
Tidal stream device reliability comparison models Tidal stream device reliability comparison models
Upload
independentCategory
view
0download
0
CSIRO PUBLISHING
www.publish.csiro.au/journals/mfr Marine and Freshwater Research, 2005, 56, 825–833
Response of stream macroinvertebrates to changes in salinityand the development of a salinity index
Nelli HorriganA,C, Satish ChoyA, Jonathan MarshallA and Friedrich RecknagelB
ADepartment of Natural Resources and Mines (NR&M), 120 Meiers Rd, Indooroopilly, Qld 4068, Australia.BSchool of Earth and Environmental Sciences, University of Adelaide, SA 5000, Australia.
CCorresponding author. Email: [email protected]
Abstract. Many streams and wetlands have been affected by increasing salinity, leading to significant changesin flora and fauna. The study investigates relationships between macroinvertebrate taxa and conductivity levels(µS cm−1) in Queensland stream systems. The analysed dataset contained occurrence patterns of frequently foundmacroinvertebrate taxa from edge (2580 samples) and riffle (1367 samples) habitats collected in spring and autumnover 8 years. Sensitivity analysis with predictive artificial neural network models and the taxon-specific meanconductivity values were used to assign a salinity sensitivity score (SSS) to each taxon (1—very tolerant, 5—tolerant, 10—sensitive). Salinity index (SI) based on the cumulative SSS was proposed as a measurement of changein macroinvertebrate communities caused by salinity increase. Changes in macroinvertebrate communities wereobserved at relatively low salinities, with SI rapidly decreasing to ∼800–1000 µS cm−1 and decreasing further at aslower rate. Natural variability and water quality factors were ruled out as potential primary causes of the observedchanges by using partial canonical correspondence analysis and subsets of the data with only good water quality.
Extra keywords: anthropogenic impacts, aquatic invertebrates, dryland salinity, ecological indicators, salt pollution.
Introduction
Increasing concentration of dissolved solids in freshwatersystems is becoming a serious problem in many parts of theworld (Hart et al. 1991; Goetsch and Palmer 1997; Lelandand Fend 1998). Many streams and wetlands have beenaffected by rising salinity leading to significant changes inflora and fauna (Hart et al. 1991). Secondary salinisationcan affect aquatic systems in multiple ways, including directtoxic effects, changed chemical processes and the loss ofhabitat in streams, riparian zones and adjacent floodplains.Current understanding of the resilience of freshwater biota tothese impacts is limited and more research is needed (Jameset al. 2003).
Several authors have studied the changes in macroinver-tebrate communities along the salinity gradient in streamsand rivers (Williams et al. 1991; Bunn and Davies 1992;Metzeling 1993; Kefford 1998; Leland and Fend 1998;Kay et al. 2001). The major difficulty associated with thisapproach is dissociating the effects of salinity from those ofthe other factors. Different responses have been observed, butthere is a general acceptance that freshwater ecosystemsexperience little ecological stress when the electric conduc-tivity (EC) level is below 1500 µS cm−1 (Hart et al. 1991).Marshall and Bailey (2004) conducted field experiments toexamine the effect of short-term releases of saline wastew-ater on stream macroinvertebrate communities. Significant
reduction in the abundance of some species (Ferrissiatasmanica, Baetis sp. 5) were observed at 1500 mg L−1
(∼2205 µS cm−1).Kefford et al. (2003) estimated the relative salinity toler-
ance to artificial seawater of macroinvertebrate species underacute 72-h exposure. Even though many macroinvertebratesin older stages appear to be highly tolerant (mean LC50
31 000 µS cm−1 ranging from 5500 to 76 000 µS cm−1), eggsand hatchlings had a salinity tolerance between 5% and 100%of 72-h LC50 of the older stages, with less than 50% for insects(Kefford et al. 2004). Long-term or indirect effects of salinityare still poorly understood and might cause biological effectsat lower salinity levels than those causing direct toxic effects(Kefford et al. 2003). In this respect, analysis of field datacollected on a wide geographical scale and over many yearscan provide an important insight into the effects of salini-sation including all potential mechanisms by which salinitymay impact on freshwater organisms.
Statistical methods, such as correlations, multiple linearregression, generalised additive models, logistic regression,principal component analysis and cluster analysis, are com-monly used to study species distribution along an environ-mental gradient. However, there are two potential problemsassociated with this approach. First, it is often difficult toknow for certain that observed changes in biota are causedby changes in salinity and not by a multitude of the other
© CSIRO 2005 10.1071/MF04237 1323-1650/05/060825

826 Marine and Freshwater Research N. Horrigan et al.
underlying factors. Second, our confidence in the resultsis often limited by the method’s inability to meet severalassumptions, such as statistical distribution, independenceof variables and linearity of the relationships.
Use of machine learning methods as artificial neuralnetworks (ANNs) overcomes the limitations (e.g. data dis-tributions or non-linearity) of many traditional statisticalmethods. Artificial neural networks are considered univer-sal and highly flexible approximators for any data and arepowerful tools for ecological modelling, especially when thedata relationships are unknown (Lek and Guegan 1999). Sev-eral researchers found superior performance of ANNs whencompared with the other methods (linear regression, multipleregression, PCA and combination of PCA with least-squaresregression) (Ball et al. 2000; Brosse et al. 2001), particu-larly when the data were known to contain strong non-linearrelationships (Olden and Jackson 2002).
Sensitivity analysis with ANNs is a relatively newmethod for the estimation of complex non-linear relation-ships between environmental factors and taxon distribution.It allows simulation of the changes in biota specific to a vari-able in question when all the other variables in the model arekept static. This means that we can have some confidencethat predicted changes are caused by the factor in ques-tion (e.g. conductivity) and not underlying natural or otheranthropogenic gradients. Sensitivity analysis usingANNs hasbeen successfully applied to the investigation of relationshipsbetween habitat properties and the occurrence of macroin-vertebrates in Queensland streams (Hoang et al. 2001, 2003;Marshall et al. 2002).
