
Different Eucalyptus Species Show Different Mechanisms of Tolerance to Salinity and Salinity ×...
14
PLEASE SCROLL DOWN FOR ARTICLE This article was downloaded by: [Nasim, Muhammad] On: 6 August 2009 Access details: Access Details: [subscription number 913571864] Publisher Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Journal of Plant Nutrition Publication details, including instructions for authors and subscription information: http://www.informaworld.com/smpp/title~content=t713597277 Different Eucalyptus Species Show Different Mechanisms of Tolerance to Salinity and Salinity × Hypoxia Muhammad Nasim a ; Riaz H. Qureshi b ; Tariq Aziz a ; M. Saqib b ; Shafqat Nawaz a ; J. Akhtar b ; M. A. Haq b ; Shahbaz Talib Sahi a a Sub-Campus Depalpur, University of Agriculture, Faisalabad, Depalpur, Pakistan b Saline Agriculture Research Centre, University of Agriculture, Faisalabad, Depalpur, Pakistan Online Publication Date: 01 September 2009 To cite this Article Nasim, Muhammad, Qureshi, Riaz H., Aziz, Tariq, Saqib, M., Nawaz, Shafqat, Akhtar, J., Haq, M. A. and Sahi, Shahbaz Talib(2009)'Different Eucalyptus Species Show Different Mechanisms of Tolerance to Salinity and Salinity × Hypoxia',Journal of Plant Nutrition,32:9,1427 — 1439 To link to this Article: DOI: 10.1080/01904160903092648 URL: http://dx.doi.org/10.1080/01904160903092648 Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf This article may be used for research, teaching and private study purposes. Any substantial or systematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expressly forbidden. The publisher does not give any warranty express or implied or make any representation that the contents will be complete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should be independently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with or arising out of the use of this material.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Different Eucalyptus Species Show Different Mechanisms of Tolerance to Salinity and Salinity ×...

PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Nasim, Muhammad]On: 6 August 2009Access details: Access Details: [subscription number 913571864]Publisher Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
Journal of Plant NutritionPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713597277
Different Eucalyptus Species Show Different Mechanisms of Tolerance toSalinity and Salinity × HypoxiaMuhammad Nasim a; Riaz H. Qureshi b; Tariq Aziz a; M. Saqib b; Shafqat Nawaz a; J. Akhtar b; M. A. Haq b;Shahbaz Talib Sahi a
a Sub-Campus Depalpur, University of Agriculture, Faisalabad, Depalpur, Pakistan b Saline AgricultureResearch Centre, University of Agriculture, Faisalabad, Depalpur, Pakistan
Online Publication Date: 01 September 2009
To cite this Article Nasim, Muhammad, Qureshi, Riaz H., Aziz, Tariq, Saqib, M., Nawaz, Shafqat, Akhtar, J., Haq, M. A. and Sahi,Shahbaz Talib(2009)'Different Eucalyptus Species Show Different Mechanisms of Tolerance to Salinity and Salinity ×Hypoxia',Journal of Plant Nutrition,32:9,1427 — 1439
To link to this Article: DOI: 10.1080/01904160903092648
URL: http://dx.doi.org/10.1080/01904160903092648
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

Journal of Plant Nutrition, 32: 1427–1439, 2009
Copyright © Taylor & Francis Group, LLC
ISSN: 0190-4167 print / 1532-4087 online
DOI: 10.1080/01904160903092648
Different Eucalyptus Species Show DifferentMechanisms of Tolerance to Salinity
and Salinity × Hypoxia
Muhammad Nasim,1 Riaz H. Qureshi,2 Tariq Aziz,1 M. Saqib,2
Shafqat Nawaz,1 J. Akhtar,2 M. A. Haq,2 and Shahbaz Talib Sahi1
1Sub-Campus Depalpur, University of Agriculture, Faisalabad,Depalpur, Pakistan
2Saline Agriculture Research Centre, University of Agriculture, Faisalabad,Depalpur, Pakistan
ABSTRACT
We studied the effect of sodium chloride (NaCl) salinity and oxygen deficiency stresson growth and leaf ionic composition of three Eucalyptus species [E. tereticornis, E.camaldulensis (Silverton), and E. camaldulensis (Local)]. Species were grown withcontrol (no NaCl) and salinity (150 mol m−3 NaCl) under hypoxic and non-hypoxicconditions in nutrient solution with five replications following CRD. Species differedsignificantly in their response to salinity and hypoxia. Absolute shoot dry matter wassignificantly better in E. camaldulensis (Silverton) in salinity and in E. camaldulensis(Local) in saline hypoxic treatment. E. tereticornis was the most sensitive species tosalinity and salinity × hypoxia in the root environment. Sodium (Na+) and chloride (Cl−)concentrations were significantly lower in E. camaldulensis (Local) in non-hypoxicsaline treatment compared to the other two species. E. camaldulensis (Silverton) seemsto have better tissue compartmentalization, whereas E. camaldulensis (local) seems tohave better exclusion of Na+ at the root level.
