
9 The Earliest Putative Homo Fossils
43
Handbook of Paleoanthropology
Transcript of 9 The Earliest Putative Homo Fossils

Handbook of Paleoanthropology

Prof. Dr. Dr. h. c. Winfried Henke
Institut fur Anthropologie (1050)
Fachbereich 10 ‐ BiologieJohannes Gutenberg‐Universitat Mainz
D‐55099 Mainz
Germany
email: [email protected]
Prof. Dr. Ian Tattersall
Division of Anthropology
American Museum of Natural History
New York, NY 10024‐5192USA
email: [email protected]
Dipl. Biol. Thorolf Hardt
Institut fur Anthropologie (1050)
Fachbereich 10 ‐ BiologieJohannes Gutenberg‐Universitat Mainz
D‐55099 Mainz
Germany
email: [email protected]
ISBN‐13: 978‐3‐540‐32474‐4
This publication is available also as:Electronic publication under ISBN 978‐3‐540‐33761‐4 andPrint and electronic bundle under ISBN 978‐3‐540‐33858‐1
Library of Congress Control Number: 2006936414
This work is subject to copyright. All rights are reserved, whether the whole or part of thematerial is concerned, specifically the rights of translation, reprinting, reuse of illustrations,recitation, broadcasting, reproduction on microfilms or in other ways, and storage in databanks. Duplication of this publication or parts thereof is only permitted under the provisions ofthe German Copyright Law of September 9, 1965, in its current version, and permission for usemust always be obtained from Springer‐Verlag. Violations are liable for prosecution under theGerman Copyright Law.
Springer is part of Springer Science+Business Media
springer.com
# Springer‐Verlag Berlin Heidelberg New York 2007
The use of registered names, trademarks, etc. in this publication does not imply, even in theabsence of a specific statement, that such names are exempt from the relevant protective lawsand regulations and therefore free for general use.
Product liability: The publishers cannot guarantee the accuracy of any information about theapplication of operative techniques and medications contained in this book. In every individualcase the user must check such information by consulting the relevant literature.
Editor: Dieter Czeschlik, Heidelberg/Sandra Fabiani, HeidelbergDevelopment Editor: Susanne Friedrichsen, HeidelbergProduction Editor: Frank Krabbes, HeidelbergCover Design: Frido Steinen‐Broo, Spain
Printed on acid‐free paper SPIN: 2109 ‐ 5 4 3 2 1 0

We dedicate these volumes to our long-time colleagues
Hartmut Rothe and Theodoros Pitsios
in appreciation of their friendship and unique contributions to
primatology and paleoanthropology.

Preface to the Series
Palaeoanthropology is perhaps the most multidisciplinary of all the sciences. Any
complete account of the evolution and of the cultural and biological contexts
of Homo sapiens must combine information from geology, paleoecology, prima-
tology, evolutionary biology and a host of other fields. Above all, historical
information garnered from the fossil record needs to be combined with, and
interpreted in the light of, what we know of the living world. In these volumes we
have brought together contributions by a variety of leading specialists that reflect
the broad spectrum of modern paleoanthropology, in an attempt to provide a
resource that we hope will be useful to professionals and students alike.
Volume I of this three-volume Handbook deals with principles, methods,
and approaches. In recent years enormous advances have been made in such areas
as phylogenetic analysis, evolutionary theory and philosophy, paleoecology, and
dating methods. The contributions aim to present the state of the art in these
and other relevant fields, as well as to furnish succinct introductions to them and
to reflect the many ways in which they interact. Human beings are primates,
and Volume II is devoted to primate origins, evolution, behavior, and adaptive
variety. In this compilation the emphasis is on the integration of fossil data with
the vast amount that is now known of the behavior and ecology of living primates
in natural environments. The third and final volume deals directly with the fossil
and molecular evidence for the evolution of Homo sapiens and its fossil relatives
(the family Hominidae or subfamily Homininae, according to taste, a matter that
we have left to each individual contributor). Paleoanthropology is a pluralistic
and actively developing field in which much remains to be settled, and we have
not tried to impose any uniformity of viewpoint on our authors. Instead, while
maintaining an emphasis on the data, we have encouraged them to express their
individual interpretations rather than to cover all possible points of view. This has
inevitably led to a certain degree of heterogeneity of opinion between the covers
of this Handbook; but we believe that this is the best way of reflecting the
excitement and momentum of the field and that it is best for the reader to be
left to reach his or her own conclusions. Science is, after all, a process rather than
a static product, and one of our primary aims here is to reflect the ongoing
dynamism of that process in paleoanthropology.
We thank all of the contributors to these volumes for their participation.
Some initially responded enthusiastically while others needed convincing about
the basic strategy of the Handbook, but all responded marvellously to the

viii Preface to the Series
particular needs of a corporate effort such as this one. We are particularly grateful
to those authors who responded at short notice to needs that became apparent
only as the project progressed. This series was conceived in collaboration with
Prof. Hartmut Rothe of the University of Gottingen, who was later forced to
withdraw for reasons beyond his control. We thank him most warmly for his
creativity in the conceptual stages and for his subsequent moral support. The
laborious process of putting together the volumes could not have been accom-
plished without the cheerful help of Thorolf Hardt, whose active involvement was
indispensable throughout.
This project could never have come to fruition without the enthusiastic
support of Dr. Dieter Czeschlik, editor life sciences at Springer Publishing, and
the efficient assistance of Mrs. Ursula Gramm. We express our deep gratitude to
Mrs. Susanne Friedrichsen and Mrs. Caroline Simpson, who showed both care
and commitment during the phases of copyediting and product development.
The continuous cooperation and dialogue with them and their professionalism
gave us the courage to see the project through. Further thanks go to Mrs. Sandra
Fabiani and her colleagues at Springer Publishing, who prepared the eReference.
Ms. Nitya Swaruba, compositor at SPi Technologies, deserves warm thanks for
her efficient help, and finally our gratitude goes in addition to Britta Hardt, Peter
Menke and Monika Sandfuhr, who also rendered much valuable assistance.
Winfried Henke and Ian Tattersall
Mainz and New York City
November, 2006

Table of Contents
Volume 1
1 Historical Overview of Paleoanthropological Research . . . . . . . . . . . . . . . 1Winfried Henke
2 Evolutionary Theory in Philosophical Focus . . . . . . . . . . . . . . . . . . . . . . . 57Philippe Huneman
3 The Ontogeny–Phylogeny Nexus in a Nutshell: Implications for
Primatology and Paleoanthropology . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103Peter R. Menke
4 Principles of Taxonomy and Classification: Current Procedures
for Naming and Classifying Organisms . . . . . . . . . . . . . . . . . . . . . . . . . . 141Michael Ohl
5 Quantitative Approaches to Phylogenetics . . . . . . . . . . . . . . . . . . . . . . . 167Kaila E. Folinsbee . David C. Evans . Jorg Frobisch . Linda A. Tsuji .
Daniel R. Brooks
6 Homology: A Philosophical and Biological Perspective . . . . . . . . . . . . . 217Olivier Rieppel
7 Taphonomic and Diagenetic Processes . . . . . . . . . . . . . . . . . . . . . . . . . . 241Gisela Grupe
8 Archeology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 261Miriam N. Haidle
9 Contribution of Stable Light Isotopes to Paleoenvironmental
Reconstruction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 289
Julia Lee‐Thorp . Matt Sponheimer10 Chronometric Methods in Paleoanthropology . . . . . . . . . . . . . . . . . . . . 311Gunther A. Wagner

xxvi
11 Geological Background of Early Hominid Sites in Africa . . . . . . . . . . . . 339Ottmar Kullmer
12 Paleoclimate . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 357Keith Alverson
13 Paleosols . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 383Gregory Retallack
14 Quaternary Deposits and Paleosites . . . . . . . . . . . . . . . . . . . . . . . . . . . . 409Klaus‐Dieter Jager
15 Zoogeography: Primate and Early Hominin Distribution and
Migration Patterns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 421Alan Turner . Hannah O’Regan
16 Patterns of Diversification and Extinction . . . . . . . . . . . . . . . . . . . . . . . . 441Walter Etter
17 Paleoecology: An Adequate Window on the Past? . . . . . . . . . . . . . . . . . 503Thorolf Hardt . Britta Hardt . Peter R. Menke
18 Hominin Paleodiets: The Contribution of Stable Isotopes . . . . . . . . . . . 555Matt Sponheimer . Julia Lee ‐Thorp
19 Estimation of Basic Life History Data of Fossil Hominoids . . . . . . . . . . 587Helmut Hemmer
20 Population Genetics and Paleoanthropology . . . . . . . . . . . . . . . . . . . . . 621John H. Relethford
21 Ancient DNA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 643Susanne Hummel
22 Paleodemography of Extinct Hominin Populations . . . . . . . . . . . . . . . . 673Janet Monge . Alan Mann
23 Modeling the Past: The Primatological Approach . . . . . . . . . . . . . . . . . 701R. W. Sussman . Donna Hart
Table of Contents
24 Modeling the Past: The Paleoethnological Evidence . . . . . . . . . . . . . . . 723Paolo Biagi

xxvii
25 Modeling the Past: The Linguistic Approach . . . . . . . . . . . . . . . . . . . . . 747Bernard Comrie
26 General Principles of Evolutionary Morphology . . . . . . . . . . . . . . . . . . . 769Gabriele A. Macho
27 Computer‐Based Reconstruction: Technical Aspects
and Applications . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 787Lilian Ulhaas
Table of Contents
28 Prospects and Pitfalls . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 815Jean-Jacques Hublin
Volume 2
1 Primate Origins and Supraordinal Relationships: Morphological
Evidence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 831Mary T. Silcox . Eric J. Sargis . Jonathan I. Bloch . Doug M. Boyer
2 Molecular Evidence on Primate Origins and Evolution . . . . . . . . . . . . . 861Hans Zischler
3 Fossil Record of the Primates from the Paleocene to the Oligocene . . . . . 889D. Tab Rasmussen
4 Fossil Record of Miocene Hominoids . . . . . . . . . . . . . . . . . . . . . . . . . . . . 921David R. Begun
5 The Biotic Environments of the Late Miocene Hominids . . . . . . . . . . . . 979Jordi Agustı
6 Postcranial and Locomotor Adaptations of Hominoids . . . . . . . . . . . . 1011Carol V. Ward
7 Hominoid Cranial Diversity and Adaptation . . . . . . . . . . . . . . . . . . . . . 1031Alan Bilsborough . Todd C. Rae
8 Dental Adaptations of African Apes . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1107.
Mark F. Teaford Peter S. Ungar9 Evolution of the Primate Brain . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1133Dean Falk

xxviii
10 Primate Life Histories . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1163Elke Zimmermann . Ute Radespiel
11 The Biology and Evolution of Ape and Monkey Feeding . . . . . . . . . . . 1207Joanna E. Lambert
12 Great Ape Social Systems . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1235Angela Meder
13 Primate Intelligence . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1273Richard W. Byrne
14 Chimpanzee Hunting Behavior . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1295Nicholas E. Newton‐Fisher
15 Cooperation, Coalition, and Alliances . . . . . . . . . . . . . . . . . . . . . . . . . . 1321Charlotte K. Hemelrijk . Jutta Steinhauser
Volume 3
1 Potential Hominoid Ancestors for Hominidae . . . . . . . . . . . . . . . . . . . . 1347George D. Koufos
2 Defining Hominidae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1379Jeffrey H. Schwartz
3 Origins of Homininae and Putative Selection Pressures
Acting on the Early Hominins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1409Bogusław Pawłowski
4 Role of Environmental Stimuli in Hominid Origins . . . . . . . . . . . . . . . . 1441Elisabeth S. Vrba
5 The Origins of Bipedal Locomotion . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1483William E. H. Harcourt‐Smith
6 The Earliest Putative Hominids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1519Brigitte Senut
Table of Contents
7 The Species and Diversity of Australopiths . . . . . . . . . . . . . . . . . . . . . . 1539William H. Kimbel

