
An Aggressive Surgical Approach Leads to Improved Survival in Patients With Gallbladder Cancer

Bradykinin B2 Receptors Mediate Contraction in the Normaland Inflamed Human Gallbladder In Vitro
MARCELLO TREVISANI,* SILVIA AMADESI,‡ FABIEN SCHMIDLIN,‡ MARIA T. POBLETE,§
ELISABETTA BARDELLA,� BARBARA MAGGIORE,* SELENA HARRISON,* CARLOS D. FIGUEROA,§
MICHELE TOGNETTO,* GIUSEPPE NAVARRA,� ALESSANDRO TURINI,� NIGEL W. BUNNETT,‡
PIERANGELO GEPPETTI,* and ROBERTO DE GIORGIO¶
*Department of Experimental Medicine and Clinical Medicine, Pharmacology Unit, University of Ferrara, Ferrara, Italy; ‡Departments ofSurgery and Physiology, University of California, San Francisco, California; §Department of Pathology, Institute of Histology and Pathology,Austral University, Isla Teja, Valdivia, Chile; �Department of Surgery, University of Ferrara, Ferrara, Italy; and ¶Department of Internal Medicineand Gastroenterology, University of Bologna, Bologna, Italy
Background & Aims: The components of the kinin sys-tem, including kinongens, kininogenases, and B2 and B1
receptors, are expressed and activated during inflamma-tion. Here, we investigated the expression of the kinin B2
receptor messenger RNA, kininogen and kallikrein im-munoreactivity, and the ability of kinins to contract con-trol and inflamed gallbladders in vitro. Methods: Humangallbladders, obtained from patients undergoing chole-cystectomy either for acute cholecystitis secondary togallstone disease or during elective gastro-entero-pancreatic surgery (controls), were processed for re-verse-transcription polymerase chain reaction analysis,kallikrein and kininogen immunohistochemistry, bindingstudies, and in vitro contractility studies. Results: Tissueexpression of B2 receptor messenger RNA and specificbinding of [3H]-bradykinin increased significantly inacute cholecystitis compared to controls. Kallikrein im-munoreactivity was detected in the epithelium and infil-trating leukocytes, whereas kininogen immunoreactivityin the lumen of blood vessels and interstitial space.Bradykinin contracted isolated strips of control andacute cholecystitis gallbladders. In acute cholecystitistissue, efficacy of bradykinin was higher than that ofcontrol gallbladders and similar to that of cholecystoki-nin. The contraction induced by bradykinin was signifi-cantly attenuated by B2 receptor antagonism but not bycyclooxygenase inhibition and B1, muscarinic, or tachy-kinin receptor antagonism. Conclusions: All the compo-nents of the kinin system are expressed in the humangallbladder. Bradykinin is a powerful spasmogen via B2
receptor activation in the normal and, especially, in theinflamed human gallbladder.
Bradykinin (BK) belongs to a family of peptides,including kallidin (Lys-BK), des-Arg9-BK and des-
Arg9-kallidin, which are formed from tissue and plasmaprecursors, the kininogens by the action of proteases,termed kallikreins.1,2 Kinins, generated locally at the site
of inflammation, produce their biological effects via 2receptor subtypes, termed B1 and B2.1–4 BK shows highaffinity for B2 and a low affinity for B1 receptors, whereasthe opposite occurs for des-Arg9-BK.2 B2 receptors areconstitutively present in a large array of cells includingsmooth muscle, endothelial, and epithelial as well asmany others. In contrast, B1 receptors are rarely ex-pressed in healthy tissues, and their expression is classi-cally promoted by inflammatory cytokines or after invitro incubation of the tissue.3,5 However, there is recentevidence that, under specific proinflammatory condi-tions, B2 receptor expression is up-regulated.6 B2 recep-tor activation evokes inflammatory responses either viadirect stimulation of effector cells or through the releaseof other mediators, including prostanoids, nitric oxide(NO), and peptide neurotransmitters.1,2,7,8
In humans, BK produces a variety of motor effects,including bronchoconstriction9,10 and contraction of uri-nary bladder.11,12 BK has been shown to exert a motorfunction in guinea pig gallbladder,13,14 where it causes acontraction that is largely dependent on prostanoid re-lease.15 However, despite the important role of BK ininflammation, no information is available regarding thepresence of the various components of the kinin systemand the effect of these peptides in the human gallbladder.
In this study, we measured the expression of B2 re-ceptor messenger RNA by reverse-transcription poly-merase chain reaction (RT-PCR) and examined kallikrein
Abbreviations used in this paper: BK, bradykinin; CCh, carbachol;CCK, cholecystokinin; Emax, maximal effect; GAPDH, glyceraldehyde3-phosphate dehydrogenase; NO, nitric oxide; pD2, negative logarithmto base 10 of the EC50; RT-PCR, reverse-transcription polymerase chainreaction.
© 2003 by the American Gastroenterological Association0016-5085/03/$30.00
doi:10.1016/S0016-5085(03)00694-2
GASTROENTEROLOGY 2003;125:126–135

and kininogen tissue expression by immunohistochem-istry in human specimens of normal gallbladder and inacute cholecystitis. We have also investigated the abilityof BK to contract human isolated gallbladder. Finally,we investigated the mechanism of the gallbladder con-tractile response to BK using B2 and B1 receptor antag-onists (HOE 14016 and R-715,17 respectively), cycloox-ygenase inhibitors (indomethacin), antimuscarinic agents(atropine), and a combination of NK1 (SR 140333), NK2
(SR 48968), and NK3 (SR 142801) tachykinins receptorantagonists.18–20
Materials and MethodsHuman Gallbladders
Human gallbladders were obtained from patients (13men, 15 women; 43–75 years) undergoing laparoscopic cho-lecystectomy for acute cholecystitis secondary to gallstonedisease. All patients complained of acute symptoms includingupper right quadrant abdominal pain, fever, nausea, and vom-iting. A thickened gallbladder wall (swollen viscus) was atypical feature observed at ultrasonography and macroscopicanalysis performed after surgery. Twenty-three control gall-bladders (12 men, 11 women; 43–76 years) were removedduring elective abdominal surgery for gastric resection, rightlobe hepatectomy, and cephalopancreatic duodenectomy(Whipple’s procedure) because of neoplastic diseases. Thesegallbladders were normal at ultrasonography, and postsurgicalmacroscopic analysis revealed apparently normal organs with-out gallstones in the lumen. Three gallbladders with macro-scopic signs of severe tissue damage (i.e., diffuse areas ofnecrosis and fibrosis), which was confirmed by microscopicexamination, were excluded from the study. All patients gavetheir informed consent before surgery, and the present studywas approved by the Ethical Committee of the University ofFerrara.
