
emt-curriculum.pdf - Wisconsin Department of Health Services

E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
ava i l ab l e a t www.sc i enced i r ec t . com
www.e l sev i e r . com/ loca te /yexc r
Research Article
VEGF elicits epithelial-mesenchymal transition (EMT) in prostateintraepithelial neoplasia (PIN)-like cells via an autocrine loop
Oscar Gonzalez-Morenoa, Jon Lecandab, Jeffrey E. Greenc, Victor Segurad, Raul Catenaa,Diego Serranoa, Alfonso Calvoa,⁎aLaboratory of Novel Therapeutic Targets, Division of Oncology, Center for Applied Medical Research (CIMA), University of Navarra,Avda. Pio XII, 55, 31008 Pamplona, SpainbDigna Biotech, and Department of Biochemistry, University of Navarra, Pamplona, SpaincLaboratory of Cancer Biology and Genetics, NCI, NIH, Bethesda , MD, USAdUnit of Proteomics, Genomics and Bioinformatics, CIMA, University of Navarra, Pamplona, Spain
A R T I C L E I N F O R M A T I O N
⁎ Corresponding author. Fax: +34 948 194714E-mail address: [email protected] (A. Calvo).
0014-4827/$ – see front matter © 2009 Elseviedoi:10.1016/j.yexcr.2009.11.020
A B S T R A C T
Article Chronology:
Received 8 July 2009Revised version received24 November 2009Accepted 30 November 2009Available online 16 December 2009
Vascular endothelial growth factor (VEGF) is overexpressed during the transition from prostateintraepithelial neoplasia (PIN) to invasive carcinoma. We have mimicked such a process in vitro
using the PIN-like C3(1)/Tag-derived Pr-111 cell line, which expresses low levels of VEGF andexhibits very low tumorigenicity in vivo. Elevated expression of VEGF164 in Pr-111 cells led to asignificant increase in tumorigenicity, invasiveness, proliferation rates and angiogenesis.Moreover, VEGF164 induced strong changes in cell morphology and cell transcriptome throughan autocrine mechanism, with changes in TGF-beta1- and cytoskeleton-related pathways, amongothers. Further analysis of VEGF-overexpressing Pr-111 cells or following exogenous addition ofrecombinant VEGF shows acquisition of epithelial–mesenchymal transition (EMT) features, withan increased expression of mesenchymal markers, such as N-cadherin, Snail1, Snail2 (Slug) and
vimentin, and a decrease in E-cadherin. Administration of VEGF led to changes in TGF-beta1signaling, including reduction of Smad7 (TGF-beta inhibitory Smad), increase in TGF-betaR-II, andtranslocation of phospho-Smad3 to the nucleus. Our results suggest that increased expression ofVEGF in malignant cells during the transition from PIN to invasive carcinoma leads to EMT throughan autocrine loop, which would promote tumor cell invasion andmotility. Therapeutic blockade ofVEGF/TGF-beta1 in PIN lesions might impair not only tumor angiogenesis, but also the earlydissemination of malignant cells outside the epithelial layer.
© 2009 Elsevier Inc. All rights reserved.
Keywords:
Prostate intraepithelial neoplasia (PIN)VEGFEMTSnail
SlugTGF-β1
Introduction
Prostate cancer is the leading cause of cancer in men in theWestern world [1]. Prostate intraepithelial neoplasia (PIN) hasbeen found in a large proportion of men over 50 years of age. PINlesions are thought to progress to invasive carcinoma overtime,
.
r Inc. All rights reserved.
although the mechanisms causing such transformation are poorlyunderstood [2]. Aging, carcinogens, genetic predisposition, diet,and levels of steroid hormones are factors that may influenceprogression from PIN to invasive carcinoma [2].
The development of prostate cancer includes a multi-stepprogram of genetic alterations in pathways affecting cell cycle,

555E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
apoptosis, adhesion, motility and angiogenesis. The angiogenicswitch is a critical step that allows for the growth of neoplasticlesions [3]. Although overexpression of different growth factorshave been linked to the angiogenic switch, the vascular endothelialgrowth factor (VEGF) plays a key role in this molecular event [3].Three major isoforms generated by alternative splicing areexpressed in mouse tissues (VEGF120, 164, 188), but VEGF164 is byfar the most abundant and proangiogenic one [4]. VEGF actsthrough their receptors VEGF-R1 (Flt-1) and VEGF-R2 (Flk-1/KDR)in endothelial cells to promote cell proliferation, migration andtube formation [5]. Interestingly, both VEGF receptors have alsobeen found in prostate tumor cells in vitro and an autocrinemechanism has been described to induce cell proliferation [6]. Inthe carcinogenic events leading to prostate cancer, the angiogenicswitch takes place during the transition from PIN to invasivecarcinoma [7]. The expression of VEGF and its receptors isincreased in high grade PIN (HGPIN), compared to the normalprostate [7], which results in the development of a vascularnetwork within HGPIN lesions [8]. In the TRAMPmodel of prostatecancer, the angiogenic switch has also been shown to occur as aresult of an up-regulation of HIF-1α and VEGF-R1 in high grade PINlesions preceding invasion [9].
The advent of genetically modified mouse models for prostatecancer has allowed understanding the process of prostatecarcinogenesis in a well defined way. In the C3 (1)/Tag model,PIN lesions can be found in 2-month-old mice, while invasivecarcinoma is detected in 7-month-old animals [10]. A series of celllines isolated from this model have been developed and thor-oughly characterized [11,12]. Pr-111 is a cell line representative ofa low-grade PIN lesion, whereas Pr-14 and its metastatic variantsPr-14C1 and Pr-14C2 resemble invasive carcinoma and distantmetastasis, respectively [11]. Pr-111 cells are very low tumori-genic, androgen receptor positive, and show low VEGF levels,unlike the other Pr-derived tumor cells [11,12]. These cellsconstitute a unique in vitro model to analyze prostate carcinogen-esis [11]. In the present study we have taken advantage of the PIN-like features of Pr-111 cells to analyze the autocrine andangiogenic effect of VEGF164.
Epithelial–mesenchymal transition (EMT) is a process ofdifferentiation that is crucial for adequate gastrulation in embryo-genesis and takes place in fibrogenic events, and carcinomainvasion [13,14]. Epithelial cells undergoing EMT show a reductionin both polarity and adhesive properties, and acquire fibroblast-like (spindle) morphology. EMT favors the acquisition of motility,invasion, dedifferentiation and metastasis [14]. The outcome ofEMT is the acquisition of a novel mesenchymal phenotype, withdown-regulation of epithelial markers, and concomitant up-regulation of markers such as vimentin and N-cadherin [14].
Transforming growth factor-β1 (TGF-β1) has been shown tocause EMT and stimulate cell migration in different epithelial celllines [15]. TGF-β1 is a potent growth inhibitor in most epithelialcells [16]. In later stages of malignant progression, however, thereis a loss of the growth inhibitory response to TGF-β1 which theninduces EMT [17]. Other factors, including the basic fibroblastgrowth factor (bFGF), epidermal growth factor (EGF), hepatocytegrowth factor (HGF), and Wnt ligands are also able to induce EMT[14]. Recent studies have demonstrated that VEGF produces EMTfeatures in pancreatic cancer cells [18]. Since the angiogenic switchis characterized by an abrupt rise in VEGF expression, wehypothesized that such increase would contribute to the acquisi-
tion of mesenchymal properties by the preinvasivemalignant cells.This could foster the escape of such cells from the epithelialcompartment to the stroma through increased migration. Wedemonstrate here that VEGF enhances the angiogenic andtumorigenic capacity of Pr-111 cells and causes transcriptomicchanges leading to the acquisition of EMT features and increasedmotility. Therefore, VEGF orchestrates non-angiogenic events thatplay a key role in early tumor spread.
Materials and methods
Cell culture
The C3(1)/Tag Pr cancer cells have previously been isolated andcharacterized [11,12]. Pr-111 cells were grown in MEGM medium(Calbiochem, Walkersville, MD) supplemented with 2% fetal calfserum (FCS), in collagen PureCol™-coated plates (Nutacon BV,Leimuiden, Holland). GP-293 cells were obtained from ATCC andgrown in DMEMwith 10% FCS. VEGF164 was added at 10 ng/mL (Rand D Systems, MN) for the indicated times. TGF-β1 (R and DSystems, MN) was added at 10 pM for the indicated times. Bothtreatments were conducted in RPMI containing 1% FCS in 6-wellplates.
Establishment of cell clones overexpressing VEGF
The murine isoform VEGF164 was cloned into the retroviral vectorPCLNEX, which was kindly donated by Dr. S. Libutti (TumorAngiogenesis Section, National Cancer Institute, NIH, USA). 106
cells were plated into 6-well plates and cultured in DMEM withblasticidin (20 μg/mL). GP-293 cells were transfected with Fugene6 (Roche, Basel, Switzerland)with 1 μg of either the empty PCLNEXplasmid or the plasmid carrying VEGF164, plus 1 μg of the MDGenvelope plasmid. The next day, culture media were replaced withfresh media. On days 3 and 4 after transfection, conditioned mediawere collected, filtered through a 0.45 μm filters and used forretroviral infection of Pr-111 cells. Transductions were carried outin 6-well plates after seeding 105 Pr-111 cells. Cell medium wasreplaced by 1 mL of supernatant plus 8 μg/mL polybrene (Sigma,St. Louis, MO), incubated for 4 h at 37 C, and then replaced withMGEM medium. The procedure was repeated for two subsequentdays, after which cells were treated with G-418 (300 μg/mL) for15 days. Nine G-418-resistant clones carrying the VEGF-containingplasmid and 5 clones carrying the empty vector were selected,cultured into new plates and checked for VEGF expression in theconditioned medium, using an ELISA method (R and D Systems,MN). Three cloneswere used for further assays: One clone carryingthe empty vector with similar VEGF levels to those of the parentalcells (called hereafterMock), and two clones carrying the VEGF164-containing plasmid (hereafter referred as to clone 1 and clone 2),both with high expression of VEGF.
