
Production of microcystins in calcareous Mediterranean streams: The Alharabe River, Segura River...
13
Journal of Applied Phycology (2005) 17: 231–243 DOI: 10.1007/s10811-005-2999-z C Springer 2005 Production of microcystins in calcareous Mediterranean streams: The Alharabe River, Segura River basin in south-east Spain Marina Aboal 1,∗ , M. ´ Angeles Puig 2 & Antonia D. Asencio 3 1 Laboratorio de Algolog´ ıa, Departamento de Biolog´ ıa Vegetal, Facultad de Biolog´ ıa, Universidad de Murcia, Campus de Espinardo, E-30100 Murcia, Spain; 2 Departamento de Biogeoqu´ ımica Aquatica, Centro de Estudios Avanzados de Blanes (CEAB-CSIC), Carrer d’acc` es a la Cala Sant Francesc, 14, 17300 Blanes, Girona, Spain; 3 Departamento de Biolog´ ıa Aplicada (Bot ´ anica), Edificio La Galia, Universidad Miguel Hern ´ andez, 03202 Elche, Alicante, Spain ∗ Author for correspondence: e-mail: [email protected]; fax: +34-968-363963 Received 20 November 2004; revised and accepted 25 February 2005 Key words: cyanotoxins, calcareous rivers, microcystins, benthic cyanobacteria, Mediterranean, SE Spain, toxicity Abstract The development of epilithic cyanobacteria communities in a Mediterranean calcareous stream in the province of Murcia (SE Spain) was studied during the course of one year in an attempt to clarify the environmental variables that influence the production of microcystins. The predominant cyanobacteria were species of Rivularia, which formed conspicuous colonies throughout the year. Seasonally, other species were abundant: Schizothrix fasciculata, Toly- pothrix distorta and Phormidium splendidum. All the species collected produced microcystins to a varying degree (up to five varieties), while the benthic community as a whole produced concentrations as high as 20.45 mg m −2 . At the same time, the presence of microcystins dissolved in water was confirmed. Among environmental variables, air temperature and silicate content were positively and strongly correlated with total microcystins, while nitrite, nitrate, orthophosphate, calcium and flow were negatively correlated with them. Dissolved microcystins were negatively correlated with microcystin LR, P.A.R. and total phosphorus and positively with rainfall. The production of micro- cystin YR seems to be regulated by different factors from those regulating the other main varieties (microcystin LR and microcystin RR). The data obtained indicate that all the tested benthic cyanobacteria produced microcystins in this shallow calcareous stream, which may contribute to their predominance in the prevailing conditions. The accumulation of microcystins in mucilaginous colonies of other groups of algae poses new questions concerning the possible ecological function of these compounds and needs further study. Introduction The health risks involved in consuming water contam- inated by cyanotoxins, which may be lethal in acute cases and carcinogenic following the long term con- sumption of small quantities, have led to countries en- courage their study so that they may be readily iden- tified and quantified (Falconer, 1999). The increased number of cases of toxicity has led the WHO to propose protocols for the identification of the different types of toxins (hepatotoxins and neurotoxins) and to recom- mend maximum levels in waters destined for different uses, mainly drinking (Chorus & Bartram, 1999; Codd et al., 1999; Figueiredo et al., 2004). More recently, the genetic identification of toxic strains has been under- taken (Bolch et al., 1999; Dyble et al., 2002) but the factors promoting microcystin synthesis are still not known with precision. In most cases, the available data refer to the toxi- city produced by a relatively small number of plank- tonic species, while benthic communities have been less studied since the overall number of intoxica- tions produced by them is much lower (Edwards et al., 1992; Owen, 1994). Furthermore, all the known studies of benthic cyanobacteria toxicity have dealt with lakes, reservoirs or water depuration systems
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Production of microcystins in calcareous Mediterranean streams: The Alharabe River, Segura River...

Journal of Applied Phycology (2005) 17: 231–243DOI: 10.1007/s10811-005-2999-z C© Springer 2005
Production of microcystins in calcareous Mediterranean streams:The Alharabe River, Segura River basin in south-east Spain
Marina Aboal1,∗, M. Angeles Puig2 & Antonia D. Asencio3
1Laboratorio de Algologıa, Departamento de Biologıa Vegetal, Facultad de Biologıa, Universidad de Murcia,Campus de Espinardo, E-30100 Murcia, Spain; 2Departamento de Biogeoquımica Aquatica, Centro de EstudiosAvanzados de Blanes (CEAB-CSIC), Carrer d’acces a la Cala Sant Francesc, 14, 17300 Blanes, Girona, Spain;3Departamento de Biologıa Aplicada (Botanica), Edificio La Galia, Universidad Miguel Hernandez, 03202 Elche,Alicante, Spain
∗Author for correspondence: e-mail: [email protected]; fax: +34-968-363963
Received 20 November 2004; revised and accepted 25 February 2005
Key words: cyanotoxins, calcareous rivers, microcystins, benthic cyanobacteria, Mediterranean, SE Spain, toxicity
Abstract
The development of epilithic cyanobacteria communities in a Mediterranean calcareous stream in the province ofMurcia (SE Spain) was studied during the course of one year in an attempt to clarify the environmental variables thatinfluence the production of microcystins. The predominant cyanobacteria were species of Rivularia, which formedconspicuous colonies throughout the year. Seasonally, other species were abundant: Schizothrix fasciculata, Toly-pothrix distorta and Phormidium splendidum. All the species collected produced microcystins to a varying degree(up to five varieties), while the benthic community as a whole produced concentrations as high as 20.45 mg m−2. Atthe same time, the presence of microcystins dissolved in water was confirmed. Among environmental variables, airtemperature and silicate content were positively and strongly correlated with total microcystins, while nitrite, nitrate,orthophosphate, calcium and flow were negatively correlated with them. Dissolved microcystins were negativelycorrelated with microcystin LR, P.A.R. and total phosphorus and positively with rainfall. The production of micro-cystin YR seems to be regulated by different factors from those regulating the other main varieties (microcystin LRand microcystin RR). The data obtained indicate that all the tested benthic cyanobacteria produced microcystinsin this shallow calcareous stream, which may contribute to their predominance in the prevailing conditions. Theaccumulation of microcystins in mucilaginous colonies of other groups of algae poses new questions concerningthe possible ecological function of these compounds and needs further study.