Keywords: salinity, hypoxia, Eucalyptus, leaf expansion, growth, ionic composition
INTRODUCTION
Desertification of arable lands is a serious threat to agriculture around the globe.Salinization is an important factor contributing to the degradation of the arable
Received 29 October 2007; accepted 17 July 2008.Address correspondence to Dr. Muhammad Nasim, University of Agriculture,
Faisalabad, Sub-Campus Depalpur, Depalpur, Okara, Pakistan. E-mail: [email protected]
1427
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

1428 M. Nasim et al.
lands particularly in arid and semi-arid regions (Ashraf, 1994). Approximately6.3 million hectares of agricultural land in Pakistan is affected by varyingdegrees of salinity/sodicity (Ghafoor et al., 2004) that adversely affects plantgrowth and yield in a number of species (Eom et al., 2007). Soil salinity affectsplant growth mainly through osmotic effects and specific ion toxicities (Grattanand Grieve, 1999; Munns, 2002; 2005) resulting from accumulation of sodiumand chloride in the plant tissues (Saqib et al., 2004a; 2005a; Rezaei et al.,2006). Sodium (Na) concentration in tissues and potassium (K+): Na+ ratiosare extensively used in screening mass germplasms as selection parameters(Saqib et al., 2004a; Tahir et al., 2006).
Salt affected soils (mainly those with high sodium) are generally dispersedsoils having low soil permeability, porosity, and hydraulic conductivity (Qadiret al., 2005). Low water infiltration and soil dispersion poses a serious drainageproblem and may lead to waterlogging. Plant roots affected by waterlogging aredeprived of sufficient oxygen resulting in a change in the mode of respirationfrom aerobic to an-aerobic and low energy production (Marschner, 1995). Lowenergy production in roots disturbs the nutrient and water uptake (Jackson,1979; Morard and Silvestre, 1996). Waterlogging also reduces nutrient uptakethat results in nutrient deficiency in shoot leading to reduced shoot growth(Trought and Drew, 1980). Higher abscisic acid production and movementto younger leaves under oxygen (O2) deficiency causes stomatal closure inplants (Zhang and Zhang, 1994) affecting photosynthesis and ultimately yield.Waterlogging also affects sodium exclusion from plant roots, which is a majorsalinity tolerance mechanism in various glycophytes (Barrett-Lennard, 1986;Saqib et al. 2005a). A combined effect of salinity and waterlogging on plantgrowth is more damaging than caused by salinity and waterlogging alone(Qureshi and Barrett-Lennard, 1998; Saqib et al., 2004a).
Although several relatively salt tolerant cultivars of different agronomiccrops are available to grow on moderately salt affected soils yet these culti-vars fail to produce economical yields on highly degraded salt affected soils.Reclamation of such soils through physical and chemical approaches is alsonot feasible. Revegetation of these lands with salt tolerant tree species is a vi-able approach mainly because of low input and increased demand of wood forfuel and furniture. Selection of suitable tree species is a pre-requisite for suchbiological remediation approach. However, very little work has been reportedon this aspect particularly on tree species. The present paper reports the effectof salinity and hypoxia interaction on growth and leaf ionic composition ofthree Eucalyptus species.
MATERIALS AND METHODS
Site and Environmental Conditions
The experiment was conducted in a rain-protected wire house at the Insti-tute of Soil & Environmental Sciences, University of Agriculture, Faisalabad,
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

Salinity × Hypoxia Interaction in Eucalyptus 1429
Pakistan. Faisalabad is situated at 73.4◦ longitude and 31.5◦ latitude. The aver-age day and night temperatures during the study were 32 and 20◦C, respectively,while the average maximum relative humidity was 70%, and there was 7 h av-erage daily sunshine during the growth period.