Messenger RNA and RT–PCR
Total RNA was isolated as described previously.21 Twograms of tissue were homogenized in 3 mL Trizol (Gibco,Grand Island, NY). Total RNA was extracted from the ho-mogenate as described by the manufacturer. The RNA pelletwas resuspended into water and concentrated by precipitationwith sodium acetate and ethanol.
Total RNA (1 �g) for each of the isolated tissues was reversetranscribed in cDNA at 42°C (1 hour) with the use of hex-anucleotides and the avian myeloblastoma virus reverse tran-scriptase (Promega, Madison, WI). A 525-base pair (bp) prod-uct specific to the human BK B2 gene was amplified by PCRusing specific primers (forward primer, 5-CCCTTTGTAT-GTCG TGAAGCAGAC-3; and reverse primer, 5-TTCCTG-GAGTGTTTCTTTGAGGTG-3). Cycling conditions were asfollows: 92°C for 1.5 minutes, 53°C for 2 minutes, and 72°Cfor 1.5 minutes. Cycling under these conditions was repeated30 times followed by a final extension step at 72°C for 8
minutes. As an internal standard for the efficiency of amplifi-cation, the RT product was also used to amplify glyceralde-hyde 3-phosphate dehydrogenase (GAPDH) messenger RNAsequences.
[3H]-Bradykinin–Binding Studies
Human gallbladder specimens from patients withacute cholecystitis and control gallbladders were used forbinding studies. After scraping off the mucosa, tissue speci-mens were homogenized in ice-cold buffer (50 mmol/L sodiumtrimethylamino-ethansulphonate [TES] and 1 mmol/L 1,10-phenantroline, pH 6.8) with a Polytron homogenizer (Kine-matica, Gmbh, Luzern, Switzerland). The homogenate wascentrifuged (1000 � g, 20 minutes, 4°C) to remove cellulardebris and supernatant was centrifuged (100,000 � g, 60 min,4°C) to obtain a membrane pellet. The pellet was then resus-pended in ice-cold assay buffer composed of 50 mmol/L TES,1 mmol/L 1,10-phenantroline, 140 �g/mL bacitracin, 1mmol/L dithiothreitol, 1 �mol/L captopril, and 0.1% bovineserum albumin at pH 6.8. A concentration of 160–200 �gprotein/100 �L was used for binding experiments. The proteinconcentration was determined according to a Bio-Radmethod22 using bovine albumin as reference standard.
The specific binding of [3H]-BK to plasma membranes ofhuman gallbladder was assayed in a total volume of 250 �Lcontaining the assay buffer (see composition earlier), accordingto the method previously described23,24 with minor modifica-tions. In saturation experiments, human gallbladder homoge-nates were incubated with 8 to 10 different concentrations of[3H]-BK ranging from 0.05–5 nmol/L. Nonspecific bindingwas defined as the binding obtained in the presence of 1�mol/L unlabeled BK. Incubation time was 60 minutes atroom temperature. Bound and free radioactivity were separatedby filtering the assay mixture through a Whatman GF/B glassfiber filter previously treated with 0.1% polyethyleneimine forat least 45 minutes in a Brandel cell harvester. The incubationmixture was diluted with 3 mL of ice-cold buffer (50 mmol/LTris-HCl) and then rapidly filtered under vacuum and thefilter was washed 3 times with 3 mL of buffer. The filter-boundradioactivity was counted in a Beckman LS-1800 Spectrometer(efficiency 55%).
Histochemistry and Immunohistochemistry
Gallbladder specimens from controls and acute chole-cystitis patients were washed free of bile immediately afterremoval and fixed in 10% formol saline at room temperaturefor 24–48 hours. Tissue blocks were dehydrated in a gradedseries of ethanol and embedded in paraffin wax. Sections, 5 �mthick, were mounted on glass slides previously coated withpoly-lysine (Sigma Aldrich, St. Louis, MO). For routine his-tological evaluation, tissue sections were dewaxed with xylene,rehydrated through a graded series of ethanol, and stained withH&E.
For immunohistochemistry, sections were dewaxed as pre-viously described and treated with absolute methanol and 10%hydrogen peroxide to block endogenous peroxidase activity.
July 2003 B2 RECEPTOR ACTIVATION IN HUMAN GALLBLADDER 127

After rinsing several times in 0.05 mol/L Tris-HCl buffer pH7.8, sections were incubated overnight with either antiurinarykallikrein antibodies or antihuman kininogens. Three poly-clonal antibodies raised against human urinary kallikrein weretested: 2 rabbit antisera (1:3000)25,26 and a goat antiserum(1:8000, Calbiochem, San Diego, CA). The antikininogensantibody was raised in the goat against both high– and low–molecular-weight kininogens (1:1000, Boehringer, Germany).Once incubation was completed, sections were rinsed 3 timesfor 5 minutes each in Tris-HCl buffer and incubated withbiotinylated antirabbit/goat antibody and streptavidin-perox-idase (Kit LSAB�; Dako, Carpinteria, CA) for 15 minuteseach. Peroxidase was developed for 5 minutes using a com-mercial liquid diaminobenzidine-substrate-chromogen system(Dako). All incubations were carried out at 22°C in a waterbath that was used as a moist chamber. Once immunostainingwas completed, sections were rinsed with distilled water andcounterstained with hematoxylin for 30 seconds.
Finally, sections were dehydrated in ethanol, cleared withxylene, and mounted using Canada balsam. Controls of theimmunostaining procedure included omission of the specificantibody, replacement by nonimmune rabbit or goat serum,and incubation with the specific antibody in the presence of anexcess of human urinary kallikrein (50 �g/mL), human pros-tate-specific antigen (50 �g/mL), or human kininogens (80�g/mL, Calbiochem). In addition, consecutive sections wereincubated with antiurinary kallikrein and antiprostate specificantigen (Dako, 1:1000). Sections were analyzed by using anOlympus AX 70 microscope (Olympus, Orlando, FL).
