
Enantioselectivity of polychlorinated biphenyl atropisomers in sediment and biota from the...
8
Enantioselectivity of polychlorinated biphenyl atropisomers in sediment and biota from the Turtle/Brunswick River estuary, Georgia, USA Matthew S. Ross a , Erin L. Pulster b , Malgorzata B. Ejsmont a , Elaine A. Chow a , Colin M. Hessel a , Keith A. Maruya c , Charles S. Wong a,d,⇑ a Department of Chemistry, University of Alberta, Edmonton, AB, Canada T6G 2G2 b Mote Marine Laboratory Center for Ecotoxicology, Sarasota, FL 34236, USA c Southern California Coastal Water Research Project, Costa Mesa, CA 92626, USA d Environmental Studies Program and Department of Chemistry, Richardson College for the Environment, University of Winnipeg, Winnipeg, MB, Canada R3B 2E9 article info Keywords: Aroclor 1268 Chiral PCB Bottlenose dolphin Microbial dechlorination abstract To investigate the potential for enantioselective transformation and accumulation, the enantiomer distributions of seven polychlorinated biphenyl (PCB) atropisomers were measured in the sediment and biota from a sub-tropical estuary heavily contaminated with Aroclor 1268, a technical mixture of highly chlorinated PCB congeners. Enantiomer fractions (EFs) of PCBs 91, 95, 136, 149, 174, 176, and 183 in marsh sediment, invertebrate, forage and predatory fish species, and bottlenose dolphins were determined. Non-racemic EFs greater than 0.75 were found in sediments for PCBs 136 and 174, likely the result of microbial dechlorination. Although enantiomer fractions in grass shrimp (Palaemonetes spp.) mirrored those of sediment, fish species had EFs that differed significantly from sediment or grass shrimp. Similarly, bottlenose dolphins were also found to contain non-racemic quantities of PCBs 91, 136, 174, 176, and 183. Non-racemic EFs in these biota were likely a result of both uptake of non-racemic pro- portions of PCBs from the diet and enantioselective biotransformation. Ó 2011 Elsevier Ltd. All rights reserved. 1. Introduction Due to their persistence within the environment, polychlori- nated biphenyls (PCBs) continue to pose risks to humans and biota more than 30 years after their use was discontinued. High concen- trations of PCBs are still found in some environmental compart- ments. In aquatic environments, their hydrophobicity causes PCBs to partition into sediments, where they may be may be taken up by lower trophic level organisms and enter the food web. Microbially-mediated reductive dehalogenation is one possible transformation process for highly chlorinated PCBs in anaerobic sediments. Sediment microbes remove chlorines from more highly chlorinated congeners, resulting in an enrichment of lower chlori- nated congeners in the environment (Bedard, 2001). The presence of in situ dechlorination activity can be inferred by analyzing con- gener and homolog patterns in sediments and comparing those to congener and homolog patterns found in the original Aroclor mix- tures (Bedard and May, 1996; Brown et al., 1987; Magar et al., 2005). While such techniques provide strong evidence for micro- bial activity, they do no necessarily prove that it exists, as changes in congener patterns may also result from vaporization or selective sorption. Furthermore, dechlorination rates may also be suffi- ciently slow to make it difficult to detect changes in the congener patterns over short durations of time. The use of enantioselective analytical techniques and the detec- tion of non-racemic proportions of chiral molecules can provide unequivocal evidence for biological activity. Nineteen of the 209 PCB congeners are chiral and exist in the environment as stable atropisomers (Kaiser, 1974). These congeners were released into the environment as racemic mixtures, and although individual enantiomers possess identical physical and chemical properties, non-racemic distributions of PCB enantiomers have been found in sediment and biota samples (Wong et al., 2001a,b). Biological processing may change the enantiomer distribution of a compound within the environment due to differential interaction between enantiomers with other chiral molecules, such as proteins. Previ- ous studies have found that dechlorination of PCBs by sediment microbes and in vivo biotransformation by biota can occur in an enantioselective manner (Buckman et al., 2006; Pakdeesusuk et al., 2003; Singer et al., 2002; Warner and Wong, 2006; Wong et al., 2002b). Therefore, the use of stereoisomer analysis and the detection of non-racemic enantiomer distributions provides a sensitive means of detecting and tracing biologically-mediated 0025-326X/$ - see front matter Ó 2011 Elsevier Ltd. All rights reserved. doi:10.1016/j.marpolbul.2011.02.001 ⇑ Corresponding author at: Environmental Studies Program and Department of Chemistry, Richardson College for the Environment, University of Winnipeg, Winnipeg, MB, Canada R3B 2E9. Tel.: +1 204 786 9335; fax: +1 204 775 2114. E-mail address: [email protected] (C.S. Wong). Marine Pollution Bulletin 63 (2011) 548–555 Contents lists available at ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Enantioselectivity of polychlorinated biphenyl atropisomers in sediment and biota from the...

Marine Pollution Bulletin 63 (2011) 548–555
Contents lists available at ScienceDirect
Marine Pollution Bulletin
journal homepage: www.elsevier .com/locate /marpolbul
Enantioselectivity of polychlorinated biphenyl atropisomers in sediment and biotafrom the Turtle/Brunswick River estuary, Georgia, USA
Matthew S. Ross a, Erin L. Pulster b, Malgorzata B. Ejsmont a, Elaine A. Chow a, Colin M. Hessel a,Keith A. Maruya c, Charles S. Wong a,d,⇑a Department of Chemistry, University of Alberta, Edmonton, AB, Canada T6G 2G2b Mote Marine Laboratory Center for Ecotoxicology, Sarasota, FL 34236, USAc Southern California Coastal Water Research Project, Costa Mesa, CA 92626, USAd Environmental Studies Program and Department of Chemistry, Richardson College for the Environment, University of Winnipeg, Winnipeg, MB, Canada R3B 2E9
a r t i c l e i n f o
Keywords:Aroclor 1268Chiral PCBBottlenose dolphinMicrobial dechlorination
0025-326X/$ - see front matter � 2011 Elsevier Ltd.doi:10.1016/j.marpolbul.2011.02.001
⇑ Corresponding author at: Environmental StudiesChemistry, Richardson College for the EnvironmeWinnipeg, MB, Canada R3B 2E9. Tel.: +1 204 786 933
E-mail address: [email protected] (
a b s t r a c t
To investigate the potential for enantioselective transformation and accumulation, the enantiomerdistributions of seven polychlorinated biphenyl (PCB) atropisomers were measured in the sedimentand biota from a sub-tropical estuary heavily contaminated with Aroclor 1268, a technical mixture ofhighly chlorinated PCB congeners. Enantiomer fractions (EFs) of PCBs 91, 95, 136, 149, 174, 176, and183 in marsh sediment, invertebrate, forage and predatory fish species, and bottlenose dolphins weredetermined. Non-racemic EFs greater than 0.75 were found in sediments for PCBs 136 and 174, likelythe result of microbial dechlorination. Although enantiomer fractions in grass shrimp (Palaemonetesspp.) mirrored those of sediment, fish species had EFs that differed significantly from sediment or grassshrimp. Similarly, bottlenose dolphins were also found to contain non-racemic quantities of PCBs 91, 136,174, 176, and 183. Non-racemic EFs in these biota were likely a result of both uptake of non-racemic pro-portions of PCBs from the diet and enantioselective biotransformation.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Due to their persistence within the environment, polychlori-nated biphenyls (PCBs) continue to pose risks to humans and biotamore than 30 years after their use was discontinued. High concen-trations of PCBs are still found in some environmental compart-ments. In aquatic environments, their hydrophobicity causesPCBs to partition into sediments, where they may be may be takenup by lower trophic level organisms and enter the food web.
