
Urokinase versus VATS for treatment of empyema: a randomized multicenter clinical trial
Upload
independentCategory
view
0download
0
Structural and functional characterization of the humanPAX7 5 0-flanking regulatory region
Yana V. Syagailoa,1, Olga Okladnovaa,2, Ella Reimera, Marcus Graßlea, Rainald Mossnera,Stefan Gattenlohnerb, Alexander Marxb, Jobst Meyera, Klaus-Peter Lescha,*
aDepartment of Psychiatry and Psychotherapy, University of Wurzburg, Fuchsleinstrasse 15, 97080 Wurzburg, GermanybInstitute of Pathology, University of Wurzburg, Wurzburg, Germany
Received 28 July 2001; received in revised form 11 June 2002; accepted 19 June 2002
Received by W.-H. Li
Abstract
The human PAX7 gene is a member of the paired box containing gene family of transcription factors implicated in development of the
skeletal muscle of the trunk and limbs as well as elements of the central nervous system. To understand the molecular mechanisms involved
in its expression, we have localized the transcription start sites in adult skeletal muscle and functionally characterized the 5 0-flanking
regulatory region responsible for PAX7 expression in this tissue. The major transcription start was identified 664 bp upstream from the
ATG codon using primer extension and 5 0-rapid amplification of cDNA ends (5 0-RACE). Analysis of the 5 0-flanking sequence revealed the
absence of a TATA-box and the presence of an inverted CCAAT-box. Several consensus sites for common transcriptional regulators
including Oct-1, NF1, AP2, AP4, CREB, Sp1, Nkx2.5, and MyoD are present in the promoter region. To determine the sites critical for
the function of the PAX7 promoter, a series of deletion fragments of the 5 0-flanking region were cloned adjacent to luciferase reporter gene
and expressed in RD, Cos-7 and JAR cell lines. The maximal promoter activity was achieved by a fragment extending from the position
2403 to 1373. No strong positive or negative regulatory elements were discovered by adding of further sequences (up to 2.97 kb). A
polymorphic (CCT)n repeat sequence was found 107 bp upstream of the transcription initiation site. PCR-based systematic screening for
length variations in 227 unrelated individuals of a Caucasian population showed a bimodal distribution of three alleles containing 8, 10 or 11
repeat units. When different variants of this PAX7 gene-linked polymorphic region (PAX7-LPR) were fused to a luciferase reporter gene and
transfected into RD cells, the variant with 11 repeat units revealed higher transcriptional efficiency compared to the 8 or 10 repeat alleles.
q 2002 Elsevier Science B.V. All rights reserved.
Keywords: Transcription regulation; Promoter; Paired box genes; Trinucleotide repeat polymorphism
1. Introduction
Nine members of the Pax gene family, Pax1 to Pax9,
have been identified in mice and humans (Gruss and
Walther, 1992; Tremblay and Gruss, 1994). These genes
exhibit spatial and temporal regulation during development
and encode proteins acting as transcriptional regulators. The
common feature of these proteins is a conserved 128-amino
acid DNA-binding domain, the paired domain, initially
found in some Drosophila segmentation genes (Bopp et
al., 1986; Walther et al., 1991). Mutations in several Pax
genes have been found in both congenital mouse and human
diseases, and have been shown to disrupt the developmental
processes in tissues in which they are expressed (Gruss and
Walther, 1992; Chalepakis et al., 1993; Strachan and Read,
1994).
In the developing mouse embryo, Pax7 is expressed from
Gene 294 (2002) 259–268
0378-1119/02/$ - see front matter q 2002 Elsevier Science B.V. All rights reserved.
PII: S0378-1119(02)00798-9
www.elsevier.com/locate/gene
Abbreviations: ANOVA, analysis of variance; bp, base pair(s); cDNA,
DNA complementary to RNA; cpm, counts per minute; dNTP, deoxyribo-
nucleoside triphosphate; d.p.c., day post coitum; DTT, dithiothreitol;
EDTA, ethylenediaminetetraacetic acid; ENase, restriction endonuclease;
EtdBr, ethidium bromide; FCS, fetal calf serum; FKHR, fork head domain
in rhabdomyosarcoma gene; kb, kilobase(s) or 1000 bp; nt, nucleotide(s);
Pax, paired box-containing gene; PAX7-LPR, PAX7 gene-linked poly-
morphic region; PBS, phosphate-buffered saline; PCR, polymerase chain
reaction; PMA, phorbol 12-myristate 13-acetate; RACE, rapid amplifica-
tion of cDNA ends; RNase, ribonuclease; RT-PCR, reverse transcription
PCR; SDS, sodium dodecyl sulfate; UTR, untranslated region
* Corresponding author. Tel.: 149-931-201-77-600; fax: 149-931-201-
77-620.
E-mail address: [email protected] (K.-P. Lesch).1 Present address: German Cancer Research Center; Heidelberg,
Germany.2 Present address: Institute of Oncological Chemistry, University of
Dusseldorf, Dusseldorf, Germany.

day 9 post coitum (d.p.c.) in the ventricular zone of the
neural tube, which generates a stem cell population for the
entire central nervous system, and dermomyotomal parts of
the somites, from which muscle cells derive (Jostes et al.,
1991). At early stages Pax7 expression is also detected in all
brain vesicles, but later (11 d.p.c.) this expression is
retracted to the mesencephalon (Jostes et al., 1991; Stoy-
kova and Gruss, 1994). No spontaneous mutations in Pax7/
PAX7 were found. Mice with a targeted inactivation of Pax7
have been generated (Mansouri et al., 1996). Homozygous
knockout animals die within the first three postnatal weeks
and show malformations in facial skeletal structures. Cell
culture and electron microscopic analysis revealed a
complete absence of satellite cells in Pax72/2 skeletal
muscle, whereas the proportion of muscle-derived stem
cells was unaffected. In contrast to its paralogue, Pax3,
which plays an essential role in regulating the developmen-
tal program of embryonic myoblasts (Maroto et al., 1997;
Tajbakhsh et al., 1997), Pax7 is expressed in satellite cells
residing in the adult muscle (Seale et al., 2000). Interest-
ingly, differential expression of alternately spliced Pax7
transcripts correlates with muscle regenerative efficiency
in different strains of mice (Kay et al., 1995, 1998).
A specific chromosome translocation, t(1;13)(p36;q14),
resulting in the fusion of PAX7 to a fork head domain
gene (FKHR), has been implicated in the genesis of alveolar
rhabdomyosarcoma (Davis et al., 1994). The resulting
chimeric protein is a more potent transcriptional activator
compared to the wild-type Pax7 protein and possibly acts by
suppressing the apoptotic program normally responsible for
elimination of aberrant cells (Bernasconi et al., 1996). Cells
of other tumors, embryonal rhabdomyosarcoma and
Ewing’s sarcoma, were found to express wild-type PAX7
at elevated levels in comparison with primary human
myoblasts (Bernasconi et al., 1996; Barr et al., 1999).
