
Haptophytes of the Nervión River estuary, northern Spain
13
Botanica Marina 52 (2009): 47–59 2009 by Walter de Gruyter • Berlin • New York. DOI 10.1515/BOT.2009.027 Article in press - uncorrected proof 2009/63 Haptophytes of the Nervio ´ n River estuary, northern Spain Sergio Seoane 1, *, Wenche Eikrem 2 , Jon Arluzea 3 and Emma Orive 1 1 Laboratorio de Ecologı´a, Departamento de Biologı´a Vegetal y Ecologı´a,Facultad de Ciencia y Tecnologı ´a, Universidaddel Paı´s Vasco, Apdo. 644, 48080 Bilbao, Spain, e-mail: [email protected] 2 Norwegian Institute for Water Research, Gaustadalle ´ en 21, 0349 Oslo, Norway 3 Departamentode Biologı´a Celular e Histologı´a, Facultad de Medicina e Histologı ´a, Universidad del Paı ´s Vasco, Apdo. 644, 48080 Bilbao, Spain * Corresponding author Abstract The meso-macrotidal Nervio ´ n River estuary in northern Spain was sampled monthly from March to September 2002 and 2003 to study species composition and abun- dance of haptophytes in the phytoplankton. From natural samples and uni-algal cultures, 17 haptophyte species were identified using light and electron microscopy. The species were found across a wide range of temperatures and salinities. The most frequently occurring haptophytes in the estuary were species of the genus Chrysochro- mulina; however, Isochrysis galbana reached highest abundances. Chrysochromulina lanceolata and Phaeo- cystis globosa were occasionally present at high abun- dances. Another bloom-forming species, Prymnesium parvum, was recorded only once. Of the genera identi- fied, eight species belonged to Chrysochromulina, two to Prymnesium and the rest of the genera were represented by one species only. The majority of the species were recorded for the first time in the Cantabrian Sea and one of them for the first time in the Atlantic Ocean. Keywords: Chrysochromulina; distribution; Haptophyta; Nervio ´ n estuary. Introduction Haptophytes are one of the main components of marine phytoplankton as revealed by light and electron micro- scopy, and pigment and molecular analysis (Thomsen et al. 1994, Andersen et al. 1996, Carreto et al. 2003, DiTullio et al. 2003, Lohrenz et al. 2003, Rodrı ´guez et al. 2003, Latasa et al. 2005, Not et al. 2005). Some hapto- phytes, such as Emiliania huxleyi (Lohmann) Hay et Moh- ler occur regularly in the North Atlantic Ocean forming extensive blooms (Lampert et al. 2002), while Chryso- chromulina polylepis Manton et Parke has been respon- sible for sporadic bloom events in Scandinavian waters (Dahl et al. 1998). The preponderance of haptophytes in oligotrophic waters may be explained by their small cell size and ability to feed phagotrophically (Jones et al. 1993, Hansen and Hjorth 2002, Lessard et al. 2005), or, in the case of the genus Phaeocystis Lagerheim, by its ability to form gel-like colonies (Schoemann et al. 2005). Many haptophyte species have been reported from a wide latitudinal range and, therefore, may be regarded as cosmopolitan (Estep et al. 1984, Birkhead and Pienaar 1995a, Jensen 1998, Eikrem and Edvardsen 1999, Eik- rem and Throndsen 1999, Puigserver et al. 2003, LeRoi and Hallegraeff 2004, Hu et al. 2005, Nakayama et al. 2005). The filiform organelle, known as the haptonema, char- acterizes the algal phylum Haptophyta Hibberd ex Edvardsen et Eikrem and, when present, it is located between two flagella of equal or unequal length. Hapto- phyte cells belonging to the class Coccolithophyceae Rothmaler ex Silva et Throndsen are covered with organ- ic scales and/or coccoliths, whose shapes and patterns are useful for species identification (Silva et al. 2007). In fact, the examination of scales and coccoliths by electron microscopy is usually required to discriminate between species. The abundance of haptophytes is often under- estimated when observed by light microscopy, as is frequently the case when enumerating other small phy- toflagellates. Haptophytes can also be underemphasized when using pigment analysis to measure their contribu- tion to total phytoplankton biomass due to their complex pigment composition. Recent improvements in high performance liquid chro- matography pigment analyses have allowed identification of several different pigment patterns among hapto- phytes, with some species (e.g., Isochrysis galbana Parke, Pavlova gyrans Butcher) lacking 199hexanoyloxy- fucoxanthin (199hex) (Van Lenning et al. 2003, Zapata et al. 2004), which until recently was considered the main pigment signature for the phylum. Keeping in mind that there is no marker pigment for all haptophytes, pigment analysis may still be a valuable tool in estimating the relative abundance of haptophytes in marine phyto- plankton. Although most haptophytes are described from marine environments, several species are euryhaline (Rhodes and Burke 1996) and occur in coastal waters including estuaries (Dahl et al. 2005). In the Nervio ´ n River estuary in northern Spain, Seoane et al. (2005, 2006) reported the relative importance of haptophytes in terms of cell abundances, although the concentration of 199hex was generally low, which can be explained by the fact that Isochrysis galbana, which lacks 199hex, frequently dom- inated the haptophyte assemblage. The main objective of this paper is to report on the taxonomic composition of haptophytes recorded in the Nervio ´ n River estuary and to describe their morphological characteristics.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Haptophytes of the Nervión River estuary, northern Spain

Botanica Marina 52 (2009): 47–59 � 2009 by Walter de Gruyter • Berlin • New York. DOI 10.1515/BOT.2009.027
Article in press - uncorrected proof
2009/63
Haptophytes of the Nervion River estuary, northern Spain
Sergio Seoane1,*, Wenche Eikrem2, JonArluzea3 and Emma Orive1
1 Laboratorio de Ecologıa, Departamento de BiologıaVegetal y Ecologıa, Facultad de Ciencia y Tecnologıa,Universidad del Paıs Vasco, Apdo. 644, 48080 Bilbao,Spain, e-mail: [email protected] Norwegian Institute for Water Research, Gaustadalleen21, 0349 Oslo, Norway3 Departamento de Biologıa Celular e Histologıa,Facultad de Medicina e Histologıa, Universidad del PaısVasco, Apdo. 644, 48080 Bilbao, Spain
* Corresponding author
Abstract
The meso-macrotidal Nervion River estuary in northernSpain was sampled monthly from March to September2002 and 2003 to study species composition and abun-dance of haptophytes in the phytoplankton. From naturalsamples and uni-algal cultures, 17 haptophyte specieswere identified using light and electron microscopy. Thespecies were found across a wide range of temperaturesand salinities. The most frequently occurring haptophytesin the estuary were species of the genus Chrysochro-mulina; however, Isochrysis galbana reached highestabundances. Chrysochromulina lanceolata and Phaeo-cystis globosa were occasionally present at high abun-dances. Another bloom-forming species, Prymnesiumparvum, was recorded only once. Of the genera identi-fied, eight species belonged to Chrysochromulina, two toPrymnesium and the rest of the genera were representedby one species only. The majority of the species wererecorded for the first time in the Cantabrian Sea and oneof them for the first time in the Atlantic Ocean.
Keywords: Chrysochromulina; distribution; Haptophyta;Nervion estuary.
Introduction
Haptophytes are one of the main components of marinephytoplankton as revealed by light and electron micro-scopy, and pigment and molecular analysis (Thomsen etal. 1994, Andersen et al. 1996, Carreto et al. 2003,DiTullio et al. 2003, Lohrenz et al. 2003, Rodrıguez et al.2003, Latasa et al. 2005, Not et al. 2005). Some hapto-phytes, such as Emiliania huxleyi (Lohmann) Hay et Moh-ler occur regularly in the North Atlantic Ocean formingextensive blooms (Lampert et al. 2002), while Chryso-chromulina polylepis Manton et Parke has been respon-sible for sporadic bloom events in Scandinavian waters
(Dahl et al. 1998). The preponderance of haptophytes inoligotrophic waters may be explained by their small cellsize and ability to feed phagotrophically (Jones et al.1993, Hansen and Hjorth 2002, Lessard et al. 2005), or,in the case of the genus Phaeocystis Lagerheim, by itsability to form gel-like colonies (Schoemann et al. 2005).Many haptophyte species have been reported from awide latitudinal range and, therefore, may be regarded ascosmopolitan (Estep et al. 1984, Birkhead and Pienaar1995a, Jensen 1998, Eikrem and Edvardsen 1999, Eik-rem and Throndsen 1999, Puigserver et al. 2003, LeRoiand Hallegraeff 2004, Hu et al. 2005, Nakayama et al.2005).
The filiform organelle, known as the haptonema, char-acterizes the algal phylum Haptophyta Hibberd exEdvardsen et Eikrem and, when present, it is locatedbetween two flagella of equal or unequal length. Hapto-phyte cells belonging to the class CoccolithophyceaeRothmaler ex Silva et Throndsen are covered with organ-ic scales and/or coccoliths, whose shapes and patternsare useful for species identification (Silva et al. 2007). Infact, the examination of scales and coccoliths by electronmicroscopy is usually required to discriminate betweenspecies. The abundance of haptophytes is often under-estimated when observed by light microscopy, as isfrequently the case when enumerating other small phy-toflagellates. Haptophytes can also be underemphasizedwhen using pigment analysis to measure their contribu-tion to total phytoplankton biomass due to their complexpigment composition.
