
Selective Suicide Gene Therapy of Colon Cancer Exploiting the Urokinase Plasminogen Activator...
12
Selective Suicide Gene Therapy of Colon Cancer Cell Lines Exploiting Fibroblast Growth Factor 18 Promoter Ladan Teimoori-Toolabi, 1 Kayhan Azadmanesh, 2,3 and Sirous Zeinali 1 Abstract Fibroblast growth factor 18 (FGF18) is one of the genes downstream of Wnt, one of the most important signaling pathways activated in colon cancer. An FGF18 promoter containing a single T-cell factor=lymphocyte enhancing factor 1 (TCF=LEF1) binding site was inserted upstream of a thymidine kinase (TK) suicide gene module, while a bacterial b-Gal (LacZ) element served as the reporter gene. Following transient transfection with pUCFG- F18LacZ, b-Gal staining showed that 5% of SW480, 10% of HCT116, 0% of human umbilical vein endothelial cells (HUVECs) and 0% of normal colon cells (NCCs) had expressed LacZ. b-Gal enzyme-linked immunosorbent assay revealed that the ratio of pUCFGF18LacZ activity to that of positive control was 0.09 and 0.25 in SW480 and HCT116, respectively (significantly higher than mock plasmid), while there were no significant changes in the b-Gal expression in HUVEC and NCC cells transfected with pUCFGF18LacZ or mock plasmid. Following transfection with pUCFGF18TK and pUCCMVTK (positive control), cytotoxicity analysis of transfected cells showed that treatment with ganciclovir (GCV) significantly decreased SW480 and HCT116 cell survival at GCV concentrations above 20 mg=mL. An inverse correlation between GCV concentration and cell viability was evi- dent in both colon cancer cell lines following transfection with these suicide plasmids. pUCFGF18TK and pUCCMVTK induced apoptosis after the administration of GCV in HCT116, but not in SW480, as demonstrated by M30 cytodeath antibody. This discrepancy may stem from differences in the mechanisms of TK=GCV- induced apoptosis in p53-proficient (HCT116) and -deficient (SW480) cells. The specific activity of the FGF18 promoter in HCT116 and SW480 may reflect the advantage of this promoter over artificial promoters containing artificial TCF=LEF binding sites. Key words: genes, transgenic, suicide, neoplasm, colonic neoplasm, gene therapy Introduction C olon cancer is the third and fourth most prevalent cancer among Iranian women and men, respectively, 1 whereas in the United States, it is rated the third most common cancer for both men and women. 2 Conventional therapies cannot fully eradicate cancerous cells, and therefore, gene therapy has attracted much attention in recent years. 3 Efficacy of chemotherapy in complete eradication of a few types of can- cers, such as leukemia, has led to the hypothesis that all types of cancer can potentially be eradicated by higher doses of chemotherapy, 4 an assumption that is questionable given the side-effects associated with high-dose regimens of chemo- therapeutic agents. In suicide gene therapy, a gene construct that encodes an enzyme is introduced to the target tissues. The enzyme converts a nontoxic prodrug to a toxic drug. There- fore, suicide gene therapy opens a new horizon for the local administration of these toxic drugs. Thymidine kinase (TK) is the first, and the most studied, suicide gene, 5 which accounts for about 10% of all clinical trials evaluating cancer gene therapy. 6 The mechanism of its action exploits a unique characteristic of herpes simplex virus thy- midine kinase (HSV-TK), which, contrary to normal mamma- lian thymidine kinase, preferentially monophosphorylates gancyclovir (GCV), rendering it toxic to normal mammalian cells. Further phosphorylation of GCV-monophosphate by Departments of 1 Molecular Medicine, Biotechnology Research Center, 2 Hepatitis and AIDS, and 3 Virology, Pasteur Institute of Iran, Tehran, Iran. Address correspondence to: Sirous Zeinali; Department of Molecular Medicine, Biotechnology Research Center, Pasteur Institute of Iran; 69th Pasteur Avenue, Kargar Street, Tehran 13169-43551, Iran E-mail: [email protected] CANCER BIOTHERAPY AND RADIOPHARMACEUTICALS Volume 25, Number 1, 2010 ª Mary Ann Liebert, Inc. DOI: 10.1089=cbr.2009.0643 105
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Selective Suicide Gene Therapy of Colon Cancer Exploiting the Urokinase Plasminogen Activator...

Selective Suicide Gene Therapy of Colon CancerCell Lines Exploiting Fibroblast Growth Factor 18 Promoter
Ladan Teimoori-Toolabi,1 Kayhan Azadmanesh,2,3 and Sirous Zeinali1
Abstract
Fibroblast growth factor 18 (FGF18) is one of the genes downstream of Wnt, one of the most important signalingpathways activated in colon cancer. An FGF18 promoter containing a single T-cell factor=lymphocyte enhancingfactor 1 (TCF=LEF1) binding site was inserted upstream of a thymidine kinase (TK) suicide gene module, while abacterial b-Gal (LacZ) element served as the reporter gene. Following transient transfection with pUCFG-F18LacZ, b-Gal staining showed that 5% of SW480, 10% of HCT116, 0% of human umbilical vein endothelialcells (HUVECs) and 0% of normal colon cells (NCCs) had expressed LacZ. b-Gal enzyme-linked immunosorbentassay revealed that the ratio of pUCFGF18LacZ activity to that of positive control was 0.09 and 0.25 in SW480and HCT116, respectively (significantly higher than mock plasmid), while there were no significant changes inthe b-Gal expression in HUVEC and NCC cells transfected with pUCFGF18LacZ or mock plasmid. Followingtransfection with pUCFGF18TK and pUCCMVTK (positive control), cytotoxicity analysis of transfected cellsshowed that treatment with ganciclovir (GCV) significantly decreased SW480 and HCT116 cell survival at GCVconcentrations above 20 mg=mL. An inverse correlation between GCV concentration and cell viability was evi-dent in both colon cancer cell lines following transfection with these suicide plasmids. pUCFGF18TK andpUCCMVTK induced apoptosis after the administration of GCV in HCT116, but not in SW480, as demonstratedby M30 cytodeath antibody. This discrepancy may stem from differences in the mechanisms of TK=GCV-induced apoptosis in p53-proficient (HCT116) and -deficient (SW480) cells. The specific activity of the FGF18promoter in HCT116 and SW480 may reflect the advantage of this promoter over artificial promoters containingartificial TCF=LEF binding sites.
Key words: genes, transgenic, suicide, neoplasm, colonic neoplasm, gene therapy
Introduction
Colon cancer is the third and fourth most prevalent canceramong Iranian women and men, respectively,1 whereas
in the United States, it is rated the third most common cancerfor both men and women.2 Conventional therapies cannotfully eradicate cancerous cells, and therefore, gene therapyhas attracted much attention in recent years.3 Efficacy ofchemotherapy in complete eradication of a few types of can-cers, such as leukemia, has led to the hypothesis that all typesof cancer can potentially be eradicated by higher doses ofchemotherapy,4 an assumption that is questionable given theside-effects associated with high-dose regimens of chemo-
therapeutic agents. In suicide gene therapy, a gene constructthat encodes an enzyme is introduced to the target tissues. Theenzyme converts a nontoxic prodrug to a toxic drug. There-fore, suicide gene therapy opens a new horizon for the localadministration of these toxic drugs.
Thymidine kinase (TK) is the first, and the most studied,suicide gene,5 which accounts for about 10% of all clinical trialsevaluating cancer gene therapy.6 The mechanism of its actionexploits a unique characteristic of herpes simplex virus thy-midine kinase (HSV-TK), which, contrary to normal mamma-lian thymidine kinase, preferentially monophosphorylatesgancyclovir (GCV), rendering it toxic to normal mammaliancells. Further phosphorylation of GCV-monophosphate by
Departments of 1Molecular Medicine, Biotechnology Research Center, 2Hepatitis and AIDS, and 3Virology, Pasteur Institute of Iran, Tehran,Iran.
