaneously with either Ab1–40 or the samevolume of vehicle alone. Human glioblastoma tumorxenografts from vehicle treated mice grew to an averagesize of 308 ± 16 mm3 whereas glioblastoma tumorsinjected with Ab1–40 grew only to an average volume of134 ± 16 mm3 (Figure 5c). Thus, in this human glio-blastoma tumor model, Ab1–40 also appears to potentlyinhibit tumor growth. Vascularization of the tumors wasevaluated by CD31 immunostaining and revealed thatAb1–40 treatment reduces by approximately 50% thevascular density of these glioblastoma tumors suggestingthat Ab1–40 inhibits angiogenesis in vivo in U87MGtumor xenografts (Figure 5d).These anti-tumoral and anti-angiogenic effects of Ab1–
40 were observed following the direct intra-tumoralinjection of Ab. We went on to investigate whetheranother delivery route of the Ab peptide would also affecttumor growth. Nudemice were implantedwith A549 cellsand when tumors reached approximately 150 mm3 someanimals received a single intraperitoneal injection of Ab1–40, or a daily intraperitoneal injection of Ab1–40 or thevehicle alone during the duration of the treatment.Following intraperitoneal injection of Ab, a rapid diffu-sion of the peptide was observed in the blood circulationof themice (15 min after the IP injection) (Figure 5f). Thehuman Ab1–40 peptide is rapidly eliminated but remaineddetectable in the blood circulation of the animals 72 hfollowing a single intraperitoneal injection of 50 mg Ab/kg of body weight. Tumors from vehicle treated animalsgrew to an average size of 289 ± 44 mm3 whereastumors from mice treated daily with an intraperitonealinjection of Ab1–40 grew to an average size of only95 ± 18 mm3 (Figure 5e). Immunostaining of Ab-trea-ted tumors with the anti-Ab antibody 4G8 did not revealany deposits of Ab in the tumoral masses or in thevasculature of the tumors (data not shown).
Discussion
Numerous structural and functional cerebromicrovas-cular abnormalities have been identified in AD subjects,
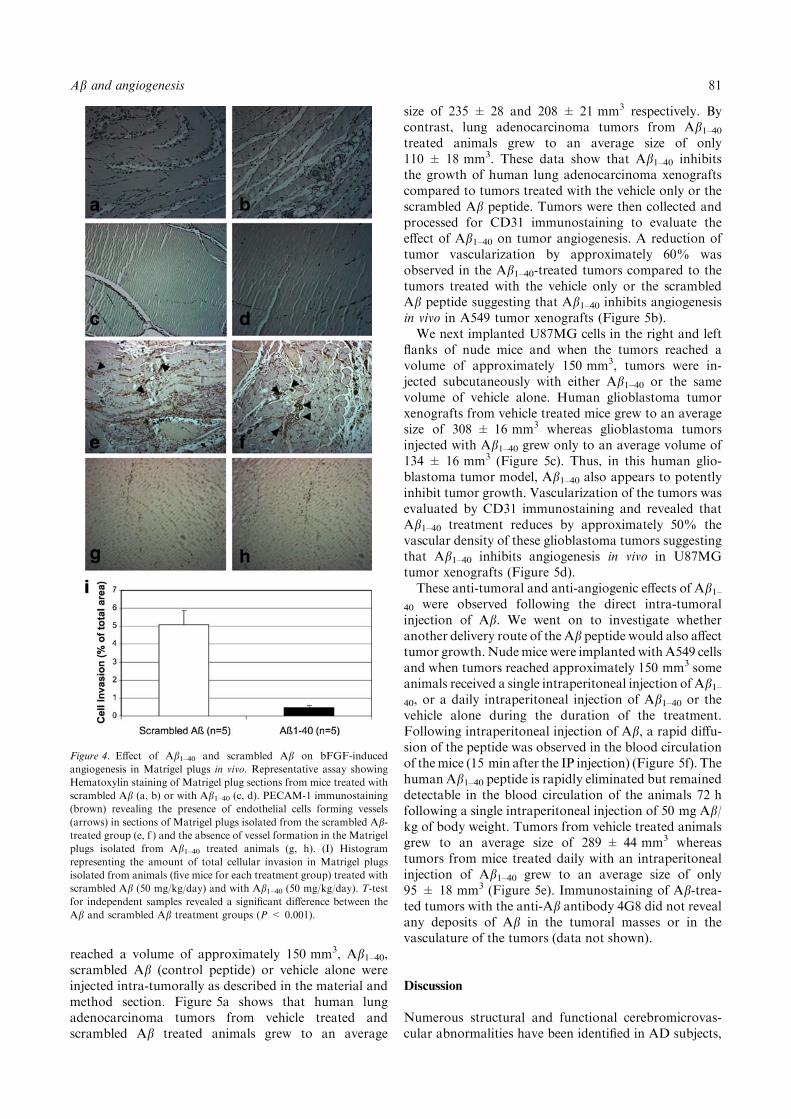
Figure 4. Effect of Ab1–40 and scrambled Ab on bFGF-induced
angiogenesis in Matrigel plugs in vivo. Representative assay showing
Hematoxylin staining of Matrigel plug sections from mice treated with
scrambled Ab (a, b) or with Ab1–40 (c, d). PECAM-1 immunostaining
(brown) revealing the presence of endothelial cells forming vessels
(arrows) in sections of Matrigel plugs isolated from the scrambled Ab-treated group (e, f ) and the absence of vessel formation in the Matrigel
plugs isolated from Ab1–40 treated animals (g, h). (I) Histogram
representing the amount of total cellular invasion in Matrigel plugs
isolated from animals (five mice for each treatment group) treated with
scrambled Ab (50 mg/kg/day) and with Ab1–40 (50 mg/kg/day). T-test
for independent samples revealed a significant difference between the
Ab and scrambled Ab treatment groups (P < 0.001).
Ab and angiogenesis 81

Figure 5. Effect of intratumoral injection of Ab1–40 on human lung adenocarcinoma xenografts in nude mice. (a) Ab1–40 suppresses human lung