Thus, PAX7 gene expression, which is rigorously controlled
during normal tissue development, is abnormally regulated
in certain pathological states. Therefore, in order to eluci-
date the molecular mechanisms controlling the expression
of the human PAX7 gene in normal and pathological tissues,
we have determined the transcription start site(s) and carried
out the structural and functional analysis of the regulatory
region responsible for its expression in adult skeletal
muscle. Finally, a polymorphic repetitive sequence located
within the basal promoter and its functional assessment are
reported.
2. Materials and methods
2.1. Isolation and sequencing of the 5 0-flanking regulatory
region
The human genomic PAC library no. 704 (Resource
Center/Primary Database of the German Human Genome
Project, Berlin), constructed by ligation of DNA from a
human male fibroblast cell line into pCYPAC-2 vector,
was screened under high-stringency hybridization condi-
tions with a 386 bp 32P-labeled EcoRI-SspI DNA fragment
encompassing a part of the human PAX7 promoter sequence
deposited in the EMBL/GenBank database (Accession
number: X96744). PAC DNA preparation from a single
positive clone (RPCIP704B248Q25), restriction endonu-
clease (ENase) mapping, and subcloning of restriction frag-
ments were carried out according to established protocols.
Fragments containing exon 1 and the 5 0-flanking sequences
were identified by Southern analysis and subcloned into
pBluescript II KS(1) vector (Stratagene). The 3 0 end of
the PAC clone was identified by Southern analysis using
four different short cDNA fragments containing the exons
1, 2, 3 and 4 as probes. Double-stranded DNA was
sequenced by the dideoxy chain termination method with
flanking reverse and forward primers as well as by primer
walking. Sequence analysis of the 5 0-regulatory region was
performed using on-line program MatInspector V2.2 (http://
transfac.gbf.de/TRANSFAC/index.html). The sequence
was deposited in the EMBL/GenBank database (Accession
number: AJ130875).
2.2. Northern blot
Human Multiple Tissue Northern (MTN) Blot membrane
(Clontech) with equal amounts of poly(A)1 RNA (2 mg)
from various peripheral tissues was hybridized with a 573
bp 32P-labeled fragment of human PAX7 cDNA using the
protocol provided by the manufacturer. The cDNA probe
was amplified by PCR with primers P7-1 (5 0-GCGCAAG-
CAGCGACGCAGTCG) and P7-2 (5 0-GAGAAGTCAGC-
CTGTGGCTGCG). Hybridized membrane was exposed to
X-ray film (Kodak XR) at 2708C.
2.3. RT-PCR amplification
For RT-PCR analysis, 5 mg of total RNA isolated from
frontal, temporal, occipital and entorhinal cortex, cerebel-
lum, thalamus, subthalamic nuclei, substantia nigra, hippo-
campus, putamen, amygdala and caudate nucleus were
reverse transcribed with 200 units of Superscript II reverse
transcriptase (Gibco BRL) in a 20 ml reaction volume
following the manufacturer’s recommended protocol. 3 ml
of this reaction was used as template for PCR with the
paired-box-specific primers P7-3 (5 0-TTGCCG-CTAC-
CAGGAGACCG) and P7-4 (5 0-CCAGGATGCC-
GTCGATGCTG) and b-actin specific primers (5 0-CCT-
CGCCTTTGCCGATCC; 5 0-GGATCTTCATGAGGTAG-
TCAGTC). PCR (40 s at 948C, 30 s at 588C, 60 s at 728C
for 35 cycles) was performed in a final volume of 50 ml
containing 20 pmol of each primer, 200 mM of each
dNTP in 1.5 mM MgCl2, 20 mM (NH4)2SO4, 75 mM
Tris–HCl (pH 9.0), 0.01% Tween-20, and 0.5 units of Taq
DNA polymerase (Eurogentec). Fragments were separated
on a 2% agarose gel stained with EtdBr.
Y.V. Syagailo et al. / Gene 294 (2002) 259–268260

2.4. Primer extension analysis
Antisense oligonucleotides P7-5 (5 0-AGCCGGCCGCA-
TCATTCTCGGTAC) and P7-6 (5 0-GACCACACTCGCT-
TCTCCCTCCTC) were end-labeled with [g-32P]ATP (3000
Ci/mmol) using T4 polynucleotide kinase (Promega). 0.5
mg of poly(A)1 RNA from human adult skeletal muscle
or 30 mg of yeast tRNA were mixed with the labeled primer
(1.0 £ 105 cpm) in 20 ml of hybridization buffer (225 mM
KCl and 10 mM Tris–HCl (pH 8.3)), denatured at 858C for
10 min, and incubated at 658C for 30 min. The hybridization
mix was slowly cooled to 378C for 60 min, and 40 ml of the
reverse transcription mixture (50 mM Tris–HCl (pH 8.3), 75
mM KCl, 3 mM MgCl2, 7.5 mM DTT, 0.75 mM dNTPs, and
30 units of RNasin) was then added. Primer was extended by
200 units of Superscript II reverse transcriptase (Gibco
BRL) at 428C for 60 min, and reactions were terminated
by addition of 5 ml of 0.5 M EDTA (pH 8.0) and 0.5 ml
of 10 mg/ml RNase A1. After incubation at 378C for 30 min,
extended fragments were purified by phenol-chloroform
extraction, analyzed on 6% polyacrylamide/8 M urea gels,
and sized against sequencing reactions.
2.5. 5 0-Rapid amplification of cDNA ends (5 0-RACE)
5 0-RACE was performed with 1 mg of poly(A)1 RNA
using the Marathon RACE kit (Clontech) according to the
manufacturer’s instructions. Initial amplification was
carried out with gene-specific primer P7-7 (5 0-GCCTTCT-
TTCTCCGGACCACACTC), followed by nested PCR
using gene-specific primer P7-6. PCR products were cloned
in pGEM-T vector (Promega) and automatically sequenced.