xxix
8 Defining the Genus Homo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1575Mark Collard . Bernard Wood
9 The Earliest Putative Homo Fossils . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1611Friedemann Schrenk . Ottmar Kullmer . Timothy Bromage
10 Homo ergaster and Its Contemporaries . . . . . . . . . . . . . . . . . . . . . . . . . 1633Ian Tattersall
11 Defining Homo erectus: Size Considered . . . . . . . . . . . . . . . . . . . . . . . . 1655Susan C. Anton . Fred Spoor . Connie D. Fellmann . Carl C. Swisher III
12 Later Middle Pleistocene Homo . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1695G. Philip Rightmire
13 Neanderthals and Their Contemporaries . . . . . . . . . . . . . . . . . . . . . . . 1717Katerina Harvati
14 Origin of Modern Humans . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1749Gunter Brauer
15 Analyzing Hominid Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1781David Strait . Frederick E. Grine . John G. Fleagle
16 Phylogenetic Relationships (Biomolecules) . . . . . . . . . . . . . . . . . . . . . . 1807Todd R. Disotell
17 Population Biology and Population Genetics of
Pleistocene Hominins . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1825Alan R. Templeton
18 Species Concepts and Speciation: Facts and Fantasies . . . . . . . . . . . . 1861Colin Groves
19 Human Environmental Impact in the Paleolithic and Neolithic . . . . . 1881Wolfgang Nentwig
20 The Dentition of American Indians: Evolutionary Results
Table of Contents
and Demographic Implications Following Colonization
from Siberia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1901Christy G. Turner II . G. Richard Scott

xxx
21 Overview of Paleolithic Archeology . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1943Nicholas Toth . Kathy Schick
22 The Network of Brain, Body, Language, and Culture . . . . . . . . . . . . . . 1965Steven Mithen
23 Cultural Evolution in Africa and Eurasia During the Middle
and Late Pleistocene . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2001Nicholas Conard
24 Paleoanthropology and the Foundation of Ethics: Methodological
Table of Contents
Remarks on the Problem of Criteriology . . . . . . . . . . . . . . . . . . . . . . . . 2039Mathias Gutmann . Michael Weingarten

About the Editors
Editors in Chief
Prof. Dr. Dr. h. c. Winfried Henke
Institut fur Anthropologie (1050)
Fachbereich 10 - Biologie
Johannes Gutenberg-Universitat Mainz
D‐55099 Mainz
Germany
email: [email protected]
Prof. Dr. Ian Tattersall
Division of Anthropology
American Museum of Natural History
New York, NY 10024–5192
USA
email: [email protected]
Editorial Assistant
Dipl. Biol. Thorolf Hardt
Institut fur Anthropologie (1050)
Fachbereich 10 - Biologie
Johannes Gutenberg-Universitat Mainz
D‐55099 Mainz
Germany
email: [email protected]
Winfried Henke is currently Professor of Anthropology at the Johannes
Gutenberg University of Mainz. He was born in 1944 in Ludwigshorst/
Pomerania, Germany, and studied biology, anthropology and geosciences in
Kiel and Braunschweig. He received his Ph.D. from the University of Kiel in
1971, his thesis focusing on a prehistoric anthropological topic. In 1990 he
habilitated with a monograph on the ‘‘Anthropology of Early Paleolithics and
Mesolithics’’ at the University of Mainz.
Research activities in various countries (Iceland, Israel, Jordan, US, Greece)
and extensive teaching assignments in the Erasmus exchange program at
numerous European Universities followed. From 1996 to 2004, he acted as
anthropology referee for the German Research Foundation (DFG). He served at
the advisory boards of many scientific journals, and was advisory consultant to
museums, e.g. the Neanderthal Museum (Mettmann, Germany). In 2006 he was
awarded with the honorary doctorate of the National and Kapodistrian
University of Athens and is an elected member of the German Academy of

2072 About the Editors
Sciences Leopoldina. Areas of research and teaching: paleoanthropology,
primatology, prehistoric anthropology, comparative morphology, systematics,
demography and sociobiology.
He has published approximately 180 original papers in scientific journals and
anthologies, and over 600 book reviews. He is author, co-author (together with
H. Rothe) and editor of several books, including such standard works as
‘‘Palaoanthropologie’’ and ‘‘Stammesgeschichte des Menschen’’ (published at
Springer-Verlag).
Ian Tattersall is currently Curator in the Division of Anthropology of the
American Museum of Natural History in New York City. Born in England and
raised in East Africa, he has carried out both primatological and paleontological
fieldwork in countries as diverse as Madagascar, Vietnam, Surinam, Yemen and
Mauritius. Trained in archeology and anthropology at Cambridge, and in
geology and vertebrate paleontology at Yale, Tattersall has concentrated his
research since the 1960s in two main areas: the analysis of the human fossil
record and its integration with evolutionary theory, and the study of the
ecology and systematics of the lemurs of Madagascar. Tattersall is also a
prominent interpreter of human paleontology to the public, with several trade
books to his credit, among them The Monkey in the Mirror (2002), Extinct
Humans (with Jeffrey Schwartz, 2000), Becoming Human: Evolution and Human
Uniqueness (1998) and The Last Neanderthal: The Rise, Success and Mysterious
Extinction of Our Closest Human Relatives (1995; rev. 1999) as well as several
articles in Scientific American and the co-editorship of the definitive Encyclopedia
of Human Evolution and Prehistory. He lectures widely, and, as curator, has also
been responsible for several major exhibits at the American Museum of Natural
History, including Ancestors: Four Million Years of Humanity (1984); Dark Caves,
Bright Visions: Life In Ice Age Europe (1986); Madagascar: Island of the Ancestors
(1989); The First Europeans: Treasures from the Hills of Atapuerca (2003); and the
highly acclaimed Hall of Human Biology and Evolution (1993).
Thorolf Hardt graduated in biology and is currently a PhD student at the
Institute of Anthropology, University of Mainz. He was born in 1973 in
Neustadt (RhP/Germany) and studied anthropology, paleontology and zoology
in Kiel and Mainz. At present his research activities are focused on Geometric
Morphometrics, functional morphology and evolutionary biology in Primates.

Contributors
J. Agustı Ballester
ICREA-Institut of Human
Paleoecology,
Universitat Rovira i Virgili,
Pl. Imperial Tarraco, 1,
43005-Tarragona,
Spain
K. Alverson
Ocean Observations and Services,
IOC/UNESCO,
Global Ocean Observing System,
1 rue Miollis,
75732 Paris Cedex 15,
France
S. C. Anton
Center for the Study of Human
Origins,
Department of Anthropology NYU,
25 Waverly Place,
New York, NY 10003,
USA
D. R. Begun
Department of Anthropology,
100 St George Street, Rm 1037,
University of Toronto,
Toronto, ONT M5S 3G3,
Canada
P. Biagi
Department of Science of Antiquities
and the Near East,
Ca’ Foscari University, Venice,
Palazzo Bernardo, S. Polo 1977,
30125 Venezia,
Italy
A. Bilsborough
Department of Anthropology,
University of Durham,
43 Old Elvet, Durham,
DH 1 3HN,
UK
J. I. Bloch
Vertebrate Paleontology,
Florida Museum of Natural History,
Dickenson Hall, University
of Florida,
Gainesville, Florida 32611‐7800,USA
D. M. Boyer
Department of Anatomical
Sciences,
Stony Brook University,
Stony Brook, NY 11794‐8081,USA
G. Brauer
Institut fur Humanbiologie,
Universitat Hamburg,
Allende-Platz 2,
D‐20146 Hamburg,
Germany

2074 Contributors
T. Bromage
Department of Biomaterials and
Biomimetics,
New York University College of
Dentistry,
345 East 24th Street, Room 804-S,
New York, NY 10010,
USA
D. R. Brooks
Department of Ecology &
Evolutionary Biology,
University of Toronto,
25 Harbord Street,
Toronto, Ontario M5S 3G5,
Canada
R. W. Byrne
School of Psychology,
University of St. Andrews,
St. Andrews, Fife KY16 9JU,
Scotland
M. Collard
Laboratory of Biological
Anthropology,
Department of Anthropology,
University of British Columbia,
6303 NW Marine Drive,
Vancouver, British Columbia V6T 1Z1,
Canada
B. Comrie
Department of Linguistics,
Max Planck Institute of Evolutionary
Anthropology,
Deutscher Platz 6,
D‐04103 Leipzig,
Germany
N. J. Conard
Institut fur Ur- und Fruhgeschichte,
Abteilung Altere Urgeschichte und
Quartarokologie,
Eberhard-Karls-Universitat Tubingen,
Schloss Hohentubingen,
D-72070 Tubingen,
Germany
T. R. Disotell
Center for the Study of Human
Origins,
Department of Anthropology,
New York University,
25 Waverly Place,
New York, NY 10003,
USA
W. Etter
Naturhistorisches Museum Basel,
Abteilung Geowissenschaften,
Augustinergasse 2,
CH-4001 Basel,
Switzerland
D. Evans
Department of Biology,
University of Toronto at Mississauga,
3359 Mississauga Road,
Mississauga, ON L5L 1C6,
Canada
D. Falk
Department of Anthropology,
Florida State University,
Tallahassee,
FL 32306‐7772, 1847 West,
Tennessee St.,
USA

Contributors 2075
C. D. Fellmann
Center for the Study of Human
Origins,
Department of Anthropology,
New York University,
25 Waverly Place,
New York City, NY 10003,
USA
J. G. Fleagle
Department of Anatomical Sciences,
Health Sciences Centre, Stony
Brook University,
Stony Brook, New York 11794‐8081,USA
K. E. Folinsbee
Department of Biology,
University of Toronto at Mississauga,
3359 Mississauga Road,
Mississauga, ON L5L 1C6,
Canada
J. Frobisch
Department of Biology,
University of Toronto at Mississauga,
3359 Mississauga Road North,
Mississauga, ON L5L 1C6,
Canada
F. E. Grine
Department of Anthropology,
State University of New York,
Stony Brook, NY 11794‐4364,USA
C. P. Groves
School of Archaeology and
Anthropology,
Australian National University
Canberra, ACT 0200,
Australia
G. Grupe
Dept. I fur Biologie,
Bereich Biodiversitatsforschung/
Anthropologie,
Universitat Munchen,
Grosshaderner Straße 2,
D-82152 Planegg-Martinsried,
Germany
M. Gutmann
Institut fur Philosophie,
Philipps-Universitat Marburg,
Wilhelm-Ropke-Strasse 6, Block B
D‐35032 Marburg,
Germany
M. N. Haidle
Institut fur Ur- und Fruhgeschichte
und Archaologie des Mittelalters,
Abt. Altere Urgeschichte und
Quartarokologie,
Universitat Tubingen,
Burgsteige 11,
D-72070 Tubingen,
Germany
W. E. H. Harcourt-Smith
Division of Paleontology,
American Museum of Natural History,
Central Park West and 79th Street,
New York, NY 10024,
USA
B. Hardt
Institut fur Anthropologie (1050),
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat

