
Bradykinin-related peptides from Phyllomedusa hypochondrialis
10
Bradykinin-related peptides from Phyllomedusa hypochondrialis G.D. Brand a,b , F.C. Krause a , L.P. Silva a , J.R.S.A. Leite a , J.A.T. Melo a , M.V. Prates a , J.B. Pesquero c , E.L. Santos c , C.R. Nakaie c , C.M. Costa-Neto d , C. Bloch Jr. a, * a Laborato ´ rio de Espectrometria de Massa, EMBRAPA, Recursos Gene ´ticos e Biotecnologia, Brası´lia, DF 70770-900, Brazil b Programa de po ´ s-graduac ¸a ˜o em Biologia Animal, IB, Universidade de Brası´lia, Brası ´lia, DF, Brazil c Departamento de Biofı ´sica, Escola Paulista de Medicina, UNIFESP, 04023-062 Sa ˜ o Paulo, Brazil d Departamento de Bioquı ´mica e Imunologia, Faculdade de Medicina de Ribeira ˜o Preto, USP, 14049-900 Ribeira ˜ o Preto, Brazil 1. Introduction Amphibian skin secretions are a rich source of bioactive molecules which cannot only act as a barrier against infectious agents but also disrupt the homeostasis of potential amphibian predators [20,26]. Several families of opioid and neurotransmitter-like peptides present in anuran skin are capable of inducing hypotension, hallucination, smooth-muscle contractions, and changes in the ingestion behavior of snakes, birds and mammals [17,19,21]. A common target of these peptides is the kallikrein–kinin system, responsible for maintaining the vascular tone and responses to tissue injury. In mammals, activation of this system ultimately leads to the secretion of endogenous pro-inflam- matory peptides such as bradykinin (BK), which exerts its effects via the activation of the BK B 2 receptor, inducing arteriolar dilation, nociceptor hypersensitivity and gastro- intestinal smooth-muscle contraction [3,10,21]. It is now well documented that anuran skin secretions contain large amounts of bradykinin related peptides (BRPs) that potentiate the effects of endogenous BK of predatory species, possibly altering their vascular and gastrointestinal functions [6– 8,15,18]. Since amphibian BRPs are molecules secreted in the skin, it has been suggested that they are in fact acting as part of a defense strategy rather than as endogenous hormones like their BK mammalian analogs [9]. Consistent with this idea is the apparent lack of a kallikrein–kinin system in the plasma of amphibians [9]. peptides 27 (2006) 2137–2146 article info Article history: Received 2 March 2006 Received in revised form 11 April 2006 Accepted 11 April 2006 Published on line 22 June 2006 Keywords: Imaging mass spectrometry Frog skin BRP Phyllomedusa abstract Bradykinin related peptides (BRPs) present in the water-soluble secretion and freshly dissected skin fragments of Phyllomedusa hypochondrialis were investigated by mass spectro- metry techniques. Eighteen BRPs, along with their post-translational modifications, were characterized in the secretion by de novo MS/MS sequencing and direct MALDI imaging experiments of the frog skin. These molecules revealed strong sequence similarities to the main plasma kinin of some mammals and reptiles. Such a diversity of molecules, within the same peptide family, belonging to a single amphibian species may be related to functional specializations of these peptides and a variety of corresponding receptors that might be present in a number of different predators. Also, a novel analog, [Val] 1 ,[Thr] 6 -bradykinyl- Gln,Ser had its biological activity positively detected in cell culture expressing the human bradykinin B 2 receptor and in guinea pig ileum preparations. # 2006 Elsevier Inc. All rights reserved. * Corresponding author at: Laborato ´ rio de Espectrometria de Massa, EMBRAPA, Recursos Gene ´ ticos e Biotecnologia, Estac ¸a ˜o Parque Biolo ´gico, Final W5, Asa Norte, Brası´lia, DF 70770-900, Brazil. Tel.: +55 61 3448 4636; fax: +55 61 3340 3658. E-mail address: [email protected] (C. Bloch Jr.). available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/peptides 0196-9781/$ – see front matter # 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.peptides.2006.04.020
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Bradykinin-related peptides from Phyllomedusa hypochondrialis

Bradykinin-related peptides from Phyllomedusahypochondrialis
G.D. Brand a,b, F.C. Krause a, L.P. Silva a, J.R.S.A. Leite a, J.A.T. Melo a, M.V. Prates a,J.B. Pesquero c, E.L. Santos c, C.R. Nakaie c, C.M. Costa-Neto d, C. Bloch Jr.a,*a Laboratorio de Espectrometria de Massa, EMBRAPA, Recursos Geneticos e Biotecnologia, Brasılia, DF 70770-900, Brazilb Programa de pos-graduacao em Biologia Animal, IB, Universidade de Brasılia, Brasılia, DF, BrazilcDepartamento de Biofısica, Escola Paulista de Medicina, UNIFESP, 04023-062 Sao Paulo, BrazildDepartamento de Bioquımica e Imunologia, Faculdade de Medicina de Ribeirao Preto, USP, 14049-900 Ribeirao Preto, Brazil
p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 6
a r t i c l e i n f o
Article history:
Received 2 March 2006
Received in revised form
11 April 2006
Accepted 11 April 2006
Published on line 22 June 2006
Keywords:
Imaging mass spectrometry
Frog skin
BRP
Phyllomedusa
a b s t r a c t
Bradykinin related peptides (BRPs) present in the water-soluble secretion and freshly
dissected skin fragments of Phyllomedusa hypochondrialis were investigated by mass spectro-
metry techniques. Eighteen BRPs, along with their post-translational modifications, were
characterized in the secretion by de novo MS/MS sequencing and direct MALDI imaging
experiments of the frog skin. These molecules revealed strong sequence similarities to the
main plasma kinin of some mammals and reptiles. Such a diversity of molecules, within the
same peptide family, belonging to a single amphibian species may be related to functional
specializations of these peptides and a variety of corresponding receptors that might be
present in a number of different predators. Also, a novel analog, [Val]1,[Thr]6-bradykinyl-
Gln,Ser had its biological activity positively detected in cell culture expressing the human
bradykinin B2 receptor and in guinea pig ileum preparations.