2.6. Trinucleotide repeat genotyping
Genomic DNA from 227 unrelated healthy volunteers
(133 males, mean age 28.7 ^ 7.9 years; 94 females,
32.3 ^ 11.6 years) was extracted from whole blood. The
study was approved by the ethics committee of the Univer-
sity of Wurzburg, and informed consent was obtained from
all subjects. Primers P7-8 (5 0-CGTTATTGGTCCTCCG-
CTC) and P7-9 (5 0-CGCGAGTGATCAGCTGGGTA)
flanking the PAX7 gene-linked polymorphic region
(PAX7-LPR) located directly upstream of the transcription
start site were used to generate fragments containing the
(CCT)n trinucleotide repeat. PCR (40 s at 948C, 30 s at
598C, 60 s at 728C for 38 cycles) was performed in a final
volume of 25 ml containing 60 ng genomic DNA, 10 pmol
of each primer, 200 mM of each dNTP in 1.5 mM MgCl2, 20
mM (NH4)2SO4, 75 mM Tris–HCl (pH 9.0), 0.01% Tween-
20, 2 mCi of [a-32P]dCTP (10 mCi/ml, 3000 Ci/mmol), and
0.2 units of Taq DNA polymerase (Eurogentec). Products
were fractionated by electrophoresis through 6% sequen-
cing gels.
2.7. Construction of reporter gene plasmids
The 2.97 kb XbaI-NaeI fragment including part of exon 1
and its upstream region was subcloned into the promoter-
less, luciferase reporter gene vector pGL3 basic (Promega)
in sense orientation and designated pXN. Serial deletion
fragments of pXN, HindIII-NaeI (21605 to 1373, pHN),
BglII-NaeI (21388 to 1373, pBgN), PstI-NaeI (21159 to
1373, pPN), EcoRI-NaeI (2633 to 1373, pRIN), BamHI-
NaeI (2403 to 1373, pBN), PvuII-NaeI (270 to 1373,
pPvN), and SmaI-NaeI (142 to 1373, pSN) were also
studied. To obtain construct pXND, pXN was digested
with PstI and re-ligated resulting in a plasmid containing
the 2.97 kb XbaI-NaeI fragment lacking 1.18 kb. Fragments
BamHI-NaeI (pNB), and PvuII-NaeI (pNPv) were also
fused to the luciferase gene in an antisense orientation.
The 1.0 kb EcoRI-NaeI (pRIN) fragment contains 10 CCT
repeats (allele 10, (CCT)10). For generation of constructs
with different length of repetitive sequence, genomic
DNAs with alleles containing length variants of the
(CCT)n repeat were amplified with primers P7-10 (5 0-
GCTGCAGGAATTCCTCCCTTC) and P7-11 (5 0-GCGA-
GTGATCAGCTGGGTAAG). EcoRI and PvuII-digested
products (alleles 8 and 11) were then ligated into pRIN.
All constructs were verified by cycle sequencing.
2.8. Cell culture
Cos-7 (African green monkey kidney, DSMZ ACC60)
cells were grown in Dulbecco’s modified Eagle’s medium
supplemented with 10% fetal calf serum (FCS), 50 mg/ml
streptomycin and 50 units/ml penicillin. JAR cells (human
placental choriocarcinoma, ATCC HTB-144) were grown in
RPMI 1640 medium supplemented with 10% FCS, 50 mg/
ml streptomycin and 50 units/ml penicillin. RD cells
(human embryonal rhabdomyosarcoma, ECACC 85111
502) were maintained in EMEM medium supplemented
with 2 mM glutamine, 2% NEAA, 2% vitamins, 10%
FCS, 50 mg/ml streptomycin and 50 units/ml penicillin.
All cells were cultured at 378C in a humidified 5% CO2
atmosphere.
2.9. Transfection and luciferase assays
Transfections were performed by the lipofection method
(Roche) according to standard procedures. Briefly, approxi-
mately 6 £ 105 cells on 60 mm plates were cotransfected
with 2.5 mg of luciferase constructs and 0.5 mg of
pRSV40-lacZ plasmid to control for transfection efficiency.
Cells were harvested in 250 ml lysis buffer (Roche) 40 h
after transfection. Luciferase assays were conducted with
100 ml of cell extracts using luciferin reagent (Promega).
Chemoluminescence was counted for 10 s in a liquid scin-
tillation spectrophotometer. The extracts (10 ml) were also
tested for b-galactosidase activity, and luciferase activities
were normalized to b-galactosidase activities using equal
Y.V. Syagailo et al. / Gene 294 (2002) 259–268 261
amounts of total protein determined by the method of Brad-
ford.
3. Results
3.1. Cloning of the 5 0-flanking regulatory region of the
human PAX7 gene
Screening of the human PAC library with a probe target-
ing a part of the human PAX7 5 0-flanking region yielded a
single ,70 kb clone covering the first three exons of PAX7
including its 5 0-flanking regulatory region. The partial
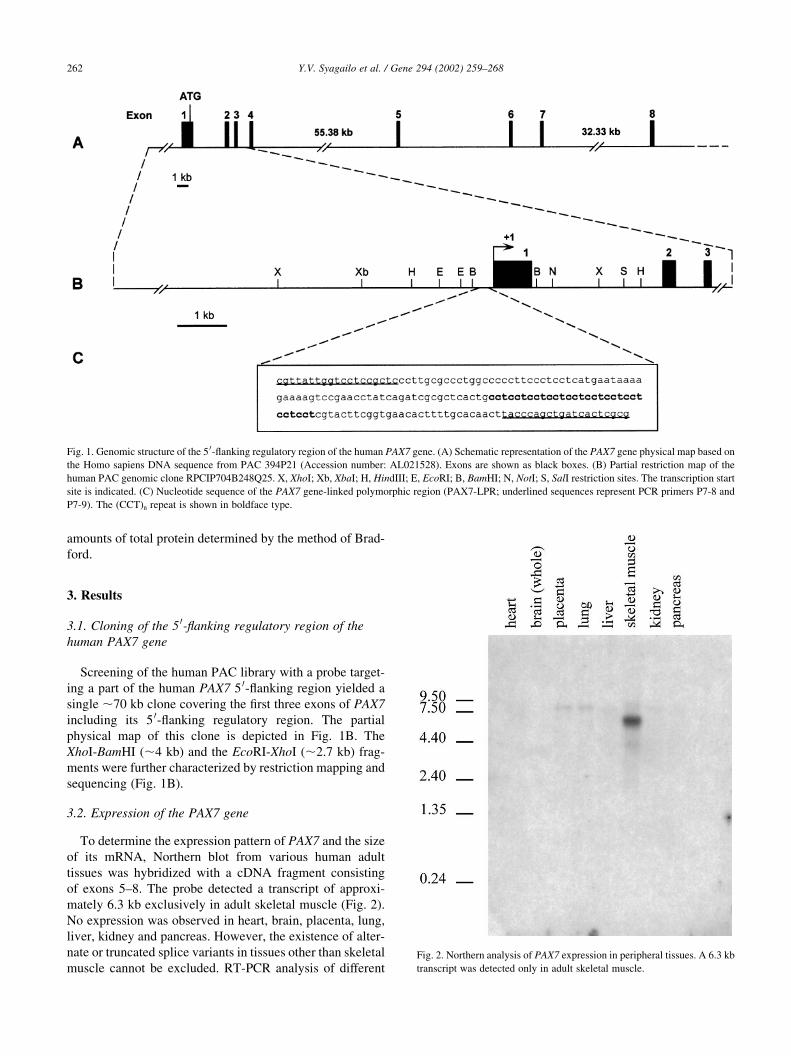
physical map of this clone is depicted in Fig. 1B. The
XhoI-BamHI (,4 kb) and the EcoRI-XhoI (,2.7 kb) frag-
ments were further characterized by restriction mapping and
sequencing (Fig. 1B).