Microbially-mediated reductive dehalogenation is one possibletransformation process for highly chlorinated PCBs in anaerobicsediments. Sediment microbes remove chlorines from more highlychlorinated congeners, resulting in an enrichment of lower chlori-nated congeners in the environment (Bedard, 2001). The presenceof in situ dechlorination activity can be inferred by analyzing con-gener and homolog patterns in sediments and comparing those tocongener and homolog patterns found in the original Aroclor mix-tures (Bedard and May, 1996; Brown et al., 1987; Magar et al.,2005). While such techniques provide strong evidence for micro-
All rights reserved.
Program and Department ofnt, University of Winnipeg,5; fax: +1 204 775 2114.C.S. Wong).
bial activity, they do no necessarily prove that it exists, as changesin congener patterns may also result from vaporization or selectivesorption. Furthermore, dechlorination rates may also be suffi-ciently slow to make it difficult to detect changes in the congenerpatterns over short durations of time.
The use of enantioselective analytical techniques and the detec-tion of non-racemic proportions of chiral molecules can provideunequivocal evidence for biological activity. Nineteen of the 209PCB congeners are chiral and exist in the environment as stableatropisomers (Kaiser, 1974). These congeners were released intothe environment as racemic mixtures, and although individualenantiomers possess identical physical and chemical properties,non-racemic distributions of PCB enantiomers have been foundin sediment and biota samples (Wong et al., 2001a,b). Biologicalprocessing may change the enantiomer distribution of a compoundwithin the environment due to differential interaction betweenenantiomers with other chiral molecules, such as proteins. Previ-ous studies have found that dechlorination of PCBs by sedimentmicrobes and in vivo biotransformation by biota can occur in anenantioselective manner (Buckman et al., 2006; Pakdeesusuket al., 2003; Singer et al., 2002; Warner and Wong, 2006; Wonget al., 2002b). Therefore, the use of stereoisomer analysis and thedetection of non-racemic enantiomer distributions provides asensitive means of detecting and tracing biologically-mediated

Fig. 1. Map of the study area.
M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555 549
processes, for instance the presence of microbial dehalogenationactivity, that may otherwise be difficult to detect using achiralmethods (Wong et al., 2004, 2007).
The LCP Chemicals Superfund site, a marshfront industrial sitelocated on the margins of the Turtle/Brunswick estuary in south-eastern Georgia, USA (Fig. 1), was home to an oil refinery, a powerplant, and a paint manufacturing plant over the past 70 years. Mostrecently, a chlor-alkali plant operated on the site from 1955 to1994. During this time, industrial wastes were discharged directlyinto the surrounding marsh areas known as Purvis Creek. This re-sulted in the sediment from the marsh and creek areas surround-ing the LCP site being highly contaminated with mercury andAroclor 1268, an uncommon PCB formulation comprised primarilyof octa- to decachlorinated biphenyls (Table 1; Kannan et al.,1997). Previous studies on the site have found that the highly chlo-rinated PCBs are bioavailable to organisms, which has also resultedin widespread contamination of local biota (Kannan et al., 1998;Maruya and Lee, 1998a). However, due to the recalcitrance of thesecongeners, these studies were not designed to investigate whetherbiotransformation was occurring, either in sediments or biota.
In this study, we analyzed the enantiomer distribution of sevenchiral PCB congeners in sediment cores from the contaminatedestuary around the LCP Superfund site to determine whether ornot microbial dechlorination was occurring at this site. Addition-ally, the high concentrations of heptachlorinated biphenyls foundin biota at this site provide a unique opportunity to investigate
Table 1Polychlorinated biphenyl relative homolog distributions (%) in Aroclor 1268 andmarsh sediments from the LCP Superfund site, Brunswick, GA.
Homolog Aroclor 1268a Marsh sedimenta,b
Di- 0 0Tri- 0.006 0.16Tetra- 0.12 0.30Penta- 0.19 1.57Hexa- 4.38 7.2Hepta- 10.1 14.6Octa- 45 33.1Nona- 35 37.9Deca- 4.8 5.2
a Data from Kannan et al. (1997).b Average of left and right south marsh transect.
the enantioselective behavior of highly chlorinated PCB homologs.Therefore, enantiomer distributions were determined in inverte-brates, fish species, and resident dolphins to determine the extentof biotransformation occurring within these species. This studyprovides insight into the biological processing occurring at theLCP Superfund site, and allows for a further understanding of theenvironmental behavior of highly chlorinated PCBs.
2. Materials and methods
2.1. Sample collection
Three sediment core samples were collected using a polyvinyl-chloride push core (7.6 cm diameter � 61 cm length) from twoareas of the intertidal marsh (Fig. 1). Intact cores were sealed withend caps lined with solvent-rinsed aluminum foil and were sec-tioned using solvent-rinsed stainless implements upon return tothe lab. All sediment samples were stored frozen at �20 �C inpre-cleaned glass (I-Chem) jars prior to analysis. The south marsharea (cores NSM and LCP) is located adjacent to waste pits on theLCP Superfund site, and is characterized by high concentrationsof PCBs and mercury (Kannan et al., 1997; Winger et al., 1993).A causeway bisects the marsh area, and sediments collected fromthe north side of the causeway (core 311A) contain significantlylower concentrations of PCBs.
Biota was collected from various locations around the LCP sitein an effort to represent the basic trophic levels of the local estua-rine food web. Grass shrimp (Palaemonetes pugio), an abundantcrustacean macroinvertebrate in estuaries of this region, were col-lected from Purvis Creek by dip netting (Maruya and Lee, 1998a).Silver perch (Bairdiella chrysoura), spot (Leiostomus xanthurus),spotted seatrout (Cynoscion nebulosus), and striped mullet (Mugilcephalus) were collected from Purvis Creek by trawl, gill and castnetting (Pulster et al., 2005). These fish species were collected torepresent likely prey items of the bottlenose dolphins (Tursiopstruncatus), the predominant marine mammal species that frequentthe region’s estuaries (Pulster and Maruya, 2008). Blubber samplesfrom free ranging bottlenose dolphins were collected from theTurtle/Brunswick estuary by means of a dart biopsy (Pulster andMaruya, 2008; Pulster et al., 2009).