Plant Material and Experimental Details
Seeds of three Eucalyptus species [E. tereticornis, E. camaldulensis (Silver-ton), and E. camaldulensis (Local)] were collected from Australian Revegeta-tion Corporation Limited, Western Australia and Saline Agriculture ResearchCentre, University of Agriculture, Faisalabad, Pakistan. Fifty seeds of eachspecies were sown in polyethylene lined iron trays containing silica gravel andsynthetic vermiculite (mixed in 1:1 ratio). The canal water was used to moistenthe seeds for proper germination. Electrical conductivity of canal water was0.29 d Sm−1. One week after germination, half strength Hoagland’s nutrientsolution (Hoagland and Arnon, 1950) was used for the seedling establishment.Seedlings were grown for three months in these trays. Uniform sized seedlingswere then transplanted in foam plugged holes in polystyrene sheets floatingon continuously aerated half strength Hoagland’s nutrient solution containedin polyethylene lined iron tubs of 200 L capacity (100 × 100 × 30 cm3).The experiment was replicated five times following completely randomizeddesign (CRD). Solution pH was monitored and maintained daily at 6 ± 0.5.There were four treatments viz i) non-saline non-hypoxic (control), ii) salinenon-hypoxic, iii) non-saline hypoxic, and iv) saline-hypoxic. Salinity level forsaline treatment was 150 mol m−3 sodium chloride (NaCl). The salinity wasdeveloped in three equal splits in a week (each of 50 mol m−3 NaCl after 2d). The hypoxia was imposed by surface sealing of the nutrient solution. Threedays after surface sealing, substrate oxygen concentration (measured with O2
electrode) was 3 mg dm−3. The treatment solutions were replaced thereafterweekly till harvesting and in hypoxic treatments the new solution was flushedwith N2 for 15 minutes to remove oxygen.
Growth Measurement
To study leaf expansion (increase in leaf length) 2-day old leaf of each plantfrom shoot apex was marked and leaf length was measured at 2 day (d) intervalup to 11th day. After 8 weeks of salinity and hypoxia stress plant height wasrecorded. At this time plants were harvested and their dry weights of shootsand roots were recorded after oven drying the samples at 70◦C in a vacuumoven for 48 hrs.
Leaf Ionic Analysis
Plants were harvested after 8 weeks and leaves of plants were divided into topleaves (young leaves) and lower fully expanded leaves (mature leaves). The
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

1430 M. Nasim et al.
separated leaves were immediately washed with distilled water and blotted drywith tissue paper. The samples were then dried at 70◦C in a forced air drivenoven for 48 h. The oven dried plant samples were fine ground in a wily mill topass through 1 mm sieve. The fine ground plant samples (1 g) were digested intri-acid mixture (sulfuric acid, nitric acid, and perchloric acid) (Miller, 1998).Potassium and Na+ were determined on a flame photometer (Jenway PFP-7, Bibby Scientific LTD., Essex England). For chloride determination, plantsamples were extracted with HNO3 and chloride was determined from thisextract using chloride analyzer (Corning Chloride Analyzer 926, Corning Inc.,Corning, NY, USA).
Statistical Analysis
The data were analyzed statistically following the methods of Gomez andGomez (1984) using MSTAT-C (Russell and Eisensmith, 1983). The signif-icance of differences among the means was compared using standard errorcomputed as s/
√n, where s is the standard deviation and n shows the number
of observations.
RESULTS
Shoot and Root Growth
Salinity significantly reduced shoot dry matter of all the species at both non-hypoxic and hypoxic conditions (Figure 1). E. camaldulensis (Silverton) pro-duced the maximum absolute and relative shoot dry matter (36%) under non-hypoxic salinity followed by E. camaldulensis (Local) and E. tereticornis.However, under hypoxic salinity treatment E. camaldulensis (Local) performedbest followed by E. camaldulensis (Silverton) and E. tereticornis, in absoluteas well as relative terms. Hypoxia alone reduced the shoot and root dry matterof E. tereticornis only. Root dry matter (RDM) of Eucalyptus species was alsosignificantly reduced by salinity in hypoxic conditions (Figure 2). E. camal-dulensis (Local) and E. tereticornis produced respectively, the maximum andminimum root dry matter in this treatment. However, in non-hypoxic salinitytreatment E. camaldulensis (Silverten) and E. camaldulensis (Local) differednon-significantly. Root: shoot ratio (RSR) of Eucalyptus species was increasedsignificantly under saline conditions both in hypoxic and non-hypoxic treat-ments, whereas hypoxia alone did not affect it significantly (data not shown).Salinity also significantly reduced leaf expansion of E. tereticornis and E.camaldulensis (Local) under non-hypoxic as well as hypoxic conditions (Fig-ure 3). Plant height of all the species was also reduced significantly by salinityand salinity × hypoxia (data not shown).