3.2. Expression of the PAX7 gene
To determine the expression pattern of PAX7 and the size
of its mRNA, Northern blot from various human adult
tissues was hybridized with a cDNA fragment consisting
of exons 5–8. The probe detected a transcript of approxi-
mately 6.3 kb exclusively in adult skeletal muscle (Fig. 2).
No expression was observed in heart, brain, placenta, lung,
liver, kidney and pancreas. However, the existence of alter-
nate or truncated splice variants in tissues other than skeletal
muscle cannot be excluded. RT-PCR analysis of different
Y.V. Syagailo et al. / Gene 294 (2002) 259–268262
Fig. 1. Genomic structure of the 5 0-flanking regulatory region of the human PAX7 gene. (A) Schematic representation of the PAX7 gene physical map based on
the Homo sapiens DNA sequence from PAC 394P21 (Accession number: AL021528). Exons are shown as black boxes. (B) Partial restriction map of the
human PAC genomic clone RPCIP704B248Q25. X, XhoI; Xb, XbaI; H, HindIII; E, EcoRI; B, BamHI; N, NotI; S, SalI restriction sites. The transcription start
site is indicated. (C) Nucleotide sequence of the PAX7 gene-linked polymorphic region (PAX7-LPR; underlined sequences represent PCR primers P7-8 and
P7-9). The (CCT)n repeat is shown in boldface type.
Fig. 2. Northern analysis of PAX7 expression in peripheral tissues. A 6.3 kb
transcript was detected only in adult skeletal muscle.

regions of human adult brain using paired-box-specific
primers revealed transcripts in cerebellum and subthalamic
nuclei, as well as weak bands in temporal and frontal cortex,
thalamus, amygdala and putamen (Fig. 3).
3.3. Mapping of transcription start sites
Murmann et al. (2000) determined a major transcription
start site 70 bp upstream of the first ATG in the C2C12 mouse
myoblasts and tumor A673 cell lines, while the longest 5 0-
UTR (599 bp) was described for RNA isolated from alveolar
rhabdomyosarcoma tumor (Vorobyov et al., 1997). There-
fore, in order to define the transcription initiation site(s) of
PAX7 in adult skeletal muscle, primer extension analysis was
performed with primers (P7-5 and P7-6) from two putative
candidate regions. The primer extension analysis with distal
P7-5 primer revealed no extension product within the region
presented by Murmann et al. (2000), but showed an extension
product of a higher molecular weight, out of range of the
sequencing ladder, suggesting the presence of a possible
start site in the region described for alveolar rhabdomyosar-
coma (Vorobyov et al., 1997) (data not shown). The primer
extension analysis with primer P7-6 from the proximal
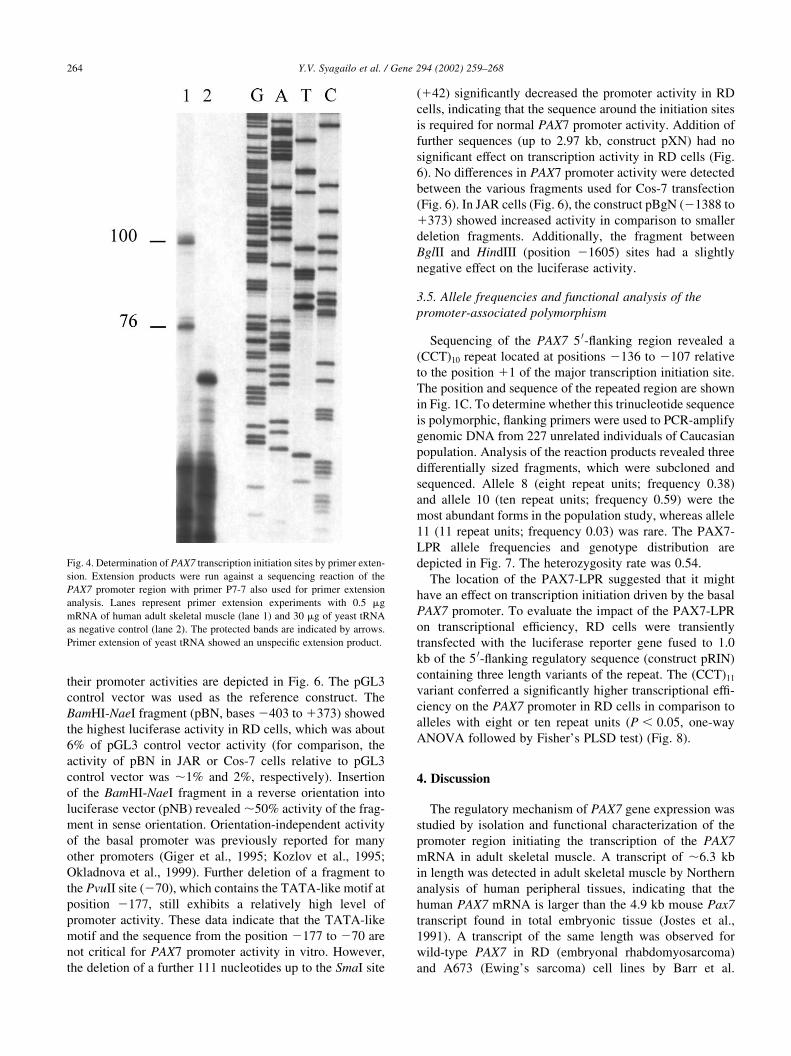
region yielded two extended products corresponding to size
of 76 and 100 nt (Fig. 4, lane 1). No specific signals were
observed in the negative control using yeast tRNA (Fig. 4,
lane 2). In order to confirm the results obtained with primer
extension, 5 0-RACE analysis using antisense primers speci-
fic for the proximal region was performed. 5 0-RACE consis-
tently produced a 91 bp fragment in two independent
experiments. The 5 0 end of this fragment revealed by
sequence analysis of ten clones and two primer extension
products are indicated in Fig. 5. The most upstream nucleo-
tide position found for the 5 0 end of the PAX7 cDNA initia-
tion site is labeled 11 in subsequent figures.