2076 Contributors
Mainz,
D‐55099 Mainz,
Germany
T. Hardt
Institut fur Anthropologie (1050),
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat
Mainz,
D‐55099 Mainz,
Germany
D. L. Hart
Department of Anthropology,
University of Missouri at St. Louis,
St. Louis, MO 63130, USA
K. Harvati
Department of Human Evolution,
Max Planck Institute for
Evolutionary Anthropology,
Deutscher Platz 6,
D‐04103 Leipzig,
Germany
C. K. Hemelrijk
Theoretical Biology,
Centre for Ecological and
Evolutionary Studies,
University of Groningen,
Biological Centre,
Kerklaan 30,
9751 NN Haren,
The Netherlands
H. Hemmer
Anemonenweg 18,
D‐55129 Mainz,
Germany
or
Institut fur Zoologie,
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat
Mainz,
Johannes v. Muller-Weg 6,
D‐55128 Mainz,
Germany
W. Henke
Institut fur Anthropologie (1050),
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat
Mainz,
D‐55099 Mainz,
Germany
J.-J. Hublin
Department of Human Evolution,
Max Planck Institute for Evolutionary
Anthropology,
Deutscher Platz 6,
D‐4103 Leipzig,
Germany
S. Hummel
Johann Friedrich Blumenbach-
Institut fur Zoologie und
Anthropologie, Historische
Anthropologie und Humanokologie,
Georg August‐Universitat Gottingen,Burgerstrasse 50,
D-37037 Gottingen,
Germany
P. Huneman
Institut d’Histoire et de Philosophie
des Sciences et des Techniques
CNRS/Universite Paris I
Sorbonne,

Contributors 2077
13 rue du Four,
75006 Paris,
France
K.-D. Jager
Institut fur Prahistorische
Archaologie,
Martin Luther-Universitat Halle,
Brandbergweg 23c,
D‐06099 Halle,
Germany
W. H. Kimbel
Institute of Human Origins,
Arizona State University,
P.O. Box 874101,
Tempe, AZ 85287‐4101,USA
G. D. Koufos
Department of Geology,
University of Thessaloniki,
GR‐54124 Thessaloniki,
Greece
O. Kullmer
Forschungsinstitut und
Naturmuseum,
Abt. Palaoanthropologie und
Quartarpalaontologie,
Senckenberganlage 25,
D-60325 Frankfurt am Main,
Germany
J. E. Lambert
Departments of Anthropology and
Zoology,
University of Wisconsin-Madison,
5317 Wm Sewell Social Science
Building,
Madison, WI 53706,
USA
J. Lee-Thorp
Department of Archaeological
Sciences,
University of Bradford,
Bradford, West Yorkshire,
BD7 1DP,
UK
G. A. Macho
Palaeoanthropology Research Group,
Centre for Research of Evolutionary
Anthropology,
Whitelands College,
Roehampton University,
London SW15 4JD,
England
A. E. Mann
Department of Anthropology,
Princeton University,
Princeton,
NJ 08544,
USA
A. Meder
Augustenstrasse 122,
D‐70197 Stuttgart,
Germany
P. Menke
Institut fur Anthropologie (1050),
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat
Mainz,
D‐55099 Mainz,
Germany

2078 Contributors
S. Mithen
School of Human and
Environmental Sciences,
The University of Reading,
Whiteknights, P.O. Box 217,
Reading, Berkshire,
RG6 6AHm UK
J. Monge
Department of Anthropology,
University of Pennsylvania,
University Museum,
3260 South Street,
Philadelphia, PA 19104-6398,
USA
W. Nentwig
Zoologisches Institut,
Universitat Bern,
Baltzer Strasse 6,
CH‐3012 Bern,
Switzerland
N. E. Newton-Fisher
Department of Anthropology,
Marlowe Building,
University of Kent,
Canterbury, CT2 7NR,
UK
H. O’Regan
School of Biological and Earth
Sciences,
Liverpool John Moores University,
Liverpool L3 3AF,
UK
M. Ohl
Humboldt-Universitat zu Berlin,
Museum fur Naturkunde,
D‐10115 Berlin,
Invalidenstraße 43,
Germany
B. Pawłowski
Department of Anthropology,
University of Wrocław,
ul. Kuznica 35,
Wrocław 50‐138,Poland
U. Radespiel
Institut fur Zoologie,
Tierarztliche Hochschule Hannover,
Bunteweg 17,
D-30559 Hannover,
Germany
T. C. Rae
Department of Anthropology,
University of Durham,
43 Old Elvet, Durham,
DH1 3HN,
UK
D. T. Rasmussen
Department of Anthropology,
CB 1114, Washington University,
One Brookings Drive,
St. Louis, Missouri 63130,
USA
J. H. Relethford
Department of Anthropology,
SUNY College at Oneonta,
Oneonta, NY 13820,
USA
G. J. Retallack
Department of Geology,
University of Oregon,

Contributors 2079
Eugene, OR 97403‐1272,USA
O. Rieppel
Department of Geology,
The Field Museum,
1400 S. Lake Shore Drive,
Chicago, IL 60605‐2496,USA
G. P. Rightmire
Department of Anthropology,
Peabody Museum,
Harvard University,
Cambridge,
MA 02138,
USA
E. J. Sargis
Department of Anthropology,
Yale University,
P.O. Box 208277,
New Haven, CT 06520,
USA
K. Schick
Stone Age Institute,
1392 W. Dittemore Road,
Gosport, IN 47433,
USA
F. Schrenk
Forschungsinstitut und
Naturmuseum,
Abt. Palaoanthropologie und
Quartarpalaontologie,
Senckenberganlage 25,
D-60325 Frankfurt am Main,
Germany
J. H. Schwartz
Department of Anthropology,
3302 WWPH,
University of Pittsburgh,
Pittsburgh, PA 15260,
USA
G. R. Scott
Department of Anthropology,
University of Nevada,
Reno, NV 89557,
USA
B. Senut
Laboratoire de Paleontologie,
Museum National d’Histoire
Naturelle & UMR 8569,
CNRS,
8, rue Buffon,
75005 Paris,
France
M. T. Silcox
University of Winnipeg,
Department of Anthropology,
515 Portage Ave.,
Winnipeg, Manitoba R3B 2E9,
Canada
M. Sponheimer
Department of Anthropology,
University of Colorado at Boulder,
Boulder, CO 80309,
USA
F. Spoor
Department of Anatomy &
Developmental Biology,

2080 Contributors
University College London,
Gower St.,
London WC1E 6BT,
UK
J. Steinhauser
Theoretical Biology Group,
Centre for Ecological and
Evolutionary Studies,
University of Groningen,
P.O. Box 14,
9750 AA Haren,
The Netherlands
D. Strait
Department of Anthropology,
University at Albany,
1400 Washington Avenue,
Albany, New York 12222,
USA
R. W. Sussman
Department of Anthropology,
Washington University at St. Louis,
St. Louis, MO 63130,
USA
C. C. Swisher III
Department of Geological Sciences,
Rutgers University,
610 Taylor Road,
Piscataway,
USA
I. Tattersall
Division of Anthropology,
American Museum of Natural
History,
New York, NY 10024‐5192,USA
M. F. Teaford
Center for Functional Anatomy and
Evolution,
Johns Hopkins University School of
Medicine,
1830 E. Monument St., Room 303,
Baltimore, MD 21205,
USA
A. R. Templeton
Department of Biology,
Washington University in
St. Louis,
1 Brookings, Campus Box 1137,
St. Louis, MO 63130,
USA
N. Toth
Stone Age Institute,
1392 W. Dittemore Road,
Gosport, IN 47433,
USA
L. A. Tsuji
Museum fur Naturkunde,
Humboldt-Universitat zu Berlin,
D‐10099 Berlin,
Germany
A. Turner
School of Biological and
Earth Sciences,
Liverpool John Moores University,
Liverpool L3 3AF,
UK
C. G. Turner II
School of Human Evolution and
Social Change,
Arizona State University,

Contributors 2081
Tempe, AZ 85287‐2402,USA
L. Ulhaas
Forschungsinstitut und
Naturmuseum,
Abt. Palaoanthropologie und
Quartarpalaontologie,
Senckenberganlage 25,
D-60325 Frankfurt am Main,
Germany
P. S. Ungar
Department of Anthropology,
University of Arkansas,
Old Main 330,
Fayetteville, AR 72701,
USA
E. S. Vrba
Department of Geology and
Geophysics,
Yale University,
New Haven, CT 06520,
USA
G. A. Wagner
Geographisches Institut,
Universitat Heidelberg,
Im Neuenheimer Feld 348,
D‐69120 Heidelberg,
Germany
C. V. Ward
Department of Pathology and
Anatomical Sciences,
M263 Medical Sciences Building,
University of Missouri,
One Hospital Drive,
Columbia, MO 65212,
USA
M. Weingarten
Institut fur Philosophie,
Philipps-Universitat Marburg,
Wilhelm-Ropke-Strasse 6, Block B,
D‐35032 Marburg,
Germany
B. Wood
Center for the Advanced Study of
Human Paleobiology, Department of
Anthropology,
The George Washington University,
2110 G St. NW,
Washington, DC 20052,
USA
E. Zimmermann
Institut fur Zoologie,
Tierarztliche Hochschule
Hannover,
Bunteweg 17,
D-30559 Hannover,
Germany
H. Zischler
Institut fur Anthropologie (1050),
Fachbereich 10 – Biologie,
Johannes Gutenberg-Universitat
Mainz,
D‐55099 Mainz,
Germany

9 The Earliest Putative HomoFossilsFriedemann Schrenk . Ottmar Kullmer . Timothy Bromage
Abstract
The earliest fossil remains of the genus Homo have been discovered in eastern,
southeastern, and southern Africa. The sample comprises about 200 skeletal
fragments attributable to about 40 individuals and assigned to two species:
Homo rudolfensis (2.5–1.8 Ma) showing a combination of primitive dentition
with Homo‐like locomotion andHomo habilis (2.1–1.5 Ma) exhibiting a progres-
sive reduction of tooth roots but resembling great apes rather than humans in
the postcranial skeleton. Another significant difference between early Homo and
the australopithecines is the brain size, which was larger in early Homo than
Australopithecus but smaller than in Homo erectus. Endocasts of H. habilis from
Olduvai Gorge and Koobi Fora reveal a number of distinctive features some of
which are recognized as Homo autapomorphies. Differences in tooth wear
betweenH. rudolfensis, with megadont teeth and more horizontal tooth abrasion,
and H. habilis, with more gracile molars and higher relief in worn teeth indicate
significant differences in diet and ecology of early Homo species. The origin of
the genusHomo coincided with the onset of material culture. Between ca. 2.8 and
2.5 Ma, extensive open habitats comprising more arid‐tolerant vegetation devel-
oped in Africa. The selective pressures of this habitat change resulted in the
increased survival of more megadont species varieties. Megadonty allowed these
species to feed on tougher fruit and open woodland–open savannah food items
resulting in the phyletic splitting of Australopithecus afarensis into Paranthropus
and Homo lineages by ca. 2.5 Ma. An evolutionary scenario that complies with
both the Habitat Theory and early hominid biogeography is provided. It deline-
ates the association between faunal turnover and climate change, and suggests a
single origin for the Paranthropus lineage but separate origins for H. rudolfensis
and H. habilis from A. afarensis and A. africanus ancestors, respectively.
# Springer-Verlag Berlin Heidelberg 2007