Cell proliferation assay
7000 cells were seeded into 96-well plates in presence of 2% FCS-containing MEGM medium and let grow for several time points.Cell proliferation rates were determined with the MTT CellProliferation Kit I (Roche) and measured at 540/690 nm with aSunRise (Tecan, Salzburg, Austria) ELISA plate reader.

Table 1 – List of primers used for real time RT-PCRs.
Gene Primer Primer sequence (5′–3′)
mVEGF164 mVEGF164 S ACAGGACAAAGCCAGAAAAACmVEGF164 AS GTTTAACTCAAGCTGCCTCGCC
mVEGF mVEGF S GCGGATCAAACCTCACCAAAmVEGF AS TTCACATCGGCTGTGCTGTAG
mFlt1 mFlt1 S GCCCTGCTGTGGTCTCACTACmFlt1 AS CAAAGCATTGCCCATTCGAT
mKdr mKdr S GAGGAGGATGAGGGTGTCTATAGGTmKdr AS GTGATCAGCTCCAGGTTTGACT
mGAPDH mGAPDH S ACTTTGTCAAGCTCATTTCCmGAPDH AS TGCAGCGAACTTTATTGATG
mSnail1 mSnail1 S CTTCTCTAGGCCCTGGCTGCTTmSnail1 AS CAGACTCTTGGTGCTTGTGG
mSnail2 mSnail2 S AAGCCCAACTACAGCGAACTGGmSnail2 AS ACTTACACGCCCCAAGGATGAG
mSmad7 mSmad7 S TCGGACAGCTAATTCGGACmSmad7 AS GGTAACTGCTGCGGTTGTAA
mE-cadherin mE-cadherin S CACAGACGCGGACGATGATmE-cadherin AS GATCTTGGCTGAGGATGGTGTAA
mVimentin mVimentin S TTCTCTGCCTCTTCCAAACTTTTCmVimentin AS GGGTATCAACCAGAGGGAGTGA
556 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
Anchorage-independent cell growth assay
Soft agar experiments were performed in six-well plates using5×104 cells/mL per well. The bottom layer contained 2mL of 0.6%noble agar (BD, San Jose, CA), penicillin (100 U/mL), streptomycin(100 mg/mL), 2% FBS and 300 μg/mL G-418 in RPMI 1640medium. The top layer contained 1 mL of 0.3% noble agar,penicillin (100 U/ mL), streptomycin (100 mg/ mL), 2% FBS,300 μg/ mL G-418 and 5×104 cells. Parental Pr-111 cells werecultured in the same conditions but without G-418. Plates wereincubated for 2 weeks at 37 °C with 5% CO2 to allow cellproliferation in soft agar. The presence of cell colonies wasrevealed by adding 10 mg/ mL MTT (500 μL) for 16 h, and 1 mLDMSO. Images of the agar plates were taken with an hp scanjet4600 scanner and cell colony number was counted for each well.Assays were repeated at least 3 times, and results are shown asthe average of colony number.
Migration assay
Pr-111 stable transfectants (Mock, clone 1 and clone 2)were platedin the upper part of Boyden chambers (BD Biosciences) at a densityof 5×104 cells in serum free RPMI medium, in 24-well plates. Thelower part of the chamber contained 20% serum in RPMI. Cells werelet migrate for 24 h, washed and fixed in 4% formaldehyde. Nonmigrated cells from the upper part of the chamber were removedwith a cotton scrub; migrated cells were stained with 1% crystalviolet. The cell-containing membrane was cut from the insert andmounted on slides with PBS-Glycerol. Four randomized imageswere taken using a Nikon Y-THS microscope (Nikon, Japan)equipped with the ACT-20 software, and the number of migratedcells was counted.
Real time RT-PCR
Total RNA from cells was isolated using the RNAeasy Mini Kit(Qiagen, Valencia, CA), according to the manufacturer's protocol.The quality of RNA was determined by running aliquots through1% agarose gels. Two μg DNAse-treated RNA was reversedtranscribed into cDNA using the First Strand SuperScript II RT kit(Invitrogen). PCR primer sequences for mouse (m) total VEGF,VEGF164, Flt1, KDR [19], E-cadherin, vimentin, Smad7, Snail1, andSnail2 is shown in Table 1. Melting curves and gel analyses wereperformed to confirm the presence of single amplified species.Quantitative analysis of gene expression was carried out usingSYBR Green Core Reagents (PE Biosystems, Warrington, UK) usingthe Applied Biosystems 7300 real-time detection system. Geneexpression levels were calculated after normalizing the samples toGAPDH levels.
Microarray analysis and gene expression profiles
Microarray analysis was used to profile gene expression patternsafter VEGF overexpression in Pr-111 cells. The cDNA from theparental Pr-111 cells was used as the reference sample againstwhich cDNA from Mock 1, Mock 2, and VEGF-overexpressing cellclones 1, 2, 3, 6, and 7 was competitively hybridized. For thispurpose, total RNA from cultured cells was isolated as describedabove and cDNA-labeled probes were prepared with the MICRO-MAX™ system (NEN Life Science Products, Boston, MA) as
previously described [11]. Hybridizations were carried out on theIncyte Genomics mouse GEM1 set of cDNA clones, spotted at theNational Cancer Institute Advanced Technology Center. The genelist is available at http://nciarray.nci.nih.gov. LIMMA (LinearModels for Microarray Data) [20] was used to find out the genesthat showed significant differential expression between differentPr-111 clones with VEGF overexpression (clones 1, 2, 3, 6, and 7)and controls (Mock 1 and Mock 2). Genes were selected assignificant using a criteria of false discovery rate FDR [20] <5%.Biological interpretation of the selected genes was carried out byGene Ontology (GO) enrichment analysis using the GARBANsoftware (Genomic Analysis for Rapid Biological Annotation,http://garban.tecnun.es/garban2/index.php) [21]. Signaling path-wayswere analyzedwith Ingenuity™ software (Ingenuity Systems,Redwood City, CA).
In order to analyze transcription factors that might be activatedin Pr-111-overexpressing cells, several bioinformatic resourceswere used. Proximal promoter sequences of the murine genomewere taken from the EnsMart database and position weightmatrixes of known transcription factor binding sites (TFBS) fromthe public version of Jaspar [22] and Transfac [23] databases.Detection of known TFBSs in the proximal promoters wasperformed with MotifScanner. Both promoter sequences anddetected TFBS were stored in a MySQL database. The FactorYsoftware [24] was used to access the database and to compare theTFBS distribution in our set of selected genes with the distributionin the murine genome. Finally, a TFBS enrichment p-value wascalculated using the hypergeometric distribution. Multiple hy-pothesis correction was performance and FDR lower than 0.05were considered statistically significant.
Western blot
Total proteins were obtained from parental cells and stabletransfectants (Mock, clone 1 and clone 2) in RIPA buffer (50 mMTris-HCl pH 7.4, 150 mM NaCl, 1% NP-40, 0.25% Na-deoxycholate,and 1 mM EDTA), supplemented with 1 mM PMSF, 1 mM Na-
557E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
orthovanadate, 1 mM NaF, and proteases inhibitor cocktail(Sigma). Nuclear/cytoplasmic proteins were isolated using theNE-PER kit (Pierce, Rockford, IL), followingmanufacturer's instruc-tions. Protein concentration for each sample was determined usingQuick Start Bradford Dye Reagent (Bio-Rad, Hercules, CA), andequal amounts of proteins were loaded in 10% SDS-PAGE (10%acrylamide). Blocking buffer included 1% BSA/1% milk in TBS and0.1% Tween, and 20 mM NaF. The following antibodies were used:VEGF (Santa Cruz, Temecula, CA), Flt-1/VEGF-R1 (Neomarkers),VEGF-R2 (Santa Cruz), E-cadherin (BD Transduction Laboratories),N-cadherin (Invitrogen), Vimentin (Biocare Medical, Concord, CA)
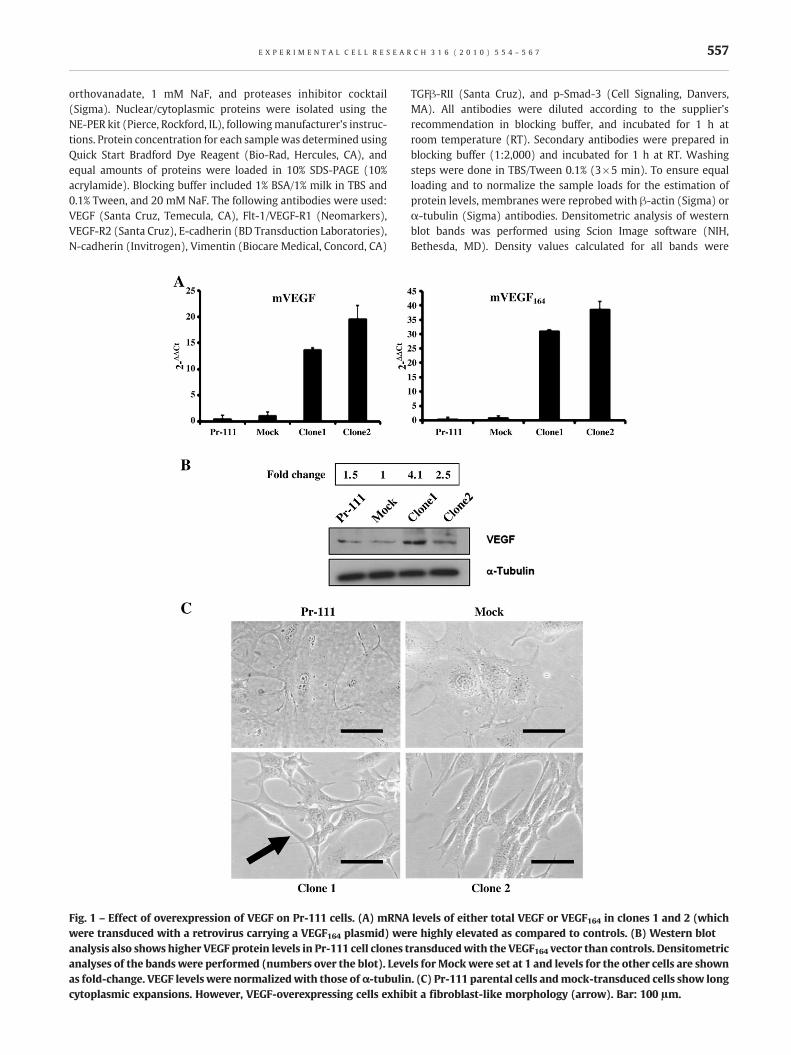
Fig. 1 – Effect of overexpression of VEGF on Pr-111 cells. (A) mRNAwere transduced with a retrovirus carrying a VEGF164 plasmid) weanalysis also shows higher VEGF protein levels in Pr-111 cell clones tanalyses of the bands were performed (numbers over the blot). Leveas fold-change. VEGF levels were normalized with those of α-tubulincytoplasmic expansions. However, VEGF-overexpressing cells exhib
TGFβ-RII (Santa Cruz), and p-Smad-3 (Cell Signaling, Danvers,MA). All antibodies were diluted according to the supplier'srecommendation in blocking buffer, and incubated for 1 h atroom temperature (RT). Secondary antibodies were prepared inblocking buffer (1:2,000) and incubated for 1 h at RT. Washingsteps were done in TBS/Tween 0.1% (3×5 min). To ensure equalloading and to normalize the sample loads for the estimation ofprotein levels, membranes were reprobed with β-actin (Sigma) orα-tubulin (Sigma) antibodies. Densitometric analysis of westernblot bands was performed using Scion Image software (NIH,Bethesda, MD). Density values calculated for all bands were
levels of either total VEGF or VEGF164 in clones 1 and 2 (whichre highly elevated as compared to controls. (B) Western blotransducedwith the VEGF164 vector than controls. Densitometricls for Mockwere set at 1 and levels for the other cells are shown. (C) Pr-111 parental cells andmock-transduced cells show longit a fibroblast-like morphology (arrow). Bar: 100 μm.