Address correspondence to: Sirous Zeinali; Department of Molecular Medicine, Biotechnology Research Center, Pasteur Institute of Iran;69th Pasteur Avenue, Kargar Street, Tehran 13169-43551, IranE-mail: [email protected]
CANCER BIOTHERAPY AND RADIOPHARMACEUTICALSVolume 25, Number 1, 2010ª Mary Ann Liebert, Inc.DOI: 10.1089=cbr.2009.0643
105

cellular kinases produces a metabolite, which, upon inte-gration into DNA, terminates DNA-strand elongation.7 Inaddition to the direct toxicity induced by this suicide gene, abystander effect also plays an important role in enhancing itsefficacy.8 Earlier clinical trials in using this system have hadcontroversial results. For example, suicide gene therapy inmalignant mesothelioma increased the median survival ofpatients,9 and clinical trials involving localized prostatecarcinoma resulted in a lengthened doubling time for thecancer-marker prostate-specific antigen10,11 (PSA). However,the results of clinical trials on brain tumors were unsatis-factory.12
Several strategies have been designed to overcome thetoxic effects of the systemic administration of vectors con-taining suicide genes,13 such as targeting the construct to thedesired cells by inserting monoclonal antibodies (mAbs)specific for tumor cells on the surface of the viral vectors14 orrestricting the expression of suicide gene by tissue- or cancer-specific promoters.15,16 Cancer-specific promoters are betterchoices, since they can generally be used in different typesof cancers and can evade downregulation in case the can-cerous cells dedifferentiate.16 There are many examples ofcancer-specific suicide gene therapy, such as the PSA pro-moter for prostate adenocarcinoma17 or carcinoembryogenicantigen (CEA) promoter for colon carcinoma.18
One of the more successful strategies in selecting the bestcancer-specific promoters for suicide gene therapy is to in-vestigate deregulated signaling pathways in cancerous cellsin order to identify downstream promoters abnormally up-regulated by these pathways.
The most important constitutive pathway showing ab-normal activation in colon cancer cells is Wnt.3,19 Inactivatingmutations in the adenomatous polyposis coli (APC) gene areseen in 70%–80% of familial and sporadic cases of coloncancer.20 If this signaling pathway is activated, for example,by mutations that inactivate the APC gene or b-catenin geneproducts, proteasome-mediated degradation of b-cateninthrough axin and GSK3 (glycogen synthetase kinase 3)complex would be disrupted. This would lead to increased b-catenin in the cytoplasm and, subsequently, to its migrationinto the nucleus. Free b-catenin in the nucleus associates withthe HMG-box transcription factor, T-cell factor 4 (TCF4); thiscomplex then activates promoters that contain TCF bindingsites.20 Some examples of genes downstream to this pathwaybeen identified by microarray21 analysis are vascular endo-thelial growth factor (VEGF),22 neural cell adhesion molecule(Nr-CAM),23 and fibroblast growth factor 18 (FGF18).24
Considering the importance of this pathway in trans-forming the epithelial cells of the colon, it has been hypoth-esized that the Wnt signaling pathway can be exploited incolon-cancer–specific suicide gene therapy. Various studieshave utilized this pathway; for instance, swapping out TCFbinding sites for the adenoviral E2 promoter in cell lines withan activated Wnt signaling pathway25 yielded activity com-parable to the wild-type E2 promoter. Insertion of five TCFbinding sites adjacent to the minimal SV40 promoter showedpromising results; however, there was a promiscuous ex-pression in normal cells,3 which was overcome by replacingthe minimal SV40 promoter with a TATA box.26 Both ofthese promoters were active in colon cancer cell lines withhigh activation levels of Wnt. Other studies have reportedthe use of a minimal cytomegalovirus (CMV) promoter27 or
minimal thymidine kinase promoter28 besides using artificialTCF=LEF binding sites.29 These constructs showed bothspecificity and activity in cell lines with activated Wnt sig-naling pathways; however, their activity in various coloncancer cell lines has not been confirmed.28,30
Expression profile data of colon carcinoma have shownthat in cancerous tissues, the FGF18 gene is upregulateddownstream of the Wnt signaling pathway; therefore, theFGF18 promoter can be used to construct cancer-specificsuicide vectors. FGF18 is a heparin-binding extracellular se-creting factor,29 which normally plays a role in chondrogenesisand osteogenesis31; it is only expressed in adult heart tissue.24
In this study, the unaltered native FGF18 promoter con-taining one TCF=LEF1 binding site24 was cloned upstream ofthe bacterial b-Gal (LacZ) reporter gene and thymidine kinase(TK) suicide gene. The influence of these constructs on twocolon cancer cell lines (SW480 and HCT116), which exhibitinherently different levels of activation of the Wnt signal-transduction pathway, and on the normal human umbilicalvein endothelial cell line (HUVEC) and normal colon cells(NCCs), was studied.
The application of natural promoters might carry the ad-vantage of mimicking the natural state of the promoters inthe cells, utilizing repressor elements to restrict expression inthe normal cells. Results obtained in this study may providean insight into the comparative performances of natural andartificial promoters studied, so far.
Materials and Methods
DNA extraction
Genomic DNA was extracted from venous blood lym-phocytes of a healthy person by the proteinase K method.32
In accord with guidelines, signed consent was obtained fromthe blood donor; she had no history of familial cancers or ofcancer onset before the age of 60 in her relatives. PlasmidDNA and viral DNA from herpes simplex virus were ex-tracted by a miniprep plasmid extraction kit and theQiaAmp Viral DNA Extraction Kit, respectively (Qiagen,Germany).
Fragment amplifications
The CMV and FGF18 promoters were amplified frompcDNA3.1þ (Invitrogen) and human genomic DNA, re-spectively. The gene was amplified from the herpes simplexvirus 1 genome. Primer sequences are given in Table 1.
Plasmid design and construction
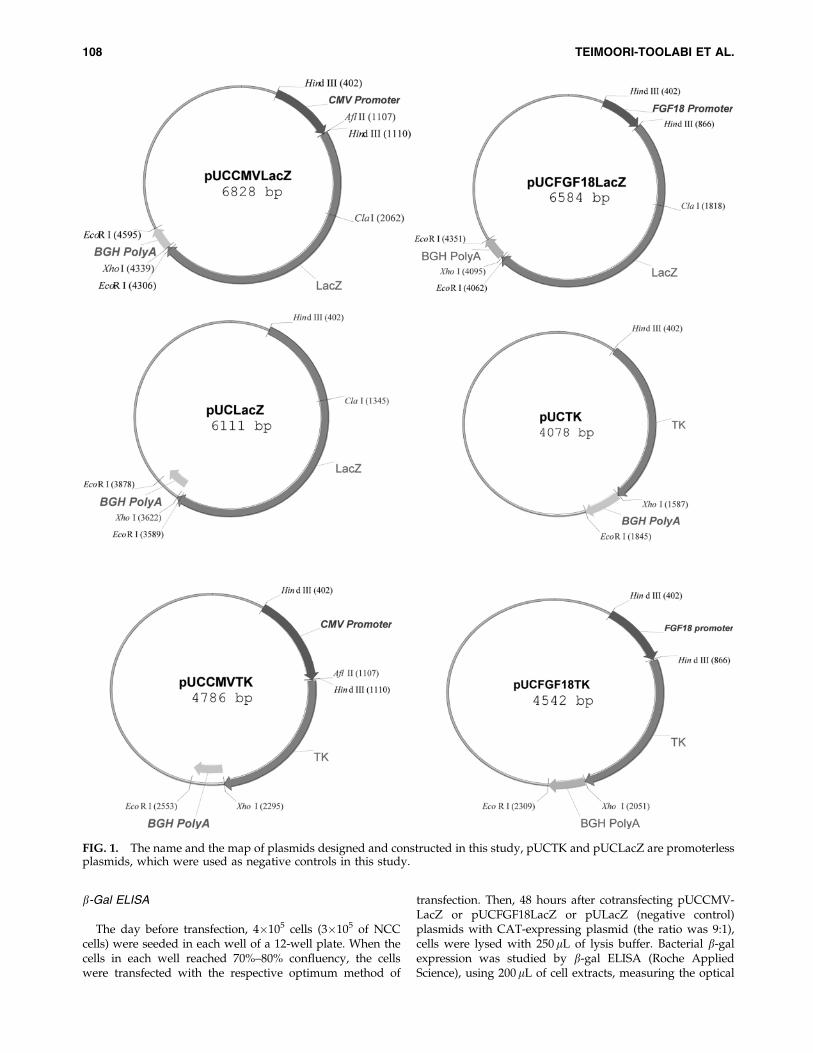
In this study, six vectors were constructed from usingpUCLTRLacZ, which encodes the complete bacterial b-Galgene between the HTLV-1 long terminal repeat (LTR)promoter and BGH polyA.33 The LTR was digested out andreplaced with CMV or FGF18 promoters to obtainpUCCMVLacZ and pUCFGF18LacZ constructs. The HindIIIdigested plasmid was also relegated, resulting in pUCLacZplasmid. In addition, LacZ was digested out from pUClacZplasmid by XhoI and NcoI, keeping the PolyA tail intact, andthe TK gene was inserted in its place to construct pUCTK.CMV or FGF18 promoters were also placed upstream of theTK gene to obtain pUCCMVTK or pUCFGF18TK. Schematicfigures of constructed plasmids are given in Figure 1.