Aquatic systems affected by secondary salinisation oftenalso experience poor water quality, for example higher con-centration of nutrients, suspended particulate matter andtoxicants (Hart et al. 2003). In this respect, it might be advan-tageous to consider the effect of EC in combination with theother water quality factors. Canonical correspondence anal-ysis (CCA) allows description and visualisation of relation-ships between multiple environmental variables and biota (terBraak and Verdonschot 1995). Partial CCA (ter Braak 1988)is of particular interest to us because it makes it possible topartial out the effect of covariables (e.g. temporal and naturalvariability) and use the residuals to examine variables of inter-est (e.g. EC and other water quality variables). The methodis also relatively robust in terms of statistical distribution.
The aim of this study was to investigate changes inmacroinvertebrate communities associated with a conductiv-ity gradient in Queensland, Australia using a semi-qualitativeapproach. The study was conducted using a large dataset con-taining occurrences of macroinvertebrate families collectedover eight years. Analysis of data collected over a wide geo-graphical area can provide an interesting broad-scale insightinto the differences between structures of macroinvertebratecommunities under varied levels of stress from salinisation.This study utilises sensitivity analysis usingANNs to estimate
conductivity tolerances of specific macroinvertebrate taxaand to develop an index reflecting changes in macroinver-tebrate communities as a result of changes in conductivitylevel. Partial CCA is used to provide an additional insightinto the effect of EC and the other water quality factors onthe structure of macroinvertebrate communities.
Materials and methods
Data
Data were collected each spring and autumn from 1994 to 2002 from1008 sites (Fig. 1) by the Department of Natural Resources and Mines(NR&M). A total of 2580 samples from edge habitat and 1367 samplesfrom riffle habitat were used for the analysis. Data from the two dif-ferent habitats were considered separately in order to reduce the effectof natural variability; there is also a geographical difference betweensubsets because many samples from the edge habitat were collectedin western inland areas and the majority of samples from riffle habi-tat were collected in coastal regions. Salinity level was measured asEC (µS cm−1) adjusted for temperature. We used salinity categoriesgiven in the median range guidelines for surface water conductivity(DEH and NR&M 1999): fresh water <800 µS cm−1, marginal 800–2500 µS cm−1, brackish 2500–8000 µS cm−1, saline >8000 µS cm−1.
The purpose of this study was to identify general trends using fre-quently found macroinvertebrate taxa with state-wide distributions. Weused taxa occurring in more than 5% of all samples. Several taxa withlimited geographic distributions were excluded from the analysis inorder to avoid confounding the results with their natural geographical
N
FreshMarginalBrackishSaline
Kilometres200 0 200 400
Fig. 1. Sampling sites marked according to the median range guide-lines for surface water conductivity (DEH and NR&M 1999).

Response of stream macroinvertebrates to salinity Marine and Freshwater Research 827
distribution range. This left 57 taxa (mostly family level) from edgehabitat and 60 taxa from riffle habitat for the analysis.
The sites were sampled according to the standard Australian RiverAssessment System (AusRivAS) rapid assessment protocols and pro-cessed by NR&M ecologists using standard procedures (Conrick andCockayne 2000). The method involves sampling 10 m of habitat usinga 250-µm mesh dip net with live picking in the field and identificationof samples in the laboratory.
Data analysis and modelling
Sensitivity analysis with multilayer perceptron
Artificial neural network models have been widely applied to ecolog-ical problems (Lek and Guegan 1999) and have proven to be a valuabletool when dealing with multidimensional noisy data with complicatednon-linear relationships between variables.
Multilayer perceptron (MLP) is the most commonly used type ofpredictive ANN. Multilayer perceptron is described in several publica-tions (Haykin 1999; Principe et al. 2000), and only a brief outline isprovided here. An ANN is composed of two main units: a processingelement or neuron and connections or weights between the neurons. Theneuron receives signals from other neurons or outside and processes thisinformation. If cumulative input exceeds a certain threshold, an outputsignal is sent to other neurons in the network. Typically, MLP consistsof input, hidden and output layers of neurons. The input layer contains pneurons, each of which represents one of the p predictor variables. Theoptimal number of hidden neurons in the neural network is determinedempirically by choosing the number of hidden neurons that produces thelowest misclassification rate (Bishop 1995). The output layer containsone neuron representing the probability of species occurrence. Eachneuron of a given layer is connected to all neurons in the next layer.In the process of learning, a neuron receives inputs from the input orprevious layer, inputs are multiplied by weight values (which is initiallya random number) and combines these weighted inputs.
netj =∑
wijxi,
where netj is the summed weighted input for jth neuron, wij is the weightfrom ith neuron in the previous layer to the jth neuron at the currentlayer, and xi is the input from the ith to the jth neuron (Poff 1996). Thenet is passed through a transfer function, most commonly a sigmoidalor hyperbolic tangent (tanh) function. In this study, tanh function wasused:
tanh z = ez − e−z/ez + e−z,
where e is the base of the natural algorithm.The network calculates the outputs for all neurons and compares the
total output to the actual or target output. The goal of learning is to finda set of weights that minimises the mean-squared error (MSE) betweenthe target and calculated output. Weights are modified after each itera-tion (epoch) and output is calculated depending on the magnitude anddirection of the error. The iterative technique of minimising error isknow as gradient descent, where weights are modified in the directionof greatest descent, travelling ‘downhill’ in the direction of the mini-mum (Olden and Jackson 2002). It is possible to overtrain the network,producing excellent fit for the training data but poor predictions for thevalidation (generalisation) set. Early stopping or stopping with cross-validation method is used to ensure the maximum generalisation of themodel (Principe et al. 2000). The cross-validation set of data is usuallytaken as 10% data not used for training. Training is stopped every 5 to 10epochs and the model is tested against the cross-validation set. Trainingshould be stopped when the MSE in the cross-validation set starts toincrease. When training is successfully completed, MLP can be used fora prediction or the sensitivity analysis.
