
diversity of edible fish species in adayar estuary, chennai ...

PRIMARY RESEARCH PAPER
Fish habitat use response to anthropogenic induced changesof physical processes in the Elwha estuary, Washington,USA
J. Anne Shaffer • M. Beirne • T. Ritchie •
R. Paradis • D. Barry • P. Crain
Received: 5 February 2009 / Revised: 2 September 2009 / Accepted: 11 September 2009 / Published online: 19 October 2009
� Springer Science+Business Media B.V. 2009
Abstract The Elwha River estuary has been signif-
icantly influenced by anthropogenic changes to the
river, including two large dams upriver and rock
dikes installed in the estuary. Together these have
disrupted hydrodynamic processes and subsequent
sediment delivery throughout the watershed. This
article defines the functional response of fish distri-
bution within the estuary as a result of these changes.
We assessed fish distribution of three main areas of
the Elwha estuary using standard beach seining
techniques from March to August 2007. Species
composition, ecological indices, and relative propor-
tion of all salmonids, and in particular Chinook
salmon (Oncorhynchus tshawytscha), were consis-
tently significantly different across the estuary. Dif-
ferences corresponded to a rock dike installed
30 years ago, and a sediment lens that was observed
to form at the entrance to the east estuary. Sediment
lenses are documented to be a common occurrence in
the Elwha nearshore, and symptomatic of docu-
mented, severely disrupted sediment processes of the
Elwha River. Combining the fish distribution docu-
mented in this study with the rock dike and observed
sediment lens and the sediment processes docu-
mented by other researchers we, therefore, conclude:
(1) Fish use within the Elwha River estuary is
complex, and even fragments of connected estuary
are critically important for migrating salmon; (2)
Anthropogenic effects, including in river damming
and diking of the estuary, can be an important
ecological driver in nearshore habitat function that
should be appropriately considered in estuary habitat
research, management, and restoration; and (3)
Juvenile salmonids appear to be able to respond to
dynamic sediment environments if there are habitat
options available.
Keywords Nearshore � Estuary � Sediment �Hydrodynamics � Salmon � Chinook � Elwha � Habitat
Handling editor: Pierluigi Viaroli
J. A. Shaffer (&) � T. Ritchie
Washington Department of Fish and Wildlife,
332 E. 5th Street, Port Angeles, WA 98362, USA
e-mail: [email protected]
M. Beirne
Lower Elwha Klallam Tribe, Fish Hatchery Road,
Port Angeles, WA 98362, USA
e-mail: [email protected]
R. Paradis � D. Barry
Western Washington University, Huxley College of the
Environment, Port Angeles, WA 98362, USA
e-mail: [email protected]
D. Barry
e-mail: [email protected]
P. Crain
Olympic National Park, 600 Park Avenue, Port Angeles,
WA 98362, USA
e-mail: [email protected]
123
Hydrobiologia (2009) 636:179–190
DOI 10.1007/s10750-009-9947-x

Introduction
Hydrodynamic processes, including channel morphol-
ogy and sediment delivery, define estuarine habitat
form (Marshall & Elliot, 1998; Williams et al., 2002).
Hydrodynamic processes can result in significant
impacts to habitat function and fish population
dynamics. For example, Gregory (1993) and Gregory
& Levings (1998) found that turbidity defined preda-
tion success by juvenile Chinook in the Fraser River,
and Hood (2002) documented the importance of
hydrodynamic flow in habitat utilization in estuaries.
Sediment delivery is another component of estuarine
hydrodynamics that forms nearshore habitats.
Although an important component of estuarine hydro-
dynamics, less is known about the role that sediment
delivery plays in estuarine habitat function for fish.
Similarly, human alterations, including diking and
filling of nearshore environments, can significantly
disrupt hydrodynamic processes and, as a result,
habitat function (Hood, 2004; Toft et al., 2007).
Located on the north Olympic Peninsula in Wash-
ington state, USA, the Elwha River supports no fewer
than three federally listed salmon (Winter & Crain,
2008). The Elwha estuary, which connects the Elwha
watershed with the Strait of Juan de Fuca, is a
classified a dynamic stream delta estuary that is
defined by high-energy riverine and marine processes
(Todd et al., 2006). The current estuary is relatively
small, and includes approximately 90 acres of tidal
channel estuary. Prior to the early 1900s, the Elwha
estuary was much larger, more diverse structurally,
and included complex tidal channels and distributaries
as well as tidal lagoons. The river mouth was dynamic,
and defined by shifting sand bars that formed offshore
at the river mouth (Todd et al., 2006).