Organ Bath Studies
Forty to 50 minutes after removal, gallbladder tissuesdissected from the body of acute cholecystitis and controlgroups were carefully cleaned and strips (5–6 mm wide and1.1–1.2 cm long) containing smooth muscle and mucosa wereprepared. Tissues were placed in 5-mL organ baths containinga Krebs buffer solution ([mmol/L]: NaCl 119, NaHCO3 25,KH2PO4 1.2, MgSO4 1.5, CaCl2 2.5, KCl 4.7, and glucose 11)maintained at 37°C and oxygenated (96% O2 and 4% CO2).To maintain the physiologic osmolarity, when a high KClsolution was used, the NaCl was substituted with an identicalconcentration of KCl. For recording the mechanical activity,gallbladder strips were connected to an isometric force trans-ducer (Unirecord 7050, Ugo Basile, Comerio, Italy) withresting tension of 1.5 g. In preliminary studies, performedaccording to the guidelines indicated previously,27 we testedtensions between 0.5 and 2 g and showed that 1.5 g was theoptimal tension for KCl-induced contraction.
After an initial stabilization period (90 minutes) cumulativeconcentration-response curves were performed with high KCl(15–240 mmol/L) solution and 60 minutes later with carba-chol (CCh, 0.3–30 �mol/L). After 90-minute cumulative con-centration-response curves were performed with BK (0.1 nmol/L–3 �mol/L) or cholecystokinin (CCK, 0.1 nmol/L–1 �mol/L). Tissues were extensively washed every 15 minutes at theend of each concentration-response curve. Preliminary experi-
ments showed that the response to BK was not different intissues with or without the mucosa (data not shown). Accord-ingly, to avoid further tissue manipulation, mucosa-intacttissues were used.
In a series of experiments, after a first concentration-re-sponse curve performed with BK (0.1 nmol/L–3 �mol/L), asecond cumulative concentration-response curve was per-formed in the presence of HOE140 (1 �mol/L); R-715 (1�mol/L); indomethacin (5 �mol/L); atropine; the combinationof SR 140333, SR 48968, and SR 142801 (all 1 �mol/L); ortheir respective vehicles. All compounds were added 15 min-utes before BK with the exception of indomethacin, given 45minutes before the agonist.
Reagents
BK, atropine, captopril, TES, bacitracin, BSA, dithio-threitol, and indomethacin were obtained from Sigma (Italy).Cholecystokinin was obtained from Bachem (Switzerland).HOE140 and R-715 were kind gifts of Dr. B. Schoelkens(Aventis, Frankfurt, Gemany) and Dr. D. Regoli (University ofSherbrooke, Sherbrooke, Canada). [3H]-BK (specific activity76 Ci mmol�1) was from Amersham (UK). SR 140333, SR48968, and SR 142801 were kindly donated by Dr. X.Emonds-Alt (Sanofi-Synthlabo, Montpellier, France). Agentswere dissolved in Krebs buffer, except for indomethacin, SR140333, SR48968, and SR142801, which at stock concentra-tions (1 mmol/L) were dissolved in 100% dimethyl sulfoxide.
Statistical Analysis
All data are expressed as mean � standard error of themean. Variability of responses to the different spasmogens wasminor. This allowed the expression of data as absolute values(mg of tension). Results from in vitro studies are expressed asabsolute values (mg). The maximum effect (Emax) and thepotency of the agonist, expressed as the negative logarithm tobase 10 of the molar concentration of agonist that produceshalf-maximum response (pD2), were obtained with an iterativecurve-fitting package (Origin Software, Microcal Software,Northampton, MA). Statistical analysis was performed bymeans of Student t test or the analysis of variance and Bonfer-roni’s test when required. If P � 0.05 the results were con-sidered significant.
ResultsHistological Analysis
Histopathological analysis on normal gallbladderand acute cholecystitis was performed by 2 expert pa-thologists in a blind manner. Microscopic examinationon control gallbladders showed a mucosa that was ar-ranged in closely set and intersecting folds with a neg-ligible infiltrate of inflammatory cells. In contrast, gall-bladders of patients with acute cholecystitis showedfeatures characterized by wall thickening with a denseacute inflammatory cell infiltration, mainly characterized
128 TREVISANI ET AL. GASTROENTEROLOGY Vol. 125, No. 1
by polymorphonuclear cells, granulomas with cholesterolclefts, and mucosal outpouchings. Gallbladders (n � 3)showing foci of purulent bile along with areas of ulcer-ation, necrosis, thick fibrosis, and shallow and scarredmucous membranes were discarded from the presentstudy.