Introduction
The health risks involved in consuming water contam-inated by cyanotoxins, which may be lethal in acutecases and carcinogenic following the long term con-sumption of small quantities, have led to countries en-courage their study so that they may be readily iden-tified and quantified (Falconer, 1999). The increasednumber of cases of toxicity has led the WHO to proposeprotocols for the identification of the different types oftoxins (hepatotoxins and neurotoxins) and to recom-mend maximum levels in waters destined for differentuses, mainly drinking (Chorus & Bartram, 1999; Codd
et al., 1999; Figueiredo et al., 2004). More recently, thegenetic identification of toxic strains has been under-taken (Bolch et al., 1999; Dyble et al., 2002) but thefactors promoting microcystin synthesis are still notknown with precision.
In most cases, the available data refer to the toxi-city produced by a relatively small number of plank-tonic species, while benthic communities have beenless studied since the overall number of intoxica-tions produced by them is much lower (Edwardset al., 1992; Owen, 1994). Furthermore, all the knownstudies of benthic cyanobacteria toxicity have dealtwith lakes, reservoirs or water depuration systems

232
(Aboal & Puig, 2005; Baker et al., 2001; Marsaleket al., 1997; Mez et al., 1998). The information referringto the production of toxins in rivers is scarce despitethe fact that cyanophytes (cyanobacteria) may be thepredominant group throughout the year in calcareousstreams and that many authors consider it more thanprobable that the capacity to produce toxic compoundsis a universal characteristic of cyanobacteria (Dow &Swoboda, 2000). In some calcareous streams the pro-duction of toxins and their influence on the decreaseof diversity of macroinvertebrate and diatom commu-nities has been studied (Aboal et al., 2000, 2002).
Besides the huge amount of data produced in re-cent years, the ecological or physiological function ofmicrocystins is still not known. Antigrazing (Henninget al., 2001; Kurmayer & Juttner, 1999) and allelo-pathic activities (Gromov et al., 1991; Kearns & Hunter,2001; Schagerl et al., 2002; Valdor, 2003) have beenproposed. Some authors have observed an increase inthe synthesis of microcystins in the presence of herbi-vores in planktic communities (Jang et al., 2004) andeven toxic or lethal effects on aquatic macroinverte-brates (M. Puig, unpublished data).
The Mediterranean region will surely be one of themost affected by global warming, a view that, togetherwith the abundance of calcareous materials, could wellfavour cyanobacterial growth (Margalef, 1983). Theproduction of microcystins may not only have an effecton ecological succession, but also on human health ifcontaminated waters are used, for example, to providedrinking water to country villages, to water cattle orirrigate horticultural crops.
In view of the scarcity of data concerning this prob-lem, we decided to investigate whether river benthiccyanobacteria produce microcystins and, if so, whetherthere is a predictable seasonal dynamic involved andwhat environmental factors might modulate their syn-thesis and release.
Study area
The Alharabe is a tributary of the Segura River in thenorth-west of the province of Murcia, where the highestrainfall of the province occurs (Figure 1). The samplingsites (ALH1 and ALH2) were situated in a forestedzone of medium sized mountains (Table 1), where agri-culture, cattle grazing and human population have littleinfluence.
The substrate is rocky with a predominance ofblocks and boulders, although some stretches are siltyor sandy. The geological materials are basically lime-
Table 1. Localization and main hydrological and climaticcharacteristics of the Alharabe River during the annual cyclestudied (May 2001–May 2002)a.
Alharabe ALH 1 ALH 2
Altitude (m) 900 1000
Orientation NW SE
Flow (L s−1) 0.8–112.0 0.3–155.1
Mean 29.4 42.5
Maximum depth (cm) 15 20
Maximum width (m) 3 6
Rainfall (mm) 0–1095 0–1095
Mean 521.2 521.2
Maximum air temperature (◦C) 14–34.3 14–34.3
Mean 21.4 21.4
Minimum air temperature (◦C) 3.2–19.2 3.2–19.2
Mean 10.0 10.0
aMinimum, maximum and means are indicated.
Figure 1. Location of the sampling points.
stone and dolomites. The river runs through a pine for-est and bank-side vegetation is well conserved. Themean temperature during the study was 15.7 ◦C withminimum and maximum means of 10 and 21.4 ◦C, inDecember and July respectively. The annual rainfall forthe years in question (2001 and 2002) was 521.2 mm,with December being the rainiest month. Climaticdata corresponded to the weather station nearest theAlharabe basin (Benizar, Albacete) and were providedby the Spanish National Meteorological Centre.
The area is well preserved and is one of the mostbio-diverse of SE Spain from a zoological and botanicalpoint of view (Aboal, 1989; Aboal et al., 2003).

233
Material and methods
Sampling
Sampling took place monthly from May 2001 to May2002. Depth and current were measured every 20 cm ateach sampling site using a rigid meter and a FLOMATE2000 flow meter. The calculation of discharge wasmade according to the method decribed by Gordon et al.(2004).
The pH and water temperature were measured usinga WTW 95/1 pHmeter, conductivity with a WTW LF95 conductivimeter and oxygen with a CRISON OXY96 oxymeter. Light intensity (PAR) was measured witha LICOR LI-92 SA underwater sensor coupled to aLICOR LI-1000–32 datalogger, storing the maximum,minimum and mean values for periods of 15 min duringthe time that sampling lasted.
A 12-cm2 section of the epilithic community wasscraped off and frozen in liquid nitrogen in the fieldfor later extraction of microcystins in the laboratory.Single colonies of organisms forming conspicuouscolonies were also collected and frozen in the fieldfor later extraction. Additional samples were collectedand transported refrigerated to the laboratory where,after a preliminary examination, they were fixedwith 4% formaline for later microscopic study orcultured.