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009
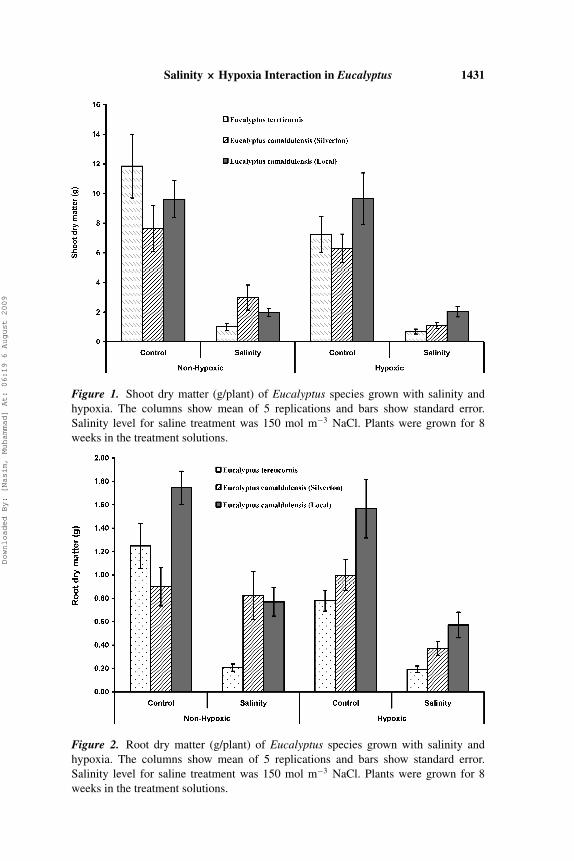
Salinity × Hypoxia Interaction in Eucalyptus 1431
Figure 1. Shoot dry matter (g/plant) of Eucalyptus species grown with salinity andhypoxia. The columns show mean of 5 replications and bars show standard error.Salinity level for saline treatment was 150 mol m−3 NaCl. Plants were grown for 8weeks in the treatment solutions.
Figure 2. Root dry matter (g/plant) of Eucalyptus species grown with salinity andhypoxia. The columns show mean of 5 replications and bars show standard error.Salinity level for saline treatment was 150 mol m−3 NaCl. Plants were grown for 8weeks in the treatment solutions.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009
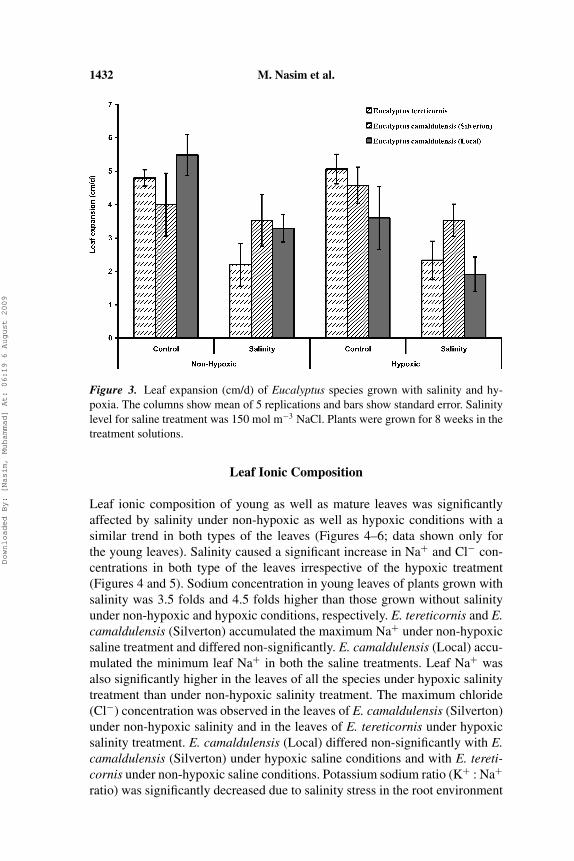
1432 M. Nasim et al.
Figure 3. Leaf expansion (cm/d) of Eucalyptus species grown with salinity and hy-poxia. The columns show mean of 5 replications and bars show standard error. Salinitylevel for saline treatment was 150 mol m−3 NaCl. Plants were grown for 8 weeks in thetreatment solutions.