In order to estimate the salinity tolerance of each macroinvertebratetaxon, we built 117 taxon-specific MLP models (57 for edge habitat and
60 for riffle habitat) with the following architecture: 25 neurons in theinput layer associated with predictor variables, six neurons in the hid-den layer and one neuron in the output. The variables used as predictorsare shown in Table 1. Data were range standardised between 0 and 1.Optimum number of epochs during which overtraining does not occurwas determined using cross-validation method with 10% of the data.Thequality of each model was estimated as the percentage of correct predic-tion in the cross-validation subset. For sensitivity analysis, we changedthe mean value of conductivity by 10 standard deviations and calculated50 steps in each direction, while all the other predictor variables werekept at their respective means. Each taxon-specific model was simulatedand continuous output for each taxon was plotted against EC to forma response curve. Output of the model ranges between 0 and 1 and isinterpreted as probability of taxon occurrence, higher value indicatinghigher probability of occurrence. The response curves were classifiedinto one of three shapes: (i) monotonically decreasing; (ii) unimodal;and (iii) monotonically increasing. These shapes were used to classifydifferent taxa as sensitive or tolerant to the conductivity level. As con-ductivity increases, a sensitive taxon would be less likely to occur and,therefore, would exhibit a monotonically decreasing response curve.A tolerant taxon would exhibit a monotonically increasing relationshipalong the conductivity gradient because it would be more likely to occuras conductivity increases.
Salinity sensitivity score and salinity index
All taxa were assigned the following salinity sensitivity scores (SSS):‘10’ for sensitive, ‘5’ for generally tolerant, and ‘1’ for very tolerant.The range from 1 to 10 was used as for SIGNAL (Stream InvertebrateGrade Number—Average Level) index proposed by Chessman (2003).
Table 1. List of variables used as predictors for trainingmultilayer perceptron (MLP) models
Variable Units
Maximum water velocity m s−1
Habitat depth mSeason (spring and autumn)YearBedrock %Boulder %Cobble %Pebble %Gravel %Sand %Silt/clay %Latitude decimalAltitude mStream orderSlope m km−1
Distance from source kmRatio of mean wet season monthly
rainfall to mean dry seasonmonthly rainfall
Mean annual rainfall mmWater temperature ◦CConductivity µS cm−1
Dissolved oxygen mg L−1
pHTurbidity NTUAlkalinity mg L−1 CaCO3
Total phosphorus mg L−1 as P

828 Marine and Freshwater Research N. Horrigan et al.
The following rules were used to determine which score should beassigned (quantitative cut-off values are based on the mean conduc-tivities in a given habitat dataset with a gap of 50 µS cm−1 to providefor some additional distance between sensitive and very tolerant taxa):
Sensitive: shape of sensitivity curve = ‘Decreasing’ and MEAN con-ductivity ≤ 300 µS cm−1 (edge), 250 µS cm−1 (riffle)
Very tolerant: shape of sensitivity curve = ‘Increasing’ and MEANconductivity >350 µS cm−1 (edge), 300 µS cm−1 (riffle)
Generally tolerant: all taxa that fell neither into sensitive nor very
tolerant categories were assigned score ‘5’.
These rules were used only as a general guide, exceptions were madein some cases. For example, we had to take into consideration maximalconductivity at which taxa occur because some taxa show an increasingtrend but were not found in high conductivities. We labelled taxa asgenerally tolerant in all cases where results were inconclusive or con-troversial. Using the SSS of all taxa in a sample, we calculated a salinityindex similar to a chloride contamination index suggested by Williamset al. (2000).
Salinity index (SI) = (∑
Xi × SSSi)/n
where Xi = 1 if taxon i was present, Xi = 0 if absent, SSSi = saltsensitivity score of taxon i and n = the total number of taxa in the sample.
Salinity index can theoretically vary from a value of 1 when all thetaxa in a sample are highly tolerant, to a value of 10 when all taxa aresensitive. In practice, we would expect opportunistic taxa to be present inboth unimpacted and impacted sites and contribute to keeping the totalscore to less then 10 and higher than 1. We used box-and-whisker plotsto demonstrate the changes in percentage of groups with different salin-ity preferences along the conductivity gradient. The data were sortedby ascending conductivity and split into 12 equal-sized bins with noconsideration of the site location, year, etc. Number of bins was chosenarbitrarily.
To test whether SI indeed reflects changes in macroinvertebratecommunities mainly due to the changes in conductivity and not theeffect of other wide-spread stressors, such as concentration of nutri-ents, we isolated a subset of data with otherwise good water quality:turbidity <5 NTU, total nitrogen <0.375 mg L−1, total phosphorus<0.05 mg L−1, pH between 6.5 and 9 and dissolved oxygen >5 mg L−1.These values are taken from water quality guidelines for the protection ofaquatic ecosystems (Bloedel et al. 2000). This analysis excluded waterquality parameters other than conductivity as a potential cause of theobserved changes in macroinvertebrate communities. However, it is stillpossible that these changes related to natural factors like rainfall, dis-tance from source, flow, etc. In order to account for this possibility, weused partial CCA.
Partial canonical correspondence analysis
Partial CCA was used to examine relationships of water quality vari-ables to each other and macroinvertebrate communities, having excludedthe influence of natural factors, flow and temporal variability. First weused CCA to determine which variables out of the set of 25 (see Table 1)were significant in structuring macroinvertebrate communities. Forwardselection procedure (ter Braak and Verdonschot 1995) with 999 MonteCarlo permutations was used to test the significance of each variable.Only variables with P < 0.01 were used for the subsequent analysis. Thesignificant variables were divided into two sets: (i) covariables describ-ing temporal and natural variability (e.g. season, year, rainfall, distancefrom source, etc.); and (ii) water quality variables (e.g. EC, water tem-perature, pH, turbidity, etc.). Using partial CCA we excluded temporaland natural variability and built two separate biplots for each habitat,reflecting only the effect of water quality variables.