To quantify chlorophylls and phycobilins, six repli-cate samples of 4 cm2 were scraped (three for chloro-phylls and three for phycobilins). These samples werefrozen in the field with liquid nitrogen.
The water samples for detecting microcystins werecollected by syringe from the vicinity of the colonies,filtered through a 0.45 µm Millipore filter and frozenin the field with liquid nitrogen, while water samplesfor chemical analysis were collected and transportedin cool conditions to the laboratory, where they werefiltered and frozen. An additional sample was fixed withchloroform in the field for calcium and total alkalinityquantification.
Analysis of waters
Chemical analyses were all completed within aweek of field sample collection. Ammonia, nitrate,nitrite, orthophosphate and silicate were analysedon a Bran-Luebbe TRAACS 2000 Autoanalyser(methods 98-70W, J-002-88E, J-003-88E, G-033-92Cand J-005-88D, respectively), using the hypochlorite-oxalic acid method for NH4-N (Sawyer & Grisley,
1967), the cadmium-copper reduction method forNO3-N (APHA, 1998), the sulphanilamide-NEDdihydrochloride colorimetric method for NO2-N(APHA, 1998) and the molybdenum blue colorimetricmethod for PO4-P (Murphy & Riley, 1962). Totalphosphorus concentration was analysed with the samemethod as ortophosphate after digestion using thenitric acid-sulphuric acid method (APHA, 1998).The level of detection was 0.01 µM L−1 in all cases.Organic phosphorus was calculated as the differenceof total phosphorus and orthophosphate. Whenorthophosphate was below the detection limit, a valueequal to half the detection limit was used for statisticalpurposes (Turner et al., 2003a). Dissolved inorganicnitrogen (DIN) was estimated as nitrate and nitrite,and the DIN: P molar ratio from total phosphorus.
The concentration of calcium was measured bythe selective ion analyser 830 ORION equipped with9720BN ORION sensor. Total alkalinity was estimatedby titration with sulphuric acid and phenolphtalein ac-cording to method 2320 (APHA, 1998).
Microscopic studies, cultures and identification
The colonies collected in the field and refrigerated wereseparated upon arrival at the laboratory with an OLYM-PUS TL2 stereomicroscope. A portion of the materialwas fixed for later microscopic study. All samples weretreated with 5% EDTA to eliminate carbonate depositsand then embedded in glycerine before thin sectionswere cut with a microtome (when necessary). Obser-vations were made with bright - light and phase contrastmicroscopy.
Some cyanobacteria colonies were isolated inBG11, BBM and CHU 10D media according to Zehn-der (1985) and deposited in the culture collection ofthe Laboratory of Algology of Murcia University. Thecultures obtained were tested for microcystins usingimmunological methods (see below).
Pigment extraction and quantification
Chlorophylls were extracted after sonication in the darkwith acetone (90%) at 4 ◦C for 24 h and quantifiedspectrophotometrically according to the trichromaticmethod (Jeffrey & Humphrey, 1975). Carotenoids wereextracted with 90% acetone and quantified spectropho-tometrically. Readings at 430 nm were used to calcu-late total carotenoids and the Margalef index (Mar-galef, 1983). The phycobilins were extracted withsaline phosphate buffer (PBS) of pH 7 for 4 h at

234
4 ◦C and quantified spectrophotometrically accordingto Bennett and Bogorad (1973).
Intracellular microcystin quantification
Microcystins were extracted with methanol accordingto methods described by Gjolme and Utkilen (1996)and the intracellular microcystins content was mea-sured using high performance liquid chromatographywith an internal surface reverse-phase column (Gjolme& Utkilen, 1996; Meriluoto et al., 1990). The mobilephase was in a gradient from 70% TFA (0.5 mL L−1)to 0%, and 30% TFA in acetonitrile (mL L−1) to 100%in 45 min and back to 70% TFA in water and 30% TFAin acetonitrile at 46 until 55 min. The flow rate was1 mL min−1. The detector was a photodiode array setbetween 200 and 300 nm. The toxin content was quan-tified by calibrating against purified toxin standards(microcystin RR, microcystin LR and microcystin YR)from Calbiochem
R©. Unidentified microcystin varieties
were detected by their UV-spectra and quantified asmicrocystin LR equivalents. The detection level was0.02 µg mL−1.
Quantification dissolved microcystins
Microcystins dissolved in water were quantified withthe ELISA Plate Kit 75400 from Strategic DiagnosticInc. Envirogard
R©kit, a more precise method than the
chromatographic method and with a level of detectionof 0.1 ppb.
Statistical analysis
The STATISTICA program v. 5.5 was used to calculatesimple statistics and the correlation matrix.
Results
The Alharabe River is a shallow permanent stream andit has a typical Mediterranean flow regime (Table 1)with peaks that usually coincide with rainfall: the high-est discharge in the year studied was detected in spring(April) with a second peak in December.
Total alkalinity and calcium concentration were1.24–5.25 meq L−1 and 11.7–99.5 ppm, respectively,with the highest values in winter and summer (Table 2).
The concentration of nitrate and nitrite was 0.17–6.41 mg L−1 and 3.13–22.59 µg L−1, respectively, bothwith minimum values in summer and maximum values
Table 2. Physichal and chemical characteristics of the Alharabe River.