558 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
normalized to background signal of each blot. Intensity levels of thetarget protein were then normalized to those of the housekeepingprotein.
Angiogenesis Matrigel plug assay
Pr-111 and the cell clones (0.5×106 cells for each mouse, in0.2 mL final volume) were subcutaneously injected with Matrigel(BD Biosciences) into athymic nude mice (4-week-old males).Two weeks after cell implantation, intracardiac injection of200 μL 25 mg/mL FITC-dextran-500000-conjugate (Fluka) for15 min were conducted, and Matrigel plugs were collected andfixed in 4% formaldehyde for 24 h in darkness. Fluorescence wascaptured with a LSM 510 META confocal microscope (Zeiss,Germany).
In vivo tumor growth
Cells were subcutaneously injected with Matrigel (1×106 cells permouse) into athymic nude male mice (4 weeks old). Tumor sizewas measured twice a week with a calliper and volumes werecalculated using the following formula: Volume= (width2×length)/2. All mice were treated in accordance with the guidelinesfor the Animal Care Ethics Commission of our institution(University of Navarra) under an approved animal protocol.
Statistical analysis
An ANOVA test was performed to analyze differences betweengroups. The Bonferroni test was used for post-hoc comparisons. TheSPSS software was used for statistical comparisons. Results with ap-vale lower than 0.05 were considered statistically significant.
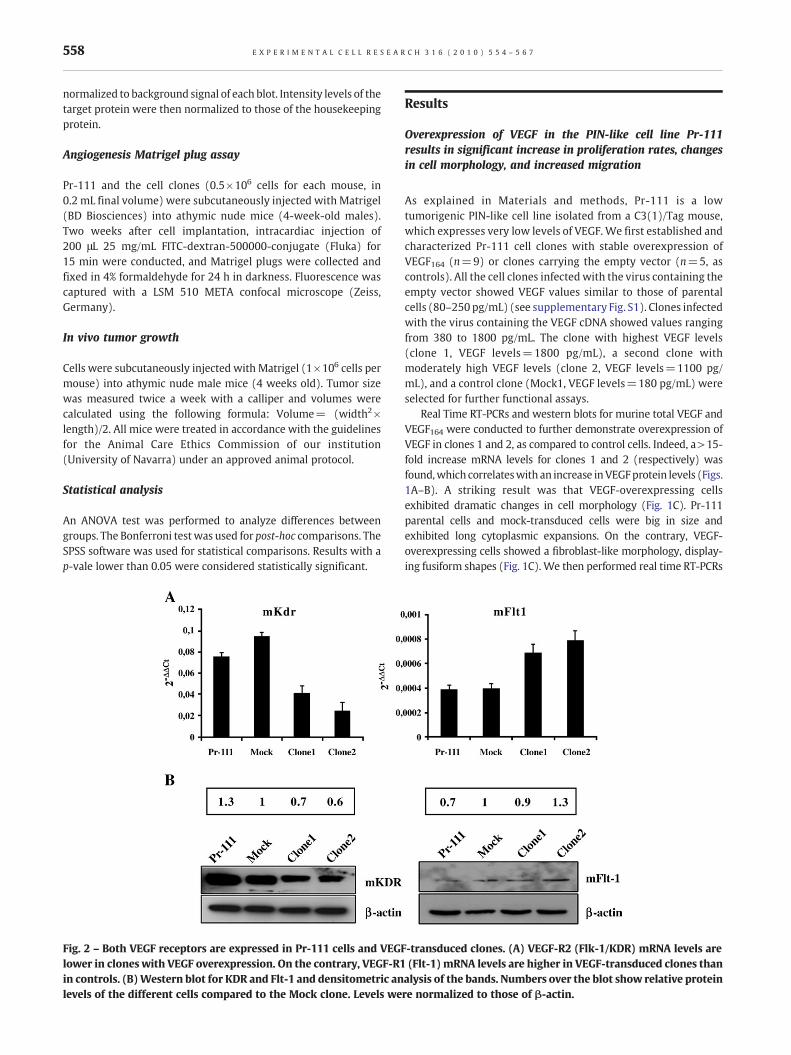
Fig. 2 – Both VEGF receptors are expressed in Pr-111 cells and VEGlower in clones with VEGF overexpression. On the contrary, VEGF-R1in controls. (B)Western blot for KDR and Flt-1 and densitometric anlevels of the different cells compared to the Mock clone. Levels we
Results
Overexpression of VEGF in the PIN-like cell line Pr-111results in significant increase in proliferation rates, changesin cell morphology, and increased migration
As explained in Materials and methods, Pr-111 is a lowtumorigenic PIN-like cell line isolated from a C3(1)/Tag mouse,which expresses very low levels of VEGF. We first established andcharacterized Pr-111 cell clones with stable overexpression ofVEGF164 (n=9) or clones carrying the empty vector (n=5, ascontrols). All the cell clones infected with the virus containing theempty vector showed VEGF values similar to those of parentalcells (80–250 pg/mL) (see supplementary Fig. S1). Clones infectedwith the virus containing the VEGF cDNA showed values rangingfrom 380 to 1800 pg/mL. The clone with highest VEGF levels(clone 1, VEGF levels=1800 pg/mL), a second clone withmoderately high VEGF levels (clone 2, VEGF levels=1100 pg/mL), and a control clone (Mock1, VEGF levels=180 pg/mL) wereselected for further functional assays.
Real Time RT-PCRs and western blots for murine total VEGF andVEGF164 were conducted to further demonstrate overexpression ofVEGF in clones 1 and 2, as compared to control cells. Indeed, a>15-fold increase mRNA levels for clones 1 and 2 (respectively) wasfound,which correlateswith an increase inVEGFprotein levels (Figs.1A–B). A striking result was that VEGF-overexpressing cellsexhibited dramatic changes in cell morphology (Fig. 1C). Pr-111parental cells and mock-transduced cells were big in size andexhibited long cytoplasmic expansions. On the contrary, VEGF-overexpressing cells showed a fibroblast-like morphology, display-ing fusiform shapes (Fig. 1C). We then performed real time RT-PCRs
F-transduced clones. (A) VEGF-R2 (Flk-1/KDR) mRNA levels are(Flt-1) mRNA levels are higher in VEGF-transduced clones thanalysis of the bands. Numbers over the blot show relative proteinre normalized to those of β-actin.
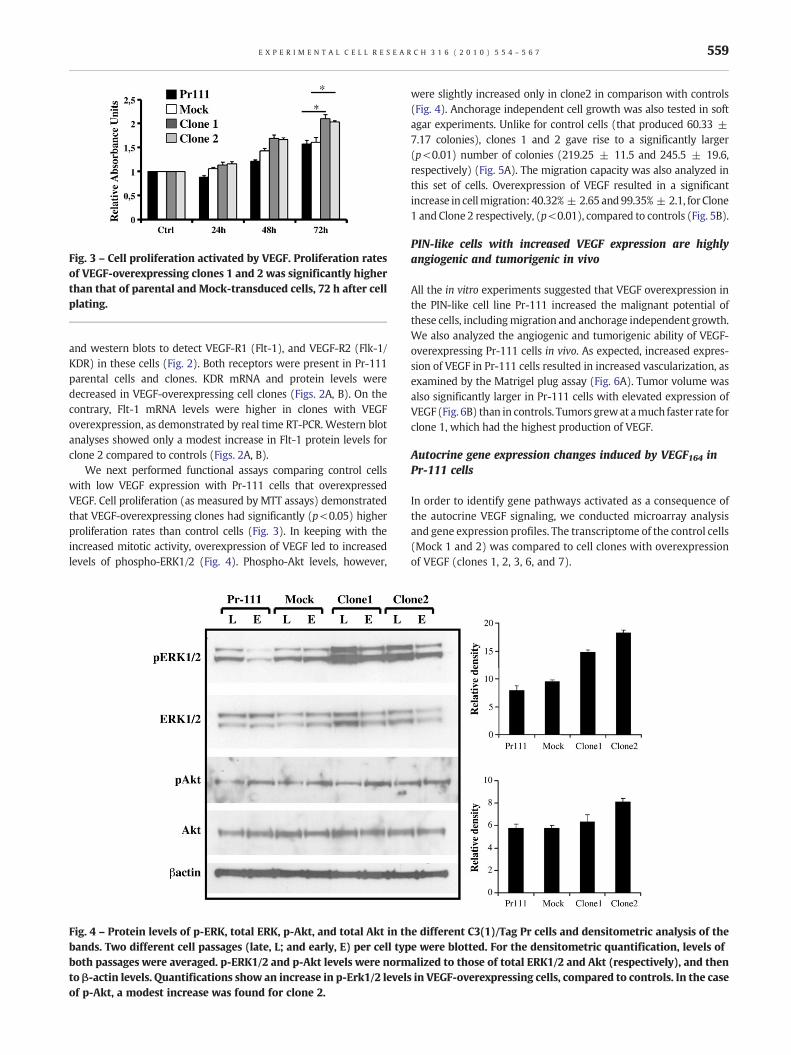
Fig. 3 – Cell proliferation activated by VEGF. Proliferation ratesof VEGF-overexpressing clones 1 and 2 was significantly higherthan that of parental and Mock-transduced cells, 72 h after cellplating.