(A-549) tumor growth in nude mice. A-549 cells were implanted subcutaneously in nude mice and tumor volumes were measured over time. Bars
represent standard error and data are representative of six tumors for scrambled Ab treatment and eight tumors for both control and Abtreatment conditions: (d) control group (j) scrambled Ab and (m) Ab1–40-treated group. The arrow on the graph indicates when the treatments
were initiated. ANOVA revealed significant main effect for Ab1–40 treatment (P < 0.001), time (P < 0.001) as well as an interactive term between
time and Ab1–40 treatment (P < 0.003) but no significant main effect for scrambled Ab (P¼ 0.365). Post-hoc analysis showed significant
differences between tumor volumes in the control group compared to the Ab1–40-treated group (P < 0.001) but no difference between the control
group and the scrambled Ab group (P¼ 0.735). Representative pictures depicting the vascularization of A-549 tumors detected by labeling
82 D. Paris et al.

including reduction of the cerebral blood flow, de-creased capillary diameter and microvessel density,thinning of capillary basement membrane, attenuationof capillary endothelium and cerebrovascular muscleatrophy [27–30] supporting the possibility of an alter-ation of the angiogenic process in AD brain.Interestingly, the levels of VEGF are increased in ADbrain, suggesting the existence of compensatory mech-anisms to counter insufficient or damaged vascularityapparent in AD [31]. If the brain attempts to create newcollateral channels by stimulating the production of pro-angiogenic factors, microvessel density should increasein AD brain. However, despite increased levels of pro-angiogenic factors, microvessel densities appear reducedin AD brain [32–35] suggesting a failure of a reparativeangiogenic mechanism. Microvessels undergoing angio-genesis and sprouting, normally proliferate by growth ofthe endothelium, which subsequently secretes the base-ment membrane [36]. The opposite result has beenobserved in AD, where microvessels associated with Abdeposits displayed non-functional endothelium alongwith thinning of the basement membrane [27–29]. Manynormal biological functions depend critically onangiogenesis, as do some pathologic ones such astumor growth, arthritis, and diabetic retinopathy.Anti-angiogenic approaches may be increasingly use-ful in anti-tumor therapies and the search foradditional anti-angiogenic agents continues. Theprimary sequence of several anti-angiogenic peptidesshows an abundance of hydrophobic and cationicresidues. For instance, comparison of the three-dimensional structures of endostatin [8, 9], plateletfactor-4 [10], TNF-a [11] and bactericidal-permeabil-ity-increasing protein [12] shows they are comprisedprimarily of anti-parallel b-sheets. Recently, the anti-angiogenic activity of Anginex, a designed peptide,has been shown to be dependent on its b-sheetconformation [6]. Ab mainly consists of b-sheetstructure, which prompted us to investigate the effectof Ab peptides on angiogenesis.