ALH 1 ALH 2
Silicate-Si (mg L−1)
Min–max 2.4–8.1 3.1–8.0
Mean 4.8 5.3
PO4-P (µg L−1)
Min–max <0.01–71.2 <0.01–67.4
Mean 23.7 18.1
Total P (µg L−1)
Min-max 14.3–65.2 13.7–59.6
Mean 34.07 33.76
Organic P (µg L−1)
Min–max 1.7–62.9 0.5–57.9
Mean 26.3 27.6
NO2-N (µg L−1)
Min–max 3.1–22.6 3.7–15.6
Mean 9.7 7.8
NO3-N (mg L−1)
Min–max 0.27–5.3 0.3–6.8
Mean 2.2 2.2
Total alkalinity (meq L−1)
Min–max 1.8–5.3 1.2–5.0
Mean 3.7 3.4
DIN: P
Min–max 2.9–217.4 2.9–224.5
Mean 44.53 44.81
Calcium (mg L−1)
Min–max 18.1–97.8 31.7–111.7
Mean 57.7 65.2
Flow (L s−1)
Min–max 0.8–112.0 0.3–155.1
Mean 29.4 42.5
PAR (µmol photon cm−1 s−1)
Min–max 41.0–366.4 41.0–117.2
Mean 117.2 117.2
Chlorophyll a (mg m−2)
Min–max 0.3–4.6 0.5–7.9
Mean 2.0 2.4
Chlorophyll c (mg m−2)
Min–max 0.1–1.5 0.1–1.3
Mean 0.6 0.6
Total phycobilin (µg m−2)
Min–max <0.01–1.5 <0.01–1.6
Mean 0.1 0.1
Total intracellular microcystin (mg m−2)
Min–max 1.7–19.7 2.0–22.1
Mean 10.7 12.7
Dissolved microcystin (µg L−1)
Min–max <0.01–0.7 <0.01–0.2
Mean 0.3 0.1

235
Figure 2. Changes in flow and nitrate concentration during the year studied in the Alharabe River.
in winter (Figure 2). Ammonium was not detected atany time during the sampling period. Orthophosphatewas below the detection limit in summer and reachedmaximum values (67.43 µg L−1) in winter (ALH 2) orspring (ALH 1), whilst the lowest organic phospho-rus concentration was detected in winter and the high-est in spring-summer (Figure 3). The total phosphorusconcentration in stream water varied between 13.69and 65.2 µg L−1, with a maximum value in spring. TheDIN: P ratio showed considerable seasonal variation,reaching the lowest values in summer and the highestin winter (Table 2). The silicate concentration variedfrom 2.42 to 7.98 mg L−1 with a maximum value insummer and minimum in winter.
The predominant group of algae on all lithic sur-faces belonged to the Cyanophyta, especially Rivulariabiasolettiana Meneghini and R. haematites (D. C.)Agardh, both of which formed extensive populationsthroughout the year (Figure 4). Conspicuous growthsof other cyanophytes developed seasonally, includ-ing Tolypothrix distorta Kutzing (September) andPhormidium splendidum (Grev. ex Gom.) Anagn. etKom.(March and August-September). The chlorophyll
a concentration reached the highest values in summerwhilst total phycobilin and chlorophyll c concentra-tions peaked in spring (Table 2).
Intracellular microcystins were detected throughoutthe year with peaks in May-June and mean values of1.70–22.10 µg m−2 for total intracellular microcystins(Figure 5). Microcystin LR was below the detectionlevel in December and reached the highest concentra-tion in June-July (Figure 6). Microcystins RR and YRwere always detectable and reached their maxima inautumn and spring, respectively. Total intracellular mi-crocystins were positively correlated with all micro-cystin forms identified, except microcystin YR, and airtemperature (Figure 7) and negatively with nitrate, ni-trite, calcium, silicate (Figure 8) and flow (Table 3).Microcystin YR was positively correlated with DIN: Pratio whilst microcystin RR showed a slight positivecorrelation with carotenoids and some of the phyco-bilins (Table 4). Dissolved microcystins were alwayspresent and the highest values were detected at theend of summer (Table 2); they were positively corre-lated with rainfall and negatively with total phosphorus,chlorophyll a, PAR and microcystin LR (Table 3).

236
Figure 3. Annual change of ortophosphate and organic phosphorus in the Alharabe River.
During the year studied, a constant production ofmicrocystins by both Rivularia biasolettiana and R.haematites was detected, with values ranging from 1.12to 16.47 µg g−1 and 2.49 to 15.62 µg g−1, respectively.All species seemed to produce different proportionsof microcystins in the different seasons of the year:the predominant form was microcystin LR except inautumn when microcystin YR showed the highest con-centration in Tolypothrix distorta and microcystin RRin Rivularia biasolettiana or microcystin 2 in Phormid-ium splendidum (Table 4).
The mucilaginous colonies of diatoms (Cymbellaaffinis Kutzing) and chlorophytes (Chaetophora in-crassata (Hudson) Hazen) contained substantial quan-tities of microcystins (5.977 µg g−1) of which micro-cystin LR was predominant with 1.897 µg g−1.
Discussion
The highest concentrations of nitrate and nitrite wererecorded in the Alharabe River in winter, with a secondpeak in spring, while the total P concentration reached
the maximum in spring-summer; however inorganicphosphorus (PO4-P) was below the detection limit insummer. This pattern of variation may be comparedwith that reported for other shallow calcareous streamsdominated by cyanophytes in which the maximum val-ues of nitrate and total P have been reported in sum-mer (Chapman et al., 2001) or spring (Turner et al.,2003a). The undetectable levels of ammonium foundhere respond to a common pattern in most calcareousMediterranean rivers (Prat, 2000, 2002). The main lim-itation for primary production (chlorophyll a) seems tobe the low light intensity due to the river’s locationin the gully of a forested area and the low levels ofphosphorus, as is indicated by correlations among en-vironmental variables.