Fig. 2. Enantiomer fractions (EFs) at various depths in sediment cores collectedfrom north marsh area and south marsh areas from the LCP Superfund site inGeorgia. Horizontal dashed lines represent the racemic EF value of 0.5 and the 95%confidence interval of racemic standards (±0.032).
550 M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555
2.2. Sample analysis
The procedures for the extraction and achiral analysis of sedi-ment and biota samples have been described in detail elsewhere(Maruya and Lee, 1998a,b; Pulster and Maruya, 2008; Pulsteret al., 2005). Briefly, sediment samples were extracted by Soxhletextraction in CH2Cl2 and cleaned up by activated silica columnchromatography, followed by sulfur removal by activated copper.Whole-body composite grass shrimp samples were extracted sim-ilarly, but cleaned up by Florisil column chromatography. Compos-ite fish samples (�5 g) and dolphin blubber samples (0.5–1.0 g)were extracted in triplicate with CH2Cl2 using a Dionex 200 ASEsystem (Salt Lake City, UT). Lipids were removed by gel permeationchromatography, and the fraction containing the PCBs was subse-quently cleaned up on a glass column packed with 1% water-deactivated Florisil (Fisher Scientific, Fair Lawn, NJ, USA).
Total PCB analysis of was carried out by GC coupled to an elec-tron capture detector. A DB-1 (30 m � 0.25 mm � 0.25 lm df, J&WScientific) or DB-XLB (30 m � 0.25 mm � 0.25 lm df, J&W Scien-tific) fused silica column was used for separation of individualPCB congeners.
Stereoisomer analysis was performed on an Agilent 5890GC–MS using electron impact ionization (EI) and selected ionmonitoring (SIM). Separation of enantiomers was achieved usingan array of chiral columns and previously published conditions(Wong and Garrison, 2000; Wong et al., 2002a). A Chirasil-Dex col-umn (25 m � 0.25 mm i.d. � 0.25 lm df, Varian, Walnut Creek, CA,USA) was used for the separation of PCBs 91, 95, 136, 149, 174 and176, and results were confirmed on a Cyclosil-B column(30 m � 0.25 mm i.d. � 0.25 lm df, J&W Scientific, Folsom, CA,USA). A BGB-172 column (30 m � 0.25 mm i.d. � 0.18 lm df, Anal-ytik, Adiswil, Switzerland) was used for the separation of PCBs 132and 183.
In order to assure that no coelutions with other PCB homologswas occurring, all columns were calibrated with standard solu-tions containing all 209 PCB congeners (Wong and Garrison,2000). In addition, results of the enantiomer separations of PCBs91, 95, 136, 149 and 176 were compared between the Chirasil-Dex and Cyclosil-B columns. Data points with enantiomer frac-tions (see below) that were not in agreement between the twocolumns within ±0.032, based on the 95% confidence interval ofrepeated measurements of racemic standards on these columns(Wong et al., 2004), were assumed to have interferences and wereremoved from the data set. Replicate injections of standardsyielded racemic compositions for all congeners, with EFs rangingfrom 0.495 to 0.505.
2.3. Data analysis
For enantiomer analyses, all peaks were integrated usingmodel-fitting software (PeakFit v.4.0, Systat, San Jose, CA, US) toallow for more accurate quantification of partially coeluting peaks(Asher et al., 2009).
Results are expressed as enantiomer fractions (EFs), which areused to quantify the stereoisomer distribution of a sample (Harneret al., 2000). The EF (Eq. (1)) is defined as the peak area of the firsteluted enantiomer (E1) divided by the sum area of both enantio-mers for those congeners where the elution order of the enantio-mers on a particular column is unknown (PCBs 95 and 183).
EF ¼ E1E1þ E2
or EF ¼ ðþÞðþÞ þ ð�Þ ð1Þ
However, if the elution order is known (all other congeners),then the EF was determined using the peak areas of the (+) and(�) enantiomers.
Comparisons amongst all groups (standards, sediment, and bio-ta) were carried out by one-way analysis of variance (ANOVA) witha = 0.05.
3. Results and discussion
3.1. Sediment cores
All PCB congeners investigated were found in non-racemic pro-portions in at least one of the sediment core samples (Fig. 2). PCBs174 and 136 were non-racemic in all cores and at all depths. PCB149 was generally racemic, but was found to be non-racemic intwo cores (311A, 12 cm; NSM, 15 cm). Likewise, PCB 91 was onlynon-racemic at a single depth (311A, 7 cm). Lower chlorinatedcongeners (<6 chlorines) were found sporadically within the cores,and were detectable at some depths, but not others. Heptachlori-nated congeners, with the exception of PCB 176 in the 311A core,were detectable in all cores.

M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555 551
The measurement of non-racemic EFs in sediment samplesindicates that biotransformation has occurred within the marshsediments. Biotransformation was likely the result of microbially-mediated reductive dechlorination, given the anaerobic conditionsof the sediment at these depths. Previous studies have concludedthat microbial dechlorination activity towards Aroclor 1268 (Pale-kar et al., 2003) and higher PCB homologs in general (Kuipers et al.,1999) is limited. Microcosm studies investigated the dechlorina-tion activity of microbes in sediments from the marsh area andfound significant dechlorination of the primer (2,3,4,5,6-pentachl-orbiphenyl), indicating that microbial dechlorination at this site ispossible. However, during the 1-year incubation period, no mea-surable decrease in the concentration of hexa- to decachlorinatedbiphenyls was found (Palekar et al., 2003). This was attributed tothe high concentrations of heavy metals found at this location orthe reduced bioavailability of the more highly chlorinated congen-ers, rather than a limitation in the dechlorination capability of thesediment microbes (Kannan et al., 1997; Palekar et al., 2003). Theformation of non-racemic EFs in the sediment confirms that evencongeners with more than six chlorines were bioavailable to mi-crobes, and that microbial dechlorination had occurred at this site.
For all cores, there was little change in EF with depth, and nocorrelation between total PCB concentrations and EF. Furthermore,despite differences in total PCB concentrations, EFs among thelocations were similar, regardless of depth. PCB 136 is an excep-tion, with a spike in enantiomer enrichment in the north marshcore, and a decrease in enrichment in the south marsh core. Wonget al. (2007) found EFs to vary with depth in sediment cores fromLake Hartwell, SC, and in some instances, found reversals in the EFfor some congeners. No reversals in EF were found in this study,possibly indicating a consistency in the microbial communities atdifferent depths. Alternatively, the lack of any variation in EF withdepth in this study may be due to a cessation of microbial dechlo-rination activity in the sediments. The decline or cessation ofreductive dechlorination activity over time has been found in othersediment cores from other locations (Bzdusek et al., 2006; Magaret al., 2005; Pakdeesusuk et al., 2005; Wong et al., 2007), and thelack of reductive dechlorination found in previous studies from thissite may be attributable to this decline in activity.