# 2006 Elsevier Inc. All rights reserved.
avai lab le at www.sc iencedi rect .com
journal homepage: www.elsev ier .com/ locate /pept ides
1. Introduction
Amphibian skin secretions are a rich source of bioactive
molecules which cannot only act as a barrier against
infectious agents but also disrupt the homeostasis of
potential amphibian predators [20,26]. Several families of
opioid and neurotransmitter-like peptides present in anuran
skin are capable of inducing hypotension, hallucination,
smooth-muscle contractions, and changes in the ingestion
behavior of snakes, birds and mammals [17,19,21]. A common
target of these peptides is the kallikrein–kinin system,
responsible for maintaining the vascular tone and responses
to tissue injury. In mammals, activation of this system
ultimately leads to the secretion of endogenous pro-inflam-
* Corresponding author at: Laboratorio de Espectrometria de Massa,Biologico, Final W5, Asa Norte, Brasılia, DF 70770-900, Brazil. Tel.: +55
E-mail address: [email protected] (C. Bloch Jr.).
0196-9781/$ – see front matter # 2006 Elsevier Inc. All rights reserveddoi:10.1016/j.peptides.2006.04.020
matory peptides such as bradykinin (BK), which exerts its
effects via the activation of the BK B2 receptor, inducing
arteriolar dilation, nociceptor hypersensitivity and gastro-
intestinal smooth-muscle contraction [3,10,21]. It is now well
documented that anuran skin secretions contain large
amounts of bradykinin related peptides (BRPs) that potentiate
the effects of endogenous BK of predatory species, possibly
altering their vascular and gastrointestinal functions [6–
8,15,18]. Since amphibian BRPs are molecules secreted in the
skin, it has been suggested that they are in fact acting as part
of a defense strategy rather than as endogenous hormones
like their BK mammalian analogs [9]. Consistent with this
idea is the apparent lack of a kallikrein–kinin system in the
plasma of amphibians [9].
EMBRAPA, Recursos Geneticos e Biotecnologia, Estacao Parque61 3448 4636; fax: +55 61 3340 3658.
.

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 62138
Since the first report describing the presence of BK in the
skin secretion of Rana temporaria, several structural variants of
BKs have been identified in other anuran species. The present
work describes the characterization of eighteen bradykinin
related peptides isolated from the skin secretions of the tree
frog Phyllomedusa hypochondrialis. Mass spectrometric strate-
gies were used to determine their complete primary structures
and identify their post-translational modifications (PTMs). In
addition, the peptides’ direct spatial distribution in the frog
skin was established by matrix-assisted laser desorption/
ionization mass spectrometry (MALDI-MS) imaging [4,23]. The
biological activity of one novel analog, [Val]1,[Thr]6-bradyki-
nyl-Gln,Ser, was also determined in cell cultures expressing
the human B2 receptor and in guinea pig smooth-muscle
preparations.
2. Material and methods
2.1. Amphibians
Frog skin secretions were obtained from adult specimens of P.
hypochondrialis, taken from the cerrado region of Brazil. Frogs
were captured according to the Instituto Brasileiro do Meio
Ambiente e dos Recursos Renovaveis, IBAMA, under the
license number 240/2005-CGFAU (process no. 034/06-COFAN).
2.2. Peptide purification
Frog secretion was obtained by mild electric stimulation of the
skin glands of P. hypochondrialis and collected in distilled water
as a soluble extract. The extract was filtered by gravity through
filter paper, frozen, and lyophilized (Centrivap Concentrador,
Labconco, Kansas City, MO, USA). Peptide separation was
performed by injecting the crude extract onto a semi-
preparative Vydac 218 TP 510 (Hesperia, CA, USA) reverse-
phase chromatographic column in a HPLC system (Class LC-
10VP from Shimadzu Corp., Kyoto, Japan). Peptides were
purified using a linear gradient from 0 to 70% acetonitrile
containing 0.1% trifluoroacetic acid. RP-HPLC experiments
were monitored at two different wavelengths (216 and
280 nm), and fractions were manually collected and subse-
quently lyophilized. When necessary, the isolated fractions
were submitted to further steps of purification using Vydac 218
TP 54 and GE Healthcare C2C18 columns with optimized
gradients. Sample purity and mass analyses were evaluated by
mass spectrometry.
2.3. Mass spectrometry analysis and de novo sequencing
Peptides were analyzed by MALDI-TOF/TOF on an ABI 4700
Proteomics Analyzer (Applied Biosystems, Framingham,
MA, USA), an Ultraflex II (Bruker Daltonics, Germany) and
a Q-TOF Ultima API (Micromass, UK). For the MALDI-MS
analyses, samples were mixed with a saturated matrix
solution of a-cyano-4-hydroxycinnamic acid (1:3) and
spotted on a sample plate. The MS and the MS/MS spectra
were carried out in the reflector mode with external
calibration, using the 4700 Standard kit (Applied Biosys-
tems) and Peptide Calibration Standard—Starter Kit 4
(Bruker Daltonics). Peptide de novo sequencing was per-
formed by precursor ion fragmentation using argon as
collision-induced dissociation gas in both spectrometers.
Electrospray ionization (ESI) analyses were performed in a
Q-TOF Ultima API mass spectrometer operating in W mode.
The sample dissolved in a mixture consisting of water/
methanol/acetic acid (50:50:0.1) was applied at 2 mL/min.