1612 9 The earliest putative Homo fossils
9.1 Introduction
The search for the roots of the genus Homo is of particular interest in the field of
paleoanthroplogy. The taxonomic determination of the earliest putative Homo
fossils provides the basis for the definition of the taxonHomo to which all modern
humans belong.
Carolus Linnaeus (1758) established the genus Homo in the tenth revision of
his Systema Naturae. In his opinion, Homo subsumed six groups: H. sylvestris
H. troglodytes—a mixture of orangutan and myths, H. sapiens, and four geo-
graphical variants from Africa, America, Asia, and Europe. In the two centuries
that followed H. neanderthalensis (King 1864), H. heidelbergensis (Schoetensack
1908), H. erectus (Dubois 1892; Mayr 1944), H. habilis (Leakey et al. 1964),
H. ergaster (Groves and Mazak 1975), H. rudolfensis (Alexeev 1986), H. antecessor
(Bermudez de Castro et al. 1997), and others were included in the genus Homo.
The history of research, the order of discoveries, and existing paradigms
heavily influence the formation and change in interpretations of human evolu-
tion. This holds true especially regarding ideas on the origin of the genus Homo
since the 1960s. Debates on the attribution of fossil specimens and the definition
of the genus Homo continue today (Wood 1992).
Opinions differ regarding the number of species and also the specimens
included in the genus. Some even assign all putative Homo specimens to living
humans (H. sapiens). Species names in paleoanthropology are labels rather than
natural species, and the taxonomic determination of fossils is more or less a
question of the philosophy followed by the authors. There are both theoretical
and practical reasons to erect taxa, as chronospecies for time equivalent appear-
ance or morphospecies for a complex of shared anatomical features, and in the
worst case there are political reasons for the allocation of species.
9.2 Fossil evidence
Early research on the origin of the genus Homo is closely related to the African
fieldwork of Louis Leakey (1903–1972). He strongly believed in Africa as
the cradle of humankind and in 1932 discovered the first evidence of early
Homo at Kanam, (Kenya) east of Lake Victoria—a specimen, which today is
attributed to H. erectus. He also undertook archeological surveys in Olduvai
Gorge, Tanzania (> Figure 9.1), where later he discovered early pebble tools in
Bed I (ca. 1.8 Ma)—remains of what he termed the ‘‘Oldowan’’ industry. The
search for the artifacts creator led to the discovery of robust australopithecine
remains(Zinjanthropus boisei) (Leakey 1959). However, due to its small brain size,
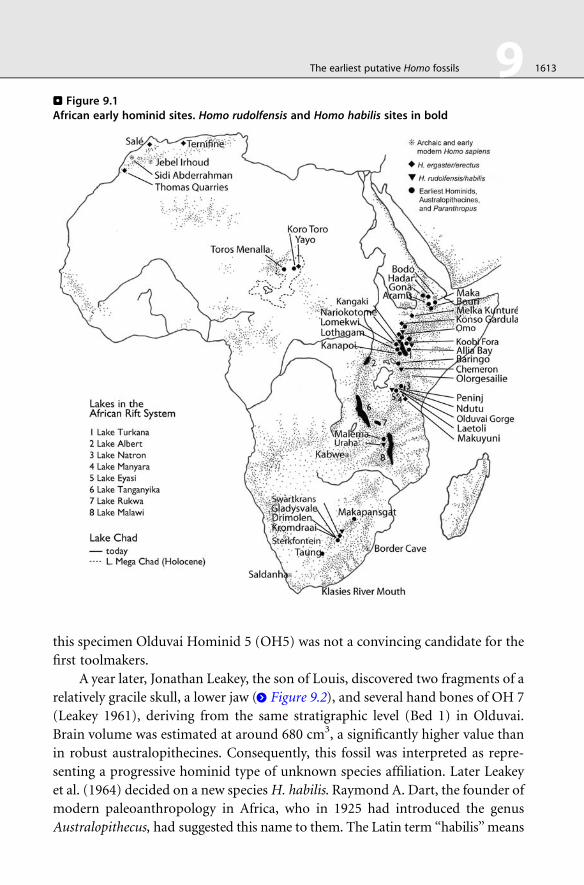
. Figure 9.1African early hominid sites. Homo rudolfensis and Homo habilis sites in bold
The earliest putative Homo fossils 9 1613
this specimen Olduvai Hominid 5 (OH5) was not a convincing candidate for the
first toolmakers.
A year later, Jonathan Leakey, the son of Louis, discovered two fragments of a
relatively gracile skull, a lower jaw (> Figure 9.2), and several hand bones of OH 7
(Leakey 1961), deriving from the same stratigraphic level (Bed 1) in Olduvai.
Brain volume was estimated at around 680 cm3, a significantly higher value than
in robust australopithecines. Consequently, this fossil was interpreted as repre-
senting a progressive hominid type of unknown species affiliation. Later Leakey
et al. (1964) decided on a new speciesH. habilis. Raymond A. Dart, the founder of
modern paleoanthropology in Africa, who in 1925 had introduced the genus
Australopithecus, had suggested this name to them. The Latin term ‘‘habilis’’ means

. Figure 9.2Type specimen of Homo habilis: mandible OH 7, Olduvai Gorge, Tansania
1614 9 The earliest putative Homo fossils
‘‘handy, skillful, able’’: finally, the producer of the Oldowan culture seemed to have
been identified.
Apart from OH 7, the species description of H. habilis included skull frag-
ments and teeth (OH 4 and OH 6), part of an adult foot (OH 8), and the
incomplete skull of an adolescent (OH 13). Further, Leakey et al. (1964) referred
a collection of juvenile cranial pieces (OH 14) and the fragmented cranial vault
and dentition (OH 16) of a young adult to the new species.
Since then, numerous additional fossils of H. habilis have been discovered at
Olduvai Gorge: 9 fragmentary skulls, 4 mandible fragments, 19 teeth, and 8 post-
cranial fragments. Among these fragments was the squashed skull OH 24
(Twiggy), which was found in 1968. In 1986, a partial female skeleton (OH 62)
was assembled from a number of fragments (Johanson et al. 1987). This specimen
showed that H. habilis was fully bipedal and had a brain larger than all australo-
pithecines. For many years, the H. habilis remains from Olduvai Gorge were seen
as the most important early Homo specimens and consequently played the
leading role in most hypotheses regarding the origin of genus Homo.

The earliest putative Homo fossils 9 1615
Leakey et al. (1964) originally discussed cranial and mandibular traits, to
distinguish the Homo specimens of Olduvai from Australopithecines and
H. erectus. Maxillary and mandibular size is smaller than in Australopithecus
and tends in size to H. erectus and H. sapiens. The surface of the skull shows
slight to strong muscular markings and the parietal curvature in the sagittal plane
varies from slight to moderate. The frontal bone is more vertical and the torus
supraorbitalis is less developed than in australopithecines. In the occipital region,
the relatively open‐angled external sagittal curvature differs markedly from
Australopithecus.
In 1970 the picture of earliest Homo began to change significantly with the
success of the Koobi Fora Research Project in northern Kenya and led by Richard
Leakey, Louis’ son. In just a few years on the eastern shores of Lake Turkana, his
team recovered 9 skulls, 10 mandibles, 6 isolated teeth, and 5 postcranial frag-
ments (Leakey 1973a,b). Originally, all of the early Homo finds from East
Turkana, with an age similar to those from Olduvai (1.9–1.8 Ma), were inter-
preted as bearing similarities to H. habilis—then the only early species of Homo.
One cranial fragment from the Nachukui Formation on the western shores of
Lake Turkana was also assigned to H. habilis. However, two of the best preserved
skulls from Koobi Fora (KNM‐ER1470,> Figure 9.4; KNM‐ER 1813, > Figure 9.5)
later gave rise to an extended debate among researchers about the heterogeneity
of the H. habilis hypodigm, and finally led to the recognition of a new species,
H. rudolfensis.
In the 1970s a large number of isolatedHomo teeth were discovered north of
Koobi Fora, near the Omo‐River in southern Ethiopia, in Member G and H of the
Shungura Formation. From these it became clear that the origin of the genus
Homo extended well beyond 2 Ma. Already in 1965, a temporal bone had been
discovered by John Martyn at Chemero, Kenya, which nearly two decades later
was described by Hill et al. (1992) as a very early member of the genus Homo,
dated to around 2.4 Ma.
In 1976, an early Homo fossil was found at Sterkfontein, South Africa
(Stw 53), which belonged neither to H. erectus nor to Australopithecus (Hughes
and Tobias 1977). A partial facial skull (SK 847 from Swartkrans), assembled from
several fragments, originally attributed to a different species, is further evidence
for H. habilis, which probably migrated into southern Africa around 2 Ma (see
below).
The geographical gap between the southern and eastern African early homi-
nid sites was filled in the early 1990s through discoveries in the ‘‘Hominid
Corridor’’ of the northernMalawi Rift (Schrenk et al. 1993; Bromage and Schrenk
1995). In 1992, at Uraha, the Hominid Corridor Research Project (HCRP) has
1616 9 The earliest putative Homo fossils
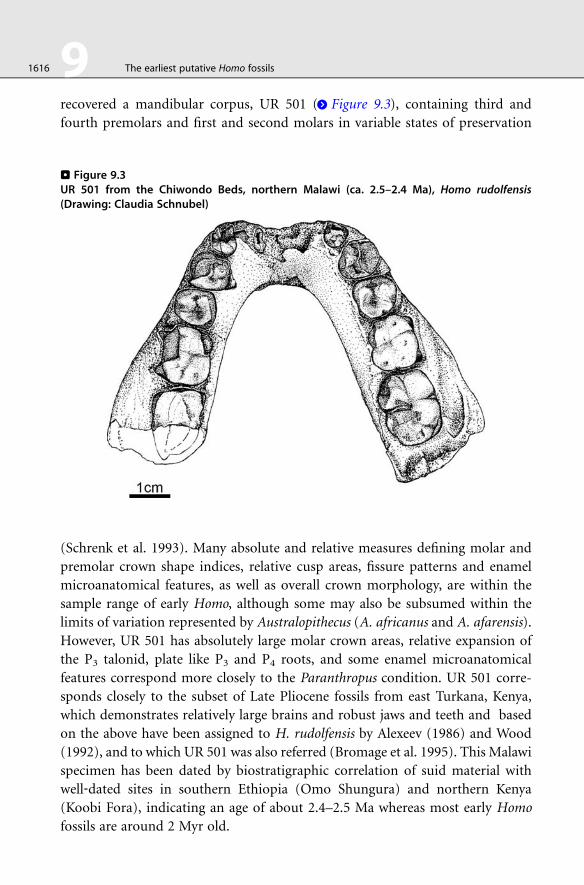
recovered a mandibular corpus, UR 501 (> Figure 9.3), containing third and
fourth premolars and first and second molars in variable states of preservation
. Figure 9.3UR 501 from the Chiwondo Beds, northern Malawi (ca. 2.5–2.4 Ma), Homo rudolfensis(Drawing: Claudia Schnubel)
(Schrenk et al. 1993). Many absolute and relative measures defining molar and
premolar crown shape indices, relative cusp areas, fissure patterns and enamel
microanatomical features, as well as overall crown morphology, are within the
sample range of early Homo, although some may also be subsumed within the
limits of variation represented by Australopithecus (A. africanus and A. afarensis).
However, UR 501 has absolutely large molar crown areas, relative expansion of
the P3 talonid, plate like P3 and P4 roots, and some enamel microanatomical
features correspond more closely to the Paranthropus condition. UR 501 corre-
sponds closely to the subset of Late Pliocene fossils from east Turkana, Kenya,
which demonstrates relatively large brains and robust jaws and teeth and based
on the above have been assigned to H. rudolfensis by Alexeev (1986) and Wood
(1992), and to which UR 501 was also referred (Bromage et al. 1995). This Malawi
specimen has been dated by biostratigraphic correlation of suid material with
well‐dated sites in southern Ethiopia (Omo Shungura) and northern Kenya
(Koobi Fora), indicating an age of about 2.4–2.5 Ma whereas most early Homo
fossils are around 2 Myr old.