The non-racemic quantities of PCBs 91, 136, 174, 176 and 183 inthe sediments is indicative that these congeners were either beingdegraded or formed enantioselectively. The preference for mi-crobes to degrade one enantiomer over the other under anaerobicconditions has been previously shown in controlled microcosmstudies (Pakdeesusuk et al., 2003). The direction and magnitudeof enantioselective microbial degradation of PCB congeners occursin a site-specific manner in situ. We found EFs of PCB 91 in the sed-iment cores to be of similar magnitude and direction of enrichmentto that which had previously been seen in the Housatonic Riverand Lake Hartwell, but opposite of that seen in the Hudson River(Wong et al., 2001a). In addition, PCB 95, which was found to beracemic in this study, was extremely enriched in the second elutedenantiomer in microcosm studies and sediment cores from LakeHartwell (Pakdeesusuk et al., 2003; Wong et al., 2007).
Literature data for the enantioselective dechlorination of morehighly chlorinated congeners is sparse. Non-racemic EFs of PCB174 and 176 have been measured in several sites across the USAin a large survey, and in sediment cores from Lake Hartwell, SC(Wong et al., 2001a, 2007). For PCB 174, all of these locations werefound to be slightly enriched in the (+)-enantiomer. Surficialsediment samples from Lake Hartwell, however, contained highlynon-racemic EFs of PCB 174 (EF = 0.788, converted from enantio-mer ratio) in a single site (Wong et al., 2001a), which is moreclosely in line with the EFs observed in this study. Enantiomer frac-tions of PCB 176 are generally enriched in the (�)-enantiomer, theexception being Lake Hartwell sediment, which was slightly en-
riched in the (+)-enantiomer (Wong et al., 2001a). Observationsof PCB 183 in non-racemic proportions are limited, with onlytwo observations occurring in the White River Basin of Indiana,and in Hudson River Sediment Standard Reference Material (SRM1939a) (Morrissey et al., 2007; Wong et al., 2001a). All of the EFsobserved to date for PCB 183 show only a slight enrichment, whichis in stark contrast to the results seen in this study, which foundsediment samples to be highly enriched in the first eluting enantio-mer. Such differences in the magnitude and direction of EF shiftsamong sites has been attributed differences in species and/or activ-ities of the microbial communities found at those sites (Wonget al., 2001a, 2007).
A key difference between this and previous studies is the envi-ronment in which the sediments were collected. Most other stud-ies investigated sites in freshwater environments, which may differsignificantly in the microbial populations from those in saltwateror estuarine environments. In particular, sulfate reducing bacteriaare much more abundant in estuarine and marine sediments com-pared to freshwater environments. Non-racemic EFs of PCBs 95 and149 have been found in the Hudson River estuary, although theirpresence was attributed to the transport of contaminated sedimentcontaining non-racemic EFs from the upper Hudson River (Asheret al., 2007). A lack of enantioselective biodegradation has beenfound in marine sediments for chlordanes and toxaphene congen-ers (Li et al., 2007; Vetter and Maruya, 2000), which may implythat marine microorganisms lack the ability to degrade chlorinatedorganic pollutants enantioselectively. Our results contrast this,with the finding of non-racemic EFs, particularly of more highlychlorinated congeners, in the estuarine sediment from thislocation.
In general, reductive dechlorination pathways favor theremoval of flanked meta and para chlorines first, resulting indecreased abundance of highly chlorinated congeners, and anincrease in the less chlorinated congeners (Bedard, 2001). Basedon Palekar et al. (2003), several reductive dechlorination pathwaysmay be occurring at this site. The dechlorination of 2,3,4,5,6-pentachlorobiphenyl in sediments from this site followed twodechlorination pathways, depending on whether 2,3,4,5,6-pentachlorbiphenyl was initially meta- or para-dechlorinated. Themeta-initiated route appears to proceed through process N, whichpreferentially removes flanked meta-chlorines from penta- tononachlorinated biphenyls. Processes P and H may also be occur-ring at this site, as both processes remove flanked para chlorinesfrom tetra- to heptachlorinated biphenyls.
The use of enantioselective analysis alone makes it difficult toconclude which dechlorination processes were occurring. Theenantioselective dechlorination of several congeners in this studymay yield the formation of other chiral congeners in non-racemicamounts; for example, the formation of PCBs 91 or 95 from PCB149 (Wong et al., 2001a). These chiral daughter congeners mayin turn be enantioselectively degraded, making it difficult to deter-mine whether the non-racemic EFs observed for a particular con-gener arose from enantioselective formation or degradation.Therefore, observation of a non-racemic parent congener andnon-racemic daughter compound does not necessarily mean thatconversion of a chiral parent to a chiral daughter congener hasoccured. However, the use of chiral analysis and the comparisonof possible parent–daughter congeners does make it possibledetermine which processes are not occurring at a particular site.
This can be exemplified at this site, where chiral analysis sug-gests that it is unlikely that the reductive dechlorination of themore highly chlorinated congeners proceeds through the preferen-tial removal of flanked para chlorines. The removal of a flankedpara chlorine form PCB 183 would have formed PCB 149, whichwas found to be racemic, and which in turn can form PCBs 91 or95. The observation of non-racemic proportions of PCB 183 and
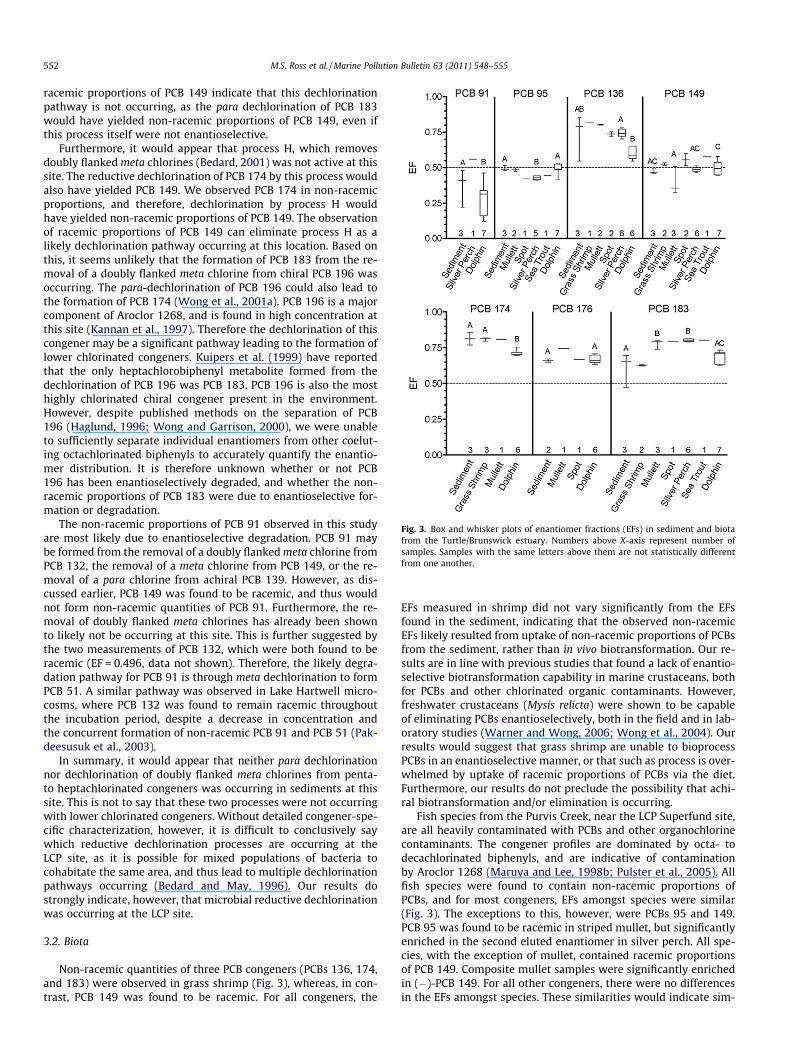
Fig. 3. Box and whisker plots of enantiomer fractions (EFs) in sediment and biotafrom the Turtle/Brunswick estuary. Numbers above X-axis represent number ofsamples. Samples with the same letters above them are not statistically differentfrom one another.