The Elwha estuary has been hydromorphologically
disrupted by two anthropogenic events:
(1) Two hydroelectric dams installed in the
watershed have resulted in approximately
100 years of sediment starvation throughout the
lower watershed and nearshore areas. The dams
are both managed as run-of-the-river and do not
regulate water flow, but have severely disrupted
sediment processes by inhibiting transport of sand
and gravel sediment to the lower river. Disruption
of sediment delivery has had a number of effects
on the hydrodynamics of the lower river and
estuary of the Elwha (Draut et al., 2008). Conse-
quently the estuary is now much smaller and less
complex, and has a smaller, foreshortened, and
unstable river mouth and tidal lagoon area. The
Elwha lower river that bisects the estuary is much
more channelized, and has repeatedly migrated
laterally to the east and west over the last 67 years
(Draut et al., 2008; Todd et al., 2006). Although
not quantified, ephemeral sediment bars/lenses
continue to form at the mouth (Lower Elwha
Klallam Tribe, pers obs.; Todd et al., 2006).
(2) An earthen and rock dike along the west estuary
installed in the 1960s has resulted in a truncated
and functionally disconnected estuary; that has
disconnected approximately one-third of the west
estuary from the rest of the lower river habitat and
continues to modify riverine flow patterns.
The current Elwha estuary can be split into three
sections that are at the same tidal elevation and
connection to the Strait shoreline: (1) The east
estuary, located east of the river mouth, which
includes approximately 63 acres (71% of the total
estuary); (2) The west estuary, which includes 18
acres of habitat (19% of the total estuary), which is
connected directly to the west Elwha river mouth on
its east side, and is bordered to the west by a rock
levee; and (3) The impounded estuary, which
includes 9 acres (10% of total estuary) of estuary,
and is separated from the unimpounded west estuary
by the dike. This west levee creates a total fish barrier
between the river and the impounded estuary (Fig. 1).
The Elwha River watershed is slated to undergo a
large-scale restoration event via the removal of two
large hydroelectric dams that were installed in the
river over a 100 years ago and that have severely
disrupted the sediment processes of the Elwha River.
Damming has had a significant impact on the Elwha
River ecosystem, including increasing the erosion of
the river bed, a decrease in recruitment of gravels
needed to create suitable habitats for spawning,
which have been trapped in the sediment load behind
the dams, and an increase in water temperature which
negatively impacts fish spawning and rearing. A
special edition of Northwest Science has been
dedicated to the science and restoration of the Elwha
watershed (Duda et al., 2008).
Dam removal is slated to begin in 2011 (Winter &
Crain, 2008), and will result in the transport of
180 Hydrobiologia (2009) 636:179–190
123

approximately 1.5–2.3 million m3 of coarse (sand and
gravel) sediment and (4–5 million m3) of fine (sand/
silt) sediment from the upper river to the nearshore,
including the Elwha estuary, within 5 years of dam
removal (Randle et al., 2004).
Given the critical importance of estuarine systems
for juvenile fish survival (Beamer et al., 2003, 2005;
Fresh, 2006), the historic importance of the Elwha
watershed to federally listed salmon species, and the
large scale restoration that is to occur, understanding
how the Elwha estuary functions is central to defining
and understanding the full ecosystem recovery (Shaffer
et al., 2008). At present, very little is known about basic
fish distribution of the Elwha estuary. The Washington
Department of Fish and Wildlife and the Lower Elwha
Klallam Tribe, with numerous partners including
Western Washington University and Peninsula Col-
lege, have therefore begun monitoring fish distribution
of the Elwha estuary to assess habitat function. In this
study, we attempt to understand fish distribution of the
three areas of the Elwha estuary and, if different,
identify what role the physical processes that have been
disrupted by anthropogenic features of sediment
starvation and diking may play in fish distribution.
We also define the next priority steps in ecosystem
monitoring, management, and restoration.
Methods and materials
We define habitat function based on fish distribution
as reflected by various ecological indices, including
diversity, abundance, and morphological metrics. Our
hypotheses for this study are as follows:
H01 Species diversity is not significantly different
between impounded, west, and east portions of the
Elwha estuary;
H02 Total fish and salmon abundance is not signif-
icantly different between impounded, west, and east
portions of the Elwha estuary, and;
H03 Fish length for major salmon species is not
significantly different between east and west estuary.
S T R A I T O F J U A N D E F U C AS T R A I T O F J U A N D E F U C A
Elwha River Main Channel
Elwha River Main Channel
ImpoundedWest Estuary
4.1 ha.
EastEstuary28.5 ha.
West Estuary8.7 ha.
0 200100
Meters
1996 Shoreline
LEKT Reservation
2001 LiDaR Elevation (ft.)
�2 � 4
5 � 10
11 � 15
16 � 25
26 � 1,304
Lower Elwha Klallam Tribal G.I.S. Dept.Map prepared by: Randall E. McCoy
Elwha River Estuary 09 (hydrobiologia).mxd
Location MapLocation Map
Project SiteProject Site
WashingtonState
WashingtonState
Fig. 1 Map of the Elwha Estuary with elevational profile and the three sampling areas delinated. Asterisks indicate sample sites
Hydrobiologia (2009) 636:179–190 181
123

The east, west, and impounded areas of the Elwha
estuary were sampled using standard beach seining
methodology during the salmon outmigration period
(March–August) 2007. Large and small Puget Sound
Protocol (PSP) nets were used in the west estuary.