The capillary and cone voltages were set at 2.8 kV and 30 V,
respectively. For the MS/MS experiments, collision cell
voltage was set to 30–70 eV. CapLC (Micromass, Manchester,
UK) analysis was performed using a microbore Symmetry1
C18 5 mm (0.32 � 150) column at a constant flow rate of 1 mL/
min using a gradient spanning from 5 to 100% of reagent B in
400 min. The mobile phases were a mixture of water/
acetonitrile/trifluoroacetic acid, in the proportions of
90:10:0.1 and 10:90:0.1, for reagents A and B, respectively.
Mass detection was performed in ESI+ mode on a Q-TOF
Ultima API (Waters, UK) operating in W mode using the DDA
Survey acquisition mode.
2.4. N-terminal sequencing
Amino acid sequencing was performed by the automated
EDMAN degradation method on a PPSQ-23 protein peptide
sequencer (Shimadzu Co., Japan).
2.5. Determination of [Hyp]3-phyllokinin sulphation
[Hyp]3-phyllokinin was incubated with D2O/D-methanol/D-
acetic acid (50:50:0.1) for 10 min, which is a sufficient time for
full H/D exchange (data not shown). MS/MS experiments were
conducted using the previously described parameters for the
Q-TOF Ultima API MS analysis.
2.6. Sample preparation for imaging by massspectrometry
Dorsal frog skin was dissected and air-dried under a glass slide
for 10 min. Images of tissue pieces were acquired by light
microscopy and small pieces (1–2 mm) were directly attached
to a MALDI plate. 1 mL of matrix (a-cyano-4-hydroxycinnamic
acid, 10 mg/mL in acetonitrile/water/3% trifluoroacetic acid
5:4:1) was applied with a pipette and spread over the skin
fragment and left to dry at room temperature for 20 min prior
to imaging mass spectrometry (IMS) analyses.
2.7. Imaging by mass spectrometry
Molecular ions profiles were initially generated using a
Voyager DE STR (ABI, Framingham, MA, USA) operating in
positive mode. Automatic scanning steps were accomplished
using the MALDI-MS Imaging Tool 2.2.1 Software for the
Voyager DE STR spectrometer. The pulsed N2 laser was
rastered across the sample surface in 60 mm single steps.
Global spectra were generated for each image from 10 to 15
laser shots at each position in the global maps consisting of
600 spectra. BioMAP 3� Functional Image Analysis Software
was used to generate intensity-based ion maps of the
molecular components present in the skin surface ranging
from 1000 to 4000 Da. Similar conditions were used to perform

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 6 2139
MS/MS imaging analysis using the Ultraflex II equipped with
Smartbeam Laser technology and Flex Imaging Software.
2.8. Synthesis of [Val]1,[Thr]6-bradykinyl-Gln,Ser
Val-Hyp-Pro-Gly-Phe-Thr-Pro-Phe-Arg-Gln-Ser was synthe-
sized manually according to the standard Boc/Bzl strategy
[1,2]. Boc (Bzl)-Ser-PAM resin was used as the solid support
and the side-chain protecting groups employed were Bzl (Thr
and Ser) and Tos (Arg). After HPLC purification, the homo-
geneity of the peptide was confirmed by analytical HPLC
(Waters), mass spectrometry on RP-HPLC/MS (Micromass),
and amino acid analysis (Biochrom 20 Plus, Amersham
Biosciences).
2.9. Cell culture and transfections
Chinese hamster ovary (CHO) cells were cultivated in
Dulbecco’s modified Eagle’s medium (DMEM) supplemented
with 10% fetal calf serum and grown in a 5%-CO2 environment.
The expression plasmid containing the BK B2 receptor was
permanently transfected into CHO cells using lipofectin (Life
Technologies, Gaithersburg, MD, USA) as described by the
manufacturer. Cells containing the inserted plasmid were
selected by addition of G418 (500 mg/mL) in DMEM.
2.10. Microphysiometric analysis of the [Val]1,[Thr]6-bradykinyl-Gln,Ser interaction with BK B2 receptors
Cells expressing the B2 receptor were transferred 24 h prior
to the assays to transwells made of polycarbonate mem-
brane with 12 mm diameter and 3 mm pores (5 � 105 cells/
well). The Cytosensor microphysiometer (Molecular Devices,
Sunnyvale CA, USA) was used to measure the acidification
rate in the extracellular microenvironment due to cellular
metabolic activity [5,22]. Cells in the transwell were perfused
with DMEM, pH 7.4, containing BSA 0.1% and 44.4 mM NaCl
using a peristaltic pump, with alternating cycles of perfusion
at 100 mL/min (1 min:40 s) and pauses (20 s). During stimula-
tion the medium was replaced by DMEM containing BK in
concentrations varying from 10�9 to 10�5 M. Experiments
were done in quadruplicate, and the generated dose-
response curves were analyzed by nonlinear regression
analysis using PRISM 3.02 (Graph-Pad Software, San Diego,
CA, USA).
2.11. Smooth muscle assay
Female guinea pigs were killed by decapitation (according to
the Animal Ethics Committee protocol) and the 20-cm
terminal portion of the ileum was removed and washed in
Tyrode solution (NaCl 137 mM, KCl 2.7 mM, CaCl2 1.4 mM,
MgCl2 0.5 mM, NaHCO3 12 mM, NaH2PO4 0.4 mM and D-glucose
5.6 mM). Segments of 3 cm were cut and mounted in 5-mL
chambers containing Tyrode solution at 37 8C and bubbled
with a gas mixture of 95% O2 and 5% of CO2. After a 2-h
equilibration period isometric responses were recorded with a
force-displacement transducer (Hewlett-Packard FTA-100)
and a potentiometric recorder (ECB RB-102). Experiments
were done in duplicate, and the generated dose–response
curves were analyzed by nonlinear regression analysis using
PRISM 3.02 (Graph-Pad Software, San Diego, CA, USA).