3.4. PAX7 promoter activity
Analysis of the 5 0-flanking sequence revealed a TATA-
like motif localized at position 2177 nt relative to the major
transcription initiation site (11). No obvious CCAAT
consensus was found within 100 bp 5 0 to the start site, but
one CCAAT box in reverse complementary orientation was
found 222 bp upstream of the 11 site (Fig. 5). Several
consensus sites for common transcription regulators includ-
ing Oct-1 (consensus binding motif NNGAATATKCANN-
NN), NF1 (core string TGGC), AP2 (MKCCCSCNGGCG),
AP4 (WGARYCAGCTGYGGNCNK), CREB (NNGNT-
GACGYNN), Sp1 (KRGGCGKRRY), Nkx2.5 (TYAAG-
TG), and MyoD (NNCANTGNY) (Quandt et al., 1995)
were mapped to the promoter region. The second consensus
site for a TATA box (ataTATAaataaata) motif was found at
position 1153 relative to the initiation start site. To identify
the DNA regions responsible for the control of PAX7 gene
transcription we analyzed the ability of serially truncated
fragments from the 5 0 end of PAX7 to drive the expression
of a luciferase reporter gene. Previous data (Schafer et al.,
1994) prompted us to choose the rhabdomyosarcoma cell
line (RD), which displays robust constitutive PAX7 expres-
sion, as an in vitro model for reporter gene experiments.
Additionally, JAR cells which are devoid of endogenous
PAX7 and Cos-7 cells expressing low levels of PAX7
mRNA (data not shown) were used for transient transfection
assays. Absolute activities of promoter constructs were up to
six-fold lower in JAR cells and up to three-fold lower in
Cos-7 compared to RD cells. The deletion constructs and
Y.V. Syagailo et al. / Gene 294 (2002) 259–268 263
Fig. 3. Expression of the PAX7 gene in human adult brain. Total RNA from temporal (1), occipital (9), frontal (11) and entorhinal (12) cortex, hippocampus (2),
thalamus (3), cerebellum (4), amygdala (5), substantia nigra (6), caudate nucleus (7), subthalamic nuclei (8) and putamen (10) was reverse-transcribed and
subjected to cDNA amplification using primers for PAX7 and b-actin. PCR products were separated on 2% agarose gel stained with EtdBr. DNA ladder
(Fermentas), which contains pUC19 plasmid DNA cut with MspI, was used as a size marker (M).
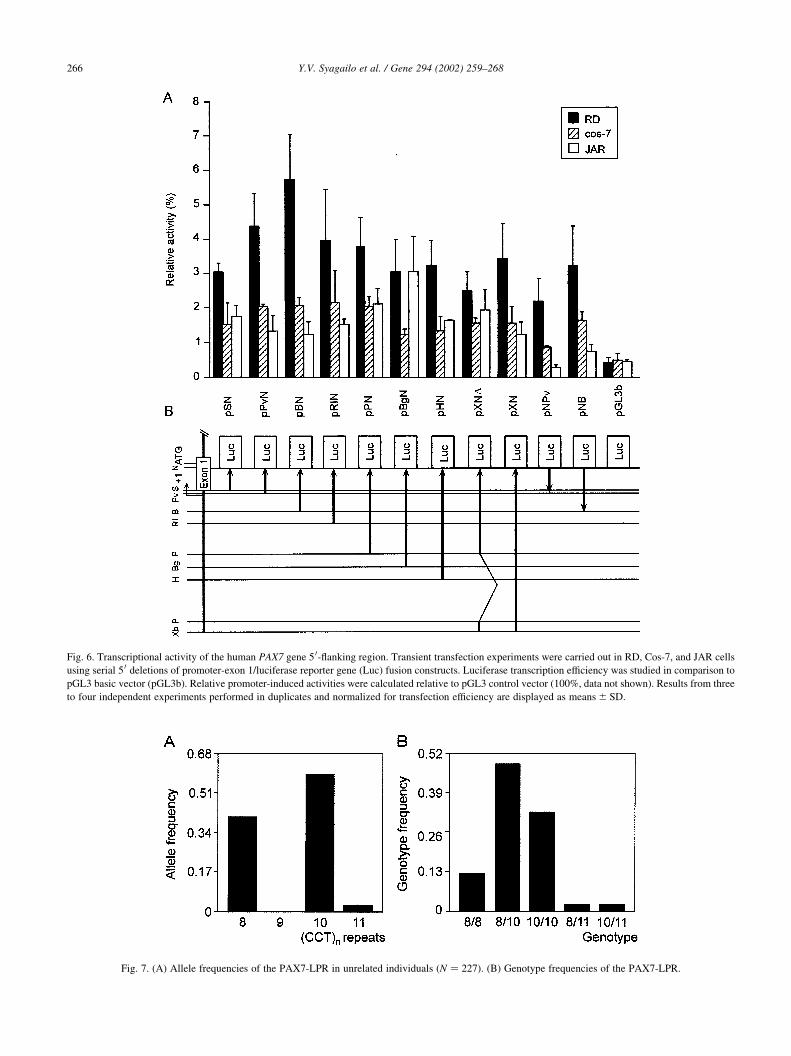
their promoter activities are depicted in Fig. 6. The pGL3
control vector was used as the reference construct. The
BamHI-NaeI fragment (pBN, bases 2403 to 1373) showed
the highest luciferase activity in RD cells, which was about
6% of pGL3 control vector activity (for comparison, the
activity of pBN in JAR or Cos-7 cells relative to pGL3
control vector was ,1% and 2%, respectively). Insertion
of the BamHI-NaeI fragment in a reverse orientation into
luciferase vector (pNB) revealed ,50% activity of the frag-
ment in sense orientation. Orientation-independent activity
of the basal promoter was previously reported for many
other promoters (Giger et al., 1995; Kozlov et al., 1995;
Okladnova et al., 1999). Further deletion of a fragment to
the PvuII site (270), which contains the TATA-like motif at
position 2177, still exhibits a relatively high level of
promoter activity. These data indicate that the TATA-like
motif and the sequence from the position 2177 to 270 are
not critical for PAX7 promoter activity in vitro. However,
the deletion of a further 111 nucleotides up to the SmaI site
(142) significantly decreased the promoter activity in RD
cells, indicating that the sequence around the initiation sites
is required for normal PAX7 promoter activity. Addition of
further sequences (up to 2.97 kb, construct pXN) had no
significant effect on transcription activity in RD cells (Fig.