The large PSP seine net was used in the east estuary.
A total of 4–6 seines were conducted across the three
stations of the east estuary every other week. A total
of 1–2 seines were conducted along the west estuary
weekly. Two seines were conducted in the
impounded estuary weekly. By the end of May,
dense accumulations of green algal mats in the
impounded estuary made the area unworkable, and
potentially unsafe for human health. The impounded
portion of the west estuary was therefore only
sampled during March–May 2007. During each
sampling, all the fish in the net were identified to
the lowest possible taxa and counted. The first
twenty-five individuals of each species were mea-
sured in both fork and total fish length and recorded
in millimeters. Species richness and Shannon–Wiener
diversity (H0) indices were calculated (Zar, 1984) for
each month, and for each portion of the estuary.
ANOVA and t-tests were used to evaluate differ-
ences in study variables between the three estuary
areas using the coin package (Hothorn et al., 2008) in
the R system for statistical computing (ver. 2.8.0, R
Development Core Team, 2008) and RT4Win soft-
ware (Huo et al., 2006). Monte Carlo simulation with
10,000 replications was used to generate approximate
P-values for ANOVAs and systematic permutation
was used to generate exact P-values for t-tests
(Edgington & Onghena, 2007). Also using R, Welch’s
correction for the t-distribution was used to obtain 95%
confidence intervals on means as well as the differ-
ences between means. Percent similarity indices were
calculated to compare sites by evaluating the density
of each species that occurred in at least one site with
the other two sites. For each species, the lowest
relative species density value is selected, and the sum
of those species density percentages among sites
reflects the degree of similarity among sites (McCune
et al., 2002).
Results
A total of 118 seines were conducted from March to
August 2007 (Table 1). Salmon, particularly Chinook
(Oncorhynchus tshawytscha), coho (O. kisutch),
chum (O. keta), cutthroat (Salmo clarki clarki), and
steelhead (Salmo gairdneri), smelt (primarily Hy-
pomesus pretiousus pretiousus), threespine stickle-
back (Gasterosteus aculeatus), and staghorn sculpin
(Leptocottus armatus) were the dominant fish species
collected (Tables 2, 3). Fish species composition
varied in each area. Ecological indices and individual
species densities also varied by month, with higher
densities and diversities observed during June, July,
and August (Figs. 2, 3; Tables 4, 5). In total, 47% of
the fish were collected in the west estuary. Over 90%
of juvenile salmonids (all species) and 94% of
juvenile Chinook salmon were collected in the west
estuary (Tables 2, 4). The total number of fish
collected in the impounded estuary was comparable
to the other sites in March and April, and had the
highest percentage of fish in May. However, the
species composition of the impounded estuary dif-
fered from that of the east and west estuary. Juvenile
salmonids were not collected in the impounded
estuary, which was instead dominated by large
populations of three spine sticklebacks.
Species diversity (F = 10.338, P = 0.004) and
richness (F = 19.513, P = 0.0001) were statistically
different between the three sites (Fig. 2; Table 6).
The west estuary had the highest values for all the
indices. In general, the greatest differences in diver-
sity and richness were between the west estuary and
the impounded estuary, followed by the east and west
estuary. Percent similarity analysis indicated the
greatest similarity between the east and impounded
estuary, and the lowest similarity between the west
and impounded estuary (Table 7).
Frequency of occurrence of all the fish species
combined was not significantly different among the
three sites (Fig. 2). Densities of Chinook, coho, and
Table 1 Seining summary Elwha estuary 2007
Date Number of Seines by site Total
East West Impounded
March 10 2 2 14
April 12 6 6 24
May 15 6 10 31
June 8 6 ns 14
July 10 8 ns 18
August 4 5 ns 9
182 Hydrobiologia (2009) 636:179–190
123

chum salmon were all significantly greater in the west
estuary than in the other areas of the Elwha estuary
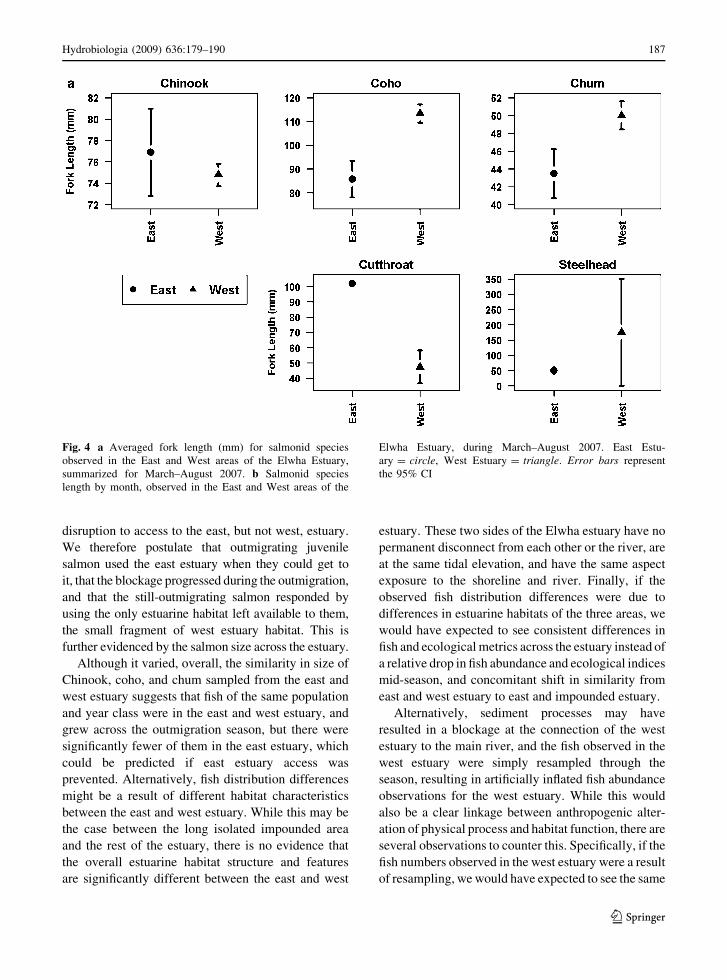
(Fig. 4; Table 4).