The earliest putative Homo fossils 9 1617
It is important to note that the appearance of earliest Homo is contempora-
neous with the origin of hyperrobust australopithecines (Paranthropus). The
earliest evidence for this co‐existence is based on further hominid discoveries in
the Chiwondo Beds of Northern Malawi. A maxillary fragment (RC 911) pre-
serves part of the left alveolar process, with badly damaged M1 crown and
fragmentary M2 crown. Size, morphology and abrasion occlusal pattern on the
surface suggest that RC 911 should be assigned to Paranthropus boisei (Kullmer
et al. 1999).
The biogeographic significance of these Malawi Rift hominids lay in their
association with the eastern African endemic faunal group. The associated bovid
and suid faunas show a small amount of overlap with southern African animals
and a greater overlap with eastern African faunal elements. Biogeographic varia-
tion, in the Malawi Rift may be linked to habitat change occurring due to climate
shifts, with maximum change occurring around 2.5 Ma.
9.3 Changing taxonomy
Hominid fossils are generally assigned to the genus Homo if they fulfill four main
criteria (Keith 1948; Tobias 1991; Wood and Collard 2001): a brain size above
600 cm3, ability for speech and tool‐making, as well as an opposable pollux. To
date, the hypodigm of earliest Homo attributed to H. habilis sensu stricto and
H. rudolfensis contains about 200 skeletal fragments attributable to about 40 indi-
viduals (> Tables 9.1 and > 9.2). Despite—or even due to—the large number of
specimens, the taxonomic interpretation of earliest Homo is highly controversial
(Wood 2000).
Originally, the interpretation of the early Homo hand as ‘‘modern’’ (Leakey
et al. 1964) supported the view of H. habilis as an early but ‘‘able’’ human as
opposed to the rather ‘‘clumsy’’ australopithecines. However, later skeletal finds
at Olduvai Gorge (OH 62) (Johanson et al. 1987) demonstrated that the postcra-
nial skeleton ofH. habilis indeed resembled Australopithecus africanus rather than
Homo. Yet the most distinctive character of H. habilis remains its relatively and
absolutely higher brain volume compared to that of Australopithecus. The fore-
head of H. habilis is more vertical and a weak supraorbital torus is present.
Whereas the morphological characters are quite uniform in the Olduvai sample,
the discussion started to heat up mainly over two very distinct skull fragments
from Koobi Fora: KNM‐ER 1470 (> Figure 9.4) (Leakey 1973a) and KNM‐ER1813 (> Figure 9.5) (Leakey 1973b).
In a comprehensive character analysis of all available putative H. habilis
fossils from Koobi Fora, Wood (1991) concluded that the variability exhibited
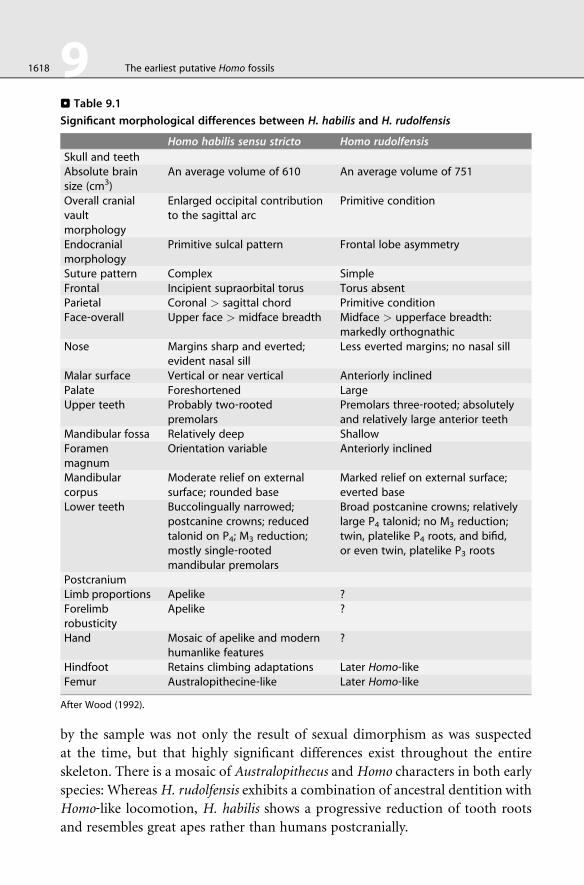
. Table 9.1
Significant morphological differences between H. habilis and H. rudolfensis
Homo habilis sensu stricto Homo rudolfensis
Skull and teethAbsolute brainsize (cm3)
An average volume of 610 An average volume of 751
Overall cranialvaultmorphology
Enlarged occipital contributionto the sagittal arc
Primitive condition
Endocranialmorphology
Primitive sulcal pattern Frontal lobe asymmetry
Suture pattern Complex SimpleFrontal Incipient supraorbital torus Torus absentParietal Coronal > sagittal chord Primitive conditionFace‐overall Upper face > midface breadth Midface > upperface breadth:
markedly orthognathicNose Margins sharp and everted;
evident nasal sillLess everted margins; no nasal sill
Malar surface Vertical or near vertical Anteriorly inclinedPalate Foreshortened LargeUpper teeth Probably two‐rooted
premolarsPremolars three‐rooted; absolutelyand relatively large anterior teeth
Mandibular fossa Relatively deep ShallowForamenmagnum
Orientation variable Anteriorly inclined
Mandibularcorpus
Moderate relief on externalsurface; rounded base
Marked relief on external surface;everted base
Lower teeth Buccolingually narrowed;postcanine crowns; reducedtalonid on P4; M3 reduction;mostly single‐rootedmandibular premolars
Broad postcanine crowns; relativelylarge P4 talonid; no M3 reduction;twin, platelike P4 roots, and bifid,or even twin, platelike P3 roots
PostcraniumLimb proportions Apelike ?Forelimbrobusticity
Apelike ?
Hand Mosaic of apelike and modernhumanlike features
?
Hindfoot Retains climbing adaptations Later Homo‐likeFemur Australopithecine‐like Later Homo‐like
After Wood (1992).
1618 9 The earliest putative Homo fossils
by the sample was not only the result of sexual dimorphism as was suspected
at the time, but that highly significant differences exist throughout the entire
skeleton. There is a mosaic of Australopithecus and Homo characters in both early
species: WhereasH. rudolfensis exhibits a combination of ancestral dentition with
Homo‐like locomotion, H. habilis shows a progressive reduction of tooth roots
and resembles great apes rather than humans postcranially.
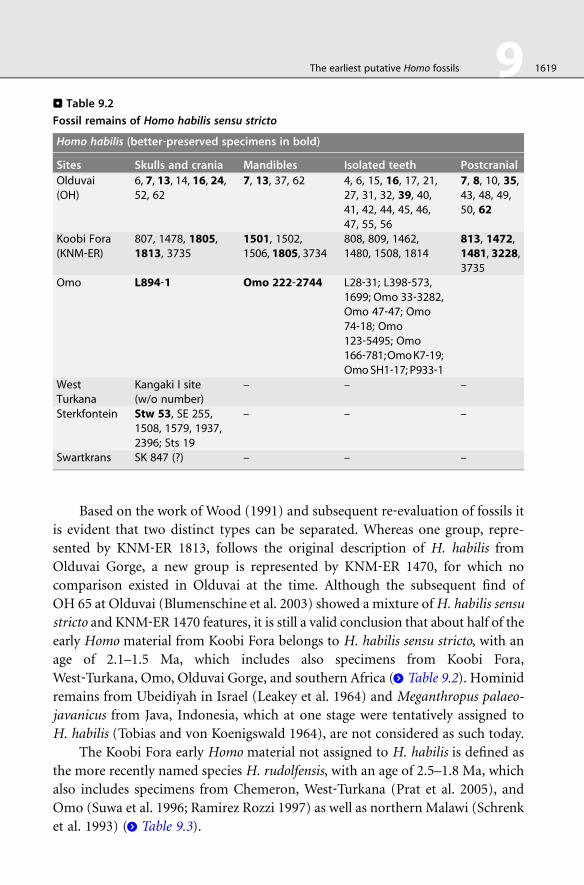
. Table 9.2
Fossil remains of Homo habilis sensu stricto
Homo habilis (better‐preserved specimens in bold)
Sites Skulls and crania Mandibles Isolated teeth Postcranial
Olduvai(OH)
6, 7, 13, 14, 16, 24,52, 62
7, 13, 37, 62 4, 6, 15, 16, 17, 21,27, 31, 32, 39, 40,41, 42, 44, 45, 46,47, 55, 56
7, 8, 10, 35,43, 48, 49,50, 62
Koobi Fora(KNM‐ER)
807, 1478, 1805,1813, 3735
1501, 1502,1506, 1805, 3734
808, 809, 1462,1480, 1508, 1814
813, 1472,1481, 3228,3735
Omo L894‐1 Omo 222‐2744 L28‐31; L398‐573,1699; Omo 33‐3282,Omo 47‐47; Omo74‐18; Omo123‐5495; Omo166‐781;OmoK7‐19;OmoSH1‐17; P933‐1
WestTurkana
Kangaki I site(w/o number)
– – –
Sterkfontein Stw 53, SE 255,1508, 1579, 1937,2396; Sts 19
– – –
Swartkrans SK 847 (?) – – –
The earliest putative Homo fossils 9 1619
Based on the work of Wood (1991) and subsequent re‐evaluation of fossils it
is evident that two distinct types can be separated. Whereas one group, repre-
sented by KNM‐ER 1813, follows the original description of H. habilis from
Olduvai Gorge, a new group is represented by KNM‐ER 1470, for which no
comparison existed in Olduvai at the time. Although the subsequent find of
OH 65 at Olduvai (Blumenschine et al. 2003) showed a mixture ofH. habilis sensu
stricto and KNM‐ER 1470 features, it is still a valid conclusion that about half of the
early Homo material from Koobi Fora belongs to H. habilis sensu stricto, with an
age of 2.1–1.5 Ma, which includes also specimens from Koobi Fora,
West‐Turkana, Omo, Olduvai Gorge, and southern Africa (> Table 9.2). Hominid
remains from Ubeidiyah in Israel (Leakey et al. 1964) and Meganthropus palaeo-
javanicus from Java, Indonesia, which at one stage were tentatively assigned to
H. habilis (Tobias and von Koenigswald 1964), are not considered as such today.
The Koobi Fora early Homo material not assigned to H. habilis is defined as
the more recently named species H. rudolfensis, with an age of 2.5–1.8 Ma, which
also includes specimens from Chemeron, West‐Turkana (Prat et al. 2005), and
Omo (Suwa et al. 1996; Ramirez Rozzi 1997) as well as northern Malawi (Schrenk
et al. 1993) (> Table 9.3).
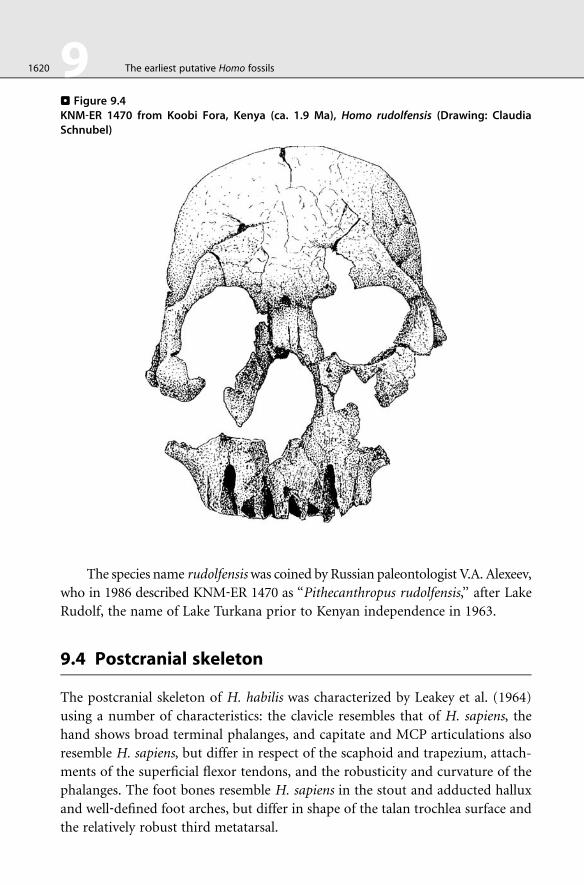
. Figure 9.4KNM‐ER 1470 from Koobi Fora, Kenya (ca. 1.9 Ma), Homo rudolfensis (Drawing: ClaudiaSchnubel)
1620 9 The earliest putative Homo fossils
The species name rudolfensiswas coined by Russian paleontologist V.A. Alexeev,
who in 1986 described KNM‐ER 1470 as ‘‘Pithecanthropus rudolfensis,’’ after Lake
Rudolf, the name of Lake Turkana prior to Kenyan independence in 1963.
9.4 Postcranial skeleton
The postcranial skeleton of H. habilis was characterized by Leakey et al. (1964)
using a number of characteristics: the clavicle resembles that of H. sapiens, the
hand shows broad terminal phalanges, and capitate and MCP articulations also
resemble H. sapiens, but differ in respect of the scaphoid and trapezium, attach-
ments of the superficial flexor tendons, and the robusticity and curvature of the
phalanges. The foot bones resemble H. sapiens in the stout and adducted hallux
and well‐defined foot arches, but differ in shape of the talan trochlea surface and
the relatively robust third metatarsal.
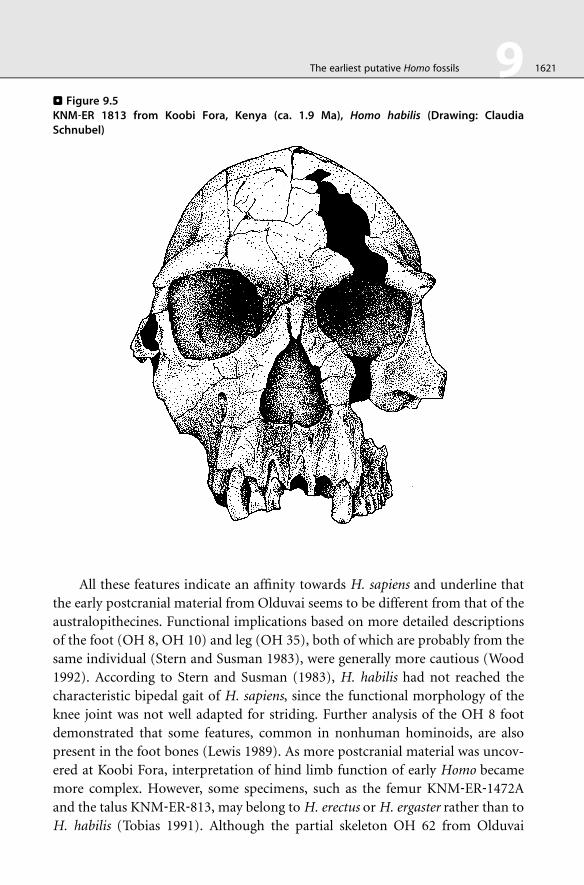
. Figure 9.5KNM‐ER 1813 from Koobi Fora, Kenya (ca. 1.9 Ma), Homo habilis (Drawing: ClaudiaSchnubel)
The earliest putative Homo fossils 9 1621
All these features indicate an affinity towards H. sapiens and underline that
the early postcranial material from Olduvai seems to be different from that of the
australopithecines. Functional implications based on more detailed descriptions
of the foot (OH 8, OH 10) and leg (OH 35), both of which are probably from the
same individual (Stern and Susman 1983), were generally more cautious (Wood
1992). According to Stern and Susman (1983), H. habilis had not reached the
characteristic bipedal gait of H. sapiens, since the functional morphology of the
knee joint was not well adapted for striding. Further analysis of the OH 8 foot
demonstrated that some features, common in nonhuman hominoids, are also
present in the foot bones (Lewis 1989). As more postcranial material was uncov-
ered at Koobi Fora, interpretation of hind limb function of early Homo became
more complex. However, some specimens, such as the femur KNM‐ER‐1472Aand the talus KNM‐ER‐813, may belong toH. erectus orH. ergaster rather than to
H. habilis (Tobias 1991). Although the partial skeleton OH 62 from Olduvai
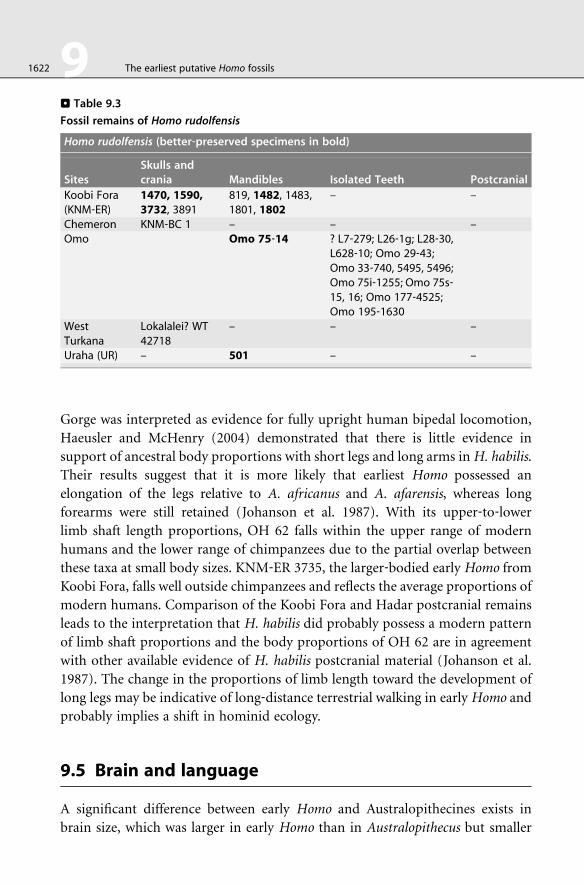
. Table 9.3
Fossil remains of Homo rudolfensis
Homo rudolfensis (better‐preserved specimens in bold)
SitesSkulls andcrania Mandibles Isolated Teeth Postcranial
Koobi Fora(KNM‐ER)
1470, 1590,3732, 3891
819, 1482, 1483,1801, 1802
– –
Chemeron KNM‐BC 1 – – –Omo Omo 75‐14 ? L7‐279; L26‐1g; L28‐30,
L628‐10; Omo 29‐43;Omo 33‐740, 5495, 5496;Omo 75i‐1255; Omo 75s‐15, 16; Omo 177‐4525;Omo 195‐1630
WestTurkana
Lokalalei? WT42718
– – –
Uraha (UR) – 501 – –
1622 9 The earliest putative Homo fossils
Gorge was interpreted as evidence for fully upright human bipedal locomotion,
Haeusler and McHenry (2004) demonstrated that there is little evidence in
support of ancestral body proportions with short legs and long arms inH. habilis.
Their results suggest that it is more likely that earliest Homo possessed an
elongation of the legs relative to A. africanus and A. afarensis, whereas long
forearms were still retained (Johanson et al. 1987). With its upper‐to‐lowerlimb shaft length proportions, OH 62 falls within the upper range of modern
humans and the lower range of chimpanzees due to the partial overlap between
these taxa at small body sizes. KNM‐ER 3735, the larger‐bodied early Homo from
Koobi Fora, falls well outside chimpanzees and reflects the average proportions of
modern humans. Comparison of the Koobi Fora and Hadar postcranial remains
leads to the interpretation that H. habilis did probably possess a modern pattern
of limb shaft proportions and the body proportions of OH 62 are in agreement
with other available evidence of H. habilis postcranial material (Johanson et al.
1987). The change in the proportions of limb length toward the development of
long legs may be indicative of long‐distance terrestrial walking in early Homo and
probably implies a shift in hominid ecology.
9.5 Brain and language
A significant difference between early Homo and Australopithecines exists in
brain size, which was larger in early Homo than in Australopithecus but smaller