Microcystin production seems to be a year-roundphenomenon in the stream studied. In arid zones,basin substrata may liberate nutrients, specially nitrate(Holloway et al., 1998), which could explain the rela-tively high nitrate load detected in the Alharabe River.Other sources of nitrogen may arise from the leach-ing of nearby pastureland (Turner et al., 2003a) or dif-fuse points (Turner et al., 2003b). External sources of
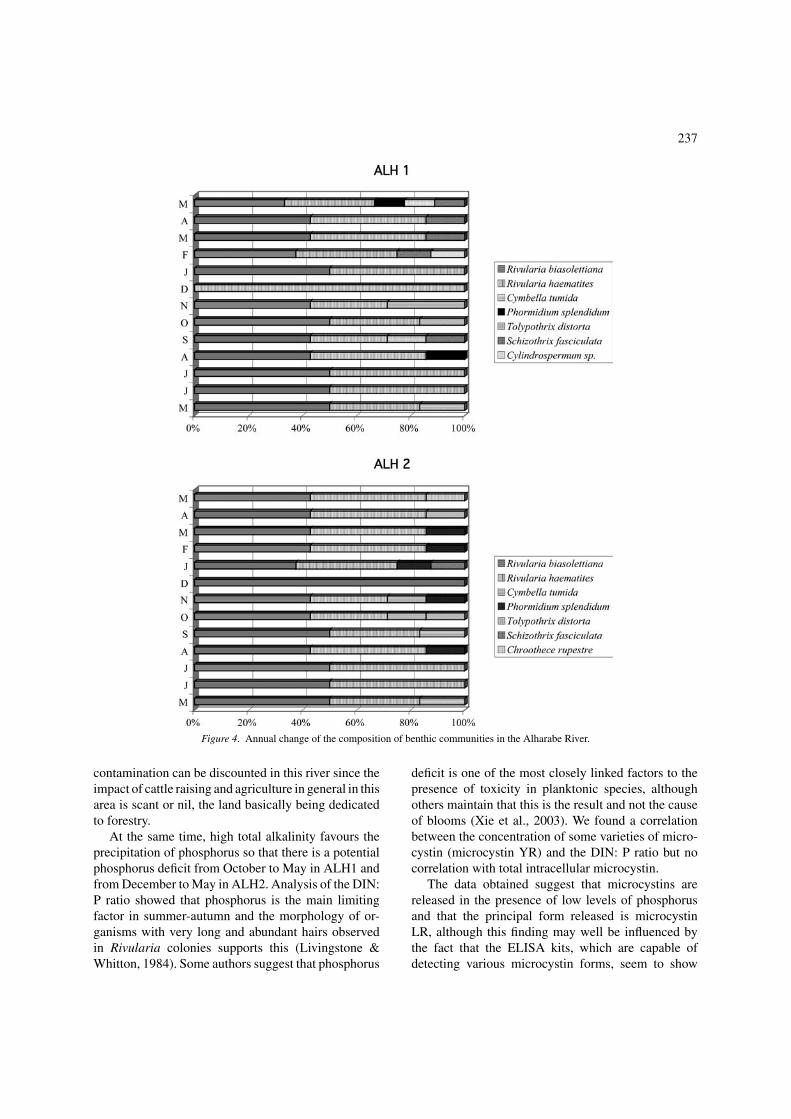
237
Figure 4. Annual change of the composition of benthic communities in the Alharabe River.
contamination can be discounted in this river since theimpact of cattle raising and agriculture in general in thisarea is scant or nil, the land basically being dedicatedto forestry.
At the same time, high total alkalinity favours theprecipitation of phosphorus so that there is a potentialphosphorus deficit from October to May in ALH1 andfrom December to May in ALH2. Analysis of the DIN:P ratio showed that phosphorus is the main limitingfactor in summer-autumn and the morphology of or-ganisms with very long and abundant hairs observedin Rivularia colonies supports this (Livingstone &Whitton, 1984). Some authors suggest that phosphorus
deficit is one of the most closely linked factors to thepresence of toxicity in planktonic species, althoughothers maintain that this is the result and not the causeof blooms (Xie et al., 2003). We found a correlationbetween the concentration of some varieties of micro-cystin (microcystin YR) and the DIN: P ratio but nocorrelation with total intracellular microcystin.
The data obtained suggest that microcystins arereleased in the presence of low levels of phosphorusand that the principal form released is microcystinLR, although this finding may well be influenced bythe fact that the ELISA kits, which are capable ofdetecting various microcystin forms, seem to show

238
Figure 5. Annual change of the concentration of total intracellular microcystins of benthic communities in the Alharabe River.
Figure 6. Mean concentrations of the different forms of intracellular microcystins in the total benthic community in the Alharabe River duringthe year studied.
greater affinity for microcystin LR. Dissolved micro-cystins show an inverse correlation with chlorophylla, which may be considered as an indicator of toxinrelease during cell senescence. The growth of mostcolonies usually implies the senescence of lower partsand apical growth in an almost continuous process(Pentecost, 1991; Riding, 1991).
The concentration of intracellular microcystins waspositively correlated with temperature and silicate andnegatively correlated with nitrate, nitrite, calcium andflow, all variables linked to geologic and climatic fac-tors. The dissolved inorganic nitrogen concentrationwas directly related with flow, which could be ex-plained by the origin of nutrients from geological mate-rial as in other arid regions (Holloway et al.,1998) and
not from anthropogenic activities. A relationship be-tween the presence of different forms of nitrogen andthe concentration of microcystins has generally beenaccepted (Sivonen, 1990; Yin et al., 1997). In the Al-harabe River a close relationship between temperatureand total intracellular microcystin concentration wasobserved. Cyanobacteria culture studies suggest thatmicrocystin concentrations reach a maximum at around25 ◦C in the laboratory and decreases sharply above thistemperature (Sivonen, 1990; Yin et al., 1997). The pre-vailing weather conditions of this region are thereforehighly suitable for synthesis, which explains the valuesrecorded. The positive correlation with silicate, alsofound in south-east Spain reservoirs (Aboal & Puig,2005), may imply a relation with a decline in diatoms. A

239
Figure 7. Relationship between total intracellular microcystin and maximum air temperature during the year of the study.
similar relationship was found by other authors (Repkaet al., 2004).
The different varieties of microcystins seem torespond differently to environmental variables, ex-plaining the changing relative proportions observedthroughout the year: microcystin YR is linked to lowlevels of ortophosphate and total alkalinity but micro-cystin LR and microcystin RR are more closely relatedwith low levels in nitrogen and calcium. If not all micro-cystins are produced in the same conditions they maywell have different roles in cyanophyte persistence, butthis fact needs to be more deeply studied.