559E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
and western blots to detect VEGF-R1 (Flt-1), and VEGF-R2 (Flk-1/KDR) in these cells (Fig. 2). Both receptors were present in Pr-111parental cells and clones. KDR mRNA and protein levels weredecreased in VEGF-overexpressing cell clones (Figs. 2A, B). On thecontrary, Flt-1 mRNA levels were higher in clones with VEGFoverexpression, as demonstrated by real time RT-PCR. Western blotanalyses showed only a modest increase in Flt-1 protein levels forclone 2 compared to controls (Figs. 2A, B).
We next performed functional assays comparing control cellswith low VEGF expression with Pr-111 cells that overexpressedVEGF. Cell proliferation (as measured by MTT assays) demonstratedthat VEGF-overexpressing clones had significantly (p<0.05) higherproliferation rates than control cells (Fig. 3). In keeping with theincreased mitotic activity, overexpression of VEGF led to increasedlevels of phospho-ERK1/2 (Fig. 4). Phospho-Akt levels, however,
Fig. 4 – Protein levels of p-ERK, total ERK, p-Akt, and total Akt in thbands. Two different cell passages (late, L; and early, E) per cell typboth passages were averaged. p-ERK1/2 and p-Akt levels were normto β-actin levels. Quantifications show an increase in p-Erk1/2 levelsof p-Akt, a modest increase was found for clone 2.
were slightly increased only in clone2 in comparison with controls(Fig. 4). Anchorage independent cell growth was also tested in softagar experiments. Unlike for control cells (that produced 60.33 ±7.17 colonies), clones 1 and 2 gave rise to a significantly larger(p<0.01) number of colonies (219.25 ± 11.5 and 245.5 ± 19.6,respectively) (Fig. 5A). The migration capacity was also analyzed inthis set of cells. Overexpression of VEGF resulted in a significantincrease in cellmigration:40.32% ± 2.65 and99.35% ± 2.1, for Clone1 and Clone 2 respectively, (p<0.01), compared to controls (Fig. 5B).
PIN-like cells with increased VEGF expression are highlyangiogenic and tumorigenic in vivo
All the in vitro experiments suggested that VEGF overexpression inthe PIN-like cell line Pr-111 increased the malignant potential ofthese cells, includingmigration and anchorage independent growth.We also analyzed the angiogenic and tumorigenic ability of VEGF-overexpressing Pr-111 cells in vivo. As expected, increased expres-sion of VEGF in Pr-111 cells resulted in increased vascularization, asexamined by the Matrigel plug assay (Fig. 6A). Tumor volume wasalso significantly larger in Pr-111 cells with elevated expression ofVEGF (Fig. 6B) than in controls. Tumors grewat amuch faster rate forclone 1, which had the highest production of VEGF.
Autocrine gene expression changes induced by VEGF164 inPr-111 cells
In order to identify gene pathways activated as a consequence ofthe autocrine VEGF signaling, we conducted microarray analysisand gene expression profiles. The transcriptome of the control cells(Mock 1 and 2) was compared to cell clones with overexpressionof VEGF (clones 1, 2, 3, 6, and 7).
e different C3(1)/Tag Pr cells and densitometric analysis of thee were blotted. For the densitometric quantification, levels ofalized to those of total ERK1/2 and Akt (respectively), and thenin VEGF-overexpressing cells, compared to controls. In the case
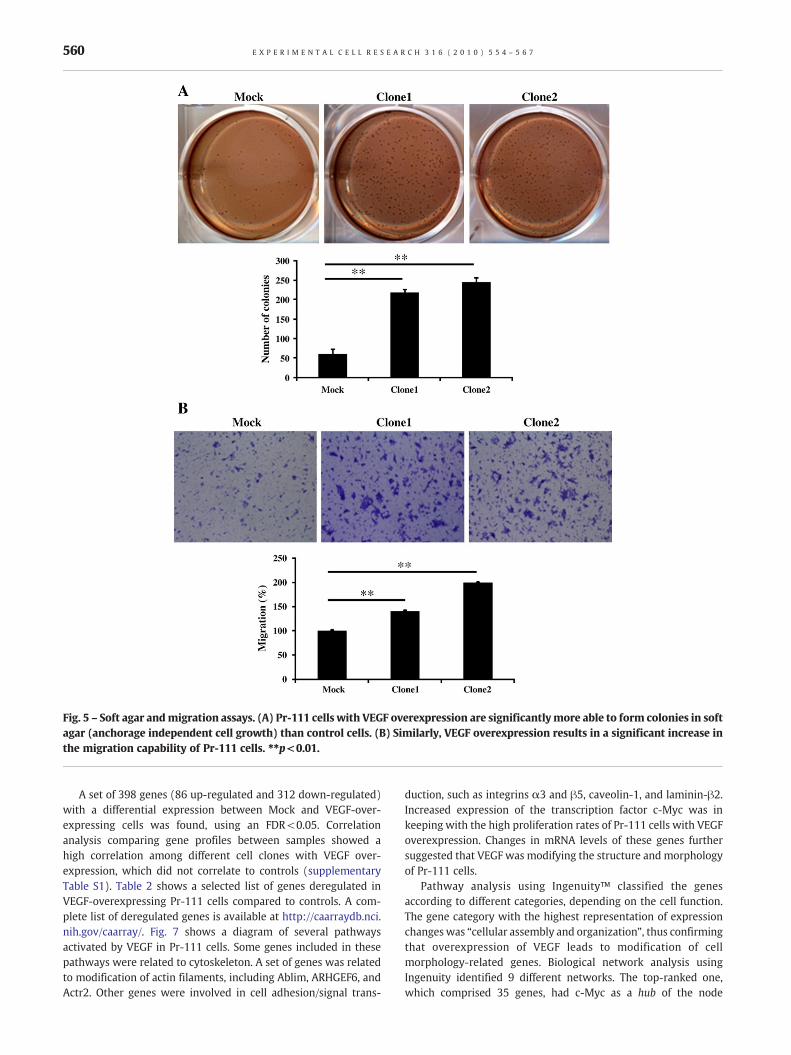
Fig. 5 – Soft agar andmigration assays. (A) Pr-111 cells with VEGF overexpression are significantlymore able to form colonies in softagar (anchorage independent cell growth) than control cells. (B) Similarly, VEGF overexpression results in a significant increase inthe migration capability of Pr-111 cells. ⁎⁎p<0.01.
560 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
A set of 398 genes (86 up-regulated and 312 down-regulated)with a differential expression between Mock and VEGF-over-expressing cells was found, using an FDR<0.05. Correlationanalysis comparing gene profiles between samples showed ahigh correlation among different cell clones with VEGF over-expression, which did not correlate to controls (supplementaryTable S1). Table 2 shows a selected list of genes deregulated inVEGF-overexpressing Pr-111 cells compared to controls. A com-plete list of deregulated genes is available at http://caarraydb.nci.nih.gov/caarray/. Fig. 7 shows a diagram of several pathwaysactivated by VEGF in Pr-111 cells. Some genes included in thesepathways were related to cytoskeleton. A set of genes was relatedto modification of actin filaments, including Ablim, ARHGEF6, andActr2. Other genes were involved in cell adhesion/signal trans-
duction, such as integrins α3 and β5, caveolin-1, and laminin-β2.Increased expression of the transcription factor c-Myc was inkeeping with the high proliferation rates of Pr-111 cells with VEGFoverexpression. Changes in mRNA levels of these genes furthersuggested that VEGF was modifying the structure and morphologyof Pr-111 cells.