3. Results
3.1. Purification of BKs and primary structure analyses
Samples of the crude skin secretion obtained from P.
hypochondrialis were fractionated by RP-HPLC using a semi-
preparative C18 column and on-line capillary LC/MS/MS
experiments in order to have a comprehensive identifica-
tion/sequencing strategy. Fractions containing peptides iden-
tified as BRPs are indicated in both HPLC and capillary LC
chromatograms in Fig. 1A and B, respectively. Semi-prepara-
tive RP-HPLC fractions, showing the characteristic elution
times of the peptides belonging to the BK family, were further
analyzed by mass spectrometry and fragmented using CID [6–
9,15,18]. Novel analogs were de novo sequenced by MS/MS
experiments and also had their chromatographic fractions
subjected to further purification steps up to homogeneity for
N-terminal Edman degradation and PTMs confirmation.
Sequences were categorized according to their primary
structure similarities based on the main plasma BKs analogs
of some animal groups, and are summarized in Table 1 [9]. MS/
MS spectra for all BRPs can be found as Supplemental material.
3.1.1. Mammalian BK analogsThe peptides [Thr]6-bradykinin, [Hyp]3-bradykinin, and
[Hyp]3,[Thr]6-bradykinin, with measured molecular masses
of [M + H]+ = 1074.56, 1076.63, and 1090.61 Da, respectively,
were identified in the skin extract. These were grouped, along
with the C-terminally cleaved peptide des-Arg-[Thr]6-brady-
kinin, [M + H]+ = 918.48 Da, under the mammalian BKs group.
3.1.2. [Val]1,[Thr]6-bradykinin analogs, or snake BKsThe BRP [Val]1,[Thr]6-bradykinin, [M + H]+ = 1017.54 Da, along
with its cleaved form, des-Arg-[Val]1,[Thr]6-bradykinin,
[M + H]+ = 861.48 Da, and also [Val]1,[Hyp]2,[Thr]6-bradykinin,
[M + H]+ = 1033.54 Da, were identified in the skin secretion of P.
hypochondrialis. The molecules listed above have been pre-
viously described as the main plasma BK in some viperidae
snakes and also in the skin secretion of Rana nigromaculata
[9,13]. However, this is the first report of a hydroxyproline
containing form, such as [Val]1,[Hyp]2,[Thr]6-bradykinin.
Indeed, a variety of BRPs sharing sequence similarities to
these peptides, with characteristic C-terminal extensions,
were also reported for the first time. [Val]1,[Hyp]2,[Thr]6-
bradykinyl-Gln,Ser, [M + H]+ = 1248.69 Da, [Val]1,[Thr]6-brady-
kinyl-Gln,Ser, [M + H]+ = 1232.66 Da and [Val]1,[Hyp]2,[Thr]6-
bradykinyl-Gln, [M + H]+ = 1161.65 Da. Other C-terminally
extended forms, such as [Val]1,[Thr]6-bradykinyl-Val,Asp,
[M + H]+ = 1231.69 Da, and [Val]1,[Hyp]2,[Thr]6-bradykinyl-
Val,Asp, [M + H]+ = 1247.73 were also observed.
3.1.3. PhyllokininsP. hypochondrialis also synthesizes and secretes some of the
phyllokinin variants previously identified in otherPhyllomedusid
species [11,18]. The peptides phyllokinin, [M + H]+ = 1416.70 Da,
[Hyp]3-phyllokinin, [M + H]+ = 1432.70 Da, and [Thr]6-phylloki-
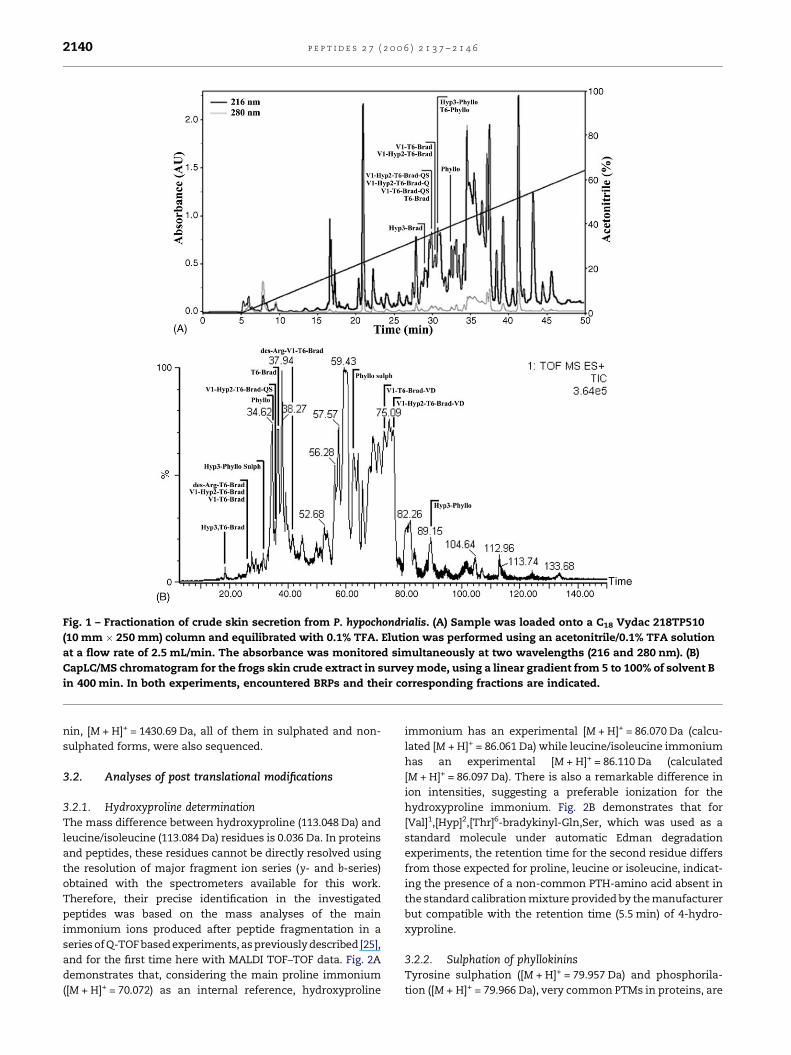
p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 62140
Fig. 1 – Fractionation of crude skin secretion from P. hypochondrialis. (A) Sample was loaded onto a C18 Vydac 218TP510
(10 mm � 250 mm) column and equilibrated with 0.1% TFA. Elution was performed using an acetonitrile/0.1% TFA solution
at a flow rate of 2.5 mL/min. The absorbance was monitored simultaneously at two wavelengths (216 and 280 nm). (B)
CapLC/MS chromatogram for the frogs skin crude extract in survey mode, using a linear gradient from 5 to 100% of solvent B
in 400 min. In both experiments, encountered BRPs and their corresponding fractions are indicated.
nin, [M + H]+ = 1430.69 Da, all of them in sulphated and non-
sulphated forms, were also sequenced.