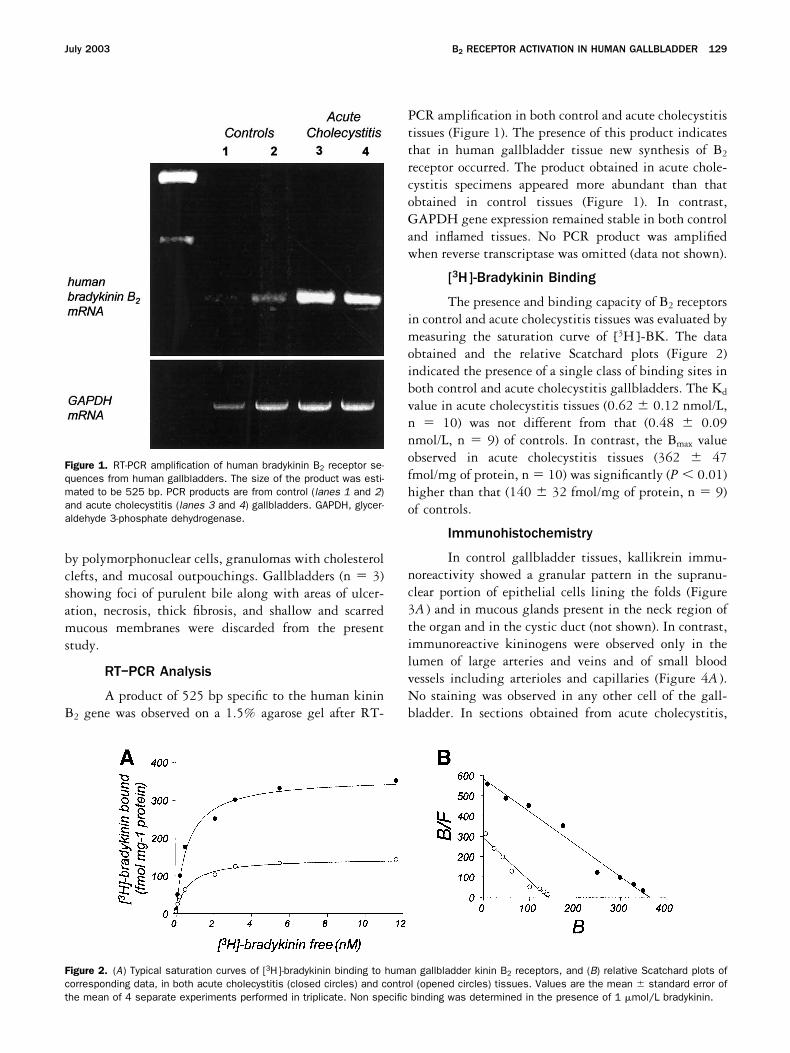
RT–PCR Analysis
A product of 525 bp specific to the human kininB2 gene was observed on a 1.5% agarose gel after RT-
PCR amplification in both control and acute cholecystitistissues (Figure 1). The presence of this product indicatesthat in human gallbladder tissue new synthesis of B2
receptor occurred. The product obtained in acute chole-cystitis specimens appeared more abundant than thatobtained in control tissues (Figure 1). In contrast,GAPDH gene expression remained stable in both controland inflamed tissues. No PCR product was amplifiedwhen reverse transcriptase was omitted (data not shown).
[3H]-Bradykinin Binding
The presence and binding capacity of B2 receptorsin control and acute cholecystitis tissues was evaluated bymeasuring the saturation curve of [3H]-BK. The dataobtained and the relative Scatchard plots (Figure 2)indicated the presence of a single class of binding sites inboth control and acute cholecystitis gallbladders. The Kd
value in acute cholecystitis tissues (0.62 � 0.12 nmol/L,n � 10) was not different from that (0.48 � 0.09nmol/L, n � 9) of controls. In contrast, the Bmax valueobserved in acute cholecystitis tissues (362 � 47fmol/mg of protein, n � 10) was significantly (P � 0.01)higher than that (140 � 32 fmol/mg of protein, n � 9)of controls.
Immunohistochemistry
In control gallbladder tissues, kallikrein immu-noreactivity showed a granular pattern in the supranu-clear portion of epithelial cells lining the folds (Figure3A ) and in mucous glands present in the neck region ofthe organ and in the cystic duct (not shown). In contrast,immunoreactive kininogens were observed only in thelumen of large arteries and veins and of small bloodvessels including arterioles and capillaries (Figure 4A ).No staining was observed in any other cell of the gall-bladder. In sections obtained from acute cholecystitis,
Figure 1. RT-PCR amplification of human bradykinin B2 receptor se-quences from human gallbladders. The size of the product was esti-mated to be 525 bp. PCR products are from control (lanes 1 and 2)and acute cholecystitis (lanes 3 and 4) gallbladders. GAPDH, glycer-aldehyde 3-phosphate dehydrogenase.
Figure 2. (A) Typical saturation curves of [3H]-bradykinin binding to human gallbladder kinin B2 receptors, and (B) relative Scatchard plots ofcorresponding data, in both acute cholecystitis (closed circles) and control (opened circles) tissues. Values are the mean � standard error ofthe mean of 4 separate experiments performed in triplicate. Non specific binding was determined in the presence of 1 �mol/L bradykinin.
July 2003 B2 RECEPTOR ACTIVATION IN HUMAN GALLBLADDER 129

kallikrein immunoreactivity appeared reduced in epithe-lial mucosal cells (Figures 3B and C ). In contrast, anintense kallikrein immunoreactivity was observed ininfiltrating polymorphonuclear leukocytes dispersedthroughout the inflamed mucosa (Figures 3C and D). Asignificant degree of human kininogen immunoreactivitywas seen in the lamina propria and in the interstitialspace between epithelial cells of inflamed gallbladders(Figures 4B and C ).
Control experiments showed no kallikrein immunore-activity when the specific antibody was replaced by non-immune serum or when the sections were incubated withthe antibody in the presence of an excess of humanurinary kallikrein (Figure 3E ) or with an antibody raisedagainst prostate-specific antigen (Figure 3F ), a closelyrelated member of the human kallikrein family. On theother hand, the immunostaining pattern was not alteredwhen the antikallikrein antibody was used in the pres-
ence of an excess of purified human prostate antigen (datanot shown). No immunostaining was detected when theantikininogens serum was replaced by nonimmune se-rum or the antiserum was used in the presence of anexcess of purified human kininogens (Figure 4D).
Bradykinin Contracts Strips of HumanGallbladder
KCl (15–240 mmol/L) produced a concentration-dependent contractile response in strips of all gallblad-ders (Figures 5 and 6) with the exception of 2 samples.Specimens from these 2 gallbladders were excluded fromthe study. Emax and pD2 for KCl are reported in Table 1.CCh (1 nmol/L–30 �mol/L) caused a concentration-dependent increase in tone in all tissues that respondedto KCl (Figures 5 and 6) with an Emax that was about36% of that of KCl. Emax and pD2 for CCh are alsoreported in Table 1.
BK caused a sustained increase in tone in strips ofhuman gallbladder in all tissues that responded to KCl.Threshold concentration to cause a visible contractionwas 1–10 nmol/L (Figures 5 and 6). In nonprecontractedstrips (n � 4) or in strips precontracted with CCh (10�mol/L, n � 4) or KCl (120 mmol/L, n � 4), BK failedto cause any measurable relaxation, whereas at the high-est concentrations it caused a contraction superimposedto the KCl-induced contraction (data not shown). BKwas found to be more potent (pD2 was more than 1
Figure 4. Kininogen immunoreactivity in acute cholecystitis (B–D) orcontrol gallbladder (A) groups. Sections were immunostained usingan antibody that recognized both high– and low–molecular-weightkininogens. (D) In control experiments, the antikininogen antibodywas used in the presence of an excess of the purified human antigen.E, surface epithelium; A, artery; C, capillary. Magnifications: A, B, andD 400� and C 1500�.