Pathway analysis using Ingenuity™ classified the genesaccording to different categories, depending on the cell function.The gene category with the highest representation of expressionchangeswas “cellular assembly and organization”, thus confirmingthat overexpression of VEGF leads to modification of cellmorphology-related genes. Biological network analysis usingIngenuity identified 9 different networks. The top-ranked one,which comprised 35 genes, had c-Myc as a hub of the node
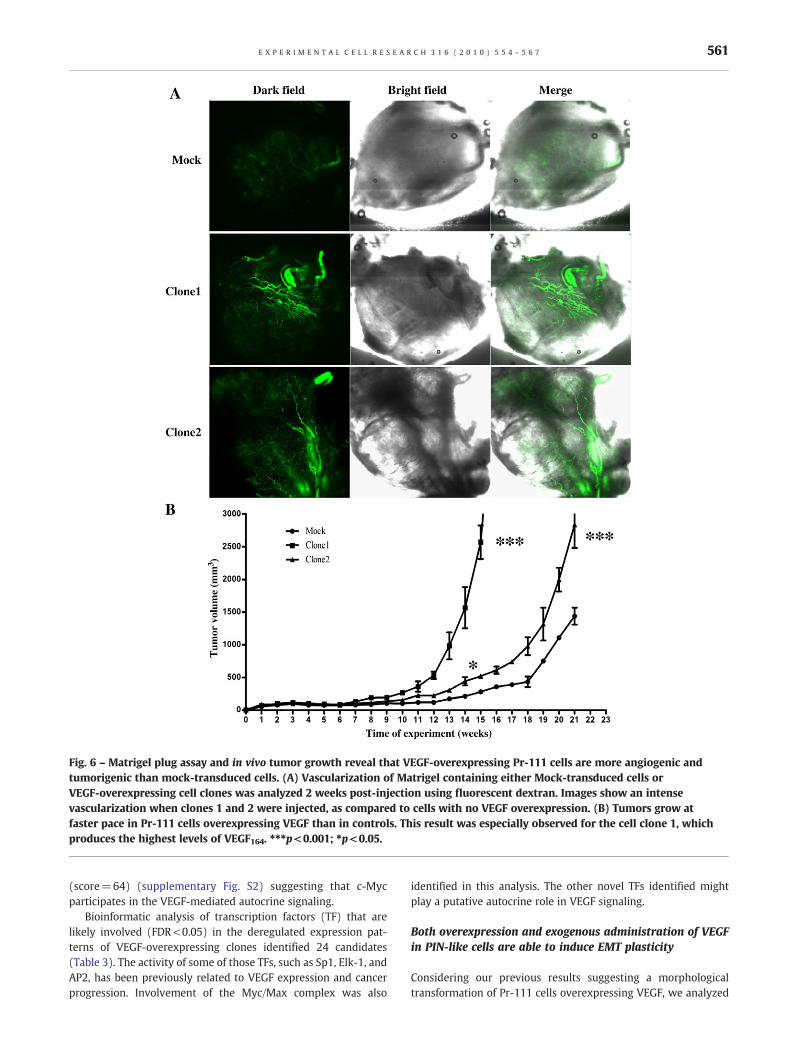
Fig. 6 – Matrigel plug assay and in vivo tumor growth reveal that VEGF-overexpressing Pr-111 cells are more angiogenic andtumorigenic than mock-transduced cells. (A) Vascularization of Matrigel containing either Mock-transduced cells orVEGF-overexpressing cell clones was analyzed 2 weeks post-injection using fluorescent dextran. Images show an intensevascularization when clones 1 and 2 were injected, as compared to cells with no VEGF overexpression. (B) Tumors grow atfaster pace in Pr-111 cells overexpressing VEGF than in controls. This result was especially observed for the cell clone 1, whichproduces the highest levels of VEGF164. ⁎⁎⁎p<0.001; ⁎p<0.05.
561E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
(score=64) (supplementary Fig. S2) suggesting that c-Mycparticipates in the VEGF-mediated autocrine signaling.
Bioinformatic analysis of transcription factors (TF) that arelikely involved (FDR<0.05) in the deregulated expression pat-terns of VEGF-overexpressing clones identified 24 candidates(Table 3). The activity of some of those TFs, such as Sp1, Elk-1, andAP2, has been previously related to VEGF expression and cancerprogression. Involvement of the Myc/Max complex was also
identified in this analysis. The other novel TFs identified mightplay a putative autocrine role in VEGF signaling.
Both overexpression and exogenous administration of VEGFin PIN-like cells are able to induce EMT plasticity
Considering our previous results suggesting a morphologicaltransformation of Pr-111 cells overexpressing VEGF, we analyzed
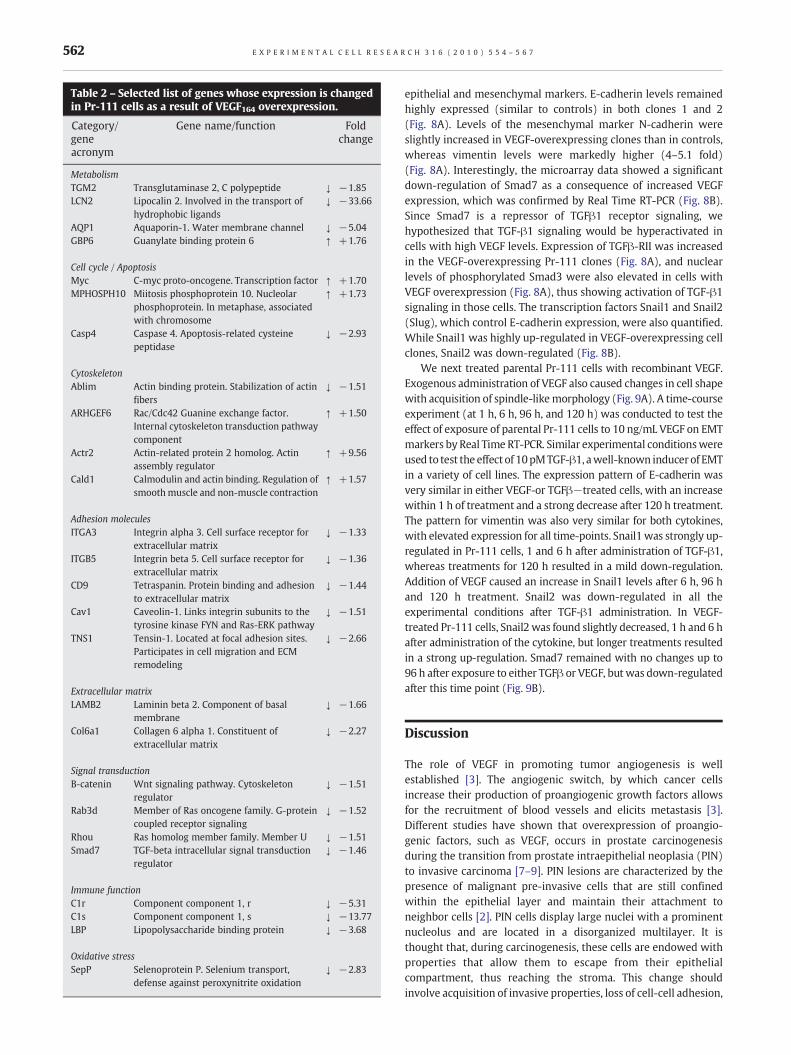
Table 2 – Selected list of genes whose expression is changedin Pr-111 cells as a result of VEGF164 overexpression.
Category/geneacronym
Gene name/function Foldchange
MetabolismTGM2 Transglutaminase 2, C polypeptide ↓ −1.85LCN2 Lipocalin 2. Involved in the transport of
hydrophobic ligands↓ −33.66
AQP1 Aquaporin-1. Water membrane channel ↓ −5.04GBP6 Guanylate binding protein 6 ↑ +1.76
Cell cycle / ApoptosisMyc C-myc proto-oncogene. Transcription factor ↑ +1.70MPHOSPH10 Miitosis phosphoprotein 10. Nucleolar
phosphoprotein. In metaphase, associatedwith chromosome
↑ +1.73
Casp4 Caspase 4. Apoptosis-related cysteinepeptidase
↓ −2.93
CytoskeletonAblim Actin binding protein. Stabilization of actin
fibers↓ −1.51
ARHGEF6 Rac/Cdc42 Guanine exchange factor.Internal cytoskeleton transduction pathwaycomponent
↑ +1.50
Actr2 Actin-related protein 2 homolog. Actinassembly regulator
↑ +9.56
Cald1 Calmodulin and actin binding. Regulation ofsmoothmuscle and non-muscle contraction
↑ +1.57
Adhesion moleculesITGA3 Integrin alpha 3. Cell surface receptor for
extracellular matrix↓ −1.33
ITGB5 Integrin beta 5. Cell surface receptor forextracellular matrix
↓ −1.36
CD9 Tetraspanin. Protein binding and adhesionto extracellular matrix
↓ −1.44
Cav1 Caveolin-1. Links integrin subunits to thetyrosine kinase FYN and Ras-ERK pathway
↓ −1.51
TNS1 Tensin-1. Located at focal adhesion sites.Participates in cell migration and ECMremodeling
↓ −2.66
Extracellular matrixLAMB2 Laminin beta 2. Component of basal
membrane↓ −1.66
Col6a1 Collagen 6 alpha 1. Constituent ofextracellular matrix
↓ −2.27
Signal transductionB-catenin Wnt signaling pathway. Cytoskeleton
regulator↓ −1.51
Rab3d Member of Ras oncogene family. G-proteincoupled receptor signaling
↓ −1.52
Rhou Ras homolog member family. Member U ↓ −1.51Smad7 TGF-beta intracellular signal transduction
regulator↓ −1.46
Immune functionC1r Component component 1, r ↓ −5.31C1s Component component 1, s ↓ −13.77LBP Lipopolysaccharide binding protein ↓ −3.68
Oxidative stressSepP Selenoprotein P. Selenium transport,
defense against peroxynitrite oxidation↓ −2.83
562 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
epithelial and mesenchymal markers. E-cadherin levels remainedhighly expressed (similar to controls) in both clones 1 and 2(Fig. 8A). Levels of the mesenchymal marker N-cadherin wereslightly increased in VEGF-overexpressing clones than in controls,whereas vimentin levels were markedly higher (4–5.1 fold)(Fig. 8A). Interestingly, the microarray data showed a significantdown-regulation of Smad7 as a consequence of increased VEGFexpression, which was confirmed by Real Time RT-PCR (Fig. 8B).Since Smad7 is a repressor of TGFβ1 receptor signaling, wehypothesized that TGF-β1 signaling would be hyperactivated incells with high VEGF levels. Expression of TGFβ-RII was increasedin the VEGF-overexpressing Pr-111 clones (Fig. 8A), and nuclearlevels of phosphorylated Smad3 were also elevated in cells withVEGF overexpression (Fig. 8A), thus showing activation of TGF-β1signaling in those cells. The transcription factors Snail1 and Snail2(Slug), which control E-cadherin expression, were also quantified.While Snail1 was highly up-regulated in VEGF-overexpressing cellclones, Snail2 was down-regulated (Fig. 8B).