Body length of salmonids among sites was vari-
able. Chum and Chinook length increased during the
sampling season. Cutthroat, steelhead, and coho
lengths generally remained consistent. Lengths for
Chinook and chum differed significantly by month
(Fig. 4a, b). Fish lengths for all the species differed
significantly between east and west portions of the
estuary (P \ 0.001).
Discussion
Fish distribution within the Elwha estuary reflects
complexity in fish life history and habitat. Fish
abundance varied through the season, corresponding
with salmon outmigration in the east and west estuary,
and strong seasonal reproduction of non-migrating
species (sculpin and sticklebacks) primarily in the
impounded and east estuary. Salmon outmigration
observed during this study includes both wild and in-
river hatchery releases of coho, Chinook, and steel-
head. Changes in salmon lengths over the sampling
period reflect growth of fish during the season.
Surprisingly, mean coho size did not increase during
the sampling period, likely due to hatchery supple-
mentation (Crain et al., pers obs.).
Statistical analysis of ecological indices and fish
metrics led us to reject all three of our null hypotheses
and conclude that the three areas of the Elwha estuary
appear to be functioning differently for fish. Percent
similarity results reveal that the east and impounded
areas of the estuary exhibited relatively more similar
ecological characteristics than the connected east and
west estuaries. The difference in Chinook and chum
densities of the east and west estuary was also found to
be biologically statistically significant, with more fish
present on the west than the east estuary.
Given the similarities in tidal elevation, proximity
to shoreline and river, and lack of other habitat
alterations of the three areas, the functional differ-
ences between the three areas of the Elwha estuary
are likely due to anthropogenic events that result in
physical obstructions to fish movement in two ways.
First, fish access to the impounded portion of the
Elwha estuary has been obstructed for decades by the
permanent dike, which is a complete fish passage
barrier, resulting in a much different fish assemblage
than observed in portions of the estuary that migrat-
ing fish can access. Second, sediment bars or lens are
a common feature offshore of the Elwha River mouth
as documented in historic accounts (Todd et al.,
2006). One such sediment lens was observed, but not
quantified, to have formed in the lower river at the
entrance to the east estuary at or just prior to the
beginning of the annual juvenile salmon outmigration
period in 2007 (Lower Elwha Klallam Tribe pers
obs.). This lens is believed to have disrupted fish
Table 2 Percent summary of dominant fish collected from Elwha estuary
Site 3-Spine
stickleback
Chinook Coho Chum Cut throat Steelhead Smelt Starry
flounder
Staghorn
sculpin
Cottids
Impounded 100 0 0 0 0 0 0 0 0 0
East 82 3 1 0 0 1 0 3 6 3
West 40 25 4 2 1 0 0 6 8 11
Table 3 Percentage of all fish collected in east, west, and
impounded areas of Elwha estuary by month
Percent of total fish
East
estuary
West
estuary
Impounded Total
Date
March 27 33 40 100
April 27 39 34 100
May 7 19 75 100
June 38 62 ns 100
July 27 73 ns 100
August 48 52 ns 100
Total percent
March–May 11 23 66 100
June–August 41 59 ns 100
All months 32 47 21 100
Note that owing to sampling limitations, the impounded
estuary was sampled during March–May only
Hydrobiologia (2009) 636:179–190 183
123

184 Hydrobiologia (2009) 636:179–190
123

access to the east estuary during, at least, part of our
study. In contrast to these two areas, the west portion
of the Elwha estuary experienced no blockages to fish
access, and as a result reflected a more diverse
estuarine ecosystem with respect to fish diversity and
abundance than the impounded area (literally only
meters away), and the east estuary.