Leaf Ionic Composition
Leaf ionic composition of young as well as mature leaves was significantlyaffected by salinity under non-hypoxic as well as hypoxic conditions with asimilar trend in both types of the leaves (Figures 4–6; data shown only forthe young leaves). Salinity caused a significant increase in Na+ and Cl− con-centrations in both type of the leaves irrespective of the hypoxic treatment(Figures 4 and 5). Sodium concentration in young leaves of plants grown withsalinity was 3.5 folds and 4.5 folds higher than those grown without salinityunder non-hypoxic and hypoxic conditions, respectively. E. tereticornis and E.camaldulensis (Silverton) accumulated the maximum Na+ under non-hypoxicsaline treatment and differed non-significantly. E. camaldulensis (Local) accu-mulated the minimum leaf Na+ in both the saline treatments. Leaf Na+ wasalso significantly higher in the leaves of all the species under hypoxic salinitytreatment than under non-hypoxic salinity treatment. The maximum chloride(Cl−) concentration was observed in the leaves of E. camaldulensis (Silverton)under non-hypoxic salinity and in the leaves of E. tereticornis under hypoxicsalinity treatment. E. camaldulensis (Local) differed non-significantly with E.camaldulensis (Silverton) under hypoxic saline conditions and with E. tereti-cornis under non-hypoxic saline conditions. Potassium sodium ratio (K+ : Na+
ratio) was significantly decreased due to salinity stress in the root environment
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

Salinity × Hypoxia Interaction in Eucalyptus 1433
Figure 4. Sodium concentration (mmoles kg−1) in young leaves of Eucalyptus speciesgrown with salinity and hypoxia. The columns show means of 5 replications and barsshow standard error. Salinity level for saline treatment was 150 mol m−3 NaCl. Plantswere grown for 8 weeks in the treatment solutions.
Figure 5. Chloride concentration (mmoles kg−1) in young leaves of Eucalyptus speciesgrown with salinity and hypoxia. The columns show means of 5 replications and barsshow standard error. Salinity level for saline treatment was 150 mol m−3 NaCl. Plantswere grown for 8 weeks in the treatment solutions.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009
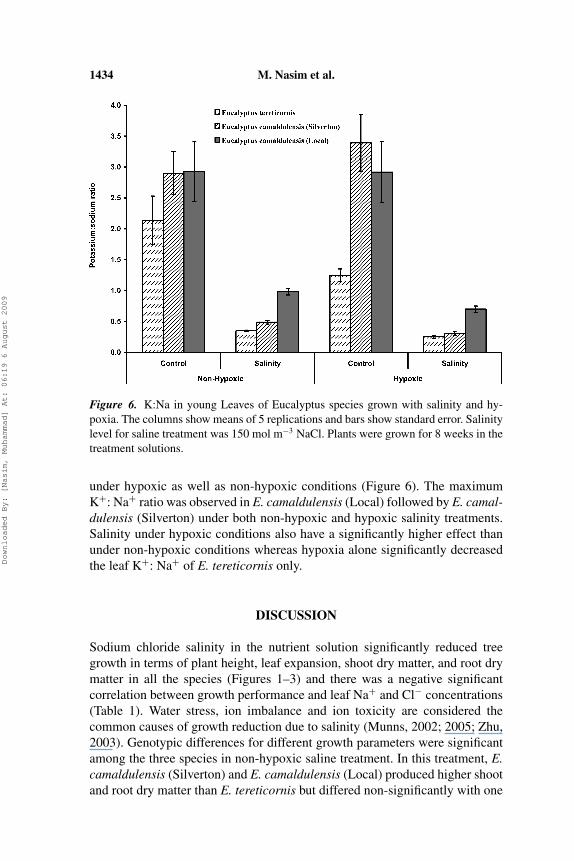
1434 M. Nasim et al.
Figure 6. K:Na in young Leaves of Eucalyptus species grown with salinity and hy-poxia. The columns show means of 5 replications and bars show standard error. Salinitylevel for saline treatment was 150 mol m−3 NaCl. Plants were grown for 8 weeks in thetreatment solutions.
under hypoxic as well as non-hypoxic conditions (Figure 6). The maximumK+: Na+ ratio was observed in E. camaldulensis (Local) followed by E. camal-dulensis (Silverton) under both non-hypoxic and hypoxic salinity treatments.Salinity under hypoxic conditions also have a significantly higher effect thanunder non-hypoxic conditions whereas hypoxia alone significantly decreasedthe leaf K+: Na+ of E. tereticornis only.