Figure 3. Kallikrein immunoreactivity in acute cholecystitis (B–F ) or(A) control gallbladder groups. (A–E) Sections were immunostainedusing an antibody raised against human urinary kallikrein or (F ) pros-tate-specific antigen. (E) In control experiments, the kallikrein anti-body was used in the presence of an excess of purified human urinarykallikrein. E, surface epithelium; PMN, infiltrating polymorphonuclearleukocytes. Magnifications: A and B 160�; C, E, and F 400�; and D1000�.
130 TREVISANI ET AL. GASTROENTEROLOGY Vol. 125, No. 1

logarithmic unit higher) and efficacious (Emax was morethan 3-fold higher) than CCh in contracting strips ofhuman gallbladder (Figure 6 and Table 1). We alsocompared the contraction produced by BK with thatinduced by CCK. Threshold concentration of CCK tocause a visible contraction (0.01–0.03 nmol/L) was lowerthan that of BK and pD2 to BK was slightly lower (lessthan 1 log unit) than CCK. However, in control gall-bladders, Emax of BK was less than half of that of CCK,whereas, in acute cholecystitis gallbladders, Emax of BKand CCK were similar (Figure 6 and Table 1).
In a specific set of experiments, the B2 receptor antag-onist, HOE 140 (1 �mol/L); the B1 receptor antagonist,R-715 (1 �mol/L); the combination of the tachykininNK1, NK2, and NK3 receptor antagonists; SR 140333,SR 48968, and SR 142801 (all 1 �mol/L); atropine (1�mol/L); and indomethacin (5 �mol/L) did not affectbaseline tone. HOE 140 shifted significantly to the rightthe concentration response curve to BK (Figure 7),whereas R-715 was without effect. Atropine (not shown),indomethacin (5 �mol/L), and the combination of the
tachykinin receptor antagonists (all 1 �mol/L) did notaffect the concentration response curve to BK (Figure 7).
DiscussionIn the present study, we showed that BK pro-
duces a robust contraction of the human gallbladder invitro. One major finding of our work is that BK showeda potency higher than that of CCh in contracting thehuman gallbladder. More surprisingly, the efficacy of BKto contract the human gallbladder was also much higher(by at least 350%) than that of CCh. Furthermore, ourresults confirmed that another peptide, CCK, was able tocontract the gallbladder tissue with high potency andefficacy. These observations suggest that BK, because ithas been proposed for CCK, is a good candidate as amajor spasmogen of the mammalian and, specifically,human gallbladder.12,28–30
Figure 5. Typical tracings of the contractile response to bradykinin,carbachol, KCl, and cholecystokinin in isolated strips of human gall-bladders from control patients and patients with acute cholecystitis.
Figure 6. Concentration-response curves to bradykinin (E), cholecys-tokinin (● ), carbachol (■ ), and KCl (�) in isolated strips of humangallbladder in vitro obtained from patients undergoing cholecystec-tomy for elective abdominal surgery (control, A) or for (B) acutecholecystitis. Values are mean � standard error of the mean of atleast 6 experiments. P � 0.05 versus controls.
July 2003 B2 RECEPTOR ACTIVATION IN HUMAN GALLBLADDER 131

A second major finding of the present study was thedifference in the contractile response to BK in gallblad-ders with acute cholecystitis versus the response seen intissues taken from control gallbladders: the efficacy ofBK in causing contraction of gallbladder tissue wasmarkedly increased in preparations obtained from acutecholecystitis patients. Notably, the Emax to BK in controlgallbladders was still higher (178%) than that of CCh inacute cholecystitis. The presence of acute cholecystitisdid not appear to affect the contractile ability of thegallbladder tissue because the response to the unspecificspasmogen KCl was unaffected. Similarly, the presence ofacute cholecystitis did not affect the potency of BK tocontract the human gallbladder receptor as estimated by
the pD2 value. However, the already robust contractionto BK seen in control gallbladders was more than double(showing efficacy similar to that of CCK) in specimenstaken from acute cholecystitis gallbladders. Either sim-ilar or decreased responses to spasmogens, includingCCK, have been reported to occur in a subgroup ofgallbladders, defined as hyporesponsive to CCK. Thereason of the reduced responsiveness is not clear, al-though severe inflammation, including histological fea-tures of purulent foci, areas of ulceration or necrosis, andshallow and scarred mucous membranes have been asso-ciated with subcontractor gallbladders. This type of gall-bladder was not included in the present study, and thisfact may be one possible explanation for the similarresponses observed to known spasmogens, including CCK,in both control and acute cholecystitis gallbladders.
In this study, we investigated the mechanisms respon-sible for the BK-induced contraction and for the in-creased efficacy of this peptide in acute cholecystitisgallbladders. The hypothesis that acetylcholine, releasedfrom postganglionic cholinergic nerves, contributes tothe BK-evoked contraction of isolated human gallblad-der strips was excluded by the observation that atropinefailed to inhibit the response to BK. In the guinea piggallbladder, the nonadrenergic noncholinergic compo-nent of the contractile response to electrical stimuliresults from the release of substance P and neurokinin Afrom sensory/intrinsic nerve terminals and the subse-quent activation of tachykinin NK2, and possibly NK3,receptors.31–34 Kinins are known to stimulate tachyki-nin-containing sensory nerve terminals in a variety oftissues and organs via B2 receptor activation.7 However,because the BK-induced motor response in the humangallbladder was not affected by the combination of NK1,NK2, and NK3 receptor antagonists, the possibility thattachykinins released from extrinsic (sensory) and/or in-trinsic nerves contribute to, or enhance, gallbladder con-tractility appears unlikely. BK contracts guinea pig-isolated gallbladder via activation of B2 receptors andprostaglandin release,15,35 and the ability of BK to release
Table 1. Emax and pD2 of Different Spasmogens in Isolated Strips of Human Gallbladder
Control Acute cholecystitis
pD2 Emax (mg) pD2 Emax (mg)
KCI 1.79 � 0.08 451.2 � 51.5 1.78 � 0.09 456.2 � 71.2Carbachol 5.38 � 0.25 168.0 � 41.5 5.55 � 0.29 165.3 � 34.8Bradykinin 6.83 � 0.27 287.6 � 42.7 6.65 � 0.24 731.5 � 80.0*,‡
Cholecystokinin 7.65 � 0.12 724.5 � 60.0 7.41 � 0.18 775.2 � 62.3
NOTE. Data are mean � SEM of at least 5 experiments.*P � 0.01 versus controls.‡P � 0.05 versus carbachol.