These findings emphasise the importance of nutri-ents, air temperature and flow in regulating the concen-tration of intracellular microcystins in some calcareousstreams. All are key factors when taking into considera-tion the possible effects of global climatic warming andillustrate the lack of dependence, at least in some cases,of microcystin synthesis on external nitrogen inputs.
The general belief about the lack of palatabilityof cyanophytes is probably related with their capac-ity to produce toxic substances, which not only affectsmacroinvertebrate communities, accumulating in the
food chain (Chen et al., 2005; Malbrouck et al., 2004;Pinho et al., 2003), but also the development of otherspecies of algae (Kearns & Hunter, 2000) and may con-tribute to their predominance in slowly flowing, shal-low calcareous streams, as was pointed out by someauthors (Whitton et al., 2005).
In calcareous streams with low levels of ambientphosphorus likely to lead to phosphorus deficiency inalgae, the presence of mucilaginous colonies of produc-ing high quantities of exopolymers and/or long hairsis relatively common. Both morphological character-istics are linked to phosphatase activity, which maycontribute to the prevalence of some taxa in these con-ditions (Whitton et al., 2005). The production of ex-opolymers is greatly enhanced in diatoms in condi-tions of phosphorus deficiency (Leandro et al., 2003;Margaletti et al., 2004; Staats et al., 1999). The sameconditions seem to promote the release of microcystins.
The data presented point to the important role playedby mucilage in the retention and accumulation oftoxic substances. Algae which produce mucilages canalso benefit from the capacity of cyanophytes to avoidgrazing by macroinvertebrates. The accumulation of

240
Table 3. Correlation matrix of physical, chemical, climatic and biological variables with main varieties of microcystins detectedin the Alharabe River.
Microcystin RR Microcystin YR Microcystin LR Total microcystin Dissolved microcystin
Silicate 0.35 0.40 0.42 0.59 −0.14PO4-P 0.01 −0.54 0.09 −0.19 −0.35TP −0.01 0.12 0.20 0.11 −0.59NO2-N −0.50 −0.02 −0.71 −0.67 0.17NO3-N −0.58 −0.04 −0.86 −0.83 0.22Total alkalinity 0.38 −0.53 0.34 0.23 0.35Calcium −0.45 −0.10 −0.66 −0.72 −0.00Flow −0.35 0.37 −0.76 −0.56 0.08Rainfall −0.10 0.33 −0.49 −0.17 0.41Max. air temperature 0.56 0.18 0.70 0.77 −0.19Min. air temperature 0.53 0.20 0.67 0.76 −0.12DIN: P 0.15 0.54 −0.06 0.28 0.33PAR 0.02 0.05 0.19 0.13 −0.45Microcystin RR 0.05 0.66 0.84 −0.15Microcystin YR 0.05 −0.18 0.23 0.23Microcystin LR 0.66 −0.18 0.86 −0.47Total microcystin 0.84 0.23 0.86 −0.20Dissolved microcystin −0.15 0.23 −0.47 −0.20Chlorophyll a 0.30 −0.03 0.17 0.18 −0.57Carotenoids 0.44 0.08 0.04 0.16 −0.17Allophycocyanin 0.62 −0.03 0.29 0.38 −0.25Phycoerythrin 0.61 −0.02 0.29 0.38 −0.25
Correlations significant p = 0.05 shown in bold font.
Figure 8. Changes in total intracellular microcystin concentration of benthic cyanophytes and silicate in the Alharabe River between May 2001and May 2002.

241
Table 4. Maximum, minimum and mean concentrations of the different variants of microcystins identified in the macroscopiccolonies of benthic cyanophytes in the Alharabe River.
Microcystin 1 Microcystin RR Microcystin YR Microcystin LR Microcystin 2Species (µg g−1) (µg g−1) (µg g−1) (µg g−1) (µg g−1)
Rivularia biasolettiana
Min–max 0.2–2.0 0.3–3.8 0.1–1.8 0.4–8.4 0.1–0.5
Mean ( n = 15) 0.6 0.8 0.6 1.7 0.2
Rivularia haematites
Min–max 0.3–1.4 0.5–2.6 0.2–2.7 0.3–10.3 0.0–0.4
Mean ( n = 14) 0.5 1.1 0.7 2.5 0.2
Phormidium splendidum
Min–max 0.4–1.1 0.4–1.7 0.1–1.4 0.6–1.3 0.3–2.2
Mean ( n = 2) 0.8 1.1 0.5 1.0 1.3
Tolypothrix distorta ( n = 1) 0.5 0.7 2.2 0.6 0.7
microcystins in mucilaginous colonies could explainthe low palatability and the seasonal dominance ofmacroscopic Cymbella colonies in some calcareousstreams. The process whereby toxins accumulated andwhether they have any kind of antigrazing activity needto be more deeply studied but could explain the durablepresence of some of these colonies in calcareousrivers.
Acknowledgments
This Project was funded by Spanish Ministry of Scienceand Technology (REN2000-1021-C03-02/HID).
References
Aboal M (1989) Flora algal del rıo Benamor (Cuenca del Segura, SEde Espana). Limnetica 5: 1–11.
Aboal M, Alcaraz FJ, Barrena J, Egidos AI (2003) El componentevegetal en los humedales de la Region de Murcia. Catalogacion,evaluacion de la rareza y propuesta de medidas para su con-servacion. Comunidad Autonoma de Murcia. Novograf, SA,Murcia, pp. 115.
Aboal M, Puig MA (2005) Intracellular and dissolved microcystinsin reservoirs of the river Segura basin, Murcia, SE Spain. Toxicon45 (4): 509–518.
Aboal M, Puig MA, Mateo P, Perona E (2002) Implications ofcyanophyte toxicity on biological monitoring of calcareousstreams in north–east Spain. J. appl. Phycol. 14: 49–56.