552 M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555
racemic proportions of PCB 149 indicate that this dechlorinationpathway is not occurring, as the para dechlorination of PCB 183would have yielded non-racemic proportions of PCB 149, even ifthis process itself were not enantioselective.
Furthermore, it would appear that process H, which removesdoubly flanked meta chlorines (Bedard, 2001) was not active at thissite. The reductive dechlorination of PCB 174 by this process wouldalso have yielded PCB 149. We observed PCB 174 in non-racemicproportions, and therefore, dechlorination by process H wouldhave yielded non-racemic proportions of PCB 149. The observationof racemic proportions of PCB 149 can eliminate process H as alikely dechlorination pathway occurring at this location. Based onthis, it seems unlikely that the formation of PCB 183 from the re-moval of a doubly flanked meta chlorine from chiral PCB 196 wasoccurring. The para-dechlorination of PCB 196 could also lead tothe formation of PCB 174 (Wong et al., 2001a). PCB 196 is a majorcomponent of Aroclor 1268, and is found in high concentration atthis site (Kannan et al., 1997). Therefore the dechlorination of thiscongener may be a significant pathway leading to the formation oflower chlorinated congeners. Kuipers et al. (1999) have reportedthat the only heptachlorobiphenyl metabolite formed from thedechlorination of PCB 196 was PCB 183. PCB 196 is also the mosthighly chlorinated chiral congener present in the environment.However, despite published methods on the separation of PCB196 (Haglund, 1996; Wong and Garrison, 2000), we were unableto sufficiently separate individual enantiomers from other coelut-ing octachlorinated biphenyls to accurately quantify the enantio-mer distribution. It is therefore unknown whether or not PCB196 has been enantioselectively degraded, and whether the non-racemic proportions of PCB 183 were due to enantioselective for-mation or degradation.
The non-racemic proportions of PCB 91 observed in this studyare most likely due to enantioselective degradation. PCB 91 maybe formed from the removal of a doubly flanked meta chlorine fromPCB 132, the removal of a meta chlorine from PCB 149, or the re-moval of a para chlorine from achiral PCB 139. However, as dis-cussed earlier, PCB 149 was found to be racemic, and thus wouldnot form non-racemic quantities of PCB 91. Furthermore, the re-moval of doubly flanked meta chlorines has already been shownto likely not be occurring at this site. This is further suggested bythe two measurements of PCB 132, which were both found to beracemic (EF = 0.496, data not shown). Therefore, the likely degra-dation pathway for PCB 91 is through meta dechlorination to formPCB 51. A similar pathway was observed in Lake Hartwell micro-cosms, where PCB 132 was found to remain racemic throughoutthe incubation period, despite a decrease in concentration andthe concurrent formation of non-racemic PCB 91 and PCB 51 (Pak-deesusuk et al., 2003).
In summary, it would appear that neither para dechlorinationnor dechlorination of doubly flanked meta chlorines from penta-to heptachlorinated congeners was occurring in sediments at thissite. This is not to say that these two processes were not occurringwith lower chlorinated congeners. Without detailed congener-spe-cific characterization, however, it is difficult to conclusively saywhich reductive dechlorination processes are occurring at theLCP site, as it is possible for mixed populations of bacteria tocohabitate the same area, and thus lead to multiple dechlorinationpathways occurring (Bedard and May, 1996). Our results dostrongly indicate, however, that microbial reductive dechlorinationwas occurring at the LCP site.
3.2. Biota
Non-racemic quantities of three PCB congeners (PCBs 136, 174,and 183) were observed in grass shrimp (Fig. 3), whereas, in con-trast, PCB 149 was found to be racemic. For all congeners, the
EFs measured in shrimp did not vary significantly from the EFsfound in the sediment, indicating that the observed non-racemicEFs likely resulted from uptake of non-racemic proportions of PCBsfrom the sediment, rather than in vivo biotransformation. Our re-sults are in line with previous studies that found a lack of enantio-selective biotransformation capability in marine crustaceans, bothfor PCBs and other chlorinated organic contaminants. However,freshwater crustaceans (Mysis relicta) were shown to be capableof eliminating PCBs enantioselectively, both in the field and in lab-oratory studies (Warner and Wong, 2006; Wong et al., 2004). Ourresults would suggest that grass shrimp are unable to bioprocessPCBs in an enantioselective manner, or that such as process is over-whelmed by uptake of racemic proportions of PCBs via the diet.Furthermore, our results do not preclude the possibility that achi-ral biotransformation and/or elimination is occurring.
Fish species from the Purvis Creek, near the LCP Superfund site,are all heavily contaminated with PCBs and other organochlorinecontaminants. The congener profiles are dominated by octa- todecachlorinated biphenyls, and are indicative of contaminationby Aroclor 1268 (Maruya and Lee, 1998b; Pulster et al., 2005). Allfish species were found to contain non-racemic proportions ofPCBs, and for most congeners, EFs amongst species were similar(Fig. 3). The exceptions to this, however, were PCBs 95 and 149.PCB 95 was found to be racemic in striped mullet, but significantlyenriched in the second eluted enantiomer in silver perch. All spe-cies, with the exception of mullet, contained racemic proportionsof PCB 149. Composite mullet samples were significantly enrichedin (�)-PCB 149. For all other congeners, there were no differencesin the EFs amongst species. These similarities would indicate sim-

M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555 553
ilarities in uptake and/or biotransformation of PCBs amongst thesefour species, which is supported by of a lack of discernable differ-ence amongst species in PCB conger profiles (Pulster et al., 2005).
The measurement of non-racemic proportions of PCBs within anorganism does not, in and of itself, indicate that biotransformationwithin the organism is occurring. Exposure and subsequent uptakeof non-racemic quantities of PCBs, followed by non-enantioselec-tive or no biotransformation may also lead to non-racemic EFs inan organism (Vetter and Maruya, 2000). In order to ascertainwhether or not biotransformation was occurring, enantiomer dis-tributions within an organism must be compared to that of itsexposure source. Due to the low number of detectable congenersin grass shrimp, fish EFs were compared to those found in thetop layer of the sediment cores, as this likely represents the distri-bution of enantiomers to which the base of the food web is ex-posed to. The exception to this was the NSM core, which lackeddetectable levels of several congeners in the uppermost layer.Due to the constancy of the EFs measured in the cores, the top mostlayer with detectable levels of PCBs 91, 95, 136, 176, and 183 (gen-erally 8–10 cm) were used for statistical testing.