Figure 7. Effect of HOE 140, R-715, indomethacin, and the combi-nation of SR 140333, SR 48968, and SR 142801 (E) or theirrespective vehicles (● ) on the concentration-response curves to bra-dykinin in strips of human gallbladders in vitro obtained from patientswith acute cholecystitis. Values are means � standard error of themean of at least 5 experiments. *P � 0.05 versus vehicle.
132 TREVISANI ET AL. GASTROENTEROLOGY Vol. 125, No. 1

prostaglandins is increased by acute inflammatory con-ditions.36 However, we found that indomethacin fails toaffect the concentration response curve to BK, thus rul-ing out the possibility that prostanoids significantlycontribute to the contraction produced by BK in thehuman gallbladder in vitro.
Regarding the kinin receptor responsible for the BK-induced gallbladder contraction, the exclusive involve-ment of B2 receptors was indicated by the observationthat the selective B2 receptor antagonist, HOE 140,shifted to the right the concentration response curve toBK, whereas the B1 selective antagonist, R-715, wasinactive. It is well known that inflammatory mediators orin vitro incubation of the tissue up-regulates the expres-sion of B1 receptors.3 However, even though the presentexperiments were performed in clearly inflamed gallblad-ders, the observation that BK is a selective B2 receptoragonist2,5 along with the evidence that R-715 failed toaffect the response to BK excluded the possibility that B1
receptors, or its possible up-regulation, plays any rele-vant role in the contraction produced by BK. Recently,it has been reported that B2 receptor expression is alsoup-regulated by inflammation.6 The present observationthat the expression of B2 receptor mRNA was increasedin acute cholecystitis as compared with control gallblad-ders supports the possibility that inflammation underliesthe acute cholecystitis up-regulation of B2 receptor ex-pression. This hypothesis was strengthened by the ob-servation that the number of binding sites for the B2
receptor ligand was increased in acute cholecystitis tis-sues, thus suggesting that an augmented number of B2
receptors may contribute to the exaggerated contractileresponse to BK in acute cholecystitis.
Kinins are short-lived bioactive peptides released fromtissue and plasma protein precursors (i.e., the kinino-gens) and derived by the action of 2 serine-proteases, thetissue and plasma kallikreins.1 Once formed, these pep-tides exert most of their actions in a paracrine fashion.Requirements for paracrine function are the presence offorming enzymes and substrates, expression of the ap-propriate kinin receptor, and presence of the kinin-metabolizing enzymes (i.e., kininases). Tissue kallikreinsand kininogens have been constitutively identified invarious cell types,1 whereas plasma kininogens are likelyto be present within tissues, particularly during inflam-mation, when plasma proteins extravasate in large quan-tities in the interstitial space.1,7,8 The existence of cellsexpressing either kallikreins or kininogens in the humangallbladder has not been reported previously, althoughboth human bile and biliary tract homogenates possessthe ability to produce kinins.37 The question we have
tried to address in the present article is whether tissuekallikrein and the kinin precursors were expressed byspecific cells of the human gallbladder. Our immunohis-tochemical findings, showing decreased kallikrein im-munoreactivity along with increased kininogen immu-nostaining in acute cholecystitis, provide a valuablemorphological basis that further supports the role ofkinin peptides in the inflamed gallbladder. The demon-stration of both kallikrein and kininogens and of theirchanges in specific areas (epithelial cells of the mucosa,lamina propria, and interstitial space between mucosalepithelial cells) of the human gallbladder suggests thatkinin formation is feasible in this tissue. The mecha-nisms leading to decreased kallikrein immunoreactivityduring inflammation remain unknown because there isno previous study on tissue kallikrein expression in thehuman gallbladder and on the specific stimuli that maytrigger its release. In this study, we were unable todiscriminate whether kallikrein diminution was becauseof cell damage, provoked by consecutive inflammatoryepisodes, or because of an increased secretion and redis-tribution of the enzyme, although both mechanisms maycontribute.
A further interesting immunohistochemical result ofthis study was the detection of kallikrein immunoreac-tivity in leukocytes infiltrating the inflamed gallbladder,a finding that expands previous evidence in the field38–40
because it indicates a new source of kallikrein in acutecholecystitis, in addition to the epithelium. Previousstudies have shown that human polymorphonuclear leu-kocytes, isolated from peripheral blood of normal sub-jects, contain within their enzymatic pool a tissue kal-likrein38–40 that can be released after stimulation withthrombin.41 In addition, various factors involved in con-tact-phase activation, such as intact high–molecular-weight kininogen, plasma prekallikrein factor XI andXII are placed on the exterior surface of the humanneutrophil indicating that, during contact-phase activa-tion, kinins may be released by the activation of plasmaprekallikrein.42,43 Thus, hydrolysis of plasma kininogensby either tissue or plasma kallikreins may lead to kininformation during acute cholecystitis. Tissue kallikreinsmay originate from infiltrating polymorphonuclear leu-kocytes and likely from epithelial gallbladder cells,whereas formation of kinins by neutrophil-bound plasmaprekallikrein may be circumscribed to the vicinity of theinflammatory cells.43 To assess the real contribution ofeach component of the kinin system in the inflamedgallbladder, ideally one would like to measure each ac-tive enzyme (factor XII, kallikreins, kininases), show theappearance of cleaved kininogens (after kinin release),
July 2003 B2 RECEPTOR ACTIVATION IN HUMAN GALLBLADDER 133

and assay BK and its receptor. However, these determi-nations are difficult to perform because some of theenzymes and BK are rapidly inactivated or destroyed andoften assays are only able to determine the level ofresidual components. Moreover, kinins can also beformed by activation of the corresponding kininogenasesduring sample preparation and blood-contaminated tis-sue extracts commonly contain significant amounts ofplasma kallikrein and kininogens.1
In conclusion, the present study shows that BK exertsan important role in evoking contraction of human gall-bladder, particularly during inflammation. Should BKplay a role in the mechanism underlying acute cholecys-titis, then newly developed, high-affinity peptide andnonpeptide B2 receptor antagonists (e.g., HOE 140,WIN 64338, and FR-173657)16,44–46 might be a usefuloption in the pharmacological treatment of this disease.