Results
The distribution of conductivity values in the dataset washighly skewed towards lower values, with the majority of sitesfalling into the fresh water category (see Fig. 1). For exam-ple, the data for the edge habitat contained 181 samples withconductivities in the marginal category, 20 in brackish andonly three in saline. The highest conductivities in the datasetwere 12 000 µS cm−1 and 5600 µS cm−1 in edge and rifflehabitat, respectively, with mean conductivities 355 µS cm−1
and 300 µS cm−1, respectively. These conductivity levelsare probably not high enough to cause visible reduction intaxonomic richness (Fig. 2).
Sensitivity analysis with multilayer perceptron
Average percentage of correct predictions of the 117 MLPmodels (tested on 10% of all data not used for training)was 72.4% ranging from 50 to 89.8%. Figure 3 shows typ-ical trends in the probability of taxon occurrence along theconductivity gradient. Monotonically increasing or decreas-ing shapes of the sensitivity curves were the most common,with only five taxa exhibiting unimodal shapes. Trends for
0
2000
4000
6000
8000
1000
0
1200
0
Conductivity (�S cm�1)
0
10
20
30
40
50
Ric
hnes
s
(a)
Conductivity (�S cm�1)
0
1000
2000
3000
4000
5000
6000
0
5
10
15
20
25
30
35
40
Ric
hnes
s
(b)
Fig. 2. Scatterplot of taxonomic richness v. conductivity for (a) edgeand (b) riffle habitats.

Response of stream macroinvertebrates to salinity Marine and Freshwater Research 829
each taxon are documented in Appendix 1 (edge habitat)and Appendix 2 (riffle habitat) (Appendices available asAccessory Material on the Marine and Freshwater Researchwebsite). We suggest that a taxon showing a decreasing trendcan be considered as sensitive because it shows preferencetowards lower conductivities. Taxa with increasing trendshave high tolerance to salinity because probability of theiroccurrence increases with increase in conductivity. Taxa withunimodal curves are considered opportunistic taxa, with widetolerances or preferences around the medium conductivityrange.
Salinity sensitivity score and salinity index
Salinity sensitivity scores resulting from the outputs of sen-sitivity analysis and mean conductivity values are shown inAppendix 1 (edge habitat) and Appendix 2 (riffle habitat)(Appendices available as Accessory Material on the Marineand Freshwater Research website).Twelve taxa were labelledas sensitive in both datasets and 16 and 21 were labelledas very tolerant in edge and riffle habitats respectively. Thegenerally tolerant taxa comprised the largest group in bothhabitats, 29 in edge and 27 in riffle.
As conductivity increases, sensitive taxa are beingreplaced by very tolerant taxa (Fig. 4).This trend is obvious inboth habitats but appears to be more prominent in riffle habi-tat, with higher proportions of sensitive taxa present underlow conductivities. For example, the sites with an EC in the
(a)
(b)
(c)
0
0.1
0.2
0.3
-1
DugesiidaeTipulidaeLeptophlebiidae
0.40.50.60.70.80.9
1
Pro
babi
lity
of o
ccur
renc
e
Copepoda
Dytiscidae
Hydraenidae
0.4
0.5
0.5
0.6
0.6
0.7
0 2000 4000 6000 8000
Ceratopogonidae
Cladocera
Protoneuridae
Conductivity (�S cm�1)
Fig. 3. Typical shapes of the sensitivity curves resulted from the sensi-tivity analysis with multilayer perceptron—edge habitat. (a) Decreasing,(b) increasing and (c) unimodal trends in the probability of occurrenceof macroinvertebrate taxa along the conductivity gradient.
range of 800–1500 µS cm−1 were observed to decrease inthe mean percentage of sensitive taxa from 33 to 16.7 rela-tive to the low conductivity category (22–99 µS cm−1) andthe percentage of very tolerant taxa increased accordinglyfrom 9.4 to 32.
Salinity index calculated using all three tolerance groupsranged from 8 to 2.14 in riffle habitat and from 6.8 to 1in the edge. Most of the sites with high SI had low con-ductivity; for example, out of 710 samples (edge habitat)with SI values between 5 and 7.3, only nine sites (1.2%)had conductivity higher than 1000 µS cm−1. However, theconductivity in sites with low SI was not always high. Outof 498 samples with SI between 2 and 4, 411 samples hadconductivity less than 800 µS cm−1. More than half of thesesamples (216) had elevated concentrations of nutrients (totalnitrogen >0.75 mg L−1 or total phosphorus >0.1 mg L−1) orturbidity higher then 50 NTU, which might explain the pres-ence of mainly opportunistic taxa in these sites. However, wecould not explain low SI (between 2 and 4) at 195 samples. Italso has become apparent that many samples with SI too high
SensitiveVery tolerant
6–51
51–6
364
–89
89–1
2512
5–15
915
9–20
520
5–27
027
1–33
833
8–43
043
0–56
056
0–77
377
4–12
000
0
10
20
30
40
50
60
70
80
%
SensitiveVery tolerant
22–4
949
–59
59–7
272
–99
100–
132
133–
175
175–
232
233–
309
310–
396
396–
530
530–
692
696–
5600
0
10
20
30
40
50
60
70
80
%
Conductivity (�S cm�1)
(a)
(b)
Fig. 4. Percentage of sensitive and very tolerant taxa in 12 equal-sizedbins along increasing conductivity gradient: (a) edge habitat, (b) rif-fle habitat. Median values with boxes corresponding to 80th and 20thpercentiles and horizontal bars to maximum and minimum.