6). No differences in PAX7 promoter activity were detected
between the various fragments used for Cos-7 transfection
(Fig. 6). In JAR cells (Fig. 6), the construct pBgN (21388 to
1373) showed increased activity in comparison to smaller
deletion fragments. Additionally, the fragment between
BglII and HindIII (position 21605) sites had a slightly
negative effect on the luciferase activity.
3.5. Allele frequencies and functional analysis of the
promoter-associated polymorphism
Sequencing of the PAX7 5 0-flanking region revealed a
(CCT)10 repeat located at positions 2136 to 2107 relative
to the position 11 of the major transcription initiation site.
The position and sequence of the repeated region are shown
in Fig. 1C. To determine whether this trinucleotide sequence
is polymorphic, flanking primers were used to PCR-amplify
genomic DNA from 227 unrelated individuals of Caucasian
population. Analysis of the reaction products revealed three
differentially sized fragments, which were subcloned and
sequenced. Allele 8 (eight repeat units; frequency 0.38)
and allele 10 (ten repeat units; frequency 0.59) were the
most abundant forms in the population study, whereas allele
11 (11 repeat units; frequency 0.03) was rare. The PAX7-
LPR allele frequencies and genotype distribution are
depicted in Fig. 7. The heterozygosity rate was 0.54.
The location of the PAX7-LPR suggested that it might
have an effect on transcription initiation driven by the basal
PAX7 promoter. To evaluate the impact of the PAX7-LPR
on transcriptional efficiency, RD cells were transiently
transfected with the luciferase reporter gene fused to 1.0
kb of the 5 0-flanking regulatory sequence (construct pRIN)
containing three length variants of the repeat. The (CCT)11
variant conferred a significantly higher transcriptional effi-
ciency on the PAX7 promoter in RD cells in comparison to
alleles with eight or ten repeat units (P , 0:05, one-way
ANOVA followed by Fisher’s PLSD test) (Fig. 8).
4. Discussion
The regulatory mechanism of PAX7 gene expression was
studied by isolation and functional characterization of the
promoter region initiating the transcription of the PAX7
mRNA in adult skeletal muscle. A transcript of ,6.3 kb
in length was detected in adult skeletal muscle by Northern
analysis of human peripheral tissues, indicating that the
human PAX7 mRNA is larger than the 4.9 kb mouse Pax7
transcript found in total embryonic tissue (Jostes et al.,
1991). A transcript of the same length was observed for
wild-type PAX7 in RD (embryonal rhabdomyosarcoma)
and A673 (Ewing’s sarcoma) cell lines by Barr et al.
Y.V. Syagailo et al. / Gene 294 (2002) 259–268264
Fig. 4. Determination of PAX7 transcription initiation sites by primer exten-
sion. Extension products were run against a sequencing reaction of the
PAX7 promoter region with primer P7-7 also used for primer extension
analysis. Lanes represent primer extension experiments with 0.5 mg
mRNA of human adult skeletal muscle (lane 1) and 30 mg of yeast tRNA
as negative control (lane 2). The protected bands are indicated by arrows.
Primer extension of yeast tRNA showed an unspecific extension product.

(1999). However, the longest wild-type PAX7 cDNA (2.2
kb), that was previously reported to contain the complete
coding region and 5 0 sequence, is considerably shorter
(Vorobyov et al., 1997). Therefore, in order to determine
the full length of the 5 0-UTR, we performed primer exten-
sion and 5 0-RACE. Primer extension analysis identified two
transcription start sites 640 bp and 664 bp upstream of the
ATG codon in adult skeletal muscle mRNA. The smaller
primer-extended product may represent an additional start
site or indicates mRNA secondary structure/degradation. A
transcriptional start site 655 bp upstream of the translation
initiation codon was mapped by 5 0-RACE with a slight
discrepancy (9 bp) in comparison to primer extension
results. The presence of GC-rich stretches in mRNA
sequences could result in the premature termination of the
reverse transcription of the long mRNA from poly(dT)
primer by 5 0-RACE procedure. Thus, PAX7 mRNA
contains a long 5 0-UTR encompassing 664 nucleotides.
Cellular mRNAs containing complex 5 0-UTR may be
post-transcriptionally regulated by their 5 0-UTR and are
Y.V. Syagailo et al. / Gene 294 (2002) 259–268 265
Fig. 5. Nucleotide sequence of the first exon in uppercase and a partial 5 0-flanking region of the human PAX7 gene (EMBL/GenBank accession number:
AJ130875). Restriction endonuclease sites used to generate deletion constructs are indicated. The transcription start sites detected by primer extension and 5 0-
RACE assays are denoted by arrows. Numbering of the sequence is relative to the major upstream transcription start site (11). Consensus sequences for DNA-
binding proteins (TATA-like motif; AP-2; AP-4; NF-1; MyoD; Oct1; Sp1; CCAAT; Nkx2.5) and the (CCT)n repeat are underlined.
Y.V. Syagailo et al. / Gene 294 (2002) 259–268266
Fig. 6. Transcriptional activity of the human PAX7 gene 5 0-flanking region. Transient transfection experiments were carried out in RD, Cos-7, and JAR cells
using serial 5 0 deletions of promoter-exon 1/luciferase reporter gene (Luc) fusion constructs. Luciferase transcription efficiency was studied in comparison to
pGL3 basic vector (pGL3b). Relative promoter-induced activities were calculated relative to pGL3 control vector (100%, data not shown). Results from three
to four independent experiments performed in duplicates and normalized for transfection efficiency are displayed as means ^ SD.
Fig. 7. (A) Allele frequencies of the PAX7-LPR in unrelated individuals (N ¼ 227). (B) Genotype frequencies of the PAX7-LPR.

often found to encode regulatory proteins, proto-oncogenes,
growth factors and their receptors, and homeodomain tran-
scription factors (van der Velden and Thomas, 1999).
In most eukaryotic genes, the TATA box is located 25–30
bp upstream of the mRNA initiation sites (Johnson and
McKnight, 1989) and directs accurate transcription initiation
through a mechanism that depends on its specific interaction
with a multiprotein complex called TFIID (Kaufmann and
Smale, 1994; Ghosh and Van Duyne, 1996). The A/T-rich
short TATA-like sequence (gaATAAAAgaaaa) found in the
PAX7 5 0-flanking region and located 177 bp upstream of the
transcription initiation site does not completely match the
TATA-box consensus sequence (STATAAAWRNNNN-
NNN) and is not critical for expression of the PAX7 gene
in vitro as was shown by functional analysis. Thus, in contrast
to the promoter of the human PAX3 gene, that contains
TATA and two CCAAT boxes upstream of the start site at
appropriate positions for classical promoter structure
(Okladnova et al., 1999), the PAX7 promoter lacks a canoni-
cal TATA box and resembles previously described TATA-
less human PAX2 and mouse Pax4 gene regulatory regions
(Stayner et al., 1998; Brink et al., 2001). Several potential
binding sites for Sp1, AP-2, AP-4, NF-1, MyoD, Oct1,
Nkx2.5, CREB, and CCAAT-binding (in negative orienta-
tion) transcription factors were identified upstream and
downstream of the transcription start site.