Recent improvements in high performance liquid chro-matography pigment analyses have allowed identificationof several different pigment patterns among hapto-phytes, with some species (e.g., Isochrysis galbanaParke, Pavlova gyrans Butcher) lacking 199hexanoyloxy-fucoxanthin (199hex) (Van Lenning et al. 2003, Zapata etal. 2004), which until recently was considered the mainpigment signature for the phylum. Keeping in mind thatthere is no marker pigment for all haptophytes, pigmentanalysis may still be a valuable tool in estimating therelative abundance of haptophytes in marine phyto-plankton.
Although most haptophytes are described from marineenvironments, several species are euryhaline (Rhodesand Burke 1996) and occur in coastal waters includingestuaries (Dahl et al. 2005). In the Nervion River estuaryin northern Spain, Seoane et al. (2005, 2006) reportedthe relative importance of haptophytes in terms of cellabundances, although the concentration of 199hex wasgenerally low, which can be explained by the fact thatIsochrysis galbana, which lacks 199hex, frequently dom-inated the haptophyte assemblage. The main objectiveof this paper is to report on the taxonomic compositionof haptophytes recorded in the Nervion River estuary andto describe their morphological characteristics.

48 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
Figure 1 Location of the sampling stations in the Nervion River estuary.
Materials and methods
The Nervion River estuary is a small estuary draining intothe Cantabrian Sea in the Bay of Biscay. Tides in the Bayof Biscay are semidiurnal, with a maximum range of4.5 m during spring tides. With a moderate freshwaterflow rate (36 m3 s-1) (Borja et al. 2006), the Nervion Riverestuary has been subjected to large-scale industrial anddomestic sewage discharges from approximately onemillion people for decades. The estuary can be dividedinto two regions: a shallow (-7 m depth) and narrow 15-km-long channel, which is highly stratified, and a 30-m-deep outer area (Abra of Bilbao) with a maximum widthof 3.8 km and which is strongly influenced by coastalwaters (Seoane et al. 2005, 2006) (Figure 1).
At the Nervion River estuary, there are 9 monitoringstations (labelled 0–8). We focussed on the exterior partof the estuary (stations 0, 1, 2 and 3) and selected onerepresentative station of the middle part (station 5) andof the inner part (station 8) of the estuary. Water samplingwas conducted monthly from March to September at sta-tions 1, 2, and 3 in 2002 and at stations 0, 1, 2, 3, 5, and8 in 2003. One additional sampling was carried out inAugust 2004 at stations 0, 1, 2, 3, 5, and 8. Surface watersamples (0.5 m) were always collected in the morningduring slack water of high tide. Sampling was carried outfrom a small fishing boat and water samples were col-lected with 1-l Nalgene� bottles (Nalge Nunc Internation-al Corporation, Rochester, NY, USA). A 250-ml sub-sample was fixed in 1% final concentration glutaralde-
hyde, while the remaining 750-ml subsample wasbrought to the laboratory alive. Surface water salinity andtemperature were measured at each station with a WTWLF 197 (WTW Wissenschaftlich-Technische WerkstattenGmbH, Weilheim, Germany) microprocessor conductivitymeter and a YSI 58(5739) (Fondriest environmental mon-itoring products, Alpha, OH, USA) oxygen meter,respectively.
Three different medium solutions were prepared withseawater from the Nervion River estuary to maintain thehaptophyte cultures: modified Erd-Schreiber (Throndsen1978), f/2 (Guillard and Ryther 1962), and modified f/2medium (Table 1). Germanium dioxide was added to thehaptophyte cultures to prevent diatom growth. Serialdilution cultures (SDCs) were established (Throndsen1978) and single haptophyte cells were isolated from theculture flasks by micropipetting under a Leitz fluovert FSmicroscope (Leica, Wetzlar, Germany) fitted with a micro-manipulator. The haptophyte cultures and SDCs weremaintained at a light intensity of 60 mmol photons m-2 s-1
at 18–228C with a photoperiod of 14:10 h light/darkcycle.
Field samples and culture material were examined inlight microscopy (LM) with an inverted and traditionallight microscope (Nikon TMD, Tokyo, Japan and LeicaDMRB (Leica, Wetzlar, Germany, respectively), equippedwith a digital camera system (Leica Q500 image analysissoftware (Leica, Wetzlar, Germany). Culture material wasfixed in 1% glutaraldehyde, mounted on polylysine-coat-ed coverslips and stored overnight in a moist chamber

S. Seoane et al.: Haptophytes of the Nervion River estuary 49
Article in press - uncorrected proof
Table 1 Composition of modified f/2 medium.
Stock solutions
NaNO3 7.5 g dissolved in 100 ml Milli-Qwater
C3H7Na2O6PØ5H2O 1.11 g in 100 ml Milli-Q waterC6H8O7ØH2O 0.096 g in 100 ml Milli-Q waterNa2SeO3 0.173 mg in 100 ml Milli-Q water
Micronutrients Add to 1000 ml of Milli-Q water:Na2EDTA 4.360 gFeCl3Ø6H2O 3.150 gCuSO4Ø5H2O 0.010 gZnSO4Ø7H2O 0.022 gCoCl2Ø6H2O 0.010 gMnCl2Ø4H2O 0.180 gNa2MoO4Ø2H2O 0.006 g
Vitamins Add to 1000 ml of Milli-Q water:Thiamine 10 mgBiotin 50 mgCyanocobalamine 50 mg
To prepare medium, add 1 ml NaNO3, 1 ml C3H7Na2O6PØ5H2O,1 ml C6H8O7ØH2O, 1 ml Na2SeO3, 1 ml micronutrients and 1 mlvitamins to 1000 ml of sea water.
at room temperature. Coverslips were then rinsed in fil-tered culture medium for 20 min, post-fixed in 1% osmi-um tetroxide for 5 min and rinsed in distilled water.Finally, they were dehydrated in an ethanol series fol-lowed by two 5-min changes in hexamethyldisilazane.Coverslips were mounted on aluminum stubs, coatedwith Au and examined by scanning electron microscopy(SEM) (S-4800 Hitachi, Hitachi, Tokyo, Japan) at the Elec-tron Microscopy Facility of the University of the BasqueCountry. Additional culture material was placed on form-var and carbon-coated copper grids, fixed in 1% osmium
tetroxide vapor and left to dry. After rinsing in distilledwater, the grids were either contrasted on drops of sat-urated aqueous uranyl acetate and subsequently rinsedin distilled water or shadow-cast with Au/Pd (SpeedivacEdwards Coating Unit 12E6/1629, Edwards Vacuum,Crawley, UK) at an angle of 308. Preparations were exam-ined by transmission electron microscopy (TEM) (EM 208S, CM 120 Philips STEM and CM 100 Philips, Philips,Eindhoven, The Netherlands).
Cell size, haptonema and flagellum length, and scaledimensions were measured in 10 to 20 specimens ofeach species. Cell shapes, flagella, haptonema and largescales as well as swimming behavior were examined inLM. Cell size and the lengths of flagella and haptonemawere measured in LM, except for the very short append-ages that were measured in TEM. The shape and orna-mentation of the scales covering the cell were examinedin both TEM and SEM.
Results
We recorded a total of more than 20 haptophytes (Table2), of which 17 taxa were maintained in culture and iden-tified at the species level or lower from phytoplanktonsurveys conducted in the inner and outer regions of theNervion River estuary in northern Spain (Table 3, Figures2–59). Surface water temperature and salinity rangedfrom 15–258C to 18–35 psu, respectively, during oursampling (Table 3). Several specimens of Chrysochro-mulina Lackey and some coccolithophorids, such asCalciopappus Gaarder et Ramsfjell and RhabdosphaeraHaeckel, were only observed in natural samples by LMand were never maintained in culture. In 2002 and 2003,

50 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
Table 3 Surface water salinity range, temperature range and17 haptophyte taxa recorded at the five sampling stations onthe Nervion River estuary in 2002, 2003 and August 2004.
Taxon Temperature(8C)
Salinity(psu)
Chrysochromulina acantha 13–20 24–33C. brevifilum 12–15 18–24C. cf. cymbium 21 31C. hirta 17–21 27–32C. lanceolata 24–26 25–35C. pringsheimii 19–24 28–35C. simplex 10–26 17–34C. throndsenii 13–21 24–34Emiliania huxleyi 12–24 27–35Imantonia rotunda 12–25 21–35Isochrysis galbana 13–24 23–31Pavlova gyrans 19–25 35Phaeocystis globosa 13–21 24–33Platychrysis pienaarii 21 20Pleurochrysis roscoffensis 17–19 19–33Prymnesium parvum f. patelliferum 21 35Prymnesium sp. 17–24 32–34
haptophytes were found at all stations along the estuary,except at the innermost station 8. The number of taxawas highest during the summer months and Emilianiahuxleyi and Imantonia rotunda Reynolds were the mostfrequently recorded species in the serial dilution cultures.In August 2004, Prymnesium parvum f. patelliferum(Green, Hibberd et Pienaar) Larsen was recorded atstation 0.