The earliest putative Homo fossils 9 1623
than H. erectus. Endocasts of H. habilis from Olduvai and Koobi Fora reveal a
number of distinctive features, some of which are recognized as autapomorphies
of the genus Homo. Tobias (1987) defined the principal morphological trait to
distinguishH. habilis from Australopithecus as a larger mean endocranial capacity
in the former (640 cm3) than in A. africanus (441 cm3), A. boisei (513 cm3), and
A. robustus (530 cm3). This suggests that the evolutionary trend toward brain
expansion was already well under way more than 2 Ma. The H. habilis mean
(640 cm3) is close to the lower limit (647 cm3) of the 95% population range of
H. erectus but well above the upper limit of the A. africanus range (492 cm3). The
brain capacity of the H. rudolfensis type specimen (750 cm3) is larger than the
known range for H. habilis from Olduvai Gorge and Koobi Fora and falls within
the lower range of H. erectus.
A prominent feature of the H. habilis brain is the bilateral transverse expan-
sion of the cerebrum, especially in the frontal and parieto‐occipital areas, and a
posterior heightening. The increased bulk of cerebral frontal and parietal lobes
and the sulcal and gyral pattern of the lateral frontal lobe have been interpreted as
derived features for the genusHomo (Tobias 1987). TheH. habilis brain showed a
well‐developed left superior parietal lobule, and a prominent development of the
inferior parietal lobule.
The endocast of KNM‐ER 1470 shows a left frontal lobe sulcal pattern that is
associated with Broca’s area in living people (Falk 1987), a finding that has led to
the conclusion that H. rudolfensismay have been capable speech. This conclusion
is in accordance with Holloway’s observation of a pronounced left‐occipital‐right‐frontal petalia pattern in the KNM‐ER‐1470 endocast that may indicate
functional cortical asymmetry (Holloway 1983). Surprising corroborative evi-
dence has been provided by Toth’s analyses of stone flakes, which indicate that
hominids were predominantly right‐handed by 2 Ma (Toth 1985). Adjacent areas
in the left frontal lobe control the speech organs and the right hand. Tobias (1991)
stated that H. habilis is the earliest hominid to show prominent enlargement of
Broca’s and Wernicke’s areas. If so, the same should be seen in H. rudolfensis.
Australopithecus endocasts show Broca’s area, but not Wernicke’s region,
while anthropoid apes display neither of the two. The prominent develop-
ment of the two speech areas may thus be seen as an important autapomor-
phy of the genus Homo (Tobias 1991). Even if H. habilis and H. rudolfensis
possessed the neurological basis of speech, there is no evidence that either of them
used spoken language. The areas of the brain that control spoken communication
probably manifested themselves only when brain enlargement occurred and
marked encephalization started.