While we were not able to quantify the sediment bar
formation during this study, previous studies by others
that documents variation in sediment delivery to the
lower Elwha River support the idea that the sedimen-
tation lens disrupted fish access to the east estuary.
Between September 2006 and April 2007, the lower
0.5 km of the river channel, including the area of our
study, migrated eastward approximately 10 m, and the
river bed aggraded by as much as 1 m with sand and
gravel deposits (Draut et al., 2007). There is also a
large body of evidence that supports our conclusion of
sedimentation being a driving factor in fish distribu-
tion of the estuary. Sediment processes in the Elwha
River are documented to be significantly disrupted,
and large scale sediment starvation has changed the
hydrodynamic processes of the lower river, causing
large seasonal changes in the river bed (Draut et al.,
2008; Randle et al., 2004). Disruption of physical
processes has played a dominant role in Elwha lower
river and nearshore habitat (Draut et al., 2008; Warrick
et al., 2008; Warrick et al., 2009). Unconstrained, low
gradient channel reaches in the lower Elwha River
historically contained extensive side channels (Pess
et al., 2008) and estuarine slack-water habitats with
suitable substrate for critical fish distribution, includ-
ing eulachon spawning. Truncation of sediment
transport to the lower river, along with channelization
and the systematic removal of large woody debris
(LWD), has caused channel incision and an increase in
bed substrate size (Pohl, 2004). Nearshore effects of
this disruption likely include a significant reduction in
Table 4 Difference in mean densities of salmonids between
the east and west estuaries (fish/m3 ± 95% CI)
Species Difference
(fish/m3 ± 95% CI)
t P
Chinook 0.2127 ± 0.1047 3.36 0.001
Coho 0.0307 ± 0.0278 1.91 0.05
Chum 0.0131 ± 0.0118 1.94 0.04
Table 5 Salmonid abundance summary, total number of fish
and percent, Elwha estuary
East
estuary
West
estuary
Impounded
estuary
Combined
estuary
Total salmon
Number 250 2,527 2 2,780
Percent 9 91 0 100
Chinook
Number 130 1,942 1 2,073
Percent 6 94 0 75
Coho
Number 37 335 0 372
Percent 10 90 0 14
Chum
Number 37 180 0 217
Percent 17 83 0 10
Cutthroat
Number 2 65 0 67
Percent 3 97 0 2
Steelhead
Number 44 5 1 50
Percent 88 10 2 2
Table 6 Comparison of Shannon–Wiener ecological diversity
(H) values for the three areas of Elwha estuary, by month
Month Groups na F or t P
March 3 2/10/2 2.282 0.16
April 3 6/12/6 9.413 0.001
May 3 7/15/10 8.571 0.001
June 2 8/6 2.881 0.018
July 2 10/8 3.956 0.001
August 2 4/6 1.215 0.23
a n values are for East/West/Impounded areas respectively
Table 7 Percent similarity results, fish species and densities,
Elwha estuary
Comparison Percent
similarity
West vs. Impounded 39
East vs. West 61
East vs. Impounded 69
Fig. 2 Comparison of mean values for fish distribution indices
for the three areas of the Elwha Estuary, for the entire
migration season (left side) and by month (right side), 2007: (a)
diversity (H0), (b) species richness, and (c) density (fish/m3).
Error bars represent the 95% CI. a Impounded site sampled
during March–May
b
Hydrobiologia (2009) 636:179–190 185
123

side channel habitat and a reduction in suitable
eulachon spawning habitat (Shaffer et al., 2007).
Sediment delivery to the estuary is also extremely
variable and is documented to result in localized
rising of the river bed (Draut et al., 2007, 2008). This
aggradation appears to have driven habitat function
of the east estuary during the course of this study by
forming a sediment lens at the entrance to the east
estuary that prevented many fish from using the east
estuary.
When combined, the hydrologic disruptions of
sediment deposition and diking appear to be contrib-
uting to reduced ecological indices and salmon species
densities in a significant portion of the Elwha estuary
by preventing fish from accessing areas of the estuary.
The presence of fish in both the east and impounded
areas of the estuary, and high numbers of fish in the
impounded estuary indicates both areas are suitable
habit for fish, provided they can access the habitat. The
intermediate nature of indices and salmon densities in
the east estuary indicate that the barrier to fish
distribution to this part of the estuary is recent, and
given how it formed, likely to be temporary. The
significantly lower species richness, and dominance of
stickleback (a long-lived non-migratory species), and
lack of juvenile salmonids in the impounded estuary
indicates that, due to the permanent and total physical
access barrier for both river and marine fish, it is
functioning differently from the other sites, with lower
long-term ecological function for fish.