Class Coccolithophyceae Rothmaler ex Silva etThrondsenOrder Prymnesiales Papenfuss, emend. Edvardsenet EikremChrysochromulina acantha Leadbeater et Manton(Figures 2, 24–27)
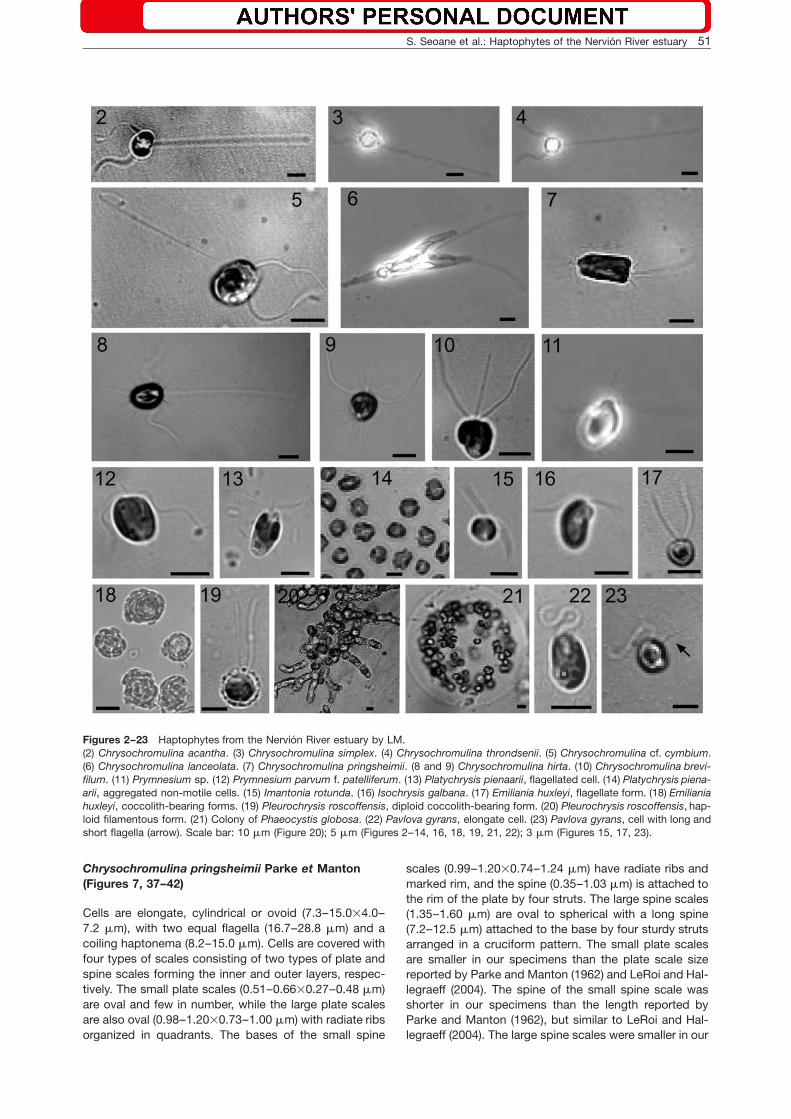
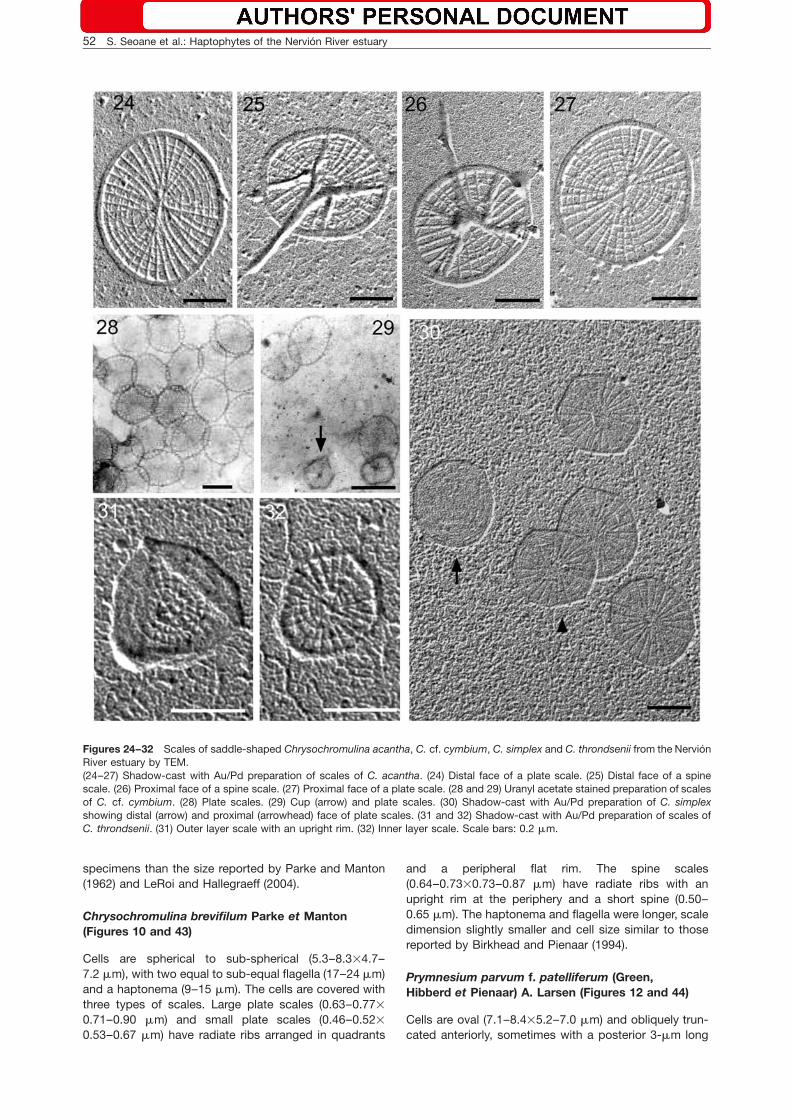
Cells are saddle-shaped (4.1–7.4=5.0–8.8 mm), with twoequal flagella (15–21 mm) and a long coiling haptonema(33–53 mm). Cell covering consists of two types ofscales: basal plate scales (0.45–0.53=0.62–0.70 mm)and spine scales (0.46–0.54=0.43–0.50 mm). The spine(0.42–0.53 mm) is almost equal in length to the scalediameter and is attached to the basal plate by four strutsreaching the scale rim. Both types of scales have radiateand concentric rings. Our cell measurements are slightlyshorter than those reported for the type material by Lead-beater and Manton (1971), whereas the scale dimensionswere similar.
Chrysochromulina cf. cymbium Leadbeater etManton (Figures 5, 28, 29)
Cells are saddle-shaped (5.6–8.6=5.0–8.6 mm), with twoequal flagella (19.5–26.0 mm) and a coiling haptonemaof variable size (42–110 mm). The cells swam very rap-idly. Cell covering is composed of two types of scales:cup scales (0.13–0.17 mm) and plate scales (0.23–0.29=0.16–0.21 mm), which have radiate ribs and a sin-
gle row of marginal perforations. Our cell and plate scalemeasurements agreed with those reported for the typespecies (Leadbeater and Manton 1969), while the size ofcup scales in our specimens were larger.
Chrysochromulina simplex Estep, Davis, Hargraveset Sieburth emend. Birkhead et Pienaar (Figures 3and 30)
Cells are saddle-shaped (4.3–7.0=4.0–6.5 mm), with twoequal flagella (16.6–19.0 mm) and a coiling haptonema3–4 times longer (45–66 mm) than the flagella. The cellsare covered with plate scales only (0.33–0.46=0.40–0.49 mm), whose dimensions are similar to those report-ed by Estep et al. (1984) and Birkhead and Pienaar(1995b).
Chrysochromulina throndsenii Eikrem(Figures 4, 31, 32)
Cells are saddle-shaped (4.4–6.3=3.7–4.6 mm), with twoequal flagella (11–15 mm) and a coiling haptonema(29.0–42.5 mm). Two types of oval to spherical platescales cover the cell in two layers. Scales of the outerlayer have a raised rim and are larger (0.29–0.33=0.32–0.37 mm) than the scales of the inner layer(0.28–0.31=0.25–0.31 mm). The center of the scales hasa cruciform structure surrounded by an open ring withradiate ribs extending out of the central ring to the rimand crossed by spiraling ribs. Our specimens are smallerthan those reported for the type species, but our scaledimensions were similar (Eikrem 1996, Puigserver et al.2003).