References1. Bhoola K, Figueroa CD, Worthy K. Bioregulation of kinins: kal-
likreins, kininogens, and kininases. Pharmacol Rev 1992;44:1–80.
2. Regoli D, Barabe’ J. Pharmacology of bradykinin and relatedkinins. Pharmacol Rev 1980;32:1–46.
3. Marceau F. Kinin B1 receptor: a review. Immunopharmacology1995;30:1–26.
4. Regoli D. Kinins. Br Med Bull 1987;43:270–284.5. Marceau F, Hess JF, Bacharov DR. The B1 receptors for kinins.
Pharmacol Rev 1998;50:357–386.6. Schmidlin F, Scherrer D, Daeffler L, Bertrand C, Landry Y, Gies JP.
Interleukin-1beta induces bradykinin B2 receptor gene expres-sion through a prostanoid cyclic AMP-dependent pathway in hu-man bronchial smooth muscle cells. Mol Pharmacol 1998;53:1009–1015.
7. Geppetti P. Sensory neuropeptide release by bradykinin: mecha-nisms and pathophysiological implications. Regul Pept 1993;47:1–23.
8. Proud D, Kaplan AP. Kinin formation: mechanisms and role ininflammatory disorders. Annu Rev Immunol 1988;6:49–84.
9. Fuller R, Dixon C, Cuss F, Barnes P. Bradykinin-induced broncho-constriction in humans: mode of action. Am Rev Respir Dis1987;135:176–180.
10. Ricciardolo FLM, Di Maria GU, Sapienza MA, Mistretta A, GeppettiP. Impairment of bronchoprotection by NO in severe asthma.Lancet 1997;350:1297–1298.
11. Figueroa CD, Marchant A, Novoa U, Forstermann U, Jarnagin K,Scholkens B, Muller-Esterl W. Differential distribution of bradyki-nin B(2) receptors in the rat and human cardiovascular system.Hypertension 2001;37:110–120.
12. Meini S, Patacchini R, Giuliani S, Lazzeri M, Turini D, Maggi CA,Lecci A. Characterization of bradykinin B(2) receptor antagonistsin human and rat urinary bladder. Eur J Pharmacol 2000;388:177–182.
13. Cabrini DA, Silva AM, Calixto JB. Mechanisms of bradykinin-induced contraction of the guinea-pig gallbladder in vitro. Br JPharmacol 1995;114:1549–1556.
14. Woods M, Baird AW. Increased sensitivity of guinea pig gallblad-der in vitro to bradykinin. Agents Actions Suppl 1992;38:371–377.
15. Falcone RC, Hubbs SJ, Vanderloo JD, Prosser JC, Little J, GomesB, Aharony D, Krell RD. Characterization of bradykinin receptors
in guinea pig gall bladder. J Pharmacol Exp Ther 1993;266:1291–1299.
16. Hock F, Wirth K, Albus U, Linz W, Gerhards H, Wiemer G, HenkeG, Breipohl G, Konig W, Knolle J, Scholkens B. HOE 140 a newpotent and long acting bradykinin-antagonist: in vitro studies. Br JPharmacol 1991;102:769–773.
17. Regoli D, Nsa Allogho S, Rizzi A, Gobeil FJ. Bradykinin receptorsand their antagonists. Eur J Pharmacol 1998;348:1–10.
18. Emonds-Alt X, Vilain P, Goulaouic P, Proietto V, Van Broeck D,Advenier C, Naline E, Neliat G, Le Fur G, Breliere JC. A potent andselective non-peptide antagonist of the neurokinin A (NK2) recep-tor. Life Sci 1992;50:L101–106.
19. Emonds-Alt X, Doutremepuich JD, Heaulme M, Neliat G, SantucciV, Steinberg R, Vilain P, Bichon D, Ducoux JP, Proietto V, VanBroeck D, Soubrie P, Le Fur G, Breliere JC. In vitro and in vivobiological activities of SR140333, a novel potent non-peptidetachykinin NK1 receptor antagonist. Eur J Pharmacol 1993;250:403–413.
20. Emonds-Alt X, Bichon D, Ducoux JP, Heaulme M, Miloux B, Pon-celet M, Proietto V, Van Broeck D, Vilain P, Neliat G, Soubrie P, LeFur G, Breliere JC. SR 142801, the first potent non-peptideantagonist of the tachykinin NK3 receptor. Life Sci 1995;56:L27–32.
21. Trevisani M, Schmidlin F, Tognetto M, Nijkamp FP, Gies JP,Frossard N, Amadesi S, Folkerts G, Geppetti P. Evidence for invitro expression of B1 receptor in the mouse trachea and urinarybladder. Br J Pharmacol 1999;126:1293–1300.
22. Bradford MM. A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of pro-tein-dye binding. Anal Biochem 1976;72:248–254.
23. Manning DC, Snyder SH. 3H-bradykinin binding site localization inguinea pig urinary system. Adv Exp Med Biol 1986;198:563–570.
24. Gessi S, Rizzi A, Calo G, Agnello G, Jorizzo G, Mollica G, Borea PA,Regoli D. Human vascular kinin receptors of the B2 type charac-terized by radioligand binding. Br J Pharmacol 1997;122:1450–1454.
25. Bagshaw AF, Bhoola KD, Lemon MJ, Whicher JT. Developmentand characterization of a radioimmunoassay to measure humantissue kallikrein in biological fluids. J Endocrinol 1984;101:173–179.
26. Vio CP, Figueroa CD, Caorsi I. Anatomical relationship betweenkallikrein-containing tubules and the juxtaglomerular apparatusin the human kidney. Am J Hypertens 1988;1:269–271.