Significant differences between sediment and fish EFs werefound only for PCBs 95 and 183, indicating that these speciesmay be biotransforming these congeners enantioselectively. Race-mic proportions of PCB 95 were found in mullet, while EFs in perchwere enriched in the second eluted enantiomer of PCB 95 and sig-nificantly different from those found in sediment. A slight enrich-ment in the second eluted enantiomer of PCB 95 was alsodetected in spot and seatrout, although EFs were only measurablein single samples of each species, and consequently enrichment ofthis congener should be interpreted with caution. An oppositetrend was found for PCB 149, with mullet being significantly moreenriched in (�)-PCB 149, with all other fish species mirroring EFsfound in the sediment. Thus, mullet appeared unable to biotrans-form PCB 95 enantioselectively, but are able to biotransform PCB149 in a enantioselective manner.
Fish species are generally considered to have low biotransfor-mation capacity towards PCBs, although they have been found tobiotransform some congeners enantioselectively (Buckman et al.,2006; Wiberg et al., 2006; Wong et al., 2002b). Field studies havefound non-racemic EFs of congeners in fish species that differedfrom those found in their prey or sediment, and enantioselectiveelimination has been confirmed in laboratory experiments. Our re-sults are consistent with results from Lake Superior and rivers fromthe United States (Wong et al., 2001b, 2004), where forage fishwere found to be capable of biotransforming PCB 95. In contrastto our findings, these studies, as well as laboratory based studies,found some of the fish species investigated can biotransform PCB136 in an enantioselective manner. Enantiomer fractions of PCBshave yet to be reported in any of the fish species investigated inthis study, and it is difficult to draw conclusions based on compar-isons to other species, as it is clear that species differ in their capa-bility for enantioselective biotransformation (Wong et al., 2004).
PCB 183 was found to be non-racemic, with EFs shifted relativeto sediment by all fish species in this study (Fig. 3). Generally, PCB183 is found to be non-biotransformed in fish. The exception tothis is trout from Lake Superior, which have been found to be en-riched in the second eluting enantiomer. PCB 183 has also beenfound in non-racemic proportions in mammalian and avian spe-cies. PCB 183 is often considered difficult to metabolize, with lowrates of elimination from organisms. This congener lacks vicinalmeta–para hydrogens, the presence of which makes a congenermore susceptible to biotransformation by CYP 2B-like enzymes.However, the lack of vicinal meta–para hydrogens does not neces-sarily preclude metabolism by CYP 2B-like enzymes, as only a sin-gle unsubstituted meta or para position is necessary for theformation of hydroxylated metabolites. Furthermore, the finding
of enantioselective biotransformation of PCB 183 in these andother species, while other congeners which posses vicinal meta–para hydrogens are transformed non-enantioselectively may indi-cate other process or other enzymes which may be responsiblefor the observed non-racemic distributions. Alternatively, thenon-racemic proportions of PCB 183 in fish may be the result of up-take from prey that also exhibited a non-racemic EF and that werenot analyzed in this study.
With the exception of PCBs 136 and 149, dolphins were foundto have non-racemic distributions of all PCB congeners investi-gated (Fig. 3). Non-racemic EFs have previously been found in ceta-ceans. Striped dolphins from the Mediterranean were found tocontain non-racemic quantities of PCBs 95, 149, 174, and 176(Reich et al., 1999) and PCBs 95, 149 and 176 were found to benon-racemic in bottlenose dolphins found stranded along the Ital-ian coast (Jiménez et al., 2000). Non-racemic EFs of PCBs 95, 132,and 149 have also been found in adult harbour porpoises (Phocoenaphocoena) from the North Sea (Chu et al., 2003b). These studies,however, did not characterize the enantiomer distribution in anyprey species, and therefore, it is unknown whether or not thesenon-racemic EFs are due to biotransformation, or uptake of non-racemic proportions from prey.
The EFs measured in this study were enriched in the samedirection for PCBs 95 and 176 as those found in other dolphins,although in the opposite direction than EFs of PCB 95 found in har-bour porpoises (Chu et al., 2003b). In addition, this study foundthat EFs are considerably more enriched in a single enantiomerthan those previously found in cetaceans. Because of the enantio-mer enrichment observed in prey species, this is likely due to acombination of exposure to highly non-racemic EFs in the foodweb from this estuary, as well as in vivo biotransformation.Although these animals are likely year round residents of this re-gion (Pulster and Maruya, 2008), the duration and seasonality ofexposure to Aroclor 1268 contaminated prey for these animalsmay also play a role in interpreting these EF signatures.
A comparison of EFs in dolphins to those in prey species indi-cates that dolphins may be biotransforming several PCB congenersenantioselectively. Significant differences between EFs in dolphinsand those in fish species were found for PCBs 95, 136, 149, and 183.In general, EFs in dolphins were less enriched (closer to racemic)than EFs observed in fish. This may indicate that the biotransfor-mation processes occurring in dolphins are enantioselective to-wards the antipode of the enantiomers favored in fish. Theability of cetaceans to biotransform PCBs enantioselectively hasbeen found in bowhead whale (Balaena mysticetus), which had sig-nificantly different EFs for PCBs 91, 135, 149, 174, 176, and 183that those found in zooplankton (Calanus spp.), their primary preyitem (Hoekstra et al., 2002).
The observation that dolphins may be enantioselectively bio-transforming highly chlorinated PCBs, based on comparisons ofEFs between dolphins and likely prey, is a novel and significantfinding. Dolphins are generally regarded as having a low capabilityto biotransform PCBs with six or more chlorines, or those congen-ers with three or more ortho chlorines (Tanabe et al., 1988). How-ever, all of the congeners investigated in this study contain morethan three ortho chlorines, and 5 of the 7 congeners contain sixor more chlorines. Furthermore, congeners with no vicinal meta–para hydrogens and more than three ortho chlorines were foundto have no susceptibility towards biotransformation. PCB 183 fitsthis criterion, yet dolphins appear to be biotransforming this con-gener enantioselectively. Our results are corroborated by the find-ing of methylsulfonyl PCBs, the primary end product of PCBmetabolism, in highly non-racemic proportions in harbour por-poises (Chu et al., 2003a).
An alternate explanation may be the influence of other preyspecies that were not measured in this study and that contain race-

Fig. 4. Linear regression of total PCB concentrations versus enantiomer fraction (EF)of PCB 91 and PCB 149 in bottlenose dolphin blubber from the Turtle/Brunswickestuary.