106 TEIMOORI-TOOLABI ET AL.

In this study, the promoterless plasmids, pUCLacZ andpUCTK, were used as negative controls.
Cell lines and culturing
HUVEC and SW480 cell lines were obtained from Na-tional Cell Bank of Iran (NCBI; Pasteur Institute of Iran,Tehran, Iran). The HCT116 cell line was obtained fromATCC (American Type Culture Collection, Manassas, VA).SW480 and HCT116 cell lines were cultured in high-glucoseDulbecco’s modified Eagle’s medium (DMEM) with 10%fetal bovine serum (FBS) plus 100 unit=mL of penicillin,100 mg=mL of streptomycin, and 2 mM of l-glutamine. TheHUVEC cell line was cultured in a medium containingHAM’s F12: high glucose DMEM (1:1), with the same sup-plements as described above. The media for HCT116, SW480,and HUVEC cell lines were changed every 3, 5, and 6 days,respectively. All reagents were purchased from Gibco(Grand Isle, NY).
Obtaining primary cells from normal colon tissue
The normal colon tissue specimen was freshly obtainedfrom a 27-year-old man under surgery at Imam KhomeiniHospital (Tehran University of Medical Sciences). He hadundergone the surgical procedure because of a volvulus inhis colon. He had no history of familial cancer, and none ofhis first-degree relatives had died from colorectal carcinoma.Before donating his tissue, he had signed the ethical consent.This specimen was proved to be normal in pathologic studies.The CEA was not positive in the donor. This procedurewas approved by the ethical committee of Pasteur Instituteof Iran.
Immediately upon transferring the tissue to the lab, it waswashed with phosphate-buffered saline (PBS). Then, it waschopped into 1-mm pieces. The chopped pieces were washedwith PBS and then treated with 200 unit=mL of collagenase(Gibco-BRL) in a CO2 incubator for about 2 hours. There-after, the cells were seeded into two to four wells of a 12-wellplate (Nunc, Denmark). They were cultured in a mixture ofDMEM:Ham’s F12(1:1) media (Gibco-BRL), supplementedwith 4 mM of l-glutamine, 200 unit=mL of penicillin,200 mg=mL of streptomycin, 2.5 mG=mL of fungizone (Gibco-BRL), and 40mg=mL of gentamycin for about 5 days. In thenext four passages, fungizone and gentamycin were re-moved from the supplement and the concentration of peni-cillin and streptomycin was decreased to 100 and 100 mg=mL,respectively.
The cells were seeded onto a plate treated with collagenfrom rat tail (Roche Applied Science, Germany). After twopassages, they were trypsinized from the plate and seededonto noncollagenated plates or flasks for further experi-ments. In this article, this mixture of adherent cells obtainedfrom normal colon tissue has been named NCCs.
Optimization of transfection methods
One (1) day prior to transfection, 2�105 of SW480,HCT116, and HUVEC cell lines and 1.5�105 of NCCs wereseeded into each well of a 24-well plate. When the cells ineach well reached 70%–80% confluency, they were trans-fected with positive control plasmid (pUCCMVLacZ), usingLipofectamine 2000 (Invitrogen), polyfect, and effectene(Qiagen). Different ratios of DNA and reagents examined inthe optimization procedure are given in Table 2. The per-centage of transfected cells was calculated by a semiquanti-tative method, using b-Gal staining. The average ratio ofstained cells to the total number of cells was examined in atleast five different fields to determine the transfection rate.The death rate for each method was estimated by stainingthe transfected cells in another well (treated with the samereagent and DNA: reagent ratio) by the trypan blue method.The optimum transfection method was chosen according tothe maximum transfection rate and the minimum death rateof the cells. If the death rate of the cells in a method washigher than 30% (high), the method would not be consideredas an optimum method. Transfection rate and death rate ofcells were compared with nontransfected wells.
For normalization of transfection in b-Gal enzyme-linkedimmunorsorbent assay (ELISA), cells were transfected with amixture of the main reporter and chloramphenicol acetyl-transferase (CAT)-expressing construct (pRc=CMV2CAT;Invitrogen, USA) in the ratio of 9:1.
b-Gal staining
Forty-eight (48) hours after transfection, the cells werefixed with 0.5% glutaraldehyde (Sigma, St. Louis, MO). Afterwashing with (PBS), the cells in a 24-well plate (Nunc) werestained with 3 mL of 400 mM of ferrocyanide potassium(Fluka, Switzerland), 3 mL of 400 mM of ferricyanide potas-sium (Fluka), 3 mL of 200 mM of MgCl2 (Merck, Germany),15mL of 20 mg=mL of Xgal (Fermentas, Lithuania) and276mL of PBS. The percentage of positively stained cells ineach well was estimated by counting the blue cells amongthe total number of cells in at least five different fields underthe inverted microscope (400�zoom).
Table 1. Sequences of Primers: Primers for Real-Time Polymerase Chain Reaction and Cloning
b-actin forward primer for Real-Time 50-CCAAGGCCAACCGCGAGAAG-30
b-actin reverse primer for Real-Time 50-CACCGGAGTCCATCACGATGC-30
Thymidine kinase forward primer for real-time 50-AAACGCCTCCGTCCCATG-30
Thymidine kinase reverse primer for real-time 50-GGTCGCAGATCGTCGGTATG-30
Thymidine kinase forward primer for cloning 50-ATCCATGGCTTCGATCCCCTGCCA-30
Thymidine kinase reverse primer for cloning 50-TCCTCGAGTCATAGCGCGGGTTCCTTC-30
CMV promoter forward primer 50-AATAAGCTTCGATGTACGGGCCAGA-30
CMV promoter reverse primer 50-GGTAAGCTTAAGTTTAAACGCTAG-30
FGF18 promoter forward primer 50-TGCAAGCTTAGCATCGCCTGCCCAATTTG-30
FGF18 promoter reverse primer 50-AGCAAGCTTTCATGTCTCCTCCTCCGCGTCTCTC-30
CMV, cytomegalovirus.
FGF18 PROMOTER IN COLON CANCER SUICIDE GENE THERAPY 107
b-Gal ELISA
The day before transfection, 4�105 cells (3�105 of NCCcells) were seeded in each well of a 12-well plate. When thecells in each well reached 70%–80% confluency, the cellswere transfected with the respective optimum method of
transfection. Then, 48 hours after cotransfecting pUCCMV-LacZ or pUCFGF18LacZ or pULacZ (negative control)plasmids with CAT-expressing plasmid (the ratio was 9:1),cells were lysed with 250 mL of lysis buffer. Bacterial b-galexpression was studied by b-gal ELISA (Roche AppliedScience), using 200 mL of cell extracts, measuring the optical
FIG. 1. The name and the map of plasmids designed and constructed in this study, pUCTK and pUCLacZ are promoterlessplasmids, which were used as negative controls in this study.
108 TEIMOORI-TOOLABI ET AL.
density (OD) at 405 nm (490 nm as background) every 2minutes by a microplate reader (Biotek) and calculating themaximum slopes of OD change.
CAT ELISA
First 50mL of the cell extracts used for b-Gal ELISA weremixed with 150 mL of sample buffer and then added to eachwell of a CAT ELISA plate (Roche Applied Science). Afterfollowing the ELISA protocol detailed in the manufacturer’sinstructions, the OD was measured at 405 nm (490 nm asbackground) every 2 minutes and the maximum slopes ofOD change were calculated.
Cytotoxicity assay
Four (4) and 8 hours after transfection with polyfect andeffectene reagents, cells were trypsinized and 104 cells wereplated into each well of a flat-bottomed 96-well cell-cultureplate (Nunc). On the next day, GCV (Roche, Switzerland)was added to cell medium at final concentrations of 0, 20, 40,60, 80, and 100 mg=ml. Four (4) days after transfection, themedia were replaced with 100 mL of fresh DMEM with 10%FBS plus 50mL of XTT mixture (Roche Applied Science), asper the manufacturer’s instructions. After 4 hours of incu-bation in a CO2 incubator, the OD of cell medium was
measured at 490 nm with the background of 690 nm by amicroplate reader (Biotek).