We next treated parental Pr-111 cells with recombinant VEGF.Exogenous administration of VEGF also caused changes in cell shapewith acquisition of spindle-likemorphology (Fig. 9A). A time-courseexperiment (at 1 h, 6 h, 96 h, and 120 h) was conducted to test theeffect of exposure of parental Pr-111 cells to 10 ng/mL VEGF on EMTmarkers by Real Time RT-PCR. Similar experimental conditionswereused to test the effect of 10pMTGF-β1, awell-known inducer of EMTin a variety of cell lines. The expression pattern of E-cadherin wasvery similar in either VEGF-or TGFβ−treated cells, with an increasewithin 1 h of treatment and a strong decrease after 120 h treatment.The pattern for vimentin was also very similar for both cytokines,with elevated expression for all time-points. Snail1was strongly up-regulated in Pr-111 cells, 1 and 6 h after administration of TGF-β1,whereas treatments for 120 h resulted in a mild down-regulation.Addition of VEGF caused an increase in Snail1 levels after 6 h, 96 hand 120 h treatment. Snail2 was down-regulated in all theexperimental conditions after TGF-β1 administration. In VEGF-treated Pr-111 cells, Snail2 was found slightly decreased, 1 h and 6 hafter administration of the cytokine, but longer treatments resultedin a strong up-regulation. Smad7 remained with no changes up to96 h after exposure to either TGFβ or VEGF, but was down-regulatedafter this time point (Fig. 9B).
Discussion
The role of VEGF in promoting tumor angiogenesis is wellestablished [3]. The angiogenic switch, by which cancer cellsincrease their production of proangiogenic growth factors allowsfor the recruitment of blood vessels and elicits metastasis [3].Different studies have shown that overexpression of proangio-genic factors, such as VEGF, occurs in prostate carcinogenesisduring the transition from prostate intraepithelial neoplasia (PIN)to invasive carcinoma [7–9]. PIN lesions are characterized by thepresence of malignant pre-invasive cells that are still confinedwithin the epithelial layer and maintain their attachment toneighbor cells [2]. PIN cells display large nuclei with a prominentnucleolus and are located in a disorganized multilayer. It isthought that, during carcinogenesis, these cells are endowed withproperties that allow them to escape from their epithelialcompartment, thus reaching the stroma. This change shouldinvolve acquisition of invasive properties, loss of cell-cell adhesion,

Fig. 7 – Example of signaling pathways altered by VEGF in Pr-111 cells. Expression of cytoskeletal genes, such as Ablim, ARHGEF6, andActr2 are modified by VEGF. Changes also include Smad7 down-regulation and increased expression of c-Myc. Arrows pointing upfor a given gene mean up-regulation, whereas arrows pointing down, down-regulation.
Table 3 – Enrichment analysis of TFBSs performed withFactorY (FDR<0.05) for the 398 deregulated genes foundin Pr-111 cells with VEGF overexpression. N is the number ofgenes in the mouse genome with a predicted binding site inits promoter region, and n is the number of deregulatedgenes with a predicted binding site.
Motif name N n p-value FDR
SP1 10649 181 2.40E-10 0GC 11466 186 6.97E-09 0USF 5273 91 3.82E-05 0.004Elk-1 5750 97 4.92E-05 0.004Dorsal_1 2936 57 6.14E-05 0.004MZF1 6482 105 1.18E-04 0.007SPI-1 6275 102 1.34E-04 0.007MINI19 5458 91 1.40E-04 0.007E74A 6146 100 1.54E-04 0.007ATF 3390 62 1.64E-04 0.006NRF2 5221 87 2.08E-04 0.007MZF_5-13 7655 119 2.09E-04 0,007MYCMAX 6239 97 1.02E-03 0.029Bsap 4467 73 1.26E-03 0.034AP2 4839 78 1.27E-03 0.032GBP 4043 67 1.41E-03 0.033MZF_1-4 5765 90 1.42E-03 0.032CREB 3254 56 1.47E-03 0.031c-ETS 3686 62 1.48E-03 0.03GABP 4375 71 1.78E-03 0.034NF-Y 5515 86 1.92E-03 0.036ELK1 5410 84 2.47E-03 0.044AP4 4515 72 2.61E-03 0.045PAX5 3339 56 2.62E-03 0.043
563E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
and expression of collagenases to disrupt the basal membrane andmove through the surrounding stroma [2]. However, the precisemolecular mechanisms of this process are still elusive.
In the present study we have analyzed phenotypic and geneticchanges caused by VEGF that lead PIN-like cells to acquire EMTfeatures and become invasive, through an autocrine loop. We haveused a unique in vitro system consisting of a well established andcharacterized cell line (Pr-111) that was isolated from a C3(1)/Tag-derived PIN lesion [11,12]. These cells express low levels ofVEGF and are poorly invasive and tumorigenic [11]. As expected,overexpression of VEGF164 (the main VEGF isoform present inmouse [4]) in Pr-111 cells leads to an increased vascular andtumorigenic ability of these cells. Strikingly, increased expressionof VEGF also shifts the phenotype of these cells to a highly invasivefibroblastoid-like stage. We have shown that Pr-111 cells expressboth VEGF receptors VEGF-R1 (Flt-1) and VEGF-R2 (Flk-1/KDR)and respond to VEGF with the following effects: (a) an increase incell proliferation and activation of ERK-1/2 signaling pathways; (b)by activating a genetic program that involves changes in numerousgenes related to cytoskeleton remodeling, adhesion, signaltransduction and metabolism; (c) by acquiring a mesenchymal-like morphology and expressing EMT markers.
Our microarray analysis identified a set of genes that wasrelated to modification of the actin filaments, including Ablim,ARHGEF6, and Actr2. A group of adhesion-related genes (integrinsα3 and β5, caveolin-1, and laminin-β2 was down-regulated in Pr-111 cells overexpressing VEGF. These changes suggested modifi-cation of the cellular architecture, which are in keeping with theirincreased invasive potential. Bioinformatic classification ofderegulated genes analyzed by Ingenuity showed that the genecategory with the highest representation was “cellular assemblyand organization”. All these data, together with the profound
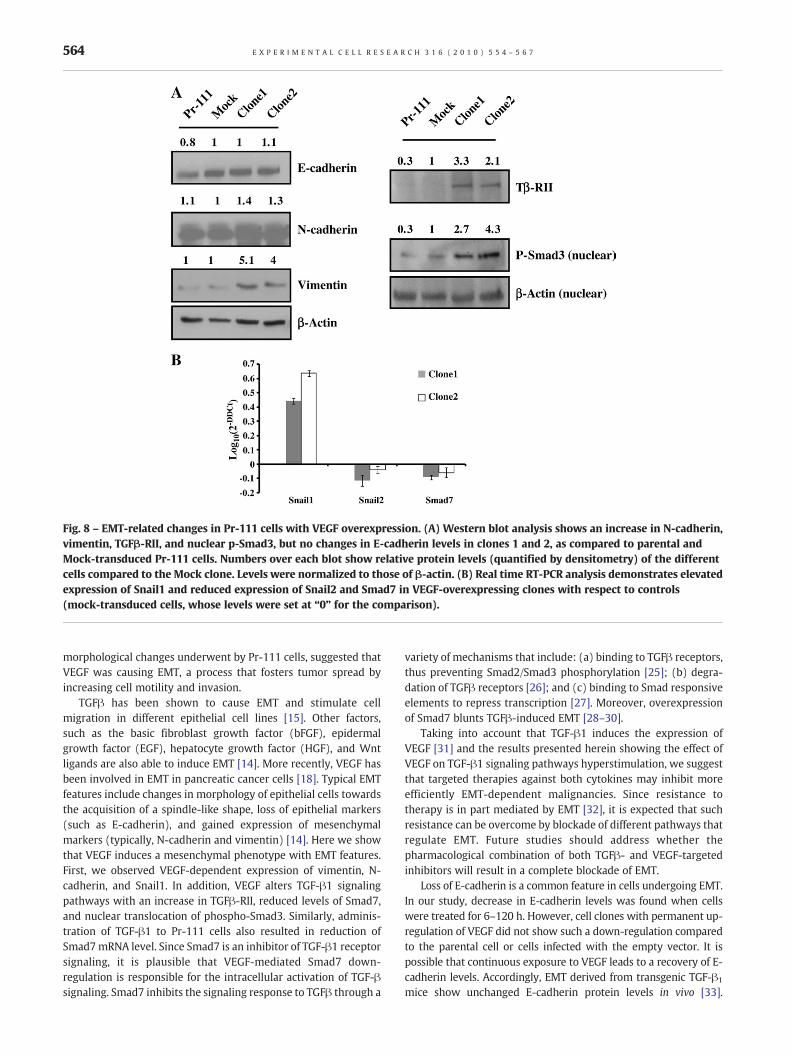
Fig. 8 – EMT-related changes in Pr-111 cells with VEGF overexpression. (A) Western blot analysis shows an increase in N-cadherin,vimentin, TGFβ-RII, and nuclear p-Smad3, but no changes in E-cadherin levels in clones 1 and 2, as compared to parental andMock-transduced Pr-111 cells. Numbers over each blot show relative protein levels (quantified by densitometry) of the differentcells compared to the Mock clone. Levels were normalized to those of β-actin. (B) Real time RT-PCR analysis demonstrates elevatedexpression of Snail1 and reduced expression of Snail2 and Smad7 in VEGF-overexpressing clones with respect to controls(mock-transduced cells, whose levels were set at “0” for the comparison).
564 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
morphological changes underwent by Pr-111 cells, suggested thatVEGF was causing EMT, a process that fosters tumor spread byincreasing cell motility and invasion.