There may be factors other than the changes in
sediment deposition and connectivity, which could
have resulted in the observed differences in fish
distribution within the Elwha estuary. Differences in
observations of the east and west estuary may be
attributed to sampling effort. The west estuary is much
smaller than the east estuary. Fish in the east estuary
had a larger area to inhabit, and may have had a lower
probability of capture due to limited sampling area
relative to the habitat available. Trends in ecological
indices and fish abundance, however, do not support
this alternate explanation. Ecological indices and
salmon densities in the east estuary initially were
similar to those in the west estuary, and also increased
with the salmon migration periods. The trends then
drop off within the east estuary only, indicating a
Fig. 3 Mean density (fish/m3) of the five salmonid species and total salmonids, grouped by location within the Elwha Estuary,
March–August 2007. Error bars represent the 95% CI
186 Hydrobiologia (2009) 636:179–190
123
disruption to access to the east, but not west, estuary.
We therefore postulate that outmigrating juvenile
salmon used the east estuary when they could get to
it, that the blockage progressed during the outmigration,
and that the still-outmigrating salmon responded by
using the only estuarine habitat left available to them,
the small fragment of west estuary habitat. This is
further evidenced by the salmon size across the estuary.
Although it varied, overall, the similarity in size of
Chinook, coho, and chum sampled from the east and
west estuary suggests that fish of the same population
and year class were in the east and west estuary, and
grew across the outmigration season, but there were
significantly fewer of them in the east estuary, which
could be predicted if east estuary access was
prevented. Alternatively, fish distribution differences
might be a result of different habitat characteristics
between the east and west estuary. While this may be
the case between the long isolated impounded area
and the rest of the estuary, there is no evidence that
the overall estuarine habitat structure and features
are significantly different between the east and west
estuary. These two sides of the Elwha estuary have no
permanent disconnect from each other or the river, are
at the same tidal elevation, and have the same aspect
exposure to the shoreline and river. Finally, if the
observed fish distribution differences were due to
differences in estuarine habitats of the three areas, we
would have expected to see consistent differences in
fish and ecological metrics across the estuary instead of
a relative drop in fish abundance and ecological indices
mid-season, and concomitant shift in similarity from
east and west estuary to east and impounded estuary.
Alternatively, sediment processes may have
resulted in a blockage at the connection of the west
estuary to the main river, and the fish observed in the
west estuary were simply resampled through the
season, resulting in artificially inflated fish abundance
observations for the west estuary. While this would
also be a clear linkage between anthropogenic alter-
ation of physical process and habitat function, there are
several observations to counter this. Specifically, if the
fish numbers observed in the west estuary were a result
of resampling, we would have expected to see the same
Fig. 4 a Averaged fork length (mm) for salmonid species
observed in the East and West areas of the Elwha Estuary,
summarized for March–August 2007. b Salmonid species
length by month, observed in the East and West areas of the
Elwha Estuary, during March–August 2007. East Estu-
ary = circle, West Estuary = triangle. Error bars represent
the 95% CI
Hydrobiologia (2009) 636:179–190 187
123

trends in total fish numbers in the west and impounded
estuary. Instead, we saw very different trends in fish
numbers in the impounded and west sections of the
estuary, and somewhat different trends between the
east and west estuary. For example, chum abundances
increased steadily in the west estuary (but not in the
impounded or east portions of the estuary), indicating
that the west estuary had regular fish access while the
other areas of the estuary did not. Among the three
areas, the impounded estuary (for the months sampled)
and east estuary appeared to be similar—and in some
cases almost identical—for ecological indices and fish
distribution, indicating that the east estuary and
impounded estuary are similar in fish distribution
due to lack of fish access. The disruption to the east
estuary is presumed to be due to a sediment lens,
formed from well-documented and very disrupted
sediment delivery processes that block fish access.
Fig. 4 continued
188 Hydrobiologia (2009) 636:179–190
123

Because this lens was created by river hydrodynamics
in an extremely variable system, it is a temporary
habitat alteration. Ecological implications of tempo-
rary blockage are at this time unknown, but worthy of
further study. For example, these observations are
consistent with those of Harrison & Whitfield (1995),
who defined similar trends in fish distribution and
ecological linkages within an estuary that is regularly
occluded due to sedimentation. The disruption to the
impounded estuary, on the other hand, is due to the
dike, which is a permanent habitat feature. Ecological
implications of this permanent barrier are likely much
more severe than those associated with temporary
barriers, and a priority for further detailed study.
Our results illustrate the important relationship
between anthropogenic changes to a watershed, and
the role diking and disruption of physical processes of
sediment transport and hydrologic connectivity play in
estuarine habitat utilization by fish. We believe that
fish distribution observed in this study indicates (1)
Diking clearly changes fish distribution of an estuary;
(2) Sediment delivery can result in the same barrier to
fish as a diking; and (3) Juvenile salmonids can
respond fairly quickly to a dynamic sediment land-
scape. If our third conclusion is correct, providing the
fish with the most alternatives for estuarine habitat is
the best alternative for estuarine management and
salmon recovery. The most obvious opportunity for
providing more habitat is to provide hydrologically
restored fish access to the impounded west estuary.
This restoration should be conducted sooner than later.