3.2. Analyses of post translational modifications
3.2.1. Hydroxyproline determinationThe mass difference between hydroxyproline (113.048 Da) and
leucine/isoleucine (113.084 Da) residues is 0.036 Da. In proteins
and peptides, these residues cannot be directly resolved using
the resolution of major fragment ion series (y- and b-series)
obtained with the spectrometers available for this work.
Therefore, their precise identification in the investigated
peptides was based on the mass analyses of the main
immonium ions produced after peptide fragmentation in a
series of Q-TOF based experiments, as previously described [25],
and for the first time here with MALDI TOF–TOF data. Fig. 2A
demonstrates that, considering the main proline immonium
([M + H]+ = 70.072) as an internal reference, hydroxyproline
immonium has an experimental [M + H]+ = 86.070 Da (calcu-
lated [M + H]+ = 86.061 Da) while leucine/isoleucine immonium
has an experimental [M + H]+ = 86.110 Da (calculated
[M + H]+ = 86.097 Da). There is also a remarkable difference in
ion intensities, suggesting a preferable ionization for the
hydroxyproline immonium. Fig. 2B demonstrates that for
[Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser, which was used as a
standard molecule under automatic Edman degradation
experiments, the retention time for the second residue differs
from those expected for proline, leucine or isoleucine, indicat-
ing the presence of a non-common PTH-amino acid absent in
the standard calibration mixture provided by the manufacturer
but compatible with the retention time (5.5 min) of 4-hydro-
xyproline.
3.2.2. Sulphation of phyllokininsTyrosine sulphation ([M + H]+ = 79.957 Da) and phosphorila-
tion ([M + H]+ = 79.966 Da), very common PTMs in proteins, are

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 6 2141
Table 1 – BRPs in the skin secretion of P. hypochondrialis
Peptide sequence [M + H]e+ [M + H]t
+
Bradykinin analogs
[Thr]6-bradykinin R P P G F T P F R 1074.56 1074.58
des-Arg-[Thr]6-bradykinin R P P G F T P F 918.48 918.48
[Hyp]3-bradykinin R P P G F S P F R 1076.63 1076.60
[Hyp]3,[Thr]6-bradykinin R P P G F T P F R 1090.61 1090.62
[Val]1,[Thr]6 bradykinin analogs
[Val]1,[Thr]6-bradykinin V P P G F T P F R 1017.54 1017.55
des-Arg-[Val]1,[Thr]6-bradykinin V P P G F T P F 861.48 861.45
[Val]1,[Hyp]2,[Thr]6-bradykinin V P P G F T P F R 1033.54 1033.58
[Val]1,[Thr]6-bradykinyl-Gln,Ser V P P G F T P F R Q S 1232.66 1232.64
[Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln V P P G F T P F R Q 1161.65 1161.64
[Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser V P P G F T P F R Q S 1248.69 1248.67
[Val]1,[Thr]6-bradykinyl-Val,Asp V P P G F T P F R V D 1231.69 1231.65
[Val]1,[Hyp]2,[Thr]6-bradykinyl-Val,Asp V P P G F T P F R V D 1247.73 1247.68
Phyllokinins
Phyllokinin R P P G F S P F R I Y 1336.74 1336.72
Phyllokinin sulphated R P P G F S P F R I Y (HSO3) 1416.70 1416.68
[Thr]6-phyllokinin R P P G F T P F R I Y 1350.74 1350.73
[Thr]6-phyllokinin sulphated R P P G F T P F R I Y (HSO3) 1430.69 1430.70
[Hyp]3-phyllokinin R P P G F S P F R I Y 1352.74 1352.75
[Hyp]3-phyllokinin sulphated R P P G F S P F R I Y (HSO3) 1432.70 1432.71
P represents hydroxyproline residues; [M + H]e+ and [M + H]t
+ correspond to experimental and theoretical monoisotopic masses, respectively.
rather rare in peptides, existing only in some hormones [8].
Although their occurrence can be clearly identifiable by MS,
they cannot be distinguished by conventional peptide frag-
mentation, for their masses diverge by only 0.009 Da. Fig. 3
demonstrates the MS strategy used for the differentiation
between sulphation and phosphorilation of [Hyp]3-phylloki-
nin. This is based on the mass differences ([M + H]+ = 79.95 Da)
of the non-sulphated form obtained by fragmenting the
singly-charged sulphated [Hyp]3-phyllokinin, compared with
the same ions using deuterated solvents ([M + D]+ = 79.96 Da).
When a phosphorilated peptide is deuterated, mass differ-
ences arising from phosphoryl groups are incremented by
1 Da, corresponding to hydrogen/deuterium exchange, while
the sulphonyl group does not exchange hydrogen with the
solvent [14].