TGFβ has been shown to cause EMT and stimulate cellmigration in different epithelial cell lines [15]. Other factors,such as the basic fibroblast growth factor (bFGF), epidermalgrowth factor (EGF), hepatocyte growth factor (HGF), and Wntligands are also able to induce EMT [14]. More recently, VEGF hasbeen involved in EMT in pancreatic cancer cells [18]. Typical EMTfeatures include changes in morphology of epithelial cells towardsthe acquisition of a spindle-like shape, loss of epithelial markers(such as E-cadherin), and gained expression of mesenchymalmarkers (typically, N-cadherin and vimentin) [14]. Here we showthat VEGF induces a mesenchymal phenotype with EMT features.First, we observed VEGF-dependent expression of vimentin, N-cadherin, and Snail1. In addition, VEGF alters TGF-β1 signalingpathways with an increase in TGFβ-RII, reduced levels of Smad7,and nuclear translocation of phospho-Smad3. Similarly, adminis-tration of TGF-β1 to Pr-111 cells also resulted in reduction ofSmad7mRNA level. Since Smad7 is an inhibitor of TGF-β1 receptorsignaling, it is plausible that VEGF-mediated Smad7 down-regulation is responsible for the intracellular activation of TGF-βsignaling. Smad7 inhibits the signaling response to TGFβ through a
variety of mechanisms that include: (a) binding to TGFβ receptors,thus preventing Smad2/Smad3 phosphorylation [25]; (b) degra-dation of TGFβ receptors [26]; and (c) binding to Smad responsiveelements to repress transcription [27]. Moreover, overexpressionof Smad7 blunts TGFβ-induced EMT [28–30].
Taking into account that TGF-β1 induces the expression ofVEGF [31] and the results presented herein showing the effect ofVEGF on TGF-β1 signaling pathways hyperstimulation, we suggestthat targeted therapies against both cytokines may inhibit moreefficiently EMT-dependent malignancies. Since resistance totherapy is in part mediated by EMT [32], it is expected that suchresistance can be overcome by blockade of different pathways thatregulate EMT. Future studies should address whether thepharmacological combination of both TGFβ- and VEGF-targetedinhibitors will result in a complete blockade of EMT.
Loss of E-cadherin is a common feature in cells undergoing EMT.In our study, decrease in E-cadherin levels was found when cellswere treated for 6–120 h. However, cell clones with permanent up-regulation of VEGF did not show such a down-regulation comparedto the parental cell or cells infected with the empty vector. It ispossible that continuous exposure to VEGF leads to a recovery of E-cadherin levels. Accordingly, EMT derived from transgenic TGF-β1
mice show unchanged E-cadherin protein levels in vivo [33].
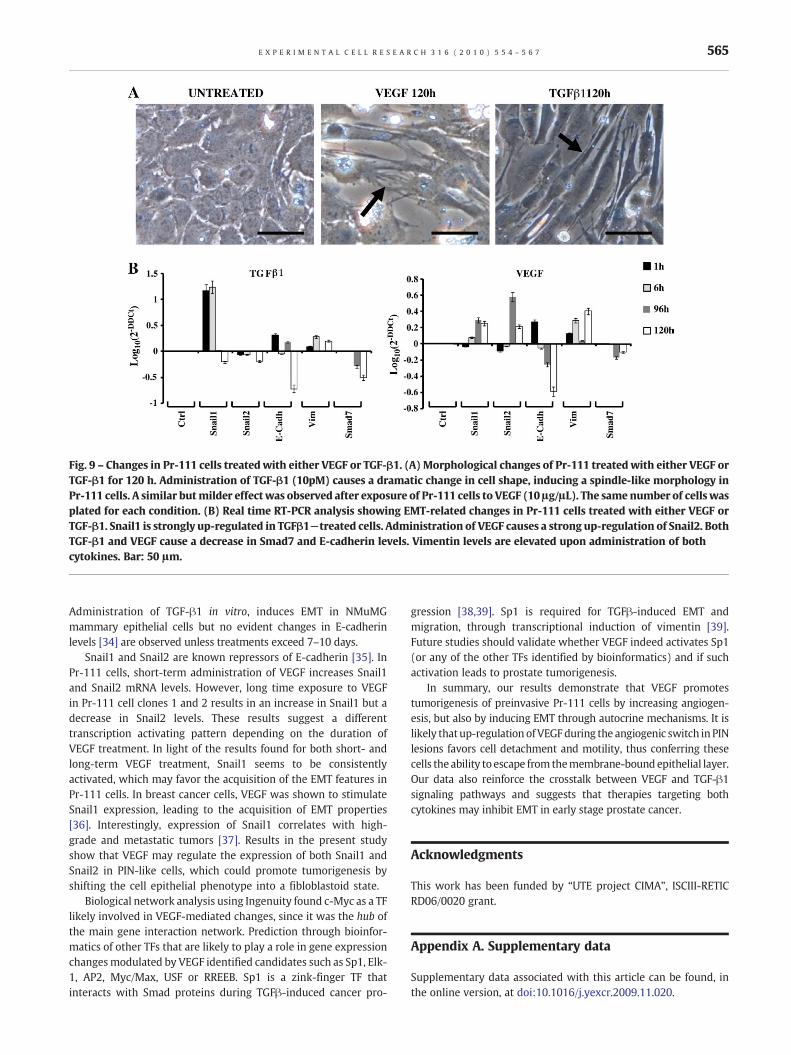
Fig. 9 – Changes in Pr-111 cells treated with either VEGF or TGF-β1. (A) Morphological changes of Pr-111 treated with either VEGF orTGF-β1 for 120 h. Administration of TGF-β1 (10pM) causes a dramatic change in cell shape, inducing a spindle-like morphology inPr-111 cells. A similar butmilder effectwas observed after exposure of Pr-111 cells to VEGF (10 μg/μL). The same number of cells wasplated for each condition. (B) Real time RT-PCR analysis showing EMT-related changes in Pr-111 cells treated with either VEGF orTGF-β1. Snail1 is strongly up-regulated in TGFβ1−treated cells. Administration of VEGF causes a strong up-regulation of Snail2. BothTGF-β1 and VEGF cause a decrease in Smad7 and E-cadherin levels. Vimentin levels are elevated upon administration of bothcytokines. Bar: 50 μm.
565E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
Administration of TGF-β1 in vitro, induces EMT in NMuMGmammary epithelial cells but no evident changes in E-cadherinlevels [34] are observed unless treatments exceed 7–10 days.
Snail1 and Snail2 are known repressors of E-cadherin [35]. InPr-111 cells, short-term administration of VEGF increases Snail1and Snail2 mRNA levels. However, long time exposure to VEGFin Pr-111 cell clones 1 and 2 results in an increase in Snail1 but adecrease in Snail2 levels. These results suggest a differenttranscription activating pattern depending on the duration ofVEGF treatment. In light of the results found for both short- andlong-term VEGF treatment, Snail1 seems to be consistentlyactivated, which may favor the acquisition of the EMT features inPr-111 cells. In breast cancer cells, VEGF was shown to stimulateSnail1 expression, leading to the acquisition of EMT properties[36]. Interestingly, expression of Snail1 correlates with high-grade and metastatic tumors [37]. Results in the present studyshow that VEGF may regulate the expression of both Snail1 andSnail2 in PIN-like cells, which could promote tumorigenesis byshifting the cell epithelial phenotype into a fibloblastoid state.
Biological network analysis using Ingenuity found c-Myc as a TFlikely involved in VEGF-mediated changes, since it was the hub ofthe main gene interaction network. Prediction through bioinfor-matics of other TFs that are likely to play a role in gene expressionchangesmodulated by VEGF identified candidates such as Sp1, Elk-1, AP2, Myc/Max, USF or RREEB. Sp1 is a zink-finger TF thatinteracts with Smad proteins during TGFβ-induced cancer pro-
gression [38,39]. Sp1 is required for TGFβ-induced EMT andmigration, through transcriptional induction of vimentin [39].Future studies should validate whether VEGF indeed activates Sp1(or any of the other TFs identified by bioinformatics) and if suchactivation leads to prostate tumorigenesis.
In summary, our results demonstrate that VEGF promotestumorigenesis of preinvasive Pr-111 cells by increasing angiogen-esis, but also by inducing EMT through autocrine mechanisms. It islikely that up-regulation ofVEGF during the angiogenic switch in PINlesions favors cell detachment and motility, thus conferring thesecells the ability to escape from themembrane-bound epithelial layer.Our data also reinforce the crosstalk between VEGF and TGF-β1signaling pathways and suggests that therapies targeting bothcytokines may inhibit EMT in early stage prostate cancer.
Acknowledgments
This work has been funded by “UTE project CIMA”, ISCIII-RETICRD06/0020 grant.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.yexcr.2009.11.020.

566 E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
R E F E R E N C E S
[1] A. Jemal, M.M. Center, E. Ward, M.J. Thun, Cancer occurrence,Methods Mol. Biol. 471 (2009) 3–29.
[2] J.I. Epstein, Precursor lesions to prostatic adenocarcinoma,Virchows Arch. 454 (2009) 1–16.
[3] J. Folkman, D. Hanahan, Switch to the angiogenic phenotypeduring tumorigenesis, Princess Takamatsu Symp. 22 (1991)339–347.
[4] R. Catena, V. Muniz-Medina, B. Moralejo, B. Javierre, C.J. Best, M.R.Emmert-Buck, J.E. Green, C.C. Baker, A. Calvo, Increasedexpression of VEGF(121)/VEGF(165-189) ratio results in asignificant enhancement of human prostate tumor angiogenesis,Int. J. Cancer 120 (2007) 2096–2109.
[5] N. Ferrara, Vascular endothelial growth factor: basic science andclinical progress, Endocr. Rev. 25 (2004) 581–611.
[6] M.W. Jackson, J.S. Roberts, S.E. Heckford, C. Ricciardelli, J. Stahl, C.Choong, D.J. Horsfall, W.D. Tilley, A potential autocrine role forvascular endothelial growth factor in prostate cancer, Cancer Res.62 (2002) 854–859.
[7] J. Pallares, F. Rojo, J. Iriarte, J. Morote, L.I. Armadans, I. de Torres,Study of microvessel density and the expression of the angiogenicfactors VEGF, bFGF and the receptors Flt-1 and FLK-1 in benign,premalignant and malignant prostate tissues, Histol. Histopathol.21 (2006) 857–865.