1624 9 The earliest putative Homo fossils
9.6 Material culture and food processing
Around 2.5 Ma, simultaneously with an increase in drier and harder food stuffs
due to increasing aridity in eastern Africa (de Menocal 2004), there occurred
the first hyperrobust australopithecines (Paranthropus) and the genus Homo in
the fossil record (Bromage et al. 1995). This demonstrates an evolutionary
alternative to the massive masticationary system of Paranthropus, which was
capable of dealing with very tough foods. The evolutionary alternative to mega-
donty was the manufacturing and use of stone tools. The oldest chopper tools
are known from Ethiopia (Hata, Bouri Formation) and from Tanzania, approxi-
mately 2.5 Ma (Kaiser et al. 1995). From Gona, east of Hadar in the Afar Triangle,
primitive pebble tools are dated to 2.6 Ma (Harris 1986). New discoveries from
west of Lake Turkana confirm the existence of an early tool culture around 2.5 Ma
(Roche et al. 2003). In the Hadar area of Ethiopia, stone tools were found
associated with early Homo remains (Kimbel et al. 1996). At many sites the
presence of more than one hominid species, occurring in the same horizon as
early Oldowan pebbles does not give clear evidence of who were the first tool-
makers. However, the distinct specialization of the skull and dental morphology
in robust australopithecines and brain expansion in earlyHomo point to the latter
as the most likely tool manufacturer.
Implements are widely used by higher primates. Yet during marked habitat
shifts, which led to pronounced changes in food resources, it probably was the
invention of stone tools, which supported the origin of the genus Homo around
2.5 Ma. Increasing independence from the environment led to an increase in the
dependence on culture.
If early Homo utilized stone tools to prepare food, the dentition might
actually indicate these behavioral changes in food acquisition. However, the
morphology of early Homo teeth does not seem to suggest extensive food
preparation before ingestion. The incisors are large compared to those of Aus-
tralopithecus and H. erectus, and the canines are large relative to the premolar
crown surfaces. The premolars are narrower than in Australopithecus and fall
within the range of H. erectus. Molar size overlaps the ranges for Australopithecus
and H. erectus. The cheek teeth of H. rudolfensis are enlarged and show affinities
to Paranthropus molars. All teeth are relatively narrow buccolingually and elon-
gated mesiodistally, especially the mandibular molars and premolars. InH. habilis
we see well‐developed third molars, while in H. rudolfensis the third molar forms
a smaller crown than the second molar. The occlusal surface of the cheek teeth is
not as broad as in australopithecine molars and indicates differences in chewing.
Tobias (1987) states that the crown’s cusps relief is still present even when the
teeth are in advanced wear and dentine is visible. This means that the attrition of

The earliest putative Homo fossils 9 1625
enamel is less pronounced than in earlier hominids. Differences in tooth wear
between H. rudolfensis, with megadont teeth and a more horizontal tooth abra-
sion, and H. habilis, with its more gracile molars and higher relief in worn teeth
are clearly visible. This indicates significant differences in diet and ecology of early
H. species. H. rudolfensis and the robust australopithecines share some cranial
and dental features regarding the morphology of the masticatory apparatus
(Wood 1992), which indicates that these hominids were able to cope with
tough fruits and plants. Since those features are judged as an adaptation to
drier climatic conditions, they also show that H. rudolfensis was relatively conser-
vative nutritionally and probably followed a more herbivorous strategy in food
acquisition.
9.7 Biogeographic scenario
The scenario presented here is derived mainly from hypotheses about early
hominid evolution in the context of environmental change and faunal biogeog-
raphy. It is thus a biogeographic perspective against which we understand the
relevance to studies of early Homo systematics in general, and morphology and
character transformation more specifically.
The behavioral inclination of the earliest hominids distributed along the
margin of the tropical rain forest, was to maintain a connection to and remain
near the borders between broad riparian habitats and open woodlands during the
ascendancy of more warm and humid times. Over short geological timescales, this
was typically a local, nondispersing tendency, but by approximately 4 Ma, several
species of Australopithecus had successfully dispersed throughout the reaches of
the African Rift Valley and into western Africa (> Figure 9.7). Over longer time
frames, this included dispersal through the riparian ‘‘corridor’’ connecting
eastern and southern Africa, permitting population dispersal into southern Africa
by 3 Ma. This dispersing population maintained habitat specificities to forested
environments (Rayner et al. 1993), though in more environmentally temperate
climes and in relative geographical isolation at the extreme distal edge of its
distribution. The dispersion along changing latitudinal circumstances covar-
ied with its transformation into, first, a geographic variant and, subsequently,
into A. africanus, joining ranks with other southern African endemic faunas.
Thus, A. afarensis was essentially an eastern African endemic and it follows that
no typical representatives are likely to be recovered from southern African
deposits older than 3.5 Ma. (> Figures 9.6 and > 9.7).
By approximately 2.8 Ma, the initiation of cooler and drier conditions
prevailed upon the African landscape, its vegetation, and its faunas, until
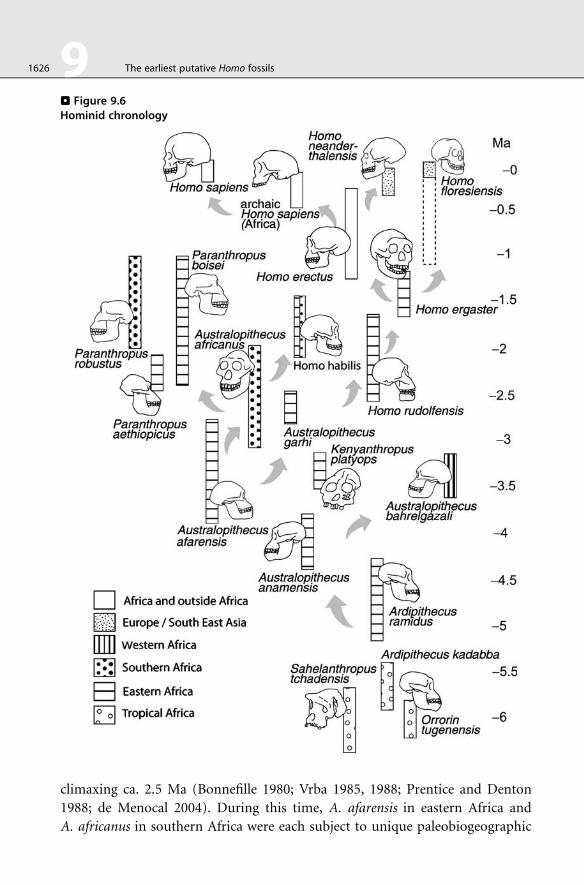
. Figure 9.6Hominid chronology
1626 9 The earliest putative Homo fossils
climaxing ca. 2.5 Ma (Bonnefille 1980; Vrba 1985, 1988; Prentice and Denton
1988; de Menocal 2004). During this time, A. afarensis in eastern Africa and
A. africanus in southern Africa were each subject to unique paleobiogeographic
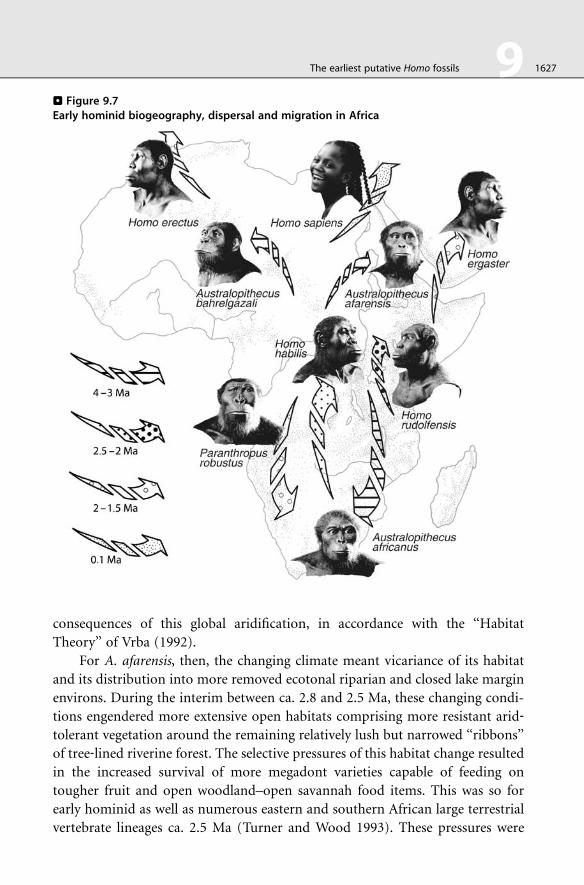
. Figure 9.7Early hominid biogeography, dispersal and migration in Africa
The earliest putative Homo fossils 9 1627
consequences of this global aridification, in accordance with the ‘‘Habitat
Theory’’ of Vrba (1992).
For A. afarensis, then, the changing climate meant vicariance of its habitat
and its distribution into more removed ecotonal riparian and closed lake margin
environs. During the interim between ca. 2.8 and 2.5 Ma, these changing condi-
tions engendered more extensive open habitats comprising more resistant arid‐tolerant vegetation around the remaining relatively lush but narrowed ‘‘ribbons’’
of tree‐lined riverine forest. The selective pressures of this habitat change resulted
in the increased survival of more megadont varieties capable of feeding on
tougher fruit and open woodland–open savannah food items. This was so for
early hominid as well as numerous eastern and southern African large terrestrial
vertebrate lineages ca. 2.5 Ma (Turner and Wood 1993). These pressures were