3.3. Detection and spatial profiling of BRPs on skin
Conventional molecular characterization methodologies,
such as chromatography and mass spectrometry, require
samples to be homogenized prior to analysis, potentially
losing morphologically and physiologically relevant informa-
tion in the process. On the other hand, immunohistochemical
and in situ hybridization methods may retain morphological-
functional aspects but do not permit large-scale analyses. IMS
of frog skin introduces the possibility of imaging molecular
ions [4,23] under almost native conditions, preserving mor-
phological and molecular information, and may represent an
important tool for the investigation of small-sized molecules
and possible PTMs. Indeed, a small dorsal skin fragment of one
specimen of P. hypochondrialis was dissected, as shown in
Fig. 4A and B, mounted onto a MALDI plate, and data were
acquired in the indicated area (Fig. 4B, green box), generating a
global spectra (Fig. 4C) of the components detected in the BRPs
mass range (m/z = 1000–1700 Da). Fig. 4C shows that this
technique is capable of detecting most of the phyllokinins
described herein, probably because these represent the most
abundant kinins in P. hypochondrialis skin, but fails with less
concentrated analogs. Also, it demonstrates that phyllokinins
are found in sulphated and in non-sulphated forms in high
yields under native conditions. The spatial profiles of BRPs
were generated to relate peptide expression to certain skin
sites and co-relate their occurrence to definite areas of the
skin. Indeed, the relation between gland morphological
position and the detection of peptides is demonstrated by
the superimposition of Fig. 4D (a light microscope image),
which constitutes an inset of the imaged area, with the
occurrence of a still unsequenced but identified ion at
m/z = 6121 Da. Therefore, this ion was used as a molecular
reference in order to get an appropriate co-register between
light microscopy and MALDI-IMS (Fig. 4E). Fig. 4F–N shows the
spatial distribution of all detected BRPs in the skin fragment. It
is worth noting that sulphated and non-sulphated phylloki-
nins have very close spatial profiles, suggesting that the same
glands synthesize these molecules.
3.4. Determination of the biological activity of[Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser
Synthetic [Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser and human
BK activated the BK B2 receptor expressed in CHO cell culture
and produced smooth-muscle contraction in preparations of
guinea pig ileum in a dose-responsive manner. Fig. 5A
demonstrates that both peptides produce similar maximum
acidification in cells expressing B2 receptors, but their
affinities showed a 10-fold difference, for BK displays an
EC50 of 10 nM, compared to 100 nM for the synthetic analog.
Comparable results were found for guinea pig ileum prepara-

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 62142
Fig. 2 – Determination of hydroxyproline. (A) Mass spectra demonstrating that leucine/isoleucine
(immoniums = [M + H]calc+ = 86.097 Da and [M + H]exp
+ = 86.109 Da) and hydroxyproline
(immoniums = [M + H]calc+ = 86.061 Da and [M + H]exp
+ = 86.070 Da) residues can be correctly assigned in BRPs by immonium
mass analysis. (B) N-terminal degradation of the three first cycles of [Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser demonstrating
that hydroxyproline2 has a different retention time from other standard residues.
tions, where the EC50 for BK is around 60 nM and 100 times
higher for the amphibian analog (Fig. 5B). These indicate that
the [Val]1,[Hyp]2,[Thr]6-bradykinyl-Gln,Ser affinity for the
human B2 receptors is residual and results from the primary
structure identity shared with mammalian BKs.
4. Discussion
The present work deals with the identification of BRPs
expressed and secreted in the skin of P. hypochondrialis,
focusing on the analyses of PTMs by MS as well as their
incidence under native conditions, as probed by IMS studies
on ions obtained directly from fresh skin segments. The
biological activity of a novel analog [Val]1,[Hyp]2,[Thr]6-
bradykinyl-Gln,Ser was positively detected on cell cultures
expressing the BK B2 receptor and guinea pig illeum prepara-
tions. However, comparisons of relative potency using BK as
the positive control revealed a weaker response of the analog,
possibly due to species-specific receptor targeting. As has
already been suggested, the diversity of the primary structures
of BRPs in frog skin secretions may reflect the pressure exerted
by natural selection for the generation of specific kinins for
each predator [6,9]. Therefore, studies on the structural
diversity of these molecules may shed light on the molecular
events that target kinins towards their specific receptors.
BRPs were arbitrarily classified according to their similarity
to the main plasma kinins of some vertebrates [9]. Molecules
sharing sequence similarity to BK, specifically peptide
sequences with an N-terminal Arg residue, have been
identified as mammalian BKs. Among these, [Thr]6-bradykinin
is present in a number of different frog skin secretions.

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 6 2143
Fig. 3 – Determination of [Hyp]3-phyllokinin sulphation. Fragmentation of the precursor ion [M + H]+ = 1432.70 Da causes
[Hyp]3-phyllokinin to generate a peak at [M + H]+ = 1352.75 Da, corresponding to the loss of 79.95 Da. Using deuterated
solvents, [Hyp]3-phyllokinin incorporates 16.23 Da, resulting in a [M + D]+ = 1448.93 Da, that generates a peak 79.96 Da apart
from the first ([M + D]+ = 1368.96 Da) upon its fragmentation. The unchanged mass difference of 79.96 Da corresponds to
sulphation, for phosphoryl groups are incremented by 1 Da upon deuteration.
Although structurally similar to BK, it may be functionally
diverse, as it is found not only in mammals but also as the
main plasma kinin in crocodilians, chelonians, and varanid
lizards [9]. Peptides with the [Val]1,[Thr]6 motif and associated
C-terminal extensions were grouped under the name of snake
BKs, due to the fact that [Val]1,[Thr]6-bradykinin is the main
plasma kinin in rattlesnakes such as Crotalus atrox, and also in
bullsnakes [9,13]. The diversity of snake BRPs found in P.
hypochondrialis points to an equal variety of predators and
probably different receptor subpopulations, but the precise
biological function of the C-terminal extensions shown herein
is an interesting issue to be evaluated. Apart from the
[Hyp]3,[Thr]6-phyllokinin, all of the phyllokinins previously
identified in other Phyllomedusids were also found in the skin
secretion of P. hypochondrialis, and similarly to other species
from the same genera, these analogs represent the most
abundant kinins in their secretions [8,11,12].