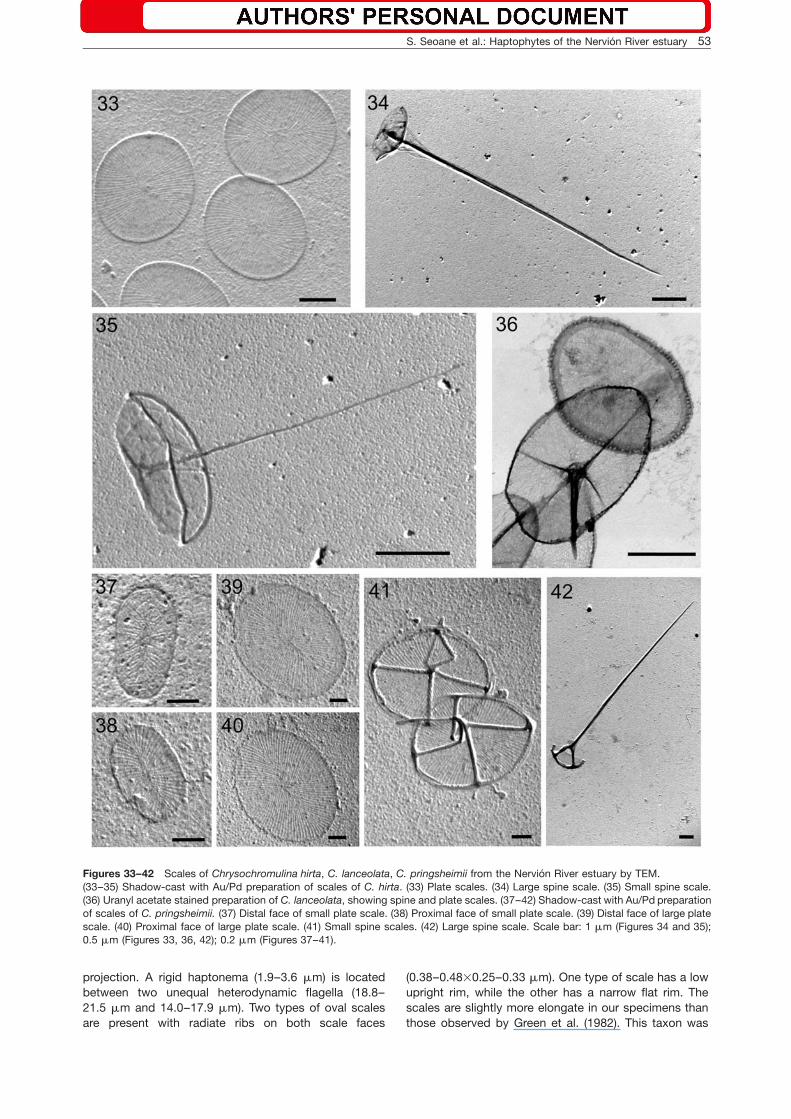
Chrysochromulina hirta Manton (Figures 8, 9, 33–35)
Cells are spherical to sub-spherical (5.0–8.2 mm), withtwo equal flagella (16.0–22.8 mm) and a coiling hapto-nema (31–45 mm). Three types of sub-spherical scaleswere observed, viz. plate scales (1.09–1.43=1.00–1.19 mm) and two types of spine scales. The small spinescales (1.3–2.0 mm) have short spines (3.5–4.8 mm) andthe large spine scales (1.7–2.4 mm) have long spines(12.4–14.7 mm) attached to a concave base. Dimensionsof cells, flagella, and haptonema are within the size rangereported by Manton (1978) and Eikrem (1999), but thescale size was smaller than those reported by Manton(1978) and LeRoi and Hallegraeff (2004).
Chrysochromulina lanceolata Chretiennot-Dinet,Nezan et Puigserver (Figures 6 and 36)
Cells are lancet-shaped w20–36(61)=7.3–13.0 mmx, withtwo almost equal flagella (48–51 mm) and a haptonema(28–33 mm). Two types of scales are present, viz. platescales (0.91–1.06=0.50–0.83 mm) and spine scales(0.93–1.19=0.65–0.77 mm), with a spine of 0.58–0.75 mm in length. The spine is attached to an oval baseby four struts extending out to the rim. Both types ofscales have radiate ribs arranged in quadrants on theproximal scale face. Dimensions of cells, haptonema,and flagella coincided with those of Puigserver et al.(2003), but the scales were slightly smaller.
S. Seoane et al.: Haptophytes of the Nervion River estuary 51
Article in press - uncorrected proof
Figures 2–23 Haptophytes from the Nervion River estuary by LM.(2) Chrysochromulina acantha. (3) Chrysochromulina simplex. (4) Chrysochromulina throndsenii. (5) Chrysochromulina cf. cymbium.(6) Chrysochromulina lanceolata. (7) Chrysochromulina pringsheimii. (8 and 9) Chrysochromulina hirta. (10) Chrysochromulina brevi-filum. (11) Prymnesium sp. (12) Prymnesium parvum f. patelliferum. (13) Platychrysis pienaarii, flagellated cell. (14) Platychrysis piena-arii, aggregated non-motile cells. (15) Imantonia rotunda. (16) Isochrysis galbana. (17) Emiliania huxleyi, flagellate form. (18) Emilianiahuxleyi, coccolith-bearing forms. (19) Pleurochrysis roscoffensis, diploid coccolith-bearing form. (20) Pleurochrysis roscoffensis, hap-loid filamentous form. (21) Colony of Phaeocystis globosa. (22) Pavlova gyrans, elongate cell. (23) Pavlova gyrans, cell with long andshort flagella (arrow). Scale bar: 10 mm (Figure 20); 5 mm (Figures 2–14, 16, 18, 19, 21, 22); 3 mm (Figures 15, 17, 23).
Chrysochromulina pringsheimii Parke et Manton(Figures 7, 37–42)
Cells are elongate, cylindrical or ovoid (7.3–15.0=4.0–7.2 mm), with two equal flagella (16.7–28.8 mm) and acoiling haptonema (8.2–15.0 mm). Cells are covered withfour types of scales consisting of two types of plate andspine scales forming the inner and outer layers, respec-tively. The small plate scales (0.51–0.66=0.27–0.48 mm)are oval and few in number, while the large plate scalesare also oval (0.98–1.20=0.73–1.00 mm) with radiate ribsorganized in quadrants. The bases of the small spine
scales (0.99–1.20=0.74–1.24 mm) have radiate ribs andmarked rim, and the spine (0.35–1.03 mm) is attached tothe rim of the plate by four struts. The large spine scales(1.35–1.60 mm) are oval to spherical with a long spine(7.2–12.5 mm) attached to the base by four sturdy strutsarranged in a cruciform pattern. The small plate scalesare smaller in our specimens than the plate scale sizereported by Parke and Manton (1962) and LeRoi and Hal-legraeff (2004). The spine of the small spine scale wasshorter in our specimens than the length reported byParke and Manton (1962), but similar to LeRoi and Hal-legraeff (2004). The large spine scales were smaller in our
52 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
Figures 24–32 Scales of saddle-shaped Chrysochromulina acantha, C. cf. cymbium, C. simplex and C. throndsenii from the NervionRiver estuary by TEM.(24–27) Shadow-cast with Au/Pd preparation of scales of C. acantha. (24) Distal face of a plate scale. (25) Distal face of a spinescale. (26) Proximal face of a spine scale. (27) Proximal face of a plate scale. (28 and 29) Uranyl acetate stained preparation of scalesof C. cf. cymbium. (28) Plate scales. (29) Cup (arrow) and plate scales. (30) Shadow-cast with Au/Pd preparation of C. simplexshowing distal (arrow) and proximal (arrowhead) face of plate scales. (31 and 32) Shadow-cast with Au/Pd preparation of scales ofC. throndsenii. (31) Outer layer scale with an upright rim. (32) Inner layer scale. Scale bars: 0.2 mm.
specimens than the size reported by Parke and Manton(1962) and LeRoi and Hallegraeff (2004).