DISCUSSION
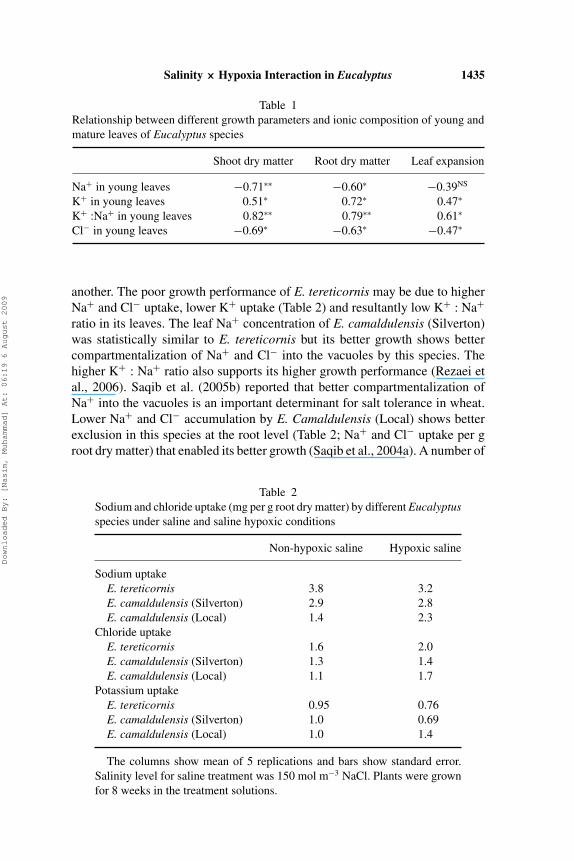
Sodium chloride salinity in the nutrient solution significantly reduced treegrowth in terms of plant height, leaf expansion, shoot dry matter, and root drymatter in all the species (Figures 1–3) and there was a negative significantcorrelation between growth performance and leaf Na+ and Cl− concentrations(Table 1). Water stress, ion imbalance and ion toxicity are considered thecommon causes of growth reduction due to salinity (Munns, 2002; 2005; Zhu,2003). Genotypic differences for different growth parameters were significantamong the three species in non-hypoxic saline treatment. In this treatment, E.camaldulensis (Silverton) and E. camaldulensis (Local) produced higher shootand root dry matter than E. tereticornis but differed non-significantly with one
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009
Salinity × Hypoxia Interaction in Eucalyptus 1435
Table 1Relationship between different growth parameters and ionic composition of young andmature leaves of Eucalyptus species
Shoot dry matter Root dry matter Leaf expansion
Na+ in young leaves −0.71∗∗ −0.60∗ −0.39NS
K+ in young leaves 0.51∗ 0.72∗ 0.47∗
K+ :Na+ in young leaves 0.82∗∗ 0.79∗∗ 0.61∗
Cl− in young leaves −0.69∗ −0.63∗ −0.47∗
another. The poor growth performance of E. tereticornis may be due to higherNa+ and Cl− uptake, lower K+ uptake (Table 2) and resultantly low K+ : Na+
ratio in its leaves. The leaf Na+ concentration of E. camaldulensis (Silverton)was statistically similar to E. tereticornis but its better growth shows bettercompartmentalization of Na+ and Cl− into the vacuoles by this species. Thehigher K+ : Na+ ratio also supports its higher growth performance (Rezaei etal., 2006). Saqib et al. (2005b) reported that better compartmentalization ofNa+ into the vacuoles is an important determinant for salt tolerance in wheat.Lower Na+ and Cl− accumulation by E. Camaldulensis (Local) shows betterexclusion in this species at the root level (Table 2; Na+ and Cl− uptake per groot dry matter) that enabled its better growth (Saqib et al., 2004a). A number of
Table 2Sodium and chloride uptake (mg per g root dry matter) by different Eucalyptusspecies under saline and saline hypoxic conditions
Non-hypoxic saline Hypoxic saline
Sodium uptakeE. tereticornis 3.8 3.2E. camaldulensis (Silverton) 2.9 2.8E. camaldulensis (Local) 1.4 2.3
Chloride uptakeE. tereticornis 1.6 2.0E. camaldulensis (Silverton) 1.3 1.4E. camaldulensis (Local) 1.1 1.7
Potassium uptakeE. tereticornis 0.95 0.76E. camaldulensis (Silverton) 1.0 0.69E. camaldulensis (Local) 1.0 1.4
The columns show mean of 5 replications and bars show standard error.Salinity level for saline treatment was 150 mol m−3 NaCl. Plants were grownfor 8 weeks in the treatment solutions.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

1436 M. Nasim et al.
researchers have reported significant variations in the growth and salt toleranceamong different species of the woody plants (Bell et al., 1994; Marcar et al.,1995; Rawat and Banerjee, 1998). Many earlier researchers have used Na+ :K+ ratio as an indicator of salinity tolerance (Saqib et al., 2004b; Munns, 2005;Rezaei et al., 2006) as adequate Na+ : K+ ratio in the cytosol is essential fornormal cellular functions of plants (Marschner, 1995; Chinnusamy et al., 2005).Higher levels of external Na+ in saline soils interfere with K+ acquisition byplants (Subbarao et al., 1990). The species with ability to maintain K+ uptakeor to exclude Na+ can tolerate salinity stress such as E. camaldulensis (local).The lower Na+ uptake and higher K+ uptake at root level, and higher K+ :Na+ ratio in this species is in accordance with its better performance at salineenvironment (Table 2; Figure 6).