Functional mapping of the gene regulatory region
revealed the sequence between 270 and 1373 bp essential
for minimal basal activity of the PAX7 promoter. The maxi-
mal promoter activity was detected for pBN construct
(2403 to 1373 bp). Progressive addition of further
sequences (up to 2.97 kb, construct pXN) does not effec-
tively change the transcriptional activity detected with this
construct. Only the fragment between BamHI and EcoRI
sites (pRIN construct) has some regulatory elements,
which have a slightly negative effect on the promoter activ-
ity in RD cells. Our results suggest that the large region of
the human PAX7 promoter between XbaI and BamHI does
not contain any strong positive or negative regulatory
elements affecting the PAX7 control of transcription,
however, it is not unusual for tissue-specific regulatory
sequences to be located at a long distance from the tran-
scription start (Hardison et al., 1997; Jaakkola et al., 1997;
Maul et al., 1998). Further studies are required to define
factors necessary for PAX7 activation and regulation.
Our findings provide the first evidence for the expression
of the PAX7 gene in several regions of adult human brain.
The Pax7 gene is implicated in neurogenesis during devel-
opment and its expression was detected in the specific
regions of embryonic brain (Jostes et al., 1991; Stoykova
and Gruss, 1994) as well as in the adult brain of the mouse
(Ziman et al., 1997). Though Northern analysis with a speci-
fic probe from the 3 0 region of the gene showed no expres-
sion of PAX7 in adult human brain, the existence of alternate
or truncated transcripts could not be excluded. Indeed, RT-
PCR analysis with paired-box-specific primers indicated
PAX7 transcripts in cerebellar, temporal and frontal cortex,
subthalamic nuclei, thalamus, amygdala and putamen. Inter-
estingly, transcripts containing only the paired domain
encoding region of the PAX3 gene were found in many
human adult tissues including different brain regions
(Tsukamoto et al., 1994).
Results of our study have also demonstrated that the poly-
morphic (CCT)n repeat located 107 bp upstream of the tran-
scription start site within the core regulatory region shows
length variation in a western European population. Tandem
arrays of trinucleotides (triplet repeats) are abundant in
eukaryotic genomes and a subset of these repeats is asso-
ciated with the triplet expansion disorders (Mitas, 1997).
Intrastrand or interstrand folded structures, such as hairpins,
tetraplexes and triplexes have been described for the triplet
expansion disease-associated repeats (Kashi et al., 1997).
Such structures may promote triplet expansion by favoring
strand-slippage during DNA replication (Sinden and Wells,
1992). The presence of these structures on the template strand
may block the forward progress of DNA polymerase, thus
promoting dissociation of the polymerase and increasing the
likelihood that the nascent strand and the template strand will
slip relative to one another. Structures promoting the triplet
expansion may also block the forward progression of RNA
polymerase during transcription. Therefore, the PAX7
(CCT)n repetitive element described here may influence the
transcriptional efficiency of the PAX7 promoter. Population
and family-based association studies are currently under way
to determine the role of PAX7-LPR genotypes in neurode-
velopmental and neuromuscular disorders.
Acknowledgements
We thank G. Ortega and M. Schad for technical expertise,
G. Stober for sharing DNA samples, and J. Nicol for secre-
Y.V. Syagailo et al. / Gene 294 (2002) 259–268 267
Fig. 8. Transcriptional activity of PAX7-LPR alleles. The 1.0 kb fragments
EcoRI-NaeI containing 8, 10, or 11 repeat units had been fused to the
luciferase expression vector and transfected into RD cells. Luciferase activ-
ities of allelic constructs are calculated relatively to allele 10 (construct
pRIN) and displayed as means ^ SD. Allele 11 vs. allele 8 or allele 10,
**P , 0:05, one-way ANOVA followed by Fisher’s PLSD test.

tarial assistance. This work was supported by the Bundes-
ministerium fur Bildung, Wissenschaft, Forschung und
Technologie (BMBF, 512-4001-01KS9603 and
01GA9802/5).
References
Barr, F.G., Fitzgerald, J.C., Ginsberg, J.P., Vanella, M.L., Davis, R.J.,
Bennicelli, J.L., 1999. Predominant expression of alternative PAX3
and PAX7 forms in myogenic and neural tumor cell lines. Cancer
Res. 59, 5443–5448.
Bernasconi, M., Remppis, A., Fredericks, W.J., Rauscher, F.J., Schafer,
B.W., 1996. Induction of apoptosis in rhabdomyosarcoma cells through
down-regulation of PAX proteins. Proc. Natl. Acad. Sci. USA 93,
13164–13169.
Bopp, D., Burri, M., Baumgartner, S., Frigerio, G., Noll, M., 1986. Conser-
vation of a large protein domain in the segmentation gene paired and in
functionally related genes of Drosophila. Cell 47, 1033–1040.
Brink, C., Chowdhury, K., Gruss, P., 2001. Pax4 regulatory elements
mediate beta cell specific expression in the pancreas. Mech. Dev.
100, 37–43.
Chalepakis, G., Stoykova, A., Wijnholds, J., Tremblay, P., Gruss, P., 1993.
Pax: gene regulators in the developing nervous system. J. Neurobiol. 24,
1367–1384.
Davis, R.J., D’Cruz, C.M., Lovell, M.A., Biegel, J.A., Barr, F.G., 1994.
Fusion of PAX7 to FKHR by the variant t(1;13)(p36;q14) translocation
in alveolar rhabdomyosarcoma. Cancer Res. 54, 2869–2872.
Ghosh, G., Van Duyne, G.D., 1996. Pieces of the puzzle: assembling the
preinitiation complex of Pol II. Structure 4, 891–895.
Giger, R.J., Vogt, L., Zuellig, R.A., Rader, C., Henehen-Beatty, A., Wolfer,
D.P., Sonderegger, P., 1995. The gene of chicken axonin: complete
structure and analysis of the promoter. Eur. J. Biochem. 227, 617–628.
Gruss, P., Walther, C., 1992. Pax in development. Cell 69, 719–722.