830 Marine and Freshwater Research N. Horrigan et al.
2
34
5
6
78
9
SI
2
3
4
5
6
7
8
9
0 1000 2000 3000 4000 5000 6000
SI
(a)
(b)
Conductivity (�S cm�1)
0 1000 2000 3000 4000 5000 6000
Conductivity (�S cm�1)
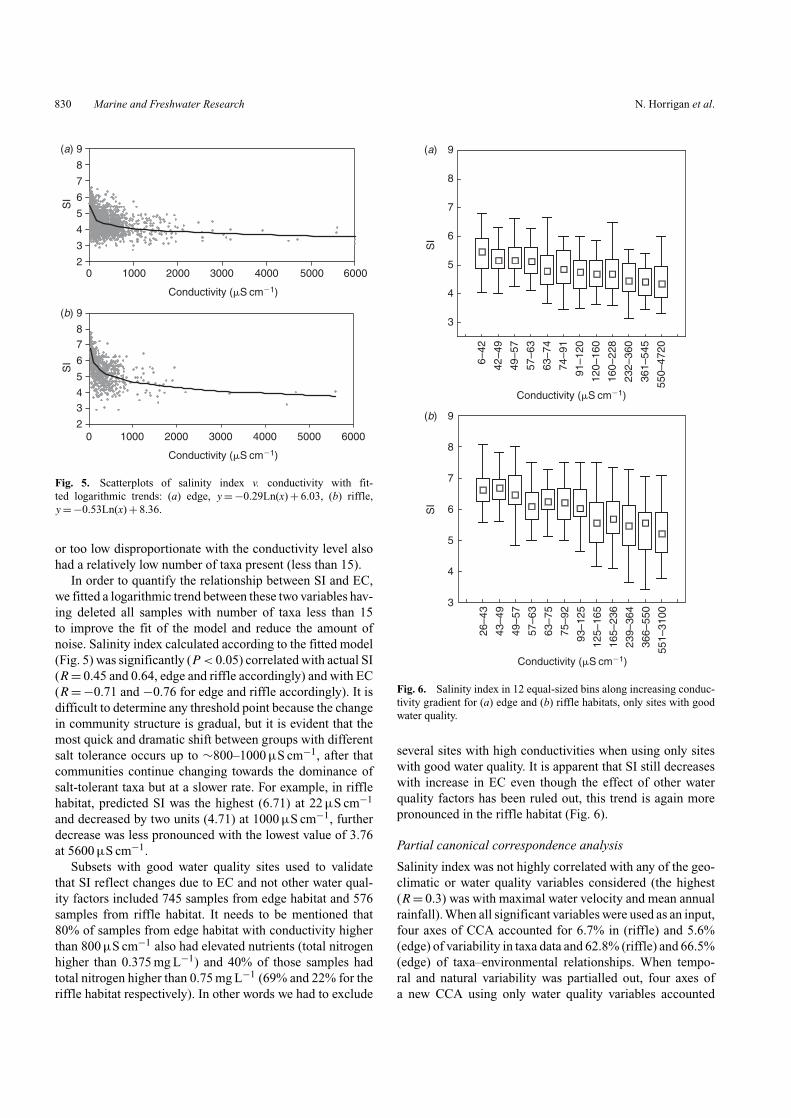
Fig. 5. Scatterplots of salinity index v. conductivity with fit-ted logarithmic trends: (a) edge, y = −0.29Ln(x) + 6.03, (b) riffle,y = −0.53Ln(x) + 8.36.
or too low disproportionate with the conductivity level alsohad a relatively low number of taxa present (less than 15).
In order to quantify the relationship between SI and EC,we fitted a logarithmic trend between these two variables hav-ing deleted all samples with number of taxa less than 15to improve the fit of the model and reduce the amount ofnoise. Salinity index calculated according to the fitted model(Fig. 5) was significantly (P < 0.05) correlated with actual SI(R = 0.45 and 0.64, edge and riffle accordingly) and with EC(R = −0.71 and −0.76 for edge and riffle accordingly). It isdifficult to determine any threshold point because the changein community structure is gradual, but it is evident that themost quick and dramatic shift between groups with differentsalt tolerance occurs up to ∼800–1000 µS cm−1, after thatcommunities continue changing towards the dominance ofsalt-tolerant taxa but at a slower rate. For example, in rifflehabitat, predicted SI was the highest (6.71) at 22 µS cm−1
and decreased by two units (4.71) at 1000 µS cm−1, furtherdecrease was less pronounced with the lowest value of 3.76at 5600 µS cm−1.
Subsets with good water quality sites used to validatethat SI reflect changes due to EC and not other water qual-ity factors included 745 samples from edge habitat and 576samples from riffle habitat. It needs to be mentioned that80% of samples from edge habitat with conductivity higherthan 800 µS cm−1 also had elevated nutrients (total nitrogenhigher than 0.375 mg L−1) and 40% of those samples hadtotal nitrogen higher than 0.75 mg L−1 (69% and 22% for theriffle habitat respectively). In other words we had to exclude
26–4
3
43–4
9
49–5
7
57–6
3
63–7
5
75–9
2
93–1
25
125–
165
165–
236
239–
364
366–
550
551–
3100
3
4
5
6
7
8
9S
I
6–42
42–4
9
49–5
7
57–6
3
63–7
4
74–9
1
91–1
20
120–
160
160–
228
232–
360
361–
545
550–
4720
3
4
5
6
7
8
9
SI
(a)
(b)
Conductivity (�S cm�1)
Conductivity (�S cm�1)
Fig. 6. Salinity index in 12 equal-sized bins along increasing conduc-tivity gradient for (a) edge and (b) riffle habitats, only sites with goodwater quality.
several sites with high conductivities when using only siteswith good water quality. It is apparent that SI still decreaseswith increase in EC even though the effect of other waterquality factors has been ruled out, this trend is again morepronounced in the riffle habitat (Fig. 6).