Analyzing the dead cells by staining the cells with M30cytodeath antibody
Twenty-four (24) hours after transfecting HCT116 andSW480 cell lines with suicide and negative control plasmids,40mg=mL GCV was added to the cell medium. Then, 24hours later, cells were trypsinized and washed with PBS andstained with M30 cytodeath antibody (Roche Applied Sci-ence), according to the manufacturer’s instructions. There-after, cells were analyzed by a flowcytometer with at least10,000 events per reading in a PAS machine, using flowmaxsoftware (Partec, Germany).
The GCV concentrations and cell-harvesting time forflowcytometry experiments were optimized by collecting thecells at different time intervals with different GCV concen-trations.
RNA extraction
Twenty-four (24) hours after transfection in a 12-wellplate, the cells were washed with PBS and then RNA wasextracted with 1 mL of Tripure (Roche Applied Science).32
Table 2. Optimization of Transfection in HCT116, SW480, HUVEC, and Normal Colon Cells (NCCs)
ReagentDNA(ng)
Reagent(mL)
Transfectionrate in
HCT116 (%)
Deathrate in
HCT116
Transfectionrate in
SW480 (%)
Deathrate inSW480
Transfectionrate in
HUVEC (%)
Deathrate in
HUVEC
Transfectionrate in
NCC (%)
Deathrate inNCC
PolyFect 400 2 30 Low 10 Low 2–5 Low 2 LowPolyFect 400 4 35 Low 10 Low <5 Low >2 LowPolyFect 400 6 35–40 Low 10 Low <5 Low >2 LowPolyFect 800 4 <50 Low 4 Moderate <5 Low 3 LowPolyFect 800 8 <50 Low 5 Moderate 5 Low 1 ModeratePolyFect 800 12 45 Low 7 High 10 Low 1 ModeratePolyFect 1600 8 50 Low 4 Moderate <5 Moderate 4 LowPolyFect 1600 16 45 Low 2 Moderate 5 Moderate 3 ModeratePolyFect 1600 24 40 Low 1 High 10 High 4 HighEffectene 100 1 5–10 Low 10 Low <0.5 Low >0 LowEffectene 100 2.5 15 Low 15 Low <0.5 Low >0 LowEffectene 100 5 15 Low 20 Low 0.5 Low <0.5 lowEffectene 200 2 7–10 Low 20 Low 0.5 Low 0.7 lowEffectene 200 5 12–17 Low 20–25 Low 0.5–1 Low 0.5 lowEffectene 200 10 <20 Low 15–20 Low 1 Low 0.5 lowEffectene 400 4 <20 Low 30–40 Moderate 0.5 Moderate 0.5 lowEffectene 400 10 30 Moderate 40 High 1 Moderate 0.7 moderateEffectene 400 20 20–25 High 30 High 1–1.5 High 0.7 highLipofectemine 800 1.2 1 Low 1 Low <0.5 Low <0.5 lowLipofectemine 800 2 <1 Low <1 Low <0.5 Low 0.5 lowLipofectemine 800 2.8 1 Low 1 Low <0.5 Low 0.5 lowLipofectemine 1600 2.4 1.5 Low 1.5 Low 0.5 Low 1 lowLipofectemine 1600 4 1.5 Low 1.5 Moderate 0.5–1 High 2 lowLipofectemine 1600 5.6 6 Moderate 6 Moderate 0.5–1 High 1–2 moderateLipofectemine 3200 4.8 7 Moderate 7 Moderate 0.5–1 Moderate <1 moderateLipofectemine 3200 8 15 High 15 High 0.5–1 High 1 moderateLipofectemine 3200 11.2 15 High 15 High 1 High <1 high
Different rates of DNA and reagents were used and summarized in this table, transfection rates were calculated by b-Gal staining(described in Materials and Methods), and death rate of cells were calculated by trypan blue staining of another well with the same condition.The death rate was assumed to be low when it was lower than 10%, moderate when it was between 10 and 30%, and high when it was higherthan 30%.
HUVEC, human endothelial vein endothelial cell.
FGF18 PROMOTER IN COLON CANCER SUICIDE GENE THERAPY 109

The OD ratio of extracted RNA was measured at 260 and280 nm, and the quality of RNA was considered suitablewhen the ratio of 260:280 was between 1.5 and 1.8.
cDNA synthesis
The extracted RNA was treated with DNAase to removethe contaminating plasmid DNA. The RNA was converted tocDNA by using M-MuLV as a reverse-transcriptase enzymeand oligo dT as the primer.32 All reagents were obtainedfrom Roche Applied Science.
Real-time polymerase chain reaction (PCR)
The expression of TK in cells transfected with TK-expressing plasmids was assessed at the cDNA level, usingSYBR Green mastermix (AB), in an AB 7300 machine (AB).This amplified a 141-bp fragment of thymidine kinase fromthe cDNA. b-actin expression was also evaluated at the cDNAlevel by amplifying a 134-bp fragment, which served as thereference. The sequences of primers for amplifying cDNAfrom both genes are given in Table 1.
The efficiency of amplification was evaluated by testingserial dilutions of cDNA on which the real-time PCR ex-periments were performed.
Statistical methods
All experiments were repeated at least three times. Theresults were analyzed by the Student’s t-test and Pearsoncorrelation coefficient, using SPSS software, version 12 (SPSS,Inc., Chicago, IL).
Results
Transfection optimization
An optimized transfection rate for SW480, HCT116,HUVEC, and NCC was achieved by using Effectene atthe DNA=reagent ratio of 400 ng=4 mL, and PolyFect at theDNA=reagent ratios of 1600 ng=8mL, 800 ng=12mL, and1600 ng=8mL, respectively. Table 2 summarizes the trans-fection and death rates of these cell lines after applying dif-ferent reagents.
The maximum transfection rates for HCT116, SW480,HUVEC, and NCC cells were 50%, 35%, 10%, and below10%, respectively.
Semiquantitative analysis showed that FGF18promoter was active in both colon cancer cell linesbut not in HUVEC cell line
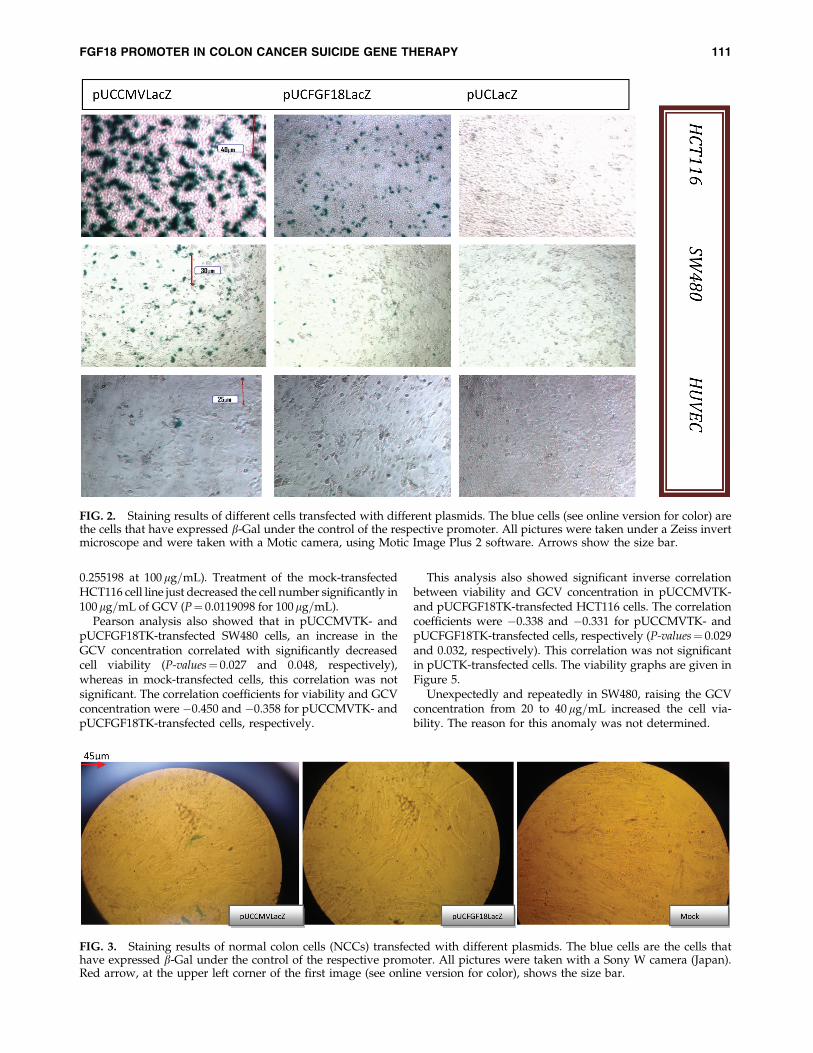
Staining results of HCT116, SW480, and HUVEC celllines transfected with different plasmids are shown inFigure 2. The staining results of NCC are also given inFigure 3.