Hardison, R., Slightom, J.L., Gumucio, D.L., Goodman, M., Stojanovic, N.,
Miller, W., 1997. Locus control regions of mammalian beta-globin gene
clusters: combining phylogenetic analyses and experimental results to
gain functional insights. Gene 205, 73–94.
Jaakkola, P., Vihinen, T., Maatta, A., Jalkanen, M., 1997. Activation of an
enhancer on the syndecan-1 gene is restricted to fibroblast growth factor
family members in mesenchymal cells. Mol. Cell. Biol. 17, 3210–3219.
Johnson, P.F., McKnight, S.L., 1989. Eukaryotic transcriptional regulatory
proteins. Annu. Rev. Biochem. 58, 799–839.
Jostes, B., Walther, C., Gruss, P., 1991. The murine paired box gene, Pax7,
is expressed specifically during the development of the nervous and
muscular system. Mech. Dev. 33, 27–37.
Kashi, Y., King, D., Soller, M., 1997. Simple sequence repeats as a source
of quantitative genetic variation. Trends Genet. 13, 74–78.
Kaufmann, J., Smale, S.T., 1994. Direct recognition of initiator elements by
a component of the transcription factor IID complex. Genes Dev. 8,
821–829.
Kay, P.H., Mitchell, C.A., Akkari, A., Papadimitriou, J.M., 1995. Associa-
tion of an unusual form of a Pax7-like gene with increased efficiency of
skeletal muscle regeneration. Gene 163, 171–177.
Kay, P.H., Harmon, D., Fletcher, S., Robertson, T., Ziman, M., Papadimi-
triou, J.M., 1998. Pax7 includes two polymorphic homeoboxes which
contain rearrangements associated with differences in the ability to
regenerate damaged skeletal muscle in adult mice. Int. J. Biochem.
Cell Biol. 30, 261–269.
Kozlov, S.V., Giger, R.J., Hasler, T., Korvatska, E., Schiorderet, D.F.,
Sonderegger, P., 1995. The human TAX1 gene encoding the axon-
associated cell adhesion molecule TAG-1/Axonin-1: genomic structure
and basic promoter. Genomics 30, 141–148.
Mansouri, A., Stoykova, A., Torres, M., Gruss, P., 1996. Dysgenesis of
cephalic neural crest derivatives in Pax7 2 /2 mutant mice. Develop-
ment 122, 831–838.
Maroto, M., Reshef, R., Munsterberg, A.E., Koester, S., Goulding, M.,
Lassar, A.B., 1997. Ectopic Pax-3 activates MyoD and Myf-5 expres-
sion in embryonic mesoderm and neural tissue. Cell 89, 139–148.
Maul, R.S., Zhang, H., Reid, J.D.T., Pedigo, N.G., Kaetzel, D.M., 1998.
Identification of a cell type-specific enhancer in the distal 5 0-region of
the platelet-derived growth factor A-chain gene. J. Biol. Chem. 273,
33239–33246.
Mitas, M., 1997. Trinucleotide repeats associated with human disease.
Nucleic Acids Res. 25, 2245–2254.
Murmann, O.V., Niggli, F., Schafer, B.W., 2000. Cloning and character-
ization of the human PAX7 promoter. Biol. Chem. 381, 331–335.
Okladnova, O., Syagailo, Y.V., Tranitz, M., Riederer, P., Stober, G., Moss-
ner, R., Lesch, K.P., 1999. Functional characterization of the human
PAX3 gene regulatory region. Genomics 57, 110–119.
Quandt, K., Frech, K., Karas, H., Wingender, E., Werner, T., 1995. MatInd
and MatInspector: new fast and versatile tools for detection of consen-
sus matches in nucleotide sequence data. Nucleic Acids Res. 23, 4878–
4884.
Schafer, B.W., Czerny, T., Bernasconi, M., Genini, M., Busslinger, M.,
1994. Molecular cloning and characterization of a human PAX-7
cDNA expressed in normal and neoplastic myocytes. Nucleic Acids
Res. 22, 4574–4582.
Seale, P., Sabourin, L.A., Girgis-Gabardo, A., Mansouri, A., Gruss, P.,
Rudnicki, M.A., 2000. Pax7 is required for the specification of
myogenic satellite cells. Cell 102, 777–786.
Sinden, R.R., Wells, R.D., 1992. DNA structure, mutations, and human
genetic disease. Curr. Opin. Biotechnol. 3, 612–622.
Stayner, C.K., Cunliffe, H.E., Ward, T.A., Eccles, M.R., 1998. Cloning and
characterization of the human PAX2 promoter. J. Biol. Chem. 273,
25472–25479.
Stoykova, A., Gruss, P., 1994. Roles of Pax-genes in developing and adult
brain as suggested by expression patterns. J. Neurosci. 14, 1395–1412.
Strachan, T., Read, A.P., 1994. PAX genes. Curr. Opin. Genet. Dev. 4, 427–
438.
Tajbakhsh, S., Rocancourt, D., Cossu, G., Buckingham, M., 1997. Redefin-
ing the genetic hierarchies controlling skeletal myogenesis: Pax-3 and
Myf-5 act upstream of MyoD. Cell 89, 127–138.
Tremblay, P., Gruss, P., 1994. Pax: genes for mice and men. Pharmacol.
Ther. 61, 205–226.
Tsukamoto, K., Nakamura, Y., Niikawa, N., 1994. Isolation of two
isoforms of the PAX3 gene transcripts and their tissue-specific alter-
native expression in human adult tissues. Hum. Genet. 93, 270–274.
van der Velden, A.W., Thomas, A.A., 1999. The role of the 5 0 untranslated
region of an mRNA in translation regulation during development. Int. J.
Biochem. Cell Biol. 31, 87–106.
Vorobyov, E., Mertsalov, I., Dockhorn-Dworniczak, B., Dworniczak, B.,
Horst, J., 1997. The genomic organization and the full coding region of
the human PAX7 gene. Genomics 45, 168–174.
Walther, C., Guenet, J.L., Simon, D., Deutsch, U., Jostes, B., Goulding,
M.D., Plachov, D., Balling, R., Gruss, P., 1991. Pax: a murine multi-
gene family of paired box-containing genes. Genomics 11, 424–434.
Ziman, M.R., Fletcher, S., Kay, P.H., 1997. Alternate Pax7 transcripts are
expressed specifically in skeletal muscle, brain and other organs of adult
mice. Int. J. Biochem. Cell Biol. 29, 1029–1036.
Y.V. Syagailo et al. / Gene 294 (2002) 259–268268
Copyright © 2022 FDOKUMEN




