Partial canonical correspondence analysis
Salinity index was not highly correlated with any of the geo-climatic or water quality variables considered (the highest(R = 0.3) was with maximal water velocity and mean annualrainfall). When all significant variables were used as an input,four axes of CCA accounted for 6.7% in (riffle) and 5.6%(edge) of variability in taxa data and 62.8% (riffle) and 66.5%(edge) of taxa–environmental relationships. When tempo-ral and natural variability was partialled out, four axes ofa new CCA using only water quality variables accounted

Response of stream macroinvertebrates to salinity Marine and Freshwater Research 831
�0.6 1.0
�0.
80.
6
S
S
*
TTT
T
T
* T*
T
T* *S
*
*
SS
** T
*
*T
*
*
*
*
T
TT
*
TT
S
*T
T
S
S
** ** TS
*
T
T**
T*
*
*
S
SS
Water temperature
Conductivity
DO
pH
Turbidity
Total P
�0.6 1.0
�0.
80.
8
S
S
*
T
T
T
*
*
*
T*
T
*
*
T
S
*
*
*
S
S
*T
*
*
*
T
T
*
*
T
**T
*
T
S
*
*
T*
T
*
*
S *
TS
T *
*
S
*
S*
S
S
Water temperatureConductivity
DOpH
Turbidity
Alkalinity
Total P
(a)
(b)
Fig. 7. Canonical correspondence analysis biplots showing effect ofwater quality variables after the effect of natural and temporal variabilitywas accounted for (a) edge, (b) riffle. S, Sensitive taxa; T, very toleranttaxa; *generally tolerant taxa; DO, dissolved oxygen (mg L−1); Total P,total phosphorus (mg L−1).
for 2% (riffle) and 1.3% (edge) of variability in taxa dataand 80.9% (riffle) and 77.5% (edge) of taxa–environmentalrelationships. Figure 7 clearly shows that conductivity, watertemperature, total phosphorus and pH affect macroinverte-brate communities in a similar way. It appears that totalphosphorus and EC are correlated stronger in the riffle subset.Alkalinity is not shown in the riffle biplot because it was foundto be insignificant in the previous stages of the analysis. Thestarting point of each arrow (Fig. 7) represents mean value ofthe variable. The majority of the taxa labelled as very tolerant(T) are located along the gradient of increasing, higher than
mean EC, and the taxa labelled as sensitive (S) are locatedmostly on the opposite side. This confirms that most of thetaxa were labelled correctly, with slightly better accuracy inthe case of riffle habitat. However, it is also evident that someconfounding by the other water quality variables is possible.
Discussion
The aim of this study was to investigate changes in themacroinvertebrate communities associated with changes inthe conductivity level in streams and rivers. It has been shownthat steady substitute of salt-sensitive taxa by opportunis-tic and salt-tolerant taxa does occur, even with relativelysmall increase in EC. The data analysis was intended to ruleout other possible causes for the observed changes, such asnatural variability, flow or other water quality parameters.Even though we have a reasonable confidence that describedchanges in macroinvertebrate communities are caused by thechanges in the conductivity as a primary stressor, it is impos-sible to rule out all the possibilities. Partial CCA showed thatincrease in conductivity can also be associated with increasein water temperature and nutrient level, combination of thesefactors might have a compounding effect on the macroinver-tebrate communities and is currently poorly understood. Thisconcern has also been expressed by other authors (James et al.2003) and more research in this direction is needed.
We have not observed any reduction in taxonomic rich-ness with the increase in conductivity within the limits of theavailable data. Several other authors have found no changein richness along a salinity gradient although the communitycomposition differed (Williams et al. 1990; Metzeling 1993;Kefford 1998). Taxonomic richness might not be a sensitiveenough indicator to detect the effect of secondary salinisa-tion because sensitive species are simply replaced by moretolerant ones with overall number of species remaining at thesame level.
According to our results, the most dramatic shift betweengroups of different salinity tolerance occurs as conductivityreaches 800–1000 µS cm−1 and this shift seems to be morepronounced in riffle habitat.This threshold value is lower thanthe generally accepted value of 1500 µS cm−1, above whichfreshwater ecosystems are likely to experience salinity relatedstress (Hart et al. 1991). One possible explanation for this dif-ference is that we used the state-wide dataset containing manysamples from streams in very good condition and with verylow conductivities (in range of 6–40 µS cm−1), this is par-ticularly relevant in the case of riffle dataset. In other words,changes in stream macroinvertebrate communities affectedby secondary salinisation may be more obvious in compari-son with stream systems in near-pristine condition, whereaswe might not observe any drastic changes when comparingsystems already impacted to some degree by a variety ofanthropogenic stressors and dominated by opportunistic taxawith likewise. Even though the changes in freshwater biota

832 Marine and Freshwater Research N. Horrigan et al.
might be initially subtle, this could lead to further instabilityof the ecosystem structure and function and compoundingeffects of other potential stressors.
The value of SI was decreasing with the increase inconductivity indicating changes in the community struc-ture towards domination of salt tolerant taxa. This patternremained the same when SI and conductivity were plottedfor the data subset containing stream sites with otherwisegood water quality, which confirms that SI indeed most likelyreflects changes in community structure associated with thechanges in conductivity. However, there were sites with lowSI and low conductivity. One possible explanation for strangeSI values is generally low number of taxa in the sample(lower than 15). Salinity index appears to be less reliablewhen calculated for the communities with a low number oftaxa. Another probable explanation is that sites with unex-pectedly low SI might be under stress unaccounted for inthe scope of this study (impoverished habitats, presents oftoxicants, etc.). There is also a possibility that some low-score taxa were assigned incorrectly. This may be the casebecause the distribution of conductivity values in the datasetwas highly skewed towards lower values, with the majorityof sites falling into the fresh water category. This means thatsome taxa graded as tolerant on the basis of this data might notbe so if more data from the higher salinity range was available.For example, some molluscs (Thiaridae, Lymnaeidae, Planor-bidae) and crustaceans in the family Parastacidae, which aregenerally considered salinity sensitive (Hart et al. 1991), werescored as very tolerant in this study, which might only be truefor the conductivity range considered in this study. However,we do not know which particular species were in those fam-ilies, which makes it difficult to compare our results withthe other studies considering sensitivities of species. It alsoneeds to be mentioned that life activities of molluscs at lowsalinity (below 50 mg L−1) are inhibited because they need asufficient calcium and sodium uptake to counterbalance theirmetabolic loss and build shells (Berezina 2003). This mighthave an effect on our results because many sites consideredin this study had very low conductivities.