Aboal M, Puig MA, Rıos H, Lopez-Jimenez E (2000) Relationshipbetween macroinvertebrate diversity and cyanobacteria toxicityin streams. Verh. int. Verein. Limnol. 27: 555–559.
APHA (1998) Standard Methods for the Examination of Water andWastewater. 19th Edition. American Public Health Association,American Water Works Association, Water Pollution ControlFederation, Washington, DC, p. 769.
Baker PD, Steffensen DA, Humpage AR, Nicholson BC, FalconerIR, Lanthois B, Fergusson KM, Saint CP (2001) Preliminaryevidence of toxicity associated with the benthic cyanobacteriumPhormidium in south Australia. Environ. Toxicol. 16(6): 506–511.
Bennett A, Bogorad L (1973) Complementary chromatic adaptationin a filamentous blue-green alga. J. Cell Biol. 58: 419–435.
Bolch CJ, Orr PT, Jones GJ, Blackburn SI (1999) Genetic, mor-phological and toxicological variations among globally dis-tributed strains of Nodularia (Cyanobacteria). J. Phycol. 35: 339–355.
Chapman PJ, Edwards AC, Cresser MS (2001) The nitrogen compo-sition of streams in upland Scotland: Some regional and seasonaldifferences. Sci. Total Environ. 265: 65–83.
Chen J, Xie P, Guo L, Zheng K, Ni L (2005) Tissue distributions andseasonal dynamics of the hepatotoxic microcystins-LR and -RRin a freshwater snail (Ballamya aeruginosa) from a large shallow,eutrophic lake of the subtropical China. Environ. Pollution 134:423–430.
Chorus J, Bartram J (eds) (1999) Toxic Cyanobacteria in Water.A Guide to their Public Health Consequences, Monitoringand Management. World Health Organization, E & F N Spon,p. 416.
Codd GA, Ward CJ, Beattie KA, Bell SG (1999) Widening per-ceptions of the occurrence and significance of cyanobacterialtoxins. In Peschek GA, Loffelhardt W, Schmetterer G (eds).The Phototrophic Prokaryotes. Kluwer Academic/PlenumPublishers, Dordrecht, The Netherlands, p. 836.
Dow CS, Swoboda UK (2000) Cyanotoxins. In Whitton BA, PottsM (eds). The Ecology of Cyanobacteria. Their Diversity inTime and Space. Kluwer Academic Publishers, Dordrecht, TheNetherlands, pp. 613–632.
Dyble J, Paerl HW, Neilan BA (2002) Genetic characterizationof Cylindrospermopsis raciborskii (Cyanobacteria) isolatesfrom diverse geographic origins based on nifH and cpcBA-IGSnucleotide sequence analysis. Appl. Environ. Microbiol. 68(5):2567–2571.
Edwards C, Beattie KA, Scrimgeour CM, Codd GA (1992) Iden-tification of Anatoxin-a in benthic cyanobacteria (Blue-GreenAlgae) and in associated dog poisonings at Loch Insh, Scotland.Toxicon 30: 1165–1175.

242
Falconer IR (1999) An overview of problems caused by toxicblue-green algae (Cyanobacteria) in drinking and recreationalwater. Environ. Toxicol. 14: 5–12.
Figueiredo DR, Azeitiero M, Esteves SM, Goncalves FJM, Pereira,MJ (2004) Microcystin-producing blooms – a serious globalpublic health issue. Ecotoxicol. Environ. Safety 59: 151–163.
Gjolme N, Utkilen H (1996) The extraction and the stability ofmicrocystin-RR in different solvents. Phycologia 35(6): 80–82.
Gordon ND, McMahon TA, Finlayson BL (2004) Stream Hydrology:An introduction for ecologists. Wiley, NY, Ed. p. 444.
Gromov BV, Vepritskiy AA, Titova NN, Mamkayeva KA,Alexandrova OV (1991) Production of the antibiotic cyanobac-teria Lu-1 by Nostoc linckia CALU-892 (Cyanobacteria). J.Appl. Phycol. 3(1): 55–59.
Henning M, Rohrlack T, Kohl, JG (2001) Responses of Daphniagaleata fed with Microcystis strains with rat hepatocyte andcell-line assays. In Chorus I (ed.), Cyanotoxins – Occurrence,Causes, Consequences. Springer, Berlin, pp. 317–324.
Holloway JM, Dahlgren RA, Hansen B, Casey WH (1998) Con-tribution of bedrock nitrogen to high nitrate concentrations instream water. Nature 395: 785–788.
Jang MH, Ha K, Lucas MC, Joo GJ, Takamuira N (2004) Changesin microcystins production by Microcystis aeruginosa exposedto phytoplanktivorous and omnivorous fish. Aquatic Toxicology68: 51–55.
Jeffrey SW, Humphrey GF (1975). New spectrophotometric equa-tions for determining chlorophylls a, b, c1 and c2 in higherplants, algae and natural phytoplankton. Biochem. Physiol.Pflanzen 167: 191–194.
Kearns KD, Hunter MD (2000) Green algal extracellular productsregulate antialgal toxin production in a cyanobacterium. Environ.Microbiol. 2: 291–297.
Kurmayer R, Juttner F (1999) Strategies for the co-existenceof zooplankton with the toxic cyanobacterium Planktothrixrubescens in Lake Zurich. J. Plankton Res. 21: 659–683.
Leandro SM, Gil MC, Delgadillo I (2003) Partial characterisation ofexopolysaccharides exudated by planktonic diatoms maintainedin batch cultures. Acta Oecologica 24: 49–55.
Livingstone D, Whitton BA (1984) Water chemistry and phosphataseactivity of the blue-green alga Rivularia in Upper Teesdalestreams. J. Ecol. 72: 405–421.
Margalef R (1983) Limnologıa. Omega, Barcelona, p. 1010.Margaletti E, Urbani R, Sist P, Ferrari CR, Cicero AM (2004)
Abundance and chemical characterization of extracellu-lar carbohydrates released by the marine Cylindrothecafusiformis under N- and P- limitation. Eur. J. Phycol. 39: 133–142.