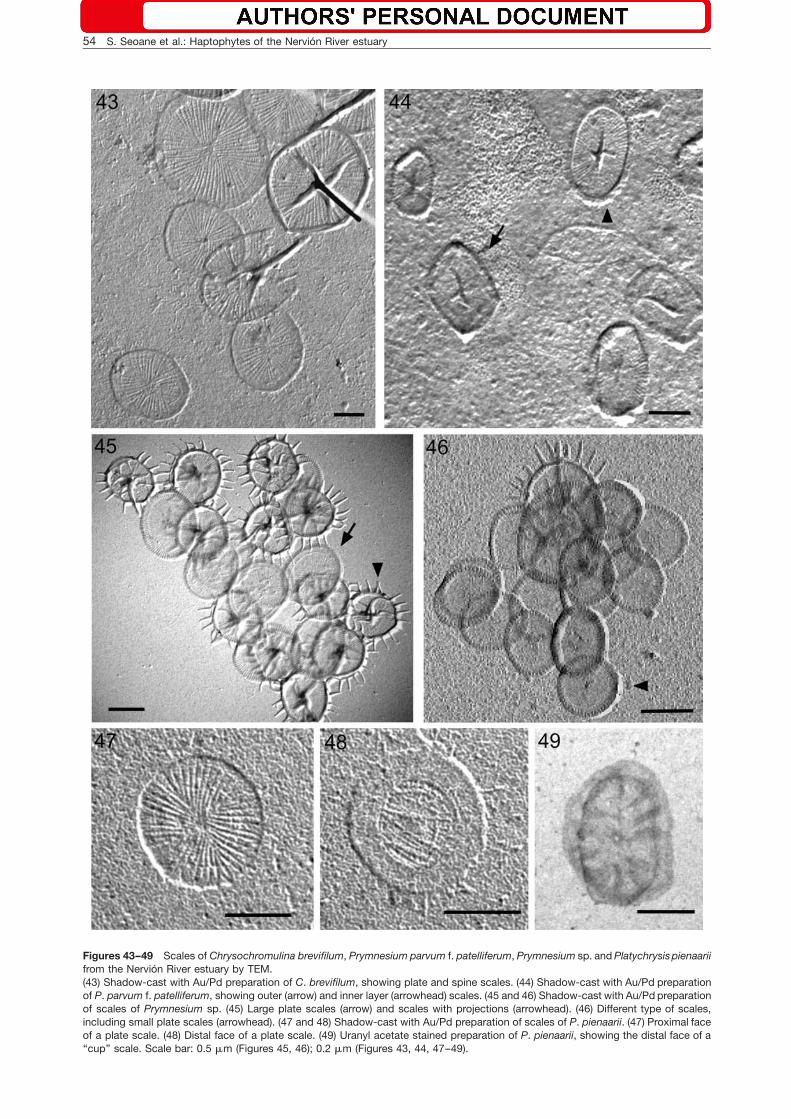
Chrysochromulina brevifilum Parke et Manton(Figures 10 and 43)
Cells are spherical to sub-spherical (5.3–8.3=4.7–7.2 mm), with two equal to sub-equal flagella (17–24 mm)and a haptonema (9–15 mm). The cells are covered withthree types of scales. Large plate scales (0.63–0.77=0.71–0.90 mm) and small plate scales (0.46–0.52=0.53–0.67 mm) have radiate ribs arranged in quadrants
and a peripheral flat rim. The spine scales(0.64–0.73=0.73–0.87 mm) have radiate ribs with anupright rim at the periphery and a short spine (0.50–0.65 mm). The haptonema and flagella were longer, scaledimension slightly smaller and cell size similar to thosereported by Birkhead and Pienaar (1994).
Prymnesium parvum f. patelliferum (Green,Hibberd et Pienaar) A. Larsen (Figures 12 and 44)
Cells are oval (7.1–8.4=5.2–7.0 mm) and obliquely trun-cated anteriorly, sometimes with a posterior 3-mm long
S. Seoane et al.: Haptophytes of the Nervion River estuary 53
Article in press - uncorrected proof
Figures 33–42 Scales of Chrysochromulina hirta, C. lanceolata, C. pringsheimii from the Nervion River estuary by TEM.(33–35) Shadow-cast with Au/Pd preparation of scales of C. hirta. (33) Plate scales. (34) Large spine scale. (35) Small spine scale.(36) Uranyl acetate stained preparation of C. lanceolata, showing spine and plate scales. (37–42) Shadow-cast with Au/Pd preparationof scales of C. pringsheimii. (37) Distal face of small plate scale. (38) Proximal face of small plate scale. (39) Distal face of large platescale. (40) Proximal face of large plate scale. (41) Small spine scales. (42) Large spine scale. Scale bar: 1 mm (Figures 34 and 35);0.5 mm (Figures 33, 36, 42); 0.2 mm (Figures 37–41).
projection. A rigid haptonema (1.9–3.6 mm) is locatedbetween two unequal heterodynamic flagella (18.8–21.5 mm and 14.0–17.9 mm). Two types of oval scalesare present with radiate ribs on both scale faces
(0.38–0.48=0.25–0.33 mm). One type of scale has a lowupright rim, while the other has a narrow flat rim. Thescales are slightly more elongate in our specimens thanthose observed by Green et al. (1982). This taxon was
54 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
Figures 43–49 Scales of Chrysochromulina brevifilum, Prymnesium parvum f. patelliferum, Prymnesium sp. and Platychrysis pienaariifrom the Nervion River estuary by TEM.(43) Shadow-cast with Au/Pd preparation of C. brevifilum, showing plate and spine scales. (44) Shadow-cast with Au/Pd preparationof P. parvum f. patelliferum, showing outer (arrow) and inner layer (arrowhead) scales. (45 and 46) Shadow-cast with Au/Pd preparationof scales of Prymnesium sp. (45) Large plate scales (arrow) and scales with projections (arrowhead). (46) Different type of scales,including small plate scales (arrowhead). (47 and 48) Shadow-cast with Au/Pd preparation of scales of P. pienaarii. (47) Proximal faceof a plate scale. (48) Distal face of a plate scale. (49) Uranyl acetate stained preparation of P. pienaarii, showing the distal face of a‘‘cup’’ scale. Scale bar: 0.5 mm (Figures 45, 46); 0.2 mm (Figures 43, 44, 47–49).
S. Seoane et al.: Haptophytes of the Nervion River estuary 55
Article in press - uncorrected proof
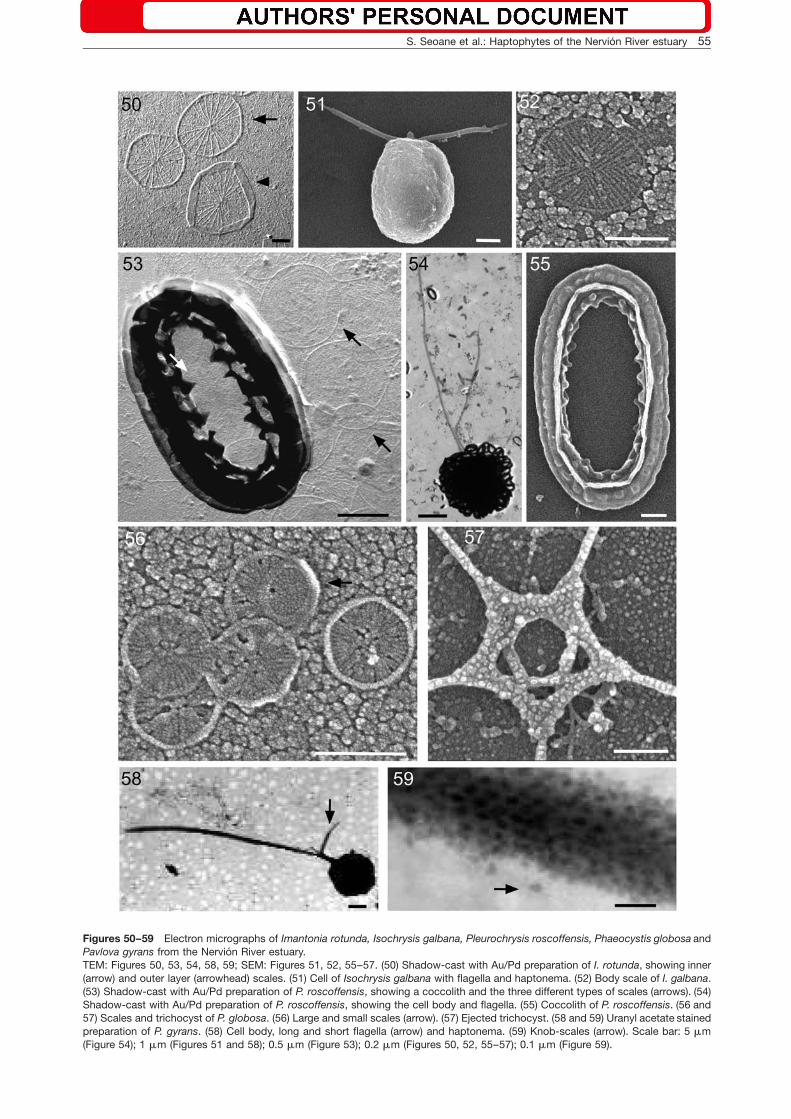
Figures 50–59 Electron micrographs of Imantonia rotunda, Isochrysis galbana, Pleurochrysis roscoffensis, Phaeocystis globosa andPavlova gyrans from the Nervion River estuary.TEM: Figures 50, 53, 54, 58, 59; SEM: Figures 51, 52, 55–57. (50) Shadow-cast with Au/Pd preparation of I. rotunda, showing inner(arrow) and outer layer (arrowhead) scales. (51) Cell of Isochrysis galbana with flagella and haptonema. (52) Body scale of I. galbana.(53) Shadow-cast with Au/Pd preparation of P. roscoffensis, showing a coccolith and the three different types of scales (arrows). (54)Shadow-cast with Au/Pd preparation of P. roscoffensis, showing the cell body and flagella. (55) Coccolith of P. roscoffensis. (56 and57) Scales and trichocyst of P. globosa. (56) Large and small scales (arrow). (57) Ejected trichocyst. (58 and 59) Uranyl acetate stainedpreparation of P. gyrans. (58) Cell body, long and short flagella (arrow) and haptonema. (59) Knob-scales (arrow). Scale bar: 5 mm(Figure 54); 1 mm (Figures 51 and 58); 0.5 mm (Figure 53); 0.2 mm (Figures 50, 52, 55–57); 0.1 mm (Figure 59).