In SW480, HCT116, HUVEC, and NCC cells transfectedwith pUCFGF18LacZ, 5%, 10%, 0%, and 0% of the cells werepositively stained, respectively.
b-Gal expression under the control of the FGF18 promoterwas not observed in the HUVEC cell line. Low rates oftransfection in this cell line made it necessary to confirm alack of b-Gal expression by b-Gal ELISA, which is a quan-titative method.
b-Gal ELISA confirmed that FGF18 promoter drivesexpression of reporter gene in colon cancer cell linesbut not in HUVEC and NCC
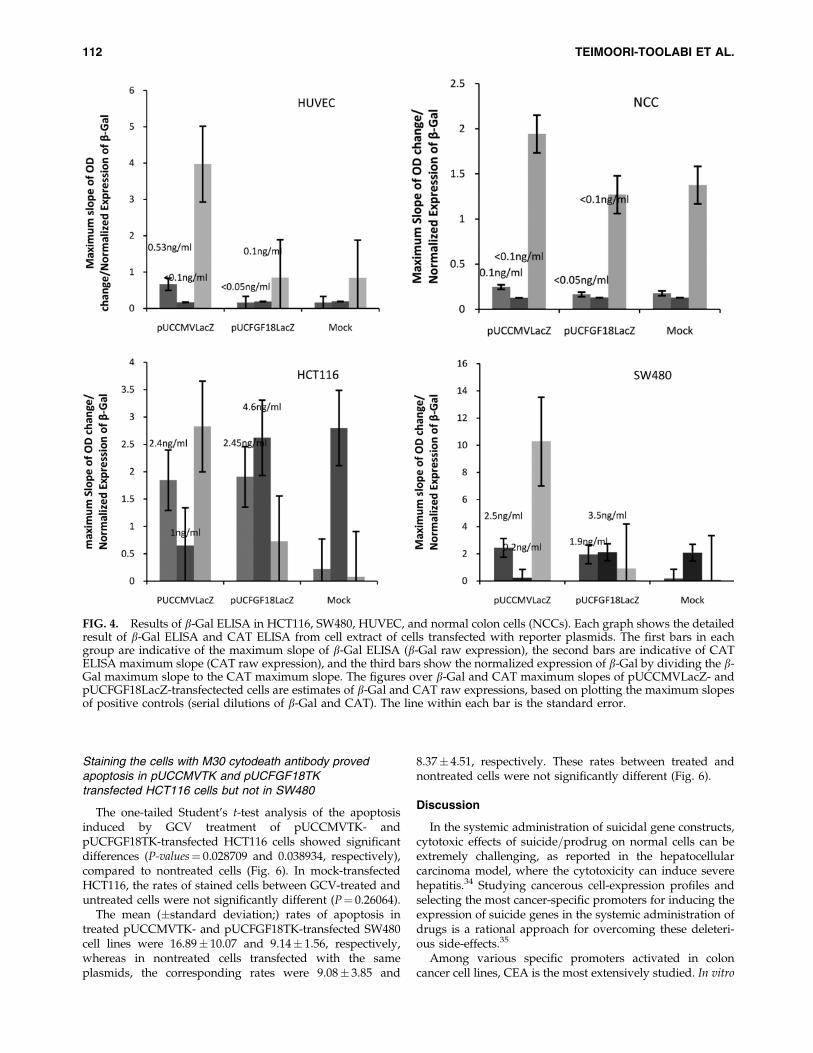
The transfection rates of all cell lines with the main re-porter plasmid (pUCCMVLacZ, pUCFGF18LacZ, or nega-tive control) were normalized by the expression of CATthrough cotransfection with pRc=CMV2CAT. For this pur-pose, the b-Gal maximum slope was divided by the CATmaximum slope of the same cell lysate. Normalized expres-sion of b-Gal, induced by different plasmids in SW480,HCT116, HUVEC, and NCC cells, is shown in Figure 4.
HCT116 and SW480 cell lines transfected with pUCCMVLacZ exhibited significantly higher expression of b-Galthan mock-transfected cells (P-values¼ 0.003 and 0.007, re-spectively). Expression of this gene under FGF18 promoterwas significantly higher than the negative control in theHCT116 and SW480 cell lines (P-values¼ 0.001 and 0.004,respectively). The ratios of FGF18 promoter activity, incomparison to CMV promoter activity, in HCT116 andSW480 cell lines were 0.25 and 0.09, respectively.
The lower transfection rate for the HUVEC cell line wascompensated for by normalization with CAT expression; theresults showed that b-Gal was not expressed by pUCFG-F18LacZ in this cell line, at levels comparable to the negativecontrol. However, pUCCMVLacZ transfected cells presentedsignificantly higher b-Gal expression than pUCFGF18LacZ(P¼ 0.01242) and pUCLacZ (P¼ 0.01248). From these data,it could be concluded that the FGF18 promoter was notactive in this cell line, while CMV promoter activityreached a significant level after normalization with CATexpression.
In NCC cells, which were a mixture of different cells ob-tained from normal colon tissue, b-Gal expression under thecontrol of CMV promoter was significantly higher than mockplasmid (P¼ 0.01227) and pUCFGF18LacZ (P¼ 0.01529). Incontrast, the FGF18 promoter did not induce significantlevels of b-Gal expression (P¼ 0.5117), in comparison tomock plasmid.
Real-time PCR assay further proved the expressionof TK by FGF18 promoter in both colon cancercell lines
Considering that the efficiency of real-time PCR for TKand b-actin genes was similar, mean expression � standarddeviation of TK expression induced by pUCFGF18TK, incomparison to pUCCMVTK, was 0.036623� 0.003984 inSW480 and 0.025599� 0.006353 in HCT116, respectively.
Cytotoxicity assays showed that FGF18TK constructinduces cell death in both colon cancer cell lines
Cell viability of SW480 cells transfected with pUCCMVTKand pUCFGF18TK in the presence of 20mg=mL or higher of aGCV concentration was significantly lower than in untreatedcells (P-values¼ 0.036 and 0.031, respectively).
In HCT116 cells, the viability of pUCCMVTK- andpUCFGF18TK-transfected cells in the presence of 20mg=mLand higher GCV concentrations was significantly lower thanin untreated cells (P-values¼ 0.011 and 0.009).
In SW480 cells, even 100mg=mL of GCV could not reducethe viability of mock-transfected cells significantly (P¼
110 TEIMOORI-TOOLABI ET AL.
0.255198 at 100mg=mL). Treatment of the mock-transfectedHCT116 cell line just decreased the cell number significantly in100mg=mL of GCV (P¼ 0.0119098 for 100mg=mL).
Pearson analysis also showed that in pUCCMVTK- andpUCFGF18TK-transfected SW480 cells, an increase in theGCV concentration correlated with significantly decreasedcell viability (P-values¼ 0.027 and 0.048, respectively),whereas in mock-transfected cells, this correlation was notsignificant. The correlation coefficients for viability and GCVconcentration were �0.450 and �0.358 for pUCCMVTK- andpUCFGF18TK-transfected cells, respectively.