554 M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555
mic or near racemic proportions of these congeners. The EF contri-bution from prey species to the body burden of a predator speciesis a weighted average of EFs found in prey species and the fre-quency at which that prey item is consumed (Wong et al., 2004).Given the limited biotransformation capability of dolphins ob-served in other studies, exposure to more racemic sources of PCBsmay ‘‘dilute’’ the observed EF, shifting it closer to racemic. Whileevidence suggest that these dolphins are feeding almost exclu-sively on prey from the Turtle/Brunswick estuary, they may inhabitmore open water regions of the estuary for unknown periods oftime, as opposed to the inshore tidal creeks from which fish sam-ples were collected. This difference may be enough to expose dol-phins to prey items that contain more racemic distributions ofPCBs.
A significant correlation between total PCB concentration andEFs of PCBs 91 (r2 = 0.79, p = 0.007) and 149 (r2 = 0.58, p = 0.05)was also found (Fig. 4). EFs of PCB 91 became more enriched inthe first eluting enantiomer as concentrations increase, while EFsof PCB 149 became less enriched. Correlations between EF andPCB concentration have been found in lake trout and for male bow-head whales (Hoekstra et al., 2002; Wong et al., 2004). It is cur-rently unclear why such a correlation exists, although a similarcorrelation between EF and concentration has been linked to vari-ation in feeding habits in other species (Ross et al., 2008).
4. Conclusions
Non-racemic EFs measured in marsh sediments confirms thepresence of reductive dechlorination activity at this site. Further-more, biota contained highly non-racemic EFs, likely the result ofa combination of uptake of non-racemic proportions of these cong-eners from the diet, and in vivo biotransformation. While furtherstudies should be conducted to determine how PCB concentration,trophic level, and feeding ecologies affect EFs, this study exempli-fies how the use of chiral analysis may be used to detect biotrans-formation processes that were otherwise thought not to beoccurring.
Acknowledgements
Funding for this work was provided to C.S.W. by the Natural Sci-ences and Engineering Research Council of Canada, and the CanadaResearch Program. We also thank P. Peronard and L. Francendesefor access to the LCP site and E. Zolman, T. Speakman and L.
Schwacke for their assistance in obtaining bottlenose dolphin sam-ples under MMPA Permit No. 779-1633-00.
References
Asher, B.J., D’Agostino, L.A., Way, J.D., Wong, C.S., Harynuk, J.J., 2009. Comparison ofpeak integration methods for the determination of enantiomeric fraction inenvironmental samples. Chemosphere 75, 1042–1048.
Asher, B.J., Wong, C.S., Rodenburg, L.A., 2007. Chiral source apportionment ofpolychlorinated biphenyls to the Hudson River estuary atmosphere and foodweb. Environmental Science and Technology 41, 6163–6169.
Bedard, D., 2001. Microbial dechlorination of PCBs in aquatic sediment. In:Robertson, L.W., Hansen, L.G. (Eds.), PCBs: Recent Advances in EnvironmentalToxicology and Health Effects. The University Press of Kentucky, Lexington, KY,pp. 27–33.
Bedard, D.L., May, R.J., 1996. Characterization of the polychlorinated biphenyls inthe sediments of Woods Pond: evidence for microbial dechlorination of Aroclor1260 in situ. Environmental Science and Technology 30, 237–245.
Brown, J.F., Bedard, D.L., Brennan, M.J., Carnahan, J.C., Feng, H., Wagner, R.E., 1987.Polychlorinated biphenyl dechlorination in aquatic sediments. Science 236,709–712.
Buckman, A.H., Wong, C.S., Chow, E.A., Brown, S.B., Solomon, K.R., Fisk, A.T., 2006.Biotransformation of polychlorinated biphenyls (PCBs) and bioformation ofhydroxylated PCBs in fish. Aquatic Toxicology 78, 176–185.
Bzdusek, P.A., Christensen, E.R., Lee, C.M., Pakdeesusuk, U., Freedman, D.L., 2006.PCB congeners and dechlorination in sediments of Lake Hartwell, SouthCarolina, determined from cores collected in 1987 and 1998. EnvironmentalScience and Technology 40, 109–119.
Chu, S.G., Covaci, A., Haraguchi, K., Voorspoels, S., Van de Vijver, K., Das, K.,Bouquegneau, J.-M., De Coen, W., Blust, R., Schepens, P., 2003a. Levels andenantiomeric signatures of methyl sulfonyl PCB and DDE metabolites in livers ofharbor porpoises (Phocoena phocoena) from the southern North Sea.Environmental Science and Technology 37, 4573–4578.
Chu, S.G., Covaci, A., Van de Vijver, K., De Coen, W., Blust, R., Schepens, P., 2003b.Enantiomeric signatures of chiral polychlorinated biphenyl atropisomers inlivers of harbour porpoises (Phocoena phocoena) from the southern North Sea.Journal of Environmental Monitoring 5, 521–526.
Haglund, P., 1996. Isolation and characterisation of polychlorinated biphenyl (PCB)atropisomers. Chemosphere 32, 2133–2140.
Harner, T., Wiberg, K., Norstrom, R., 2000. Enantiomer fractions are preferred toenantiomer ratios for describing chiral signatures in environmental analysis.Environmental Science and Technology 34, 218–220.
Hoekstra, P.F., Wong, C.S., O’Hara, T.M., Solomon, K.R., Mabury, S.A., Muir, D.C.G.,2002. Enantiomer-specific accumulation of PCB atropisomers in the bowheadwhale (Balaena mysticetus). Environmental Science and Technology 36, 1419–1425.
Jiménez, O., Jiménez, B., Gonzalez, M.J., 2000. Isomer-specific polychlorinatedbiphenyl determination in cetaceans from the Mediterranean Sea:enantioselective occurrence of chiral polychlorinated biphenyl congeners.Environmental Toxicology and Chemistry 19, 2653–2660.
Kaiser, K.L.E., 1974. Optical-activity of polychlorinated biphenyls. EnvironmentalPollution 7, 93–101.
Kannan, K., Maruya, K.A., Tanabe, S., 1997. Distribution and characterization ofpolychlorinated biphenyl congeners in soil and sediments from a Superfund sitecontaminated with Aroclor 1268. Environmental Science and Technology 31,1483–1488.
Kannan, K., Nakata, H., Stafford, R., Masson, G.R., Tanabe, S., Giesy, J.P., 1998.Bioaccumulation and toxic potential of extremely hydrophobic polychlorinatedbiphenyl congeners in biota collected at a Superfund site contaminated withAroclor 1268. Environmental Science and Technology 32, 1214–1221.
Kuipers, B., Cullen, W.R., Mohn, W.W., 1999. Reductive dechlorination ofnonachlorobiphenyls and selected octachlorobiphenyls by microbialenrichment cultures. Environmental Science and Technology 33, 3579–3585.
Li, X., Yang, L., Jans, U., Melcer, M.E., Zhang, P.F., 2007. Lack of enantioselectivemicrobial degradation of chlordane in Long Island Sound sediment.Environmental Science and Technology 41, 1635–1640.