Mass spectrometry enables the determination of PTMs in
kinins in a fast and unequivocal way. It has some advantages
over other sequencing methods such as the construction of
cDNA libraries and Edman degradation and could be highly
beneficial if used in conjunction with them. Unfortunately,
although peptide fragmentation patterns are highly informa-
tive, they are often complex to interpret and to extract
meaningful data, especially when dealing with PTMs. The data
presented here demonstrate that hydroxyproline can be
successfully distinguished from leucine/isoleucine residues
in kinins in a single MS/MS experiment. However it should be
noted that peptides containing both residues in their primary
structure might present a mixed population of both immo-
niums. Proline hydroxylation has different functions in
different proteins, ranging from granting greater stability to
sensing oxygen [16]. Its high occurrence in kinins is commonly
attributed to the modulation of peptide-ligand interactions,
but this still remains to be demonstrated.
The discrimination between isobarous modifications such
as peptide phosphorylation and sulphation is also a matter of
great importance. Both modifications are able to modulate
molecular interactions, being common in proteins but less
frequent in peptides. On the other hand, whereas phosphor-
ylation is tightly linked to regulatory events, the role of
sulphation is not clear [16,24]. Amphibian skin peptides often
share substantial identity with mammalian hormones and
these can contain the same PTMs, as in CCK and caeruleins,
both of which are sulphated [6].
The incidence of PTMs in peptides is often correlated with
regulatory events, placing a demand for their identification
and spatial-profiling in situ with little sample handling.
Indeed, mixed populations of post-translationally modified
and non-modified molecules in the same biological extract
are fairly common raising questions as to whether these exist
in different populations or are the by-product of technical
artifacts. The advent of MALDI-IMS, which generates mole-
cular profiles and two-dimensional ion density maps of
peptide and protein signals directly from the surface of thin
tissue sections, has added a suitable tool, and may represent a
great innovation in the field. Indeed, Fig. 4C demonstrates
that the presence or absence of sulphation in phyllokinins is
not a storage artifact as previously stated for P. sauvageii, since
in the present study it was demonstrated that these kinins are
natively found in sulphated and non-sulphated forms
when directly ionized from the skin of P. hypochondrialis [8].
On the other hand, it does not exclude the possibility that de-
sulphation can be an artifact originating from the preparation
of the sample for mass spectrometry, e.g. matrix deposition.
Since it is highly probable that the P. hypochondrialis
phyllokinin has the same genetic structure as P. sauvageii,
consisting of a single precursor, we can state that phyllokinin
sulphation is either facultative or naturally lost in frog skins.
The only moderate correlation observed between the spatial

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 62144
Fig. 4 – MALDI-imaging analysis of P. hypochondrialis skin. (A) Adult specimen of P. hypochondrialis. (B) Light micrograph of a
small skin fragment where the imaged area is indicated. (C) Global spectra of detected ions; those identified as BRPs are
indicated. (D) Inset of the imaged area. (E) Co-localization of glands and the marker molecule (m/z = 6121 Da) used for BRPs
spatial alignment. Mass spectral images of: (F) [Hyp]3-phyllokinin, (G) [Thr]6-phyllokinin, (H) [Hyp]3-phyllokinin sulphated,
(I) [Thr]6-phyllokinin sulphated, (J) [Hyp]3-phyllokinin + [Hyp]3-phyllokinin sulphated, (K) [Thr]6-phyllokinin + [Thr]6-
phyllokinin sulphated, and (L) [Thr]6-bradykinin and (M) [Val]1,[Hyp]2,[Thr]6-bradykinyl-QS.

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 6 2145
Fig. 5 – Biological activity of [Val]1,[Hyp]2,[Thr]6-bradykinyl-
Gln,Ser and BK. (A) BK (~) and [Val]1,[Hyp]2,[Thr]6-
bradykinyl-Gln,Ser (&) dose–response acidification rate in
permanently transfected CHO cells expressing the human
B2 receptors. (B) Dose–response contractile effect of
assayed peptides in a guinea pig ileum preparation.
distribution of sulphated and non-sulphated phyllokinins
indicates that some regions either do not have the machinery
used for sulphation or that this modification is lost in
some parts of the skin due to unknown processes. Indeed,
similar hypotheses can be formulated regarding the distribu-
tion of the other BRPs in the skin, indicating the presence of
tissue specialization and/or selectivity in the secretion of
individual BRPs.
The relative potency of the peptide [Val]1,[Hyp]2,[Thr]6-
bradykinyl-Gln,Ser was in agreement with the lower activity of
Val1-substituted analogs as compared to BK in mammalian
smooth-muscle preparations, as depicted in Fig. 5B [6,21]. The
same pattern is observed in the acidification produced by the
activation of B2 receptors expressed in CHO cell cultures
(Fig. 5A). This must be due to the Arg! Val substitution, for
alanine scan studies on BK analogs have demonstrated that
Arg1, Pro2, Gly4, Phe5, Pro7, Phe8 and Arg9 are very important for
mammalian smooth-muscle activation [21]. It is currently
believed that the diversity of BRPs in the skin secretion of P.
hypochondrialis reflects the variability of predators with which
it cohabits [9]. Therefore, the resemblance of [Val]1,[-
Hyp]2,[Thr]6-bradykinyl-Gln,Ser to the main plasma kinin of
colubrid and crotalid snakes indicates that the biological
activity of this peptide would be better evaluated when
compared to [Val]1,[Thr]6-bradykinin using snake smooth-
muscle preparations.
Acknowledgments
The authors wish to thank Dr. Michael Richardson for revising
this manuscript, the Empresa Brasileira de Pesquisa Agrope-
cuaria (EMBRAPA), Recursos Geneticos e Biotecnologia, for
facilities and equipments, CNPq for scholarship funds, and
IBAMA for assistance in the capture of amphibians.
Appendix A. Supplementary data
Supplementary data associated with this article can be
found, in the online version, at doi:10.1016/j.peptides.