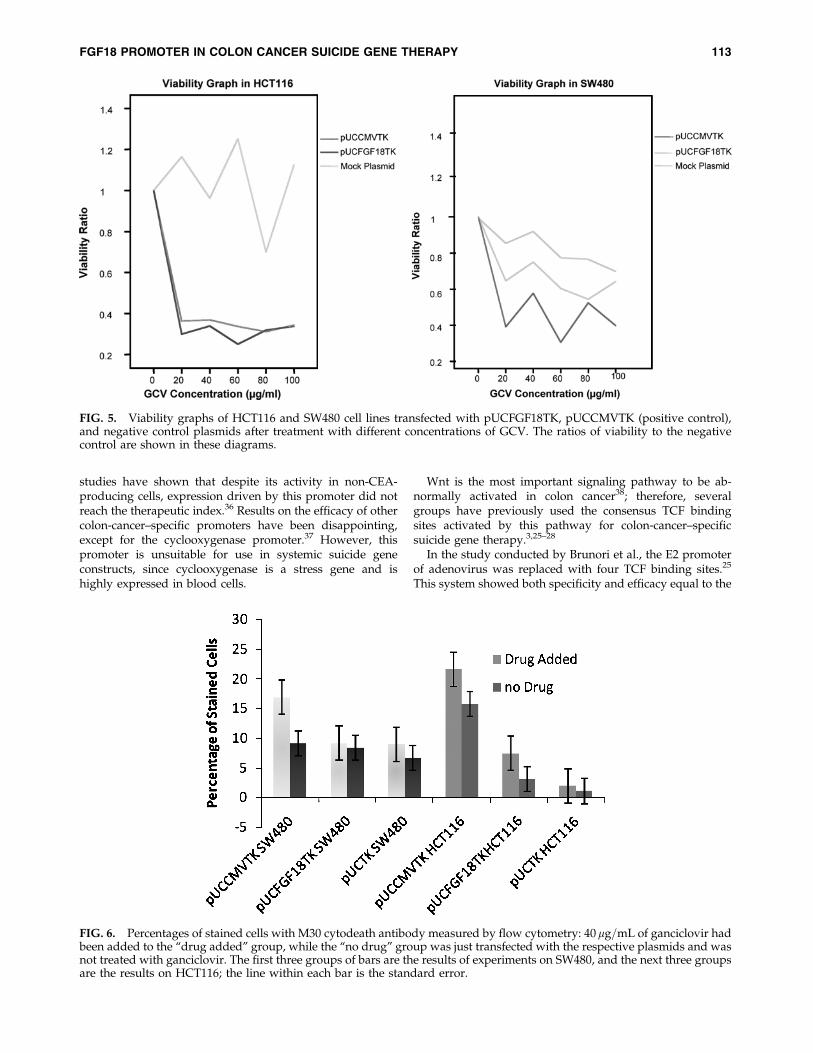
This analysis also showed significant inverse correlationbetween viability and GCV concentration in pUCCMVTK-and pUCFGF18TK-transfected HCT116 cells. The correlationcoefficients were �0.338 and �0.331 for pUCCMVTK- andpUCFGF18TK-transfected cells, respectively (P-values¼ 0.029and 0.032, respectively). This correlation was not significantin pUCTK-transfected cells. The viability graphs are given inFigure 5.
Unexpectedly and repeatedly in SW480, raising the GCVconcentration from 20 to 40 mg=mL increased the cell via-bility. The reason for this anomaly was not determined.
FIG. 3. Staining results of normal colon cells (NCCs) transfected with different plasmids. The blue cells are the cells thathave expressed b-Gal under the control of the respective promoter. All pictures were taken with a Sony W camera (Japan).Red arrow, at the upper left corner of the first image (see online version for color), shows the size bar.
FIG. 2. Staining results of different cells transfected with different plasmids. The blue cells (see online version for color) arethe cells that have expressed b-Gal under the control of the respective promoter. All pictures were taken under a Zeiss invertmicroscope and were taken with a Motic camera, using Motic Image Plus 2 software. Arrows show the size bar.
FGF18 PROMOTER IN COLON CANCER SUICIDE GENE THERAPY 111
Staining the cells with M30 cytodeath antibody provedapoptosis in pUCCMVTK and pUCFGF18TKtransfected HCT116 cells but not in SW480
The one-tailed Student’s t-test analysis of the apoptosisinduced by GCV treatment of pUCCMVTK- andpUCFGF18TK-transfected HCT116 cells showed significantdifferences (P-values¼ 0.028709 and 0.038934, respectively),compared to nontreated cells (Fig. 6). In mock-transfectedHCT116, the rates of stained cells between GCV-treated anduntreated cells were not significantly different (P¼ 0.26064).
The mean (�standard deviation;) rates of apoptosis intreated pUCCMVTK- and pUCFGF18TK-transfected SW480cell lines were 16.89� 10.07 and 9.14� 1.56, respectively,whereas in nontreated cells transfected with the sameplasmids, the corresponding rates were 9.08� 3.85 and
8.37� 4.51, respectively. These rates between treated andnontreated cells were not significantly different (Fig. 6).
Discussion
In the systemic administration of suicidal gene constructs,cytotoxic effects of suicide=prodrug on normal cells can beextremely challenging, as reported in the hepatocellularcarcinoma model, where the cytotoxicity can induce severehepatitis.34 Studying cancerous cell-expression profiles andselecting the most cancer-specific promoters for inducing theexpression of suicide genes in the systemic administration ofdrugs is a rational approach for overcoming these deleteri-ous side-effects.35
Among various specific promoters activated in coloncancer cell lines, CEA is the most extensively studied. In vitro
FIG. 4. Results of b-Gal ELISA in HCT116, SW480, HUVEC, and normal colon cells (NCCs). Each graph shows the detailedresult of b-Gal ELISA and CAT ELISA from cell extract of cells transfected with reporter plasmids. The first bars in eachgroup are indicative of the maximum slope of b-Gal ELISA (b-Gal raw expression), the second bars are indicative of CATELISA maximum slope (CAT raw expression), and the third bars show the normalized expression of b-Gal by dividing the b-Gal maximum slope to the CAT maximum slope. The figures over b-Gal and CAT maximum slopes of pUCCMVLacZ- andpUCFGF18LacZ-transfectected cells are estimates of b-Gal and CAT raw expressions, based on plotting the maximum slopesof positive controls (serial dilutions of b-Gal and CAT). The line within each bar is the standard error.
112 TEIMOORI-TOOLABI ET AL.
studies have shown that despite its activity in non-CEA-producing cells, expression driven by this promoter did notreach the therapeutic index.36 Results on the efficacy of othercolon-cancer–specific promoters have been disappointing,except for the cyclooxygenase promoter.37 However, thispromoter is unsuitable for use in systemic suicide geneconstructs, since cyclooxygenase is a stress gene and ishighly expressed in blood cells.
Wnt is the most important signaling pathway to be ab-normally activated in colon cancer38; therefore, severalgroups have previously used the consensus TCF bindingsites activated by this pathway for colon-cancer–specificsuicide gene therapy.3,25–28
In the study conducted by Brunori et al., the E2 promoterof adenovirus was replaced with four TCF binding sites.25
This system showed both specificity and efficacy equal to the
FIG. 5. Viability graphs of HCT116 and SW480 cell lines transfected with pUCFGF18TK, pUCCMVTK (positive control),and negative control plasmids after treatment with different concentrations of GCV. The ratios of viability to the negativecontrol are shown in these diagrams.
FIG. 6. Percentages of stained cells with M30 cytodeath antibody measured by flow cytometry: 40mg=mL of ganciclovir hadbeen added to the ‘‘drug added’’ group, while the ‘‘no drug’’ group was just transfected with the respective plasmids and wasnot treated with ganciclovir. The first three groups of bars are the results of experiments on SW480, and the next three groupsare the results on HCT116; the line within each bar is the standard error.
FGF18 PROMOTER IN COLON CANCER SUICIDE GENE THERAPY 113

wild-type promoter; there was a 50–1000-fold reduction ofvirus replication in cell lines with low activation levels ofWnt, such as lung carcinoma (H1299) and normal fibroblast(WI38), when compared to SW480, which proved its speci-ficity.25
Placing five TCF binding sites adjacent to the minimalSV40 promoter showed measurable activity in several hu-man colon tumors; however, there was a low level of ex-pression in the nuclei of cells with normal b-catenin levels aswell.3 This promiscuous expression posed a challenge in thecase of highly lethal suicide genes, such as ricin and diph-theria toxin. Replacing the minimal SV40 promoter with theTATA box decreased the promiscuous expression of thispromoter and increased its specificity.26
On the other hand, a synthetic TCF=LEF1 binding siteplace upstream of the FADD (Fas-associated death domainprotein) suicide gene was reported to be highly effective incolon cancer cell lines, with no activity in normal cells.28
The present study was undertaken to assess the similari-ties and differences (if any) between a native, unalteredpromoter downstream of the Wnt signaling pathway andartificial promoters containing responsive elements to thispathway. Although Wnt is the most important pathway ac-tivated in colon cancer, there have been no reports analyzinga natural promoter downstream of this pathway for suicidegene therapy. Selection of the FGF18 promoter was based onan analysis of expression profiles that showed over-expression of this gene in adenomas and adenocarcinomaswith a mean ratio of 23.9� 11, compared to normal sectionsof colon.29,39 In addition, normal expression of this gene isrestricted to embryogenesis and developmental stages, andits lack of expression has been shown by northern blotanalysis24 in all adult tissues except the heart.