Our in vitro data suggest that the anti-angiogenicactivity of Ab might be related to its b-sheet content.For instance, preparations of aged Ab1–42 (which has arelatively high b-sheet content) consistently displaymore potent anti-angiogenic activity than preparationsof freshly solubilized Ab1–42. Consistent with thishypothesis is the observation that low concentrationsof Ab enhance capillary network formation. Suchconcentrations are low in b-sheet content and manyprevious observers have suggested that Ab is trophic atthese doses in several cell types [37, 38]. In addition, wehave previously shown that low doses of Ab stimulatethe phospholipase A2-MAPK-Arachidonic acid-lipoxy-genase/cyclooxygenase pathway which is known tomediate inflammatory angiogenesis [39, 40]. Again,stimulation of this pathway requires low doses of freshlysolubilized Ab and not high dose pre-aggregated formsof the peptide [41, 42].In the in vivo models of angiogenesis we employed it
is difficult to predict the exact conformation adoptedby Ab but the conditions required to form the hydronpellets and the methylcellulose disks used to deliver Ab(respectively in the corneal and the CAM assays) favorb-sheet formation since Ab has to be progressivelydried (and therefore concentrated) in these deliverysystems.It is presently unclear what the anti-angiogenic
mechanism of Ab is in these experiments. Proposedmechanisms of anti-angiogenic b-sheet peptides includeapoptotic endothelial cell destruction (for which we findno evidence for doses of Ab lower than 10 lM),inhibition of cell cycling (which we have not examinedfor Ab on endothelial cells), or disruption of celladhesion (for which we find no evidence with Ab1–40up to 5 lM, data not shown). At least in the cornealmodel of angiogenesis and in the in vivo Matrigel plugassay, Ab blocked the angiogenesis stimulated by bFGFsuggesting that Ab could interfere with the bFGF-signaling pathway but it is equally likely that Abinterferes with downstream effects of bFGF. Moreover,
Figure 5. Continued.