1628 9 The earliest putative Homo fossils
likewise sufficient to result in the phyletic splitting of A. afarensis into Paranthro-
pus and Homo lineages by ca. 2.5 Ma (Vrba 1988) (> Figures 9.6 and > 9.7).
Ensuing cooler and drier conditions favored a tougher savannah vegetation
composed of plant species better able to retain their moisture under such condi-
tions. Selection favored more facially robust and large molar‐toothed mammals,
including early hominids, capable of efficiently processing the tougher, more
durable vegetation of the savannah. Our evidence suggests that the tropical
equatorial animals, including the hominids, of eastern Africa stayed in the
tropical African ecological domain, while during the drying and cooling of global
climates ca. 2.5 Ma, the southern and more temperate African faunas followed
their northward‐drifting vegetation zones. Thus, Homo and Paranthropus may
have emerged in tropical Africa as a result of the ca. 2.5‐Ma climatic cooling event
and remained there.
The beginnings of the Paranthropus lineage maintained a reliance on fruiting
resources on the riverine side of its ecotone, particularly during the dry season,
but it was equally adept at grinding on the postcanine dentition those food items
it required from more open habitats during more hospitable times of the year.
The beginnings of the Homo lineage, also ca. 2.5 Ma, and represented by
H. rudolfensis, was an endorsement of its recency of common ancestry with
A. afarensis, a distinction it shared with Paranthropus (Bromage et al. 1995).
However, while Paranthropus was principally adapted by means of a robust
masticatory system to its tough and abrasive diet, H. rudolfensis exhibited an
increased behavioral flexibility as its adaptation to climatic circumstances includ-
ed a larger and more provoking, inquiring, and capable brain. This included a
shift to proportionately less abrasive foodstuffs and more omnivorous habits.
Material culture ameliorated the effects of climate change to the degree that it
enabled H. rudolfensis to take advantage of other resources more efficiently than
was ever possible before.
P. boisei and H. rudolfensis remained endemic to tropical latitudes during
this time (Bromage et al. 1995). The eastern African tropical faunas, having
habitable alternatives, remained within their biogeographic domain rather than
brave the relative deterioration and paucity of habitats south of the African Rift
Valley.
The faunas of southern Africa were subject to a different set of environmental
sequelae during the ca. 2.5‐Ma cooling event. Waning of the forests and woodlands
in deference to more open arid grasslands invigorated not only evolutionary
adaptations to savannah life in tropical eastern Africa but also resulted in
the distribution drift northward of faunas tracking the equator ward shift of
grassland and woodland biomes into eastern Africa from the south ca. 2.5 Ma
(Bromage et al. 1995). The temperate zone ca. 2.5 Ma experienced more seasonal

The earliest putative Homo fossils 9 1629
extremes and many organisms unwittingly maintained their inherited preference
for moderate seasonal climes and temperate vegetation physiognomy by moving
northward with the shrinking of this biome toward the equator, effectively
transgressing the Zambezi Ecozone. Among these migrants was A. africanus
who, having been adapted to a modest temperate ecology, now found its suitable
habitats shifted to the north toward the African Rift Valley. While dispersing
toward the eastern African tropical domain, selection for increased behavioral
flexibility was related to the habitat diversity of the tropics and the presence of
other nonvegetative food resources available in their new region. This emerging
taxon, H. habilis, rapidly established itself as a categorical omnivore and found
that it could buffer itself more resolutely from environmental changes. This
enabled it to cross habitat boundaries more easily and also to advantage itself
of more resources with its material culture.
By approximately 2 Ma, Africa was rebounding from its relatively cool and dry
climate to return to slightly more warm and humid conditions (Bromage et al.
1995). A phase of biome expansion ensued that facilitated dispersions away from
the equator, ending nearly 1 Myr of relative endemism dominated by tropical
equatorial speciations. P. boisei dispersed southward along reestablished ecotonal
habitats into southern Africa, varied there as a geographic variant under more
temperate conditions, and evolved into Paranthropus robustus (> Figure 9.6).
H. habilis expanded southward into the southern African temperate domain,
but it maintained a very much broader niche and increased its distributional area
as a single species.H. rudolfensis remained endemic to the eastern African tropical
domain due partly to its preference for more open habitats around the rain
shadows of the African Rift Valley and partly, perhaps, to some small measure
of competitive exclusion from geographic realms occupied by H. habilis.
References
Alexeev VP (1986) The origin of the human
race. Progress Publishers, Moscow
Bermudez de Castro JM, Arsuaga JL, Carbonell
E, Rosas A, Martinez I, Mosqueria M (1997)
A hominid from the Lower Pleistocene of
Atapuerca, Spain possible ancestor to Nean-
derthals and modern humans. Science 276:
1392–1395
Blumenschine RJ, Peters CR, Masao FT, Clarke
RJ, Deino A, Hay RL, et al. (2003) Late Plio-
cene Homo and hominid land use from
western Olduvai Gorge, Tanzania. Science
299: 1217–1221
Bonnefille R (1980) Vegetation history of sa-
vanna in East Africa during the Pleistocene.
Proc IV Int Palyn Conf Lucknow 3: 75–89
Bromage TG, Schrenk F (eds) (1995) Evolu-
tionary history of the Malawi Rift. J Hum
Evol 28: 1–120
Bromage TG, Schrenk F, Zonneveld FW (1995)
Paleoanthropology of the Malawi Rift: an
early hominid mandible from the Chiwondo

1630 9 The earliest putative Homo fossils
Beds, northern Malawi. J Hum Evol 28:
71–108
de Menocal PB (2004) African climate change
and faunal evolution during the Pliocene‐Pleistocene, Earth Planet Sci Lett 220:
3–24
Dubois E (1892) Palaeontologische anderzoe-
kingen op Java. Verslag van het Mijnwezen.
Batavia 3: 10–14
Falk D (1987) Hominid paleoneurology. Annu
Rev Anthropol 16: 13–30
Groves CP, Mazak V (1975) An approach to the
taxonomy of the Hominidae: Gracile Villa-
franchian hominids of Africa. Casopis pro
Mineralogii A Geologii 20: 225–247
Haeusler M, McHenry HM (2004) Body pro-
portions of Homo habilis reviewed. J Hum
Evol 46: 433–465
Harris JWK (1986) Decouverte de materiel
archeologique oldowayen dans le rift de
l’Afar. L’Anthropologie 90: 339–357
Hill A, Ward S, Deino A, Curtis G, Drake R
(1992) The earliest Homo. Nature 355:
719–722
Holloway RL (1983) Human brain evolution: a
search for units, models and synthesis. Can J
Anthropol 3: 215–230
Hughes AR, Tobias PV (1977) A fossil skull
probably of the genus Homo from Sterkfon-
tein, Transvaal. Nature 265: 310–312
Johanson DC, Masao FT, Eck GG, White TD,
Walter RC, Kimbel WH, Asfaw B, Manega P,
Ndessokia P, Suwa G (1987) New partial
skeleton of Homo habilis from Olduvai
Gorge, Tanzania. Nature 327: 205–209
Kaiser T, Bromage TG, Schrenk F (1995) Hom-
inid corridor research project update: new
Pliocene fossil localities at Lake Manyara
and putative oldest Early Stone Age occur-
rences at Laetoli (Upper Ndolanya Beds),
northern Tanzania. J Hum Evol 28: 117–120
Keith A (1948) A new theory of human evolu-
tion. CAWatts & Company, London
Kimbel WH, Walter RC, Johanson DC, Red KE,
Aronson JL, Assefa Z, Marean CW, Eck GG,
Bobe R, Hovers E, Rak Y, Vondra C, Yemane
T, York D, Chen Y, Evensen NM, Smith PEB
(1996) Late Pliocene Homo and Oldowan
tools from the Hadar Formation (Kada
Hadar Member), Ethiopia. J Hum Evol 31:
549–562
King W (1864) The reputed fossil man of the
Neanderthal. Q J Sci 1: 88–97
Kullmer O, Sandrock O, Abel R, Schrenk F,
Bromage TG, Juwayeyi Y (1999) The first
Paranthropus from the Malawi Rift. J Hum
Evol 37: 121–127
Leakey RE (1973a) Evidence for an advanced
plio‐pleistocene hominid from East Rudolf,
Kenya. Nature 242: 447–450
Leakey RE (1973b) Further evidence of Lower
Pleistocene hominids from East Rudolf,
north Kenya. Nature 248: 653–656
Leakey LSB, Tobias PV, Napier JR (1964) A new
species of the genus Homo from Olduvai
Gorge. Nature 202: 7–10
Lewis OJ (1989) Functional morphology of the
evolving hand and foot. Oxford University
Press, Oxford
Linnaeus C (1758) Systema naturae. Laurentii
Salvii, Stockholm
Mayr E (1944) On the concepts and terminolo-
gy of vertical subspecies and species. Natl Res
Council Bull 262: 11–16
Prat S, Brugal J‐P, Tiercelin J‐J, Barrat J‐A,Boh M, Delagnes A, Harmand S, Kimeu K,
Kibunjia M, Texier P‐J, Roche H (2005) First
occurrence of early Homo in the Nachukui
Formation (West Turkana, Kenya) at 2.3–2.4
Myr. J Hum Evol 49: 230–240
Prentice ML, Denton GH (1988) In: Grine FE
(ed) Evolutionary history of the ‘‘Robust’’
Australopithecines. Aldine de Gruyter, New
York, pp 383–403
Ramirez Rozzi F (1997) Les hominides du Plio‐Pleistocene de la vallee de l’Omo. Cahiers de
paleoanthropologie CNRS editions, Paris
Rayner RJ, Moon BP, Masters JC (1993) The
Makapansgat australopithecine environment.
J Hum Evol 24: 219–231
Roche H, et al. (2003) Les sites arche ologi-
ques pliopleeistocenes de la formation de
Nachukui, Ouest Turkana: bilan synthetique
1997–2001. CR Palevol 2: 663–673
Schoetensack O (1908) Der Unterkiefer des
Homo heidelbergensis aus den Sanden
von Mauer bei Heidelberg. W Engelmann,
Leipzig

The earliest putative Homo fossils 9 1631
Schrenk F, Bromage TG, Betzler C, Ring U,
Juwayeyi Y (1993) Oldest Homo and Plio-
cene biogeography of the Malawi Rift. Nature
365: 833–836
Stern JT, Susman RL (1983) Locomotor anato-
my of Australopithecus afarensis. Am J Phys
Anthropol 60: 279–317
Suwa G, White TD, Howell FC (1996) Mandib-
ular postcanine dentition from the Shungura
Formation, Ethiopia: crown morphology,
taxonomic allocations, and Plio‐Pleistocenehominid evolution. Am J Phys Anthropol
101: 247–282
Tobias PV (1987) The brain of Homo habilis: a
new level of organization in cerebral evolu-
tion. J Hum Evol 16: 741–761
Tobias PV (1991) The skulls, endocasts and
teeth of Homo habilis. In: Olduvai Gorge,
vol 4, 2 vols. Cambridge University Press,
Cambridge
Tobias PV, von Koenigswald GHR (1964) A
comparison between the Olduvai hominines
and those of Java and some implications for
hominid phylogeny. Nature 204: 515–518
Toth N (1985) Archeological evidence for pref-
erential right‐handedness in the lower and
middle Pleistocene, and its possible implica-
tions. J Hum Evol 14: 607–614
Vrba ES (1985) Ecological and adaptive changes
associated with early hominid evolution. In:
Delson E (ed) Ancestors: the hard evidence.
Alan R. Liss, New York, pp 63–71
Vrba ES (1988) Late Pliocene climatic events
and human evolution. In: Grine FE (ed)
Evolutionary history of the ‘‘Robust’’ Austra-
lopithecines. Aldine de Gruyter, New York,
pp 405–426
Vrba ES (1992) Mammals as a key to evolution-
ary theory. J Mamm 73: 1–28
Wood BA (1991) Koobi Fora research project,
vol 4: Hominid Cranial remains. Clarendon
Press, Oxford
Wood BA (1992) Origin and evolution of the
genus Homo. Nature 355: 783–790
Wood BA, Collard M (2001) The meaning of
Homo. Ludus Vitalis vol. IX. 15: 63–74




