56 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
only observed in culture from the additional samplingcarried out in August 2004. Of the two forms known forPrymnesium parvum Carter, only f. patelliferum wasobserved in the marine phytoplankton of the NervionRiver estuary.
Prymnesium sp. (Figures 11, 45, 46)
Cells are oval (8.0–12.6=6.2–9.0 mm), with two equal fla-gella (20–27 mm) and a haptonema (7.0–14.7 mm). Threetypes of scales cover the cell. The spine scales arealmost circular (0.58–0.82=0.54–0.79 mm), with abranched central projection and a peripheral upright rimwith approximately 20 projections (0.14–0.23 mm). Bothlarge (0.78–0.93=0.58–0.76 mm) and small (0.44–0.69=0.36–0.66 mm) plate scales have proximal and distalfaces with radiate ribs and one of the faces has a periph-eral flat rim. The scale dimensions in our specimens aresimilar to those reported from other geographic areas,except for the large scales which are smaller than thosereported from South Africa and Tasmania (Pienaar andBandu 1984, LeRoi and Hallegraeff 2004). Molecularstudies have shown that this taxon is closely related tothe genus Prymnesium Massart ex Conrad (S. Seoane,personal communication). This taxon has been previous-ly recorded as the ‘‘eyelash Chrysochromulina’’ (Moe-strup 1979, Pienaar and Bandu 1984, Birkhead andPienaar 1995a).
Platychrysis pienaarii Gayral et Fresnel(Figures 13, 14, 47–49)
The life cycle of this taxon has two phases with aggre-gated non-motile cells (8.5–12.0=8.1–9.8 mm) and flag-ellated cells (8–12=3.5–4.7 mm), which have two equalflagella (8.2–12.0 mm) and a rigid haptonema(1.9–3.6 mm). The flagellated cells are covered by twotypes of scales. The plate scales (0.28–0.35=0.21–0.25 mm) are oval to spherical with a radial patternon one face, whereas the other face has a zone wherethe ribs are closely packed. The cup-like scales(0.27–0.36=0.21–0.26 mm) have a radial pattern on theproximal face, whereas the distal face has 10 broad ribsfrom the top of the cup to the central node. Dimensionsof cells, haptonema, and flagella were slightly larger thanthose given in the description of the type species. Scaledimensions corresponded to those reported by Gayraland Fresnel (1983). The proximal scale layer is composedof plate scales and the distal layer of cup-like scales(Gayral and Fresnel 1983).
Imantonia rotunda Reynolds (Figures 15 and 50)
Cells are spherical to sub-spherical (2–4 mm), with twoequal flagella (3.5–7.0 mm), covered with scales(0.50–0.67 mm) having 20–24 ribs converging at the cen-ter; some scales possess an upright rim (0.10–0.13 mm).The cell size measured in our specimens is similar to thatfound by Reynolds (1974) and Green and Pienaar (1977),except that the flagella are shorter.
Order Isochrysidales Pascher emend. Edvardsen etEikremIsochrysis galbana Parke (Figures 16, 51, 52)
Cells are oval (6.0–7.2=2.6–3.4 mm) and spherical(4.7–6.5=4.3–5.6 mm) in young and old cultures, respec-tively. In young cultures, the cells have a rounded pos-terior and a flattened anterior end from which the flagella(4–7 mm) and haptonema (0.5 mm) emerge. In old cul-tures, the flagella tend to be lost. The cells are coveredwith only one type of oval scale (0.42=0.31–0.36 mm)with radiate ribs organized in quadrants. The cell sizemeasured in our specimens is similar to the one reportedby Parke (1949) and Green and Pienaar (1977). We wereunable to observe the scales covering the haptonema,but the body scales are similar in size to those reportedby Green and Pienaar (1977).
Emiliania huxleyi (Lohmann) Hay et Mohler(Figures 17 and 18)
The life cycle of this species has three morphotypes, viz.non-motile coccolith-bearing cells, completely nakednon-motile cells and flagellated cells with organic scales(Green et al. 1996, Paasche 2002). In the present study,we observed only non-motile coccolith-bearing cells andflagellated cells, although only the latter were maintainedin culture over an extended period of time. Non-motilecoccolith-bearing cells are spherical (6.0–10.3 mm) andcovered with several layers of coccoliths. Flagellatedcells (3.2–4.4 mm) have two equal flagella (4.8–11.0 mm),but we were unsuccessful in observing the cell scales.
Order Coccolithales Schwarz emend. Edvardsenet EikremPleurochrysis roscoffensis Pringsheim (Figures 19,20, 53–55)
This taxon has a life cycle with diploid coccolith-bearingflagellated cells and a haploid phase that forms filaments(Young et al. 2003). Flagellated cells are spherical to ovalw12–15(20)=10–14 mmx, with two unequal flagella(18–31 mm and 14–22 mm). Three types of scales andone type of coccolith cover the cell. The oval large platescales (1.54–1.81=0.70–1.06 mm) formed the base ofthe coccolith. Circular large plate scales (0.76–0.80 mm)and oval small plate scales (0.23=0.43 mm) are alsopresent. The oval heterococcoliths (2.16–2.66=1.34–1.77 mm) constitute the outermost layer. Non-motile cells(12.0–17.8=11.0–15.1 mm) form filaments. The cell sizesof the flagellated cells and filaments measured in ourspecimens are similar to those reported by others,except for the flagella, which were much shorter (Dan-geard 1934, Chadefaud and Feldmann 1949, Magne1954, Gayral and Fresnel 1976). The dimensions ofscales and coccoliths are within the size range reportedby Gayral and Fresnel (1976).
Order Phaeocystales MedlinPhaeocystis globosa Scherffel (Figures 21, 56, 57)
This taxon has a complex life cycle with four phases, viz.three flagellated forms (macro-, meso- and microflagel-

S. Seoane et al.: Haptophytes of the Nervion River estuary 57
Article in press - uncorrected proof
lates) and one colony-forming phase (Peperzak et al.2000). In our study, we observed the meso- and micro-flagellate forms as well as colonies. The meso- andmicroflagellate forms (2.7–3.6 mm) have two flagella(5.4–6.5 mm) and a short haptonema (0.90–1.57 mm) andappeared together in cultures. The presence of ejecto-somes in our culture material indicates that some of theflagellated cells belong to the mesoflagellate group. Thecells (5.2–6.5=3.9–5.5 mm) of the globose colonies areembedded in mucilage. The flagellated cells have platescales of two size categories (0.18–0.21 mm and0.12–0.15 mm), both with radiate ribs and a narrow rimalong the scale edge. The scale size in our specimens iswithin the range given by Peperzak et al. (2000), but themeso- and microflagellates are smaller for the respectivesize-classes, and the haptonema is slightly longer. Chre-tiennot-Dinet et al. (1997) and Peperzak et al. (2000)reported that mesoflagellate cells produce ejectosomesin the form of a pentameric star.
Class Pavlovophyceae (Cavalier-Smith) Green etMedlinOrder Pavlovales GreenPavlova gyrans Butcher emend. Green et Manton(Figures 22, 23, 58, 59)
Cells are oval (5.0–7.7=2.5–4.0 mm), with two unequalflagella (11–13 mm and 2.1–3.0 mm) and a short hapto-nema (1.25–2.70 mm). Knob-scales (0.025–0.030 mm)are bi-lobed with a constriction in the middle and coverthe long flagellum. The morphology and dimensions ofthe cells, haptonema, and flagella were similar to thosereported previously (Butcher 1952, Green and Manton1970, Green 1980). The presence of knob-scales on thelong flagellum is a characteristic feature of the generaPavlova Butcher and Rebecca Green.
Discussion
A total of 17 taxa of haptophytes were identified in thesurface waters of the Nervion River estuary, with salinityranging from 17.4 to 34.9. Haptophytes were observedat all stations, except at the innermost station (8), whichhad salinity in the oligo-mesohaline range (0.7–6.2). Mostof the taxa recorded were observed in samples collectedover a wide salinity and temperature range, indicating theeuryhaline and eurythermal nature of these haptophytes.Our observations corroborate the worldwide distributionof this group of algae reported from brackish and marinewaters by Thomsen et al. (1994), but also from Scandi-navia (Hadju et al. 1996, Jensen 1998), New Zealand(Moestrup 1979) and Tasmania (LeRoi and Hallegraeff2004).
The most frequently occurring haptophytes in the Ner-vion River estuary were species of Chrysochromulina,whereas the remaining genera were represented by onlyone or two taxa, as in the case of Prymnesium. Isochrysisgalbana was mainly observed in the bloom period duringthe summer; high abundances of Chrysochromulina lan-ceolata and Phaeocystis globosa were also recorded in
the estuary (Orive 1989, Seoane et al. 2005). Otherbloom-forming species present included Prymnesiumparvum.