While the current sediment processes of the Elwha
estuary are dynamic, they will be even more so with
the large sediment pulses that will affect the estuary
within 5 years of dam removal. Dramatic changes in
sediment transportation and river flow are expected
before that natural functionality of the river will be
recovered. During this period, sedimentation pro-
cesses, and associated habitat accessibility, in the
estuary will likely fluctuate. Ecosystem restoration to
allow full hydrologic restoration of the Elwha estuary
should, therefore, be initiated as soon as possible to
allow fish the maximum opportunity for estuarine
habitat access during dam removal and concurrent
habitat restoration.
Concurrent ecosystem monitoring should be initi-
ated as soon as possible, including detailed longer-
term monitoring of fish movement in the estuary as
well as more detailed long-term physical habitat
mapping, including sediment mapping of the river
and estuary to the river mouth, and water quality
monitoring of all elements of the Elwha estuary and
shoreline are a priority. Monitoring should include
defining components of the Elwha estuary prior to
dam removal. Specifically, channel area and type
(blind versus open) and vegetation type (riverine
tidal, scrub–shrub, and emergent marsh), and detailed
water quality should be quantified throughout the
estuary, prior to, during, and after dam removal.
These studies should be conducted concurrently with
long-term fish distribution assessment to quantify the
relationship we have identified in this study.
Acknowledgments This study was sponsored in part by the
North Olympic Peninsula Lead Entity, and by the Salmon
Recovery Funding Board (SRFB), United States
Environmental Protection Agency (EPA), the Washington
Department of Fish and Wildlife (WDFW), and the Lower
Elwha Klallam Tribe. The Clallam Marine Resources
Committee provided funding for two college interns. Student
interns were also provided by Peninsula College Fisheries
Program and Center of Excellence’s NSF REU program, and
Western Washington University. Mr. Jack Ganzhorn and Dr.
Dwight Barry provided student supervision. Nancy Bluestein-
Johnson provided student guidance. Interns who assisted in the
project include Jesse Charles, Chris DeSisto, Bryan Hara, Erica
Hirsh, Mario Laungayan, Romy Laungayan, Ross McDorman,
Sean Oden, Tiffany Nabors, Jacob Ray, Melanie Roed, Justin
Rondeau, Trista Simmons, Ben Warren, Karen Wilkie, Eric
Wood, and Steve Wyall. Jenna Schilke, formerly WDFW,
supervised a portion of the field work. Private property owners
Pam Lowry and Malcom Dudley along with Chuck Janda
provided site access. Project in kind partners in the order of
contribution included Cathy Lear (Clallam County), Dave
Parks (DNR), Brian Winter (ONP), Ross Fuller, Mike Sharpf,
Chris Byrnes, Roger Mosley, Dan Penttila, Dan Doty, and Tim
Quinn (WDFW), Bruce McCarter (DFO),Tim Randle (BoR),
and Amy Draut (USGS). Kurt Fresh (NOAA) and two
anonymous reviewers provided valuable critical manuscript
review. This material is based on research supported in part by
the National Science Foundation under REU Grant No.
0452328 awarded jointly to Peninsula College and Western
Washington University. Any opinions, findings and conclu-
sions, or recommendations expressed in this material are those
of the authors, and do not necessarily reflect the views of the
National Science Foundation.
References
Beamer, E., R. Henderson, A. McBride & K. W. Wolf, 2003.
The importance of non-natal pocket estuaries in Skagit
Bay to wild Chinook salmon: An emerging priority for
restoration. Skagit River System Cooperative, Research
Department, La Connor, Washington.
Hydrobiologia (2009) 636:179–190 189
123

Beamer, E., A. McBride, C. Greene, R. Henderson, G. Hood,
K. Wolf, K. Larsen, C. Rice & K. L. Fresh, 2005. Delta
and nearshore restoration for the recovery of wild Skagit
River Chinook salmon: linking estuary restoration to wild
Chinook salmon populations. Supplement to Skagit Chi-
nook Recovery Plan. Skagit River System Cooperative,
La Conner, Washington.
Draut, A. E., J. B. Logan, R. E. McCoy, J. A. Warrick, E. Todd,
M. McHenry & D. M. Rubin, 2007. Channel Evolution on
the Lower Elwha River 1939–2007. Proceedings, 2007
GSA Annual Meeting. Denver, Colorado.
Draut, A. E., J. B. Logan, R. E. McCoy, M. McHenry & J.
Warrick, 2008. Channel evolution on the lower Elwha
River, Washington, 1939–2006. USGS Scientific Investi-
gations Report 2008–5127. U.S. Geological Survey,
Menlo Park, CA.
Duda, J. J., J. E. Freilich & E. G. Schriener (eds), 2008. Dam
Removal and Ecosystem Restoration in the Elwha
Watershed. Northwest Science 82(Special Issue): 1–12.
Edgington, E. & P. Onghena, 2007. Randomization Tests, 4th
ed. Chapman & Hall/CRC Press, Boca Raton, FL.
Fresh, K. L., 2006. Juvenile Pacific Salmon in Puget Sound.