Magar, V.S., Johnson, G.W., Brenner, R.C., Quensen, J.F., Foote, E.A., Durell, G., Ickes,J.A., Peven-McCarthy, C., 2005. Long-term recovery of PCB-contaminatedsediments at the Lake Hartwell Superfund site: PCB dechlorination. 1. End-member characterization. Environmental Science and Technology 39, 3538–3547.
Maruya, K.A., Lee, R.E., 1998a. Biota-sediment accumulation and trophic transferfactors for extremely hydrophobic polychlorinated biphenyls. EnvironmentalToxicology and Chemistry 17, 2463–2469.
Maruya, K.A., Lee, R.F., 1998b. Aroclor 1268 and toxaphene in fish from asoutheastern US estuary. Environmental Science and Technology 32, 1069–1075.
Morrissey, J.A., Bleackley, D.S., Warner, N.A., Wong, C.S., 2007. Enantiomer fractionsof polychlorinated biphenyls in three selected Standard Reference Materials.Chemosphere 66, 326–331.
Pakdeesusuk, U., Jones, W.J., Lee, C.M., Garrison, A.W., O’Niell, W.L., Freedman, D.L.,Coates, J.T., Wong, C.S., 2003. Changes in enantiomeric fractions duringmicrobial reductive dechlorination of PCB132, PCB149, and Aroclor 1254 inLake Hartwell sediment microcosms. Environmental Science and Technology37, 1100–1107.

M.S. Ross et al. / Marine Pollution Bulletin 63 (2011) 548–555 555
Pakdeesusuk, U., Lee, C.M., Coates, J.T., Freedman, D.L., 2005. Assessment of naturalattenuation via in situ reductive dechlorination of polychlorinated biphenyls insediments of the Twelve Mile Creek arm of Lake Hartwell, SC. EnvironmentalScience and Technology 39, 945–952.
Palekar, L.D., Maruya, K.A., Kostka, J.E., Wiegel, J., 2003. Dehalogenation of 2,6-dibromobiphenyl and 2,3,4,5,6-pentachlorobiphenyl in contaminated estuarinesediment. Chemosphere 53, 593–600.
Pulster, E.L., Maruya, K.A., 2008. Geographic specificity of Aroclor 1268 in bottlenosedolphins (Tursiops truncatus) frequenting the Turtle/Brunswick River estuary,Georgia (USA). Science of the Total Environment 393, 367–375.
Pulster, E.L., Smalling, K.L., Maruya, K.A., 2005. Polychlorinated biphenyls andtoxaphene in preferred prey fish of coastal southeastern US bottlenosedolphins (Tursiops truncatus). Environmental Toxicology and Chemistry 24,3128–3136.
Pulster, E.L., Smalling, K.L., Zolman, E., Schwacke, L., Maruya, K.A., 2009. Persistentorganochlorine pollutants and toxaphene congener profiles in bottlenosedolphins (Tursiops truncatus) frequenting the Turtle/Brunswick River Estuary,Georgia, USA. Environmental Toxicology and Chemistry 28, 1390–1399.
Reich, S., Jimenez, B., Marsili, L., Hernández, L.M., Schurig, V., González, M.J., 1999.Congener specific determination and enantiomeric ratios of chiralpolychlorinated biphenyls in striped dolphins (Stenella coeruleoalba) from theMediterranean Sea. Environmental Science and Technology 33, 1787–1793.
Ross, M.S., Verreault, J., Letcher, R.J., Gabrielsen, G.W., Wong, C.S., 2008. Chiralorganochlorine contaminants in blood and eggs of glaucous gulls (Larushyperboreus) from the Norwegian Arctic. Environmental Science andTechnology 42, 7181–7186.
Singer, A.C., Wong, C.S., Crowley, D.E., 2002. Differential enantioselectivetransformation of atropisomeric polychlorinated biphenyls by multiplebacterial strains with different inducing compounds. Applied andEnvironmental Microbiology 68, 5756–5759.
Tanabe, S., Watanabe, S., Kan, H., Tatsukawa, R., 1988. Capacity and mode of PCBmetabolism in small cetaceans. Marine Mammal Science 4, 103–124.
Vetter, W., Maruya, K.A., 2000. Congener and enantioselective analysis of toxaphenein sediment and food web of a contaminated estuarine wetland. EnvironmentalScience and Technology 34, 1627–1635.
Warner, N.A., Wong, C.S., 2006. The freshwater invertebrate Mysis relicta caneliminate chiral organochlorine compounds enantioselectively. EnvironmentalScience and Technology 40, 4158–4164.
Wiberg, K., Andersson, P.L., Berg, H., Olsson, P.-E., Haglund, P., 2006. The fate ofchiral organochlorine compounds and selected metabolites in intraperitoneallyexposed Arctic char (Salvelinus alpinus). Environmental Toxicology andChemistry 25, 1465–1473.
Winger, P.V., Lasier, P.J., Geitner, H., 1993. Toxicity of sediments and pore-waterfrom Brunswick Estuary, Georgia. Archives of Environmental Contaminationand Toxicology 25, 371–376.
Wong, C.S., Garrison, A.W., 2000. Enantiomer separation of polychlorinatedbiphenyl atropisomers and polychlorinated biphenyl retention behavior onmodified cyclodextrin capillary gas chromatography columns. Journal ofChromatography A 866, 213–220.
Wong, C.S., Garrison, A.W., Foreman, W.T., 2001a. Enantiomeric composition ofchiral polychlorinated biphenyl atropisomers in aquatic bed sediment.Environmental Science and Technology 35, 33–39.
Wong, C.S., Garrison, A.W., Smith, P.D., Foreman, W.T., 2001b. Enantiomericcomposition of chiral polychlorinated biphenyl atropisomers in aquatic andriparian biota. Environmental Science and Technology 35, 2448–2454.
Wong, C.S., Hoekstra, P.F., Karlsson, H., Backus, S.M., Mabury, S.A., Muir, D.C.G.,2002a. Enantiomer fractions of chiral organochlorine pesticides andpolychlorinated biphenyls in Standard and Certified Reference Materials.Chemosphere 49, 1339–1347.
Wong, C.S., Lau, F., Clark, M., Mabury, S.A., Muir, D.C.G., 2002b. Rainbow trout(Oncorhynchus mykiss) can eliminate chiral organochlorine compoundsenantioselectively. Environmental Science and Technology 36, 1257–1262.
Wong, C.S., Mabury, S.A., Whittle, D.M., Backus, S.M., Teixeira, C., DeVault, D.S.,Bronte, C.R., Muir, D.C.G., 2004. Organochlorine compounds in Lake Superior:chiral polychlorinated biphenyls and biotransformation in the aquatic foodweb. Environmental Science and Technology 38, 84–92.
Wong, C.S., Pakdeesusuk, U., Morrissey, J.A., Lee, C.M., Coates, J.T., Garrison, A.W.,Mabury, S.A., Marvin, C.H., Muir, D.C.G., 2007. Enantiomeric composition ofchiral polychlorinated biphenyl atropisomers in dated sediment cores.Environmental Toxicology and Chemistry 26, 254–263.




