Haptophytes can be distinguished to a certain extentat the species level in LM, but the scale and coccolithmorphology represents the main taxonomic feature foraccurate species identification. Four saddle-shapedChrysochromulina species (C. acantha, C. cf. cymbium,C. simplex, and C. throndsenii) were observed in the Ner-vion River estuary. They all appeared very similar in LM,but were subsequently identified (in electron microscopy)by the ornamentation of their organic scales. Taxa, suchas Prymnesium sp., which is also known as the ‘‘eyelashChrysochromulina’’, appears identical in LM to C. limoniaJensen et Moestrup and C. polylepis, and its identityneeds to be verified by electron microscopy. Accordingto Billard and Gayral (1972) and Green and Pienaar(1977), scale size has been used as a criterion to differ-entiate species of Isochrysis Parke. In the Nervion Riverestuary, I. galbana was recognized by the larger size ofits cell body scales compared to I. litoralis Billard et Gay-ral and I. maritime Billard et Gayral. In the case of thegenus Pavlova, the different structure and size of theknob-scales may allow separation at the species level(Green 1980). P. gyrans was differentiated from the close-ly related P. pinguis Green by the size of the bilobedknob-scales and the morphology of the short flagellum,which is abruptly tapering in P. pinguis as opposed togradually tapering in P. gyrans. Pleurochrysis roscoffensisand P. pseudoroscoffensis Gayral et Fresnel share mostmorphological features, but they can be differentiatedfrom one another by small protrusions which P. roscof-fensis has on the distal surface of its coccoliths (Younget al. 2003). Prymnesium parvum and P. patelliferumGreen, Hibberd et Pienaar were originally described astwo distinct species because of differences in scale mor-phology, but recent molecular studies demonstrated thatin fact they represent two forms of the same species(Larsen and Medlin 1997, Larsen 1999).
Acknowledgements
This work was financially supported by the University of theBasque Country (project 9/UPV00118.310-15339/2003) and acontract from the University of the Basque Country to S.Seoane. We thank A. Laza for his invaluable assistance in iso-lating some of the cultures and L. Medlin for contributing usefulcomments on the manuscript.
References
Andersen, R.A., R.R. Bidigare, M.D. Keller and M. Latasa. 1996.A comparison of HPLC pigment signatures and electronmicroscopic observations for oligotrophic waters of the NorthAtlantic and Pacific Oceans. Deep-Sea Res. II 43: 517–537.
Billard, C. and P. Gayral. 1972. Two new species of Isochrysiswith remarks on the genus Ruttnera. Br. Phycol. J. 7: 289–297.
Birkhead, M. and R.N. Pienaar. 1994. The ultrastructure of Chry-sochromulina brevifilum (Prymnesiophyceae). Eur. J. Phycol.29: 267–280.

58 S. Seoane et al.: Haptophytes of the Nervion River estuary
Article in press - uncorrected proof
Birkhead, M. and R.N. Pienaar. 1995a. The flagellar apparatusof Chrysochromulina sp. (Prymnesiophyceae). J. Phycol. 31:96–108.
Birkhead, M. and R.N. Pienaar. 1995b. The taxonomy and ultra-structure of Chrysochromulina simplex (Prymnesiophyceae).Phycologia 34: 145–156.
Borja, A., I. Galparsoro, O. Solaun, I. Muxika, E.M. Tello, A.Uriarte and V. Valencia. 2006. The European water frameworkdirective and the DPSIR, a methodological approach toassess the risk of failing to achieve good ecological status.Estuar. Coast. Shelf Sci. 66: 84–96.
Butcher, R.W. 1952. Contributions to our knowledge of thesmaller marine algae. J. Mar. Biol. Ass. UK 31: 175–191.
Carreto, J.L., N.G. Montoya, H.R. Benavides, R. Guerrero andM.O. Carignan. 2003. Characterization of spring phytoplank-ton communities in the Rio de La Plata maritime front usingpigment signatures and cell microscopy. Mar. Biol. 143:1013–1027.
Chadefaud, M. and J. Feldmann. 1949. Sur une Coccolithopho-racee des bacs de l’aquarium de la Station biologique deRoscoff. Bull. Mus. Natl. Hist. Nat., 2e Ser. 21: 617–621.
Chretiennot-Dinet, M.-J., M.-M. Giraud-Guille, D. Vaulot, J.-L.Puteaux, Y. Saito and H. Chanzy. 1997. The chitinous natureof filaments ejected by Phaeocystis (Prymnesiophyceae). J.Phycol. 33: 666–672.
Dahl, E., B. Edvardsen and W. Eikrem. 1998. Chrysochromulinablooms in the Skagerrak after 1988. In: (B. Reguera, J. Blan-co, M.L. Fernandez and T. Wyatt, eds.) Harmful algae. Xuntade Galicia and Intergovernmental Oceanographic ofUNESCO, Santiago de Compostela. pp. 104–105.
Dahl, E., E. Bagoien, B. Edvardsen and N.C. Stenseth. 2005. Thedynamics of Chrysochromulina species in the Skagerrak inrelation to environmental conditions. J. Plankton Res. 54:15–24.
Dangeard, P. 1934. Sur l’epiphytisme d’une Coccolithinee ren-contree a Roscoff. P.V. Soc. Linn. Bordeaux 10 janvier:45–47.
DiTullio, G.R., M.E. Geesey, D.R. Jones, K.L. Daly, L. Campbelland W.O. Smith Jr. 2003. Phytoplankton assemblage struc-ture and primary productivity along 1708W in the South Pacif-ic Ocean. Mar. Ecol. Prog. Ser. 255: 55–80.
Eikrem, W. 1996. Chrysochromulina throndsenii sp. nov. (Prym-nesiophyceae). Description of a new haptophyte flagellatefrom Norwegian waters. Phycologia 35: 377–380.
Eikrem, W. 1999. The class Prymnesiophyceae (Haptophyta) inScandinavian waters. Aspects of taxonomy, abundance,diversity and nutrition. PhD Thesis. University of Oslo, Nor-way. 229 pp.
Eikrem, W. and B. Edvardsen. 1999. Chrysochromulina fragariasp. nov. (Prymnesiophyceae), a new haptophyte flagellatefrom Norwegian waters. Phycologia 38: 149–155.
Eikrem, W. and J. Throndsen. 1999. The morphology of Chry-sochromulina rotalis sp. nov. (Prymnesiophyceae, Haptophy-ta) from the Skagerrak. Sarsia 84: 445–449.
Estep, K.W., P.G. Davis, P.E. Hargraves and J.M. Sieburth. 1984.Chloroplast containing microflagellates in natural populationsof north Atlantic nanoplankton, their identification and distri-bution; including a description of five new species of Chry-sochromulina (Prymnesiophyceae). Protistologica 20: 613–634.
Gayral, P. and J. Fresnel. 1976. Nouvelles observations sur deuxCoccolithophoracees marines: Cricosphaera roscoffensis (P.Dangeard) comb. nov. et Hymenomonas globosa (F. Magne)comb. nov. Phycologia 15: 339–355.
Gayral, P. and J. Fresnel. 1983. Platychrysis pienaarii sp. nov. etP. simplex sp. nov. (Prymnesiophyceae): description et ultra-structure. Phycologia 22: 29–45.
Green, J.C. 1980. The fine structure of Pavlova pinguis Greenand a preliminary survey of the order Pavlovales (Prymnesio-phyceae). Br. Phycol. J. 15: 151–191.
Green, J.C. and I. Manton. 1970. Studies in the fine structureand taxonomy of flagellates in the genus Pavlova. I. A revi-sion of Pavlova gyrans, the type species. J. Mar. Biol. Ass.UK 50: 1113–1130.
Green, J.C. and R.N. Pienaar. 1977. The taxonomy of the orderIsochrysidales (Prymnesiophyceae) with special reference tothe genera Isochrysis Parke, Dicrateria Parke and ImantoniaReynolds. J. Mar. Biol. Ass. UK 57: 7–17.
Green, J.C., D.J. Hibberd and R.N. Pienaar. 1982. The taxonomyof Prymnesium (Prymnesiophyceae) including a descriptionof a new cosmopolitan species, P. patellifera sp. nov., andfurther observations on P. parvum N. Carter. Br. Phycol. J.17: 363–382.
Green, J.C., P.A. Course and G.A. Tarran. 1996. The life-cycle ofEmiliania huxleyi: a brief review and a study of relative ploidylevels analysed by flow cytometry Check spelling. J. Mar.Syst. 9: 33–44.
Guillard, R.R.L. and J.H. Ryther. 1962. Studies of marine plank-tonic diatoms. I. Cyclotella nana Hustedt and Detonulaconfervacea Cleve. Can. J. Microbiol. 8: 229–239.
Hadju, S., U. Larsson and Ø. Moestrup. 1996. Seasonal dynam-ics of Chrysochromulina species (Prymnesiophyceae) in acoastal area and a nutrient-enriched inlet of the Northern Bal-tic proper. Bot. Mar. 39: 281–295.