In this study, a fragment encompassing �425 to þ31 nu-cleotide of this promoter (to transcription start site) con-taining TCF=LEF1 site (�190)24 was placed upstream ofsuicide (TK) and reporter (b-Gal) genes. These constructswere delivered to the cells by transient transfection methods.Since introduction of these constructs to human or animalmodels will definitely exploit transient gene-delivery meth-ods (cationic agents, liposomes or adeno=adeno-associatedviral vectors), we preferred to deliver DNA to the cellstransiently. In addition to safety concerns, the therapeuticgene in this study was designed to kill the transfected cells,which did not warrant the long-term expression of TK. Be-sides, an important anticipated mechanism of action ofTK=GCV system is the bystander effect. Stable transfectionrules out the possibility of including this effect while evalu-ating the actual potency of suicide gene therapy.
The activity of reporter and suicide constructs was studiedin SW480 and HCT116. In the HCT116 cell line with mutantb-catenin, APC has the ability of exporting b-catenin fromthe nucleus, thus the activation levels of TCF=LEF1 are lowerthan in other colon cancer lines, such as SW480. In cells suchas SW480 with mutant APC, b-catenin export does not occurnormally; therefore, TCF=LEF1 attains an enhanced activa-tion status.40
Analysis of FGF18 promoter activity in these cell linesshowed that it was capable of inducing cell death and re-porter gene expression even in the HCT116 cell line, whichhas lower levels of free b-catenin in the nucleus than inSW480.41
Comparable activity levels in HCT116 and SW480 havenot been reported in similar studies, which used artificialpromoters responsive to the Wnt signaling pathway. Al-though one of these artificial constructs showed activity inresected primary, secondary, and metastatic colon cancertissues,26 almost all these promoters were moderately effec-tive in colon cancer cell lines with lower activation levels ofWnt, such HT-29 and HCT116.25,26,28 However, this variationin results should be confirmed by direct comparison of thesetwo sets of promoters.
The strongest speculation about the cause of this dis-crepancy is that the FGF18 promoter might contain certainunknown responsive elements that may be activated byother deregulated pathways, such as k-ras (which is acti-vated in HCT116).42,43
In spite of the lower gene delivery efficiency for transfec-tion methods, compared to transduction methods, the ex-pression of the suicide gene in this study attained thetherapeutic level.
The cytotoxicity analysis confirmed the negative effect ofsuicide plasmids on cell viability, upon GCV treatment. Thereason for the increased viability of SW480 at the 40-mg=mLGCV concentration, which was observed repeatedly, re-quires elucidation.
Cytotoxicity assays showed the suicidal efficacy of plas-mids. However, to fully elucidate the mechanism behind thecell death, staining with M30 cytodeath antibody was used inpreference to Annexin-PI (propidium iodide), since the lattercan be influenced by stress associated with the trypsinizationof adherent cells.
Cell death was observed in both cell lines by cytotoxicityassays; however, staining with M30 cytodeath antibodyshowed that apoptosis had occurred in HCT116 (followingtransfection with suicide constructs and treatment withdrugs), but failed to prove this phenomenon in SW480 withthe same treatment. This discrepancy might be due to thefollowing: first, since the TK=GCV system acts on prolifer-ating cells,44 nonproliferating and=or low proliferating cellsdo not respond well to its cytotoxic effects. Therefore, in theSW480 cell line, which has a higher replication time thanHCT116, the apoptotic action of TK=GCV could not bedemonstrated by M30cytodeath antibody. Second, TK=GCVis known to induce both apoptosis and necrosis45; this mayalso rationalize the disparity between the death rates as-sessed by M30 cytodeath antibody or by cytotoxicity assay.
Further, M30 cytodeath antibody detects a neoepitope ofcytokeratine 18, when it is cleaved by caspase 346 in the earlyphase of apoptosis. In P53-deficient cell lines (nonfunctionalp53), such as SW480, the main mechanism of apoptosis in-duced by thymidine kinase=GCV is not through the activa-tion of caspase 3,47 while in P53-proficient cell lines (cell lineswith functional p53), such as HCT116, the mechanism of celldeath induced by TK=GCV is caspase 3 dependent.48 If thisassumption proves true upon further analysis, M30 cyto-death cannot be considered as a good marker of apoptosiswhen activation of caspase 3 is not the main mechanism ofapoptosis, as seen in P53-deficient cell lines.
The Wnt signaling pathway is shared in several types ofcancers; consequently, it would be useful to study the effectsof pUCFGF18LacZ and pUCFGF18TK in other cancer celllines, especially in hepatocellular carcinoma and melanoma,given that the Wnt signaling pathway is also activated in
114 TEIMOORI-TOOLABI ET AL.

these cancers.26 However, the levels of b-catenin in the nu-cleus determines how effective the FGF18 promoter wouldbe in different cell lines24,41; additionally, the effects of otherunknown elements on the activation of this promoter cannotbe ignored. Further, inserting other promoters from thispathway in reporter and suicide constructs may be useful inoptimizing Wnt signaling-based suicide gene therapy.
Conclusions
These preliminary data should be further confirmed bystudying the effect of this construct in other colon cancer celllines as well as in other normal cells. Specificity of FGF18promoter must be confirmed by its application in othernormal cells, since assessing the activity of the FGF18 pro-moter in HUVEC and NCC is not sufficient in proving itsspecificity. Also, this construct should be compared withartificial promoters containing TCF=LEF1 binding sites.
In vivo models must also be included in the design of anyfurther studies in order to realize the potential of this system.In addition, the application of viral vectors would help toapproach clinical models in a more practical way.
Acknowledgments
This study was supported by the Pasteur Institute of Iran.The authors thank Anis Jafari, Ph.D., (Molecular BiologyDepartment, Pasteur Institute of Iran) for the critical readingof the manuscript for this article and Amir Amanzadeh,D.V.M., (National Cell Bank of Iran, Pasteur Institute of Iran)for his kind technical assistance in this study and also allpersonnel of the 1st Ward of Surgery and 1st OperationRoom in Imam Khoemini Hospital, and, especially, Mrs.Maryam Noorayee Kia, Mrs. Leyli Ghaffarpoor, and AhmadKaviani, M.D., for their kind help in obtaining normal tissuesamples.
Disclosure Statement
No competing financial interests exist.
References
1. Sadjadi A, Nouraie M, Mohagheghi MA, et al. Cancer oc-currence in Iran in 2002, an international perspective. AsianPac J Cancer Prev 2005;6:359.
2. Parker SL, Tong T, Bolden S, et al. Cancer statistics, 1996. CACancer J Clin 1996;46:5.
3. Lipinski KS, Djeha AH, Ismail T, et al. High-level, beta-catenin=TCF-dependent transgene expression in secondarycolorectal cancer tissue. Mol Ther 2001;4:365.
4. Zeng ZJ, Li ZB, Luo SQ, et al. Retrovirus-mediated TK genetherapy of implanted human breast cancer in nude miceunder the regulation of Tet-On. Cancer Gene Ther2006;13:290.
5. Moolten FL. Tumor chemosensitivity conferred by insertedherpes thymidine kinase genes: Paradigm for a prospectivecancer control strategy. Cancer Res 1986;46:527.
6. Wiewrodt R, Amin K, Kiefer M, et al. Adenovirus-mediatedgene transfer of enhanced herpes simplex virus thymidinekinase mutants improves prodrug-mediated tumor cellkilling. Cancer Gene Ther 2003;10:353.
7. Konson A, Ben-Kasus T, Mahajna JA, et al. Herpes simplexvirus thymidine kinase gene transduction enhances tumor
growth rate and cyclooxygenase-2 expression in murinecolon cancer cells. Cancer Gene Ther 2004;11:830.
8. Freeman SM, Abboud CN, Whartenby KA, et al. The ‘‘by-stander effect’’: Tumor regression when a fraction of thetumor mass is genetically modified. Cancer Res 1993;53:5274.
9. Sterman DH, Recio A, Vachani A, et al. Long-term follow-upof patients with malignant pleural mesothelioma receivinghigh-dose adenovirus herpes simplex thymidine kinase=ganciclovir suicide gene therapy. Clin Cancer Res 2005;11:7444.
10. Freytag SO, Stricker H, Peabody J, et al. Five-year follow-upof trial of replication-competent adenovirus-mediated sui-cide gene therapy for treatment of prostate cancer. Mol Ther2007;15:636.