The other possible factors causing discrepancies in thesalinity index v. conductivity relationship could be: natu-ral patterns in taxa distribution (we only used taxa withstate-wide distribution, but we do not exclude that naturalvariability might be having an underlying effect), lag effect ofprevious exposure (stream had an inflow of fresh water priorto the sampling event, but fauna was still recovering from aprevious salinity level) and the effect of ionic composition.The last factor was not taken into consideration in this study.However, it has a potential to confound the results. Goetschand Palmer (1997) demonstrated that acute mortality in labo-ratory experimentation cannot be linked only to conductivitybut also the nature of the salt. Different ionic composition ofwater with otherwise equal conductivities might affect fresh-water biota in different ways. Bayly (1969) suggested that
monovalent ions (Na+ and K+) are more toxic than divalentions (Ca2+). McNeil et al. (2005) defined a series of watertypes in Queensland based on proportions of cations andanions. The proportion of sodium chloride decreases west-ward from the coast. The streams from inland catchmentsare often high in calcium bicarbonate, sulphate and othercomponents. This means that higher proportions of sensi-tive taxa could be found in calcium bicarbonate dominatedwater than in sodium chloride dominated water under equalconductivities.
Even though we observed similar changes in communitiesfrom both riffle and edge habitat, these changes are more pro-nounced in riffles.As was mentioned before, edge habitat dataincluded many sites from the western regions, including manystreams with intermittent flow. It is possible that macroin-vertebrates inhabiting these sites are better adapted to thenatural changes in salinity than those from the streams withpermanent flow. This difference deserves further research,which could give us more understanding into the mechanismof adaptations and natural resilience of freshwater biota.
Our findings provide an interesting insight into broad-scale salinity sensitivities of Queensland stream macroinver-tebrates. Even though we did not observe any decrease in theoverall taxonomic richness, our results suggest that there is astrong possibility that even a small increase in conductivitycan lead to structural changes in macroinvertebrate commu-nities. However, since the analysis was based on a coarsetaxonomic resolution and using data from mainly low tomedium conductivity range, care should be taken when apply-ing proposed sensitivity scores to the assessment of risk posedby salinity. Increased taxonomic and geographical resolu-tion combined with more data from the higher conductivitystreams (brackish–saline categories) and using abundancedata instead of presence/absence are likely to improve theaccuracy and precision of the salinity index.
Acknowledgments
Authors would like to acknowledge Glenn McGregor(NR&M), Jason Dunlop (NR&M) for their valuable com-ments and Ben Stewart Koster (Griffith University) and MarkKennard (Griffith University) for suggestions regarding par-tial CCA. Financial support for this study was provided bythe National Action Plan for Salinity and Water Quality.
References
Ball, G. R., Palmer-Brown, D., and Mills, G. E. (2000). A com-parison of artificial neuronal network and conventional statisticaltechniques for analysing environmental data. In ‘Artificial NeuralNetworks: Application to Ecology and Evolution’. (Eds S. Lek andJ. F. Guégan.) p. 262. (Springer Verlag: Berlin.)
Bayly, I. A. E. (1969). The occurrence of calanoid copepods in atha-lassic saline waters in relation to salinity and ionic proportions.Vernandlungen Internationale Vereinigung fur Theoretische andAngewandthe Limnologie 17, 449–455.

Response of stream macroinvertebrates to salinity Marine and Freshwater Research 833
Berezina, N. A. (2003). Tolerance of freshwater invertebrates tochanges in water salinity. Russian Journal of Ecology 34, 261–266.doi:10.1023/A:1024597832095
Bishop, C. M. (1995). ‘Neural Networks for Pattern Recognition.’(Oxford University Press Inc.: New York.)
Bloedel, L., Wilhelm, G., Clarke, R., Horn, T., and Churchill, R. (2000).Water quality exceedence, trend and status assessment for Queens-land. Report 42. Queensland Department of Natural Resources,Brisbane.
Brosse, S., Giraudel, J. L., and Lek, S. (2001). Utilisation of non-supervised neural networks and principal component analysisto study fish assemblages. Ecological Modelling 146, 159–166.doi:10.1016/S0304-3800(01)00303-9
Bunn, S. E., and Davies, P. M. (1992). Community structure ofthe macroinvertebrate fauna and water quality of a saline riversystem in south-western Australia. Hydrobiologia 248, 143–160.doi:10.1007/BF00006082
Chessman, B. (2003). New sensitivity grades for Australian rivermacroinvertebrates. Marine and Freshwater Research 54, 95–103.doi:10.1071/MF02114
Conrick, D., and Cockayne, B. (2000). Queensland Australian RiverAssessment System (AusRivAs) sampling and processing man-ual. Queensland Department of Natural Resources. FreshwaterBiological Monitoring Unit, Brisbane.
Department of Environment and Heritage and Department of NaturalResources (1999). Testing the waters. A report of the quality ofQueensland waters. Report 21. Queensland Government, Brisbane.
Hart, B. T., Bailey, P., Edwards, R., Hortle, K., James, K., andMcMahon, A. (1991). A review of the salt sensitivity of theAustralian freshwater biota. Hydrobiologia 210, 105–144.
Hart, B. T., Lake, P. S., Webb, J. A., and Grace, M. R. (2003). Ecologicalrisk to aquatic systems from salinity increases. Australian Journalof Botany 51, 689–702. doi:10.1071/BT02111
Haykin, S. (1999). ‘Neural Networks: A Comprehensive Foundation.’(Prentice Hall: Englewood Cliffs, NJ.)