2006.04.020.
r e f e r e n c e s
[1] Atherton E, Sheppard RC.In: Solid phase peptide synthesis:a practical approach. Oxford: ILR Press; 1989.
[2] Barany G, Merrifield RB. The peptides: analysis. In: Gross E,Meienhofer J, editors. Synthesis and biology. New York:Academic Press; 1980.
[3] Campbell DJ. Towards understanding the kallikrein-kininsystem: insights from measurement of kinin peptides. Br JMed Biol Res 2000;33:665–77.
[4] Chaurand P, Schwartz SA, Billheimer D, Xu BJ, Crecelius A,Caprioli RM. Integrating histology and imaging massspectrometry. Anal Chem 2004;76:1145–55.
[5] Chen R, Mukhin YV, Garnovskaya MN, Thielen TE, Iijima Y,Huang C, Raymond JR, Ullian ME, Paul RV. A functionalangiotensin II receptor-GFP fusion protein: evidence foragonist-dependent nuclear translocation. Am J PhysiolRenal Physiol 2000;279:F440–8.
[6] Chen T, Orr DF, Bjourson AJ, McClean S, O’Rourke M, HirstDG, Rao P, Shaw C. Novel bradykinins and their precursorcDNAs from European yellow-bellied toad (Bombinavariegata) skin. Eur J Biochem 2002;269:4693–700.
[7] Chen T, Orr DF, Bjourson AJ, McClean S, O’Rourke M, HirstDG, Rao P, Shaw C. Bradykinins and their precursor cDNAsfrom the skin of the fire-bellied toad (Bombina orientalis).Peptides 2002;23(9):1547–55.
[8] Chen T, Shaw C. Cloning of the (Thr6)-phyllokininprecursor from Phyllomedusa sauvagei skin confirms a non-consensus tyrosine O-sulfation motif. Peptides2003;24(8):1123–30.
[9] Conlon JM. Bradykinin and its receptors in non-mammalian vertebrates. Reg Pept 1999;79:71–81.
[10] Couture R, Harrisson M, Vianna RM, Cloutier F. Kininreceptors in pain and inflammation. Eur J Pharm2001;429:161–76.
[11] Erspamer V, Melchiorri P, Falconieri Erspamer G,Montecucchi PC, de Castiglione R. Phyllomedusa skin: ahuge factory and store-house of a variety of active peptides.Peptides 1985;6(3):7–12.
[12] Erspamer V. In Heatwole H., editor. Amphibian biology, vol1. Surray Beaty & Chpping Northon; 1994. p. 178–350[Chapter 09].
[13] Galli GL, Skovgaard N, Abe AS, Taylor EW, Conlon JM,Wang T. Cardiovascular actions of rattlesnakebradykinin ([Val1, Thr6]bradykinin) in the anesthetizedSouth American rattlesnake Crotalus durissusterrificus. Am J Physiol Regul Integr Comp Physiol2005;288(2):456–65.

p e p t i d e s 2 7 ( 2 0 0 6 ) 2 1 3 7 – 2 1 4 62146
[14] Kanakubo A, Isobe M. Differentiation of sulfate andphosphate by H/D exchange mass spectrometry:application to isoflavone. J Mass Spectrom 2004;39:1260–7.
[15] Li L, Bjourson AJ, He J, Cai G, Rao P, Shaw C. Bradykininsand their cDNA from piebald odorous frog, Odorranaschmackeri, skin. Peptides 2003;24(6):863–72.
[16] Mann M, Jensen ON. Proteomic analysis of post-translational modifications. Nat Biotechnol 2003;21:255–61.
[17] McClean S, Robinson RC, Shaw C, Smyth WF.Characterisation and determination of indole alkaloids infrog-skin secretions by electrospray ionisation ion trapmass spectrometry. Rapid Commun Mass Spectrom2002;16(5):346–54.
[18] Mignogna G, Severini C, Erspamer GF, Siciliano R, Kreil G,Barra D. Tachykinins and other biologically active peptidesfrom the skin of the Costa Rican phyllomedusid frogAgalychnis callidryas. Peptides 1997;18(3):367–72.
[19] Negri L, Lattanzi R, Giannini E, De Felice M, Colucci A,Melchiorri P. Bv8, the amphibian homologue of themammalian prokineticins, modulates ingestive behaviourin rats. Br J Pharmacol 2004;142(1):181–91.
[20] Nizet V, Ohtake T, Lauth X, Trowbridge J, Rudisill J,Dorschner RA, Pestonjamasp V, Piraino J, Huttner K, Gallo
RL. Innate antimicrobial peptide protects the skin frominvasive bacterial infection. Nature 2001;414:454–7.
[21] Regoli D, Barab J. Pharmacology of bradykinin and relatedkinins. Pharmacol Rev 1980;32:1–46.
[22] Santos EL, Pesquero JB, Oliveira L, Paiva ACM, Costa-NetoCM. Mutagenesis of the AT1 receptor reveals differentbinding modes of angiotensin II and [Sar1]-angiotensin II.Regul Pept 2004;119:183–8.
[23] Schwartz SA, Reyzer ML, Caprioli RM. Direct tissue analysisusing matrix-assisted laser desorption/ionization massspectrometry: practical aspects of sample preparation. JMass Spectrom 2003;38:699–708.
[24] Steen H, Kuster B, Fernandez M, Pandey A, Mann M.Detection of tyrosine-phosphorylated peptides byprecursor ion scanning quadrupole TOF mass spectrometryin positive ion mode. Anal Chem 2001;73:1440–8.
[25] Steen H, Mann M. Analysis of bromotryptophan andhydroxyproline modifications by high-resolution, high-accuracy precursor ion scanning utilizing fragment ionswith mass-deficient mass tags. Anal Chem2002;74(24):6230–6.
[26] Zasloff M. Antimicrobial peptides of multicellularorganisms. Nature 2002;415:389–95.




