endothelial cells with CD31 [(I) control tumor, (II) Scrambled Ab-treated tumor and (III) Ab1–40-treated tumor]. (b) The histogram represents the
mean microvessel count per tumor. ANOVA revealed significant main effect for Ab1–40 treatment (P < 0.001) but no significant main effect for
scrambled Ab treatment (P¼ 0.314). Post-hoc analysis showed significant differences between control tumors and Ab1–40-treated tumors
(P < 0.002) but no significant difference between control tumors and tumors treated with scrambled Ab (P¼ 0.941). (c) Effect of intratumoral
injection of Ab1–40 on human glioblastoma xenografts in nude mice. Ab1–40 suppresses human glioblastoma (U87-MG) tumor growth in nude
mice. U87-MG cells were implanted subcutaneously in nude mice and tumor volumes were measured over time. Bars represent standard error and
data are representative of eight tumors for each treatment condition: (d) control group and (m) Ab1–40-treated group. The arrow on the graph
indicates when the treatments were initiated. ANOVA revealed significant main effect for Ab1–40 treatment (P < 0.001), time (P < 0.001) as well
as an interactive term between time and Ab1–40 treatment (P < 0.007). Representative pictures depicting the vascularization of U87-MG tumors
detected by labeling endothelial cells with CD31: (I) control tumor and (II) Ab1–40-treated tumor. (d) The histogram represents the mean
microvessel count per tumor. t-test for independent sample revealed significant difference (P < 0.05) between microvessel count in control and
Ab-treated tumors. (e) Effect of intraperitoneal injection of Ab1–40 on the growth of human lung adenocarcinoma tumor xenografts. Bars
represent standard error and data are representative of 16 tumors for the vehicle treatment condition (d) and 8 tumors for the group of mice
treated daily with an intraperitoneal injection of Ab1–40 (m). The arrow on the graph indicates when the treatments were initiated. ANOVA
revealed significant main effect for Ab1–40 treatment (P < 0.001) and time (P < 0.001). Post-hoc analysis showed significant differences between
tumors from the vehicle treatment group and tumors from animals receiving daily an intraperitoneal injection of Ab1–40 (P < 0.001). (f )
Histogram representing the level of human Ab1–40 detected in the plasma of nude mice at different time points following the intraperitoneal
injection of 50 mg/kg of body weight of human Ab1–40.
b
Ab and angiogenesis 83

our data showing that Ab inhibits tumor growth andvascularization further demonstrate the anti-angiogenicactivity of Ab in vivo, raising the possibility that Ab likepeptides might be beneficial in anti-tumor therapy. Ithas been observed that Cu++ enhances b-sheet foldingof Ab which is important for its toxic properties [43] andthat Cu++ is also a potent angiogenesis signal in vivo[44]. It may be that Ab itself acts as a Cu++ chelator inthese assays although much further work will berequired to test this idea.Our observation that Ab can inhibit angiogenesis
offers a new role for Ab in the vasculature, and alsosuggests a possible link between Ab and the micro-cerebrovascular abnormalities frequently observed inAD.
Acknowledgements
We extend our gratitude to Diane and Robert Roskampfor their generous support, which helped to make thiswork possible. We also thank Sandy Livingston fromthe Pathological Core at the Moffitt Cancer Researchand Robert Smeed from the Roskamp Institute forexcellent technical assistance.
References
1. Naidu A, Quon D, Cordell B. Beta-amyloid peptide produced
in vitro is degraded by proteinases released by cultured cells. J Biol
Chem 1995; 270: 1369–74.
2. Ellis RJ, Olichney JM, Thal LJ et al. Cerebral amyloid angiopathy
in the brains of patients with Alzheimer’s disease: The CERAD
experience, Part XV. Neurology 1996; 46: 1592–6.
3. Kalaria RN. The blood-brain barrier and cerebrovascular pathol-
ogy in Alzheimer’s disease. Ann N Y Acad Sci 1999; 893: 113–
25.
4. Fezoui Y, Hartley DM, Harper JD et al. An improved method of
preparing the amyloid beta-protein for fibrillogenesis and neuro-
toxicity experiments. Amyloid 2000; 7: 166–78.
5. Halverson K, Fraser PE, Kirschner DA, Lansbury Jr PT.
Molecular determinants of amyloid deposition in Alzheimer’s
disease: Conformational studies of synthetic beta-protein frag-
ments. Biochemistry 1990; 29: 2639–44.
6. Dings RP, Arroyo MM, Lockwood NA et al. Beta-sheet is the
bioactive Conformation of the anti-angiogenic anginex peptide.
Biochem J 2003; 373: 281–8.
7. Dings RP, van der Schaft DW, Hargittai B et al. Anti-tumor
activity of the novel angiogenesis inhibitor anginex. Cancer Lett
2003; 194: 55–66.
8. Hohenester E, Sasaki T, Oslen BR, Timpl R. Crystal structure of
the angiogenesis inhibitor endostatin at 1.5 A resolution. EMBO J
1998; 17: 1656–64.