11. Shalev M, Kadmon D, Teh BS, et al. Suicide gene therapytoxicity after multiple and repeat injections in patients withlocalized prostate cancer. J Urol 2000;163:1747.
12. Rainov NG. A phase III clinical evaluation of herpes simplexvirus type 1 thymidine kinase and ganciclovir gene therapyas an adjuvant to surgical resection and radiation in adultswith previously untreated glioblastoma multiforme. HumGene Ther 2000;11:2389.
13. Zabala M, Wang L, Hernandez-Alcoceba R, et al. Optimi-zation of the Tet-on system to regulate interleukin 12 ex-pression in the liver for the treatment of hepatic tumors.Cancer Res 2004;64:2799.
14. Ikegami S, Tadakuma T, Yamakami K, et al. Selective genetherapy for prostate cancer cells using liposomes conjugatedwith IgM-type monoclonal antibody against prostate-specific membrane antigen. Hum Cell 2005;18:17.
15. Nettelbeck DM, Jerome V, Muller R. Gene therapy: Designerpromoters for tumour targeting. Trends Genet 2000;16:174.
16. Sieger S, Jiang S, Kleinschmidt J, et al. Tumor-specific geneexpression using regulatory elements of the glucose trans-porter isoform 1 gene. Cancer Gene Ther 2004;11:41.
17. Pang S, Taneja S, Dardashti K, et al. Prostate tissue speci-ficity of the prostate-specific antigen promoter isolated froma patient with prostate cancer. Hum Gene Ther 1995;6:1417.
18. Richards CA, Austin EA, Huber BE. Transcriptional regu-latory sequences of carcinoembryonic antigen: Identificationand use with cytosine deaminase for tumor-specific genetherapy. Hum Gene Ther 1995;6:881.
19. Polakis P. The oncogenic activation of beta-catenin. CurrOpin Genet Dev 1999;9:15.
20. Polakis P. Wnt signaling and cancer. Genes Dev 2000;14:1837.
21. Huang M, Wang Y, Sun D, et al. Identification of genesregulated by Wnt=beta-catenin pathway and involved inapoptosis via microarray analysis. BMC Cancer 2006;6:221.
22. Han JC, Zhang KL, Chen XY, et al. Expression of sevengastric cancer-associated genes and its relevance for Wnt,NF-kappaB, and Stat3 signaling. APMIS 2007;115:1331.
23. Conacci-Sorrell ME, Ben-Yedidia T, Shtutman M, et al. Nr-CAM is a target gene of the beta-catenin=LEF-1 pathway inmelanoma and colon cancer and its expression enhancesmotility and confers tumorigenesis. Genes Dev 2002;16:2058.
24. Shimokawa T, Furukawa Y, Sakai M, et al. Involvement ofthe FGF18 gene in colorectal carcinogenesis, as a noveldownstream target of the beta-catenin=T-cell factor complex.Cancer Res 2003;63:6116.
25. Brunori M, Malerba M, Kashiwazaki H, et al. Replicatingadenoviruses that target tumors with constitutive activationof the wnt signaling pathway. J Virol 2001;75:2857.
FGF18 PROMOTER IN COLON CANCER SUICIDE GENE THERAPY 115

26. Lipinski KS, Djeha HA, Gawn J, et al. Optimization of asynthetic beta-catenin-dependent promoter for tumor-specific cancer gene therapy. Mol Ther 2004;10:150.
27. Kwong KY, Zou Y, Day CP, et al. The suppression of coloncancer cell growth in nude mice by targeting beta-catenin=TCF pathway. Oncogene 2002;21:8340.
28. Chen RH, McCormick F. Selective targeting to the hyper-active beta-catenin=T-cell factor pathway in colon cancercells. Cancer Res 2001;61:4445.
29. Sonvilla G, Allerstorfer S, Stattner S, et al. FGF18 in colo-rectal tumour cells: Autocrine and paracrine effects. Carcino-genesis 2008;29:15.
30. Dvory-Sobol H, Sagiv E, Kazanov D, et al. Targeting theactive beta-catenin pathway to treat cancer cells. Mol CancerTher 2006;5:2861.
31. Haque T, Nakada S, Hamdy RC. A review of FGF18: Itsexpression, signaling pathways, and possible functionsduring embryogenesis and postnatal development. HistolHistopathol 2007;22:97.
32. Sambrook J, Russell DW. Molecular Cloning: A laboratoryManual, 3rd ed. Cold Spring Harbor, NY: Cold SpringHarbor Laboratory Press, 2001.
33. Azadmanesh K, Rohvand F, Amini S, et al. Evaluation ofstimulatory effects of HTLV-I tax protein on CREB andNFKB related signaling pathways two B-glycosidase basedreporter plasmids. Yakhteh Med J 2005;6:218.
34. Bilbao R, Gerolami R, Bralet MP, et al. Transduction efficacy,antitumoral effect, and toxicity of adenovirus-mediated her-pes simplex virus thymidine kinase=ganciclovir therapy ofhepatocellular carcinoma: The woodchuck animal model.Cancer Gene Ther 2000;7:657.
35. Tanyi JL, Lapushin R, Eder A, et al. Identification of tissue-and cancer-selective promoters for the introduction of genesinto human ovarian cancer cells. Gynecol Oncol 2002;85:451.
36. Hauck W, Stanners CP. Transcriptional regulation of thecarcinoembryonic antigen gene. Identification of regulatoryelements and multiple nuclear factors. J Biol Chem 1995;270:3602.
37. Yamamoto M, Alemany R, Adachi Y, et al. Characterizationof the cyclooxygenase-2 promoter in an adenoviral vectorand its application for the mitigation of toxicity in suicidegene therapy of gastrointestinal cancers. Mol Ther 2001;3:385.
38. Bienz M, Clevers H. Linking colorectal cancer to Wnt sig-naling. Cell 2000;103:311.
39. Lin YM, Furukawa Y, Tsunoda T, et al. Molecular diagnosisof colorectal tumors by expression profiles of 50 genes ex-pressed differentially in adenomas and carcinomas. Onco-gene 2002;21:4120.
40. Malerba M, Nikolova D, Cornelis J, et al. Targeting of au-tonomous parvoviruses to colon cancer by insertion of Tcfsites in the P4 promoter. Cancer Gene Ther 2006;13:273.
41. Rosin-Arbesfeld R, Cliffe A, Brabletz T et al. Nuclear exportof the APC tumour suppressor controls beta-catenin func-tion in transcription. EMBO 2003;J22:1101.
42. Ohmori M, Shirasawa S, Furuse M, et al. Activated Ki-rasenhances sensitivity of ceramide-induced apoptosis withoutc-Jun NH2-terminal kinase=stress-activated protein kinaseor extracellular signal-regulated kinase activation in humancolon cancer cells. Cancer Res 1997;57:4714.
43. Shirasawa S, Furuse M, Yokoyama N, et al. Altered growthof human colon cancer cell lines disrupted at activatedKi-ras. Science 1993;260:85.
44. Uch R, Gerolami R, Faivre J, et al. Hepatoma cell-specificganciclovir-mediated toxicity of a lentivirally transducedHSV-TkEGFP fusion protein gene placed under the controlof rat alpha-fetoprotein gene regulatory sequences. CancerGene Ther 2003;10:689.
45. Thust R, Tomicic M, Klocking R, et al. Comparison of thegenotoxic and apoptosis-inducing properties of ganciclovirand penciclovir in Chinese hamster ovary cells transfectedwith the thymidine kinase gene of herpes simplex virus-1:Implications for gene therapeutic approaches. Cancer GeneTher 2000;7:107.
46. Caulin C, Salvesen GS, Oshima RG. Caspase cleavage ofkeratin 18 and reorganization of intermediate filamentsduring epithelial cell apoptosis. J Cell Biol 1997;138:1379.
47. Beltinger C, Fulda S, Kammertoens T, et al. Herpes simplexvirus thymidine kinase=ganciclovir-induced apoptosis in-volves ligand-independent death receptor aggregation andactivation of caspases. Proc Natl Acad Sci U S A 1999;96:8699.
48. Wei SJ, Chao Y, Shih YL, et al. Involvement of Fas(CD95=APO-1) and Fas ligand in apoptosis induced byganciclovir treatment of tumor cells transduced with herpessimplex virus thymidine kinase. Gene Ther 1999;6:420.
116 TEIMOORI-TOOLABI ET AL.




