Puget Sound Nearshore Partnership Report No. 2006–06.
Seattle District, U.S. Army Corps of Engineers, Seattle,
Washington.
Gregory, R. S., 1993. The effect of turbidity on the predator
avoidance behavior of juvenile Chinook salmon (On-corhynchus tshawytscha). Canadian Journal of Fisheries
and Aquatic Sciences 50: 241–246.
Gregory, R. S. & C. D. Levings, 1998. Turbidity reduces
predation on migrating juvenile Pacific salmon. Transac-
tions of the American Fisheries Society 127: 275–285.
Harrison, T. D. & A. K. Whitfield, 1995. Fish community
structure in three temporarily open/closed estuaries on the
natal coast. Ichthyological Bulletin of the J. L. B. Smith
Institute of Ichthyology, Grahamstown, South Africa 64:
1–80.
Hood, W. G., 2002. Application of landscape allometry to
restoration of tidal channels. Restoration Ecology 10:
213–222.
Hood, W. G., 2004. Indirect environmental effects of dikes on
estuarine tidal channels: Thinking outside of the dike for
habitat restoration and monitoring. Estuaries 27: 273–282.
Hothorn, T., K. Hornik, M. A. van de Wiel & A. Zeileis, 2008.
Implementing a class of permutation tests: The coin
package. Journal of Statistical Software 28: 1–23.
Huo, M., P. Onghena & E. Edgington, 2006. RT4Win: Ran-
domization tests software for Windows, version 1.0.
Katholieke Universiteit Leuven, Belgium.
Marshall, S. & M. Elliott, 1998. Environmental influences on
the fish assemblage of the Humber estuary, UK. Estuarine,
Coastal and Shelf Science 46: 175–184.
McCune, B., J. Grace & D. Urban, 2002. Analysis of Eco-
logical Communities. MjM Software Design, Gleneden
Beach, OR.
Pess, G. R., M. L. McHenry, T. J. Beechie & J. Davies, 2008.
Biological impacts of the Elwha River dams and potential
salmonid responses to dam removal. Northwest Science
82(Special Issue): 72–90.
Pohl, M., 2004. Channel bed mobility downstream from the
Elwha dams, Washington. The Professional Geographer
56: 422–431.
R Development Core Team, 2008. R: A Language and Environ-
ment for Statistical Computing. R Foundation for Statistical
Computing, Vienna, Austria. http://www.R-project.org/.
Randle, T. J., J. Bountry, B. Jackson & G. Smillie, 2004. Elwha
River Restoration Draft Sediment Monitoring and Man-
agement Plan. U. S. Department of the Interior, Bureau of
Reclamation and National Park Service. Port Angeles,
Washington.
Shaffer, J. A., D. Penttila, M. McHenry & D. Vilella, 2007.
Observations of Eulachon, Thaleichthys pacificus, in the
Elwha River, Olympic Peninsula, Washington. Northwest
Science 81: 76–81.
Shaffer, J. A., P. Crain, B. Winter, M. McHenry, C. Lear & T.
Randle, 2008. Nearshore restoration of the Elwha River
through removal of the Elwha and Glines Canyon Dams:
An overview. Northwest Science 82(Special Issue): 48–59.
Todd, S., N. Fiztpatrick, A. Carter-Mortimer & C. Weller,
2006. Historical changes to estuaries, spits, and associated
tidal wetland habitats in the Hood Canal and Strait of Juan
de Fuca regions of Washington State. PNPTC Technical
Report 06 01. Point-No Point Treaty Council, Kingston,
Washington.
Toft, J. D., J. R. Cordell, C. A. Simenstad & L. A. Stamatiou,
2007. Fish distribution, abundance, and behavior along
city shoreline types in Puget Sound. North American
Journal of Fisheries Management 27: 465–480.
Warrick, J. A., G. R. Cochrane, Y. Sagy & G. Gelfenbaum, 2008.
Nearshore substrate and morphology offshore of the Elwha
River. Northwest Science 82(Special issue): 153–163.
Warrick, J. A., D. George, G. Gelfenbaum, P. Ruggiero, G.
Kaminsky & M. Beirne, 2009. Beach morphology and
change along the mixed grain-size delta of the Elwha
River, Washington. Geomorphology 111: 136–148.
Williams, P. B., M. Orr & N. J. Garrity, 2002. Hydraulic
geometry: A geomorphic design tool for tidal marsh
channel evolution in wetland restoration projects. Resto-
ration Ecology 10: 577–590.
Winter, B. D. & P. Crain, 2008. Making the case for ecosystem
restoration by dam removal in the Elwha River. Northwest
Science 82(Special issue): 13–29.
Zar, J. H., 1984. Biostatistical Analysis. Prentice-Hall, Engle-
wood Cliffs, NJ.
190 Hydrobiologia (2009) 636:179–190
123
Copyright © 2022 FDOKUMEN




