9. Kranenburg O, Kroon-Batenburg LM, Reijerkerk A et al.
Recombinant endostatin forms amyloid fibrils that bind and are
cytotoxic to murine neuroblastoma cells in vitro. FEBS Lett 2003;
539: 149–55.
10. Mayo KH, Roongta V, Barker S et al. NMR solution structure of
the 32-kDa platelet factor 4 ELR-motif N-terminal chimera: A
symmetric tetramer. Biochemistry 1995; 34: 11399–409.
11. Baeyens KJ, De Bondt HL, Raeymaekers A et al. The structure of
mouse tumour-necrosis factor at 1.4 A resolution: Towards
modulation of its selectivity and trimerization. Acta Crystallogr
D Biol Crystallogr 1999; 55: 772–8.
12. Beamer LJ, Carroll SF, Eisenberg D. Crystal structure of human
BPI and two bound phospholipids at 2.4 angstrom resolution.
Science 1997; 276: 1861–4.
13. Passaniti A, Taylor RM, Pili R et al. A simple, quantitative
method for assessing angiogenesis and antiangiogenic agents using
reconstituted basement membrane, heparin, and fibroblast growth
factor. Lab Invest 1992; 4: 519–28.
14. LeVine 3rd H. Quantification of beta-sheet amyloid fibril struc-
tures with thioflavin T. Meth Enzymol 1999; 309: 274–84.
15. Soto C, Sigurdsson EM, Morelli L et al. Beta-sheet breaker
peptides inhibit fibrillogenesis in a rat brain model of amyloido-
sis: Implications for Alzheimer’s therapy. Nat Med 1998; 7:
822–6.
16. Nicosia RF, Lin YJ, Hazelton D, Qian X. Endogenous regulation
of angiogenesis in the rat aorta model. Role of vascular endothelial
growth factor. Am J Pathol 1997; 151: 1379–85.
17. Kruger EA, Duray PH, Tsokos MG et al. Endostatin inhibits
microvessel formation in the ex vivo rat aortic ring angiogenesis
assay. Biochem Biophys Res Commun 2000; 268: 183–91.
18. Kuo YM, Crawford F, Mullan M et al. Elevated Abeta and
apolipoprotein E in A betaPP transgenic mice and its relationship
to amyloid accumulation in Alzheimer’s disease. Mol Med 2000; 5:
430–9.
19. Takano S, Gately S, Jiang JB, Brem S. A diaminoantraquinone
inhibitor of angiogenesis. J Pharm Exp Therap 1994; 271: 1027–33.
20. Takano S, Gately S, Neville ME et al. Suramin, an anticancer and
angiosuppressive agent, inhibits endothelial cell binding of basic
fibroblast growth factor, migration, proliferation, and induction of
urokinase-type plasminogen activator. Cancer Res 1994; 54: 2654–
60.
21. Gyorffy S, Palmer K, Gauldie J. Adenoviral vector expressing
murine angiostatin inhibits a model of breast cancer metastatic
growth in the lungs of mice. Am J Pathol 2001; 3: 1137–47.
22. Clarke K, Lee FT, Brechbiel MW et al. In vivo biodistribution of
a humanized anti-Lewis Y monoclonal antibody (hu3S193) in
MCF-7 xenografted BALB/c nude mice. Clin Cancer Res 2000; 6:
3621–8.
23. Miravalle L, Tokuda T, Chiarle R et al. Substitutions at codon 22
of Alzheimer’s abeta peptide induce diverse conformational
changes and apoptotic effects in human cerebral endothelial cells.
J Biol Chem 2000; 35: 27110–6.
24. Eisenhauer PB, Johnson RJ, Wells JM et al. Toxicity of various
amyloid beta peptide species in cultured human blood-brain
barrier endothelial cells: Increased toxicity of dutch-type mutant.
J Neurosci Res 2000; 6: 804–10.
25. Nicosia RF, Bonanno E, Villaschi S. Large-vessel endothelium
switches to a microvascular phenotype during angiogenesis in
collagen gel culture of rat aorta. Atherosclerosis 1992; 95:
191–9.