27. Ahmed R, Bird NC, Chess-Williams R, Thomas WE, Johnson AG.In vitro responses of gallbladder muscle from patients with acal-culous biliary pain. Digestion 2000;61:140–144.
28. Meyer BM, Werth BA, Beglinger C, Hildebrand P, Jansen JB, ZachD, Rovati LC, Stalder GA. Role of cholecystokinin in regulation ofgastrointestinal motor functions. Lancet 1989;2:12–15.
29. Shaffer EA. Review article: control of gall-bladder motor function.Aliment Pharmacol Ther 2000;2:2–8.
30. Tang C, Biemond I, Lamers CB. Cholecystokinin receptors inhuman pancreas and gallbladder muscle: a comparative study.Gastroenterology 1996;111:1621–1626.
31. Maggi CA, Santicioli P, Renzi D, Patacchini R, Surrenti C, Meli A.Release of substance P- and calcitonin gene-related peptide-likeimmunoreactivity and motor response of the isolated guineapig gallbladder to capsaicin. Gastroenterology 1989;96:1093–1101.
32. Mawe GM. Tachykinins as mediators of slow EPSPs in guinea-piggall-bladder ganglia: involvement of neurokinin-3 receptors.J Physiol (Lond) 1995;485:513–524.
33. Patacchini R, De Giorgio R, Giachetti A, Maggi CA. Differentmechanism of tachykinin NK2 receptor blockade by SR 48968and MEN 10,627 in the guinea-pig isolated gallbladder and co-lon. Eur J Pharmacol 1994;271:111–119.
134 TREVISANI ET AL. GASTROENTEROLOGY Vol. 125, No. 1

34. Patacchini R, De Giorgio R, Bartho L, Barbara G, Corinaldesi R,Maggi CA. Evidence that tachykinins are the main NANC excita-tory neurotransmitters in the guinea-pig common bile duct. Br JPharmacol 1998;124:1703–1711.
35. Cabrini DA, Calixto JB. Characterization of des-Arg9-bradykinin-induced contraction in guinea-pig gallbladder in vitro. Eur J Phar-macol 1997;331:31–38.
36. Bogar LJ, Bartula LL, Parkman HP, Myers SI. Enhanced bradyki-nin-stimulated prostaglandin release in the acutely inflamedguinea pig gallbladder is due to new synthesis of cyclooxygenase1 and prostacyclin synthase. J Surg Res 1999;84:71–76.
37. Moller-Nielsen H. Kinin forming and inactivating activities in hu-man bile and biliary tract homogenates. Scand J Clin Lab InvestSuppl 1969;107:73–82.
38. Figueroa CD, MacIver AG, Bhoola KD. Identification of a tissuekallikrein in human polymorphonuclear leucocytes. Br J Haematol1989;72:321–328.
39. Wu HF, Venezie RD, Cohen WM, Jenzano JW, Featherstone GL,Lundblad RL. Identification of tissue kallikrein messenger RNA inhuman neutrophils. Agents Actions 1993;38:27–31.
40. Naidoo Y, Snyman C, Raidoo DM, Bhoola KD, Kemme M, Muller-Esterl W. Cellular visualization of tissue prokallikrein in humanneutrophils and myelocytes. Br J Haematol 1999;105:599–612.
41. Cohen WM, Wu HF, Featherstone GL, Jenzano JW, Lundblad RL.Linkage between blood coagulation and inflammation: stimula-tion of neutrophil tissue kallikrein by thrombin. Biochem BiophysRes Commun 1991;176:315–320.
42. Figueroa CD, Henderson LM, Kaufmann J, De la Cadena RA,Colman RW, Muller-Esterl W, Bhoola KD. Immunovisualisation ofhigh (HK) and low (LK) molecular weight kininogens on isolatedhuman neutrophils. Blood 1992;79:754–759.
43. Henderson LM, Figueroa CD, Muller-Esterl W, Bhoola KD. Assem-
bly of contact-phase factors on the surface of the human neutro-phil membrane. Blood 1994;84:474–482.
44. Asano M, Hatori C, Sawai H, Johki S, Inamura N, Kayakiri H,Satoh S, Abe Y, Inoue T, Sawada Y, Mizutani T, Oku T, NakaharaK. Pharmacological characterization of a nonpeptide bradykininB2 receptor antagonist, FR165649, and agonist, FR190997. Br JPharmacol 1998;124:441–446.
45. Inamura N, Asano M, Hatori C, Sawai H, Hirosumi J, Fujiwara T,Kayakiri H, Satoh S, Abe Y, Inoue T, Sawada Y, Oku T, NakaharaK. Pharmacological characterization of a novel, orally active,nonpeptide bradykinin B2 receptor antagonist, FR167344. EurJ Pharmacol 1997;333:79–86.
46. Sawutz DG, Salvino JM, Dolle RE, Casiano F, Ward SJ, Houck WT,Faunce DM, Douty BD, Baizman E, Awad MM, Marceau F, SeoanePR. The nonpeptide WIN 64338 is a bradykinin B2 receptorantagonist. Proc Natl Acad Sci U S A 1994;91:4693–4697.
Received May 15, 2002. Accepted April 10, 2003.Reprint requests to: Pierangelo Geppetti, M.D., Department of Ex-
perimental & Clinical Medicine, Pharmacology Unit, University of Fer-rara, Via Fossato di Mortara 19, 44100, Ferrara, Italy. e-mail:[email protected]; fax: (39) 0532-291-205.
Supported by a grant from the Italian Ministry of the University,Research, Science and Technology, Rome (to P. G. and R. De G.).
The authors thank Dr. Elisabetta Marchetti (Department of Experi-mental and Diagnostic Medicine, University of Ferrara, Ferrara, Italy)and Dr. Daniela Renzi (Department of Gastroenterology, University ofFlorence, Florence, Italy) for their expertise in histopathological anal-ysis of human gallbladders, and Ms. P. Ehrenfeld (Department ofPathology Institute of Histology and Pathology Austral University, IslaTeja, Valdivia, Chile) for her valuable help in immunohistochemistry.
July 2003 B2 RECEPTOR ACTIVATION IN HUMAN GALLBLADDER 135
Copyright © 2022 FDOKUMEN




