Low oxygen supply in hypoxia affects root growth because energy pro-duction is decreased (Barret-Lennard, 1986). Under hypoxia, shoot and rootgrowth retardation has also been reported by several researchers (Jackson,1979; Lizaso et al., 2001). In the present study, hypoxia significantly reducedthe shoot and root growth of the salt sensitive species E. tereticornis only (Fig-ures 1-2). Aerenchyma development in the roots is an important mechanism ofhypoxia tolerant (Saqib et al., 2005a) that may not have developed considerablyin this species. Marcar et al. (1995) also observed large variations among theeucalyptus species for their tolerance to oxygen deficiency (waterlogging). Insaline-hypoxic conditions roots are not able to exclude Na+ and Cl− mainlybecause of low energy production (Qureshi and Barret-Lennard, 1998). Thislower ability to exclude toxic elements can further inhibit K+ uptake by plantroots. Saqib et al. (2004a) reported a higher accumulation of Na+ and Cl− andlower K+ in leaf sap under saline waterlogged conditions than under salineconditions. Marcar et al. (1993) and Galloway and Davidson (1993) observedsimilar depressive effects of salinity × hypoxia interaction on growth of Euca-lyptus and Atriplexes.
However, in the present study higher reduction in shoot and root growthwas observed only in E. camaldulensis (Silverton) in saline hypoxic treatmentthan in the non-hypoxic saline treatment (Figures 1–2). It may be due to itsdecreased root ability to develop aerenchyma that resulted in low Na+ exclusionand K+ uptake at the root level (Figure 4; Table 2) as aerenchyma helps in rootNa+ exclusion (Saqib et al., 2005b). In this treatment E. camaldulensis (local)performed significantly better than the other species which may be due to itsbetter aerenchyma and nodal root development and hence better salt exclusionat the root level. In conclusion, E. camaldulensis (Silverton) is better tolerantto salinity alone and E. camaldulensis (local) is better tolerant to saline andhypoxic conditions. E. camaldulensis (Silverton) seems to have better tissuecompartmentalization whereas E. camaldulensis (local) seems to have betterexclusion of Na+ at the root level.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

Salinity × Hypoxia Interaction in Eucalyptus 1437
REFERENCES
Ashraf, M. 1994. Breeding for salinity tolerance in plants. Critical Reviews inPlant Sciences 13: 17–42.
Barrett-Lennard, E. G. 1986. Effects of waterlogging on the growth and NaCluptake by vascular plants under saline conditions. Reclamation and Reveg-etation Research 5: 245–261.
Bell, D. T., J. A. McComb, P. G. Vander-Moezel, I. J. Bennett, andE. D. Kabay. 1994. Comparisons of selected and cloned plantletsagainst seedlings for rehabilitation of saline and waterlogged dischargezones in Australia Agricultural Catchments. Australian Forestry 57: 69–75.
Chinnusamy, V., A. Jagendorf, and J.-K. Zhu. 2005. Understanding and im-proving salt tolerance in plants. Crop Science 45: 437–448.
Eom, S. H., T. L. Setter, A. DiTommaso, and L. A. Weston. 2007. Differentialgrowth response to salt stress among selected ornamentals. Journal ofPlant Nutrition 30: 1109–1126.
Galloway, R., and N. J. Davidson. 1993. The response of Atriplex amnicolato the interactive effects of salinity and hypoxia. Journal of ExperimentalBotany 44: 653–663.
Ghafoor, A., M. Qadir, and G. Murtaza. 2004. Salt Affected Soils: Principlesof Management. Lahore, Pakistan: Allied Book Centre.
Gomez, K. A., and A. A. Gomez. 1984. Statistical Procedures for AgriculturalResearch. New York: Wiley.
Grattan, S. R., and C. M. Grieve. 1999. Salinity-mineral nutrient relations inhorticultural crops. Scientia Horticulturae 78: 127–157.
Hoagland, D. R., and D. I. Arnon. 1950. The water culture method for growingplants without soil. California Agricultural Experimental Station Circular347: 1–32.
Jackson, M. B. 1979. Rapid injury to peas by soil water logging. Journal of theScience of Food and Agriculture 30: 143–152.
Lizaso, J. I., L. M. Melendez, and R. Ramirez. 2001. Early flooding of twocultivars of tropical maize. I. Shoot and root growth. Journal of PlantNutrition 24: 979–995.
Marcar, N. E., D. F. Crawford, and P. M. Leppert. 1993. The potential of treesfor utilization and management of salt affected land. In: Productive Use ofSaline Land, eds. N. Davidson and R. Galloway. ACIAR Proceeding 42: 17–42.