26. Jones MK, Wang H, Peskar BM et al. Inhibition of angiogenesis
by nonsteroidal anti-inflammatory drugs: Insight into mechanisms
and implications for cancer growth and ulcer healing. Nat Med
1999; 5: 1418–23.
27. Mancardi GL, Perdelli F, Rivano C et al. Thickening of the
basement membrane of cortical capillaries in Alzheimer’s disease.
Acta Neuropathol 1980; 49: 79–83.
28. Kalaria RN, Hedera P. Differential degeneration of the cerebral
microvasculature in Alzheimer’s disease. NeuroReport 1995; 6:
477–80.
29. Kalaria RN, Pax AB. Increased collagen content of cerebral
microvessels in Alzheimer’s disease. Brain Res 1995; 705: 349–52.
30. Claudio L. Ultrastructural features of the blood-brain barrier in
biopsy tissue from Alzheimer’s disease patients. Acta Neupathol
1996; 91: 6–14.
31. Kalaria RN, Cohen DL, Premkumar DR et al. Vascular endothe-
lial growth factor in Alzheimer’s disease and experimental cerebral
ischemia. Brain Res Mol Brain Res 1998; 62: 101–5.
84 D. Paris et al.

32. Buee L, Hof PR, Delacourte A. Brain microvascular changes in
Alzheimer’s disease and other dementias. Ann N Y Acad Sci 1997;
826: 7–24.
33. Buee L, Hof PR, Bouras C et al. Pathological alterations of the
cerebral microvasculature in Alzheimer’s disease and related
dementing disorders. Acta Neuropathol (Berl) 1994; 87: 469–80.
34. Fischer VW, Siddiqi A, Yusufaly Y. Altered angioarchitecture in
selected areas of brains with Alzheimer’s disease. Acta Neuropa-
thol (Berl) 1990; 79: 672–9.
35. Suter O, Sunthorn T, Kraftsik R et al. Cerebral hypoperfusion
generates cortical watershed microinfarcts in Alzheimer disease.
Stroke 2002; 33: 1986–92.
36. Martinez-Hernandez A, Amenta PS. The basement membrane in
pathology. Lab Invest 1983; 48: 656–77.
37. Takenouchi T, Munekata E. Trophic effects of substance P and
beta-amyloid peptide on dibutyryl cyclic AMP-differentiated
human leukemic (HL-60) cells. Life Sci 1995; 56: PL479–84.
38. Atwood CS, Obrenovich ME, Liu T et al. Amyloid-beta: A
chameleon walking in two worlds: A review of the trophic and
toxic properties of amyloid-beta. Brain Res Brain Res Rev 2003;
43: 1–16.
39. Paris D, Townsend KP, Obregon DF et al. Pro-inflammatory
effect of freshly solubilized beta-amyloid peptides in the brain.
Prostaglandins Other Lipid Mediat 2002; 70: 1–12.
40. Paris D, Townsend KP, Humphrey J et al. Statins inhibit A beta-
neurotoxicity in vitro and A beta-induced vasoconstriction and
inflammation in rat aortae. Atherosclerosis 2002; 161: 293–9.
41. Paris D, Town T, Mori T et al. Soluble beta-amyloid peptides
mediate vasoactivity via activation of a pro-inflammatory path-
way. Neurobiol Aging 2000; 21: 183–97.
42. Crawford F, Soto C, Suo Z et al. Alzheimer’s beta-amyloid
vasoactivity: Identification of a novel beta-amyloid conformational
intermediate. FEBS Lett 1998; 436: 445–8.
43. Bush AI. The metallobiology of Alzheimer’s disease. Trends
Neurosci 2003; 26: 207–14.
44. Pan Q, Kleer CG, van Golen KL et al. Copper deficiency induced
by tetrathiomolybdate suppresses tumor growth and angiogenesis.
Cancer Res 2002; 62: 4854–9.
Ab and angiogenesis 85
Copyright © 2022 FDOKUMEN




