Marsalek B, Dolejs P, Sladeckova A (1997) Algal toxins in Czechdrinking water resources and treatment plants. Water Supply 18(Special Subject): 6–9.
Meriluoto JAO, Ermsson JE, Harada KI, Dahlem AM, SivonenK, Carmichael WW (1990). Internal surface reverse phasehigh-performance liquid chromatographic separation of thecyanobacterial peptide toxins microcystin-LR, -YR, -RR andnodularin. J. Chromatography 509: 390–395.
Malbrouck C, Trausch G, Deros P, Kestemont P (2004) Effect ofmicrocystin-LR on protein phosphatase activity in fed and fastedjuvenile goldfish Carassius auratus L. Toxicon 43: 295–301.
Mez K, Hanselmann K, Preisig HR (1998) Environmental conditionsin high mountain lakes containing toxic benthic cyanobacteria.Hydrobiologia 368: 1–15.
Murphy J, Riley JP (1962) A modified single solution method fordetermination of phosphate in natural waters. Anal. Chim. Acta27: 31–36.
Owen RP (1994) Biological and economic significance of benthiccyanobacteria in two Scottish Highland Lochs. In Codd GA,Jefferizs TM, Keevil CW, Potter E (eds). Detection Methodsfor Cyanobacterial Toxins. The Royal Society of Chemistry,Cambridge, pp.145–148.
Pentecost A (1991) Calcification processes in algae and cyanobac-terium. In Riding R (ed.). Calcareous Algae and Stromatolites.Springer-Verlag, Berlin-Heidelberg, pp. 3–20.
Pinho GLL, Mourada da Rosa C, Yunes JS, Luquet CM, BlanchiniA, Monserrat JM (2003) Toxic effects of microcystin in thehepatopancreas of the estuarine crab Chasmagnathus granulatus(Decapoda, Grapsidae). Comp. Biochem. Physiol. 135: 459–468.
Prat N (2000) La qualitat ecologica del Llobregat, el Besos, el Foixi la Tordera. Estudis de la qualitat ecologica dels rius. Vol 10.Ed. Diputacio de Barcelona, p. 163.
Prat N (2002) La qualitat ecologica del Llobregat, el Besos, el Foixi la Tordera. Estudis de la qualitat ecologica dels rius. Vol 12.Ed. Diputacio de Barcelona. Electronic version.
Puig MA, Aboal M, Mateo P, Perona E (2000) Interactions betweencyanobacteria and herbivore caddisflies in Spanish Mediter-ranean streams. Abstracts 10th International Symposium onTrichoptera, Postdam, Germany, 45.
Repka S, Meyerhofer M, von Brockel K, Sivonen K (2004) Asso-ciation of cyanobacterial toxin nodularin with environmentalfactors and zooplancton in the Baltic Sea. Microbial Ecology47: 350–358.
Riding R (1991) Calcified cyanobacteria. In Riding R (ed.).Calcareous Algae and Stromatolites. Springer-Verlag, Berlin,Heidelberg, pp. 55–87.
Rıos H (2000) Interacciones entre comunidades de cianobac-terias epilıticas fluviales y poblaciones de efemeropterosherbıvoros y plecopteros: Efectos de la toxicidad de lascianobacterias. Tesina de Licenciatura, Universidad de Girona,p. 78.
Sawyer R, Grisley LM (1967) The determination of ammonia in theppb level solution. In Technicon Symposia. Vol 1, Automationin analytical chemical.
Schagerl M, Unterrrieder I, Angeler, DG (2002) Allelopathyamong cyanoprokaryotes and other algae originating fromlake Neusiedlersee (Austria) Int. Rev. Hydrol. 87(4): 365–374.
Sivonen K (1990) Effects of light, temperature, nitrate, orthophos-phate, and bacteria on growth of an hepatotoxin production byOscillatoria agardhii strains. Appl. environ. Microbiol. 56 (9):2658–2666.
Staats N, Winder B, de Stal LJ, Mur LR (1999) Isolation and char-acterization of extracellular polysaccharides from the epipelicdiatoms Cylindrotheca closterium and Navicula salinarum. Eur.J. Phycol. 34: 161–169.
Turner BL, Baxter R, Whitton BA (2003a). Nitrogen and phospho-rus in soil solutions and drainage streams in Upper Teesdale,northern England: Implications of organic compounds forbiological nutrient limitation. Sci. Total Environ. 314–316: 153–170.
Turner BL, Chudek JA, Whitton BA, Baxter R (2003b) Phosphoruscomposition of upland soils polluted by long-term atmosphericnitrogen deposition. Biogeochemistry 65: 259–274.

243
Valdor R (2003) Toxicidad de extractos de cianofıceas sobre algas ybacterias en condiciones de laboratorio. Tesina de Licenciatura.Facultad de Biologıa. Universidad de Murcia, p. 70.
Whitton BA, Al-Shehri AM, Ellwood NTW, Turner BL (2005)Ecological aspects of phosphatase activity in cyanobacteria,eukaryotic algae and bryophytes. In Turner BL, Frossard E,Balwin DS (eds). Organic Phosphorus in the Environment.Commonwealth Agricultural Bureau International, Wallingford,UK, pp. 205–241.
Xie L, Xie P, Li S, Tang H, Liu H (2003) The low TN/TP ratio, a causeor a result of Microcystis blooms?. Water Res. 37: 2073–2080.
Yin Q, Carmichael WW, Evans WR (1997) Factors influencinggrowth and toxin production by cultures of the freshwatercyanobacterium Lyngbya wollei Farlow ex Gomont. J. Appl.Phycol. 9: 55–63.
Zehnder A (1985) Isolation and cultivation of large cyanophytes fortaxonomic purposes. Arch. Hydrobiol. Suppl. 71 (AlgologicalStudies 38/39): 281–289.




