Marcar, N. E., D. F. Crawford, P. M. Leppert, T. Jovanovic, R. Floyd, and R.Rarrow. 1995. Trees for Salt Land: A Guide for Selecting Native Speciesfor Australia. Canberra, Australia: CSIRO.
Marschner, H. 1995. Mineral Nutrition of Higher Plants. London: AcademicPress.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

1438 M. Nasim et al.
Miller, R. O. 1998. Nitric-percloric wet digestion in an open vessel. In: Hand-book of Reference Methods for Plant Analysis, eds. Y. P. Kalra, pp. 57–62.Boca Raton, FL: CRC Press.
Morard, P., and J. Silvestre. 1996. Plant injury due to oxygen deficiency in theroot environment of soil less culture: A review. Plant Soil 184: 243–254.
Munns, R. 2002. Comparative physiology of salt and water stress. Plant CellEnvironment 25: 239–250.
Munns, R. 2005. Genes and salt tolerance: Bringing them together. New Phy-tologist 167: 645–663.
Qadir, M., A. D. Noble, J. D. Oster, S. Schubert, and A. Ghafoor. 2005. Drivingforces for sodium removal during phytoremediation of calcareous sodicand saline sodic soils: A review. Soil Use and Management 21(2): 173–180.
Qureshi, R. H., and E. G. Barrett-Lennard. 1998. Saline Agriculture for Ir-rigated Land in Pakistan: A Handbook. Canberra, Australia: AustralianCentre for International Agricultural Research.
Rawat, J. S., and S. P. Banerjee. 1998. The influence of salinity on growth,biomass production and photosynthesis of Eucalyptus camaldulensisDehnh. and Dalbergia sissoo Roxb. seedlings. Plant and Soil 205: 163–169.
Rezaei, H., N. A. K. K. Sima, M. J. Malakouti, and M. Pessarakli. 2006. Salttolerance of canola in relation to accumulation and xylem transportationof cations. Journal of Plant Nutrition 29: 1903–1917.
Russell, D. F., and S. P. Eisensmith. 1983. MSTAT-C. Lansing, MI: MichiganState University.
Saqib, M., J. Akhtar, and R. H. Qureshi. 2004a. Pot study on wheat growth insaline and waterlogged compacted soil. I. Grain yield and yield compo-nents. Soil and Tillage Research 77: 169–177.
Saqib, M., J. Akhtar, and R. H. Qureshi. 2004b. Pot study on wheat growthin saline and waterlogged compacted soil II. Root growth and leaf ionicrelations. Soil & Tillage Research 77: 179–187.
Saqib, M., J. Akhtar and R. H. Qureshi. 2005a. Na+ exclusion and salt resis-tance of wheat (Triticum aestivum) in saline waterlogged conditions areimproved by the development of adventitious nodal roots and cortical rootaerenchyma. Plant Science 169: 125–130.
Saqib, M., C. Zorb, Z. Rengel, and S. Schubert. 2005b. Na+ exclusion and saltresistance of wheat (Triticum aestivum) are improved by the expressionof endogenous vacuolar Na+ /H+ anitporters in roots and shoots. PlantScience 169: 959–965.
Subbarao, G. V., C. Johansen, M. K. Jana, and J. F. D. K. Kumar-Rao. 1990. Ef-fects of sodium/calcium ratio in modifying salinity response of pigeonpea(Cajanus cajan). Journal of Plant Physiology 136: 439–443.
Tahir, M. A., Rahmatullah, T. Aziz, M. Ashraf, S. Kanwal, and M. A. Maqsood.2006. Beneficial effects of silicon in wheat (Triticum aestivum L.) undersalinity stress. Pakistan Journal of Botany 38: 1715–1722.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009

Salinity × Hypoxia Interaction in Eucalyptus 1439
Trought, M. C. T., and M. C. Drew. 1980. The development of waterloggingdamage in wheat seedlings (Triticum aestivum L) I. Shoot and root growthin relation to changes in the concentration of dissolved gas and solutes inthe soil solution. Plant and Soil 54: 77–94.
Zhang, J., and X. Zhang. 1994. Can early wilting of old leaves account for muchof the ABA accumulation in flooded pea plants? Journal of ExperimentalBotany 45: 1335–1342.
Zhu, J.-K. 2003. Regulation of ion homeostasis under salt stress. CurrentOpinion in Plant Biology 6: 441–445.
Downloaded By: [Nasim, Muhammad] At: 06:19 6 August 2009




