Hoang, H., Recknagel, F., Marshall, J., and Choy, S. (2001). Predic-tive modelling of macroinvertebrate assemblages for stream habitatassessments in Queensland (Australia). Ecological Modelling 146,195–206. doi:10.1016/S0304-3800(01)00306-4
Hoang, H., Recknagel, F., Marshall, J., and Choy, S. (2003). Elu-cidation of hypothetical relationships between habitat conditionsand macroinvertebrate assemblages in freshwater streams by arti-ficial neural networks. In ‘Ecological Informatics. UnderstandingEcology by Means of Biologically-inspired Computation’. (Ed.F. Reckangel.) pp. 179–192. (Springer-Verlag: New York.)
Goetsch, P. A., and Palmer, C. G. (1997). Salinity tolerance of selectedmacroinvertebrates of the Sabie River, Kruger National Park, SouthAfrica. Archives of Environmental Contamination and Toxicology32, 32–41. doi:10.1007/S002449900152
James, K. J., Cant, B., and Ryan, T. (2003). Responses of freshwa-ter biota to rising salinity levels and implications for saline watermanagement: a review. Australian Journal of Botany 51, 703–713.doi:10.1071/BT02110
Kay, W. R., Halse, S. A., Scanlon, M. D., and Smith, M. J. (2001). Dis-tribution and environmental tolerances of aquatic macroinvertebratefamilies in the agricultural zone of southwestern Australia. Journalof the North American Benthological Society 20(2), 182–199.
Kefford, B. J. (1998). The relationship between electrical conductivityand selected macroinvertebrate communities in four river systemsof south-west Victoria, Australia. International Journal of Salt LakeResearch 7, 151–170.
http://www.publish.csiro.au/journals/mfr
Kefford, B. J., and Papas, P. J., Nugegoda, D. (2003). Relativesalinity tolerance of macroinvertebrates from the Barwon River,Victoria, Australia. Marine and Freshwater Research 54, 755–765.doi:10.1071/MF02081
Kefford, B. J., Dalton, A., Palmer, C. G., and Nugegoda, D. (2004). Thesalinity tolerance of eggs and hatchlings of selected aquatic macroin-vertebrates in south-east Australia and South Africa. Hydrobiologia517, 179–192. doi:10.1023/B:HYDR.0000027346.06304.BC
Lek, S., and Guegan, J. F. (1999). Artificial neural networks as a toolin ecological modelling, an introduction. Ecological Modelling 120,65–73. doi:10.1016/S0304-3800(99)00092-7
Leland, H.V., and Fend, S.V. (1998). Benthic invertebrate distributions inthe San Joaquin River, California, in relation to physical and chemi-cal factors. Canadian Journal of Fisheries and Aquatic Sciences 55,1051–1067. doi:10.1139/CJFAS-55-5-1051
Marshall, J., Hoang, H., Choy, S., and Rechnagel, F. (2002). Relation-ships between habitat properties and the occurrence of macroinverte-brates in Queensland streams (Australia) discovered by a sensitivityanalysis with artificial neural networks. Vernandlungen Interna-tionale Vereinigung fur Theoretische and Angewandthe Limnologie28, 1–5.
Marshall, N.A., and Bailey, P. C. E. (2004). Impact of secondary salinisa-tion on freshwater ecosystems: effect of contrasting, experimental,short-term releases of saline wastewater on macroinvertebrates ina lowland stream. Marine and Freshwater Research 55, 509–523.doi:10.1071/MF03018
McNeil, V. H., Cox, M. E., and Preda, M. (2005). Assessment of chem-ical water types and their spatial variation using multi-stage clusteranalysis, Queensland,Australia. Journal of Hydrology 310, 181–200.
Metzeling, L. (1993). Benthic macroinvertebrate community structurein streams of different salinitites. Australian Journal of Marine andFreshwater Research 44, 335–351.
Principe, J. C., Euliano, N. R., and Lefebvre, W. C. (2000). ‘Neuraland Adaptive Systems. Fundamentals Through Simulations.’ (JohnWiley & Son, Inc.: New York.)
Poff, N. L. (1996). Stream hydrological and ecological responses toclimate change assessed with an artificial neural network. Limnologyand Oceanography 41(5), 857–863.
Olden, J. D., and Jackson, D. A. (2002). A comparison of statisti-cal approaches for modelling fish species distributions. FreshwaterBiology 47, 1976–1995. doi:10.1046/J.1365-2427.2002.00945.X
Williams, D. D.,Williams, N. E., andYong, C. (2000). Road salt contami-nation of groundwater in a major metropolitan area and developmentof a biological index to monitor its impact. Water Research 34(1),127–138. doi:10.1016/S0043-1354(99)00129-3
Ter Braak, C. J. F. (1988). Partial canonical correspondence anal-ysis. In ‘Classification and Related Methods of Data Analysis’.(Ed. H. H. Bock.) pp. 551–558. (Elsevier: Amsterdam.)
Ter Braak, C. J. F., and Verdonschot, P. F. M. (1995). Canonical corre-spondence analysis and related multivariate methods in aquatic ecol-ogy. Aquatic Sciences 57(3), 255–285. doi:10.1007/BF00877430
Williams, W. D., Boulton, A. J., and Taaffe, R. G. (1990). Salinity asdeterminant of salt lake fauna: a question of scale. Hydrobiologia197, 257–266. doi:10.1007/BF00026955
Williams, W. D., Taaffe, R. G., and Boulton, A. J. (1991). Longitu-dinal distribution of macroinvertebrates in two rivers subject tosalinization. Hydrobiologia 210, 151–160.
Manuscript received 9 September 2004; revised 22 March 2005; andaccepted 27 May 2005.
Copyright © 2022 FDOKUMEN




