Hansen, P.J. and M. Hjorth. 2002. Growth and grazing responsesof Chrysochromulina ericina (Prymnesiophyceae): the role ofirradiance, prey concentration and pH. Mar. Biol. 141:975–983.
Hu, X., M.Y. Yin and C.K. Tseng. 2005. Morphology of Chryso-chromulina planisquama sp. nov. (Haptophyta, Prymnesio-phyceae) isolated from Jiaozhou Bay, China. Bot. Mar. 48:52–57.
Jensen, M.Ø. 1998. The genus Chrysochromulina (Prymnesio-phyceae) in Scandinavian coastal waters: diversity, abun-dance and ecology. PhD Thesis. University of Copenhagen,Copenhagen. 129 pp.
Jones, H.L.J., B.S.C. Leadbeater and J.C. Green. 1993. Mixo-trophy in marine species of Chrysochromulina (Prymnesio-phyceae): ingestion and digestion of a small green flagellate.J. Mar. Biol. Ass. UK 73: 283–296.
Lampert, L., B. Queginer, T. Labasque, A. Pichon and N. Lebre-ton. 2002. Spatial variability of phytoplankton compositionand biomass on the eastern continental shelf of the Bay ofBiscay (north-east Atlantic Ocean). Evidence for a bloom ofEmiliania huxleyi (Prymnesiophyceae) in spring 1998. Cont.Shelf Res. 22: 1225–1247.
Larsen, A. 1999. Prymnesium parvum and Prymnesium patelli-ferum (Haptophyta) – one species. Phycologia 38: 541–543.
Larsen, A. and L.K. Medlin. 1997. Inter- and intraspecific geneticvariation in twelve Prymnesium (Haptophyceae) clones. J.Phycol. 33: 1007–1015.
Latasa, M., X.A.G. Moran, R. Scharek and M. Estrada. 2005.Estimating the carbon flux through main phytoplanktongroups in the northwestern Mediterranean. Limnol. Ocea-nogr. 50: 1447–1458.
Leadbeater, B.S.C. and I. Manton. 1969. Chrysochromulinacamella sp. nov. and C. cymbium sp. nov., two new relativesof C. strobilus Parke and Manton. Arch. Mikrobiol. 68:116–132.
Leadbeater, B.S.C. and I. Manton. 1971. Fine structure and lightmicroscopy of a new species of Chrysochromulina (C. acan-tha). Arch. Mikrobiol. 78: 58–69.
LeRoi, J.-M. and G. Hallegraeff. 2004. Scale-bearing nanofla-gellates from southern Tasmanian coastal waters, Australia.I. Species of the genus Chrysochromulina (Haptophyta). Bot.Mar. 47: 73–102.
Lessard, E.J., A. Merico and T. Tyrrell. 2005. Nitrate:phosphateratios and Emiliania huxleyi blooms. Limnol. Oceanogr. 50:1020–1024.
Lohrenz, S.E., C.L. Carroll, A.D. Weidemann and M. Tuel. 2003.Variations in phytoplankton pigments, size structure andcommunity composition related to wind forcing and water

S. Seoane et al.: Haptophytes of the Nervion River estuary 59
Article in press - uncorrected proof
mass properties on the North Carolina inner shelf. Cont. ShelfRes. 23: 1447–1464.
Magne, F. 1954. Les Chrysophycees marines de la station bio-logique de Roscoff. Rev. Gen. Bot. 61: 389–415.
Manton, I. 1978. Chrysochromulina hirta sp. nov., a widely dis-tributed species with unusual spines. Br. Phycol. J. 13: 3–14.
Moestrup, Ø. 1979. Identification by electron microscopy ofmarine nanoplankton from New Zealand including thedescription of four new species. N.Z. J. Bot. 17: 61–95.
Nakayama, T., M. Yoshida, M.-H. Noel, M. Kawachi and I.Inouye. 2005. Ultrastructure and phylogenetic position ofChrysoculter rhomboideus gen. et sp. nov. (Prymnesiophy-ceae), a new flagellate haptophyte from Japanese coastalwaters. Phycologia 44: 369–383.
Not, F., R. Massana, M. Latasa, D. Marie, C. Colson, W. Eikrem,C. Pedros-Alio, D. Vaulot and N. Simon. 2005. Late summercommunity composition and abundance of photosyntheticpicoeukaryotes in Norwegian and Barents Seas. Limnol.Oceanogr. 50: 1677–1686.
Orive, E. 1989. Differences in phytoplankton compositionbetween the Abra of Bilbao and the adjacent shelf waters.Hydrobiologia 182: 121–135.
Paasche, E. 2002. A review of the coccolithophorid Emilianiahuxleyi (Prymnesiophyceae), with particular reference togrowth, coccolith formation, and calcification-photosynthesisinteractions. Phycologia 40: 503–529.
Parke, M. 1949. Studies on marine flagellates. J. Mar. Biol. Ass.UK 28: 255–288.
Parke, M. and I. Manton. 1962. Studies on marine flagellates VI.Chrysochromulina pringsheimii sp. nov. J. Mar. Biol. Ass. UK42: 391–404.
Peperzak, L., F. Colijn, E.G. Vrieling, W.W.C. Gieskes and J.C.H.Peters. 2000. Observations of flagellates in colonies ofPhaeocystis globosa (Prymnesiophyceae); a hypothesis fortheir position in the life cycle. J. Plankton Res. 22: 2181–2203.
Pienaar, R.N. and V. Bandu. 1984. A new species of Chryso-chromulina (Prymnesiophyceae) from Natal inshore waters.Proc. Electron Micros. Soc. Southern Africa 14: 65–66.
Puigserver, M., M.-J. Chretiennot-Dinet and E. Nezan. 2003.Some Prymnesiaceae (Haptophyta, Prymnesiophyceae) fromthe Mediterranean Sea with the description of two new spe-cies Chrysochromulina lanceolata sp. nov. and C. pseudo-lanceolata sp. nov. J. Phycol. 39: 762–774.
Reynolds, N. 1974. Imantonia rotunda gen. et sp. nov., a newmember of the Haptophyceae. Br. Phycol. J. 9: 429–434.
Rhodes, L. and B. Burke. 1996. Morphology and growth char-acteristics of Chrysochromulina species (Haptophyceae-sPrymnesiophyceae) isolated from New Zealand coastalwaters. N.Z. J. Mar. Freshw. Res. 30: 91–103.
Rodrıguez, F., Y. Pazos, J. Maneiro and M. Zapata. 2003. Tem-poral variation in phytoplankton assemblages and pigmentcomposition at a fixed station of the Ria of Pontevedra (NWSpain). Estuar. Coast. Shelf Sci. 58: 499–515.
Schoemann, W., S. Becquevort, J. Stefels, W. Rousseau and C.Lancelot. 2005. Phaeocystis blooms in the global ocean andtheir controlling mechanisms: a review. J. Sea Res. 53:43–66.
Seoane, S., A. Laza, I. Urrutxurtu and E. Orive. 2005. Phyto-plankton assemblages and their dominant pigments in theNervion River Estuary. Hydrobiologia 549: 1–13.
Seoane, S., A. Laza and E. Orive. 2006. Monitoring phytoplank-ton assemblages in estuarine waters: the application of pig-ment analysis and microscopy to size-fractionated samples.Estuar. Coast. Shelf Sci. 67: 343–354.
Silva, P.C., J. Throndsen and W. Eikrem. 2007. Revisiting thenomenclature of haptophytes. Phycologia 46: 471–475.
Thomsen, H.A., K.R. Buck and F.P. Chavez. 1994. Haptophytesas components of marine phytoplankton. In: (J.C. Green andB.S.C. Leadbeater, eds.) The haptophyte algae. ClarendonPress, Oxford. pp. 187–208.
Throndsen, J. 1978. The dilution culture method. In: (A. Sournia,ed.) Phytoplankton manual. Monographs on oceanographicmethodology 6. UNESCO, Paris. pp. 218–224.
Van Lenning, K., M. Latasa, M. Estrada, A.G. Saez, L. Medlin, I.Probert, B. Veron and J. Young. 2003. Pigment signaturesand phylogenetic relationships of the Pavlovophyceae (Hap-tophyta). J. Phycol. 39: 379–389.
Young, J.R., M. Geisen, L. Cros, A. Kleijne, C. Sprengel, I. Pro-bert and J.B. Østergaard. 2003. A guide to extant Coccoli-thophore taxonomy. J. Nannoplankton Res. (Special Issue) 1:1–125.
Zapata, M., S.W. Jeffrey, S.W. Wright, F. Rodriguez, J.L. Garridoand L. Clementson. 2004. Photosynthetic pigments in 37species (65 strains) of Haptophyta: implications for ocean-ography and chemotaxonomy. Mar. Ecol. Prog. Ser. 270:83–102.
Received 21 September, 2007; accepted 7 October, 2008




