[8] R. Mazzucchelli, R. Montironi, A. Santinelli, G. Lucarini, A.Pugnaloni, G. Biagini, Vascular endothelial growth factorexpression and capillary architecture in high-grade PIN andprostate cancer in untreated and androgen-ablated patients,Prostate 45 (2000) 72–79.
[9] W.J. Huss, C.F. Hanrahan, R.J. Barrios, J.W. Simons, N.M.Greenberg, Angiogenesis and prostate cancer: identification of amolecular progression switch, Cancer Res. 61 (2001) 2736–2743.
[10] A. Calvo, O. Gonzalez-Moreno, C.Y. Yoon, J.I. Huh, K. Desai, Q.T.Nguyen, J.E. Green, Prostate cancer and the genomic revolution:advances using microarray analyses, Mutat. Res. 576 (2005)66–79.
[11] A. Calvo, N. Xiao, J. Kang, C.J. Best, I. Leiva, M.R. Emmert-Buck, C.Jorcyk, J.E. Green, Alterations in gene expression profiles duringprostate cancer progression: functional correlations totumorigenicity and down-regulation of selenoprotein-P in mouseand human tumors, Cancer Res. 62 (2002) 5325–5335.
[12] C.R. Soares, M.A. Shibata, J.E. Green, C.L. Jorcyk, Development ofPIN and prostate adenocarcinoma cell lines: a model system formultistage tumor progression, Neoplasia 4 (2002) 112–120.
[13] Y. Nakaya, G. Sheng, Epithelial to mesenchymal transition duringgastrulation: an embryological view, Dev. Growth Differ. 50(2008) 755–766.
[14] G. Berx, E. Raspe, G. Christofori, J.P. Thiery, J.P. Sleeman,Pre-EMTing metastasis? Recapitulation of morphogeneticprocesses in cancer, Clin. Exp. Metastasis 24 (2007) 587–597.
[15] J. Xu, S. Lamouille, R. Derynck, TGF-beta-induced epithelial tomesenchymal transition, Cell Res. 19 (2009) 156–172.
[16] J. Lecanda, V. Ganapathy, C. D'Aquino-Ardalan, B. Evans, C.Cadacio, A. Ayala, L.I. Gold, TGFbeta prevents proteasomaldegradation of the cyclin-dependent kinase inhibitor p27kip1 forcell cycle arrest, Cell Cycle 8 (2009) 742–756.
[17] J. Lecanda, T.V. Parekh, P. Gama, K. Lin, V. Liarski, S. Uretsky, K.Mittal, L.I. Gold, Transforming growth factor-beta, estrogen, andprogesterone converge on the regulation of p27Kip1 in thenormal and malignant endometrium, Cancer Res. 67 (2007)1007–1018.
[18] A.D. Yang, E.R. Camp, F. Fan, L. Shen, M.J. Gray, W. Liu, R. Somcio,T.W. Bauer, Y. Wu, D.J. Hicklin, L.M. Ellis, Vascular endothelialgrowth factor receptor-1 activation mediates epithelial tomesenchymal transition in human pancreatic carcinoma cells,Cancer Res. 66 (2006) 46–51.
[19] S.C. Shih, G.S. Robinson, C.A. Perruzzi, A. Calvo, K. Desai, J.E. Green,I.U. Ali, L.E. Smith, D.R. Senger, Molecular profiling of angiogenesismarkers, Am. J. Pathol. 161 (2002) 35–41.
[20] G.K. Smyth, Linear models and empirical Bayes methods forassessing differential expression in microarray experiments, Stat.Appl. Genet. Mol. Biol. 3 (2004) Article3.
[21] L.A. Martinez-Cruz, A. Rubio, M.L. Martinez-Chantar, A. Labarga, I.Barrio, A. Podhorski, V. Segura, J.L. Sevilla Campo, M.A. Avila, J.M.Mato, GARBAN: genomic analysis and rapid biological annotationof cDNA microarray and proteomic data, Bioinformatics 19(2003) 2158–2160.
[22] J.C. Bryne, E. Valen, M.H. Tang, T. Marstrand, O. Winther, I. daPiedade, A. Krogh, B. Lenhard, A. Sandelin, JASPAR, the openaccess database of transcription factor-binding profiles: newcontent and tools in the 2008 update, Nucleic Acids Res. 36(2008) D102–106.
[23] V. Matys, O.V. Kel-Margoulis, E. Fricke, I. Liebich, S. Land, A.Barre-Dirrie, I. Reuter, D. Chekmenev, M. Krull, K. Hornischer, N.Voss, P. Stegmaier, B. Lewicki-Potapov, H. Saxel, A.E. Kel, E.Wingender, TRANSFAC and its module TRANSCompel:transcriptional gene regulation in eukaryotes, Nucleic Acids Res.34 (2006) D108–D110.
[24] E. Guruceaga, V. Segura, F.J. Corrales, A. Rubio, FactorY, abioinformatic resource for genome-wide promoter analysis,Comput. Biol. Med. 39 (2009) 385–387.
[25] Y. Tajima, K. Goto, M. Yoshida, K. Shinomiya, T. Sekimoto, Y.Yoneda, K. Miyazono, T. Imamura, Chromosomal regionmaintenance 1 (CRM1)-dependent nuclear export of Smadubiquitin regulatory factor 1 (Smurf1) is essential for negativeregulation of transforming growth factor-beta signaling bySmad7, J. Biol. Chem. 278 (2003) 10716–10721.
[26] C. Suzuki, G. Murakami, M. Fukuchi, T. Shimanuki, Y. Shikauchi, T.Imamura, K. Miyazono, Smurf1 regulates the inhibitory activity ofSmad7 by targeting Smad7 to the plasma membrane, J. Biol.Chem. 277 (2002) 39919–39925.
[27] S. Zhang, T. Fei, L. Zhang, R. Zhang, F. Chen, Y. Ning, Y. Han, X.H.Feng, A. Meng, Y.G. Chen, Smad7 antagonizes transforminggrowth factor beta signaling in the nucleus by interfering withfunctional Smad–DNA complex formation, Mol. Cell. Biol. 27(2007) 4488–4499.
[28] N.M. Meindl-Beinker, S. Dooley, Transforming growthfactor-beta and hepatocyte transdifferentiation in liverfibrogenesis, J. Gastroenterol. Hepatol. 23 (Suppl. 1) (2008)S122–S127.
[29] U. Valcourt, M. Kowanetz, H. Niimi, C.H. Heldin, A. Moustakas,TGF-beta and the Smad signaling pathway supporttranscriptomic reprogramming during epithelial–mesenchymalcell transition, Mol. Biol. Cell 16 (2005) 1987–2002.
[30] N.A. Bhowmick, M. Ghiassi, A. Bakin, M. Aakre, C.A. Lundquist,M.E. Engel, C.L. Arteaga, H.L. Moses, Transforming growthfactor-beta1 mediates epithelial to mesenchymaltransdifferentiation through a RhoA-dependent mechanism,Mol. Biol. Cell 12 (2001) 27–36.
[31] I. Serganova, E. Moroz, J. Vider, G. Gogiberidze, M. Moroz, N.Pillarsetty, M. Doubrovin, A. Minn, H.T. Thaler, J. Massague, J.Gelovani, R. Blasberg, Multimodality imaging of TGF{beta}signaling in breast cancer metastases, FASEB J. (2009).
[32] M. Sabbah, S. Emami, G. Redeuilh, S. Julien, G. Prevost, A. Zimber,R. Ouelaa, M. Bracke, O. De Wever, C. Gespach, Molecularsignature and therapeutic perspective of the epithelial-to-mesenchymal transitions in epithelial cancers, Drug Resist. Updat.11 (2008) 123–151.
[33] G. Han, S.L. Lu, A.G. Li, W. He, C.L. Corless, M. Kulesz-Martin, X.J.Wang, Distinct mechanisms of TGF-beta1-mediatedepithelial-to-mesenchymal transition and metastasis during skincarcinogenesis, J. Clin. Invest. 115 (2005) 1714–1723.
[34] A.V. Bakin, A.K. Tomlinson, N.A. Bhowmick, H.L. Moses, C.L.Arteaga, Phosphatidylinositol 3-kinase function is required fortransforming growth factor beta-mediated epithelial to

567E X P E R I M E N T A L C E L L R E S E A R C H 3 1 6 ( 2 0 1 0 ) 5 5 4 – 5 6 7
mesenchymal transition and cell migration, J. Biol. Chem. 275(2000) 36803–36810.
[35] C. Come, V. Arnoux, F. Bibeau, P. Savagner, Roles of thetranscription factors snail and slug during mammarymorphogenesis and breast carcinoma progression, J. MammaryGland Biol. Neoplasia. 9 (2004) 183–193.
[36] L.S. Wanami, H.Y. Chen, S. Peiro, A. Garcia de Herreros, R.E.Bachelder, Vascular endothelial growth factor-A stimulates Snailexpression in breast tumor cells: implications for tumorprogression, Exp. Cell Res. 314 (2008) 2448–2453.
[37] M.J. Blanco, G. Moreno-Bueno, D. Sarrio, A. Locascio, A. Cano, J.Palacios, M.A. Nieto, Correlation of Snail expression with
histological grade and lymph node status in breast carcinomas,Oncogene 21 (2002) 3241–3246.
[38] K. Jungert, A. Buck, M. Buchholz, M. Wagner, G. Adler, T.M.Gress, V. Ellenrieder, Smad-Sp1 complexes mediateTGFbeta-induced early transcription of oncogenic Smad7 inpancreatic cancer cells, Carcinogenesis 27 (2006)2392–2401.
[39] K. Jungert, A. Buck, G. von Wichert, G. Adler, A. Konig, M.Buchholz, T.M. Gress, V. Ellenrieder, Sp1 is required fortransforming growth factor-beta-induced mesenchymaltransition and migration in pancreatic cancer cells, Cancer Res. 67(2007) 1563–1570.
Copyright © 2022 FDOKUMEN




















